 José Inciarte-Mundo1
José Inciarte-Mundo1 Beatriz Frade-Sosa
Beatriz Frade-Sosa Raimon Sanmartí
Raimon Sanmartí
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol. , 03 November 2022
Sec. Autoimmune and Autoinflammatory Disorders : Autoimmune Disorders
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.1001025
S100A9/S100A8 (calprotectin), a member of the S100 protein family, has been shown to play a pivotal role in innate immunity activation. Calprotectin plays a critical role in the pathogenesis of rheumatoid arthritis (RA), as it triggers chemotaxis, phagocyte migration and modulation of neutrophils and macrophages. Higher calprotectin levels have been found in synovial fluid, plasma, and serum from RA patients. Recent studies have demonstrated better correlations between serum or plasma calprotectin and composite inflammatory disease activity indexes than c-reactive protein (CRP) or the erythrocyte sedimentation rate (ESR). Calprotectin serum levels decreased after treatment, independently of the DMARD type or strategy. Calprotectin has shown the strongest correlations with other sensitive techniques to detect inflammation, such as ultrasound. Calprotectin independently predicts radiographic progression. However, its value as a biomarker of treatment response and flare after tapering is unclear. This update reviews the current understanding of calprotectin in RA and discusses possible applications as a biomarker in clinical practice.
Rheumatoid arthritis (RA) is a heterogeneous disease of unknown origin, characterized by chronic polyarthritis that may lead to joint destruction, disability, and increased mortality. Extraarticular manifestations are not uncommon. RA affects 0.5-1% of the adult population, predominantly females. Genetic and environmental factors have been implicated in the susceptibility to RA. Autoimmunity, with the presence of characteristic autoantibodies such as rheumatoid factor or anticitrullinated peptide autoantibodies, are implicated in the pathogenesis of RA, although recent data confirms the role of the innate immune system in this disease (1).
The innate immune system plays a central role in initiating local inflammation, contributing to the pathogenesis of RA by promoting the production of inflammatory cytokines and chemokines. Pattern recognition receptors (PRRs) are a family of receptors of the innate immune system that bind to damage-associated molecular pattern molecules (DAMP) (2). The most important PRRs are Toll-like receptors (TLRs), which allow the activation of monocytes, neutrophils, dendritic cells, natural killer (NK) cells and B cells (3).
S100A9/S100A8 (calprotectin) a member of the S100 protein family has been studied as an important proinflammatory factor of innate immunity as an endogenous DAMP via TLR4 activation. Calprotectin plays a critical role in the development of inflammation loops in RA as a trigger for chemotaxis, phagocyte migration and modulation of various macrophage functions (4–6).
This update reviews the current understanding of calprotectin in RA and discusses possible applications as a biomarker in clinical practice.
The first members of the S100 protein family were purified from bovine brain in the early 1980s. The protein complex was denominated “S100” because of its 100% solubility in ammonium sulphate solution (7). The S100 protein family is specifically linked to innate immune functions by their expression in cells of myeloid origin.
The S100 protein family is widely expressed, although they are not ubiquitous, and several have highly restricted distributions. The functions of these proteins vary widely between individual members, functioning as both intracellular and extracellular signaling molecules. S100 protein functions include cytoskeletal function, homeostasis, tumor-suppression, antimicrobial response, chemotactic activity, atherogenesis and protection from oxidative cell damage in brain tissue. The main tissue cell locations and functions are summarized in Table 1.
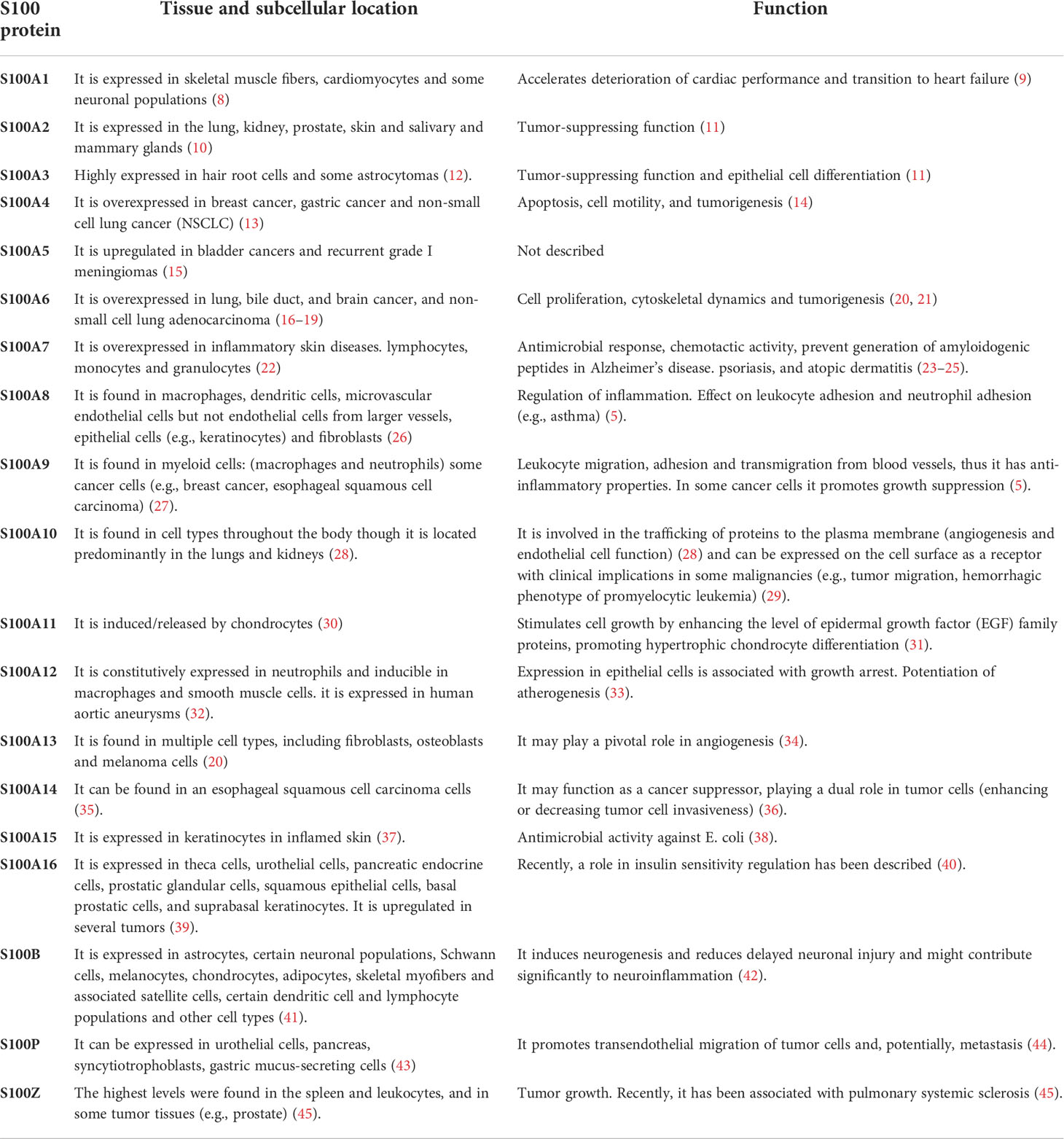
Table 1 S100 protein family. Main tissue cell localization and function of S100 protein family.
Among the S100 protein family, calprotectin (S100A8/S100A9) is primarily expressed in innate immune cells, particularly in neutrophils, monocytes, and macrophages, which constitute approximately 40% of the cytosolic proteins in these cells. However, under specific stimuli, calprotectin may be expressed in other cell lines, such as osteoclasts and keratinocytes (46–48). The gene encoding calprotectin subunits is located in the gene cluster on human chromosome 1q12-1q21 (49, 50).
Although calprotectin has been recognized for almost 40 years, it has only recently become of interest as a biomarker of disease activity and damage in rheumatology and was previously known as L1 protein (51), cystic fibrosis-associated antigen (CFA) (52), calgranulins A and B (53), S-1OOa and b (54) or myeloid related protein 8 and 14 (MRP 8/14) (48). Calprotectin is a well-established biomarker in other medical areas, such as calprotectin in faeces in inflammatory bowel disease (55–59).
Calprotectin is a calcium- and zinc-binding heterodimeric molecule of 36.5 kDa, with two heavy and one light chain non-covalently linked (51). It has two subunits, S100A8 and S100A9, which are 8.3 kDa and 13.3 kDa, respectively (60). S100A8 is the active subunit and S100A9 acts as the regulatory subunit, preventing early degradation of S100A8 (61). The EF-hand is composed of two α-helices flanking a central calcium-binding loop, resulting in classical helix-loop-helix domains (62). Calprotectin is mostly found in the form of heterodimers (S100A9-S100A8) and tetramers (S100A9-S100A8)2 in a calcium dependent-manner (63).
Two independent calprotectin activation pathways have been proposed to explain calprotectin release to extracellular compartments. The first, a canonical pathway, includes protein kinase C, which is induced by different inflammatory stimuli (e.g., bacteria). The second, a non-classical secretion avoiding the Golgi-associated pathway, requires the elevation of intracellular calcium levels, induced by contact between phagocytes and pre-activated endothelial cells by tumor necrosis factor (TNF), resulting in active secretion of calprotectin by phagocytes (63). Recently, a new activation mechanism has been proposed, namely activation via chromatin in neutrophil extracellular traps (NETs) (64). In addition, calprotectin may be secreted passively from apoptotic cells (61).
Calprotectin complexes are known to interact with cytoskeletal components such as actin filaments, keratin, vimentin, and microtubules in a calcium-dependent manner. High calcium concentrations induce a rearrangement of calprotectin into tetramers, allowing translocation to the cell membrane and tubulin polymerization (61). This process is regulated by the phosphorylation of the threonine at position 113 in S100A9 by p38 MAPK (Figure 1).
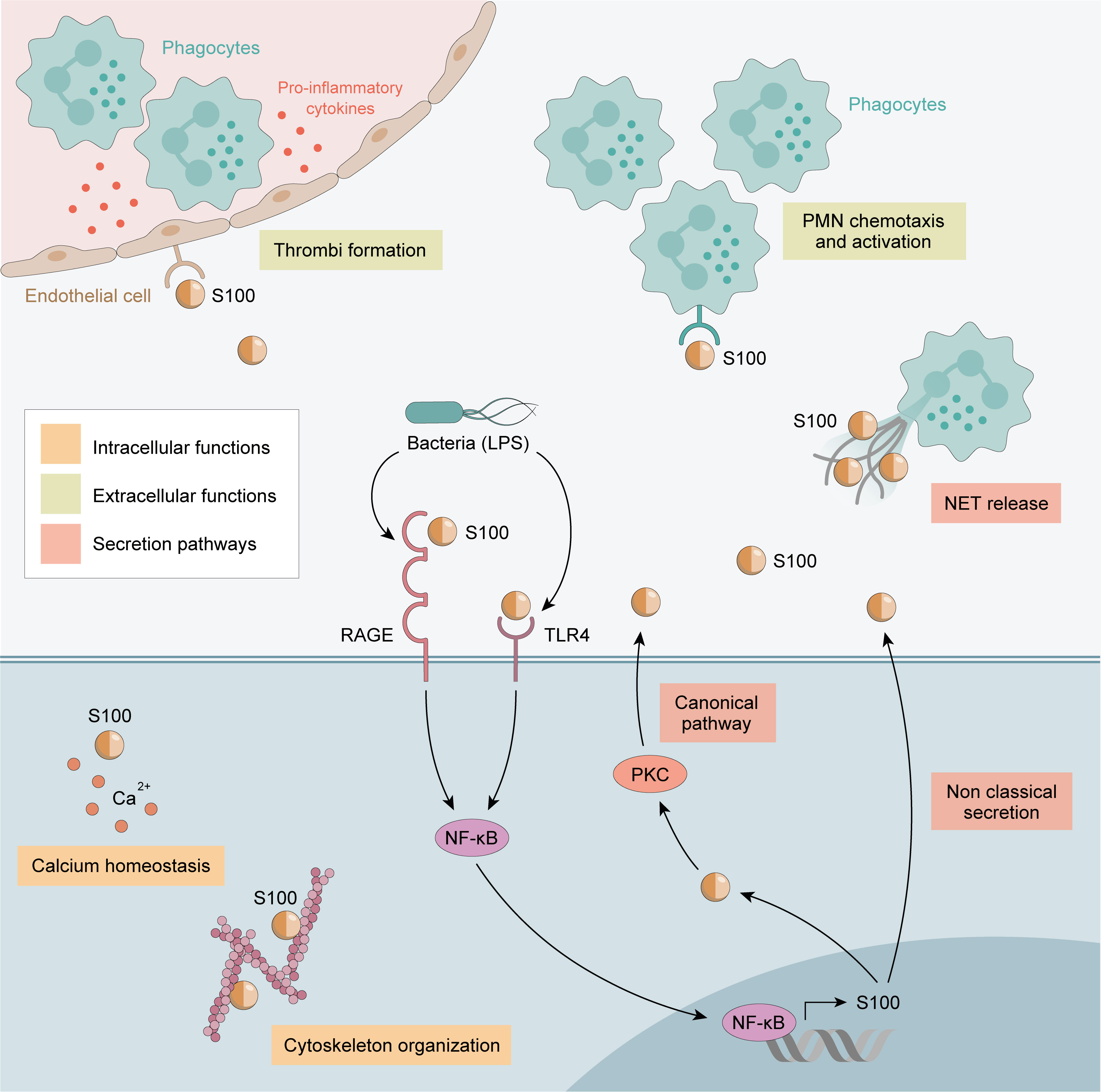
Figure 1 Calprotectin Functions. Intracellular functions are shown in orange and extracellular functions in green. Calprotectin Intracellular functions includes cytoskeleton cell migration and calcium homeostasis. Extracellular functions involve endothelial cells activation, promoting the adhesion of phagocytes to the vascular endothelium and thrombi formation. Also increase chemotaxis and the activation of PMN. Finally, calprotectin exerts a strong antimicrobial action against a variety of bacterial and fungal pathogens.
Extracellular calprotectin complexes interact with endothelial cells by binding to heparan sulfate and, specifically, carboxylated glycans, and up-regulate integrin receptors on leukocytes, resulting in the activation of endothelial cells. Activated endothelial cells express a pro-inflammatory cytokine profile (e.g., IL-1, IL-8 MCP-1), and thus calprotectin plays a central role in promoting the adhesion of phagocytes to the vascular endothelium and thrombi formation (65, 66). Calprotectin generates a positive feedback loop, increasing chemotaxis and the activation of PMN, which are the main source of calprotectin: therefore, calprotectin has an autocrine and paracrine function (67). Notably, calprotectin interaction with non-activated endothelium inhibits its secretion, meaning calprotectin is only released at sites of inflammation by activated phagocytes (Figure 1).
Calprotectin exerts a strong antimicrobial action against a variety of bacterial and fungal pathogens. Calprotectin recognizes proteins related to bacteria, such as lipopolysaccharides (LPS), up-regulating the production of pro-inflammatory profile cytokines, such as TNF-α, IL-1β, and IL-12 locally (68, 69). It is recognized as an endogenous DAMP and binds to the TLR4 receptor and RAGE, amplifying the innate immune response and inducing PMN recruitment to inflamed tissues (3). The antibacterial activity of calprotectin results from the sequestering of transition metals by chelation of nutrient Mn2+ and Zn2+ (70). Engulfment of bacteria by macrophages leads to decreased Zn2+ uptake and increased Zn2+ efflux from the cytoplasm and the efflux of Mn2+ and Fe2+ from the phagosome by NRAMP family transporters (71). Chelation is mediated through two high-affinity binding sites, both of which can bind Zn2+ with nanomolar affinity, while only one binds Mn2+ with this affinity (71).
Calprotectin also activates the MyD88-dependent and TIR domain-containing adaptor protein inducing IFNβ (TRIF; also known as TICAM1)-dependent signaling pathways downstream of TLR4, resulting in NF-κB-mediated and interferon regulatory factor (IRF)-mediated gene transcription. EMMPRIN, a transmembrane glycoprotein of the immunoglobulin superfamily is also able to bind S100A9. However, its biological function has not yet been described (72).
Studies have shown that calprotectin levels are minimal in serum and stool samples from healthy population compared with patients with inflammatory conditions (73–76). High calprotectin serum levels were also observed in patients with infectious disease and sepsis (77). Recently, the accuracy of calprotectin as a biomarker of bacterial respiratory disease was found to be even higher than procalcitonin (78).
Increased calprotectin expression has been found in patients with rheumatic diseases (See Table 2). Higher calprotectin levels have been found in plasma, serum and faecal samples from patients with RA (100), spondyloarthropathies (SpA) (101), inflammatory bowel disease (IBD) (102) and type 2 diabetes (T2D) (103). In IBD, faecal calprotectin has been shown to be a more sensitive indicator of inflammatory activity and is being used for both the diagnosis and follow-up in routine clinical practice (104). Calprotectin levels are not affected by age or gender (105).
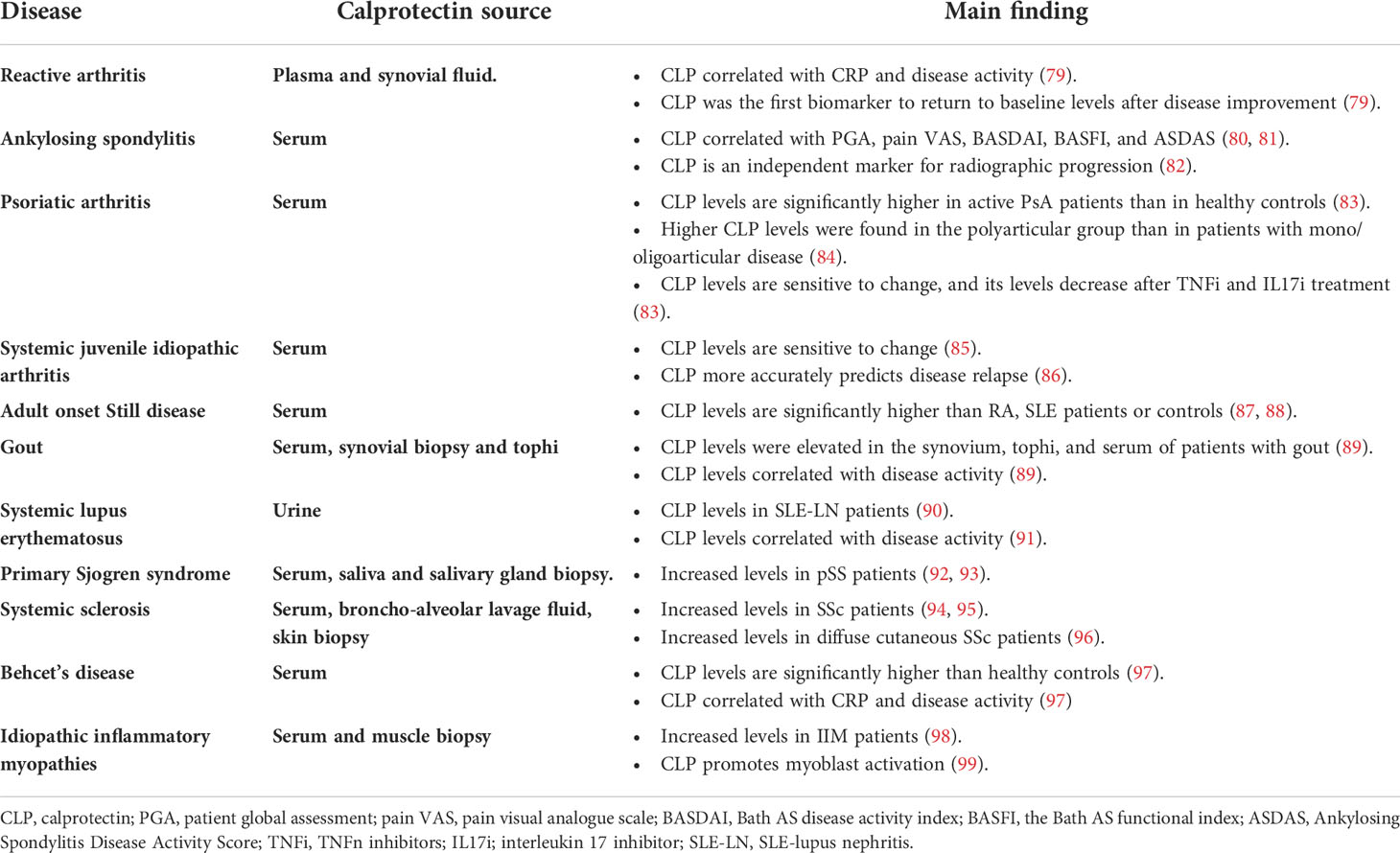
Table 2 Calprotectin in Rheumatic Diseases. This table summarizes the main findings of calprotectin levels and rheumatic diseases other than RA.
Recently, a new disorder, characterized by recurrent infections, hepatosplenomegaly, anemia, cutaneous vasculitis, and evidence of systemic inflammation has been described. These patients have shown hyperzincemia with hypercalprotectinaemia (106).
There is abundant evidence that the innate immune system is persistently activated in RA, as predominately macrophages are found in rheumatoid synovium (107).
In RA, calprotectin induces nitric oxide synthase (iNOS) by nuclear factor κB (NF-κB) activation (108). Calprotectin allows the phosphorylation of multiple protein kinase-mediated signal transduction pathways, including c-Jun-N-amino-terminal kinase (JNK), extracellular-regulated kinase 1/2 (ERK1/2), and Janus kinase/signal transducers and activators of transcription (JAK/STAT) (109). However, calprotectin activation converges in multiple pathways whose activation enhances the production of proinflammatory cytokines, namely tumor necrosis factor-α (TNF-α), IL-6, IL-8, IL-12/23, and IL-18 (110), which are known to be physiopathologically- and clinically-relevant in RA (Figure 2).
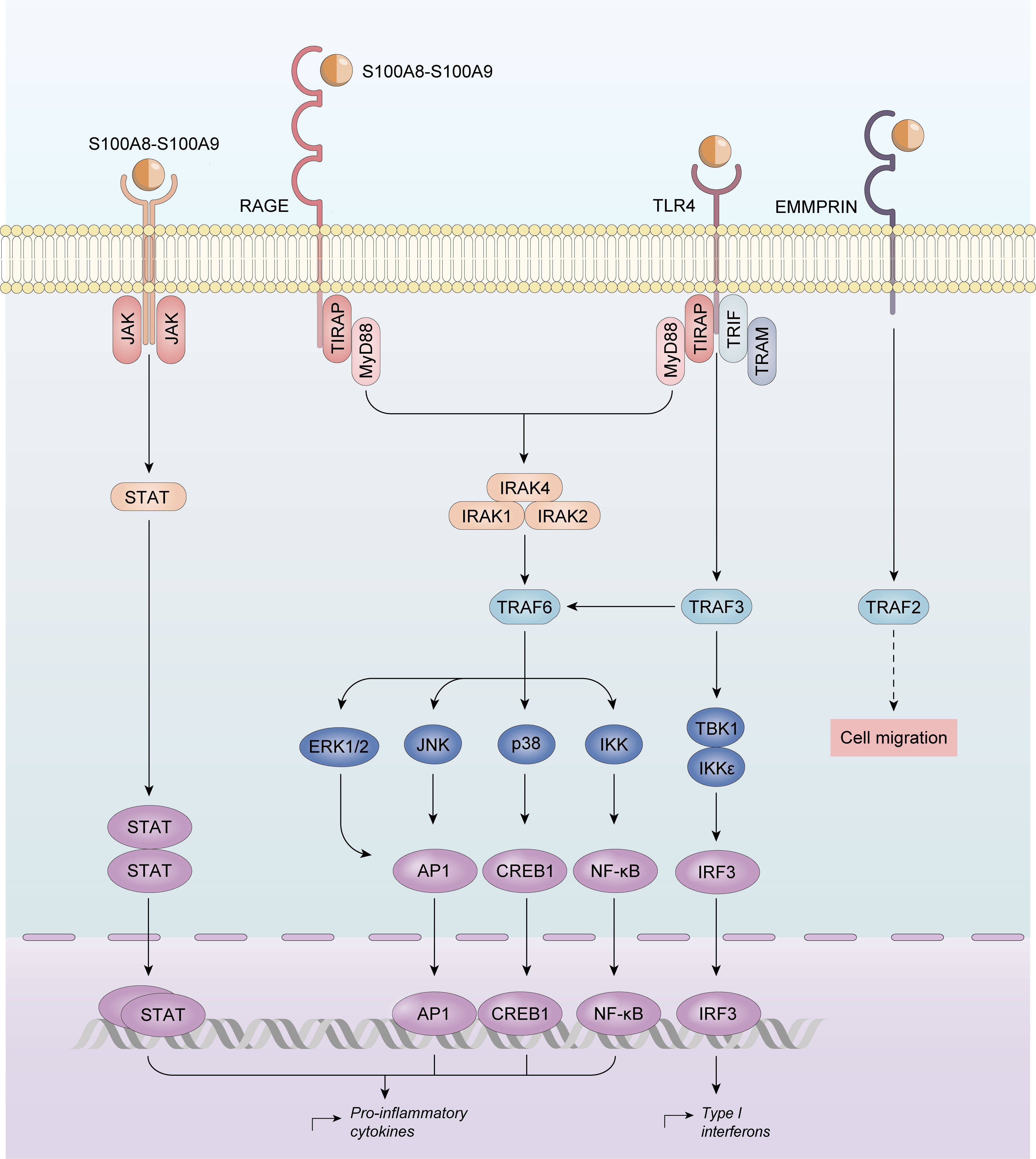
Figure 2 Calprotectin activation in Rheumatoid Arthritis. Calprotectin activates nuclear factor κB (NF-κB), and protein kinase-mediated signal transduction pathways, including c-Jun-N-amino-terminal kinase (JNK), extracellular-regulated kinase 1/2 (ERK1/2), and Janus kinase/signal transducers and activators of transcription (JAK/STAT) upregulating proinflammatory cytokines production.
Calprotectin expression is noted predominantly in the macrophages of the synovial lining layer in tissues adjacent to the cartilage-pannus junction (CPJ), suggesting altered activation and differentiation of lining layer macrophages at the CPJ, which is the site of maximum cartilage destruction in RA (111). Calprotectin is synthesized in fibroblast-like synoviocytes (FLSs), which are crucial players in the pathogenesis of synovitis in RA. IL-22 enhanced FLS proliferation and up-regulated MMP1 and S100A8/A9 production (112). Interestingly, when stimulated FLSs were treated with a JAK 2 and JAK3 inhibitor, there was a significant decrease in IL-22-induced S100A8/A9 production.
Calprotectin is released from activated leukocytes leading to increased concentrations in RA plasma and serum (113, 114). a study focused on determining the association of calprotectin in EDTA-plasma or in serum with disease activity, found stronger plasma associations with all measures of disease activity (115). However, ELISA commercial kits are designed to determine both, and most evidence available is drive by from serum determinations.
Higher calprotectin levels have been found in synovial fluid (SF), plasma and serum from RA patients (113, 114). A recent study demonstrated that the most enhanced proteins in RA SF were the S100A8, S100A9 and S00A12 proteins, using proteomic fingerprints of RA patients’ serum (116). A German study found that calprotectin and other S100 proteins were the most up-regulated proteins in SF in RA patients. Their expression was about 10-fold higher than that observed in the SF of osteoarthritis patients (OA). Not only was calprotectin the best RA biomarker identified in this study, but higher calprotectin levels were found in the SF of RA compared with SpA patients: therefore, calprotectin may differentiate RA from other inflammatory arthritis (117).
Interestingly, calprotectin levels correlated with rheumatoid factor. Furthermore, higher calprotectin plasma levels were found in seropositive than in seronegative patients. The correlation with ACPA titres remains unclear (118–126).
Calprotectin also significantly contributes to comorbid conditions in RA patients, such as cardiovascular disease. High calprotectin levels have been associated with precocious atheroma formation and the accelerated development of atherosclerosis (127).
In summary, serum calprotectin levels may provide information on macrophage activation, supporting their potential role as a biomarker of disease activity, radiographic progression and therapeutic response.
In RA, changes in the amount of synovial sublining macrophages correlate with disease activity (128): these macrophages are the major source of calprotectin, and thus their levels may provide reliable information on their activation 74). Recent studies have demonstrated better correlations between calprotectin and DAS28, swollen joint count (SJC), CDAI, SDAI and physical global VAS than C-reactive protein (CRP) or the erythrocyte sedimentation rate (ESR) (121–123, 129). This was especially pronounced in patients receiving IL-6 inhibitors, with a pronounced decrease in CRP serum levels independently of disease activity (129). These correlations were independent of the disease stage. Studies in both recent-onset patients (105, 130) and those with established disease (123) have demonstrated a significant correlation between calprotectin and SJC, DAS28, SDAI and CDAI (73, 75, 76, 131, 132). In a recent metanalysis of 16 studies, the relationship between calprotectin and disease activity was confirmed (74).
In clinical remission, CRP and ESR may be normal, although there may be residual inflammatory activity, especially in patients receiving biological therapy. Calprotectin levels, but not CRP or ESR levels, were significantly lower in patients with no swollen joints than in those with ≥ 1 swollen joint, supporting the hypothesis that calprotectin levels reflect local ongoing inflammation rather than a systemic inflammatory response (118, 120, 126). Studies by our research group have demonstrated that calprotectin more accurately stratifies disease activity in RA patients in remission or low disease activity receiving TNF inhibitors (133) or the biologic agent, tocilizumab (129). This effect might lead overestimates of the response rate when disease activity indices including ESR or CRP are used (134). In these patients, calprotectin, but not CRP or ESR, can distinguish between patients without swelling from those with ≥1 swollen joint. Calprotectin serum levels, but not CRP, are independent of trough serum tocilizumab levels (129). Recently, these results have been replicated in an independent large cohort (135).
RA patients with normal CRP pose a therapeutic challenge in daily clinical practice. Therefore, calprotectin may have a potential role in assessing disease activity in patients with remission or low disease activity, identifying patients with subclinical synovitis more accurately.
Calprotectin serum levels decreased after treatment, independently of the DMARD type or strategy (105). A significant reduction in serum calprotectin levels was observed after three months of csDMARD treatment (105, 123, 136). The same occurs during biological therapy. Infliximab significantly decreases serum calprotectin levels in RA patients, as confirmed by immunohistochemical staining for S100A8 on serial synovium sections, which showed a progressive decrease in the number of infiltrating S100 A9-positive macrophages. Similar results were observed in RA patients receiving adalimumab and etanercept (105, 137–139).
Musculoskeletal ultrasound (US) is a non-invasive diagnostic technique widely used in rheumatology to assess joint inflammation with greater sensitivity (140). A pilot study explored the associations between calprotectin and comprehensive US examination in 20 RA patients starting treatment with adalimumab and found a significant association with B-mode and the power Doppler score, and there was a correlation between calprotectin and the number of swollen joints (141) and the results were recently replicated (132, 142). In a one-year prospective cohort study of patients with established RA patients, calprotectin showed the overall strongest correlations with US scores and SJC, even after adjustment for several variables (143). We found that RA and PsA patients in remission or low disease activity with a US power Doppler signal had significantly-higher calprotectin levels than those without, and calprotectin correlated better with US power Doppler, synovial hypertrophy and US global scores than ESR or CRP (144). Taken together, serum calprotectin and power Doppler are both identifying local active synovitis in patients with inflammatory arthritis, even those with low levels of disease activity.
Murine arthritis models have shown that overexpression of IL-17 and TNFα strongly enhanced up-regulation of calprotectin, resulting in bone erosion. In contrast, calprotectin deficiency in mice protected against the IL-17/TNFα effect in cartilage (145).
In RA, calprotectin antigens were located in the synovial cartilage, suggesting a pivotal role in cartilage destruction and subchondral bone erosions, a typical hallmark in active RA patients (111). In this regard, a study of 145 RA patients showed that baseline calprotectin levels were independently associated with the van der Heijde modified Sharp score (SvH) and the Rheumatoid Arthritis Articular Damage Score (RAAD score), even when adjusted for CRP, ESR, rheumatoid factor, DAS28, sex and age (123). The prospective follow up of this cohort found that calprotectin was an independent predictor of radiographic joint damage after 10 years. Patients with normal baseline calprotectin levels had less joint damage; again, calprotectin was independently associated with progression in the SvH and RAAD scores (125). Similarly, a longitudinal study with a median of 8-years of follow-up, demonstrated that calprotectin predicts erosive disease and joint space narrowing (73). An exploratory analysis has correlated calprotectin significantly with joint bone marrow edema on MRI in RA patients in clinical remission (146). Recently, the same results have been replicated in a large early-RA cohort from the ARTIC trial, where high levels of calprotectin were associated with radiographic progression in multivariate models (130).
Prediction of the individual response to treatment has become a major clinical challenge in RA. Recent studies and post-hoc clinical trials have provided evidence of calprotectin’s accuracy in predicting the response to csDMARD and bDMARD therapy in RA. In patients receiving csDMARDs, the results are unclear, and decreases in serum calprotectin levels, rather than CRP, were associated with improvements in the SJC over time (105). Patients who achieved remission had a significant reduction in serum calprotectin levels. Likewise, a study shown that baseline serum calprotectin levels decreased rapidly in responders after csDMARD treatment but remained stable in non-responders (122). In early RA, a post-hoc analysis of the prospective ESPOIR cohort found that calprotectin was not an independent predictive factor of the response to MTX. Although the study excluded a large proportion of cohort participants, its results suggested a potential interest in calprotectin as a part of a multivariable score for personalized medicine in these patients (147). Similar results have been seen in RA patients receiving biological therapy: a prospective cohort study evaluated 170 RA patients receiving biological therapy (adalimumab, infliximab, and rituximab). Calprotectin levels were measured at 0, 4 and 16 weeks after biologic drug initiation. As previously described, responders had higher baseline calprotectin levels than non-responders. Higher baseline calprotectin levels increased the odds of being classified as a responder by up to 55-fold, and levels decreased after treatment. In contrast, non-responders had stable calprotectin levels during the study (148). The authors developed a treatment algorithm based on a prediction score using calprotectin and concluded that it may have potential in personalized treatment in RA (149).
Calprotectin levels at baseline were associated with biological treatment survival (150). A prospective cohort study did not find that baseline calprotectin levels were predictive of the response after 6 months of treatment, although a significant decrease in serum levels was observed in responders (122). A recent study demonstrated a significant decrease in calprotectin in the first month of biological therapy, which was predictive of the EULAR response at 3,6 and 12 months (143).
A recent systematic review summarized the results from 17 studies including 1065 patients and found that calprotectin levels decreased after treatment, although there was a wide range of levels and marked interstudy and intrastudy variability. Baseline calprotectin levels were a significant and independent predictor of erosive progression and therapeutic responses, particularly in patients receiving biological treatment (151).
Taken together, this data supports the idea that calprotectin could potentially help to monitor disease activity and predict the response to bDMARDs in RA, although there is no data available on patients receiving DMARDs. Randomized trials are needed to define the role of calprotectin as a predictor of treatment response, but there is a potential role for calprotectin in the follow-up of RA patients.
A clinically relevant unmet need is stratification of the risk of relapse in RA patients in remission or low disease activity. Data demonstrate that calprotectin levels are increased during relapse (152). A prospective one-year follow up cohort study found that calprotectin levels strongly and independently predicted disease relapse in RA with low levels of disease activity during TNFi treatment. In contrast, a prospective cohort study analyzing the role of calprotectin in predicting flares in RA with low disease activity by DAS28 (DAS28<3.2) found that only HAQ-DI remained a significant independent predictor of flares in the multivariate analyses. At the time of the flare, DAS28 and its components significantly correlated with calprotectin, but the correlation was low, suggesting a non-inflammatory component in most events (153).
Another situation is the risk of flare in RA patients before tsDMARD tapering. The capacity of calprotectin in predicting flares in patients undergoing tapering of biologics has been assessed. Calprotectin levels were determined in serum samples from participants in two prospective studies (DRESS and BIO-TOP). Although calprotectin has some predictive value for the clinical response after starting anti-TNF treatment, it has no added value for other clinical factors (154). In contrast, analyses from two tapering studies (IMPROVED study and the RETRO study) showed that calprotectin levels in remission on DMARDs are higher in patients who will flare upon DMARD tapering/cessation (155).
The definitive role of calprotectin as a predictor of disease relapse remains unclear, as there are no specific randomized clinical trials to assess its potential use.
Based on the fundamental role of calprotectin in the modulation of acute and chronic inflammation, its inhibition could be a novel target in the treatment of RA. An experimental study investigated the effects of calprotectin inhibition in RA using neutralizing monoclonal antibodies in a mouse collagen-induced arthritis (CIA) model, Murine S100A9 monoclonal antibody and anti-TNFα treatment were compared. Mice treated with anti-S100A9 showed markedly decreased arthritis severity scores compared to the isotype control group. Overall, anti-S100A9 treatment led to an approximately 50% reduction in disease activity, and preserved bone/collagen integrity. No significant differences in disease activity were observed between anti-S100A9 and anti-TNFα-treated animals, suggesting calprotectin might be a novel therapeutic target in RA (156, 157).
There is still an unmet need for robust biomarkers to objectively monitor inflammatory activity and response to therapy in RA and other immune mediated diseases. Based on the pivotal role of calprotectin in the pathophysiology of acute and chronic inflammation, calprotectin blood levels could be a potential biomarker of disease activity in inflammatory arthritis. In RA, there is growing evidence to support the idea that calprotectin more accurately stratifies disease activity than CRP and ESR. Furthermore, recent data has shown that calprotectin serum levels are a potential tool for monitoring disease activity and the therapeutic response in patients receiving biological therapy. However, larger studies and assay standardization are needed in RA patients to ascertain the role of serum calprotectin as a useful biomarker for monitoring disease activity or response to therapy in clinical practice, as occurs with fecal calprotectin in inflammatory bowel disease. Future applications may include potential therapeutic targets, prediction of the response to treatment, or dose-titration of biologics in a personalized medicine approach.
In conclusion, calprotectin plays a pivotal role in innate immune system activation, increasing chemotaxis and the activation of PMN, promoting the production of inflammatory cytokines and chemokines, and contributing to RA pathogenesis. There is growing evidence to support its higher accuracy in stratifying disease activity than CRP and ESR. Calprotectin has shown the strongest correlations with other sensitive techniques to detect inflammation, such as ultrasound. However, its value as biomarker of treatment response and flare after tapering still need larger, standardized studies.
JI-M: investigation, resources, writing (original draft preparation, review and editing). BF-S: investigation, resources, writing (original draft preparation, review and editing). RS: investigation, resources, writing (original draft preparation, review and editing), supervision, and project administration. All authors contributed to the article and approved the submitted version.
The authors acknowledge David Buss for technical advice, and Antonio García, PhD for Scientific Illustration.
JI-M has received honoraria from AbbVie employee.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Scherer HU, Häupl T, Burmester GR. The etiology of rheumatoid arthritis. J Autoimmun (2020) 110:102400. doi: 10.1016/j.jaut.2019.102400
2. Janeway CA. Approaching the asymptote? evolution and revolution in immunology. Cold Spring Harb Symp Quant Biol (1989) 54 Pt 1:1–13. doi: 10.1101/SQB.1989.054.01.003
3. Iwasaki A, Medzhitov R. Toll-like receptor control of the adaptive immune responses. Nat Immunol (2004) 5(10):987–95. doi: 10.1038/ni1112
4. Ryckman C, McColl SR, Vandal K, de Médicis R, Lussier A, Poubelle PE, et al. Role of S100A8 and S100A9 in neutrophil recruitment in response to monosodium urate monohydrate crystals in the air-pouch model of acute gouty arthritis. Arthritis Rheumatol (2003) 48(8):2310–20. doi: 10.1002/art.11079
5. Ryckman C, Vandal K, Rouleau P, Talbot M, Tessier PA. Proinflammatory activities of S100: proteins S100A8, S100A9, and S100A8/A9 induce neutrophil chemotaxis and adhesion. J Immunol (2003) 170(6):3233–42. doi: 10.4049/jimmunol.170.6.3233
6. Newton RA, Hogg N. The human S100 protein MRP-14 is a novel activator of the beta 2 integrin mac-1 on neutrophils. J Immunol (1998) 160(3):1427–35.
7. Moore BW. A soluble protein characteristic of the nervous system. Biochem Biophys Res Commun (1965) 19(6):739–44. doi: 10.1016/0006-291X(65)90320-7
8. Donato R. S100: a multigenic family of calcium-modulated proteins of the EF-hand type with intracellular and extracellular functional roles. Int J Biochem Cell Biol (2001) 33(7):637–68. doi: 10.1016/S1357-2725(01)00046-2
9. Rohde D, Ritterhoff J, Voelkers M, Katus HA, Parker TG, Most P. S100A1: a multifaceted therapeutic target in cardiovascular disease. J Cardiovasc Transl Res (2010) 3(5):525–37. doi: 10.1007/s12265-010-9211-9
10. Ilg EC, Sciiafer BW. Expression pattern of sloo calcium-binding proteins in human tumors. (1996) 68:325–32. doi: 10.1002/(SICI)1097-0215(19961104)68:3<325::AID-IJC10>3.0.CO;2-7
11. van Dieck J, Teufel DP, Jaulent AM, Fernandez-Fernandez MR, Rutherford TJ, Wyslouch-Cieszynska A, et al. Posttranslational modifications affect the interaction of S100 proteins with tumor suppressor p53. J Mol Biol (2009) 394(5):922–30. doi: 10.1016/j.jmb.2009.10.002
12. Kizawa K, Takahara H, Troxler H, Kleinert P, Mochida U, Heizmann CW. Specific citrullination causes assembly of a globular S100A3 homotetramer: a putative Ca2+ modulator matures human hair cuticle. J Biol Chem (2008) 283(8):5004–13. doi: 10.1074/jbc.M709357200
13. Bresnick AR, Weber DJ, Zimmer DB. S100 proteins in cancer. Nat Rev Cancer (2015) 15(2):96–109. doi: 10.1038/nrc3893
14. Wang DT, Chu WH, Sun HM, Ba HX, Li CY. Expression and functional analysis of tumor-related factor S100A4 in antler stem cells. J Histochem Cytochem (2017) 65(10):579–91. doi: 10.1369/0022155417727263
15. Hancq S, Salmon I, Brotchi J, de Witte O, Gabius HJ, Heizmann CW, et al. S100A5: a marker of recurrence in WHO grade I meningiomas. Neuropathol Appl Neurobiol (2004) 30(2):178–87. doi: 10.1046/j.0305-1846.2003.00525.x
16. Chen D, Luo L, Liang C. Aberrant S100A16 expression might be an independent prognostic indicator of unfavorable survival in non-small cell lung adenocarcinoma. PloS One (2018) 13(5):e0197402. doi: 10.1371/journal.pone.0197402
17. Filipek A, Leśniak W. S100A6 and its brain ligands in neurodegenerative disorders. Int J Mol Sci (2020) 21(11):3979. doi: 10.3390/ijms21113979
18. Dong XH, Dai D, Yang ZD, Yu XO, Li H, Kang H. S100 calcium binding protein A6 and associated long noncoding ribonucleic acids as biomarkers in the diagnosis and staging of primary biliary cholangitis. World J Gastroenterol (2021) 27(17):1973–92. doi: 10.3748/wjg.v27.i17.1973
19. Wang T, Han S, Du G. S100A6 represses calu-6 lung cancer cells growth via inhibiting cell proliferation, migration, invasion and enhancing apoptosis. Cell Biochem Funct (2021) 39(6):771–9. doi: 10.1002/cbf.3639
20. Donato R R, Cannon B, Sorci G, Riuzzi F, Hsu K, Weber D J, et al. Functions of S100 proteins. Curr Mol Med (2013) 13(1):24. doi: 10.2174/156652413804486214
21. Leśniak W, Słomnicki ŁP, Filipek A. S100A6 - new facts and features. Biochem Biophys Res Commun (2009) 390(4):1087–92. doi: 10.1016/j.bbrc.2009.10.150
22. Gläser R, Meyer-Hoffert U, Harder J, Cordes J, Wittersheim M, Kobliakova J, et al. The antimicrobial protein psorias in (S100A7) is upregulated in atopic dermatitis and after experimental skin barrier disruption. J Invest Dermatol (2009) 129(3):641–9. doi: 10.1038/jid.2008.268
23. Qin W, Ho L, Wang J, Peskind E, Pasinetti GM. S100A7, a novel alzheimer’s disease biomarker with non-amyloidogenic alpha-secretase activity acts via selective promotion of ADAM-10. PloS One (2009) 4(1):e4183. doi: 10.1371/journal.pone.0004183
24. Zheng Y, Niyonsaba F, Ushio H, Ikeda S, Nagaoka I, Okumura K, et al. Microbicidal protein psoriasin is a multifunctional modulator of neutrophil activation. Immunology (2008) 124(3):357–67. doi: 10.1111/j.1365-2567.2007.02782.x
25. Wolf R, Howard OMZ, Dong HF, Voscopoulos C, Boeshans K, Winston J, et al. Chemotactic activity of S100A7 (Psoriasin) is mediated by the receptor for advanced glycation end products and potentiates inflammation with highly homologous but functionally distinct S100A15. J Immunol (2008) 181(2):1499–506. doi: 10.4049/jimmunol.181.2.1499
26. Hsu K, Champaiboon C, Guenther B, Sorenson B, Khammanivong A, Ross K, et al. Anti-infective protective properties of s100 calgranulins. Antiinflamm Antiallergy Agents Med Chem (2009) 8(4):290–305. doi: 10.2174/187152309789838975
27. Li C, Zhang F, Lin M, Liu J. Induction of S100A9 gene expression by cytokine oncostatin m in breast cancer cells through the STAT3 signaling cascade. Breast Cancer Res (2004) 87(2):123–34. doi: 10.1023/B:BREA.0000041594.36418.f6
28. Rescher U, Gerke V. S100A10/p11: family, friends and functions. Pflugers Arch (2008) 455(4):575–82. doi: 10.1007/s00424-007-0313-4
29. O’Connell PA, Madureira PA, Berman JN, Liwski RS, Waisman DM. Regulation of S100A10 by the PML-RAR-α oncoprotein. Blood (2011) 117(15):4095–105. doi: 10.1182/blood-2010-07-298851
30. Cecil DL, Johnson K, Rediske J, Lotz M, Schmidt AM, Terkeltaub R. Inflammation-induced chondrocyte hypertrophy is driven by receptor for advanced glycation end products. J Immunol (2005) 175(12):8296–302. doi: 10.4049/jimmunol.175.12.8296
31. Sakaguchi M, Sonegawa H, Murata H, Kitazoe M, Futami JI, Kataoka K, et al. S100A11, an dual mediator for growth regulation of human keratinocytes. Mol Biol Cell (2008) 19(1):78–85. doi: 10.1091/mbc.e07-07-0682
32. Goyette J, Geczy CL. Inflammation-associated S100 proteins: new mechanisms that regulate function. Amino Acids (2011) 41(4):821–42. doi: 10.1007/s00726-010-0528-0
33. Bowman MAH, Gawdzik J, Bukhari U, Husain AN, Toth PT, Kim G, et al. S100A12 in vascular smooth muscle accelerates vascular calcification in apolipoprotein e-null mice by activating an osteogenic gene regulatory program. Arterioscler Thromb Vasc Biol (2011) 31(2):337–44. doi: 10.1161/ATVBAHA.110.217745
34. Hayrabedyan S, Kyurkchiev S, Kehayov I. FGF-1 and S100A13 possibly contribute to angiogenesis in endometriosis. J Reprod Immunol (2005) 67(1–2):87–101. doi: 10.1016/j.jri.2005.07.001
35. Jin Q, Chen H, Luo A, Ding F, Liu Z. S100A14 stimulates cell proliferation and induces cell apoptosis at different concentrations via receptor for advanced glycation end products (RAGE). PloS One (2011) 6(4):e19375. doi: 10.1371/journal.pone.0019375
36. Chen H, Yuan Y, Zhang C, Luo A, Ding F, Ma J, et al. Involvement of S100A14 protein in cell invasion by affecting expression and function of matrix metalloproteinase (MMP)-2 via p53-dependent transcriptional regulation. J Biol Chem (2012) 287(21):17109–19. doi: 10.1074/jbc.M111.326975
37. Wolf R, Lewerenz V, Büchau AS, Walz M, Ruzicka T. Human S100A15 splice variants are differentially expressed in inflammatory skin diseases and regulated through Th1 cytokines and calcium. Exp Dermatol (2007) 16(8):685–91. doi: 10.1111/j.1600-0625.2007.00587.x
38. Büchau AS, Hassan M, Kukova G, Lewerenz V, Kellermann S, Würthner JU, et al. S100A15, an antimicrobial protein of the skin: regulation by e. coli through toll-like receptor 4. J Invest Dermatol (2007) 127(11):2596–604. doi: 10.1038/sj.jid.5700946
39. Sturchler E, Cox JA, Durussel I, Weibel M, Heizmann CW. S100A16, a novel calcium-binding protein of the EF-hand superfamily. J Biol Chem (2006) 281(50):38905–17. doi: 10.1074/jbc.M605798200
40. Liu Y, Zhang R, Xin J, Sun Y, Li J, Wei D, et al. Identification of S100A16 as a novel adipogenesis promoting factor in 3T3-L1 cells. Endocrinology (2011) 152(3):903–11. doi: 10.1210/en.2010-1059
41. Donato R, Sorci G, Riuzzi F, Arcuri C, Bianchi R, Brozzi F, et al. S100B’s double life: intracellular regulator and extracellular signal. Biochim Biophys Acta (2009) 1793(6):1008–22. doi: 10.1016/j.bbamcr.2008.11.009
42. Kleindienst A, Bullock MR. A critical analysis of the role of the neurotrophic protein S100B in acute brain injury. J Neurotrauma (2006) 23(8):1185–200. doi: 10.1089/neu.2006.23.1185
43. Austermann J, Nazmi AR, Müller-Tidow C, Gerke V. Characterization of the Ca2+ -regulated ezrin-S100P interaction and its role in tumor cell migration. J Biol Chem (2008) 283(43):29331–40. doi: 10.1074/jbc.M806145200
44. Du M, Wang G, Ismail TM, Gross S, Fernig DG, Barraclough R, et al. S100P dissociates myosin IIA filaments and focal adhesion sites to reduce cell adhesion and enhance cell migration. J Biol Chem (2012) 287(19):15330–44. doi: 10.1074/jbc.M112.349787
45. Gribenko AV, Hopper JE, Makhatadze GI. Molecular characterization and tissue distribution of a novel member of the S100 family of EF-hand proteins. Biochemistry (2001) 40(51):15538–48. doi: 10.1021/bi0114731
46. Grevers LC, de Vries TJ, Vogl T, Abdollahi-Roodsaz S, Sloetjes AW, Leenen PJM, et al. S100A8 enhances osteoclastic bone resorption in vitro through activation of toll-like receptor 4: implications for bone destruction in murine antigen-induced arthritis. Arthritis Rheumatol (2011) 63(5):1365–75. doi: 10.1002/art.30290
47. Iotzova-Weiss G, Dziunycz PJ, Freiberger SN, Läuchli S, Hafner J, Vogl T, et al. S100A8/A9 stimulates keratinocyte proliferation in the development of squamous cell carcinoma of the skin via the receptor for advanced glycation-end products. PloS One (2015) 10(3):e0120971. doi: 10.1371/journal.pone.0120971
48. Hessian PA, Edgeworth J, Hogg N. MRP-8 and MRP-14, two abundant Ca(2+)-binding proteins of neutrophils and monocytes. J Leukoc Biol (1993) 53(2):197–204. doi: 10.1002/jlb.53.2.197
49. Dorin JR, Emslie E, van Heyningen V. Related calcium-binding proteins map to the same subregion of chromosome 1q and to an extended region of synteny on mouse chromosome 3. Genomics (1990) 8(3):420–6. doi: 10.1016/0888-7543(90)90027-R
50. Lagasse E, Clerc RG. Cloning and expression of two human genes encoding calcium-binding proteins that are regulated during myeloid differentiation. Mol Cell Biol (1988) 8(6):2402–10. doi: 10.1128/mcb.8.6.2402-2410.1988
51. Fagerhol MK, Dale I AT. Release and quantitation of a leukocyte derived protein (L1). Scand J Haematol (1980) 24:393–8. doi: 10.1111/j.1600-0609.1980.tb02754.x
52. Wilson GB, Jahn TL FJR. Demonstration of serum protein differences in cystic fibrosis by isoelectric focusing in thin-layer polyacrylamide gels. Clin Chim Acta (1973) 49(1):79–91. doi: 10.1016/0009-8981(73)90346-X
53. Wilkinson MM, Busuttil A, Hayward C, Brock DJ, Dorin JR, Van Heyningen V. Expression pattern of two related cystic fibrosis-associated calcium-binding proteins in normal and abnormal tissues. J Cell Sci (1988) 91(Pt 2):221–30. doi: 10.1242/jcs.91.2.221
54. Dorin JR, Novak M, Hill RE, Brock DJ, Secher DS, van Heyningen V. A clue to the basic defect in cystic fibrosis from cloning the CF antigen gene. Nature (1987) 326(6113):614–7. doi: 10.1038/326614a0
55. Waugh N, Cummins E, Royle P, Kandala NB, Shyangdan D, Arasaradnam R, et al. Faecal calprotectin testing for differentiating amongst inflammatory and non-inflammatory bowel diseases: systematic review and economic evaluation. Health Technol Assess (2013) 17(55):xv-xix, 1–211. doi: 10.3310/hta17550
56. Yang Z, Clark N, Park KT. Effectiveness and cost-effectiveness of measuring fecal calprotectin in diagnosis of inflammatory bowel disease in adults and children. Clin Gastroenterol Hepatol (2014) 12(2):253–262.e2. doi: 10.1016/j.cgh.2013.06.028
57. D’Haens G, Ferrante M, Vermeire S, Baert F, Noman M, Moortgat L, et al. Fecal calprotectin is a surrogate marker for endoscopic lesions in inflammatory bowel disease. Inflammation Bowel Dis (2012) 18(12):2218–24. doi: 10.1002/ibd.22917
58. Khaki-Khatibi F, Qujeq D, Kashifard M, Moein S, Maniati M, Vaghari-Tabari M. Calprotectin in inflammatory bowel disease. Clinica Chimica Acta (2020) 510:556–65. doi: 10.1016/j.cca.2020.08.025
59. Menees SB, Powell C, Kurlander J, Goel A, Chey WD. A meta-analysis of the utility of c-reactive protein, erythrocyte sedimentation rate, fecal calprotectin, and fecal lactoferrin to exclude inflammatory bowel disease in adults with IBS. Am J Gastroenterol (2015) 110(3):444–54. doi: 10.1038/ajg.2015.6
60. Odink K, Cerletti N, Brüggen J, Clerc RG, Tarcsay L, Zwadlo G, et al. Two calcium-binding proteins in infiltrate macrophages of rheumatoid arthritis. Nature (1987) 330(6143):80–2. doi: 10.1038/330080a0
61. Vogl T, Ludwig S, Goebeler M, Strey A, Thorey IS, Reichelt R, et al. MRP8 and MRP14 control microtubule reorganization during transendothelial migration of phagocytes. Blood (2004) 104(13):4260–8. doi: 10.1182/blood-2004-02-0446
62. Heizmann CW, Fritz G, Schäfer BW. S100 proteins: structure, functions and pathology. Front Biosci (2002) 7:d1356–68. doi: 10.2741/A846
63. Roth J, Burwinkel F, van den Bos C, Goebeler M, Vollmer E, Sorg C. MRP8 and MRP14, s-100-like proteins associated with myeloid differentiation, are translocated to plasma membrane and intermediate filaments in a calcium-dependent manner. Blood (1993) 82(6):1875–83. doi: 10.1182/blood.V82.6.1875.1875
64. McCormick A, Heesemann L, Wagener J, Marcos V, Hartl D, Loeffler J, et al. NETs formed by human neutrophils inhibit growth of the pathogenic mold aspergillus fumigatus. Microbes Infect (2010) 12(12–13):928–36. doi: 10.1016/j.micinf.2010.06.009
65. Viemann D, Strey A, Janning A, Jurk K, Klimmek K, Vogl T, et al. Myeloid-related proteins 8 and 14 induce a specific inflammatory response in human microvascular endothelial cells. Blood (2005) 105(7):2955–62. doi: 10.1182/blood-2004-07-2520
66. Srikrishna G, Panneerselvam K, Westphal V, Abraham V, Varki A, Freeze HH. Two proteins modulating transendothelial migration of leukocytes recognize novel carboxylated glycans on endothelial cells. J Immunol (2001) 166(7):4678–88. doi: 10.4049/jimmunol.166.7.4678
67. Vogl T, Tenbrock K, Ludwig S, Leukert N, Ehrhardt C, van Zoelen MAD, et al. Mrp8 and Mrp14 are endogenous activators of toll-like receptor 4, promoting lethal, endotoxin-induced shock. Nat Med (2007) 13(9):1042–9. doi: 10.1038/nm1638
68. Goebeler M, Roth J, Van Den Bos C, Ader G, Sorg C. Increase of calcium levels in epithelial cells induces translocation of calcium-binding proteins migration inhibitory factor-related protein 8 (MRP8) and MRP14 to keratin intermediate filaments. Biochem J (1995) 309(Pt 2):419–24. doi: 10.1042/bj3090419
69. Roth J, Vogl T, Sorg C, Sunderkötter C. Phagocyte-specific S100 proteins: a novel group of proinflammatory molecules. Trends Immunol (2003) 24(4):155–8. doi: 10.1016/S1471-4906(03)00062-0
70. Corbin BD, Seeley EH, Raab A, Feldmann J, Miller MR, Torres VJ, et al. Metal chelation and inhibition of bacterial growth in tissue abscesses. Science (2008) 319(5865):962–5. doi: 10.1126/science.1152449
71. Kehl-Fie TE, Chitayat S, Hood MI, Damo S, Restrepo N, Garcia C, et al. Nutrient metal sequestration by calprotectin inhibits bacterial superoxide defense, enhancing neutrophil killing of staphylococcus aureus. Cell Host Microbe (2011) 10(2):158–64. doi: 10.1016/j.chom.2011.07.004
72. Austermann J, Spiekermann C, Roth J. S100 proteins in rheumatic diseases. Nat Rev Rheumatol (2018) 14(9):528–41. doi: 10.1038/s41584-018-0058-9
73. Bach M, Moon J, Moore R, Pan T, Nelson JL, Lood C. A neutrophil activation biomarker panel in prognosis and monitoring of patients with rheumatoid arthritis. Arthritis Rheumatol (2020) 72(1):47–56. doi: 10.1002/art.41062
74. Bae SC, Lee YH. Calprotectin levels in rheumatoid arthritis and their correlation with disease activity: a meta-analysis. Postgrad Med (2017) 129(5):531–7. doi: 10.1080/00325481.2017.1319729
75. Wang Y, Liang Y. Clinical significance of serum calprotectin level for the disease activity in active rheumatoid arthritis with normal c-reactive protein. Int J Clin Exp Pathol (2019) 12(3):1009–14.
76. Van Hoovels L, Vander Cruyssen B, Bogaert L, Van Den Bremt S, Bossuyt X. Pre-analytical and analytical confounders of serum calprotectin as a biomarker in rheumatoid arthritis. Clin Chem Lab Med (2019) 58(1):40–9. doi: 10.1515/cclm-2019-0508
77. Wollmer M, Wändell P, Rosenqvist M, Larsson A, Melander O, Wessman T, et al. Plasma calprotectin in the emergency department: a potential clinical biomarker for patients with infectious diseases. Scand J Clin Lab Invest (2021) 81(7):593–7. doi: 10.1080/00365513.2021.1980223
78. Havelka A, Sejersen K, Venge P, Pauksens K, Larsson A. Calprotectin, a new biomarker for diagnosis of acute respiratory infections. Sci Rep (2020) 10(1):1–7. doi: 10.1038/s41598-020-61094-z
79. Hammer HB, Kvien TK, Glennås A, Melby K. A longitudinal study of calprotectin as an inflammatory marker in patients with reactive arthritis. Clin Exp Rheumatol (1995) 13(1):59–64.
80. Hu H, Du F, Zhang S, Zhang W. Serum calprotectin correlates with risk and disease severity of ankylosing spondylitis and its change during first month might predict favorable response to treatment. Mod Rheumatol (2019) 29(5):836–42. doi: 10.1080/14397595.2018.1519103
81. Gupta L, Bhattacharya S, Agarwal V, Aggarwal A. Elevated levels of serum MRP8/14 in ankylosing spondylitis: associated with peripheral arthritis and active disease. Clin Rheumatol (2016) 35(12):3075–9. doi: 10.1007/s10067-016-3448-x
82. Turina MC, Yeremenko N, Paramarta JE, De Rycke L, Baeten D. Calprotectin (S100A8/9) as serum biomarker for clinical response in proof-of-concept trials in axial and peripheral spondyloarthritis. Arthritis Res Ther (2014) 16(4):413. doi: 10.1186/s13075-014-0413-4
83. Sokolova MV, Simon D, Nas K, Zaiss MM, Luo Y, Zhao Y, et al. A set of serum markers detecting systemic inflammation in psoriatic skin, entheseal, and joint disease in the absence of c-reactive protein and its link to clinical disease manifestations. Arthritis Res Ther (2020) 22(1):26. doi: 10.1186/s13075-020-2111-8
84. Hansson C, Eriksson C, Alenius GM. S-calprotectin (S100A8/S100A9): A potential marker of inflammation in patients with psoriatic arthritis. (2014) 2014:696415. doi: 10.1155/2014/696415
85. Holzinger D, Frosch M, Kastrup A, Prince FHM, Otten MH, Van Suijlekom-Smit LWA, et al. The toll-like receptor 4 agonist MRP8/14 protein complex is a sensitive indicator for disease activity and predicts relapses in systemic-onset juvenile idiopathic arthritis. Ann Rheum Dis (2012) 71(6):974–80. doi: 10.1136/annrheumdis-2011-200598
86. Frosch M, Vogl T, Seeliger S, Wulffraat N, Kuis W, Viemann D, et al. Expression of myeloid-related proteins 8 and 14 in systemic-onset juvenile rheumatoid arthritis. Arthritis Rheumatol (2003) 48(9):2622–6. doi: 10.1002/art.11177
87. Mitrovic S, Fautrel B. New markers for adult-onset still’s disease. Joint Bone Spine (2018) 85(3):285–93. doi: 10.1016/j.jbspin.2017.05.011
88. Jung JY, Suh CH, Kim HA. The role of damage-associated molecular pattern for pathogenesis and biomarkers in adult-onset still’s disease. Expert Rev Mol Diagn (2019) 19(6):459–68. doi: 10.1080/14737159.2019.1615449
89. Holzinger D, Nippe N, Vogl T, Marketon K, Mysore V, Weinhage T, et al. Myeloid-related proteins 8 and 14 contribute to monosodium urate monohydrate crystal-induced inflammation in gout. Arthritis Rheumatol (2014) 66(5):1327–39. doi: 10.1002/art.38369
90. Turnier JL, Fall N, Thornton S, Witte D, Bennett MR, Appenzeller S, et al. Urine S100 proteins as potential biomarkers of lupus nephritis activity. Arthritis Res Ther (2017) 19(1):242. doi: 10.1186/s13075-017-1444-4
91. Haga HJ, Brun JG, Berntzen HB, Cervera R, Khamashta M, Hughes GRV. Calprotectin in patients with systemic lupus erythematosus: relation to clinical and laboratory parameters of disease activity. Lupus (1993) 2(1):47–50. doi: 10.1177/096120339300200108
92. Cuida M, Brun JG, Johannessen AC, Jonsson R. Immunohistochemical characterization of the cellular infiltrates in sjögren’s syndrome, rheumatoid arthritis and osteoarthritis with special reference to calprotectin-producing cells. APMIS (1996) 104(12):881–90. doi: 10.1111/j.1699-0463.1996.tb04953.x
93. Nordal HH, Brun JG, Halse AK, Madland TM, Fagerhol MK, Jonsson R. Calprotectin (S100A8/A9), S100A12, and EDTA-resistant S100A12 complexes (ERAC) in primary sjögren’s syndrome. Scand J Rheumatol (2014) 43(1):76–8. doi: 10.3109/03009742.2013.848930
94. Xu X, Wu WY, Tu WZ, Chu HY, Zhu XX, Liang MR, et al. Increased expression of S100A8 and S100A9 in patients with diffuse cutaneous systemic sclerosis. a correlation with organ involvement and immunological abnormalities. Clin Rheumatol (2013) 32(10):1501–10. doi: 10.1007/s10067-013-2305-4
95. Hesselstrand R, Wildt M, Bozovic G, Andersson-Sjöland A, Andréasson K, Scheja A, et al. Biomarkers from bronchoalveolar lavage fluid in systemic sclerosis patients with interstitial lung disease relate to severity of lung fibrosis. Respir Med (2013) 107(7):1079–86. doi: 10.1016/j.rmed.2013.03.015
96. Van Bon L, Cossu M, Loof A, Gohar F, Wittkowski H, Vonk M, et al. Proteomic analysis of plasma identifies the toll-like receptor agonists S100A8/A9 as a novel possible marker for systemic sclerosis phenotype. Ann Rheum Dis (2014) 73(8):1585–9. doi: 10.1136/annrheumdis-2013-205013
97. Torgutalp M, Dincer ABK, Yayla EM, Yurteri EU, Okatan EI, Guloksuz GEA, et al. THU0610 serum calprotectin levels in bechet’s disease: relationships between disease activity and clinical parameters. Ann Rheum Dis (2018) 77:504.
98. Nistala K, Varsani H, Wittkowski H, Vogl T, Krol P, Shah V, et al. Myeloid related protein induces muscle derived inflammatory mediators in juvenile dermatomyositis. Arthritis Res Ther (2013) 15(5):R131. doi: 10.1186/ar4311
99. Seeliger S, Vogl T, Engels IH, Schröder JM, Sorg C, Sunderkötter C, et al. Expression of calcium-binding proteins MRP8 and MRP14 in inflammatory muscle diseases. Am J Pathol (2003) 163(3):947–56. doi: 10.1016/S0002-9440(10)63454-2
100. Nys G, Cobraiville G, Servais AC, Malaise MG, de Seny D, Fillet M. Targeted proteomics reveals serum amyloid a variants and alarmins S100A8-S100A9 as key plasma biomarkers of rheumatoid arthritis. Talanta (2019) 204:507–17. doi: 10.1016/j.talanta.2019.06.044
101. Oktayoglu P, Bozkurt M, Mete N, Caglayan M, Em S, Nas K. Elevated serum levels of calprotectin (myeloid-related protein 8/14) in patients with ankylosing spondylitis and its association with disease activity and quality of life. J Investig Med (2014) 62(6):880–4. doi: 10.1097/JIM.0000000000000095
102. Meuwis MA, Vernier-Massouille G, Grimaud JC, Bouhnik Y, Laharie D, Piver E, et al. Serum calprotectin as a biomarker for crohn’s disease. J Crohns Colitis (2013) 7(12):e678–83. doi: 10.1016/j.crohns.2013.06.008
103. Pedersen L, Nybo M, Poulsen MK, Henriksen JE, Dahl J, Rasmussen LM, et al. Plasma calprotectin and its association with cardiovascular disease manifestations, obesity and the metabolic syndrome in type 2 diabetes mellitus patients. BMC Cardiovasc Disord (2014) 14(1):196. doi: 10.1186/1471-2261-14-196
104. Bressler B, Panaccione R, Fedorak RN, Seidman EG. Clinicians’ guide to the use of fecal calprotectin to identify and monitor disease activity in inflammatory bowel disease. Can J Gastroenterol Hepatol (2015) 29(7):369–72. doi: 10.1155/2015/852723
105. Andrés Cerezo L, Mann H, Pecha O, Pleštilová L, Pavelka K, Vencovský J, et al. Decreases in serum levels of S100A8/9 (calprotectin) correlate with improvements in total swollen joint count in patients with recent-onset rheumatoid arthritis. Arthritis Res Ther (2011) 13(4):R122. doi: 10.1186/ar3426
106. Resende LO, Jorge MFS, Schmitt JV. Extensive pyoderma gangrenosum-like lesions revealing a case of hyperzincemia and hypercalprotectinemia: when to suspect it? Bras Dermatol (2019) 94(6):713. doi: 10.1016/j.abd.2018.12.001
107. Youssef PP, Smeets TJ, Bresnihan B, Cunnane G, Fitzgerald O, Breedveld F, et al. Microscopic measurement of cellular infiltration in the rheumatoid arthritis synovial membrane: a comparison of semiquantitative and quantitative analysis. Br J Rheumatol (1998) 37(9):1003–7. doi: 10.1093/rheumatology/37.9.1003
108. Firestein GS. Evolving concepts of rheumatoid arthritis. Nature (2003) 423(6937):356–61. doi: 10.1038/nature01661
109. Malemud CJ. Myeloid-related protein activity in rheumatoid arthritis. Int J Inflam (2011) 2011:580295. doi: 10.4061/2011/580295
110. Malemud C, Pearlman E. Targeting JAK/STAT signaling pathway in inflammatory diseases. Curr Signal Transduct Ther (2009) 4(3):201–21. doi: 10.2174/157436209789057467
111. Youssef P, Roth J, Frosch M, Costello P, Fitzgerald O, Sorg C, et al. Expression of myeloid related proteins (MRP) 8 and 14 and the MRP8/14 heterodimer in rheumatoid arthritis synovial membrane. J Rheumatol (1999) 26(12):2523–8.
112. Carrión M, Juarranz Y, Martínez C, González-Álvaro I, Pablos JL, Gutiérrez-Cañas I, et al. IL-22/IL-22R1 axis and S100A8/A9 alarmins in human osteoarthritic and rheumatoid arthritis synovial fibroblasts. Rheumatol (Oxford) (2013) 52(12):2177–86. doi: 10.1093/rheumatology/ket315
113. Berntzen HB, Ölmez Ü, Fagerhol MK, Munthe E. The leukocyte protein L1 in plasma and synovial fluid from patients with rheumatoid arthritis and osteoarthritis. Scand J Rheumatol (1991) 20(2):74–82. doi: 10.3109/03009749109165280
114. Uchida T, Fukawa A, Uchida M, Fujita K, Saito K. Application of a novel protein biochip technology for detection and identification of rheumatoid arthritis biomarkers in synovial fluid. J Proteome Res (2002) 1(6):495–9. doi: 10.1021/pr025531w
115. Nordal HH, Fagerhol MK, Halse AK, Hammer HB. Calprotectin (S100A8/A9) should preferably be measured in EDTA-plasma; results from a longitudinal study of patients with rheumatoid arthritis. (2018) 78(1–2):102–8. doi: 10.1080/00365513.2017.1419371
116. Baillet A. Protéines S100A8, S100A9 et S100A12: marqueurs inflammatoires ou acteurs physiopathologiques de la polyarthrite rhumatoïde. Rev Medecine Interne (2010) 31(6):458–61. doi: 10.1016/j.revmed.2009.10.435
117. Baillet A, Trocmé C, Berthier S, Arlotto M, Grange L, Chenau J, et al. Synovial fluid proteomic fingerprint: S100A8, S100A9 and S100A12 proteins discriminate rheumatoid arthritis from other inflammatory joint diseases. Rheumatol (Oxford) (2010) 49(4):671–82. doi: 10.1093/rheumatology/kep452
118. Brun JG, Haga HJ, Bøe E, Kallay I, Lekven C, Berntzen HB, et al. Calprotectin in patients with rheumatoid arthritis: relation to clinical and laboratory variables of disease activity. J Rheumatol (1992) 19(6):859–62.
119. Brun JG, Jonsson R, Haga HJ. Measurement of plasma calprotectin as an indicator of arthritis and disease activity in patients with inflammatory rheumatic diseases. J Rheumatol (1994) 21(4):733–8.
120. Berntzen HB, Munthe E, Fagerhol MK. A longitudinal study of the leukocyte protein L1 as an indicator of disease activity in patients with rheumatoid arthritis. J Rheumatol (1989) 16(11):1416–20.
121. De Seny D, Fillet M, Ribbens C, Marée R, Meuwis MA, Lutteri L, et al. Monomeric calgranulins measured by SELDI-TOF mass spectrometry and calprotectin measured by ELISA as biomarkers in arthritis. Clin Chem (2008) 54(6):1066–75. doi: 10.1373/clinchem.2007.099549
122. García-Arias M, Pascual-Salcedo D, Ramiro S, Ueberschlag ME, Jermann TM, Cara C, et al. Calprotectin in rheumatoid arthritis: association with disease activity in a cross-sectional and a longitudinal cohort. Mol Diagn Ther (2013) 17(1):49–56. doi: 10.1007/s40291-013-0016-9
123. Hammer HB, Ødeg̊ard S, Fagerhol MK, Landewé R, van der Heijde D, Uhlig T, et al. Calprotectin (a major leucocyte protein) is strongly and independently correlated with joint inflammation and damage in rheumatoid arthritis. Ann Rheum Dis (2007) 66(8):1093–7. doi: 10.1136/ard.2006.064741
124. Chen YS, Yan W, Geczy CL, Brown MA, Thomas R. Serum levels of soluble receptor for advanced glycation end products and of S100 proteins are associated with inflammatory, autoantibody, and classical risk markers of joint and vascular damage in rheumatoid arthritis. Arthritis Res Ther (2009) 11(2):R39. doi: 10.1186/ar2645
125. Berner Hammer H, Ødegård S, Syversen SW, Landewé R, van der Heijde D, Uhlig T, et al. Calprotectin (a major S100 leucocyte protein) predicts 10-year radiographic progression in patients with rheumatoid arthritis. Ann Rheum Dis (2010) 69(1):150–4. doi: 10.1136/ard.2008.103739
126. Berntzen HB, Munthe E, Fagerhol MK. The major leukocyte protein L1 as an indicator of inflammatory joint disease. Scand J Rheumatol Suppl (1988) 76:251–6. doi: 10.3109/03009748809102976
127. Bisoendial RJ, Stroes ESG, Tak PP. Critical determinants of cardiovascular risk in rheumatoid arthritis. Curr Pharm Des (2011) 17(1):21–6. doi: 10.2174/138161211795049741
128. Haringman JJ, Gerlag DM, Zwinderman AH, Smeets TJM, Kraan MC, Baeten D, et al. Synovial tissue macrophages: a sensitive biomarker for response to treatment in patients with rheumatoid arthritis. Ann Rheum Dis (2005) 64(6):834–8. doi: 10.1136/ard.2004.029751
129. Inciarte-Mundo J, Ruiz-Esquide V, Hernández MV, Cañete JD, Cabrera-Villalba SR, Ramirez J, et al. Calprotectin more accurately discriminates the disease status of rheumatoid arthritis patients receiving tocilizumab than acute phase reactants. Rheumatol (United Kingdom) (2015) 54(12):2239–43. doi: 10.1093/rheumatology/kev251
130. Jonsson MK, Sundlisæter NP, Nordal HH, Hammer HB, Aga AB, Olsen IC, et al. Calprotectin as a marker of inflammation in patients with early rheumatoid arthritis. Ann Rheum Dis (2017) 76(12):2031–7. doi: 10.1136/annrheumdis-2017-211695
131. Hurnakova J, Hulejova H, Zavada J, Hanova P, Komarc M, Mann H, et al. Relationship between serum calprotectin (S100A8/9) and clinical, laboratory and ultrasound parameters of disease activity in rheumatoid arthritis: A large cohort study. PloS One (2017) 12(8):e0183420. doi: 10.1371/journal.pone.0183420
132. Mansour HE, Abdullrhman MA, Mobasher SA, El Mallah R, Abaza N, Hamed F, et al. Serum calprotectin in rheumatoid arthritis: A promising diagnostic marker, how far is it related to activity and sonographic findings? J Med Ultrasound (2017) 25(1):40–6. doi: 10.1016/j.jmu.2016.11.001
133. Inciarte-Mundo J, Hernández MV, Ruiz-Esquide V, Cabrera-Villalba SR, Ramirez J, Cuervo A, et al. Serum calprotectin more accurately discriminates the inflammatory disease activity of rheumatoid arthritis patients receiving TNF inhibitors than acute phase reactants. Arthritis Care Res (Hoboken) (2015) (Hoboken) 68(7):899–906. doi: 10.1002/acr.22795
134. Smolen JS, Aletaha D. Interleukin-6 receptor inhibition with tocilizumab and attainment of disease remission in rheumatoid arthritis: The role of acute-phase reactants. Arthritis Rheumatol (2011) 63(1):43–52. doi: 10.1002/art.27740
135. Jarlborg M, Courvoisier DS, Lamacchia C, Martinez Prat L, Mahler M, Bentow C, et al. Serum calprotectin: a promising biomarker in rheumatoid arthritis and axial spondyloarthritis. Arthritis Res Ther (2020) 22(1):18671. doi: 10.1038/s41598-021-98255-7
136. Nielsen UB, Bruhn LV, Ellingsen T, Stengaard-Pedersen K, Hornung N. Calprotectin in patients with chronic rheumatoid arthritis correlates with disease activity and responsiveness to methotrexate. Scand J Clin Lab Invest (2018) 78(1–2):62–7. doi: 10.1080/00365513.2017.1413591
137. De Rycke L, Baeten D, Foell D, Kruithof E, Veys EM, Roth J, et al. Differential expression and response to anti-TNF? treatment of infiltrating versus resident tissue macrophage subsets in autoimmune arthritis. J Pathol (2005) 206(1):17–27. doi: 10.1002/path.1758
138. Kopec-Medrek M, Kucharz EJ. Fibulin-3 and other cartilage metabolism biomarkers in relationship to calprotectin (MRP8/14) and disease activity in rheumatoid arthritis patients treated with anti-TNF therapy. Adv Clin Exp Med (2018) 27(3):383–9. doi: 10.17219/acem/68362
139. Yunchun L, Yue W, Jun FZ, Qizhu S, Liumei D. Clinical significance of myeloid-related protein 8/14 as a predictor for biological treatment and disease activity in rheumatoid arthritis. Ann Clin Lab Sci (2018) 48(1):63–8.
140. Joshua F, Edmonds J, Lassere M. Power Doppler ultrasound in musculoskeletal disease: a systematic review. Semin Arthritis Rheumatol (2006) 36(2):99–108. doi: 10.1016/j.semarthrit.2006.04.009
141. Hammer HB, Fagerhol MK, Wien TN, Kvien TK. The soluble biomarker calprotectin (a S100 protein) is associated to ultrasonographic synovitis scores and is sensitive to change in patients with rheumatoid arthritis treated with adalimumab. Arthritis Res Ther (2011) 13(5):R178. doi: 10.1186/ar3503
142. Hurnakova J, Zavada J, Hanova P, Hulejova H, Klein M, Mann H, et al. Serum calprotectin (S100A8/9): an independent predictor of ultrasound synovitis in patients with rheumatoid arthritis. Arthritis Res Ther (2015) 17:252. doi: 10.1186/s13075-015-0764-5
143. Nordal HH, Brokstad KA, Solheim M, Halse AK, Kvien TK, Hammer HB. Calprotectin (S100A8/A9) has the strongest association with ultrasound-detected synovitis and predicts response to biologic treatment: results from a longitudinal study of patients with established rheumatoid arthritis. Arthritis Res Ther (2017) 19(1):3. doi: 10.1186/s13075-016-1201-0
144. Inciarte-Mundo J, Ramirez J, Hernández MV, Ruiz-Esquide V, Cuervo A, Cabrera-Villalba SR, et al. Calprotectin and TNF trough serum levels identify power Doppler ultrasound synovitis in rheumatoid arthritis and psoriatic arthritis patients in remission or with low disease activity. (2016) 18(1):160. doi: 10.1186/s13075-016-1032-z
145. Koenders MI, Marijnissen RJ, Devesa I, Lubberts E, Joosten LAB, Roth J, et al. Tumor necrosis factor-interleukin-17 interplay induces S100A8, interleukin-1and matrix metalloproteinases, and drives irreversible cartilage destruction in murine arthritis: Rationale for combination treatment during arthritis. Arthritis Rheumatol (2011) 63(8):2329–39. doi: 10.1002/art.30418
146. Ramírez J, Narváez JA, Ruiz-Esquide V, Hernández-Gañán J, Cuervo A, Inciarte-Mundo J, et al. Clinical and sonographic biomarkers of structural damage progression in RA patients in clinical remission: A prospective study with 12 months follow-up. Semin Arthritis Rheumatol (2017) 47(3):303–9. doi: 10.1016/j.semarthrit.2017.04.007
147. Bernardy C, Lejeune S, Courtier A, Wendling D, Berenbaum F, Nguyen MVC, et al. Calprotectin alone is not sufficient to predict response to methotrexate in early ACR/EULAR 2010 rheumatoid arthritis: Analysis of the ESPOIR cohort. Joint Bone Spine (2020) 87(1):99–100. doi: 10.1016/j.jbspin.2019.07.001
148. Choi IY, Gerlag DM, Herenius MJ, Thurlings RM, Wijbrandts CA, Foell D, et al. MRP8/14 serum levels as a strong predictor of response to biological treatments in patients with rheumatoid arthritis. Ann Rheum Dis (2015) 74(3):499–505. doi: 10.1136/annrheumdis-2013-203923
149. Nair SC, Welsing PMJ, Choi IYK, Roth J, Holzinger D, Bijlsma JWJ, et al. A personalized approach to biological therapy using prediction of clinical response based on MRP8/14 serum complex levels in rheumatoid arthritis patients. PloS One (2016) 11(3):e0152362. doi: 10.1371/journal.pone.0152362
150. Martínez-Feito A, Plasencia-Rodríguez C, Navarro-Compán V, Jochems A, Hernández-Breijo B, Peiteado D, et al. Low serum calprotectin levels correlate with the presence of biological drugs after the first year of treatment in patients with rheumatoid arthritis. Scand J Clin Lab Invest (2019) 79(7):538–40. doi: 10.1080/00365513.2019.1669813
151. Abildtrup M, Kingsley GH, Scott DL. Calprotectin as a biomarker for rheumatoid arthritis: a systematic review. J Rheumatol (2015) 42(5):760–70. doi: 10.3899/jrheum.140628
152. Aghdashi MA, Seyedmardani S, Ghasemi S, Khodamoradi Z. Evaluation of serum calprotectin level and disease activity in patients with rheumatoid arthritis. Curr Rheumatol Rev (2019) 15(4):316–20. doi: 10.2174/1573397115666190122113221
153. Bechman K, Tweehuysen L, Garrood T, Scott DL, Cope AP, Galloway JB, et al. Flares in rheumatoid arthritis patients with low disease activity: Predictability and association with worse clinical outcomes. J Rheumatol (2018) 45(11):1515–21. doi: 10.3899/jrheum.171375
154. Tweehuysen L, Den Broeder N, Van Herwaarden N, Joosten LAB, Van Lent PL, Vogl T, et al. Predictive value of serum calprotectin (S100A8/A9) for clinical response after starting or tapering anti-TNF treatment in patients with rheumatoid arthritis. RMD Open (2018) 4(1):e000654. doi: 10.1136/rmdopen-2018-000654
155. De Moel EC, Rech J, Mahler M, Roth J, Vogl T, Schouffoer A, et al. Circulating calprotectin (S100A8/A9) is higher in rheumatoid arthritis patients that relapse within 12 months of tapering anti-rheumatic drugs. Arthritis Res Ther (2019) 21(1):268. doi: 10.1186/s13075-019-2064-y
156. Cesaro A, Anceriz N, Plante A, Pagé N, Tardif MR, Tessier PA. An inflammation loop orchestrated by S100A9 and calprotectin is critical for development of arthritis. PloS One (2012) 7(9):e45478. doi: 10.1371/journal.pone.0045478
Keywords: calprotectin, rheumatoid arthritis, biomarker, acute phase reactants, CRP - C-reactive protein
Citation: Inciarte-Mundo J, Frade-Sosa B and Sanmartí R (2022) From bench to bedside: Calprotectin (S100A8/S100A9) as a biomarker in rheumatoid arthritis. Front. Immunol. 13:1001025. doi: 10.3389/fimmu.2022.1001025
Received: 22 July 2022; Accepted: 19 October 2022;
Published: 03 November 2022.
Edited by:
Carmelo Carmona-Rivera, National Institute of Arthritis and Musculoskeletal and Skin Diseases (NIH), United StatesReviewed by:
Liam O’Neil, University of Manitoba, CanadaCopyright © 2022 Inciarte-Mundo, Frade-Sosa and Sanmartí. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Raimon Sanmartí, c2FubWFydGlAY2xpbmljLmNhdA==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.