 Sonia Méhault
Sonia Méhault Fabien Morandeau
Fabien Morandeau Robin Faillettaz
Robin Faillettaz Alexa Abangan
Alexa Abangan Dorothée Kopp
Dorothée Kopp
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Mar. Sci., 26 September 2022
Sec. Marine Fisheries, Aquaculture and Living Resources
Volume 9 - 2022 | https://doi.org/10.3389/fmars.2022.1009992
This article is part of the Research TopicInnovations in Fishing Technology Aimed at Achieving Sustainable FishingView all 23 articles
The French fishing industry is becoming increasingly environmentally aware and likely to adopt more sustainable fishing gears. As a result, fishers are showing a growing interest in sustainable fish pots. This experiment aimed to develop a fish pot concept specifically based on target species behavior in French coastal waters. First, the consultation led with fishers indicated the black seabream as the main species of interest. Then, the pot’s characteristics were defined to comply with fishing regulations. The conception process of the pot was led step-by-step by gradually testing a pot design appropriated to black seabream’s natural behavior. The approach and feeding behaviors of black seabream were described using underwater video combined to an ethogram. The experiment led to a prototype of fish pot, which provide fishers with an innovative fishing gear.
Active fishing gears are more and more pointed out as altering the seabed (Eigaard et al., 2016), generating bycatch (Bellido et al., 2011) and discarding unwanted catch (Morfin et al., 2017). Passive fishing gears are seen as less impacting (Suuronen et al., 2012), but studies also indicated impacts of gillnets or trammel nets notably on bycatch of birds or marine mammals (Northridge et al., 2017) and sea turtles (Lucchetti et al., 2017). Pots are traditionally used by many small-scale fisheries worldwide for catching crustaceans or fish. Though they may have adverse effects on the habitats and resources when they are used with a lack of management (Vadziutsina and Riera, 2020), this fishing technique has generally lower environmental impacts on the seabed, and generates fewer discard and bycatch than towed gears (Shester and Micheli, 2011; Suuronen et al., 2012; Kopp et al., 2020; Petetta et al., 2021). Finally, fish or invertebrates are mainly caught alive, maximizing the survival of unwanted catches (Purves et al., 2003), giving the fish high commercial value for fishers and high quality for consumers (Suuronen et al., 2012).
While fish pots are efficient in many parts of the world (Garrison et al., 2004; Marshak et al., 2008; Vadziutsina and Riera, 2020), they are not widely used in European commercial fisheries due to low catch efficiency. Thereby, many research programs have been implemented in North Atlantic waters to try increasing catchability of cod (Furevik and Løkkeborg, 1994; Jørgensen et al., 2017; Humborstad et al., 2018; Chladek et al., 2021), but in the Bay of Biscay, only low commercial valued finfish species such as conger (Conger conger) or common pout (Trisopterus luscus) were caught.
To understand the catching process of fish pots, underwater cameras are often used (Furevik et al., 2008; Anders et al., 2017b; Anders et al., 2017a; Folkins et al., 2021). Compared to traditional sampling techniques (e.g. scientific trawling, diving, or acoustics), underwater cameras minimize disturbance to species behavior and improve our ability to identify and count species, as well as analyze their behavior. This is particularly of interest in designing a pot. According to Stoner (2004), variation in fish behavior is a key factor affecting catch per unit effort. Thus, observing the behavior toward the bait, according to the shape, volume, number of chambers and color of the pot, as well as the number, position and design of entrances is crucial to understand better why a fish would enter and stay in the pot or not.
In the present study, a concertation with French artisanal fishers led to identify black seabream (Spondyliosoma cantharus) as an appropriate target species for fish pot: it is widely present in coastal waters, mainly from 10 to 100m depth (Pajuelo and Lorenzo, 1999; Collins and Mallinson, 2012) and its population is considered of least concern by the IUCN (Russel et al., 2014). According to the database of the French fisheries information system managed by Ifremer, 2392 tons of black seabream were landed by French fleet in 2020 (51% caught by demersal trawlers and 30% by pelagic trawlers). Artisanal fishers are willing to operate a diversification of their métiers to improve their capacity of adaptation to market and resource fluctuations (Prosperi et al., 2019; Pascual-Fernández et al., 2020). Black seabream is also targeted by recreational fishers (Pinder et al., 2017), who catch it with hook. The question underlying the experiment proposed in that study is: “if the black seabream can feed on a baited hook, why not on the same bait inside a pot?” We propose a step-by-step process to design a black seabream-specific pot. The methodological approach consisted in designing a pot gradually, making sure that while complexifying the gear, the target species can still enter and reach the bait. For each stage of the pot design, 2 configurations were compared: i) weighted vs floated design, ii) black vs transparent netting, iii) one vs two entrances, iv) rectangular-shaped vs triangular-shaped entrance. Underwater video cameras were used to assess the success of each configuration in terms of bait consumption and target species behavior. The process led to an operational pot prototype. The potential to transfer this methodology to target other species as well as the benefit of developing such innovative gear to diversify small scale fisheries are discussed.
The study was spread out in the Bay of Quiberon (47 °C53 N, 003 °C10 O, Bay of Biscay, East Atlantic, ICES Subarea VIIIa) over an area of about 900m2. Habitat was homogeneous all over the area with the seafloor made of sand, mud and coarse sediments. The water depth ranged from 7 m to 12 m depending on the tide hour and coefficient. The experiment took place from July to September 2019 when black seabream migrates to coastal area in summer time (Pawson, 1995).
Three observation devices were immersed each day. One device without any gear was used as control to validate the presence of fish in the study area (Figure 1, right panel). The two other devices included an experimental gear to perform comparison of fish attraction between 2 pot configurations (Figure 1, left panel). Since the deployment of the set of three devices was simultaneous, each configuration and control devices of a given comparison had the same number of replicates. Each device included one GoPro Hero 4 camera mounted on a metallic frame weighted with ballasts. This maintained a fixed distance of 2 m between the camera and the bait. The camera recorded with a 1920*1080p resolution at a frame rate of 25 fps on a 128-Gb micro-SD. It was kept in a waterproof housing designed for an extra USB battery pack of 15600 mAh and 5V allowing for more than 8 hours of video recording (limited by the micro-SD capacity). Each observation device was baited with 100 g of cockles (Cerastoderma edule) tethered on a wire. Cockles were used as bait because bivalves are part of Sparidae natural preys reported in the literature (Pita et al., 2002; Dulcic et al., 2006) and they are specifically used by recreational fishers to target black seabream.

Figure 1 Left: Experimental gear device. Right: Control device.
Four comparisons of gear configurations were carried out successively (Figure 2). The first assessed weighted versus floated gears (C1; Figure 2), hypothesizing that in a floated pot, the bait will always be aligned with the entrance regardless of tidal current direction (Furevik and Løkkeborg, 1994). Fish being attracted by odor plume (Løkkeborg, 1998), most of their approaches to the bait is likely to be up-stream. Furthermore, floating the pot avoids the catch of crustaceans such as red king crab (Paralithodes camtschaticus) and allows targeting fish species exclusively (Furevik et al., 2008). This characteristic supports the regional legislation, which prohibits the catch of crustacean while using fish pots (CRPMEM Bretagne, 2018). This experiment aims to comply with that legislation. The floated gear was thus linked up to the metal frame using a swivel 40cm above the sea floor, allowing for gyration, whereas the weighted gear was motionless attached to the metal frame. Inspired from the Canarian fish pot (Garcia-Mederos et al., 2015), the baseline frame of the experimental gear was circular (1.5m diameter and 0.6m high) to favor its rotation with the water current and avoid angular side that may prevent fish from locating the pot entrance easily. The bare metal frame was tested to make sure that it doesn’t prevent target fish from reaching the bait.
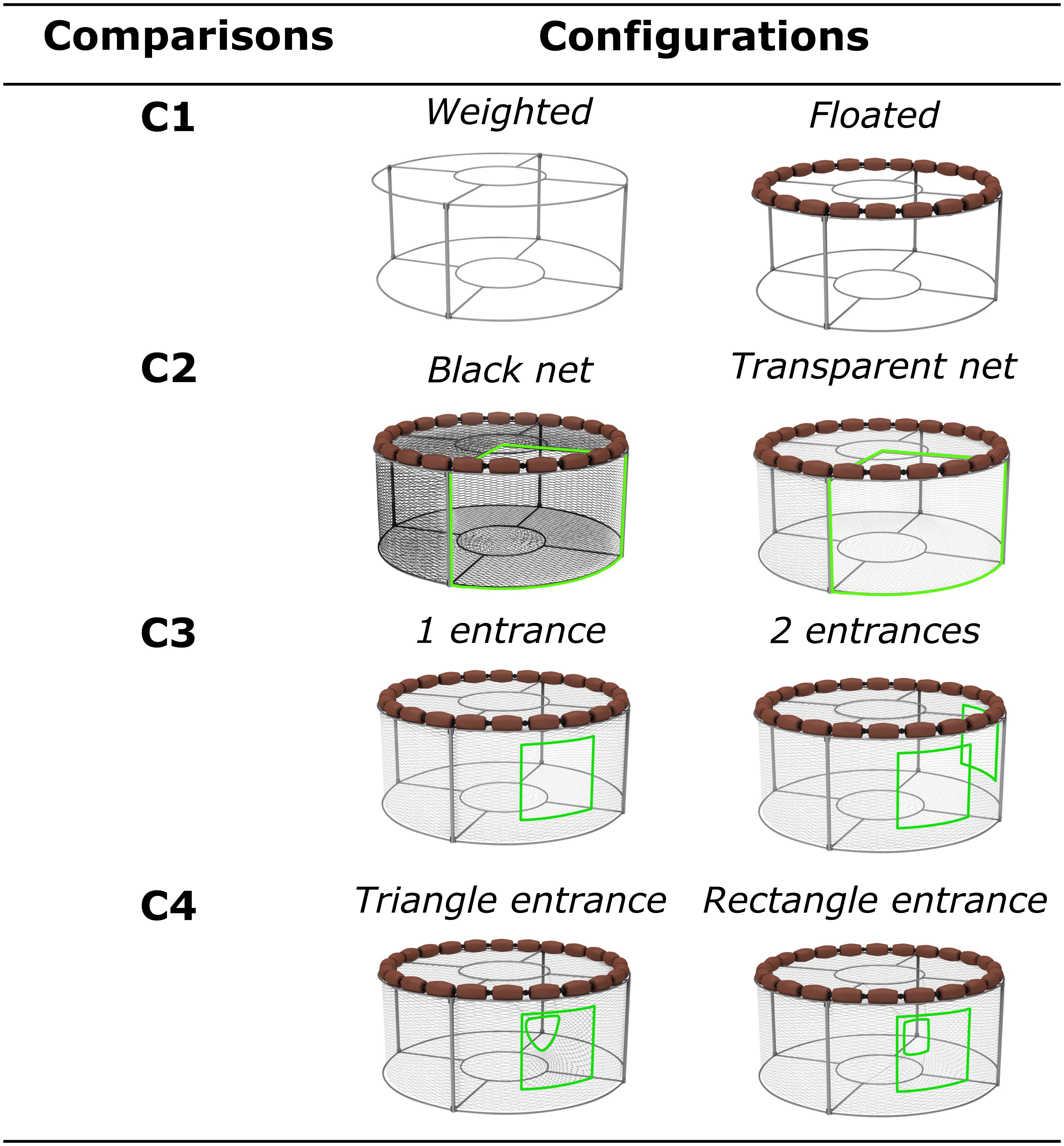
Figure 2 Configurations of experimental gear deployed to carry out each of the four comparisons.
The second comparison focuses on the color of netting (black versus transparent) (C2; Figure 2). The color of the gear net has proven to have an effect on fish species behavior (Balık and Çubuk, 2000; Orsay and Dartay, 2011). Black and transparent nets were chosen to create a strong contrast between gear configurations, with 3/4 of the vertical and top sides of each experimental gear covered by the net. Both nets were made of 50mm diamond meshes.
The third comparison evaluates the number of gear entrances (C3; Figure 2) that can affect the number of fish entering as well as the probability of fish escapement (Furevik and Løkkeborg, 1994; Jørgensen et al., 2017). For that comparison, both gear configurations were entirely covered by the net, except two gaps of 60*40cm for one configuration and one gap of 60*40cm for the other configuration.
The entrance design is also challenging as it should allow fish to enter the gear without letting it swim back to escape, such as demonstrated by Tran et al. (2020) with crustacean pots. Then, two types of entrances (triangular-shaped versus rectangular-shaped) were compared (C4; Figure 2). The two-entrance types have a funnel shape of 25cm depth. One was hanged with 4 wires designing a rectangular-shaped rectangle in the inner side. The second was made with only two hangings designing a triangular shape, which allowed slight motion of the entrance outline with the water flow in the inner side. The triangular-shaped entrance aimed to prevent fish from getting out of the experimental gear.
Each comparison aimed to assess which configuration was the most suitable to the natural behavior of the fish to keep reaching the bait.
The sampling took place during daytime, with the devices deployed in the morning to benefit from day light over the entire recording. To allow comparisons, three devices were deployed simultaneously (Figure 3): one for each configuration of the comparisons and 1 for the control leading to 24 hours of video recording for each sampling day. After each gear retrieval, the videos were quickly checked to identify which species consumed the bait, either partly or entirely. The indicator of success used for each configuration was whether or not the bait had been eaten by black seabream by the end of the 8-hour video recording. Fine video screening, which is more time-consuming, was performed afterwards to count black seabream and characterize their behavior.
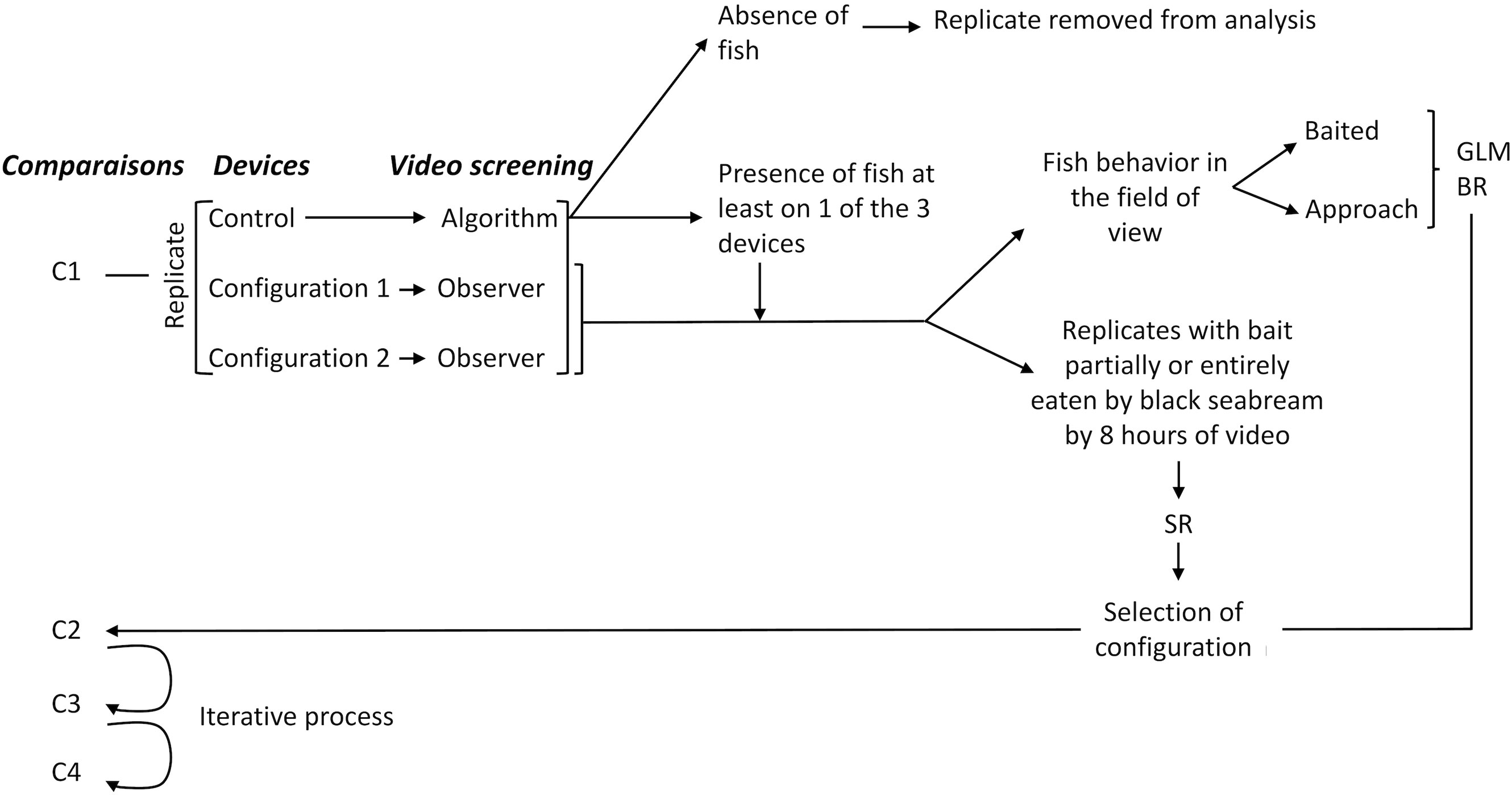
Figure 3 Experimental design of the step-by-step methodology (numbers of replicates and fish observed are presented in Table 1).
11 trials were carried out for the comparison C1, 4 for C2, 4 for C3 and 5 for C4. To process the video recorded with the control device without manually skimming their entire duration, a fully automated process was applied. Fishes around the control device were detected with a fine-tuned deep learning model (Cascade Faster R-CNN; Chen et al., 2019) trained on the videos of the experiment using the VIAME interface (Dawkins et al., 2017). VIAME is a deep learning platform where object detection and tracking models can automatically predict objects on underwater videos. This initial screening showed robust identification of the presence of fishes around the bait among sampled days. The presence of black seabream was only recorded as presence-absence since the automatic detection algorithm did not allow for considering a single individual after 20 seconds of disappearance from the screen as suggested by Anders et al. (2017a). A conservative threshold was further applied on the fish classification confidence (i.e. probability of correct classification ≥ 0.80) to focus on the rate of true positive classifications among videos (Faillettaz et al., 2016).
The immersion of the 21 sets with presence of fish produced 336 hours of underwater videos of experimental gear. Determining the attraction potential of the pot required the identification of fine-scale fish behavior, i.e. whether the fish approached or entered. This process could not be accurately performed by artificial intelligence algorithms since the floating pot moving with waves and water currents generated too many false positives in the background. The extraction of fish behavior information (rather than fish presence) was thus done manually by an observer for all recorded videos with an experimental pot.
Finally, when no fish was recorded on the control device, nor on none of the 2 experimental gear devices, the trial was considered as null and was discarded from the analysis. This situation occurred for 3 trials for C1 (probably because they took place too early before black seabream migrate to the study area). They were removed from the analysis since the absence of fish in the study area makes it impossible to conclude on gear performance.
Fish behavior was split into 2 categories. “Approach” was assigned when fish was in the field of view or heading towards the gear without entering the experimental gear. “Baited” was assigned when the fish entered the experimental gear. However, for C1, each individual had several options to enter the gear. Therefore, the swimming orientation of the baited fish was broken down into four levels which were “top” when the fish entered the gear by the top part, “side” when it entered by the side part, “bottom” when it entered by the bottom part and “entrance” when it entered by the pot entrance. Mann-Whitney test was used to determine the predominant orientation of baited fish when it enters the gear. That information was used to locate the pot entrance(s).
Juvenile fish are not targeted, therefore the small individuals that managed to enter through the meshes instead of through the entrance were classified as “approaching” individuals and not “baited”. Each single fish was followed from its entrance in the field of view of the camera until it disappeared. Black seabream could not be individually identified on the videos, therefore, according to Anders et al. (2017a) the assumption was made that an individual which disappeared for less than 20 seconds was recorded as the same individual, otherwise it was considered as another one. A unique ID number was assigned to each individual. For each of them, only one behavior was recorded: if a fish first approached and was baited afterwards, then it was recorded as “baited”. Video screening was performed until the entire consumption of the bait using BORIS 7.4.7 (Friard and Gamba, 2016).
The step-by-step methodology used to define pot design was based on two complementary indicators. First, a Success Ratio (SR) was computed for each configuration (Conf). It was computed by dividing the number of replicates with the bait eaten by black seabream by the end of the 8-hour video recording (nSuccess,Conf) by the total number of replicates with presence of fish for each configuration (nPresence,Conf).
After each gear retrieval, the videos were screened to validate that black seabream was responsible for the bait consumption. Only black seabream was observed to feed on the bait and no interaction between black seabream and any other species was observed, therefore only black seabream individuals were recorded. The SR indicator provided immediate information on fish presence inside the gear. Since black seabream is present in the study area for a limited period of time, the SR indicator was used to make a rapid decision on which pot configuration to test further: the configuration with the highest SR was kept to carry on with the subsequent comparison.
Second, the Baited Ratio (BR) was computed by dividing the number of fish baited (i.e. entered in the experimental gear) by the total number of fish observed in the field of view (i.e. either baited or approaching). Because of the low number of replicates per configuration, the BR indicator was computed by cumulating the number of fish across all replicates:
with BRConf the Baited Ratio for each configuration, nBaited,Conf the number of fish baited per replicate and configuration and nField,Conf the number of fish observed in the field of view per replicate and configuration. The BR indicator required meticulous video screening. It provides fine-scale information on fish counts and behavior but not immediate results and do not allow for a large number of replicates. BR was relevant to assess the capacity of the experimental gear to let enter the fish attracted. A Generalized Linear Model (GLM) with binomial distribution of the response (baited = 1 vs approach = 0) and log link function was fitted for each comparison to test for the effect of gear configuration on the probability that a fish is baited.
Among the 42 gear-videos analyzed, only 5 had the bait fully eaten by the end of the recording: 3 videos for C1 had the bait entirely eaten by 0.5, 2.5 and 6.1 hours of soaking time and 2 videos for C2 with the bait entirely eaten after 1.5 and 6.5 hours of soaking time. All the other videos with the bait not or partially eaten by the end of the 8 hours of recording were entirely screened. A total of 554 black seabreams were observed. On average, 16.6 (σ=9.9) individuals were recorded per deployment. The SR indicator showed that the floated, transparent, one-entrance and rectangular-shaped entrance were the most favorable configurations for black seabream predation on the bait (Table 1). The BR indicator showed a similar trend as SR except for C1. For C1, the two indicators provided an opposite trend, with BR showing a proportion of fish entering the weighted gear higher than the proportion of fish entering the floated gear. The GLM outputs confirmed the trends observed with BR, though not significantly for C2, C3 and C4 (p>0.5). For C1, GLM output indicated that the probability for a fish to be baited with a weighted gear is significantly higher than with the floated gear (p<0.001). Both configurations of C1 indicated that the frame without mesh did not repel target fish. The C1 experiment indicated that 70% of fish entered the gear to reach the bait from the side of the pot, which is significantly higher than the proportion of fish entering by the top or bottom part (Mann-Whitney, p< 0.02). According to that result, the pot entrance was positioned on the gear side.
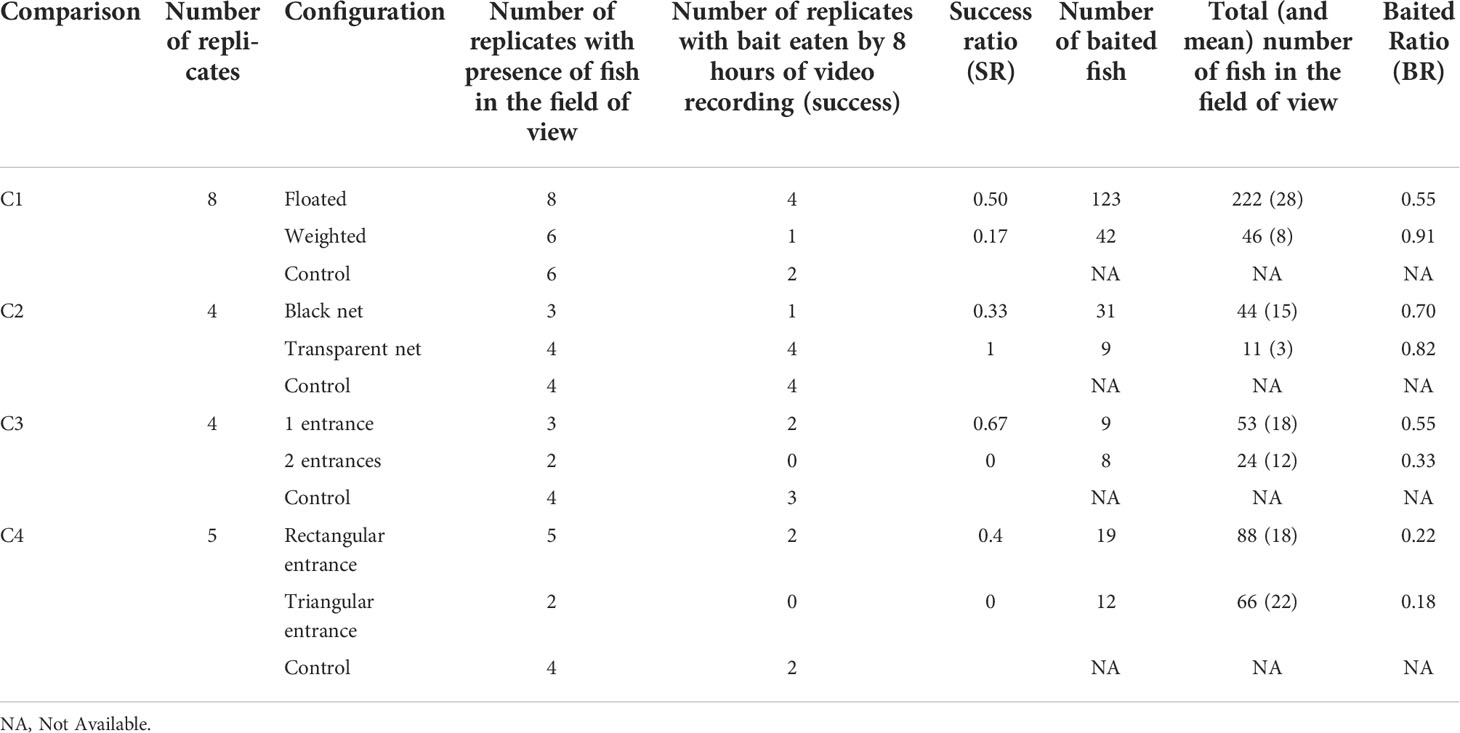
Table 1 Number of replicates, successes and baited ratios for each of the 4 comparisons.
Each comparison added a new element to the gear conception. The SR and BR indicated that the configurations selected don’t prevent fish from reaching the bait. However, the BR indicator decreased as the complexity of the experimental gear increased; except for the floated configuration of C1 which was relatively low (BR =0.55). The least BR difference was observed between the rectangular-shaped and triangular-shaped entrances. No fish were with caught with C4 configurations when the pots were hauled back, which indicates that individuals that were baited managed to escape.
The step-by-step methodology proposed to design a fish pot based on natural behavior of black seabream ended up with a floated circular pot prototype made of transparent net with one rectangular-shaped entrance. That design was based on the Success Ratio and Baited Ratio values. GLM outputs showed similar trends as BR, but larger samples size would be required to detect possible significant difference between configurations. Despite the opposing trends of SR and BR observed for the floated configuration and the higher probability of baited fish with the weighted gear (C1), the floated configuration was selected to carry on with the following comparisons (C2 to C4). Indeed, the gear set above the seafloor prevented crustaceans from reaching the bait (Furevik et al., 2008), which reduces the probability to catch them and allow to comply with the legislation (CRPMEM Bretagne, 2018). Also, the floating specificity of the gear let it orient itself with the water current, which allows its entrance to be aligned with the odor plume of the bait. Besides attracting the fish to the gear surrounding, the diffusion of the odor plume is likely to change fish swimming activity and turning behavior (Løkkeborg et al. (1995) and to increase the probability of fish entering the pot to reach the bait (Jørgensen et al., 2017).
The fish behavior in relation to the color of the net was mainly explored for trawl and set net gear (Wanner et al., 2010; Simon et al., 2020). In the case of fish pot, dark-colored net can simulate a shelter, which does not even require to bait the gear (Robichaud et al., 2000; Pravin et al., 2011). On the contrary, a transparent net reveals the bait visually from outside the gear and for Chladek et al. (2021) who tested 3 configurations of pot funnel entrance (i.e., transparent, white and green), the transparent funnels give the highest rate of cod entering the pot. The vision is one of the main senses used by fish to reach their prey (Novales Flamarique, 2019). It is known that fish swimming decision is impacted by the color and shape of the net (Glass et al., 1995). Therefore, transparent meshes are likely to facilitate the entrance of fish inside the gear. Transparent net of C2 also minimized the contrast with the bare experimental gear of C1, while converting it progressively into a pot.
The SR and BR were both higher with 1 entrance than with 2 entrances. It is known that 1 entrance improves the encounter rate, reduces the risk of escapement (Chladek et al., 2021) and therefore improves catch rate (Jørgensen et al., 2017). However, the present experiment did not account for the escapees, which tends to confirm the benefit of the floating gear: more than one entrance is not necessary if it is well aligned with the water current.
The shape of the entrance remains challenging. It should not impede the passage of the fish to enter the pot but it should prevent it from getting out. BR and SR were higher with the rectangular-shaped entrance, which indicates that this configuration does not prevent fish from entering. Since no fish were actually caught, none of the two entrance types could avoid fish escapement, therefore investigations could test more options of pot and non-return entrance designs.
On-going progress on automatic image analysis algorithms and artificial intelligence would facilitate video data treatment. Some development for trawl gear already contributed to reduce fishing gear impacts and increase their selectivity by providing rapid, accurate and standardized data on fish behavior in relation to gear (Robert et al., 2020). In parallel, the development of acoustic imaging would overcome dark and turbid conditions to describe fish behavior in relation to fishing gear on a longer period of time (Rose et al., 2005; Fujimori et al., 2018). It is especially relevant for species with diurnal feeding habit (eg. Sparus aurata (Pita et al., 2002)), which would approach the baited gear at a specific period of the day or night. This knowledge would optimize the soaking time and catch rates. Such advances in images analysis applied to pot gear and fish behavior would make the methodology proposed in this study easily transferable to other species and areas.
Further experiments on that pot selectivity, as well as an economical cost-benefit analysis and an ergonomic evaluation under commercial conditions would provide a better understanding of the pros and cons of this fishing gear. Developing species-specific pot gear would contribute to the diversification of small-scale fisheries (Prosperi et al., 2019; Pascual-Fernández et al., 2020). Most of the artisanal fishing vessels are already equipped to operate a variety of fishing gears, which would facilitate the use of fish pot. On the other hand, the adoption of this type of gear relies on the yield and market opportunities for fishers. Pots generate low discard rates compared to many other gear types (Petetta et al., 2021). Developing fish pots would help fishers to cope with new gear constraints due to the multiplication of maritime usages such as wind farms (Yates et al., 2015; Stelzenmüller et al., 2021) and marine protected areas (Cadiou et al., 2009).
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethical review and approval was not required for the animal study because fish were only observed with video cameras in their natural environment.
SM wrote the first draft of the manuscript. SM, FM, DK and JS contributed to conception and design of the study. AA and DK wrote sections of the manuscript. AC and FM performed the video analysis. SM and RF performed the analytical analysis. All authors contributed to manuscript revision, read, and approved the submitted version.
This work was part of the BAITFISH project supported by France Filière Pêche (FFP) and the Fonds Européens pour les Affaires Maritimes et la Pêche (FEAMP) (PFEA390017FA1000006).
Landings data we available thanks to the Harmonie database of the “Système d’Information Halieutique” (SIH) of Ifremer.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Anders N., Fernö A., Humborstad O.-B., Løkkeborg S., Rieucau G., Utne-Palm A. C. (2017a). Size-dependent social attraction and repulsion explains the decision of Atlantic cod gadus morhua to enter baited pots. J. Fish Biol. 91, 1569–1581. doi: 10.1111/jfb.13453
Anders N., Fernö A., Humborstad O.-B., Løkkeborg S., Utne-Palm A. C. (2017b). Species specific behaviour and catchability of gadoid fish to floated and bottom set pots. ICES J. Mar. Sci. 74, 769–779. doi: 10.1093/icesjms/fsw200
Balık İ., Çubuk H. (2000). Effect of net colours on efficiency of monofilament gillnets for catching some fish species in lake beyşehir. Turk. J. Fish. Aquat. Sci. 1, 29–32.
Bellido J. M., Santos M. B., Pennino M. G., Valeiras X., Pierce G. J. (2011). Fishery discards and bycatch: Solutions for an ecosystem approach to fisheries management? Hydrobiologia 670, 317. doi: 10.1007/s10750-011-0721-5
Cadiou G., Boudouresque C. F., Bonhomme P., Le Diréach L. (2009). The management of artisanal fishing within the marine protected area of the port-cros national park (northwest Mediterranean sea): A success story? ICES J. Mar. Sci. 66, 41–49. doi: 10.1093/icesjms/fsn188
Chen K., Wang J., Pang J., Cao Y., Xiong Y., Li X., et al. (2019). MMDetection: Open MMLab detection toolbox and benchmark. (Ithaca, USA: Cornell University. doi: 10.48550/arXiv.1906.07155
Chladek J., Stepputtis D., Hermann A., Kratzer I. M. F., Ljungberg P., Rodriguez-Tress P., et al. (2021). Using an innovative net-pen-based observation method to assess and compare fish pot-entrance catch efficiency for Atlantic cod (Gadus morhua). Fish. Res. 236, 105851. doi: 10.1016/j.fishres.2020.105851
Collins K., Mallinson J. (2012). Surveying black bream, spondyliosoma cantharus (L.), nesting sites using sidescan sonar. Underw. Technol. 30, 183–188. doi: 10.3723/ut.30.183
CRPMEM Bretagne (2018). Comité Régional des pêches Maritimes et des Elevages Marins de Bretagne. 2 Pages. It is a legislative text available at: https://www.comitedespeches-finistere.fr/upload/files/2018-015-NASSES%20A%20POISSON-B-%2030%2003%202018%281%29.pdf.
Dawkins M., Sherrill L., Fieldhouse K., Hoogs A., Richards B., Zhang D., et al. (2017). An open-source platform for underwater image and video analytics, in: 2017 IEEE winter conference on applications of computer vision (WACV). Presented at 2017 IEEE Winter Conf. Appl. Comput. Vision (WACV) pp, 898–906. doi: 10.1109/WACV.2017.105
Dulcic J., Lipej L., Glamuzina B., Bartulovic V. (2006). Diet of spondyliosoma cantharus and diplodus puntazzo (Sparidae) in the Eastern central Adriatic. Cybium 30, 115–122.
Eigaard O. R., Bastardie F., Breen M., Dinesen G. E., Hintzen N. T., Laffargue P., et al. (2016). Estimating seabed pressure from demersal trawls, seines, and dredges based on gear design and dimensions. ICES J. Mar. Sci. 73, i27–i43. doi: 10.1093/icesjms/fsv099
Faillettaz R., Picheral M., Luo J. Y., Guigand C., Cowen R. K., Irisson J.-O. (2016). Imperfect automatic image classification successfully describes plankton distribution patterns. Methods Oceanogr. Comput. Vision Oceanography 15–16, 60–77. doi: 10.1016/j.mio.2016.04.003
Folkins M. H., Grant S. M., Walsh P. (2021). A feasibility study to determine the use of baited pots in Greenland halibut (Reinhardtius hippoglossoides) fisheries, supported by the use of underwater video observations. PeerJ 9, e10536. doi: 10.7717/peerj.10536
Friard O., Gamba M. (2016). BORIS: a free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol. Evol. 7, 1325–1330. doi: 10.1111/2041-210X.12584
Fujimori Y., Ochi Y., Yamasaki S., Ito R., Kobayashi Y., Yamamoto J., et al. (2018). Optical and acoustic camera observations of the behavior of the kuril harbor seal phoca vitulina stejnegeri after invading a salmon setnet. Fish. Sci. 84, 953–961. doi: 10.1007/s12562-018-1236-z
Furevik D. M., Humborstad O.-B., Jørgensen T., Løkkeborg S. (2008). Floated fish pot eliminates bycatch of red king crab and maintains target catch of cod. Fish. Res. 92, 23–27. doi: 10.1016/j.fishres.2007.12.017
Furevik D. M., Løkkeborg S. (1994). Fishing trials in Norway for torsk (Brosme brosme) and cod (Gadus morhua) using baited commercial pots. Fish. Res. 19, 219–229. doi: 10.1016/0165-7836(94)90040-X
Garcia-Mederos A. M., Tuya F., Tuset V. M. (2015). The structure of a nearshore fish assemblage at an oceanic island: insight from small scale fisheries through bottom traps at gran canary island (Canary islands, eastern Atlantic). Aquat Living Resour 28, 1–10. doi: 10.1051/alr/2015002
Garrison V., Rogers C., Beets J., Friedlander A. (2004). The habitats exploited and the species trapped in a Caribbean island trap fishery. Environ. Biol. Fishes 71, 247–260. doi: 10.1007/s10641-004-0301-x
Glass C. W., Wardle C. S., Gosden S. J., Racey D. N. (1995). Studies on the use of visual stimuli to control fish escape from codends. i. laboratory studies on the effect of a black tunnel on mesh penetration. Fish. Res. 23, 157–164. doi: 10.1016/0165-7836(94)00330-Y
Humborstad O.-B., Utne-Palm A. C., Breen M., Løkkeborg S. (2018). Artificial light in baited pots substantially increases the catch of cod (Gadus morhua) by attracting active bait, krill (Thysanoessa inermis). ICES J. Mar. Sci. 75, 2257–2264. doi: 10.1093/icesjms/fsy099
Jørgensen T., Løkkeborg S., Furevik D., Humborstad O.-B., De Carlo F. (2017). Floated cod pots with one entrance reduce probability of escape and increase catch rates compared with pots with two entrances. Fish. Res. 187, 41–46. doi: 10.1016/j.fishres.2016.10.016
Kopp D., Coupeau Y., Vincent B., Morandeau F., Méhault S., Simon J. (2020). The low impact of fish traps on the seabed makes it an eco-friendly fishing technique. PloS One 15, e0237819. doi: 10.1371/journal.pone.0237819
Løkkeborg S. (1998). Feeding behaviour of cod,Gadus morhua: Activity rhythm and chemically mediated food search. Anim. Behav. 56, 371–378. doi: 10.1006/anbe.1998.0772
Løkkeborg S., Olla B. L., Pearson W. H., Davis M. W. (1995). Behavioural responses of sablefish, anoplopoma fimbria, to bait odour. J. Fish Biol. 46, 142–155. doi: 10.1111/j.1095-8649.1995.tb05953.x
Lucchetti A., Vasapollo C., Virgili M. (2017). Sea Turtles bycatch in the Adriatic Sea set net fisheries and possible hot-spot identification. Aquat. Conserv. Mar. Freshw. Ecosyst. 27, 1176–1185. doi: 10.1002/aqc.2787
Marshak A. R., Hill R. L., Sheridan P., Schärer M. T., Appeldoorn R. S. (2008). In-situ observations of antillean fish trap contents in southwest Puerto Rico: Relating catch to habitat and damage potential. 60th Gulf and Caribbean Fisheries Institute. 7, 447–53.
Morfin M., Méhault S., Benoît H. P., Kopp D. (2017). Narrowing down the number of species requiring detailed study as candidates for the EU common fisheries policy discard ban. Mar. Policy 77, 23–29. doi: 10.1016/j.marpol.2016.12.003
Northridge S., Coram A., Kingston A., Crawford R. (2017). Disentangling the causes of protected-species bycatch in gillnet fisheries. Conserv. Biol. 31, 686–695. doi: 10.1111/cobi.12741
Novales Flamarique I. (2019). Swimming behaviour tunes fish polarization vision to double prey sighting distance. Sci. Rep. 9, 944. doi: 10.1038/s41598-018-37632-1
Orsay B., Dartay M. (2011). Catch efficiency of monofilament gill nets configured at various colors and hanging ratios. J. Anim. Vet. Adv. 10, 1219–1226. doi: 10.3923/javaa.2011.1219.1226
Pajuelo J. G., Lorenzo J. M. (1999). Life history of black seabream, spondyliosoma cantharus, off the canary islands, central-east Atlantic. Environ. Biol. Fishes 54, 325–336. doi: 10.1023/A:1007515301745
Pascual-Fernández J. J., Pita C., Bavinck M. (Eds.) (2020). Small-scale fisheries in Europe: Status, resilience and governance, MARE publication series (Amsterdam: Springer International Publishing). doi: 10.1007/978-3-030-37371-9
Pawson M. G. (1995). “Biogeographical identification of English channel fish and shellfish stocks,” in Fisheries research technical report, vol. 99. (Ministry of Agriculture, Fisheries and Food Directorate of Fisheries Research), 72.
Petetta A., Virgili M., Guicciardi S., Lucchetti A. (2021). Pots as alternative and sustainable fishing gears in the Mediterranean Sea: an overview. Rev. Fish Biol. Fish. 31, 773–795. doi: 10.1007/s11160-021-09676-6
Pinder A. C., Velterop R., Cooke S. J., Britton J. R. (2017). Consequences of catch-and-release angling for black bream spondyliosoma cantharus, during the parental care period: Implications for management. ICES J. Mar. Sci. 74, 254–262. doi: 10.1093/icesjms/fsw151
Pita C., Gamito S., Erzini K. (2002). Feeding habits of the gilthead seabream (Sparus aurata) from the ria Formosa (southern Portugal) as compared to the black seabream (Spondyliosoma cantharus) and the annular seabream (Diplodus annularis). J. Appl. Ichthyol. 18, 81–86. doi: 10.1046/j.1439-0426.2002.00336.x
Pravin P., Meenakumari B., Baiju M., Barman J., Baruah D., Kakati B. (2011). Fish trapping devices and methods in Assam - a review. Indian J. Fish 58, 127–135.
Prosperi P., Kirwan J., Maye D., Bartolini F., Vergamini D., Brunori G. (2019). Adaptation strategies of small-scale fisheries within changing market and regulatory conditions in the EU. Mar. Policy 100, 316–323. doi: 10.1016/j.marpol.2018.12.006
Purves M. G., Agnew D. J., Moreno G., Daw T., Yau C., Pilling G. (2003). Distribution, demography, and discard mortality of crabs caught as bycatch in an experimental pot fishery for toothfish (Dissostichus eleginoides) in the south Atlantic.Fishery Bull. 101 (4), 874–88.
Robert M., Cortay A., Morfin M., Simon J., Morandeau F., Deneubourg J. L., et al. (2020). A methodological framework for characterizing fish swimming and escapement behaviors in trawls. PloS One 15, e0243311. doi: 10.1371/journal.pone.0243311
Robichaud D., Hunte W., Chapman M. R. (2000). Factors affecting the catchability of reef fishes in antillean fish traps. Bull. Mar. Sci. 67, 831–844.
Rose C. S., Stoner A. W., Matteson K. (2005). Use of high-frequency imaging sonar to observe fish behaviour near baited fishing gears. Fish. Res. 76, 291–304. doi: 10.1016/j.fishres.2005.07.015
Russel B., Pollard D., Carpenter K. E. (2014) IUCN red list of threatened species: Spondyliosoma cantharus (IUCN Red List Threat. Species). Available at: https://www.iucnredlist.org/en (Accessed 5.19.22).
Shester G. G., Micheli F. (2011). Conservation challenges for small-scale fisheries: Bycatch and habitat impacts of traps and gillnets. Biol. Conserv. Ecoregional-scale Monit. within Conserv. areas rapidly changing Climate 144, 1673–1681. doi: 10.1016/j.biocon.2011.02.023
Simon J., Kopp D., Larnaud P., Vacherot J.-P., Morandeau F., Lavialle G., et al. (2020). Using automated video analysis to study fish escapement through escape panels in active fishing gears: Application to the effect of net colour. Mar. Policy 116, 103785. doi: 10.1016/j.marpol.2019.103785
Stelzenmüller V., Gimpel A., Haslob H., Letschert J., Berkenhagen J., Brüning S. (2021). Sustainable co-location solutions for offshore wind farms and fisheries need to account for socio-ecological trade-offs. Sci. Total Environ. 776, 145918. doi: 10.1016/j.scitotenv.2021.145918
Stoner A. W. (2004). Effects of environmental variables on fish feeding ecology: Implications for the performance of baited fishing gear and stock assessment. J. Fish Biol. 65, 1445–1471. doi: 10.1111/j.0022-1112.2004.00593.x
Suuronen P., Chopin F., Glass C., Løkkeborg S., Matsushita Y., Queirolo D., et al. (2012). Low impact and fuel efficient fishing–looking beyond the horizon. Fish. Res. 119–120, 135–146. doi: 10.1016/j.fishres.2011.12.009
Tran P. D., Nguyen L. T., To P., Nguyen K. Q. (2020). Effects of the trap entrance designs on the catch efficiency of swimming crab charybdis feriata fishery. Fish. Res. 232, 105730. doi: 10.1016/j.fishres.2020.105730
Vadziutsina M., Riera R. (2020). Review of fish trap fisheries from tropical and subtropical reefs: Main features, threats and management solutions. Fish. Res. 223, 105432. doi: 10.1016/j.fishres.2019.105432
Wanner G. A., Klumb R. A., Shuman D. A., Steffensen K., Stukel S., Utrup N. J. (2010). Comparison of green and white mesh trammel nets and gill nets to assess the fish community in a Large river. North Am. J. Fish. Manage. 30, 12–25. doi: 10.1577/M07-221.1
Keywords: passive gear, Sparidae, underwater video, Bay of Biscay, fish trap
Citation: Méhault S, Morandeau F, Simon J, Faillettaz R, Abangan A, Cortay A and Kopp D (2022) Using fish behavior to design a fish pot: Black seabream (Spondyliosoma cantharus) case study. Front. Mar. Sci. 9:1009992. doi: 10.3389/fmars.2022.1009992
Received: 02 August 2022; Accepted: 05 September 2022;
Published: 26 September 2022.
Edited by:
Alessandro Lucchetti, National Research Council (CNR), ItalyReviewed by:
Andrea Petetta, National Research Council (CNR), ItalyCopyright © 2022 Méhault, Morandeau, Simon, Faillettaz, Abangan, Cortay and Kopp. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sonia Méhault, c29uaWEubWVoYXVsdEBpZnJlbWVyLmZy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.