
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Neurosci., 22 November 2019
Sec. Decision Neuroscience
Volume 13 - 2019 | https://doi.org/10.3389/fnins.2019.00724
This article is part of the Research TopicTemporal Dynamics of Reward Processing in Humans: From Anticipation to ConsummationView all 13 articles
 Sónia Ferreira1,2†
Sónia Ferreira1,2† Carlos Veiga1,2†
Carlos Veiga1,2† Pedro Moreira1,2
Pedro Moreira1,2 Ricardo Magalhães1,2
Ricardo Magalhães1,2 Ana Coelho1,2
Ana Coelho1,2 Paulo Marques1,2
Paulo Marques1,2 Carlos Portugal-Nunes1,2
Carlos Portugal-Nunes1,2 Nuno Sousa1,2,3
Nuno Sousa1,2,3 Pedro Morgado1,2,3*
Pedro Morgado1,2,3*Cognition can influence choices by modulation of decision-making processes. This cognitive regulation is defined as processing information, applying knowledge, and changing preferences to consciously modulate decisions. While cognitive regulation of emotions has been extensively studied in psychiatry, few works have detailed cognitive regulation of decision-making. Stress may influence emotional behavior, cognition, and decision-making. In addition, the brain regions responsible for decision-making are sensitive to stress-induced changes. Thus, we hypothesize that chronic stress may disrupt the ability to regulate choices. Herein, we used a functional magnetic resonance imaging task where fourteen control and fifteen chronically stressed students had to cognitively upregulate or downregulate their craving before placing a bid to obtain food. We found that stressed participants placed lower bids to get the reward and chose less frequently higher bid values for food. Nevertheless, we did not find neural and behavioral differences during cognitive regulation of craving. Our outcomes revealed that chronic stress impacts decision-making after cognitive regulation of craving by reducing the valuation of food rewards but not cognitive modulation itself. Importantly, our results need further validation with larger sample sizes.
Value-based decision-making is the ability to make a choice from competing courses of action/alternatives based on subjective values and possible outcomes attributed to them (Balleine, 2005). This process is carried out whenever a person chooses from different alternatives (e.g., choosing between eating an apple or an orange, or between going out or not). Different interacting systems are responsible for the valuation and action selection processes in the brain (Rangel et al., 2008). First, a valuation system computes the action values. A comparator system needs to evaluate the action values. An accumulator system receives and accumulates the value signals from the comparator system until the signal for one of the actions is sufficiently strong for the choice to be executed (Gold and Shadlen, 2007; Basten et al., 2010).
Values assigned to actions during the valuation process can be influenced by different factors such as the degree of risk or uncertainty of the action (Platt and Huettel, 2008; Rangel et al., 2008). Humans have a natural aversion to risky or uncertain choices and place less value on actions with temporal uncertain rewards or multiple sets of outcomes (Christopoulos et al., 2009; McGuire and Kable, 2012). Individuals often place higher values on immediate rewards rather than on future ones (Rangel et al., 2008). Social competition, cooperation, and concerns for the well-being of others also influence decision-making (Fehr and Camerer, 2007). Cognition can also influence choices through modulation of the decision-making processes. This cognitive regulation process may be defined as processing information, applying knowledge and changing preferences to consciously modulate our decisions. While cognitive regulation of emotional response has been extensively studied (Ochsner et al., 2004; Delgado et al., 2008; Wager et al., 2008), few works have detailed cognitive regulation of decision-making. A functional magnetic resonance imaging (fMRI) study where participants had to modulate their cravings for food showed that cognitive regulation affects decision-making through valuation regulation and behavioral control (Hutcherson et al., 2012). The ventromedial prefrontal cortex (vmPFC) is known to compute the value signal of decisions while the dorsolateral prefrontal cortex (dlPFC) modulates this signal during cognitive regulation tasks (Hare et al., 2009, 2011; Kober et al., 2010).
Cognitive regulation of both emotion and decision-making has a role in the treatment of several conditions (schizophrenia, bipolar disorder, depression, obesity, addiction, obsessive-compulsive disorder, and eating disorders) where emotional processing and decision-making are often impaired (Phillips et al., 2003; Ochsner et al., 2004). On the other hand, mental disorders such as schizophrenia, bipolar disorder, post-traumatic stress disorder, and depression are often associated with prolonged exposure to stress (Arnsten, 2015; Sousa, 2016). Stress impacts emotional processing leading to depressive and anxious behavior associated with alterations in amygdala-ventromedial-prefrontal pathways. Moreover, stress elicits cognitive impairments namely working memory and attentional deficits, poor decision-making (e.g., decreased reward sensitivity or increased influence of immediate rewards), behavioral inflexibility, and learning deficits. These cognitive differences are associated with changes in prefrontal and hippocampal regions (Sandi, 2013; Arnsten, 2015; Chen and Baram, 2016; Sousa, 2016). Additionally, the brain regions implicated in decision-making processes are sensitive to stress-induced changes. In fact, changes in fronto-striatal networks involved in behavioral decisions have been reported in both humans and rodents after chronic stress (Dias-Ferreira et al., 2009; Soares et al., 2012; Morgado et al., 2012, 2015a; Sousa, 2016; Magalhães et al., 2018). Thus, stress seems to influence the quality of decisions (Starcke and Brand, 2012; Morgado et al., 2015b; Bryce and Floresco, 2016; Chen and Baram, 2016) because cognitive control is diminished (Yu, 2016).
Stress has also an impact on appetite and eating behavior (Ans et al., 2018) and is one of the factors for development of eating and obesity-associated conditions (Razzoli et al., 2017). Usually, the production of adrenocorticotropic hormone (ACTH) by the anterior pituitary gland leads to the release of cortisol in the adrenal cortex to stimulate hunger and feeding behavior. High cortisol levels are associated with high insulin concentrations resulting in increased caloric intake or food craving (Adam and Epel, 2007). Stress might boost these pathways leading to an increase in food intake and appetite for high-caloric food, or also reduced reward sensitivity to low-caloric food (Razzoli et al., 2017; Ans et al., 2018; Berg Schmidt et al., 2018; Ferrer-Cascales et al., 2018), in agreement with previous stress decision-making studies demonstrating decreased reward sensitivity or increased influence of immediate rewards (Morgado et al., 2015b). Thus, stress seems to be associated with increased food reward sensitivity due to diminished self-control during food choice associated with decreased functional connectivity between the vmPFC and dlPFC (Neseliler et al., 2017) and increased connectivity between the vmPFC and subcortical regions (amygdala and striatum) (Tryon et al., 2013; Maier et al., 2015).
Herein, we used an fMRI task to clarify the impact of chronic stress on cognitive regulation of decisions. Our task consisted of cognitively upregulating or downregulating craving before placing a bid to obtain food. In addition to brain responses, we analyzed behavioral parameters (food valuation score and reaction time) associated with the task, and blood hormonal changes after the task (insulin, cortisol, and glucose). Regarding the previous findings, we hypothesize that chronic stress may disrupt the ability of individuals to regulate their choices. We expect that cognitive regulation deficits after chronic stress manifest by changes in the prefrontal cortex (vmPFC and dlPFC). Subsequently, these deficits lead to decision-making impairments, namely increased reward sensitivity, underlying brain response alterations in prefrontal and striatal regions. Moreover, we expect that chronic stress participants present augmented levels of insulin, glucose, and cortisol after the stimulation with food pictures due to an increased reward sensitivity to food.
We enrolled in this study medical students from the School of Medicine of University of Minho, Portugal. All students were healthy Caucasians, right-handed, and had a healthy body-mass index. One group was under normal academic activities [control group, n = 14; 9 females/5 males; median (range) 23.00 (3.00) years of age; education 17.00 (3.00) years] and the other included subjects on the long period of preparation for the medical licensing exam [chronic psychosocial stress condition; stress group, n = 15; 10 females/5 males; 24.00 (3.00) years of age; education 18.00 (0.00) years]. This work was conducted 1 to 3 months before the exam, but students usually start preparing 1 year before the exam. Subjects were eligible if they were at least 18 years old, reported no history of psychiatric or neurological conditions, traumatic brain lesion, or substance abuse, and were not on any psychiatric medication. The groups were matched for gender (chi-squared test χ2(1) = 0.02, p = 0.893) but not for age (Mann–Whitney test U = 169.00, p = 0.004, effect size r = 0.56) and education level (U = 210.00, p = 2.579 × 10–8, r = 0.93).
The study was performed in accordance with the Declaration of Helsinki and was approved by Ethics Subcommittee for the Life and Health Sciences of University of Minho, Portugal, and by the Ethics Committee of the Hospital of Braga, Portugal. All subjects were provided with written informed consent following description of the study goals and procedures.
Subjects filled a questionnaire to characterize gender, age, educational level, handedness, and ethnic origin. Weight and height were also measured to prevent the inclusion of participants with an unhealthy body mass index. Subjects were assessed with the 10-items Perceived Stress Scale (PSS-10) (Cohen et al., 1983; Morgado et al., 2013), the Beck Anxiety Inventory (BAI) (Beck et al., 1988), and the Beck Depression Inventory (BDI) (Beck et al., 1996). PSS-10 measures the extent to which participants perceived their life as unpredictable, uncontrollable, and overloaded during the previous month. The higher the score, the greater the intensity of perceived stress. BAI measures the severity of an individual’s anxiety during the previous week. Scores lower than 8 indicate minimal anxiety. Scores higher than 7, 15, and 25 indicate mild, moderate, and severe anxiety, respectively. BDI measures the severity of depression and can be used as a screening tool. Scores lower than 14 indicate minimal depression. Higher scores indicate more severe depressive symptoms.
Before the fMRI acquisition, samples of venous blood were collected from all participants into a 5 mL potassium ethylenediaminetetraacetic acid tube and a serum tube. We repeated the collection immediately after the fMRI acquisition. Pre-scan blood samples were used to measure cortisol, glucose, insulin, and ACTH serum levels. In post-scan samples, we repeated cortisol, glucose, and insulin serum measurements (ACTH measurement was not repeated due to technical constraints). The collection took place between 2 and 7 pm which assures small variation in cortisol levels during this period (Minkley and Kirchner, 2012). ACTH was measured based on solid-phase, two-site sequential chemiluminescent immunometric assay, and insulin with solid-phase, enzyme-labeled chemiluminescent immunometric assay (IMMULITE 2000, Siemens AG, Germany). Cortisol levels were assessed with competitive immunoassay based on direct chemiluminescent (ADVIA Centaur and Centaur XP, Siemens AG, Germany). Glucose was measured based on the hexokinase-glucose-6-phosphate method (Dimension Vista, Siemens AG, Germany). Standard procedures were applied following the manufacturer instructions.
Data related with psychological scales, laboratory values, and behavioral parameters were analyzed using IBM SPSS Statistics (version 24.0; IBM Corporation, United States). Kolmogorov–Smirnov and Shapiro–Wilk tests were used to assess for normality in the distribution of data. Comparisons between groups were carried out by parametric t-tests or repeated measures ANOVA (F-test, with Bonferroni correction for multiple comparisons for post hoc tests], or non-parametric Mann-Whitney U-tests. Differences were considered statistically significant if p < 0.05. Effect sizes were calculated for all statistically significant results.
The task was adapted from Hutcherson et al. (2012). Subjects were instructed to fast for at least 4 h before their arrival and to eat a light meal before the fasting period to increase the valuation of food pictures. We also informed that they would remain in the laboratory for 30 min at the end of the experiment to eat the food they obtained during the fMRI task. The task consisted of two parts: a pre-scan rating task that provided us with a measure of the baseline value for food, and an in-scan bidding and regulation task that measured the food value under the influence of regulation.
During the pre-scan rating task, subjects were shown 150 pictures of different snack food items (e.g., cake, chips, and candy) and rated, at their own pace, how much they would like to eat them using a four-point scale (1, “Don’t want it at all”; 4, “Want it a lot”). Our set of pictures was adapted to the Portuguese context of food.
Afterward, subjects received instructions for the in-scan bidding and regulation task (Figure 1). The 150 snack food pictures were shown again, separated into three trial conditions: indulge, distance, and natural. Each type of trial appeared 50 times, randomly interspersed over the scanning run. On each trial, before the food appeared, participants saw an abstract black-and-white symbol indicating the trial type (cue, 2 s). On indulge trials, subjects were instructed to try to increase their craving for the snack using any strategy they needed to. During distance trials, the instruction consisted on trying to decrease their craving. On natural trials, they had to allow thoughts and feelings to come naturally. Subjects had 4 s to look at the item and engage in the craving cognitive regulation task (hereinafter referred as cognitive regulation task). After the 4 s, subjects had 2 s to place a bid (0 €, 1 €, 2 €, or 3 €) for the right to eat that food at the end of the experiment. They were asked to treat each trial as if it were the only decision that counted. These bids allowed us to measure values expressed in behavior at the time of choice.

Figure 1. Timing and structure of a trial from the functional magnetic resonance task. ITI, intertrial interval.
At the end of the experiment, food was auctioned using an adapted version of the Becker-DeGroot-Marschak auction (Becker et al., 1964; Plassmann et al., 2007). We gave 3 € to each subject to spend during the auction over a maximum of three trials. Snacks and snacks prices were randomly selected by drawing a paper from a bag. The bids on those trials during the fMRI task determined whether subjects got to eat that food. Consider b the bid made by the subject during the fMRI task. During the auction, a random price a was drawn (0 €, 1 €, 2 €, and 3 € were chosen with equal probability). If b ≥ a, the participant got the item and spend a. If b < a, the subject did not get the item. The rules of the auction ensure the subjects’ best strategy to bid their true value for each food. This was explained and emphasized during the instruction period. For auction effects, omissions resulted in a bid of 0 €.
Each participant was scanned on a clinical approved 1.5 T Siemens Magnetom Avanto system (Siemens Medical Solutions, Germany) using a 12-channel receive-only head array coil. For the functional acquisition, a T2* weighted echo-planar imaging acquisition was acquired: 38 interleaved axial slices, repetition time 2750 ms, echo time 30 ms, field of view 224 mm × 224 mm, flip angle 90°, in-plane resolution 3.5 mm × 3.5 mm, slice thickness 3.5 mm, and between-slice gap 0.5 mm. To optimize the sensitivity in the orbitofrontal cortex, a tilted acquisition in an oblique orientation of 30° relative to the anterior-posterior commissure line was used. In total, 650 volumes were acquired during the task. The task stimulus was presented using the fully integrated fMRI system IFIS-SA (Invivo Corporation, United States) and the same system was used to record the subject key-press responses. One high-resolution T1-weighted Magnetization-Prepared Rapid Acquisition with Gradient Echo sequence, with 1 mm × 1 mm × 1 mm voxel size, repetition time 2.73 s, echo time 3.48 ms, flip angle 7°, field of view 234 mm × 234 mm, and 176 slices was acquired. This anatomical sequence was used to project the functional maps.
The functional scans from each participant were preprocessed using the Statistical Parametric Mapping (SPM) version 12 (Wellcome Department of Imaging Neuroscience, Institute of Neurology, United Kingdom) using MATLAB version R2018a (The MathWorks Inc.,United States). The preprocessing procedures included: slice-timing correction using the first slice as reference; realignment to the mean volume of the acquisition; nonlinear spatial normalization to Montreal Neurological Institute (MNI) standard space and resampling to 2 mm × 2 mm × 2 mm voxel size; spatial smoothing with a 8 mm full-width at half-maximum Gaussian kernel; high pass temporal filtering at 128 s. Participants with more than 3 mm of movement (1 voxel) were excluded (n = 0).
For the first-level analysis, one general linear model (GLM) was computed per participant. For this GLM, the regressors of interest included: the type of cognitive regulation trial (1 – distance, 2 – natural, and 3 – indulge) and the corresponding bid (4 – bids after distance trials, 5 – bids after natural trials, and 6 – bids after indulge trials). The bid regressors were parametrically modulated by the bid value (0, 1, 2, and 3 €), the pre-rating score before the task (1 to 4), and the reaction time. Additional regressors included: 7 – the cue; 8 – the interstimulus interval; 9 – the omission bids; 10 – 16 the motion parameters estimated during the realignment step. The onset and duration of the regressors were defined accordingly to the stimulus represented in Figure 1 with a boxcar function and the regressors were convolved with the canonical hemodynamic response function.
At the group level (second-level analysis), a random-effects analysis was performed using four different mixed-design ANOVA models: (1) represented the cognitive regulation during the task (enabled comparisons in average activation for each regulation trial between and within groups); (2) concerned the bidding/valuation during the task modulated by the bid value; (3) concerned the bidding/valuation during the task modulated by the pre-rating score; (4) concerned the bidding/valuation during the task modulated by the reaction time. Models (2) – (4) were used to test if food valuation was different between groups after distinct regulation trials. For all models, the group (stress vs. control) was introduced as the between-subject factor and each trial during cognitive regulation (distance vs. natural vs. indulge) as the within-subject factor. Age and education were used as covariates for all models. All models were implemented with the GLMFlex toolbox1 which uses partitioned error terms for within-group and between-group comparisons, enabling the estimation of all the effects of interest with a single model.
Results were considered statistically significant after correcting for multiple comparisons using cluster correction (minimum cluster size of 90 voxels). The minimum cluster size was determined with 3DClustSim (AFNI version 17.0.13; National Institute of Mental Health)2. This program determines a minimum cluster size with Monte Carlo Simulation to achieve a corrected significance of p < 0.05 with an initial voxel-wise threshold of p < 0.001. The Automated Anatomical Labeling plugin for SPM was used to classify the brain regions.
The stress group revealed higher levels of perceived stress (mean ± standard deviation 15.07 ± 5.23) than the control group (8.64 ± 5.27) as assessed by PSS-10 [t(27) = 3.30, p = 0.003, effect size d = 1.27]. No statistically significant differences were found for BAI (U = 117.50, p = 0.591) and BDI (U = 134.00, p = 0.217) between groups.
The ACTH levels before the fMRI session were similar between the two groups (U = 81.50, p = 0.310).
Cortisol serum levels were not statistically significantly different between groups [group F(1,27) = 0.45, p = 0.509] nor within group before and after the fMRI session [group × time F(1,27) = 1.00 × 10–3, p = 0.971]. However, cortisol levels decreased in both groups after the task [time F(1,27) = 10.08, p = 0.004, effect size χ2 = 0.27].
Glucose serum levels were not statistically significantly different between groups [group F(1,27) = 0.40, p = 0.531] and the pre and post-measurement were similar within groups [group × time F(1,27) = 0.18, p = 0.672]. However, glucose levels decreased in both groups after the task [time F(1,27) = 8.44, p = 0.007, χ2 = 0.24].
Insulin serum levels were not statistically significantly different between groups [group F(1,27) = 0.42, p = 0.522] and the pre and post-measurement were similar within groups [group × time F(1,27) = 3.68, p = 0.066]. However, insulin levels decreased in both groups after the task [time F(1,27) = 9.21, p = 0.005, χ2 = 0.25].
Given the differences in age and education between groups, we used these variables as covariates when analyzing behavioral parameters.
We analyzed the reaction time between and within groups during the different regulation trials (distance, natural, and indulge). We found an interaction effect between the group and the reaction time across the different regulation conditions [group × condition F(2,50) = 4.00, p = 0.024, χ2 = 0.14; Table 1 represents the results for all between and within group factors and covariate effects]. Post hoc tests with repeated measures ANOVA demonstrated statistically significant reaction time differences within the control group [F(1.42,18.43) = 7.06, p = 0.010, χ2 = 0.35, Greenhouse-Geisser correction for non-sphericity] and within the stress group [F(1.34,18.82) = 4.72, p = 0.033, χ2 = 0.25, Greenhouse-Geisser correction for non-sphericity]. Paired t-tests with Bonferroni correction showed that the reaction time for natural trials was shorter than for distance [t(13) = 4.82, p = 0.001, d = 2.67] and indulge trials [t(13) = 3.07, p = 0.027, d = 1.70], and distance and indulge trials presented similar reaction times [t(13) = 1.00, p = 1.000] in the control group. However, we did not find significant statistical differences in the stress group during post-hoc analysis [1.00 ≤ t(14) ≤ 2.36, 0.099 ≤ p ≤ 1.000] (Figure 2).
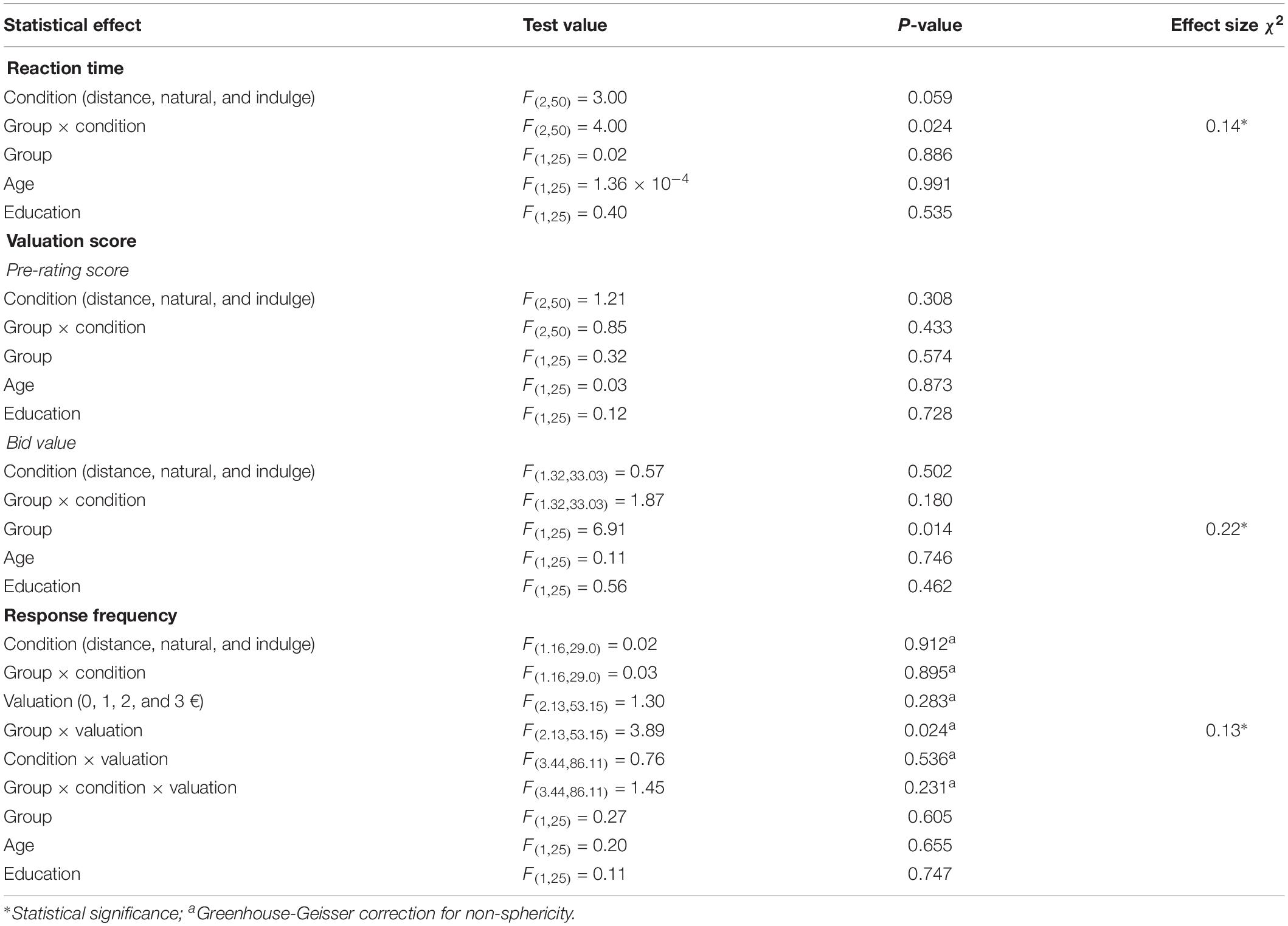
Table 1. Results for statistical tests on behavioral variables associated with the functional magnetic resonance imaging task: reaction time, valuation score, and response frequency.
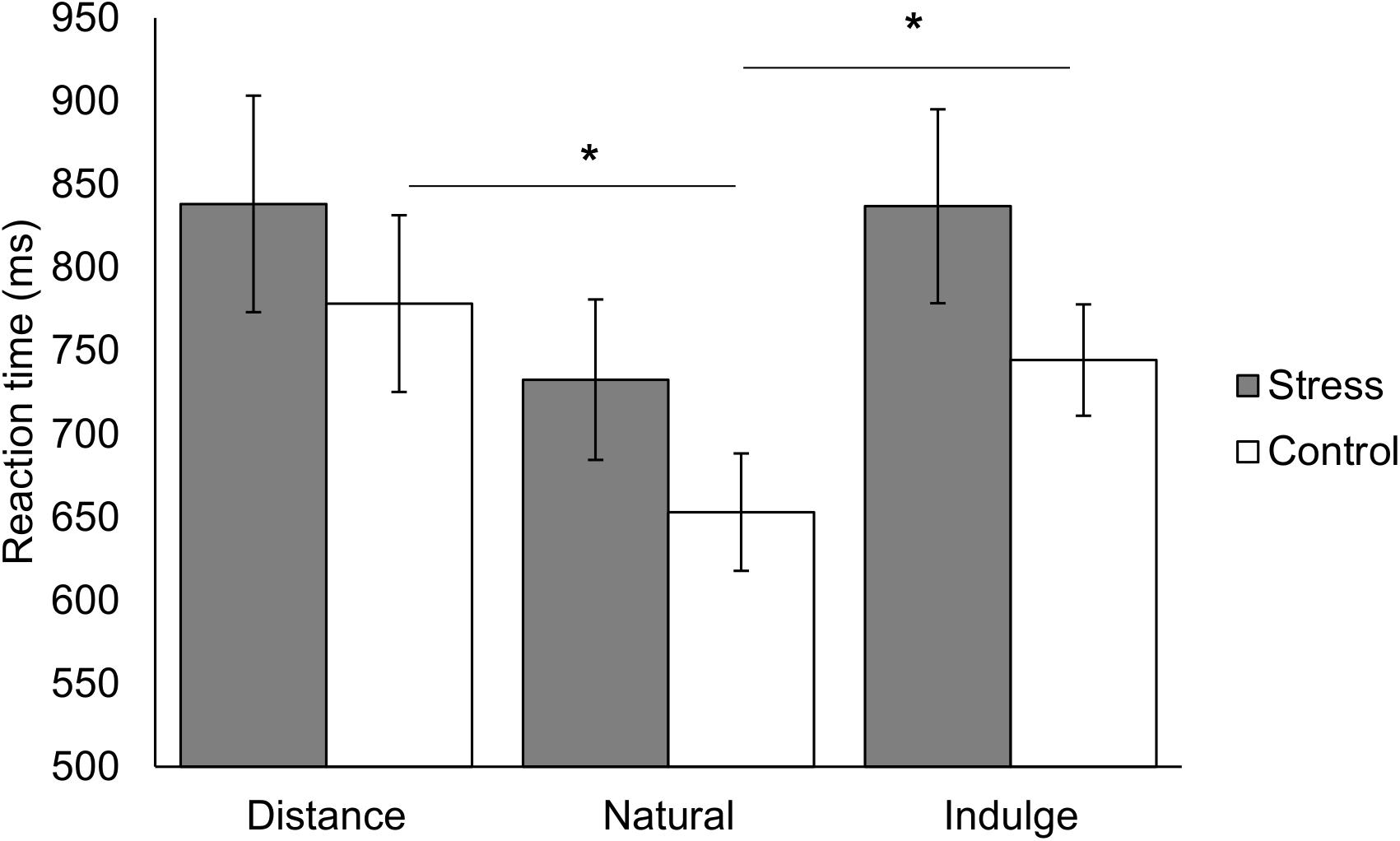
Figure 2. Representation of the reaction time for the different regulation conditions (natural, distance, and indulge) for the stress and control group. The reaction time for natural trials was shorter than for distance [t(13) = 4.82, p = 0.001, d = 2.67] and indulge trials [t(13) = 3.07, p = 0.027, d = 1.70] in the control group but no statistically significant differences occurred in the stress group. The black star represents statistically significant differences. The main bars represent the mean values and the error bars represent the standard error.
Taking into account that different instructions were given during the pre-rating (how much the participants want the food) and the bidding (how much the participants want to pay for the food), we separately analyzed differences between groups in the valuation score across the regulation conditions (distance, natural, and indulge) for pre and post-regulation scores (Table 1 represents the results for all between and within group factors and covariate effects). During pre-rating, we did not find statistically significant differences between groups or within group in terms of food valuation across the conditions. Moreover, the valuation score varied similarly among the conditions for both groups. However, during bidding, we found differences between groups [group F(1,25) = 6.91, p = 0.014, χ2 = 0.22] but not within group in terms of food valuation across the conditions. Moreover, the valuation score varied similarly among the conditions for both groups. The stress group had lower valuation scores (1.06 ± 0.36 €) during bidding in comparison to the control group (1.50 ± 0.36 €) (Figure 3).
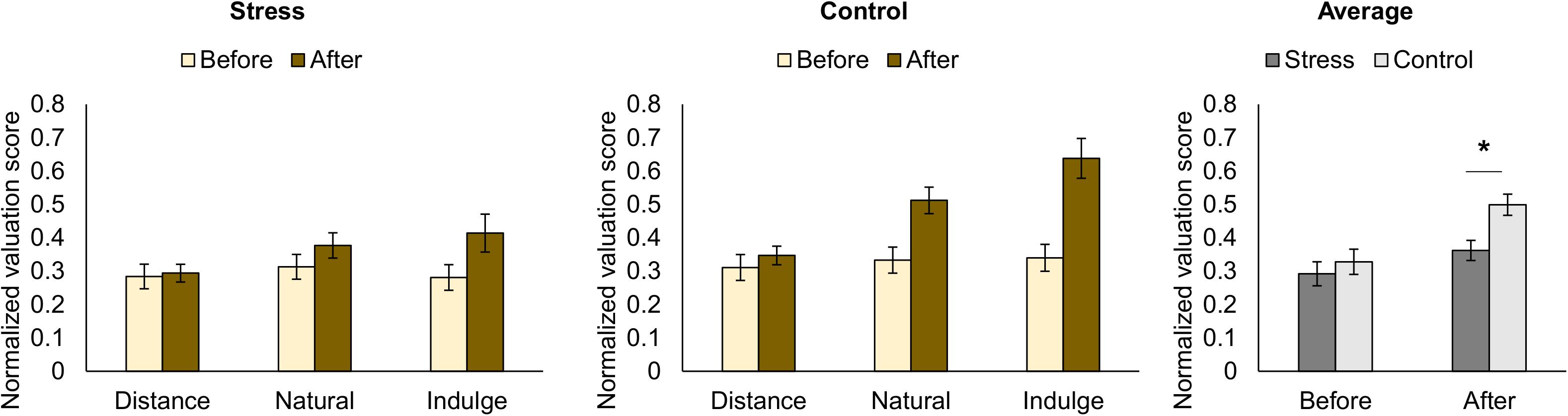
Figure 3. Representation of the normalized rating scores for food pictures for each trial condition (natural, distance, and indulge) before performing the functional magnetic resonance task (before cognitive regulation) and during the functional magnetic resonance task (after cognitive regulation) for the stress and control groups (the normalized scores represent the ratio between given score and maximum score). Before cognitive regulation, we did not find statistically significant differences between groups or within group in terms of food valuation across the conditions. After cognitive regulation, the stress group had lower average valuation scores in comparison to the control group [group F(1,25) = 6.91, p = 0.014, χ2 = 0.22]. The black star represents statistically significant differences. The main bars represent the mean values and the error bars represent the standard error.
Moreover, we also studied differences between groups in the number of responses for each bidding value after each cognitive regulation trial inside the scanner. We found a significant interaction effect between the group and the bid value [group × valuation F(2.13,53.15) = 3.89, p = 0.024, χ2 = 0.13, Greenhouse-Geisser correction for non-sphericity; Table 1 represents the results for all between and within group factors and covariate effects]. Post hoc tests with repeated measures ANOVA demonstrated that the control [F(3,39) = 9.61, p = 7.000 × 10–5, χ2 = 0.42] and the stress group [F(2.03,28.43) = 9.04, p = 0.001, χ2 = 0.39, Greenhouse-Geisser correction for non-sphericity] had a different number of responses across the bid values. Paired t-tests with Bonferroni correction demonstrated that on average the stress participants bided more often 0 [t(14) = 3.93, p = 0.009, d = 1.78] and 1 € [t(14) = 3.38, p = 0.027, d = 1.81] than 3 €, while control subjects bided more times 1 [t(13) = 3.94, p = 0.010, d = 2.18] and 2 € [t(13) = 5.91, p = 3.090 × 10–4, d = 3.28] than 3 € (Figure 4).
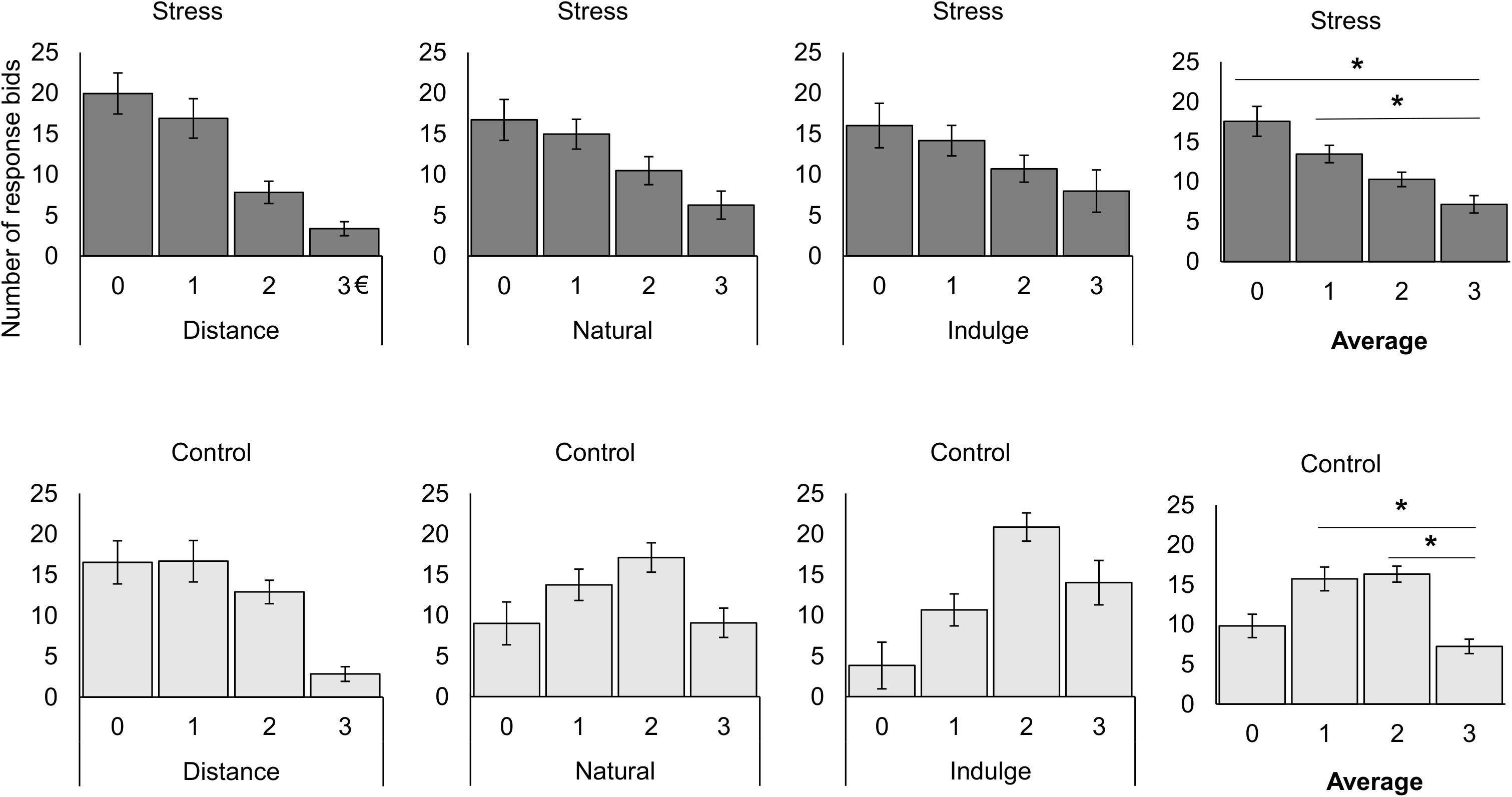
Figure 4. Representation of the number of responses for the stress and control groups during bidding for each cognitive regulation trial and for the average of all trials. On average, the stress participants bided more often 0 [t(14) = 3.93, p = 0.009, d = 1.78] and 1 € [t(14) = 3.38, p = 0.027, d = 1.81] than 3 €, while control subjects bided more times 1 [t(13) = 3.94, p = 0.010, d = 2.18] and 2 € [t(13) = 5.91, p = 3.090 × 10–4, d = 3.28] than 3 €. The black star represents statistically significant differences. The main bars represent the mean values and the error bars represent the standard error.
We tested for differences in blood-oxygen-level-dependent responses between stress and control groups during each cognitive regulation period/trial (natural, indulge, and distance) – model (1). No statistically significant brain regions were identified for overall differences between groups (main effect of group). When looking at the interaction effect between trial condition and group, there were also no statistically significant effects. Nonetheless, we found a main effect of the cognitive regulation condition in the left hemisphere in the superior (Brodmann area 22) and middle temporal gyrus (Brodmann area 21), the rolandic operculum, and the precentral gyrus (Brodmann area 6) [7.87 ≤ F(2,54) ≤ 13.97, p ≤ 0.001, 99 voxels, Montreal Neurological Institute peak voxel coordinates -60 -6 -4). Post hoc paired t-tests with Bonferroni correction demonstrated that the distance and indulge trials elicited lower activity than natural trials and that distance trials lead to higher responses than indulge trials in these regions [distance vs. natural t(28) = 2.97, p = 0.018, d = 1.12; distance vs. indulge t(28) = 2.68, p = 0.036, d = 1.01; natural vs. indulge t(28) = 4.97, p = 9 × 10–5, d = 1.88] (Figure 5).
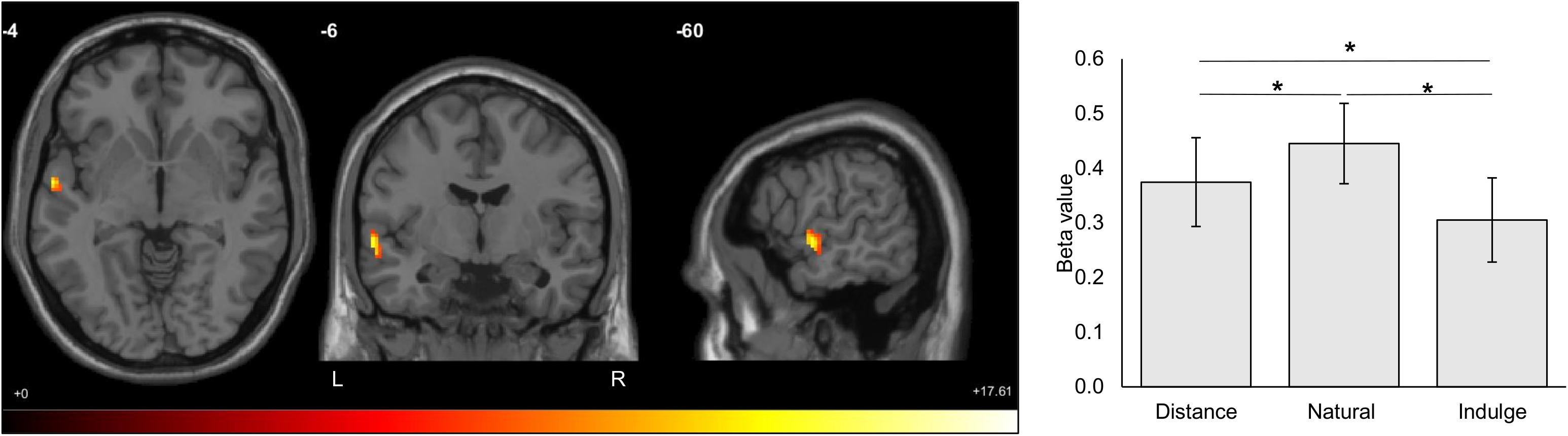
Figure 5. Statistically significant brain regions resulting from the main effect of the cognitive regulation condition (natural, indulge, and distance) for both groups (cluster correction for multiple comparisons, minimum voxel size of 90, p < 0.001, F(2,54) value between 7.87 and 13.97 represented by the colored bar). The distance and indulge trials elicited lower activity than natural trials, and the distance trials lead to higher responses than indulge trials in these regions (distance vs. natural t(28) = 2.97, p = 0.018, d = 1.12; distance vs. indulge t(28) = 2.68, p = 0.036, d = 1.01; natural vs. indulge t(28) = 4.97, p = 9 × 10– 5, d = 1.88). These regions include the superior (Brodmann area 22) and middle temporal gyrus (Brodmann area 21), the rolandic operculum, and the precentral gyrus (Brodmann area 6). The numbers above the slices represent the Montreal Neurological Institute peak voxel coordinates. The black star represents statistically significant differences. The main bars represent the mean values and the error bars represent the standard error. L, left; R, Right.
With the models (2) – (4), we tested if food valuation/bidding behavior was associated with different brain activation between groups after each regulation condition, with parametric modulation by bid value (model 2), pre-rating score (model 3), and reaction time (model 4). No statistically significant regions were identified for overall differences between groups during bidding (main effect of group) for the models (2) – (4). Additionally, no statistically significant active regions were found for interaction effects of group and the bids after each category of cognitive regulation (group × cognitive regulation condition), and the main effect of the condition was also not statistically significant for the models (2) – (4).
Given that our fMRI task was adapted from Hutcherson et al. (2012), here we compared our main results with these authors’ significant findings to study the task validity. Since we observed behavioral differences between the control and stress groups, we assessed the validity of the task only with the control group.
As observed by Hutcherson et al. (2012), we also saw that the control group took longer while bidding after distance [t(13) = 4.82, p = 0.001, d = 2.67, with Bonferroni correction] and indulge trials [t(13) = 3.07, p = 0.027, d = 1.70, with Bonferroni correction] than natural trials [group × condition F(1.42,18.43) = 7.06, p = 0.010, χ2 = 0.35, Greenhouse-Geisser correction for non-sphericity] (Figure 2). Moreover, the distance trials were also associated with the longest reaction time (778.30 ± 198.70 ms), followed by indulge (744.38 ± 125.21 ms), and natural (652.98 ± 131.97 ms) trials. These reaction times values are consistent with the previous study.
Concerning the bid value, similarly to Hutcherson et al. (2012), we observed a main effect of the cognitive regulation condition (distance, natural, and indulge) in controls [F(1.23,16.05) = 16.88, p = 4.650 × 10–4, χ2 = 0.56, Greenhouse-Geisser correction for non-sphericity]. The control participants bided higher on indulge [1.74 ± 0.43 €; t(13) = 2.89, p = 0.038, d = 1.60, with Bonferroni correction] and lower on distance [1.04 ± 0.28 €; t(13) = 4.22, p = 0.003, d = 2.34, with Bonferroni correction] compared to natural trials (1.50 ± 0.23 €). Bids after distance and indulge trials were also statistically significantly different [t(13) = 4.22, p = 0.003, d = 2.34, with Bonferroni correction] (Figure 3). The bid values in our study (0, 1, 2, and 3 €) were distinct from the original study ($ 0.0, 0.5, 1.0, 1.5, 2.0, and 2.5), thus we could not compare the average bid values after each condition trial.
For neuroimaging data, we computed the contrasts among the cognitive regulation trials in the control group to compare our results with the original study: Distance > Natural, Natural > Distance, Indulge > Natural, Natural > Indulge, Distance > Indulge, and Indulge > Distance. We applied cluster correction for multiple comparisons (90 voxels as described in the section “Materials and Methods”). We found statistically significant results only for the contrasts Distance > Natural and Indulge > Natural. These results are in agreement with Hutcherson et al. (2012)’s findings if the same minimum cluster size is considered. For the contrast Distance > Natural, similarly to the original work, we also found statistically significant activation in temporal and posterior parietal regions (Supplementary Table S1). However, results did not show statistically significant activity in medial and ventrolateral prefrontal regions. For the contrast Indulge > Natural, we found statistically significant responses in the anterior cingulate cortex, the ventral, medial, and superior prefrontal cortex, temporal and parietal regions, and the supplementary motor area (Supplementary Table S1). Thus, our results are concordant with these authors’ previous findings.
We studied how chronic stress influences decision-making on food valuation after cognitive regulation (increasing/indulge or decreasing/distance food craving) in medical students. Behavioral, biochemical and neuroimaging analysis were performed to address this question. We found that stressed participants present decreased food valuation scores. This result was reinforced by a higher number of responses for the lowest bid values for food in the stress group. The biochemical analysis (serum levels of insulin, cortisol, and glucose) did not show statistically significant differences between the control and stress group. The neuroimaging results did not demonstrate statistically significant differently activated brain regions between the stressed and control participants during cognitive regulation of craving and decision-making/bidding.
Although the acute stress response is generally beneficial, i.e., promotes adaptation to stressful stimulus, prolonged activation of the stress response produces deleterious effects on the body and brain, affecting cognitive processes such as decision-making (Mcewen, 2004; McEwen and Gianaros, 2011; Sousa, 2016). One of the main findings of the present study is that stressed individuals presented lower scores during food valuation, in contrast with our initial hypothesis. This may translate a blunted hedonic capacity or reward sensitivity (Berenbaum and Connelly, 1993; Porcelli et al., 2012; Bryce and Floresco, 2016; Porcelli and Delgado, 2017; Uy and Galván, 2017), as anhedonia has been associated with higher perceived stress scale scores (Pizzagalli et al., 2007) and stress causes changes in regions related to hedonic/rewarding behavior such as the amygdala, orbitofrontal cortex, vmPFC, and ventral and dorsal striatum (Gorwood, 2008; Porcelli et al., 2012; Bessa et al., 2013; Stark et al., 2015). Moreover, the distribution of the number of responses was higher for lower bids in the stress group, i.e., stressed subjects seemed less prone to place high bids for food. A previous work including a food-related task discovered decreased reward sensitivity associated with alterations in the putamen activity after acute stress induction (Born et al., 2010). Another report pointed out that acute stress does not potentiate craving after stimulation with food pictures (Stojek et al., 2015). Moreover, animal research indicates that acute stress reduces the motivation to work for food rewards (Bryce and Floresco, 2016). Other studies have also shown that acute and chronic stress mitigate brain responses to food stimuli in reward pathways (Wierenga et al., 2018). These results support the idea that stress participants have a reduced valuation attributed to food rewards. However, other studies have shown increased sensitivity to high-caloric food rewards in stressed individuals (Razzoli et al., 2017; Ans et al., 2018; Berg Schmidt et al., 2018; Ferrer-Cascales et al., 2018). Reward processing might be different when participants are stimulated with food pictures or real food. Moreover, the inclusion of chronic or acute stress models might also account for different results regarding reward sensitivity (Porcelli and Delgado, 2017). However, our results need confirmation with larger sample sizes.
While bidding, the stressed subjects did not present differential brain activity when compared to control subjects, despite the behavioral differences in the valuation score. We were expecting that poor cognitive self-control reflected in reduced prefrontal activation (Hare et al., 2009, 2011; Kober et al., 2010; Hutcherson et al., 2012) would lead to higher responses in striatal and amygdalar regions associated with increased reward sensitivity (Louis et al., 2009; Tryon et al., 2013; Maier et al., 2015; Neseliler et al., 2017). Other studies have found controversial results demonstrating that reduced striatal activity was associated with high levels of stress and increased food craving (Hommer et al., 2013). However, we did not observe cognitive differences between the groups and reward sensitivity was decreased. Our sample size may have limited the statistical power of this analysis. Thus, further research should be conducted to understand the neural correlates of decision-making after cognitive regulation since our results are not conclusive.
Our neuroimaging results did not show brain activity differences between groups during cognitive regulation. According to previous studies, the vmPFC and dlPFC are regions responsible for cognitive regulation in this decision-making context (Hare et al., 2009, 2011; Kober et al., 2010; Hutcherson et al., 2012). Thus, the absence of changes in these regions between groups in our work might indicate that the processes for cognitive regulation of food craving are not affected by chronic stress, or that our specific model of chronic stress might not lead to changes in cognitive modulation of craving. However, previous works revealed that cognitive control is diminished under stress, leading to emotional and habitual-biased decision-making (Yu, 2016), and increased reward sensitivity for food (Tryon et al., 2013; Maier et al., 2015; Neseliler et al., 2017). Our moderate sample size might have hindered putative differences between groups. Nonetheless, as shown in Figures 2, 3, stressed participants were able to modulate their responses, demonstrating an effective cognitive regulation, although the average food bidding score was lower than in controls. Both groups were capable of effectively using cognitive regulation to change the value placed on food during regulated trials. Moreover, both groups took longer times during bidding after regulated trials. Indeed, we found that activity in the superior and middle temporal gyrus, rolandic operculum, and precentral gyrus was differently modulated by trials with cognitive regulation of craving versus non-regulated trials in both groups. Previous authors provided evidence for a functional connection between the vmPFC and the precentral gyrus during food-related decisions, and for the correlation between food ratings and the response in the middle temporal gyrus (Kober et al., 2010; Hare et al., 2011; Hutcherson et al., 2012). Moreover, the temporal gyrus is also involved in food imagery (Hommer et al., 2013). Thus, the regulatory success does not seem to be affected by stress. During cognitive control tasks, attentional narrowing might occur after stimulation with negative pictures with threat and sadness-related content (van Steenbergen et al., 2011; Melcher et al., 2012; Papazacharias et al., 2015). Thus, the negative emotional state in the stress group (e.g., fear of falling the final exam) might have led to higher attentional focus during cognitive regulation that might compensate cognitive deficits associated with chronic stress. Nonetheless, our results need further validation with larger sample sizes to rule out a putative effect of chronic stress in cognitive regulation of craving.
Insulin, cortisol, and glucose levels are expected to decrease after fasting (Kirschbaum et al., 1997; Adam and Epel, 2007; Figlewicz, 2015; Tiedemann et al., 2017). However, peripheral concentrations of cortisol rise after stimulation with food images due to appetite enhancement, while insulin and glucose levels seem to be unaffected (Schmid et al., 2005; Schüssler et al., 2012; Kroemer et al., 2013). In our study, both groups presented a decreased in insulin, glucose, and cortisol levels after the fMRI task. Thus, the effects of fasting might have potentially surpassed the effects of stimulation with food pictures (Brede et al., 2017). Nonetheless, this hypothesis needs further testing. We were expecting increased craving in the stress group after a deficient cognitive regulation and increased reward sensitivity to food (Tryon et al., 2013; Maier et al., 2015; Neseliler et al., 2017). However, our results agree with the fact that we found reduced valuation of rewards in the absence of cognitive regulation alterations in the stress group, suggesting that overall craving was reduced. For controls, the instructions to differently modulate craving might have led to balanced changes in blood parameters after stimulation with food pictures. Thus, our results might derive from fasting since they occurred in both groups.
Importantly, our results are limited by the sample size. These results need to be replicated with larger samples to avoid false negative and positive conclusions. Moreover, the results might have been influenced by the unbalanced proportion of females and males per group, given that gender differences were found in decision-making under stress (Yu, 2016; Wemm and Wulfert, 2017). However, we focused on group differences and groups were matched for gender ratio.
Our results show that the capacity to perform cognitive regulation of craving is not impaired after prolonged stress. However, chronic stress reduces the value attributed to food rewards after craving modulation. Importantly, our conclusions are limited by the small sample size and need further validation with larger samples. These findings are relevant to guide subsequent studies on cognitive regulation of food-related decision-making for eating and obesity-associated disorders. Cognitive control techniques might be used to tackle decision-making impairments in these conditions (Louis et al., 2009; May et al., 2012).
The datasets generated for this study are available on request to the corresponding author.
This study was carried out in accordance with the Declaration of Helsinki. All subjects gave written informed consent. The protocol was approved by the Ethics Subcommittee for the Life and Health Sciences of University of Minho, Portugal, and by the Ethics Committee of the Hospital of Braga, Portugal.
SF, CV, PM, RM, AC, PMore, and CP-N acquired, analyzed, and interpreted the data and wrote the manuscript. NS and PMorg supervised the work, interpreted the data, and wrote and reviewed the manuscript. All authors participated in the design of the experiments and approved the manuscript.
This work was funded by the MyHealth project (DoIT – Desenvolvimento e Operacionalização da Investigação de Translação, Contract DoIT-13853) and the BIAL Foundation, Porto, Portugal (Grant No. PT/FB/BL-2016-206). This work was also funded by the FEDER funds, through the Competitiveness Factors Operational Programme (COMPETE), and by national funds, through the Foundation for Science and Technology (FCT), under the scope of the project POCI-01-0145-FEDER-007038. This manuscript has been developed under the scope of the project NORTE-01-0145-FEDER-000013, supported by the Northern Portugal Regional Operational Programme (NORTE 2020), under the Portugal 2020 Partnership Agreement, through the European Regional Development Fund (FEDER). PaM was supported by a fellowship of the project “SwitchBox” (Contract HEALTH-F2-2010-259772). SF was supported by a combined Ph.D. scholarship from the FCT and the company iCognitus4ALL – IT Solutions, Lda (PD/BDE/127839/2016). RM was supported by the combined FCT and Enlightenment, Lda fellowship (PDE/BDE/113604/2015) from the PhDiHES program.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnins.2019.00724/full#supplementary-material
ACTH, adrenocorticotropic hormone; BAI, Beck Anxiety Inventory; BDI, Beck Depression Inventory; dlPFC, dorsolateral prefrontal cortex; fMRI, functional magnetic resonance imaging; GLM, general linear model; MNI, Montreal Neurological Institute; PSS-10, 10-items Perceived Stress Scale; vmPFC, ventromedial prefrontal cortex.
Adam, T. C., and Epel, E. S. (2007). Stress, eating and the reward system. Physiol. Behav. 91, 449–458. doi: 10.1016/j.physbeh.2007.04.011
Ans, A. H., Anjum, I., Satija, V., Inayat, A., Asghar, Z., Akram, I., et al. (2018). Neurohormonal regulation of appetite and its relationship with stress: a mini literature review. Cureus 10:e3032. doi: 10.7759/cureus.3032
Arnsten, A. F. T. (2015). Stress weakens prefrontal networks: molecular insults to higher cognition. Nat. Neurosci. 18, 1376–1385. doi: 10.1038/nn.4087
Balleine, B. (2005). Neural bases of food-seeking: affect, arousal and reward in corticostriatolimbic circuits?. Physiol. Behav. 86, 717–730. doi: 10.1016/j.physbeh.2005.08.061
Basten, U., Biele, G., Heekeren, H. R., and Fiebach, C. J. (2010). How the brain integrates costs and benefits during decision making. Proc. Natl. Acad. Sci. U.S.A. 107, 21767–21772. doi: 10.1073/pnas.0908104107
Beck, A. T., Epstein, N., Brown, G., and Steer, R. A. (1988). An inventory for measuring clinical anxiety: psychometric properties. J. Consult. Clin. Psychol. 56, 893–897. doi: 10.1037/0022-006X.56.6.893
Beck, A. T., Steer, R. A., Ball, R., and Ranieri, W. (1996). Comparison of beck depression inventories -IA and -II in psychiatric outpatients. J. Pers. Assess. 67, 588–597. doi: 10.1207/s15327752jpa6703_13
Becker, G. M., Degroot, M. H., and Marschak, J. (1964). Measuring utility by a single-response sequential method. Behav. Sci. 9, 226–232. doi: 10.1002/bs.3830090304
Berenbaum, H., and Connelly, J. (1993). The effect of stress on hedonic capacity. J. Abnorm. Psychol. 102, 474–481. doi: 10.1037/0021-843X.102.3.474
Berg Schmidt, J., Johanneson Bertolt, C., Sjödin, A., Ackermann, F., Vibeke Schmedes, A., Lynge Thomsen, H., et al. (2018). Does stress affect food preferences? – a randomized controlled trial investigating the effect of examination stress on measures of food preferences and obesogenic behavior. Stress 21, 556–563. doi: 10.1080/10253890.2018.1494149
Bessa, J. M., Morais, M., Marques, F., Pinto, L., Palha, J. A., Almeida, O. F. X., et al. (2013). Stress-induced anhedonia is associated with hypertrophy of medium spiny neurons of the nucleus accumbens. Transl. Psychiatry 3, e266–e266. doi: 10.1038/tp.2013.39
Born, J. M., Lemmens, S. G. T., Rutters, F., Nieuwenhuizen, A. G., Formisano, E., Goebel, R., et al. (2010). Acute stress and food-related reward activation in the brain during food choice during eating in the absence of hunger. Int. J. Obes. 34, 172–181. doi: 10.1038/ijo.2009.221
Brede, S., Sputh, A., Hartmann, A.-C., Hallschmid, M., Lehnert, H., and Klement, J. (2017). Visual food cues decrease postprandial glucose concentrations in lean and obese men without affecting food intake and related endocrine parameters. Appetite 117, 255–262. doi: 10.1016/j.appet.2017.07.001
Bryce, C. A., and Floresco, S. B. (2016). Perturbations in effort-related decision-making driven by acute stress and corticotropin-releasing factor. Neuropsychopharmacology 41, 2147–2159. doi: 10.1038/npp.2016.15
Chen, Y., and Baram, T. Z. (2016). Toward understanding how early-life stress reprograms cognitive and emotional brain networks. Neuropsychopharmacology 41, 197–206. doi: 10.1038/npp.2015.181
Christopoulos, G. I., Tobler, P. N., Bossaerts, P., Dolan, R. J., and Schultz, W. (2009). Neural correlates of value. risk, and risk aversion contributing to decision making under risk. J. Neurosci. 29, 12574–12583. doi: 10.1523/JNEUROSCI.2614-09.2009
Cohen, S., Kamarck, T., and Mermelstein, R. (1983). A global measure of perceived stress. J. Health Soc. Behav. 24:385. doi: 10.2307/2136404
Delgado, M. R., Gillis, M. M., and Phelps, E. A. (2008). Regulating the expectation of reward via cognitive strategies. Nat. Neurosci. 11, 880–881. doi: 10.1038/nn.2141
Dias-Ferreira, E., Sousa, J. C., Melo, I., Morgado, P., Mesquita, A. R., Cerqueira, J. J., et al. (2009). Chronic stress causes frontostriatal reorganization and affects decision-making. Science 325, 621–625. doi: 10.1126/science.1171203
Fehr, E., and Camerer, C. F. (2007). Social neuroeconomics: the neural circuitry of social preferences. Trends Cogn. Sci. 11, 419–427. doi: 10.1016/j.tics.2007.09.002
Ferrer-Cascales, R., Albaladejo-Blázquez, N., Ruiz-Robledillo, N., Rubio-Aparicio, M., Laguna-Pérez, A., and Zaragoza-Martí, A. (2018). Low adherence to the mediterranean diet in isolated adolescents: the mediation effects of stress. Nutrients 10:1894. doi: 10.3390/nu10121894
Figlewicz, D. P. (2015). Modulation of food reward by endocrine and environmental factors. Psychosom. Med. 77, 664–670. doi: 10.1097/PSY.0000000000000146
Gold, J. I., and Shadlen, M. N. (2007). The neural basis of decision making. Annu. Rev. Neurosci. 30, 535–574. doi: 10.1146/annurev.neuro.29.051605.113038
Hare, T. A., Camerer, C. F., and Rangel, A. (2009). Self-control in decision-making involves modulation of the vmpfc valuation system. Science 324, 646–648. doi: 10.1126/science.1168450
Hare, T. A., Malmaud, J., and Rangel, A. (2011). Focusing attention on the health aspects of foods changes value signals in vmpfc and improves dietary choice. J. Neurosci. 31, 11077–11087. doi: 10.1523/JNEUROSCI.6383-10.2011
Hommer, R. E., Seo, D., Lacadie, C. M., Chaplin, T. M., Mayes, L. C., Sinha, R., et al. (2013). Neural correlates of stress and favorite-food cue exposure in adolescents: a functional magnetic resonance imaging study. Hum. Brain Mapp. 34, 2561–2573. doi: 10.1002/hbm.22089
Hutcherson, C. A., Plassmann, H., Gross, J. J., and Rangel, A. (2012). Cognitive regulation during decision making shifts behavioral control between ventromedial and dorsolateral prefrontal value systems. J. Neurosci. 32, 13543–13554. doi: 10.1523/JNEUROSCI.6387-11.2012
Kirschbaum, C., Bono, E. G., Rohleder, N., Gessner, C., Pirke, K. M., Salvador, A., et al. (1997). Effects of fasting and glucose load on free cortisol responses to stress and nicotine. J. Clin. Endocrinol. Metab. 82, 1101–1105. doi: 10.1210/jcem.82.4.3882
Kober, H., Mende-Siedlecki, P., Kross, E. F., Weber, J., Mischel, W., Hart, C. L., et al. (2010). Prefrontal-striatal pathway underlies cognitive regulation of craving. Proc. Natl. Acad. Sci. U.S.A. 107, 14811–14816. doi: 10.1073/pnas.1007779107
Kroemer, N. B., Krebs, L., Kobiella, A., Grimm, O., Vollstädt-Klein, S., Wolfensteller, U., et al. (2013). (Still) longing for food: insulin reactivity modulates response to food pictures. Hum. Brain Mapp. 34, 2367–2380. doi: 10.1002/hbm.22071
Louis, W. R., Chan, M. K.-H., and Greenbaum, S. (2009). Stress and the theory of planned behavior: understanding healthy and unhealthy eating intentions. J. Appl. Soc. Psychol. 39, 472–493. doi: 10.1111/j.1559-1816.2008.00447.x
Magalhães, R., Barrière, D. A., Novais, A., Marques, F., Marques, P., Cerqueira, J., et al. (2018). The dynamics of stress: a longitudinal MRI study of rat brain structure and connectome. Mol. Psychiatry 23, 1998–2006. doi: 10.1038/mp.2017.244
Maier, S. U., Makwana, A. B., and Hare, T. A. (2015). Acute stress impairs self-control in goal-directed choice by altering multiple functional connections within the brain’s decision circuits. Neuron 87, 621–631. doi: 10.1016/j.neuron.2015.07.005
May, J., Andrade, J., Kavanagh, D. J., and Hetherington, M. (2012). Elaborated intrusion theory: a cognitive-emotional theory of food craving. Curr. Obes. Rep. 1, 114–121. doi: 10.1007/s13679-012-0010-12
Mcewen, B. S. (2004). Protection and damage from acute and chronic stress: allostasis and allostatic overload and relevance to the pathophysiology of psychiatric disorders. Ann. N. Y. Acad. Sci. 1032, 1–7. doi: 10.1196/annals.1314.001
McEwen, B. S., and Gianaros, P. J. (2011). Stress- and allostasis-induced brain plasticity. Annu. Rev. Med. 62, 431–445. doi: 10.1146/annurev-med-052209-100430
McGuire, J. T., and Kable, J. W. (2012). Decision makers calibrate behavioral persistence on the basis of time-interval experience. Cognition 124, 216–226. doi: 10.1016/j.cognition.2012.03.008
Melcher, T., Obst, K., Mann, A., Paulus, C., and Gruber, O. (2012). Antagonistic modulatory influences of negative affect on cognitive control: Reduced and enhanced interference resolution capability after the induction of fear and sadness. Acta Psychol. 139, 507–514. doi: 10.1016/j.actpsy.2012.01.012
Minkley, N., and Kirchner, W. H. (2012). Influence of test tasks with different cognitive demands on salivary cortisol concentrations in school students. Int. J. Psychophysiol. 86, 245–250. doi: 10.1016/j.ijpsycho.2012.09.015
Morgado, P., Freitas, D., Bessa, J. M., Sousa, N., and Cerqueira, J. J. (2013). Perceived stress in obsessive–compulsive disorder is related with obsessive but not compulsive symptoms. Front. Psychiatry 4:21. doi: 10.3389/fpsyt.2013.00021
Morgado, P., Marques, F., Ribeiro, B., Leite-Almeida, H., Pêgo, J. M., Rodrigues, A. J., et al. (2015a). Stress induced risk-aversion is reverted by D2/D3 agonist in the rat. Eur. Neuropsychopharmacol. 25, 1744–1752. doi: 10.1016/j.euroneuro.2015.07.003
Morgado, P., Sousa, N., and Cerqueira, J. J. (2015b). The impact of stress in decision making in the context of uncertainty. J. Neurosci. Res. 93, 839–847. doi: 10.1002/jnr.23521
Morgado, P., Silva, M., Sousa, N., and Cerqueira, J. J. (2012). Stress transiently affects pavlovian-to-instrumental transfer. Front. Neurosci. 6:93. doi: 10.3389/fnins.2012.00093
Neseliler, S., Tannenbaum, B., Zacchia, M., Larcher, K., Coulter, K., Lamarche, M., et al. (2017). Academic stress and personality interact to increase the neural response to high-calorie food cues. Appetite 116, 306–314. doi: 10.1016/j.appet.2017.05.016
Ochsner, K. N., Ray, R. D., Cooper, J. C., Robertson, E. R., Chopra, S., Gabrieli, J. D. E., et al. (2004). For better or for worse: neural systems supporting the cognitive down- and up-regulation of negative emotion. Neuroimage 23, 483–499. doi: 10.1016/j.neuroimage.2004.06.030
Papazacharias, A., Taurisano, P., Fazio, L., Gelao, B., Di Giorgio, A., Lo Bianco, L., et al. (2015). Aversive emotional interference impacts behavior and prefronto-striatal activity during increasing attentional control. Front. Behav. Neurosci. 9:97. doi: 10.3389/fnbeh.2015.00097
Phillips, M. L., Drevets, W. C., Rauch, S. L., and Lane, R. (2003). Neurobiology of emotion perception II: implications for major psychiatric disorders. Biol. Psychiatry 54, 515–528. doi: 10.1016/S0006-3223(03)00171-179
Pizzagalli, D. A., Bogdan, R., Ratner, K. G., and Jahn, A. L. (2007). Increased perceived stress is associated with blunted hedonic capacity: Potential implications for depression research. Behav. Res. Ther. 45, 2742–2753. doi: 10.1016/j.brat.2007.07.013
Plassmann, H., O’Doherty, J., and Rangel, A. (2007). Orbitofrontal cortex encodes willingness to pay in everyday economic transactions. J. Neurosci. 27, 9984–9988. doi: 10.1523/JNEUROSCI.2131-07.2007
Platt, M. L., and Huettel, S. A. (2008). Risky business: the neuroeconomics of decision making under uncertainty. Nat. Neurosci. 11, 398–403. doi: 10.1038/nn2062
Porcelli, A. J., and Delgado, M. R. (2017). Stress and decision making: effects on valuation, learning, and risk-taking. Curr. Opin. Behav. Sci. 14, 33–39. doi: 10.1016/j.cobeha.2016.11.015
Porcelli, A. J., Lewis, A. H., and Delgado, M. R. (2012). Acute stress influences neural circuits of reward processing. Front. Neurosci. 6:157. doi: 10.3389/fnins.2012.00157
Rangel, A., Camerer, C., and Montague, P. R. (2008). A framework for studying the neurobiology of value-based decision making. Nat. Rev. Neurosci. 9, 545–556. doi: 10.1038/nrn2357
Razzoli, M., Pearson, C., Crow, S., and Bartolomucci, A. (2017). Stress, overeating, and obesity: insights from human studies and preclinical models. Neurosci. Biobehav. Rev. 76, 154–162. doi: 10.1016/j.neubiorev.2017.01.026
Sandi, C. (2013). Stress and cognition. Wiley Interdiscip. Rev. Cogn. Sci. 4, 245–261. doi: 10.1002/wcs.1222
Schmid, D. A., Held, K., Ising, M., Uhr, M., Weikel, J. C., and Steiger, A. (2005). Ghrelin stimulates appetite. imagination of food, gh, acth, and cortisol, but does not affect leptin in normal controls. Neuropsychopharmacology 30, 1187–1192. doi: 10.1038/sj.npp.1300670
Schüssler, P., Kluge, M., Yassouridis, A., Dresler, M., Uhr, M., and Steiger, A. (2012). Ghrelin levels increase after pictures showing food. Obesity 20, 1212–1217. doi: 10.1038/oby.2011.385
Soares, J. M., Sampaio, A., Ferreira, L. M., Santos, N. C., Marques, F., Palha, J. A., et al. (2012). Stress-induced changes in human decision-making are reversible. Transl. Psychiatry 2, e131–e131. doi: 10.1038/tp.2012.59
Sousa, N. (2016). The dynamics of the stress neuromatrix. Mol. Psychiatry 21, 302–312. doi: 10.1038/mp.2015.196
Starcke, K., and Brand, M. (2012). Decision making under stress: A selective review. Neurosci. Biobehav. Rev. 36, 1228–1248. doi: 10.1016/j.neubiorev.2012.02.003
Stark, E. A., Parsons, C. E., Van Hartevelt, T. J., Charquero-Ballester, M., McManners, H., Ehlers, A., et al. (2015). Post-traumatic stress influences the brain even in the absence of symptoms: a systematic, quantitative meta-analysis of neuroimaging studies. Neurosci. Biobehav. Rev. 56, 207–221. doi: 10.1016/j.neubiorev.2015.07.007
Stojek, M. K., Fischer, S., and MacKillop, J. (2015). Stress, cues, and eating behavior. Using drug addiction paradigms to understand motivation for food. Appetite 92, 252–260. doi: 10.1016/j.appet.2015.05.027
Tiedemann, L. J., Schmid, S. M., Hettel, J., Giesen, K., Francke, P., Büchel, C., et al. (2017). Central insulin modulates food valuation via mesolimbic pathways. Nat. Commun. 8:16052. doi: 10.1038/ncomms16052
Tryon, M. S., Carter, C. S., DeCant, R., and Laugero, K. D. (2013). Chronic stress exposure may affect the brain’s response to high calorie food cues and predispose to obesogenic eating habits. Physiol. Behav. 120, 233–242. doi: 10.1016/j.physbeh.2013.08.010
Uy, J. P., and Galván, A. (2017). Acute stress increases risky decisions and dampens prefrontal activation among adolescent boys. Neuroimage 146, 679–689. doi: 10.1016/j.neuroimage.2016.08.067
van Steenbergen, H., Band, G. P. H., and Hommel, B. (2011). Threat but not arousal narrows attention: evidence from pupil dilation and saccade control. Front. Psychol. 2:281. doi: 10.3389/fpsyg.2011.00281
Wager, T. D., Davidson, M. L., Hughes, B. L., Lindquist, M. A., and Ochsner, K. N. (2008). Prefrontal-subcortical pathways mediating successful emotion regulation. Neuron 59, 1037–1050. doi: 10.1016/j.neuron.2008.09.006
Wemm, S. E., and Wulfert, E. (2017). Effects of acute stress on decision making. Appl. Psychophysiol. Biofeedback 42, 1–12. doi: 10.1007/s10484-016-9347-9348
Wierenga, C. E., Lavender, J. M., and Hays, C. C. (2018). The potential of calibrated fMRI in the understanding of stress in eating disorders. Neurobiol. Stress 9, 64–73. doi: 10.1016/j.ynstr.2018.08.006
Keywords: stress, decision-making, cognition, magnetic resonance imaging, fMRI, reward, human, food
Citation: Ferreira S, Veiga C, Moreira P, Magalhães R, Coelho A, Marques P, Portugal-Nunes C, Sousa N and Morgado P (2019) Reduced Hedonic Valuation of Rewards and Unaffected Cognitive Regulation in Chronic Stress. Front. Neurosci. 13:724. doi: 10.3389/fnins.2019.00724
Received: 15 February 2019; Accepted: 28 June 2019;
Published: 10 July 2019.
Edited by:
Ruolei Gu, Institute of Psychology (CAS), ChinaReviewed by:
Sunny Dutra, William James College, United StatesCopyright © 2019 Ferreira, Veiga, Moreira, Magalhães, Coelho, Marques, Portugal-Nunes, Sousa and Morgado. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pedro Morgado, cGVkcm9tb3JnYWRvQG1lZC51bWluaG8ucHQ=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.