 Yun Lu1,2
Yun Lu1,2 Xinyi Tang
Xinyi Tang Wenxin Wang
Wenxin Wang Jun Yang
Jun Yang Shengjun Wang
Shengjun Wang
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Immunol. , 16 July 2024
Sec. Immunological Tolerance and Regulation
Volume 15 - 2024 | https://doi.org/10.3389/fimmu.2024.1422541
This article is part of the Research Topic Immune Responses in the Progression of Allergy and Asthma View all 9 articles
The silent information regulator sirtuin 1 (SIRT1) protein is an NAD+-dependent class-III lysine deacetylase that serves as an important post-transcriptional modifier targeting lysine acetylation sites to mediate deacetylation modifications of histones and non-histone proteins. SIRT1 has been reported to be involved in several physiological or pathological processes such as aging, inflammation, immune responses, oxidative stress and allergic diseases. In this review, we summarized the regulatory roles of SIRT1 during allergic disorder progression. Furthermore, we highlight the therapeutic effects of targeting SIRT1 in allergic diseases.
The silent information regulator sirtuin 1 (SIRT1) protein is a highly conserved NAD+-dependent deacetylase in the sirtuin family which mainly acts as a post-translational regulator and plays a key role in histone and non-histone deacetylation. Among the sirtuins, SIRT1 was the first one discovered in mammals. The deacetylation mediated by SIRT1 profoundly impacts numerous biological processes, including DNA damage repair (1), gene transcription, glucose and lipid metabolism (2), oxidative stress (3), inflammation (4), apoptosis (5), aging (6), and autophagy (7). In addition to regulating histone acetylation, SIRT1 is also involved in many post-translational modifications of non-histones, such as transcription factors. The expression of SIRT1 was down-regulated in many acute inflammatory responses or inflammation-related diseases. SIRT1 can directly interact with or promote the histone deacetylation in the gene promoter region of inflammatory cytokines, further inhibit the transcription of target genes, and play an anti-inflammatory role, such as deacetylated HAK16 inhibits TNF-α transcription (8–10). Moreover, SIRT1 can mediate the deacetylation of inflammation associated transcription factors such as nuclear factor κB (NF-κB), activating protein 1(AP1) and HIF-1α and further decrease the expression of pro-inflammatory genes (11–15).
Hypersensitivity denotes an abnormality of immune system, contributing to hyperactive immune cells production and aggressive inflammation, which culminates in physiological disorders and/or tissue and cell damage. In recent years, the potential roles of SIRT1 in allergic diseases has been confirmed. In this review, we take deeper insights into the role of SIRT1 in hypersensitivity reactions and conclude current therapeutic avenues that target SIRT1 to alleviate these reactions.
When allergens and harmful microorganisms enter the body, epithelial cells respond by producing and releasing cytokines such as IL-25, IL-33, and thymic stromal lymphopoietin (TSLP). These cytokines activate type II innate lymphoid cells (ILC2) and contribute to the development and maturation of Th2 cells, while, dendritic cells (DCs) activate Th2 cells in the lymph nodes in a specific manner, resulting in the production of various type II cytokines, including IL-4, IL-5, and IL-13 (16–18). Upon cytokine stimulation, plasma cells secrete IgE, initiating a cascade of events within the immune system (19). IgE and the high-affinity receptor FcϵRI on the surface of mast cells and basophils put the body in a sensitized state. Upon re-exposure to an allergen, mast cells and basophils undergo activation and degranulation, thereby releasing type II cytokines that contribute to the process of allergic inflammation (20, 21). Some recent studies have observed that SIRT1 is involved in the progression of allergic inflammatory diseases. SIRT1 deacetylates transcription factors, affecting signaling pathways like AMPK, MAPK, and NF-κB, reducing inflammatory factor secretion, thereby alleviating inflammation. Targeting SIRT1 may improve clinical symptoms and tissue damage caused by allergic inflammation.
Airway Allergic Inflammation (AAI) is a chronic inflammatory disease of the airways characterized by systemic IgE elevation, eosinophil, and lymphocyte infiltration, and increased mucus secretion and airway hyperresponsiveness (AHR) (22). Eosinophil-derived pro-inflammatory mediators are a major contributor to asthma-related inflammation, which can lead to damage of airway epithelial cells, airway dysfunction, and an excess of mucus secretion (23). The aforementioned event triggers a sequence of inflammatory responses that culminate in ischemia, hemorrhage, edema, and tissue injury. Moreover, the stimulation of antigens leads to the infiltration of effector T cells into the affected tissue, thereby inducing an inflammatory reaction that can exacerbate the tissue damage.
Research has found that patients who suffer from allergic airway inflammation (AAI) may have reduced levels of SIRT1 in their lungs but high levels of SIRT1 in their peripheral blood. This correlation is positively linked to IgE levels and negatively linked to lung function (24). Therefore, measuring the level of SIRT1 in the serum could potentially assist in the diagnosis of AAI, and increasing the level of SIRT1 in the lungs may be a way to treat AAI. However, further research is needed to fully understand the connection between SIRT1 and AAI inflammation. Recent studies have shown that individuals with asthma often have increased levels of pAMPK and SIRT1 and that AMPK/SIRT1/PGC1α plays a critical role in metabolic regulation and energy expenditure during the development of AAI (25). As a result of this discovery, it has been found that the AMPK/SIRT1/Nrf2/HO-1 signaling pathway can mitigate oxidative stress in human bronchial epithelial cells and alleviate AAI (26). Additionally, researchers have noted elevated levels of IL-6 and reduced levels of SIRT1 in bronchial epithelial cells. Further studies have shown that SIRT1 inhibits Akt-dependent expression of IL-6. Consistent with this, the use of SIRT1 agonists has been found to reduce IL-6 expression and alleviate AAI (27).
AAI is an airway inflammation in which numerous immune cells are involved in disease progression. Insufficient CD4+CD25+ Tregs were found in AAI patients with impaired function, which was positively correlated with Foxo1 and SIRT1. Therefore, targeting SIRT1 may ameliorate the deficiency of Treg quantity and function and alleviate the inflammatory response (28–30). In addition, the occurrence of AAI is associated with type 3 innate lymphocyte activation, which leads to macrophage activation and the production of neutrophil chemokines. It has been found that the expression of chemokine ligand 2(CXCL2), interleukin-1β(IL-β), and tumor necrosis factor-α(TNF-α) are reduced in myeloid-specific SIRT1 deficient macrophages (BMDM). SIRT1 inhibits the ERK/p38 MAPK pathway in BMDM.SIRT1 inhibited AAI by decreasing BMDM cytokine secretion and activation of MAPK signaling pathway (31). PPAR-γ plays an important role in the inflammatory response, inducing the oxidative metabolism of macrophages by upregulating the oxidation of fatty acids and mitochondrial biosynthesis (32). PPAR-γ increased the expression of SIRT1 in macrophages and upregulated SIRT1 increased the secretion of IL-10, thus playing an anti-inflammatory role (33).
GATA3 and STAT6 transcription factors help Th2 cells produce type II cytokines, which can exacerbate inflammation. Low acetylation levels of GATA 3 are associated with reduced Th2 function, and SIRT1 has been reported to mediate the immune response of Th2 cells through deacetylation of GATA 3 (34–36). In autoimmune diseases and chronic inflammation, Th17 cells and their effector cytokine IL-17 are known to promote inflammation. Patients with asthma have been found to have increased Th17 cells and elevated IL-17 expression. In asthmatic mice, significant increases in eosinophils and mucus secretion were observed in their alveolar lavage fluid (BLAF). The cytokines secreted by Th2 and Th17 cells can alter the structure of bronchial epithelial cells and airway smooth muscle cells, leading to severe goblet hyperplasia and increased mucin production. NF-κB is a vital transcription factor that governs the production of pro-inflammatory cytokines and the recruitment of inflammatory cells. Additionally, it regulates the expression of several genes that are critical in the inflammatory response. In a mouse model of AAI, NF-κB has a significant impact on Th2 cell-related cytokine production and inflammatory cell recruitment. If we can reduce the phosphorylation of the classical NF-κB pathway p65 in BLAF, we may be able to decrease the production of Th2 and Th17-associated pro-inflammatory factors. This could ultimately lead to an improvement in AAI (37, 38). It is plausible to mitigate AAI by reducing the phosphorylation levels of p52 and RelB in the nonclassical pathway of NF-κB, alongside ERK1/2 and p38 in the MAPK signaling pathway (Figure 1). This approach can potentially lead to the reduction of Th9 cell infiltration and an elevation in the proportion of Treg (39–41). To further investigate the specific mechanisms affecting the NF-κB signaling pathway and identify therapeutic targets for the treatment of allergic asthma, the study shows that SIRT1 can inhibit the NF-κB signaling pathway and reduce the secretion of inflammatory factors, thereby controlling the inflammatory response of AAI (42–45).
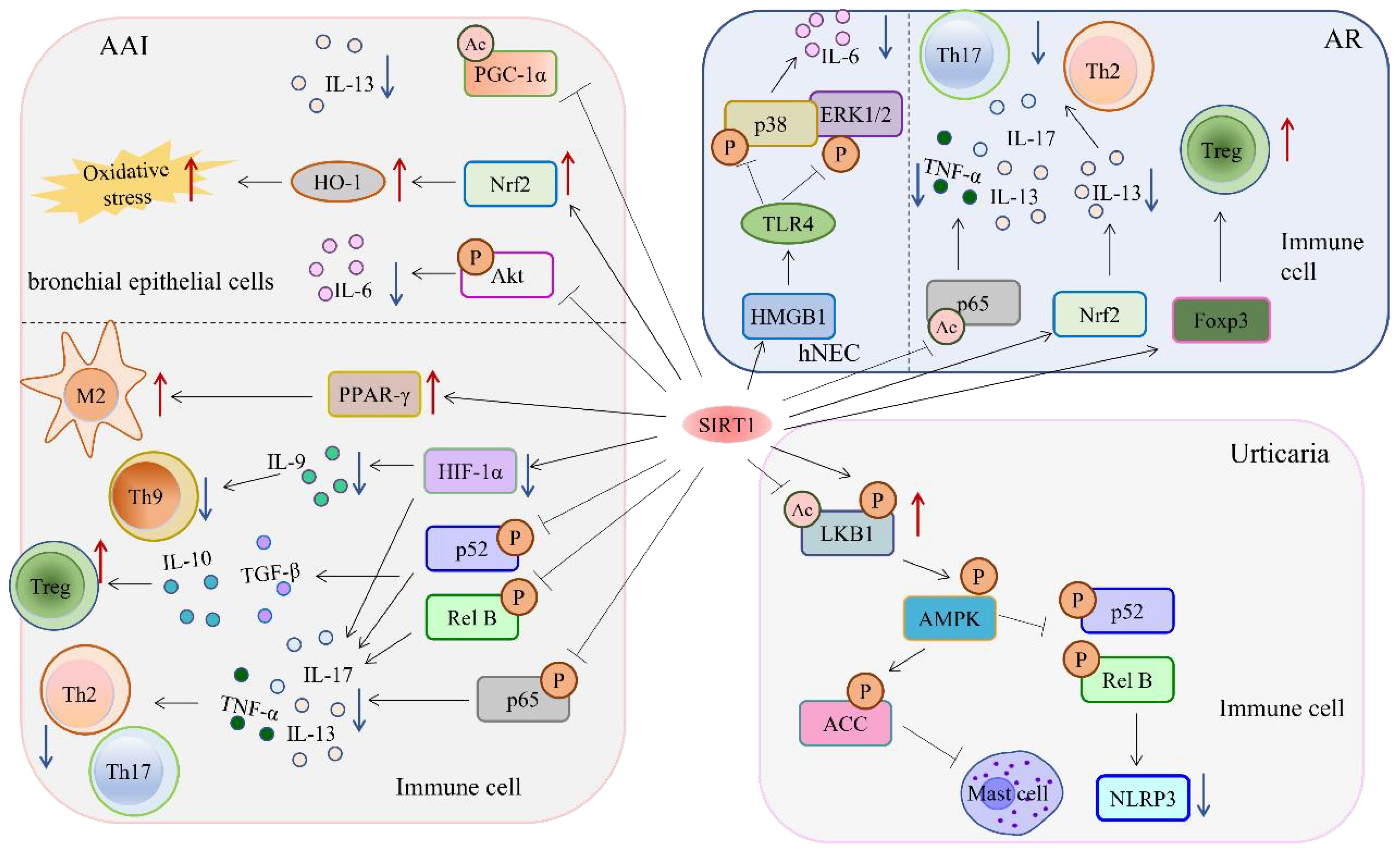
Figure 1 The target of SIRT1 in hypersensitivity reaction.
The protein SIRT1 is a vital component in reducing hypersensitivity reactions by deacetylating multiple targets. Signaling pathways involved in SIRT1 hypersensitivity in different diseases. It achieves this by suppressing inflammatory factors through its ability to inhibit the NF-κB and MAPK signaling pathways. Additionally, SIRT1 activates the AMPK pathway while inhibiting mast cell activation, which works to relieve inflammation by reducing the production of inflammatory factors. SIRT1 also regulates HIF-1α activity, leading to a decrease in the proportion of Th9 cells that cause an inflammatory response. Furthermore, it regulates PPAR-γ-associated histones to enhance the immunosuppressive function of M2-like monocytes. Lastly, SIRT1 promotes the Nrf2/HO-1 signaling pathway, which supports the body’s oxidative stress response by boosting ROS production.
HIF-1α plays a vital role in the inflammatory response, and HIF-1α activation stimulates the promotion of angiogenesis, vasodilation, and vascular permeability in inflamed tissues. SIRT1 can mediate acetylation modification of HIF-1α to decrease its activity (46). SIRT1 interacts with HIF-1α to regulate immune cells. mTOR is an evolutionarily conserved serine/threonine kinase involved in cell transcription, growth, proliferation, and survival and controls cellular autophagy (47). Studies have reported that mTOR signaling mediates epithelial cell proliferation, migration, and autophagy involved in the regulation of AAI (48–50). In allergic mice, lung levels of phosphorylated (p)-mTOR were reduced, and mTOR signaling activation suppressed AAI by inhibiting autophagy (49). It has been observed that EX-527, an inhibitor of SIRT1, can enhance this phenomenon and is favorable for the control of AAI (51). mTOR acts as an upstream signal of the HIF-1α glycolysis pathway, regulating the glycolysis pathway in immune cells (52). HIF-1α is a crucial transcription factor that regulates the expression of glycolytic enzymes and plays a central role in producing pro-inflammatory cytokines (53). HIF-1α deficiency increases Treg cell production and blocks glycolysis to inhibit Th17 cell differentiation (54, 55). Recent studies have shown that SIRT1 in mouse tissue directly inactivates HIF-1α, with or without hypoxia (56, 57). Consistent with the previous results, it was found that SIRT1 deficiency-induced upregulation of HIF-1α decreased the percentage of Treg cells and increased the rate of Th17 cells. In this case, SIRT1 deficiency induced mTOR upregulation and promoted HIF-1α expression. In addition, HIF-1α not only promotes IL-9 production by regulating the glycolytic pathway in CD4+ T cells but also directly binds to the IL-9 gene promoter to drive IL-9 transcription (Figure 1). In Th9 cells, SIRT1 suppressed IL-9 transcription by inhibiting the mTOR/HIF-1α pathway, thereby alleviating AAI (58, 59).
In summary, many studies have shown that SIRT1 plays a protective role in allergic airway inflammation. Activation of SIRT1 can cause a variety of signaling pathways to promote the activation of various transcription factors, resulting in the reduction of pro-inflammatory cytokines and chemokines associated with inflammatory cells, and finally preventing or alleviating AAI. Inconsistent with the foregoing, SIRT1 has been shown to play a pro-inflammatory role in OVA-induced mouse models of airway inflammation (60, 61). This may involve SIRT1 inhibiting PPAR-γ in DC cells and promoting the differentiation of DC cells into Th2-like cells (62). Similarly, remission of airway inflammation was observed with pharmacological inhibitors of SIRT1 (60). The current understanding of the role of SIRT1 in AAI has been challenged by recent research findings, indicating a bidirectional effect. However, the specific mechanism underlying this phenomenon has yet to be fully elucidated. Further investigations are therefore warranted to clarify the reasons why SIRT1 appears to play a bidirectional role in airway inflammation.
Allergic rhinitis (AR) manifests as sneezing, nasal congestion, nasal itchiness, and rhinorrhea (nasal discharge) due to IgE-mediated reactions to inhaled allergens. Rhinitis symptoms arise from inflammation in the nasal mucosa and/or sinuses. Research has shown that there is a decrease in SIRT1 expression in cases of AR. It is believed that SIRT1 controls the production of Th2 cell-related pro-inflammatory factors, which can be suppressed by proteins linked to the HMGB1/TLR4 pathway, ultimately reducing the severity of AR (63). In a study involving ovalbumin (OVA)-induced allergic rhinitis mice, researchers administered the SIRT1 agonist resveratrol (RSV). The results indicated a reduction in the expression of HMGB1 and TLR4 in the nasal mucosa following RSV treatment. Furthermore, SIRT1 expression was enhanced, leading to a mitigation of allergic rhinitis symptoms (64, 65).
Hypoxia-inducible factor 1α (HIF-1α) plays a crucial role in the progression of AR. Research indicates that inhibiting HIF-1α results in a notable decrease in eosinophils in BLAF, reduced levels of nasal mucosa and systemic Th2-related cytokines, and an improvement in AR symptoms (66, 67). Another important factor in AR is dendritic cells, which regulate T cell proliferation and differentiation through signaling (68, 69). Interaction between T-cell immunoglobulin and mucin domain 4(TIM4) and TIM1 increases PI3K/Akt phosphorylation in CD4+ T cells and enhances SIRT1 expression. Additionally, SIRT1 promotes Th2 CD4+ T cell proliferation by inhibiting Fas ligand and caspase-3 expression (70). Further exploration of how HIF-1α influences AR has revealed that HIF-1α-deficient dendritic cells utilize the SIRT1/NF-κB pathway to mitigate the inflammatory response, thus alleviating AR (71).
Recent studies have shown that beyond Th1 and Th2, Th9, Th17, and Treg play significant roles in the progression of AR (72, 73). Tregs interact directly with immune cells, releasing anti-inflammatory cytokines to maintain immune tolerance in the body. In AR, Tregs inhibit Th2 differentiation and restrict airway inflammation (74). Modulating SIRT1 levels can enhance Foxp3 expression, thereby boosting Treg function and differentiation (75, 76). Furthermore, SOCS1 modulates T cell activation, development, and differentiation by negatively impacting the Janus kinase (JAK)/STAT signaling pathway (77). Based on the research as mentioned above results, some researchers have stated that it has been observed in the progression of AR disease that inhibition of the SOCS1/SIRT1 pathway in CD4+ T cells promotes the proliferation of Treg, thereby inhibiting the proliferation of Th2 and alleviating AR (78). In contrast, some researchers reported that loss of Sirt1 impairs Treg survival, leading to antigen-induced T cell proliferation and inflammation (79). This difference may be caused by different disease models and research backgrounds, and the specific mechanism remains to be studied in depth.
IL-13 secreted by Th2 cells is considered to be a central mediator of allergic inflammation, stimulating mucin synthesis and secretion (80). Studies have shown that IL-13 can induce inflammatory responses and mucus secretion in human nasal epithelial cells (hNEC), and the severity of AR can be determined based on the expression of IL-13 (81). Nrf2 and Kelch-like ECH associated protein (Keap 1) combine as dimers in the cytoplasm. When stimulated, Nrf 2 dissociates from Keap 1 and translocate into the nucleus, inducing the expression of HO-1 (82). In addition, results show that activating the Nrf2/HO-1 pathway can inhibit the progression of AR (83, 84). Here, some researchers found that Formononetin relies on activation of the SIRT1/Nrf2 pathway to reduce the secretion of inflammatory factors and mucus formation caused by IL-13 (85). Consistent with this, SIRT1 mediates acetylation modification of NF-κB p65, inactivating the NF-κB pathway (86). Interfering with HDAC4 can restore the expression of SIRT1 and reduce the inflammatory response caused by IL-13 by activating the SIRT1/NF-κB pathway (81).
In conclusion, the expression of SIRT1 in nasal epithelial cells showed a protective effect on AR, and high expression of SIRT1 could reduce the inflammatory response of nasal epithelial cells and the production of mucus. In addition, during the progression of AR, the expression of SIRT1 in immune cells such as Tregs also showed protective effects on inflammation, mainly by promoting the proliferation of anti-inflammatory cells Tregs. Consistent with reports related to AAI, in AR, it is now found that SIRT1 has different effects on different immune cells, and in DCs, SIRT1 expression has been reported to show pro-inflammatory effects. SIRT1 shows different effects in different cells in the pathogenesis of AR, and the specific reasons for this are still unclear and need to be studied.
Urticaria is a type of immune response that results in localized inflammation and swelling. It is caused by the widening and increased permeability of small blood vessels in the skin and mucous membranes. This condition can present as either rubella, angioedema, or a combination of both (87, 88). The sensitization of mast cells initiates the development of urticaria through IgE, which leads to the release of inflammatory mediators like histamine and other pro-inflammatory factors (87). The progression of urticaria can also be influenced by an imbalance in immune cells, particularly CD4+ helper T cells. This heightened sensitivity can contribute to the development of urticaria (89, 90). During the inflammatory response, JNK is activated by MEK kinase 2 (MEKK2) under the influence of inflammation and stress. Subsequently, JNK participates in the activation of mast cells triggered by antigens, which leads to the expression of TNF-α and IL-6 (91, 92). Moreover, research suggests that the level of phosphorylation of ERK is closely related to the expression of TNF-α (93). When the endogenous signal is recognized by the TLR4 receptor, IκB in the NF-κB signaling pathway phosphorylates and regulates the nuclear translocation of downstream NF-κB p65, thereby promoting the transcription of NLRP3 and IL-1β (94). It is well known that the NLRP3 inflammasome is a crucial factor in regulating apoptosis (95). NF-κB and MAPK signaling pathways can activate NLRP3 (96). According to recent reports, there is a strong correlation between NLRP3 and the onset of urticaria (97). To effectively treat this condition, it’s crucial to regulate the TLR4/NF-κB/MAPK/NLRP3 inflammasome cascade. Moreover, SIRT1 can inhibit NLRP3 activation by boosting the LKB1/AMPK pathway (98). The AMPK/SIRT1 pathway is also known to curb the activation of NF-κB/NLRP3 (99, 100). In line with these findings, recent studies have pointed out that Jingfang Granules (JFG) can increase the expression of AMPK and SIRT1 by promoting the phosphorylation of LKB1 and AMPK, thereby inhibiting the activation of NLRP3, inhibiting OVA/aluminum hydroxide-induced skin inflammation, and alleviating Urticaria disease symptoms (94).
The role of SIRRT1 in urticaria is still less studied. Since SIRT1 is a key regulatory factor in the regulation of glucose metabolism and insulin secretion, researchers have observed that SIRT1 can participate in the disorder of glucose metabolism in the skin tissue of urticaria mice, and up-regulation of SIRT1 can increase insulin secretion. Promote aerobic oxidation, inhibit glycolysis, and reduce the expression of pro-inflammatory factors, effectively relieve urticaria. Whether SIRT1 is also involved in other cellular processes in the pathogenesis of urticaria is unknown.
Atopic dermatitis (AD) is a chronic skin condition characterized by persistent itching and eczema-like lesions. The development of AD has been linked to mutations in the filaggrin (FLG), as well as the presence of inflammatory factors like IL-4,IL-5and IL-13 (101). These elements can damage the skin’s barrier and trigger the migration of eosinophils (102, 103). Here, researchers investigate how SIRT1-mediated FLG or inflammatory immune cells contribute to AD inflammatory response. Studies have shown that SIRT1 levels are reduced in AD, which supports the finding that SIRT1 can help maintain the skin barrier in mouse models of AD (104, 105). Among them, FLG deficiency caused loricrin deficiency and SIRT1 pathway was destroyed (106). Deficiencies in FLG can result in a lack of loricrin and harm to the SIRT1 pathway, which is associated with inflammatory damage in AD. To combat this, targeting SIRT1 presents a potential solution. Resveratrol, an SIRT1 agonist, may serve as a therapeutic option by blocking Akt, MAPK, NF-κB, and STAT3 signaling pathways, reducing inflammatory factors, inhibiting oxidative stress and angiogenesis, and potentially alleviating the condition (107–109).
While SIRT1 levels in keratinocytes and dermal fibroblasts are upregulated, HDAC6 and CXCL13 levels are also upregulated to aggravate the inflammatory response of specific dermatitis, which is involved in the increased expression of Th1 and Th2 cytokines and the downregulation of Foxp3 and IL-10. Inconsistently, studies have suggested that SIRT1-deficient mice are sensitive to OVA’s percutaneous attack, and SIRT1 expression has a protective effect on the skin tissue of AD mice.
It has been observed that during anaphylactic shock, when SIRT1 is specifically knocked out in mast cells, there is an increase in AMPK-dependent FcϵRI signaling. This leads to an enhancement in mast cell activation both in vitro and in vivo, as confirmed by two separate studies (110, 111). At the same time, SIRT1 also weakened the inhibition of AMPK pathway through protein tyrosine phosphatase 1B (PTP1B), while also enhancing the tyrosine kinase (Syk) pathway in the spleen. As a result, it effectively inhibits allergic inflammation (111).
Recent reports have explored treatment options for allergic inflammation, including drugs that target SIRT1 to provide relief for allergic diseases (Table 1). Among the drugs that have demonstrated effectiveness in alleviating AAI are Allopurinol, GW9962, Hylocereus undatus flower (HUF), Pterostilbene (Pts), Gentiopicroside (GPS), and EX-527 (26, 33, 43, 51, 112, 113). Studies have found that SIRT1 expression in immune cells can help relieve allergic rhinitis, and the use of SIRT1 agonist resveratrol has shown promise in treating AR (64, 71). Similarly, the use of resveratrol can also alleviate AD (109). However, in nasal epithelial cells, the inhibition of SIRT1, such as with Formononetin, has been found to reduce AR, and further study is needed to understand the specific mechanism behind this (85). Studies have revealed that Jingfang granules exhibit promising outcomes in treating urticaria cases as they significantly reduce OVA/alumina-induced inflammation and lesions in mice (94).
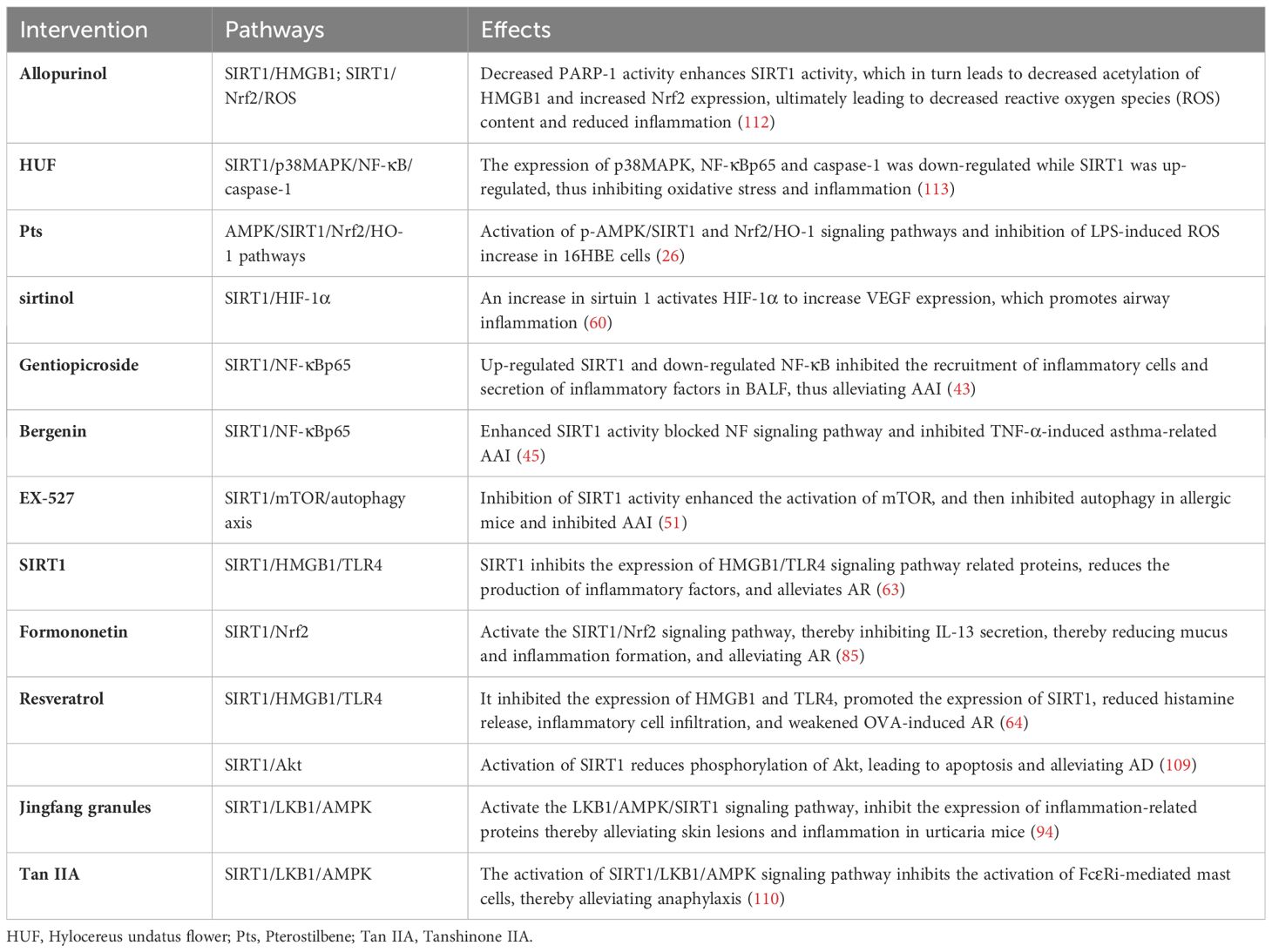
Table 1 Drug therapy targeting SIRT1.
SIRT1 is a lysine deacetylase that relies on NAD+ to function. Its impact on the secretion of inflammatory cytokines and regulation of immune cell differentiation is mediated through the deacetylation modification of transcription factors. In recent years, the role of SIRT1 in hypersensitivity has garnered significant attention, with the exact mechanism of hypersensitivity confirmed. Consequently, we shall delve into the relationship between SIRT1 and various hypersensitivity mechanisms. SIRT1 is recognized for its anti-inflammatory properties, which have been validated in multiple hypersensitivity reactions. In mouse models, certain SIRT1-targeting drugs have exhibited potential in mitigating diseases linked to hypersensitivity reactions. This presents a promising new therapeutic avenue for treating hypersensitivity inflammatory reactions. Nonetheless, the exact correlation between SIRT1 expression and the mechanism of hypersensitivity reaction remains the subject of ongoing research. Additionally, the suitability of SIRT1-targeting drugs for clinical treatment necessitates further confirmation.
YL: Writing – original draft. XT: Writing – review & editing. WW: Writing – review & editing. YJ: Writing – review & editing. SW: Conceptualization, Project administration, Supervision, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the Jiangsu Provincial Medical Key Discipline Cultivation Unit (Grant No. JSDW202241).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Yamamori T, Dericco J, Naqvi A, Hoffman TA, Mattagajasingh I, Kasuno K, et al. SIRT1 deacetylates APE1 and regulates cellular base excision repair. Nucleic Acids Res. (2010) 38:832–45. doi: 10.1093/nar/gkp1039
2. Huang Q, Su H, Qi B, Wang Y, Yan K, Wang X, et al. A SIRT1 activator, ginsenoside rc, promotes energy metabolism in cardiomyocytes and neurons. J Am Chem Soc. (2021) 143:1416–27. doi: 10.1021/jacs.0c10836
3. Li Y, Xu W, Mcburney MW, Longo VD. SirT1 inhibition reduces IGF-I/IRS-2/Ras/ERK1/2 signaling and protects neurons. Cell Metab. (2008) 8:38–48. doi: 10.1016/j.cmet.2008.05.004
4. Yang Y, Liu Y, Wang Y, Chao Y, Zhang J, Jia Y, et al. Regulation of SIRT1 and its roles in inflammation. Front Immunol. (2022) 13:831168. doi: 10.3389/fimmu.2022.831168
5. Feng Y, Liu T, Dong SY, Guo YJ, Jankovic J, Xu H, et al. Rotenone affects p53 transcriptional activity and apoptosis via targeting SIRT1 and H3K9 acetylation in SH-SY5Y cells. J neurochemistry. (2015) 134:668–76. doi: 10.1111/jnc.13172
6. Xu C, Wang L, Fozouni P, Evjen G, Chandra V, Jiang JP, et al. SIRT1 is downregulated by autophagy in senescence and ageing. Nat Cell Biol. (2020) 22:1170–9. doi: 10.1038/s41556-020-00579-5
7. Lee IH, Cao L, Mostoslavsky R, Lombard DB, Liu J, Bruns NE, et al. A role for the NAD-dependent deacetylase Sirt1 in the regulation of autophagy. Proc Natl Acad Sci U.S.A. (2008) 105:3374–9. doi: 10.1073/pnas.0712145105
8. Wu D, Shi Y, Zhang H, Miao C. Epigenetic mechanisms of Immune remodeling in sepsis: targeting histone modification. Cell Death Dis. (2023) 14:112. doi: 10.1038/s41419-023-05656-9
9. Chen GD, Yu WD, Chen XP. SirT1 activator represses the transcription of TNF−α in THP−1 cells of a sepsis model via deacetylation of H4K16. Mol Med Rep. (2016) 14:5544–50. doi: 10.3892/mmr.2016.5942
10. Ghare SS, Charpentier BT, Ghooray DT, Zhang J, Vadhanam MV, Reddy S, et al. Tributyrin mitigates ethanol-induced lysine acetylation of histone-H3 and p65-NFκB downregulating CCL2 expression and consequent liver inflammation and injury. Nutrients. (2023) 15(20):4397. doi: 10.3390/nu15204397
11. Liu K, Yang L, Wang P, Zhu J, Li F, Peng J, et al. Myricanol attenuates sepsis-induced inflammatory responses by nuclear factor erythroid 2-related factor 2 signaling and nuclear factor kappa B/mitogen-activated protein kinase pathway via upregulating Sirtuin 1. Inflammopharmacology. (2024) 32(3):1887–901. doi: 10.1007/s10787-024-01448-5
12. Ren JH, Tao Y, Zhang ZZ, Chen WX, Cai XF, Chen K, et al. Sirtuin 1 regulates hepatitis B virus transcription and replication by targeting transcription factor AP-1. J Virol. (2014) 88:2442–51. doi: 10.1128/JVI.02861-13
13. Li F, Chen Y, Li Y, Huang M, Zhao W. Geniposide alleviates diabetic nephropathy of mice through AMPK/SIRT1/NF-κB pathway. Eur J Pharmacol. (2020) 886:173449. doi: 10.1016/j.ejphar.2020.173449
14. Chen X, Chen C, Fan S, Wu S, Yang F, Fang Z, et al. Omega-3 polyunsaturated fatty acid attenuates the inflammatory response by modulating microglia polarization through SIRT1-mediated deacetylation of the HMGB1/NF-κB pathway following experimental traumatic brain injury. J Neuroinflamm. (2018) 15:116. doi: 10.1186/s12974-018-1151-3
15. He T, Bai X, Li Y, Zhang D, Xu Z, Yang X, et al. Insufficient SIRT1 in macrophages promotes oxidative stress and inflammation during scarring. J Mol Med (Berlin Germany). (2023) 101:1397–407. doi: 10.1007/s00109-023-02364-x
16. Srivastava RK, Sapra L, Bhardwaj A, Mishra PK, Verma B, Baig Z. Unravelling the immunobiology of innate lymphoid cells (ILCs): Implications in health and disease. Cytokine Growth factor Rev. (2023) 74:56–75. doi: 10.1016/j.cytogfr.2023.09.002
17. Papi A, Brightling C, Pedersen SE, Reddel HK. Asthma. Lancet (London England). (2018) 391:783–800. doi: 10.1016/S0140-6736(17)33311-1
18. Gandhi NA, Bennett BL, Graham NM, Pirozzi G, Stahl N, Yancopoulos GD. Targeting key proximal drivers of type 2 inflammation in disease. Nat Rev Drug Discov. (2016) 15:35–50. doi: 10.1038/nrd4624
19. Jirmo AC, Busse M, Happle C, Skuljec J, Dalüge K, Habener A, et al. IL-17 regulates DC migration to the peribronchial LNs and allergen presentation in experimental allergic asthma. Eur J Immunol. (2020) 50:1019–33. doi: 10.1002/eji.201948409
20. Hizawa N. The understanding of asthma pathogenesis in the era of precision medicine. Allergology international: Off J Japanese Soc Allergology. (2023) 72:3–10. doi: 10.1016/j.alit.2022.09.001
21. Shah PA, Brightling C. Biologics for severe asthma-Which, when and why? Respirology (Carlton Vic). (2023) 28:709–21. doi: 10.1111/resp.14520
22. Casaro M, Souza VR, Oliveira FA, Ferreira CM. OVA-induced allergic airway inflammation mouse model. Methods Mol Biol (Clifton NJ). (2019) 1916:297–301. doi: 10.1007/978-1-4939-8994-2_28
23. Komlósi ZI, VAN DE VEEN W, Kovács N, Szűcs G, Sokolowska M, O'Mahony L, et al. Cellular and molecular mechanisms of allergic asthma. Mol aspects Med. (2022) 85:100995. doi: 10.1016/j.mam.2021.100995
24. Wang Y, Li D, Ma G, Li W, Wu J, Lai T, et al. Increases in peripheral SIRT1: a new biological characteristic of asthma. Respirology (Carlton Vic). (2015) 20:1066–72. doi: 10.1111/resp.12558
25. Song X, Li B, Wang H, Zou X, Gao R, Zhang W, et al. Asthma alleviates obesity in males through regulating metabolism and energy expenditure. Biochim Biophys Acta Mol basis Dis. (2019) 1865:350–9. doi: 10.1016/j.bbadis.2018.10.001
26. Xu C, Song Y, Wang Z, Jiang J, Piao Y, Li L, et al. Pterostilbene suppresses oxidative stress and allergic airway inflammation through AMPK/Sirt1 and Nrf2/HO-1 pathways. Immunity Inflammation Dis. (2021) 9:1406–17. doi: 10.1002/iid3.490
27. Tang L, Chen Q, Meng Z, Sun L, Zhu L, Liu J, et al. Suppression of sirtuin-1 increases IL-6 expression by activation of the akt pathway during allergic asthma. Cell Physiol biochemistry: Int J Exp Cell physiology biochemistry Pharmacol. (2017) 43:1950–60. doi: 10.1159/000484119
28. Mohamed RA, Fakhr AE, Baioumy SA. Investigating forkhead box O transcription factor 1 gene's relation to immunoglobulin E in house dust mite-allergic asthma patients. Adv Respir Med. (2023) 91:532–45. doi: 10.3390/arm91060039
29. Mohamed RA, Fakhr AE, Baioumy SA. Investigating the T regulatory cells and Sirtuin-I serum level in immunotherapy treated house dust mite allergic asthma patients. J asthma: Off J Assoc Care Asthma. (2024) 61:348–59. doi: 10.1080/02770903.2023.2272813
30. Wei W, Liu Y, Wang Y, Zhao Y, He J, Li X, et al. Induction of CD4+CD25+Foxp3+IL-10+ T cells in HDM-allergic asthmatic children with or without SIT. Int Arch Allergy Immunol. (2010) 153:19–26. doi: 10.1159/000301575
31. Lai T, Su G, Wu D, Chen Z, Chen Y, Yi H, et al. Myeloid-specific SIRT1 deletion exacerbates airway inflammatory response in a mouse model of allergic asthma. Aging. (2021) 13:15479–90. doi: 10.18632/aging.v13i11
32. Yunna C, Mengru H, Lei W, Weidong C. Macrophage M1/M2 polarization. Eur J Pharmacol. (2020) 877:173090. doi: 10.1016/j.ejphar.2020.173090
33. Huang Q, Weng D, Yao S, Shen H, Gao S, Zhang Y, et al. Progranulin deficiency suppresses allergic asthma and enhances efferocytosis via PPAR-γ/MFG-E8 regulation in macrophages. Immunity Inflammation Dis. (2023) 11:e779. doi: 10.1002/iid3.779
34. Zhou W, Nie X. Afzelin attenuates asthma phenotypes by downregulation of GATA3 in a murine model of asthma. Mol Med Rep. (2015) 12:71–6. doi: 10.3892/mmr.2015.3391
35. Murga-Zamalloa C, Wilcox RA. GATA-3 in T-cell lymphoproliferative disorders. IUBMB Life. (2020) 72:170–7. doi: 10.1002/iub.2130
36. Colley T, Mercado N, Kunori Y, Brightling C, Bhavsar PK, Barnes PJ, et al. Defective sirtuin-1 increases IL-4 expression through acetylation of GATA-3 in patients with severe asthma. J Allergy Clin Immunol. (2016) 137:1595–7.e7. doi: 10.1016/j.jaci.2015.10.013
37. Bui TT, Piao CH, Song CH, Shin HS, Chai OH. Bupleurum chinense extract ameliorates an OVA-induced murine allergic asthma through the reduction of the Th2 and Th17 cytokines production by inactivation of NFκB pathway. Biomedicine pharmacotherapy = Biomedecine pharmacotherapie. (2017) 91:1085–95. doi: 10.1016/j.biopha.2017.04.133
38. Gu X, Zhang Q, Du Q, Shen H, Zhu Z. Pinocembrin attenuates allergic airway inflammation via inhibition of NF-κB pathway in mice. Int Immunopharmacol. (2017) 53:90–5. doi: 10.1016/j.intimp.2017.10.005
39. Huang M, Wei Y, Dong J. Epimedin C modulates the balance between Th9 cells and Treg cells through negative regulation of noncanonical NF-κB pathway and MAPKs activation to inhibit airway inflammation in the ovalbumin-induced murine asthma model. Pulmonary Pharmacol Ther. (2020) 65:102005. doi: 10.1016/j.pupt.2021.102005
40. Piao CH, Song CH, Lee EJ, Chai OH. Saikosaponin A ameliorates nasal inflammation by suppressing IL-6/ROR-γt/STAT3/IL-17/NF-κB pathway in OVA-induced allergic rhinitis. Chemico-biological Interact. (2020) 315:108874. doi: 10.1016/j.cbi.2019.108874
41. Ma SQ, Wei HL, Zhang X. TLR2 regulates allergic airway inflammation through NF-κB and MAPK signaling pathways in asthmatic mice. Eur Rev Med Pharmacol Sci. (2018) 22:3138–46. doi: 10.26355/eurrev_201805_15073
42. Liu Y, Zhang M, Zhang H, Qian X, Luo L, He Z. Anthocyanins inhibit airway inflammation by downregulating the NF-κB pathway via the miR-138-5p/SIRT1 axis in asthmatic mice. Int Arch Allergy Immunol. (2022) 183:539–51. doi: 10.1159/000520645
43. Zou B, Fu Y, Cao C, Pan D, Wang W, Kong L. Gentiopicroside ameliorates ovalbumin-induced airway inflammation in a mouse model of allergic asthma via regulating SIRT1/NF-κB signaling pathway. Pulmonary Pharmacol Ther. (2021) 68:102034. doi: 10.1016/j.pupt.2021.102034
44. Chen M, Chen Z, Huang D, Sun C, Xie J, Chen T, et al. Myricetin inhibits TNF-α-induced inflammation in A549 cells via the SIRT1/NF-κB pathway. Pulmonary Pharmacol Ther. (2020) 65:102000. doi: 10.1016/j.pupt.2021.102000
45. Chen M, Chen C, Gao Y, Li D, Huang D, Chen Z, et al. Bergenin-activated SIRT1 inhibits TNF-α-induced proinflammatory response by blocking the NF-κB signaling pathway. Pulmonary Pharmacol Ther. (2020) 62:101921. doi: 10.1016/j.pupt.2020.101921
46. Rane S, He M, Sayed D, Vashistha H, Malhotra A, Sadoshima J, et al. Downregulation of miR-199a derepresses hypoxia-inducible factor-1alpha and Sirtuin 1 and recapitulates hypoxia preconditioning in cardiac myocytes. Circ Res. (2009) 104:879–86. doi: 10.1161/CIRCRESAHA.108.193102
47. Lin J, Huo X, Liu X. "mTOR signaling pathway": A potential target of curcumin in the treatment of spinal cord injury. BioMed Res Int. (2017) 2017:1634801. doi: 10.1155/2017/1634801
48. Kudrna K, Staab EB, Eilers E, Thomes P, Maurya S, Brody SL, et al. mTOR signaling regulates aberrant epithelial cell proliferative and migratory behaviors characteristic of airway mucous metaplasia in asthma. bioRxiv: preprint server Biol. (2024). doi: 10.1101/2024.02.12.579905
49. Zou H, Wang LX, Wang M, Cheng C, Li S, Shen Q, et al. MTOR-mediated autophagy is involved in the protective effect of ketamine on allergic airway inflammation. J Immunol Res. (2019) 2019:5879714. doi: 10.1155/2019/5879714
50. Farooq MB, Walsh GM. Autophagy and asthma. Clin Exp allergy: J Br Soc Allergy Clin Immunol. (2016) 46:7–9. doi: 10.1111/cea.12633
51. Wu Y, Li W, Hu Y, Liu Y, Sun X. Suppression of sirtuin 1 alleviates airway inflammation through mTOR−mediated autophagy. Mol Med Rep. (2020) 22:2219–26. doi: 10.3892/mmr
52. Cheng SC, Quintin J, Cramer RA, Shepardson KM, Saeed S, Kumar V, et al. mTOR- and HIF-1α-mediated aerobic glycolysis as metabolic basis for trained immunity. Sci (New York NY). (2014) 345:1250684. doi: 10.1126/science.1250684
53. Cramer T, Yamanishi Y, Clausen BE, Förster I, Pawlinski R, Mackman N, et al. HIF-1alpha is essential for myeloid cell-mediated inflammation. Cell. (2003) 112:645–57. doi: 10.1016/S0092-8674(03)00154-5
54. Dang EV, Barbi J, Yang HY, Jinasena D, Yu H, Zheng Y, et al. Control of T(H)17/T(reg) balance by hypoxia-inducible factor 1. Cell. (2011) 146:772–84. doi: 10.1016/j.cell.2011.07.033
55. Shi LZ, Wang R, Huang G, Vogel P, Neale G, Green DR, et al. HIF1alpha-dependent glycolytic pathway orchestrates a metabolic checkpoint for the differentiation of TH17 and Treg cells. J Exp Med. (2011) 208:1367–76. doi: 10.1084/jem.20110278
56. Joo HY, Yun M, Jeong J, Park ER, Shin HJ, Woo SR, et al. SIRT1 deacetylates and stabilizes hypoxia-inducible factor-1α (HIF-1α) via direct interactions during hypoxia. Biochem Biophys Res Commun. (2015) 462:294–300. doi: 10.1016/j.bbrc.2015.04.119
57. Lim JH, Lee YM, Chun YS, Chen J, Kim JE, Park JW. Sirtuin 1 modulates cellular responses to hypoxia by deacetylating hypoxia-inducible factor 1alpha. Mol Cell. (2010) 38:864–78. doi: 10.1016/j.molcel.2010.05.023
58. Yu Q, Dong L, Li Y, Liu G. SIRT1 and HIF1α signaling in metabolism and immune responses. Cancer Lett. (2018) 418:20–6. doi: 10.1016/j.canlet.2017.12.035
59. Wang Y, Bi Y, Chen X, Li C, Li Y, Zhang Z, et al. Histone deacetylase SIRT1 negatively regulates the differentiation of interleukin-9-producing CD4(+) T cells. Immunity. (2016) 44:1337–49. doi: 10.1016/j.immuni.2016.05.009
60. Kim SR, Lee KS, Park SJ, Min KH, Choe YH, Moon H, et al. Involvement of sirtuin 1 in airway inflammation and hyperresponsiveness of allergic airway disease. J Allergy Clin Immunol. (2010) 125:449–60.e14. doi: 10.1016/j.jaci.2009.08.009
61. Miller BC, Sen DR, Al Abosy R, Bi K, Virkud YV, LaFleur MW, et al. Sirtuins as novel targets in the pathogenesis of airway inflammation in bronchial asthma. Eur J Pharmacol. (2019) 865:172670. doi: 10.1016/j.ejphar.2019.172670
62. Legutko A, Marichal T, Fiévez L, Bedoret D, Mayer A, de Vries H, et al. Sirtuin 1 promotes Th2 responses and airway allergy by repressing peroxisome proliferator-activated receptor-γ activity in dendritic cells. J Immunol (Baltimore Md: 1950). (2011) 187:4517–29. doi: 10.4049/jimmunol.1101493
63. Yuan Y, Liu Q, Zhao J, Tang H, Sun J. SIRT1 attenuates murine allergic rhinitis by downregulated HMGB 1/TLR4 pathway. Scandinavian J Immunol. (2018) 87:e12667. doi: 10.1111/sji.12667
64. Sekine T, Perez-Potti A, Nguyen S, Gorin JB, Wu VH, Gostick E, et al. Resveratrol-mediated SIRT1 activation attenuates ovalbumin-induced allergic rhinitis in mice. Mol Immunol. (2020) 122:156–62. doi: 10.1016/j.molimm.2020.04.009
65. Tang H, Han X, Li T, Feng Y, Sun J. Protective effect of miR-138-5p inhibition modified human mesenchymal stem cell on ovalbumin-induced allergic rhinitis and asthma syndrome. J Cell Mol Med. (2021) 25:5038–49. doi: 10.1111/jcmm.16473
66. Wang X, Liu C, Wu L, Zhu S. Potent ameliorating effect of Hypoxia-inducible factor 1α (HIF-1α) antagonist YC-1 on combined allergic rhinitis and asthma syndrome (CARAS) in Rats. Eur J Pharmacol. (2016) 788:343–50. doi: 10.1016/j.ejphar.2016.07.040
67. Zhou H, Chen X, Zhang WM, Zhu LP, Cheng L. HIF-1α inhibition reduces nasal inflammation in a murine allergic rhinitis model. PLoS One. (2012) 7:e48618. doi: 10.1371/journal.pone.0048618
68. Gao Y, Xu X, Feng J, Ma Y, Zheng D, Meng Y, et al. Effects of interleukin-1 receptor-associated kinase 1 RNA interference in dendritic cells on inflammatory cytokine release and T-cell proliferation. Mol Med Rep. (2016) 14:5685–92. doi: 10.3892/mmr.2016.5946
69. Liu G, Bi Y, Xue L, Zhang Y, Yang H, Chen X, et al. Dendritic cell SIRT1-HIF1α axis programs the differentiation of CD4+ T cells through IL-12 and TGF-β1. Proc Natl Acad Sci United States America. (2015) 112:E957–65. doi: 10.1073/pnas.1420419112
70. Hu T, Fan X, Ma L, Liu J, Chang Y, Yang P, et al. TIM4-TIM1 interaction modulates Th2 pattern inflammation through enhancing SIRT1 expression. Int J Mol Med. (2017) 40:1504–10. doi: 10.3892/ijmm.2017.3150
71. Niu Y, Wang J, Li Z, Yao K, Wang L, Song J. HIF1α Deficiency in dendritic cells attenuates symptoms and inflammatory indicators of allergic rhinitis in a SIRT1-dependent manner. Int Arch Allergy Immunol. (2020) 181:585–93. doi: 10.1159/000506862
72. Robinson DS. Regulatory T cells and asthma. Clin Exp allergy: J Br Soc Allergy Clin Immunol. (2009) 39:1314–23. doi: 10.1111/j.1365-2222.2009.03301.x
73. Lewkowich IP, Herman NS, Schleifer KW, Dance MP, Chen BL, Dienger KM, et al. CD4+CD25+ T cells protect against experimentally induced asthma and alter pulmonary dendritic cell phenotype and function. J Exp Med. (2005) 202:1549–61. doi: 10.1084/jem.20051506
74. Kearley J, Barker JE, Robinson DS, Lloyd CM. Resolution of airway inflammation and hyperreactivity after in vivo transfer of CD4+CD25+ regulatory T cells is interleukin 10 dependent. J Exp Med. (2005) 202:1539–47. doi: 10.1084/jem.20051166
75. Deng G, Song X, Fujimoto S, Piccirillo CA, Nagai Y, Greene MI. Foxp3 post-translational modifications and treg suppressive activity. Front Immunol. (2019) 10:2486. doi: 10.3389/fimmu.2019.02486
76. Beier UH, Wang L, Bhatti TR, Liu Y, Han R, Ge G, et al. Sirtuin-1 targeting promotes Foxp3+ T-regulatory cell function and prolongs allograft survival. Mol Cell Biol. (2011) 31:1022–9. doi: 10.1128/MCB.01206-10
77. Sobah ML, Liongue C, Ward AC. SOCS proteins in immunity, inflammatory diseases, and immune-related cancer. Front Med. (2021) 8:727987. doi: 10.3389/fmed.2021.727987
78. Zeng Q, Liu W, Luo R, Lu G. MicroRNA-181a and microRNA-155 are involved in the regulation of the differentiation and function of regulatory T cells in allergic rhinitis children. Pediatr Allergy immunology: Off Publ Eur Soc Pediatr Allergy Immunol. (2019) 30:434–42. doi: 10.1111/pai.13038
79. Marcel N, Perumalsamy LR, Shukla SK, Sarin A. The lysine deacetylase Sirtuin 1 modulates the localization and function of the Notch1 receptor in regulatory T cells. Sci Signaling. (2017) 10(473):eaah4679. doi: 10.1126/scisignal.aah4679
80. Rochman M, Kartashov AV, Caldwell JM, Collins MH, Stucke EM, Kc K, et al. Neurotrophic tyrosine kinase receptor 1 is a direct transcriptional and epigenetic target of IL-13 involved in allergic inflammation. Mucosal Immunol. (2015) 8:785–98. doi: 10.1038/mi.2014.109
81. Xu H, Wang L, Chen H, Cai H. HDAC4 depletion ameliorates IL-13-triggered inflammatory response and mucus production in nasal epithelial cells via activation of SIRT1/NF-κB signaling. Immunity Inflammation Dis. (2022) 10:e692. doi: 10.1002/iid3.692
82. Jung KA, Kwak MK. The Nrf2 system as a potential target for the development of indirect antioxidants. Molecules (Basel Switzerland). (2010) 15:7266–91. doi: 10.3390/molecules15107266
83. Bui TT, Fan Y, Piao CH, Nguyen TV, Shin DU, Jung SY, et al. Piper Nigrum extract improves OVA-induced nasal epithelial barrier dysfunction via activating Nrf2/HO-1 signaling. Cell Immunol. (2020) 351:104035. doi: 10.1016/j.cellimm.2019.104035
84. Van Nguyen T, Piao CH, Fan YJ, Shin DU, Kim SY, Song HJ, et al. Anti-allergic rhinitis activity of α-lipoic acid via balancing Th17/Treg expression and enhancing Nrf2/HO-1 pathway signaling. Sci Rep. (2020) 10:12528. doi: 10.1038/s41598-020-69234-1
85. Huang J, Chen X, Xie A. Formononetin ameliorates IL−13−induced inflammation and mucus formation in human nasal epithelial cells by activating the SIRT1/Nrf2 signaling pathway. Mol Med Rep. (2021) 24(6):832. doi: 10.3892/mmr
86. Kauppinen A, Suuronen T, Ojala J, Kaarniranta K, Salminen A. Antagonistic crosstalk between NF-κB and SIRT1 in the regulation of inflammation and metabolic disorders. Cell signalling. (2013) 25:1939–48. doi: 10.1016/j.cellsig.2013.06.007
87. Radonjic-Hoesli S, Hofmeier KS, Micaletto S, Schmid-Grendelmeier P, Bircher A, Simon D. Urticaria and angioedema: an update on classification and pathogenesis. Clin Rev Allergy Immunol. (2018) 54:88–101. doi: 10.1007/s12016-017-8628-1
88. Zuberbier T, Aberer W, Asero R, Abdul Latiff AH, Baker D, Ballmer-Weber B, et al. The EAACI/GA²LEN/EDF/WAO guideline for the definition, classification, diagnosis and management of urticaria. Allergy. (2018) 73:1393–414. doi: 10.1111/all.13397
89. Huilan Z, Runxiang L, Bihua L, Qing G. Role of the subgroups of T, B, natural killer lymphocyte and serum levels of interleukin-15, interleukin-21 and immunoglobulin E in the pathogenesis of urticaria. J Dermatol. (2010) 37:441–7. doi: 10.1111/j.1346-8138.2010.00805.x
90. Chen Q, Zhong H, Chen WC, Zhai Z, Zhou Z, Song Z, et al. Different expression patterns of plasma Th1-, Th2-, Th17- and Th22-related cytokines correlate with serum autoreactivity and allergen sensitivity in chronic spontaneous urticaria. J Eur Acad Dermatol Venereology: JEADV. (2018) 32:441–8. doi: 10.1111/jdv.14541
91. Garrington TP, Ishizuka T, Papst PJ, Chayama K, Webb S, Yujiri T, et al. MEKK2 gene disruption causes loss of cytokine production in response to IgE and c-Kit ligand stimulation of ES cell-derived mast cells. EMBO J. (2000) 19:5387–95. doi: 10.1093/emboj/19.20.5387
92. Chayama K, Papst PJ, Garrington TP, Pratt JC, Ishizuka T, Webb S, et al. Role of MEKK2-MEK5 in the regulation of TNF-alpha gene expression and MEKK2-MKK7 in the activation of c-Jun N-terminal kinase in mast cells. Proc Natl Acad Sci United States America. (2001) 98:4599–604. doi: 10.1073/pnas.081021898
93. Li G, Lucas JJ, Gelfand EW. Protein kinase C alpha, betaI, and betaII isozymes regulate cytokine production in mast cells through MEKK2/ERK5-dependent and -independent pathways. Cell Immunol. (2005) 238:10–8. doi: 10.1016/j.cellimm.2005.12.001
94. Sun C, Liang H, Zhao Y, Li S, Li X, Yuan X, et al. Jingfang Granules improve glucose metabolism disturbance and inflammation in mice with urticaria by up-regulating LKB1/AMPK/SIRT1 axis. J Ethnopharmacol. (2023) 302:115913. doi: 10.1016/j.jep.2022.115913
95. Jiao Y, Zhang T, Zhang C, Ji H, Tong X, Xia R, et al. Exosomal miR-30d-5p of neutrophils induces M1 macrophage polarization and primes macrophage pyroptosis in sepsis-related acute lung injury. Crit Care (London England). (2021) 25:356. doi: 10.1186/s13054-021-03775-3
96. Zhao W, Ma L, Cai C, Gong X. Caffeine inhibits NLRP3 inflammasome activation by suppressing MAPK/NF-κB and A2aR signaling in LPS-induced THP-1 macrophages. Int J Biol Sci. (2019) 15:1571–81. doi: 10.7150/ijbs.34211
97. Assrawi E, Louvrier C, Lepelletier C, Georgin-Lavialle S, Bouaziz JD, Awad F, et al. Somatic mosaic NLRP3 mutations and inflammasome activation in late-onset chronic urticaria. J Invest Dermatol. (2020) 140:791–8.e2. doi: 10.1016/j.jid.2019.06.153
98. Tufekci KU, Eltutan BI, Isci KB, Genc S. Resveratrol inhibits NLRP3 inflammasome-induced pyroptosis and miR-155 expression in microglia through sirt1/AMPK pathway. Neurotoxicity Res. (2021) 39:1812–29. doi: 10.1007/s12640-021-00435-w
99. Hu Y, Zhao M, Zhao T, Qi M, Yao G, Dong Y. The protective effect of pilose antler peptide on CUMS-induced depression through AMPK/sirt1/NF-κB/NLRP3-mediated pyroptosis. Front Pharmacol. (2022) 13:815413. doi: 10.3389/fphar.2022.815413
100. Chen L, Lan Z. Polydatin attenuates potassium oxonate-induced hyperuricemia and kidney inflammation by inhibiting NF-κB/NLRP3 inflammasome activation via the AMPK/SIRT1 pathway. Food Funct. (2017) 8:1785–92. doi: 10.1039/C6FO01561A
101. Nomura T, Kabashima K. Advances in atopic dermatitis in 2015. J Allergy Clin Immunol. (2016) 138:1548–55. doi: 10.1016/j.jaci.2016.10.004
102. Marko M, Pawliczak R. Resveratrol and its derivatives in inflammatory skin disorders-atopic dermatitis and psoriasis: A review. Antioxidants (Basel Switzerland). (2023) 12(11):1954. doi: 10.3390/antiox12111954
103. Schäbitz A, Eyerich K, Garzorz-Stark N. So close, and yet so far away: The dichotomy of the specific immune response and inflammation in psoriasis and atopic dermatitis. J Internal Med. (2021) 290:27–39. doi: 10.1111/joim.13235
104. Shin JW, Choi HR, Nam KM, Yang SH, Kim SA, Joe HJ, et al. The expression of epidermal stem cell marker and SIRT1 in atopic dermatitis: A discussion of regenerative potential. Ann Dermatol. (2019) 31:476–8. doi: 10.5021/ad.2019.31.4.476
105. Ming M, Zhao B, Shea CR, Shah P, Qiang L, White SR, et al. Loss of sirtuin 1 (SIRT1) disrupts skin barrier integrity and sensitizes mice to epicutaneous allergen challenge. J Allergy Clin Immunol. (2015) 135:936–45.e4. doi: 10.1016/j.jaci.2014.09.035
106. Nakai K, Yoneda K, Hosokawa Y, Moriue T, Presland RB, Fallon PG, et al. Reduced expression of epidermal growth factor receptor, E-cadherin, and occludin in the skin of flaky tail mice is due to filaggrin and loricrin deficiencies. Am J Pathol. (2012) 181:969–77. doi: 10.1016/j.ajpath.2012.06.005
107. Xu F, Xu J, Xiong X, Deng Y. Salidroside inhibits MAPK. NF-κB, and STAT3 pathways in psoriasis-associated oxidative stress via SIRT1 activation. Redox report: Commun Free Radical Res. (2019) 24:70–4. doi: 10.1080/13510002.2019.1658377
108. Sestito R, Madonna S, Scarponi C, Cianfarani F, Failla CM, Cavani A, et al. STAT3-dependent effects of IL-22 in human keratinocytes are counterregulated by sirtuin 1 through a direct inhibition of STAT3 acetylation. FASEB journal: Off Publ Fed Am Societies Exp Biol. (2011) 25:916–27. doi: 10.1096/fj.10-172288
109. Lee JH, Kim JS, Park SY, Lee YJ. Resveratrol induces human keratinocyte damage via the activation of class III histone deacetylase, Sirt1. Oncol Rep. (2016) 35:524–9. doi: 10.3892/or.2015.4332
110. Li X, Park SJ, Jin F, Deng Y, Yang JH, Chang JH, et al. Tanshinone IIA suppresses FcϵRI-mediated mast cell signaling and anaphylaxis by activation of the Sirt1/LKB1/AMPK pathway. Biochem Pharmacol. (2018) 152:362–72. doi: 10.1016/j.bcp.2018.04.015
111. Li X, Lee YJ, Jin F, Park YN, Deng Y, Kang Y, et al. Sirt1 negatively regulates FcϵRI-mediated mast cell activation through AMPK- and PTP1B-dependent processes. Sci Rep. (2017) 7:6444. doi: 10.1038/s41598-017-06835-3
112. Wang Y, Le Y, Wu J, Zhao W, Zhang Q, Xu G, et al. Inhibition of xanthine oxidase by allopurinol suppresses HMGB1 secretion and ameliorates experimental asthma. Redox Biol. (2024) 70:103021. doi: 10.1016/j.redox.2023.103021
113. Liao W, Liu W, Yan Y, Li L, Tong J, Huang Y, et al. Hylocereus undatus flower extract suppresses OVA-induced allergic asthma in BALb/c mice by reducing airway inflammation and modulating gut microbiota. Biomedicine pharmacotherapy = Biomedecine pharmacotherapie. (2022) 153:113476. doi: 10.1016/j.biopha.2022.113476
Keywords: silent information regulator sirtuin 1 (SIRT1), deacetylase, allergic diseases, acetylation, hypersensitivity
Citation: Lu Y, Tang X, Wang W, Yang J and Wang S (2024) The role of deacetylase SIRT1 in allergic diseases. Front. Immunol. 15:1422541. doi: 10.3389/fimmu.2024.1422541
Received: 24 April 2024; Accepted: 02 July 2024;
Published: 16 July 2024.
Edited by:
Seyyed Shamsadin Athari, Zanjan University of Medical Sciences, IranReviewed by:
Yohei Sato, Jikei University School of Medicine, JapanCopyright © 2024 Lu, Tang, Wang, Yang and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shengjun Wang, c2p3anNAdWpzLmVkdS5jbg==; Jun Yang, MzQzMzIxNTg2QHFxLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.