 Nuria Balaguer1
Nuria Balaguer1 Emilia Mateu-Brull1Jose Antonio Martínez-Conejero2Ana Cervero2Roser Navarro3Jorge Jiménez-Almazán3
Emilia Mateu-Brull1Jose Antonio Martínez-Conejero2Ana Cervero2Roser Navarro3Jorge Jiménez-Almazán3 Miguel Milán1*
Miguel Milán1*- 1Prenatal Diagnosis Department, Igenomix Spain Lab, Valencia, Spain
- 2Pre-implantation Genetic Testing for Monogenic Diseases Department, Igenomix Spain Lab, Valencia, Spain
- 3Bioinformatics Department, Igenomix Spain Lab, Valencia, Spain
Sex discordance between cell-free DNA (cfDNA) testing and ultrasound examination is rare but can cause significant patient discomfort and uncertainty. Here, we present two clinical cases where a closer examination of raw sequencing data allowed us to anticipate possible discrepancies caused by the insertion of Y-chromosome regions into the maternal genome. We used Illumina’s VeriSeq NIPT Solution v2 and a proprietary bioinformatics pipeline to analyze cfDNA in the maternal bloodstream. Paired-end sequencing data were aligned to the reference genome (hg19). Non-duplicated aligned reads were aggregated into 100-kb bins, adjusted for CG bias, and further aggregated into 5-Mb windows. Z-scores were calculated for autosomes, sex chromosomes, and 5-Mb bins. The two clinical cases were classified as low-risk male fetuses according to the primary statistics (case A: NCVx = 0.3; NCVy = 40.6; native fetal fraction (FFi) = 5.1%, and case B: NCVx = −0.3, NCVy = 40.7, FFi = 10.8%); however, the Y-chromosome-based FF (FFy) was significantly lower than the default FF estimate (FFy ≅ 2% in both cases). Plots of X and Y chromosome Z-scores for each 5-Mb bin, according to genomic position, identified bins with Z-scores significantly higher than those expected for any pregnancy with a male fetus. The genomic coordinates of these bins overlapped with the amelogenin (AMELY) and protein kinase Y-linked (PRKY) genes, respectively. Amplification of these regions in the DNA isolated from the white blood cells fraction confirmed the presence of Y-chromosome insertions in the maternal genome. This study highlights a new source of discrepancy in cfDNA testing due to maternal genomic variations. These findings suggest the need for improvements to current bioinformatics pipelines to identify and exclude possible maternal perturbations from the classification algorithms used for aneuploidy and sex calls.
Introduction
Cell-free DNA (cfDNA) testing has been established in many countries as the first-line screening method for major aneuploidies in early pregnancy, given the high level of sensitivity and specificity compared to traditional first-trimester biochemical screening (Zhang et al., 2015).
The ability of cfDNA testing to analyze sex chromosomes as early as 10 weeks of gestation has contributed to its popularity among parents desiring early information regarding fetal sex. A considerable number of patients prioritize fetal sex prediction over aneuploidy screening when deciding whether to undergo cfDNA testing (Stevens et al., 2023). Professional guidelines agree that patients undergoing cfDNA testing should receive pre- and post-test counseling, which would include a discussion of the possibility of false positives, sex discrepancies, incidental findings, and the variable expressivity and penetrance of the conditions screened for by the test, particularly sex chromosome aneuploidies and rare autosomal trisomies (Dungan et al., 2023; Wilson et al., 2013; Rose et al., 2022). This practice ensures the responsible application of cfDNA testing and enables evidence-based decision-making capacity for the prospective mother.
Discrepant results between cfDNA testing and ultrasound findings or other clinical information may originate from technical limitations and complex biological mechanisms occurring during the early stages of embryonic development. Sources of human error include blood sample mislabeling, laboratory methodological limitations, and suboptimal visualization of the external genitalia associated with limited ultrasound imaging performance at early gestational ages. Biological reasons for discordance include the presence of a vanishing twin, fetal-placental mosaicism for sex chromosome aneuploidies, a maternal transplant from a male donor, disorders of sexual development, or other fetal anomalies associated with abnormal or ambiguous external genitalia (Dhamankar et al., 2020).
Despite the numerous biological reasons already described, no scientific literature explicitly addresses cases where the insertion of Y chromosomal genetic material into the maternal genome represents the direct cause of a sex discrepancy in the context of cfDNA testing. This case report describes two instances where this specific mechanism was responsible for the observed discordance.
Case report
Patient 1 (case A)
A 38-year-old patient (gravida 1) underwent assisted reproductive treatment at the end of December 2021, in which only one embryo was transferred. There were no previous abortions and no availability of preimplantation genetic testing for aneuploidy analysis before the transfer. At 11 weeks of gestation, the patient opted for cfDNA testing (NACE 24 test, Igenomix, Valencia, Spain) to screen for aneuploidies across the 23 pairs of chromosomes and to study copy number variations (deletions/duplications) larger than 7 Mb. The native analysis algorithm (Illumina VeriSeq NIPT Solution v2) classified the pregnancy as low risk for all conditions analyzed. The fetal sex was classified as male (XY) with an estimated native fetal fraction (FFi) of 5.0%; however, the Y-chromosome-based FF (FFy) was 2.0%. The normalized chromosome values for the sex chromosomes (NCVx and NCVy) were 0.3 and 40.6. Given that these values were significantly lower than that expected for a male fetus with a 5.0% FFi (Figure 1), the possibility of underlying clinical factors interfering with the result was discussed with the physician. The prescribing physician reported none of the previously described factors, such as altered known maternal karyotype, transfusions or transplants from a male donor, or the presence of a vanishing twin.
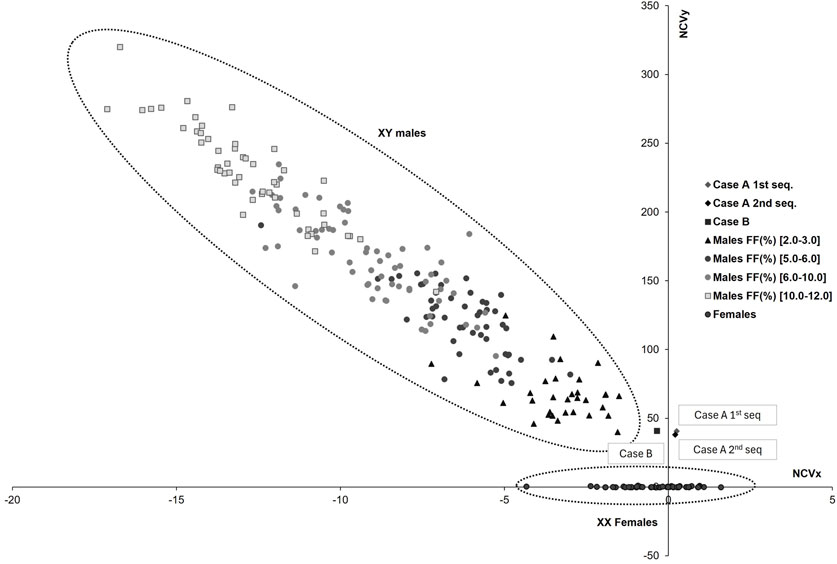
Figure 1. Sex chromosomes plot for different fetal fraction ranges. The horizontal axis represents the normalized chromosome value for the X chromosome (NCVx), and the vertical axis represents the normalized chromosome value for the Y chromosome (NCVy). The dotted lines indicate the normal range for female and male euploid samples across different fetal fraction (FF) ranges (





An ultrasound examination at 16 weeks of gestation revealed the presence of female genitalia in the fetus. To rule out a technical issue, cfDNA testing was repeated, yielding results consistent with the initial test (NCVx = 0.2; NCVy = 38.0; FFi = 5.6%; FFy = 1.9%).
To eliminate the possibility of a technical problem associated with the Illumina platform’s analysis algorithm, informed consent was obtained from the patient and the prescribing physician to re-analyze the data using an alternative bioinformatics algorithm based on that described by Bayindir et al., in 2015 (Bayindir et al., 2015). Paired-end sequencing data were aligned to the reference genome (hg19). Non-duplicated aligned reads were aggregated into 100-kb bins, adjusted for CG bias, and further aggregated into 5-Mb windows. Z-scores were calculated for autosomes, sex chromosomes, and 5-Mb bins. Sex was determined by verifying the presence of Y-chromosome-specific reads. Samples with ≥3 Y-specific bins containing >1 read were classified as male pregnancies. Two estimators were used for FF: one estimator based on the SeqFF method (Kim et al., 2015) and another estimator specific to the Y chromosome.
In the first and second sequencing, the Y chromosome Z-scores were 30 and 25, respectively, compatible with a male sex classification but still lower than expected for SeqFF-based estimated FFs (6.1% and 5.5%) [first seq: ZscoreX = −1.2; ZscoreY = 30.0; SeqFF = 6.1%; FFy = 3.0%; second seq: ZscoreX = −0.7; ZscoreY = 25.4; SeqFF = 5.5%; FFy = 2.7%].
An analysis of the distribution of DNA fragments along the Y chromosome revealed that DNA from this chromosome was not homogeneously present; Y-DNA fragments were concentrated solely to the cytogenetic region Yp11,2 covering the genomic coordinates ChrY:6,400,000_6,900,000 (GRCh37/hg19) (Figure 2A). A comparison with samples analyzed in the same set, including 50 fetuses classified as male and 50 as females without fetal sex discrepancies by ultrasound, revealed that fetuses classified by the native algorithm as female had no aligned DNA fragments in this genomic region. In contrast, fetuses classified as male exhibited a ratio of aligned Y-DNA fragments to total reads much lower than that found in the case described in this report (8 fold lower in the general male fetus sample set vs. case A) (Figure 2B).
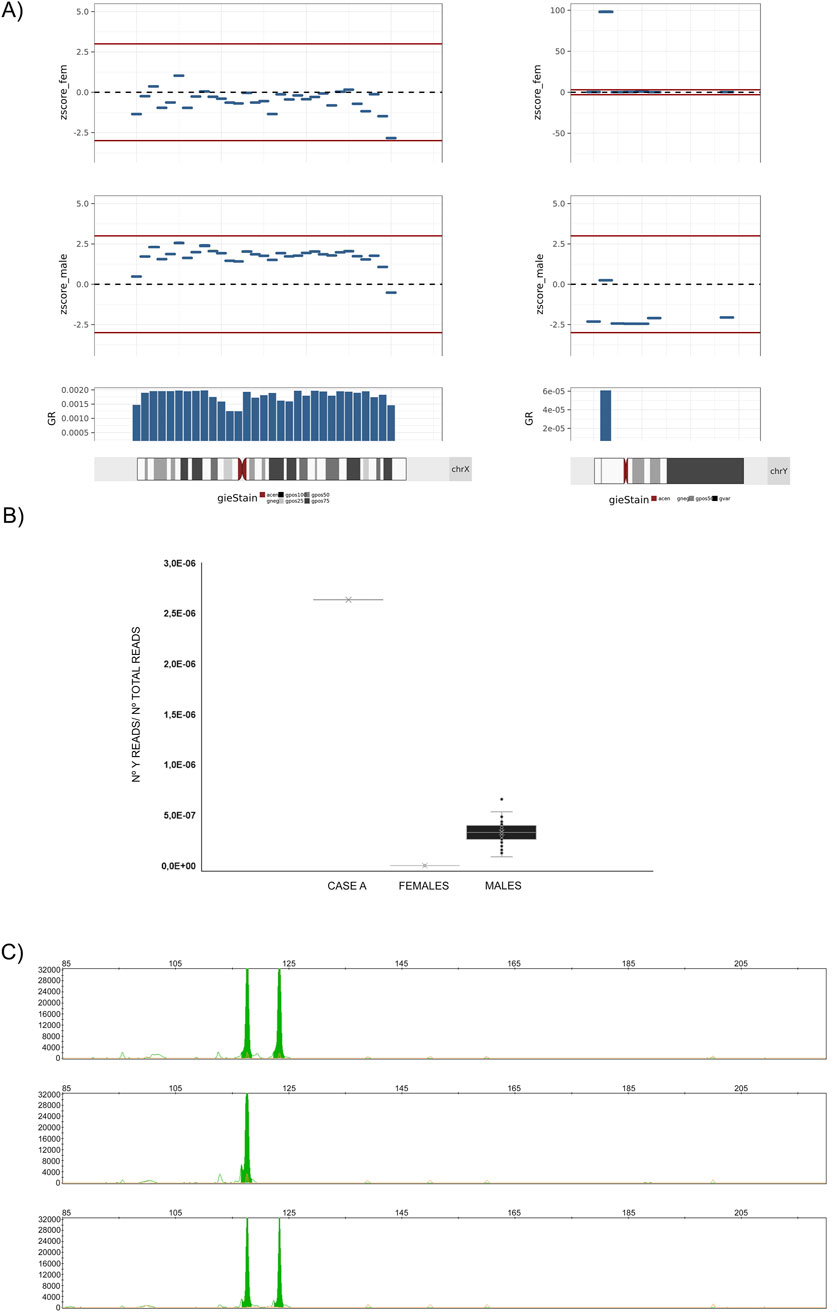
Figure 2. Analysis of the overrepresented genetic material belonging to the Y-chromosome. Case (A). (A) Left: Graphical plot of 5-Mb bin Z-scores across the X chromosome. Right: Graphical plot of Chr. Y 5-Mb z-scores across the Y chromosome. In all cases, the bin Z-score values have been calculated with regard to a female reference of normalcy (zscore_fem), or with regard to a male reference of normalcy (zscore_male). Between positions 5–10 Mb, a bin with a Z-score of approximately 100 is observed, a value 10 times higher than expected for a male fetus with a native fetal fraction (FFi) of around 5%. (B) Graphical representation of the ratio of Y chromosome reads to total reads within the genomic region of the Y chromosome spanning positions ChrY: 6,400,000–7400,000 (GRCh37/hg19) for (from left to right): CASE A, a set of 50 samples from pregnant patients with female fetuses (FFs between 5% and 12%), and a set of 50 samples from pregnant patients with male fetuses (FFs between 5% and 12%). In case A, the ratio exceeds by more than ten-fold the number of reads aligned to the Y chromosome in samples from patients with fetuses classified by the analysis algorithm as XY. (C) PCR amplification of the amelogenin (green peak) containing regions and fragment analysis of PCR products by capillary electrophoresis on AB3500 (ThermoFisher) and GeneMapper (rr). Top: patient; Middle: female reference; Bottom: male reference. The 119 bp peak represents AMELX, and the 124 bp peak represents AMELY. The marker under each allele peak represents the number of repeats.
The affected genomic location included the amelogenin gene (AMELY) and part of the Transducing-beta-like 1 (TBL1Y) gene (exons 1–6). To elucidate whether the overrepresentation of fragments was caused by an insertion of Y-chromosome genetic material into the maternal genome, fluorescent PCR was performed on white blood cell DNA using specifically designed primers. The amplification results revealed the presence of the expected female variant AMELX and the unexpected male variant AMELY in the maternal genome (Figure 2C).
Patient 2 (case B)
A 41-year-old patient (gravida 1) became spontaneously pregnant at the end of January 2023. No previous abortions or other clinically relevant information were described in the medical history. At 14 weeks of gestation, the patient opted for extended cfDNA testing (NACE 24 test, Igenomix, Valencia, Spain). A low-risk result was obtained for all conditions evaluated, and the sex of the fetus was classified as male (NCVx = −0.3; NCVy = 40.7; FFi = 10.8%; FFy = 2.0%).
As it happened in Case A, an ultrasound examination at 16 weeks of gestation revealed the presence of female genitalia in the fetus. Based on our experience with case A, we obtained consent from the physician and the patient to conduct an exploratory analysis using the alternative bioinformatics pipeline without performing a second sequencing. This analysis aimed to evaluate the presence of aligned reads in specific regions of the Y-chromosome. The Z-score value for the Y chromosome was 40.02 with a SeqFF-based estimated FF of 8.8% and an FFy of 3.5%; however, the Y-DNA fragment distribution analysis revealed alignment with the cytogenetic region Yp11.2, specifically between genomic coordinates ChrY: 7,000,000–7,500,000 (GRCh37/hg19) (Figure 3A). This region contains the pseudogene Protein Kinase Y-linked (PRKY), located in the Yp12 region. Similarly, the ratio of the number of Y-DNA readings to total readings in this region was significantly higher than that identified in a set of 50 samples classified as male fetuses (4.5 fold lower in the general male fetus sample set vs. case B) (Figure 3B).

Figure 3. Analysis of the overrepresented genetic material belonging to the Y-chromosome. Case (B). (A) Left: Graphical plot of 5-Mb bin Z-scores across the X chromosome; right: Graphical plot of Chr. Y-5 Mb Z-scores across the Y chromosome. In all cases, the bin Z-score values have been calculated with regard to a female reference of normalcy (zscore_fem), or with regard to a male reference of normalcy (zscore_male). Between positions 5–10 Mb, a bin with a Z-score of approximately 125 is observed, a value 10 times higher than expected for a male fetus with a native fetal fraction (FFi) of around 11%. (B) Graphical representation of the ratio of Y chromosome reads to total reads within the genomic region of the Y chromosome spanning positions ChrY: 6,400,000–7400,000 (GRCh37/hg19) for (from left to right): CASE B, a set of 50 samples from pregnant patients with female fetuses (FFs between 5% and 12%), and a set of 50 samples from pregnant patients with male fetuses (FFs between 5% and 12%). In case B, the ratio exceeds by more than ten-fold the number of reads aligned to the Y chromosome in samples from patients with fetuses classified by the analysis algorithm as XY. (C) PCR amplification of amelogenin and PRKY and the Y:7,350,250–7,350,345 containing regions and fragment analysis of PCR products by capillary electrophoresis on AB3500 (ThermoFisher) and GeneMapper (rr): C_1 and C_4: patient; C_2 and C_5: female reference; C_3 and C_6: male reference. Green peak: The 119 bp peak represents AMELX, and the 124 bp peak represents AMELY. The blue peak at 208 bp represents PRKY and the blue peak at 96 bp is a region in chromosome Y at Y:7,350,241–7,350,341.
After reporting the incidental findings to the prescribing physician and the patient, the decision was made to evaluate the patient for the previously described insertion. Fluorescent PCR on white blood cell DNA confirmed the insertion of the PRKY-including region into the patient’s genome (Figure 3C).
Discussion
The prenatal diagnosis of sex discrepancies has historically relied on identifying ambiguous genitalia during ultrasound evaluations or observing genotype-phenotype discordance in cases involving preimplantation genetic diagnosis or invasive prenatal testing. The advent of cfDNA testing has significantly enhanced our ability to detect such discrepancies with greater precision. Estimates indicate that sex discrepancies occur in approximately 1 in every 1,500–2,000 pregnancies (Smet et al., 2020), a figure highlighted by the widespread adoption of cfDNA testing in modern prenatal screening.
We report a novel observation: sex discrepancies between cfDNA testing and ultrasound due to the insertion of Y-chromosome genetic material into the maternal genome. By analyzing cfDNA sequencing data and partitioning the Y-chromosome into 100-kb bins aggregated into 5-Mb windows, we identified a significant overrepresentation of reads aligned to the Yp11.2 cytoregion in two cases, exceeding typical male fetal levels by ten-fold.
In the first case, the insertion encompassed Y-chromosome material containing the AMELY gene and a portion of the coding region of the TBL1Y gene. AMELY codes for a matrix protein forming tooth enamel, constituting 90% of the total organic content (Richard et al., 2007). PCR products generated from AMELX and AMELY amplification can be discriminated using primers flanking a 6-base pair deletion in the gene’s first intron and differ between the two versions based on sex chromosomes. Sequence differences between AMELX and AMELY have been used to differentiate males from females with ambiguous phenotypes or to establish the gender in biological material for various purposes (Tozzo et al., 2013; Mannucci et al., 1994; Sullivan et al., 1993). In the second case, we described an insertion involving the PRKY gene, a member of the cAMP-dependent serine/threonine protein kinase gene family that shares homology with PRKX from Xp22.3 (Schiebel et al., 1997). PRKY and its X-chromosome homolog PRKX participate in various cellular processes, including signaling pathways implicated in cell growth and differentiation.
Notably, these insertions into the maternal genome are rare, with few documented cases in the scientific literature of females with a 46, XX karyotype harboring such Y-chromosome gene insertions. Among the few reported cases, Stapleton et al. (2008) identified a family in which three females carried both the male and female variants of the AMELY gene, suggesting Mendelian inheritance and highlighting the problem of using only AMELY as a sex-specific marker (Stapleton et al., 2008). Cases of AMELY-negative males are more frequently detected worldwide; the genetic mechanisms underlying AMELY dropout involve deletions of different sizes, encompassing the AMELY locus, and mutations in the primer-binding region of the AMELY allele (Jobling et al., 2007; Butler, 2011). Deletions in the Yp11.2 region, a major cause of the AMELY null allele, are often combined with the absence of adjacent Y-STR loci DYS456 and/or DYS458 (Jobling et al., 2007; Cadenas et al., 2007; Ma et al., 2012; Chen et al., 2014; Chang et al., 2007; Ou et al., 2012; Kumagai et al., 2008; Turrina et al., 2009; Borovko et al., 2015).
The significance of these findings lies not only in the phenotypic and clinical implications for carrier patients but also in the potential to enhance fetal sex prediction in current bioinformatic analysis pipelines for cfDNA testing. The evolution of bioinformatic pipelines has led to remarkable improvements in the sensitivity and specificity of cfDNA testing for fetal sex determination and aneuploidy detection. Early PCR-based methods displayed sensitivities and specificities in the range of 95%–98% (Lo et al., 1998). The advent of massively parallel sequencing and advanced bioinformatics have improved these metrics to over 99% for fetal sex determination (Kotsopoulou et al., 2015).
Several methods have been proposed to predict fetal aneuploidies, typically involving genome binning, normalization techniques, and Z-score calculations to evaluate variation in the normalized fragment count in each bin. Comparisons are made with euploid reference samples (e.g., cn. MOPS (Talevich et al., 2016) and CNVkit (Klambauer et al., 2012), between sample variations (e.g., Wisecondor (Straver et al., 2014) and WisecondorX (Raman et al., 2019), or through reference-free approaches (e.g., FREEC (Boeva et al., 2011), QDNAseq (Scheinin et al., 2014), and BIC-seq2 (Xi et al., 2016). Although these tools facilitate cfDNA testing analysis, they remain challenging to implement in a clinical setting in many laboratories worldwide due to intensive data processing and the need for advanced bioinformatics skills. Additionally, the diversity of technologies and algorithms employed can prompt significant variability in results, and we lack a comprehensive quantification of this variation (Duboc et al., 2022). Because of the inherent differences among methodologies, clinical laboratories should leverage the bioinformatics available to help navigate complex cases with clinicians.
Therefore, we must develop a protocol for cases with an anticipated sex discrepancy, even though reaching a consensus on the best algorithm or platform for maximum sensitivity and specificity remains challenging. For prenatally reported fetal sex discrepancies despite ultrasound examination by an experienced examiner after 16 weeks of gestation, a complete history and physical examination always represent the initial steps to guide further evaluation. In parallel, we must rule out human error by confirming correct sample labeling.
We recommend conducting a detailed review of existing data before repeating sequencing on an independent sample to rule out potential sample errors. This review should focus on identifying overrepresented Y chromosome regions compared to that expected for a male fetus. In the case of identifying such overrepresentation, the prescribing physician must be informed, and the maternal genome should be investigated for possible insertions of Y-chromosome genetic material before performing any unnecessary invasive procedures.
We acknowledge there are some important limitations for the proposed protocol. The occurrences described in this article are rare, making generalizing findings to broader populations challenging. The advanced bioinformatics methods employed may not be readily accessible in all clinical settings, potentially limiting widespread adoption of the proposed protocols. And the reliance on cfDNA testing and bioinformatics could lead to misinterpretations of results if not carefully managed, particularly in the absence of confirmatory tests. Thus, major platforms offering cfDNA testing should incorporate these considerations into their decision algorithms. Based on the evidence presented here, the presence of isolated bins on the Y chromosome with Z-scores elevated to levels not expected in pregnancies bearing a male fetus should be carefully analyzed when making a final sex calling. This approach would prevent undesirable situations and alleviate the anxiety of the expectant mother concerning potential clinical issues for her or the fetus.
If none of the aforementioned conditions are confirmed, the physician should discuss the case within the context of a multidisciplinary team. This team should be aware of all developments and findings to prepare a comprehensive care pathway for the family. Based on this, a protocol of action could be defined, including a series of tests such as 1) direct assessment of the fetal sex chromosome distribution (ploidy, copy number variations, or mosaicism) through amniocentesis accompanied by a genetic counseling interview; 2) further investigation for Y-chromosome-specific material using an SRY probe which may help in evaluating sex chromosome anomalies, including translocation to the X chromosome or autosomes in XX individuals or mosaicism; and/or 3) analysis for single-gene conditions causing disorders of sexual differentiations, such as congenital adrenal hyperplasia or androgen insensitivity and a wide range of genetic syndromes that impact external genitalia.
Conclusion
These findings underscore the complexity of prenatal sex determination and the potential of genetic anomalies in influencing cfDNA testing results. By incorporating detailed bioinformatic reviews and considering maternal genomic contributions, we can improve the accuracy of fetal sex prediction and minimize unnecessary invasive procedures, ultimately enhancing prenatal care and reducing anxiety for expectant mothers.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
Ethical approval was not required for the studies involving humans because It is a case report where we report an incidental finding derived from the intended purpose of the NIPT and we have the informed consent of the patient to use the data for publishing. The studies were conducted in accordance with the local legislation and institutional requirements. The human samples used in this study were acquired from a blood sample to conduct a NIPT in a clinical context. Written informed consent to participate in this study was not required from the participants or the participants’ legal guardians/next of kin in accordance with the national legislation and the institutional requirements. Written informed consent was obtained from the individual(s) for the publication of any potentially identifiable images or data included in this article.
Author contributions
NB Cuenca: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Project administration, Supervision, Validation, Visualization, Writing–original draft, Writing–review and editing. EM-B: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Project administration, Supervision, Validation, Visualization, Writing–original draft, Writing–review and editing. JM-C: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing–original draft, Writing–review and editing. AC: Data curation, Formal Analysis, Investigation, Methodology, Resources, Validation, Visualization, Writing–original draft, Writing–review and editing. RN: Data curation, Formal Analysis, Investigation, Methodology, Software, Validation, Visualization, Writing–original draft, Writing–review and editing. JJ-A: Data curation, Formal Analysis, Investigation, Methodology, Software, Supervision, Validation, Visualization, Writing–original draft, Writing–review and editing. MM: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing–original draft, Writing–review and editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. Igenomix Lab S.L.U. financially supported the study.
Acknowledgments
We are grateful to everyone who helped us with article elaboration.
Conflict of interest
The authors declare that this research was conducted without commercial or financial relationships that could create a conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Bayindir, B., Dehaspe, L., Brison, N., Brady, P., Ardui, S., Kammoun, M., et al. (2015). Noninvasive prenatal testing using a novel analysis pipeline to screen for all autosomal fetal aneuploidies improves pregnancy management. Eur. J. Hum. Genet. 23 (10), 1286–1293. doi:10.1038/ejhg.2014.282
Boeva, V., Zinovyev, A., Bleakley, K., Vert, J. P., Janoueix-Lerosey, I., Delattre, O., et al. (2011). Control-free calling of copy number alterations in deep-sequencing data using GC-content normalization. Bioinformatics 27 (2), 268–269. doi:10.1093/bioinformatics/btq635
Borovko, S., Shyla, A., Korban, V., and Borovko, A. (2015). Amelogenin test abnormalities revealed in Belarusian population during forensic DNA analysis. Forensic Sci. Int. Genet. 15, 98–104. doi:10.1016/j.fsigen.2014.10.014
Cadenas, A. M., Regueiro, M., Gayden, T., Singh, N., Zhivotovsky, L. A., Underhill, P. A., et al. (2007). Male amelogenin dropouts: phylogenetic context, origins and implications. Forensic Sci. Int. 166 (2-3), 155–163. doi:10.1016/j.forsciint.2006.05.002
Chang, Y. M., Perumal, R., Keat, P. Y., Yong, R. Y. Y., Kuehn, D. L. C., and Burgoyne, L. (2007). A distinct Y-STR haplotype for Amelogenin negative males characterized by a large Yp11.2 (DYS458-MSY1-AMEL-Y) deletion. Forensic Sci. Int. 166 (2), 115–120. doi:10.1016/j.forsciint.2006.04.013
Chen, W., Wu, W., Cheng, J., Zhang, Y., Chen, Y., and Sun, H. (2014). Detection of the deletion on Yp11.2 in a Chinese population. Forensic Sci. Int. Genet. 8 (1), 73–79. doi:10.1016/j.fsigen.2013.07.003
Dhamankar, R., DiNonno, W., Martin, K. A., Demko, Z. P., and Gomez-Lobo, V. (2020). Fetal sex results of noninvasive prenatal testing and differences with ultrasonography. Obstet. Gynecol. 135 (5), 1198–1206. doi:10.1097/AOG.0000000000003791
Duboc, V., Pratella, D., Milanesio, M., Boudjarane, J., Descombes, S., Paquis-Flucklinger, V., et al. (2022). NiPTUNE: an automated pipeline for noninvasive prenatal testing in an accurate, integrative and flexible framework. Brief. Bioinform 23 (1), bbab380. doi:10.1093/bib/bbab380
Dungan, J. S., Klugman, S., Darilek, S., Malinowski, J., Akkari, Y. M. N., Monaghan, K. G., et al. (2023). Noninvasive prenatal screening (NIPS) for fetal chromosome abnormalities in a general-risk population: an evidence-based clinical guideline of the American College of Medical Genetics and Genomics (ACMG). Genet. Med. 25 (2), 100336. doi:10.1016/j.gim.2022.11.004
Jobling, M. A., Lo, I. C., Turner, D. J., Bowden, G. R., Lee, A. C., Xue, Y., et al. (2007). Structural variation on the short arm of the human Y chromosome: recurrent multigene deletions encompassing Amelogenin Y. Hum. Mol. Genet. 16 (3), 307–316. doi:10.1093/hmg/ddl465
Kim, S. K., Hannum, G., Geis, J., Tynan, J., Hogg, G., Zhao, C., et al. (2015). Determination of fetal DNA fraction from the plasma of pregnant women using sequence read counts. Prenat. Diagn 35 (8), 810–815. doi:10.1002/pd.4615
Klambauer, G., Schwarzbauer, K., Mayr, A., Clevert, D. A., Mitterecker, A., Bodenhofer, U., et al. (2012). cn.MOPS: mixture of Poissons for discovering copy number variations in next-generation sequencing data with a low false discovery rate. Nucleic Acids Res. 40 (9), e69. doi:10.1093/nar/gks003
Kotsopoulou, I., Tsoplou, P., Mavrommatis, K., and Kroupis, C. (2015). Non-invasive prenatal testing (NIPT): limitations on the way to become diagnosis. Diagn. Berl. 2 (3), 141–158. doi:10.1515/dx-2015-0002
Kumagai, R., Sasaki, Y., Tokuta, T., Biwasaka, H., and Aoki, Y. (2008). DNA analysis of family members with deletion in Yp11.2 region containing amelogenin locus. Leg. Med. 10 (1), 39–42. doi:10.1016/j.legalmed.2007.05.009
Lo, Y. M., Tein, M. S., Lau, T. K., Haines, C. J., Leung, T. N., Poon, P. M., et al. (1998). Quantitative analysis of fetal DNA in maternal plasma and serum: implications for noninvasive prenatal diagnosis. Am. J. Hum. Genet. 62 (4), 768–775. doi:10.1086/301800
Ma, Y., Kuang, J. Z., Zhang, J., Wang, G. M., Wang, Y. J., Jin, W. M., et al. (2012). Y chromosome interstitial deletion induced Y-STR allele dropout in AMELY-negative individuals. Int. J. Leg. Med. 126 (5), 713–724. doi:10.1007/s00414-012-0720-8
Mannucci, A., Sullivan, K. M., Ivanov, P. L., and Gill, P. (1994). Forensic application of a rapid and quantitative DNA sex test by amplification of the X-Y homologous gene amelogenin. Int. J. Leg. Med. 106 (4), 190–193. doi:10.1007/BF01371335
Ou, X., Chen, W., Chen, H., Zhao, F., Zheng, J., Tong, D., et al. (2012). Null alleles of the X and Y chromosomal amelogenin gene in a Chinese population. Int. J. Leg. Med. 126 (4), 513–518. doi:10.1007/s00414-011-0594-1
Raman, L., Dheedene, A., De Smet, M., Van Dorpe, J., and Menten, B. (2019). WisecondorX: improved copy number detection for routine shallow whole-genome sequencing. Nucleic Acids Res. 47 (4), 1605–1614. doi:10.1093/nar/gky1263
Richard, B., Delgado, S., Gorry, P., and Sire, J. Y. (2007). A study of polymorphism in human AMELX. Arch. Oral Biol. 52 (11), 1026–1031. doi:10.1016/j.archoralbio.2007.06.001
Rose, N. C., Barrie, E. S., Malinowski, J., Jenkins, G. P., McClain, M. R., LaGrave, D., et al. (2022). Systematic evidence-based review: the application of noninvasive prenatal screening using cell-free DNA in general-risk pregnancies. Genet. Med. 24 (7), 1992–2091. doi:10.1016/j.gim.2022.07.002
Scheinin, I., Sie, D., Bengtsson, H., van de Wiel, M. A., Olshen, A. B., van Thuijl, H. F., et al. (2014). DNA copy number analysis of fresh and formalin-fixed specimens by shallow whole-genome sequencing with identification and exclusion of problematic regions in the genome assembly. Genome Res. 24 (12), 2022–2032. doi:10.1101/gr.175141.114
Schiebel, K., Winkelmann, M., Mertz, A., Xu, X., Page, D. C., Weil, D., et al. (1997). Abnormal XY interchange between a novel isolated protein kinase gene, PRKY, and its homologue, PRKX, accounts for one third of all (Y+)XX males and (Y-)XY females. Hum. Mol. Genet. 6 (11), 1985–1989. doi:10.1093/hmg/6.11.1985
Smet, M. E., Scott, F. P., and McLennan, A. C. (2020). Discordant fetal sex on NIPT and ultrasound. Prenat. Diagn 40 (11), 1353–1365. doi:10.1002/pd.5676
Stapleton, P. M., Lai, D., Millar, C. D., Wu, E., Andres, M., and Love, D. R. (2008). Discovery of three related females who type XY at the amelogenin locus. Forensic Sci. Int. Genet. Suppl. Ser. 1 (1), 577–579. doi:10.1016/j.fsigss.2007.08.017
Stevens, C., Llorin, H., Gabriel, C., Mandigo, C., Gochyyev, P., and Studwell, C. (2023). Genetic counseling for fetal sex prediction by NIPT: challenges and opportunities. J. Genet. Couns. 32 (5), 945–956. doi:10.1002/jgc4.1703
Straver, R., Sistermans, E. A., Holstege, H., Visser, A., Oudejans, C. B., and Reinders, M. J. (2014). WISECONDOR: detection of fetal aberrations from shallow sequencing maternal plasma based on a within-sample comparison scheme. Nucleic Acids Res. 42 (5), e31. doi:10.1093/nar/gkt992
Sullivan, K. M., Mannucci, A., Kimpton, C. P., and Gill, P. (1993). A rapid and quantitative DNA sex test: fluorescence-based PCR analysis of X-Y homologous gene amelogenin. Biotechniques 15 (4), 636–641.
Talevich, E., Shain, A. H., Botton, T., and Bastian, B. C. (2016). CNVkit: genome-wide copy number detection and visualization from targeted DNA sequencing. PLoS Comput. Biol. 12 (4), e1004873. doi:10.1371/journal.pcbi.1004873
Tozzo, P., Giuliodori, A., Corato, S., Ponzano, E., Rodriguez, D., and Caenazzo, L. (2013). Deletion of amelogenin Y-locus in forensics: literature revision and description of a novel method for sex confirmation. J. Forensic Leg. Med. 20 (5), 387–391. doi:10.1016/j.jflm.2013.03.012
Turrina, S., Filippini, G., and De Leo, D. (2009). Evaluation of deleted region from Yp11.2 of two amelogenin negative related males. Forensic Sci. Int. Genet. Suppl. Ser. 2 (1), 240–241. doi:10.1016/j.fsigss.2009.08.157
Wilson, K. L., Czerwinski, J. L., Hoskovec, J. M., Noblin, S. J., Sullivan, C. M., Harbison, A., et al. (2013). NSGC practice guideline: prenatal screening and diagnostic testing options for chromosome aneuploidy. J. Genet. Couns. 22 (1), 4–15. doi:10.1007/s10897-012-9545-3
Xi, R., Lee, S., Xia, Y., Kim, T. M., and Park, P. J. (2016). Copy number analysis of whole-genome data using BIC-seq2 and its application to detection of cancer susceptibility variants. Nucleic Acids Res. 44 (13), 6274–6286. doi:10.1093/nar/gkw491
Keywords: sex discordance, y-chromosome, maternal abnormalities, non-invasive prenatal testing (NIPT), cell-free (fetal) DNA, case-report
Citation: Balaguer N, Mateu-Brull E, Martínez-Conejero JA, Cervero A, Navarro R, Jiménez-Almazán J and Milán M (2025) Tell me y: anticipation of sex discrepancies in cell-free DNA testing due to maternal genetic abnormalities: a case report. Front. Genet. 15:1502287. doi: 10.3389/fgene.2024.1502287
Received: 26 September 2024; Accepted: 30 December 2024;
Published: 20 January 2025.
Edited by:
Stephen J. Bush, Xi’an Jiaotong University, ChinaReviewed by:
Mathilde Pacault, CHU d'Angers, FranceMuhammad Afzal, Pak-Austria Fachhochschule Institute of Applied Sciences and Technology, Pakistan
Samantha Caldwell, Laboratory Corporation of America Holdings (LabCorp), United States
Copyright © 2025 Balaguer, Mateu-Brull, Martínez-Conejero, Cervero, Navarro, Jiménez-Almazán and Milán. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Miguel Milán, bWlndWVsLm1pbGFuQGlnZW5vbWl4LmNvbQ==