 Hanne Aarslev Jensen1*
Hanne Aarslev Jensen1* Laura Garcia Plaza1†
Laura Garcia Plaza1† Rannvá Høgnadóttir Houmann1
Rannvá Høgnadóttir Houmann1 Pernille Bak Andreasen2
Pernille Bak Andreasen2 Niels Bøknæs2
Niels Bøknæs2 Martin Laage Kragh1
Martin Laage Kragh1 Lisbeth Truelstrup Hansen1Ole Mejlholm2‡
Lisbeth Truelstrup Hansen1Ole Mejlholm2‡ Charlotte Jacobsen1
Charlotte Jacobsen1 Paw Dalgaard1
Paw Dalgaard1 Ann-Dorit Moltke Sørensen1*
Ann-Dorit Moltke Sørensen1*- 1National Food Institute, Kgs Lyngby, Denmark
- 2Royal Greenland Seafood A/S, Svenstrup J, Denmark
Immediately after the harvest of lumpfish (Cyclopterus lumpus) roe in artic regions, a common practice is to preserve the fresh roe in brine to produce a salted intermediate product (SIP), which can be transported and stored refrigerated for up to 1 year prior to retail processing. Because the roe is susceptible to microbial growth and lipid oxidation, the brines have been composed of mixes of salt (>10%) and benzoic acid to manage the quality and safety of the SIP. However, a demand for reducing the use of salt and benzoic acid is appearing on the European market. Therefore, the aim of this study was to evaluate the effect of five different brines comprised of combinations of salt, organic acids, and antioxidants on controlling microbial and chemical changes of the SIP, to be able to avoid the use of benzoic acid and reduce the salt content. Freshly harvested roe was mixed with brines to obtain 7 or 10% (w/w) NaCl and combinations of acetic, benzoic and/or lactic acids with or without addition of VivOx 7.5 (IFF Inc.) or Herbalox D-450 (Kalsec Inc.) as antioxidants. Brined roes were stored refrigerated (1.94°C ± 0.56°C) with sampling after 50, 80, 120, 210, 288 and 353 days. Regardless of the treatment applied, aerobic viable counts increased to >6.1 log CFU/g after 120 days of storage and the microbiota in all treatments became dominated by Debaromyces hansenii as identified using MALDI-TOF and metagenome sequencing. The addition of antioxidants reduced the formation of lipid hydroperoxides and thiobarbituric acid reactive substances. Both Herbalox and VivOx reduced the formation of 1-penten-3-ol, whereas VivOx increased the formation of pentanal in the roe. Concentrations of 2-methyl-butanol and 3-methyl-butanol were markedly reduced by treatments that included benzoic acid, which may be related to differences in the activity of the microbiota. Herbalox was more efficient than VivOx in reducing oxidative changes whereas the effect of organic acids on oxidative alterations remained inconclusive. In conclusion, it was possible to reduce the salt content and avoid the use of benzoic acid without affecting the microbial and oxidative stability of the SIP as well as its safety.
1 Introduction
Roe from lumpfish (Cyclopterus lumpus) is considered a delicacy in many countries and Scandinavia has a long tradition for producing different salted lumpfish roe products (Basby et al., 1998a; Johannesson, 2006). The roe makes up approximately 25% of the weight of the female fish at the time of harvest. In the North Atlantic Ocean 2,000–8,000 tonnes of roe is harvested every year with major producers located in Iceland, Greenland, Norway and Canada (Kennedy et al., 2019). Smaller amounts of fresh lumpfish roe are produced in other countries including Denmark and Sweden (Kennedy et al., 2019).
The main season for lumpfish roe fishing in the North Atlantic Ocean is generally limited to a few months during spring (Johannesson, 2006; Kennedy et al., 2019). Distribution of lumpfish roe to consumers throughout the year therefore represents a challenge, particularly, because significant quantities of lumpfish roe are landed in regions where transport to producers in e.g., Europe and the United States requires weeks/months. These logistic challenges are highly important for Greenland, where lumpfish are caught along the more than 2,000 km long coastline of West Greenland, but may also be relevant for other roe producing regions such as Canada, Iceland and Norway. Fresh lumpfish roe has a short shelf-life and at landing sites a simple process that includes mixing of roe with brines (composed of salt and organic acids) has traditionally been used for preservation (Johannesson, 2006). This refrigerated and salted intermediate product (SIP) with 10%–14% NaCl and 0.2%–0.4% sodium benzoate is subsequently transported to a factory, where it is stored for up to 1 year until further processing (Figure 1). For example, production of ready-to-eat lumpfish “caviar” involves desalting the SIP, adding a new brine, red or black natural colours, packing in glass containers and pasteurizing the product for final sale to the retail market (Bledsoe et al., 2003; Kennedy et al., 2019). As a raw material the quality attributes of SIP is expected to influence sensory properties of final roe products. Despite the value of both SIP and final lumpfish roe products, we found no published studies of SIP quality changes during storage.
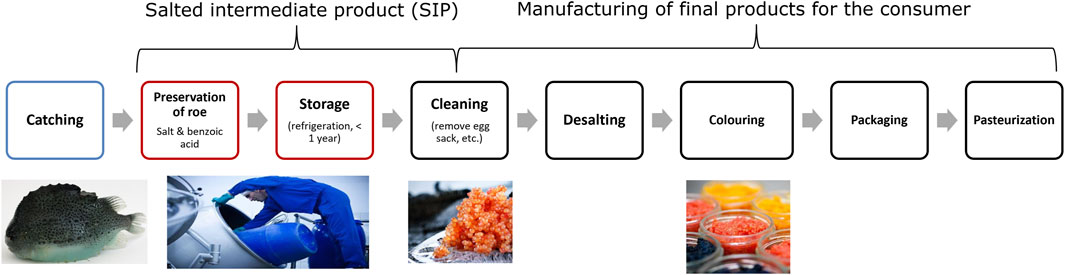
Figure 1. Illustration of the vertical production process of the lumpfish roe from catching in Greenland to the finished pasteurised consumer product.
Both microbial and oxidative changes may be relevant for quality changes in roe preserved by traditional SIP formulations with high salt and benzoic acid although this remains to be documented. Moreover, new SIP formulations with reduced salt and benzoic acid concentrations are desirable in line with requests from costumers. However, how these should be formulated to ensure sufficient preservation with respect to microbial changes is not known. Research on the ability of other organic acids to preserve SIP is therefore required. SIP with reduced salt content compared to the traditional recipy is also of interest to potentially maintain the pink colour of the fresh roe, limit product loss by dehydration and reduce transport of salt to remote regions, e.g., Greenland (Albarracín et al., 2011). Benzoic acid has previously been reported to increase the risk of food allergy in sensitive individuals (Chipley, 2020), hence replacing benzoic acid in SIP will reduce the risk of roe products causing food allergy.
Lumpfish roe contain approximately 4.5% (w/w) lipid and a high percentage of unsaturated fatty acids (Basby et al., 1998b; Vasconi et al., 2020; Pountney et al., 2022). These fatty acids are prone to oxidation, which can cause undesirable changes during storage, e.g., rancid odour and off-flavour. Rosemary extracts for preventing oxidation of other fishery products have previously been shown to be effective for a wide variety of food products (Ozogul et al., 2010) and may reduce oxidative changes in SIP. Additionally, rosemary extracts can have antimicrobial properties in foods (Campo et al., 2000), potentially giving it a dual protective effect on the lumpfish roe. To the best of our knowledge, the effect of rosemary extracts has yet to be investigated for salted lumpfish roe.
The aim of this study was to investigate the effect of brines with reduced salt and different combinations of organic acids and antioxidants based on rosemary extracts, on microbial and oxidative stability of SIP. Firstly, SIP with five different brines were produced in Greenland. Secondly, roe products were transported to the Technical University of Denmark (DTU, Denmark), chemically characterized and stored at +2°C for 1 year with sampling at regular intervals for assessment of microbial and oxidative changes.
2 Materials and methods
2.1 Lumpfish roe, brine ingredients and roe treatments
Lumpfish (Cyclopterus lumpus) were caught off the coast of Paamiut, Greenland (FAO 21, 1E) in April 2022. The lumpfish roe was manually removed from the carcass, and the roe membranes were removed. This was done for easier mixing of the roe with the brines, which is a slightly different procedure than the industrial processing (Figure 1). Fresh roe was stored refrigerated (3.5°C) for up to 4 h until further processing.
Five treatments for production of SIP using different compositions of brines were studied (Table 1). Brines were prepared according to the recipes using different concentrations of salt, benzoic acid (sodium benzoate, BAC, Sigma-Aldrich, STL, United States), acetic acid (sodium acetate, AAC, Macco Organiques, Vallyfield, Québec, Canada) and lactic acid (LAC, Purac Powder 60, Corbion, Netherlands). Additionally, for three of the treatments one of two antioxidants, i.e., Herbalox® D-450 (0.019%, HER, Kalsec Inc., CA, United States) containing rosemary (Rosmarinus officinalis) extract and VivOx 7.5 (0.046%, VIV, IFF, NY, United States) containing a mixture of rosemary (Rosmarinus officinalis) extract and maltodextrin, was also added to the brines.
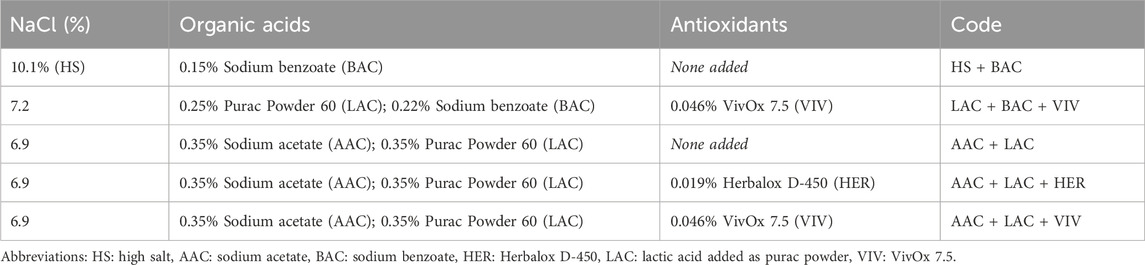
Table 1. Overview of the composition of the five studied roe treatments.
The ingredients used for each treatment (Table 1) were weighed, added to the roe and mixed with tap water. The tap water used comes from the municipal water treatment plant run by the Greenland Utility (Nukissiorfiit). The water is treated using a sand filter and UV-treatment and is regularly checked for its chemical and microbiological quality to ensure compliance with the Greenland drinking water guidelines. For each treatment two food grade polyethylene buckets were each filled with 10 kg of roe and 2 L of brine for final storage. A mesh was added to each bucket to submerge the roe in the brine during storage. The antioxidants were added so that the content of carnosic acid and carnosol was the same for all treatments containing antioxidants (LAC + BAC + VIV, AAC + LAC + HER and AAC + LAC + VIV).
2.2 Storage and sampling
Buckets with brine-salted roe were shipped refrigerated (3.5°C ± 0.9°C) to DTU, (Kgs. Lyngby, Denmark) and stored at 1.94°C ± 0.56°C. Samples were withdrawn from each bucket for further analysis on day 50, 80, 120, 210, 288 and 353 after the initial brine-salting process in Greenland. Microbial enumeration was performed on the day of sampling, whereas samples for chemical analysis were stored at −80°C until further analysis.
2.3 Product characteristics
The content of dry matter (DM) and pH were measured as described by Dalgaard et al. (1993). The salt content was determined using an automated potentiometric titration (785 DMP Tritino titrator, Methrom, Herisau, Austria) method (AOAC method 976.18 (AOAC, 2000), AOAC method 937.07 (AOAC, 1996) and AOAC method 971.27 (AOAC, 1976)). Organic acids were determined by using HPLC-DAD (Agilent 1,200 series, Agilent Technologies, CA, United States) as described by Dalgaard and Jørgensen (2000). External standards were used for identification and quantification of benzoic and lactic acids. An enzymatic acetic acid assay (Megazyme, Wicklow, Ireland) was used to quantify the concentration of acetic acids. All analyses were carried out in biological duplicates with technical replicates (n = 2 × 2).
2.4 Microbiological analysis
2.4.1 Microbial changes
At the time of sampling, 20 g of roe were aseptically withdrawn from each sample bucket and diluted 10-fold in chilled (5°C) peptone buffered saline water (PS, 0.85% NaCl and 0.1% peptone) and homogenized for 60 s in a Stomacher 400 (Seward Medical, London, UK). Further appropriate 10-fold dilutions of the homogenates were made in chilled PS. Aerobic Viable Counts (AVC) were determined by spread plating (15°C, 7 days) on Long and Hammer agar (LH) with 1% NaCl (NMKL, 2006). Lactic acid bacteria (LAB) were enumerated by pour plating (25°C, 3 days) in nitrite actidione polymyxin agar (NAP) with pH 6.2 (Davidson and Cronin, 1973). Yeasts and moulds were enumerated by spread plating (25°C, 3 days) on Oxytetracycline Glucose Yeast Extract (OGYE) agar (CM0545, Oxoid, Basingstoke, United Kingdom) with oxytetracycline commercial supplement (SR0073A, Oxoid). Presumptive Pseudomonas was enumerated by spread plating (25°C, 2 days) on CFC/Pseudomonas agar (CM0559, Oxoid) with Pseudomonas selective supplement (SR0103, Oxoid). All analyses were carried out as single determinations of biological duplicates (n = 2) and results were reported as log CFU/g of roe.
2.4.2 Identification of bacterial isolates using matrix-assisted laser desorption/ionization - time-of-flight (MALDI-TOF)
To identify the dominating microbiota in each SIP treatment after 50, 120 and 353 days of storage, representative colonies on LH agar plates were isolated. For each of the five treatments and at each of the three sampling times, eight colonies were isolated. However, more isolates were selected when colonies seemed more diverse. Colonies were grown in BHI broth (CM1135B, Oxoid) and pure cultured using LH, both incubated at 15°C during 3–7 days. Proteins from colonies on fresh agar plates were extracted with the ethanol/formic acid/acetonitrile protocol used by Nonnemann et al. (2019), as described by Bizzini et al. (2010). Mass spectra were produced with Matrix-Assisted Laser Desorption/Ionization Time of Flight (MALDI-TOF) mass spectrometry using a Maldi-Biotyper® Sirius one RUO Instrument (Bruker Daltonics, Billerica, Massachusetts, USA) with the MBT Compass library. Log scores between 0 and 3 were calculated by the Biotyper algorithm (Bruker Daltonics, Bremen, Germany). Log scores <1.7 do not lead to identification. Log scores 1.7 ≤ x < 2.0 provide genus identification, while log scores ≥2.0 provide species identification. All isolates were analysed in triplicates.
2.4.3 Culture-independent analysis of SIP samples using shotgun metagenomic sequencing
DNA was extracted from duplicate SIP samples from each of the five treatments on day 50 and 353 of the storage experiment using the 10-1 dilution in stomacher bags (see section 2.4.1) that had been stored frozen (−80°C) following the microbiological analysis. In short, aliquots of 10 mL from each sample were centrifuged at 8,960 g at 5°C for 10 min to obtain a pellet containing the microbial cells. DNA was extracted from the pellet with the DNeasy PowerFood kit (Qiagen, Hilden, Germany) following the manufacturer’s protocol. Extracted DNA was quantified using Qubit 3.0 (Invitrogen, Carlsbad, CA, United States) with the Qubit DNA HS Assay Kit (Invitrogen). DNA samples were sent on ice to Eurofins Genomics (Konstanz, Germany) for quality control and Illumina 150 bp paired-end metagenomic shotgun sequencing. One sample (Day 50, AAC + LAC treatment) failed to be sequenced. The raw metagenomic data were submitted to the European Nucleotide Archive (ENA) under project number: PRJEB78984. The raw sequencing data of the 19 samples were processed with CLC Genomics Workbench 24.0. (Qiagen, Aarhus, Denmark) and CLC Genomics Microbial Module 24.0 to trim away adapters and low-quality reads with quality scores below 0.05. After trimming, 22.6–37.4 million reads remained for each sample, with a median of 26.9 million reads/sample. For taxonomic profiling, a database containing all Fungal NCBI RefSeq genomes (RefSeq Fungal, 584 genomes, 30th January 2024, ncbi. nlm.nih.gov/refseq/) was downloaded together with QIAGEN Microbial Insights–Prokaryotic Taxonomy Database Family (QMI-PTDB, version 2.0). The latter is curated and based on reference genomes with representation of all prokaryotic families based on annotations from the Genome Taxonomy Database (GTDB, gtdp.ecogenomic.org). Both RefSeq Fungal and QMI-PTDB family databases were used to create a combined taxonomic profiling reference index, which was applied for taxonomic profiling in CLC Genomics of each sample with a minimum seed length of 30 bp and adjusted for read length variation. Taxonomic classification resulted in 0.4–20.3 million assigned reads for each sample with a median of 6.6 million classified reads and a total of 153.4 million assigned reads. Taxonomic classification results were filtered to remove sequencing noise and extremely low abundance species by removing taxa with less than 1,000 classified reads combined across all 19 samples. The filtered taxonomic abundance tables were merged, exported, and used as input for taxonomic visualizations in GraphPad Prism 10 (GraphPad Software, Boston, MA, United States).
2.5 Chemical analysis
All analyses on roe samples (five treatments, six sampling points) were carried out in biological duplicates with analytical replicates (n = 2 × 2) except for analysis of volatile oxidation products, which were carried out in triplicates (n = 2 × 3). If any exceptions, these are noted in the method description.
2.5.1 Lipid extraction
The lipids were extracted using chloroform and methanol according to the Bligh and Dyer (B&D) method with a reduced amount of solvent (Bligh and Dyer, 1959; Iverson et al., 2001). After oil extraction and the evaporation of chloroform, the lipid content was determined gravimetrically. The results were reported as a percentage of the sample weight.
2.5.2 Determination of free fatty acids (FFA)
The FFAs in the lipid extracts were titrated with NaOH using phenolphthalein as an indicator according to AOCS Method Ca 5a-40 (AOCS, 2004). Aliquots of 10–15 g of the lipid extract were mixed with ethanol (25 mL) and some drops of the indicator were added. Then, the sample was titrated with 0.1M NaOH until a faint pink colour appeared. The volume of NaOH used for titration was used to calculate the FFA content (%). The results were reported as the amount of FFAs (%) by the oleic acid content of the oil.
2.5.3 Determination of content of tocopherols
The tocopherol content was determined according to AOCS Method Ce 8–89 (AOCS, 1998). Approximately 1 g of lipid extract was evaporated to dryness under nitrogen and re-dissolved in heptane (1 mL). The extract was analysed by HPLC–FLD (Agilent 1,100 series, Agilent Technologies) as described by Lu et al. (2015). Results were quantified as µg/g sample.
2.5.4 Content of carnosic acid and carnosol
The main antioxidant compounds in the rosemary extracts were the phenolic compounds carnosic acid and carnosol. Samples were analysed for the content of carnosic acid and carnosol on days 50 and 353. Roe was weighed (0.5 g) into a centrifuge tube, and 2.5 mL solvent (acetone or ethyl acetate) was added. The sample was shaken (20 s) and sonicated for 30 min. The sample was centrifuged (3.500 rpm, ≈2.602 × g, 10 min) and the upper phase was transferred to another centrifuge tube and the solvent was evaporated. The procedure was repeated twice and the pooled evaporated upper phase was redissolved in 1 mL MeOH with iso-ascorbic acid (1 mg/mL) and filtered (0.2 µm). The extraction was performed in duplicates (n = 2). The flow on the HPLC-DAD (Agilent 1,200 series, Agilent Technologies) was 0.9 mL and a gradient method was applied with two solvents, A) Water (pH 3, adjusted with phosphoric acid) and B) Acetonitrile. The gradient method was as follows: 0–20 min 5% Solvent B, 20–25 min 100% Solvent B, 25–27 min 40% Solvent B. A Zorbax ® Eclipse XDB-C8 (5 µm 4.6 × 150 mm, Agilent Technologies) column was used for the separation. The compounds were detected at 280 nm on a UV detector. For the quantification of carnosic acid and carnosol a calibration curve was prepared in MeOH and results were quantified as µg/g sample.
2.5.5 Determination of peroxide value (PV)
PV was determined on the lipid extracts using the colourimetric ferric-thiocyanate method and measured spectrophotometrically at 500 nm, as described by Shantha and Decker (1994), with few modifications as the solvent volume was increased by 2%. The calibration curve was prepared with ferric chloride, and PV was quantified as meq. ROOH/kg oil.
2.5.6 Determination of thiobarbituric acid reactive substances (TBARS)
Samples were analysed for changes in TBARS over the storage period according to Vyncke (1970). Aliquots of 15 g of samples were mixed with 30 mL 7.5% Trichloroacetic acid using an Ultra Thurrax (10,000 rpm, 15 s) and filtrated. Five mL of the filtrated extract were transferred to each of two test tubes. Five mL of 0.02M 2-thiobarbituric acid were added to one tube, whereas 5 mL of water were added to the other tube as a blank. The tubes were then heated (90°C, 40 min) in a water bath and cooled in water. Absorbance of the samples were measured spectroscopically at 530 nm (Shimadzu UV-1280, Holm and Halby, Brøndby, Denmark). An external calibration curve was performed using 1,1,3,3-tetraethoxypropane (TEP, malondialdehyde bis (diethyl acetal)) dissolved in 7.5% trichloroacetic acid (TCA) as the standard to calculate the concentration of TBARS. The results were quantified as µmol/kg sample.
2.5.7 Determination of volatile compounds by dynamic headspace GC–MS
Approximately 10 g of the roe were weighed and mixed with 25 mL water and 30 mg of internal standard solution (30 μg/g of 4-methyl-1-pentanol in rapeseed oil). The samples were purged for 30 min at 37°C with a nitrogen flow of 340 mL/min and volatile compounds were collected on Tenax® GR tubes. After collection, the Tenax® tubes were flushed with a nitrogen flow of 50 mL/min for 20 min. The collected volatiles were desorbed from the Tenax® tubes by an automatic thermal desorber (ATD Turbomatrix 450, PerkinElmer, Norwalk, CT, USA) at 230°C combined with Agilent 6,890 (Palo Alto, CA, USA) GC coupled with a mass spectrometer (HP 5973 Network Mass Selective Detector, Agilent Technologies). The initial GC oven temperature was 55°C for 1.5 min, with increment at 2.0°C/min to 90°C, then increment at 8.0°C/min to 230°C, where it was held for 8 min. The MS conditions were: Electron ionisation mode, 70 eV, mass-to-charge ratio scan between 30 and 250). External standards (2-ethyl-furan, pentanal, 1-penten-3-ol, 3-methyl-butanol, 2-methyl-butanol, 2-pentenal, hexenal, 2-penten-2-ol, 2-hexenal, 4-heptanal, 1-octen-3-ol, octanal, 2,4-heptadienal, nonanal and 2,6-nonadienal (Sigma Aldrich, MO, United States)) were used for calibration curves. The standards were dissolved in ethanol, added to Tenax® tubes in concentrations ranging from 5 to 1,500 ng/g, and analysed by the same method described above. The results were given in ng/g of roe. The LOQ was determined as the lowest concentration used for the standard curve.
2.6 Data analysis
Graphical representations and statistical analysis were performed with GraphPad Prism 10 software. The analysis for increases in microbial concentrations, PV and TBARS were performed by using the built-in tool for simple linear regression including F-ratio and P-value to test if the slope was significantly different from zero. In case of a slope significantly different from zero, the mean of the values was analysed with two-way ANOVA using a post hoc analysis in the form of the Bonferroni t-test to analyse which treatments were significantly (p < 0.05) different over time. Correlations between microbial concentrations, FFA, PV, TBARS and concentrations of volatile compounds data were investigated by a Principal Component Analysis (PCA) using RStudio version 2023.03.1 + 446 (R-Core Team, 2023).
3 Results
3.1 Product characteristics
As expected, similar pH and salt contents were achieved in roe samples from the LAC + BAC + VIV, AAC + LAC, AAC + LAC + HER and AAC + LAC + VIV treatments. Moreover, similar contents of acetic acid and lactic acid were found for roe in the AAC + LAC, AAC + LAC + HER and AAC + LAC + VIV treatments (Table 2).
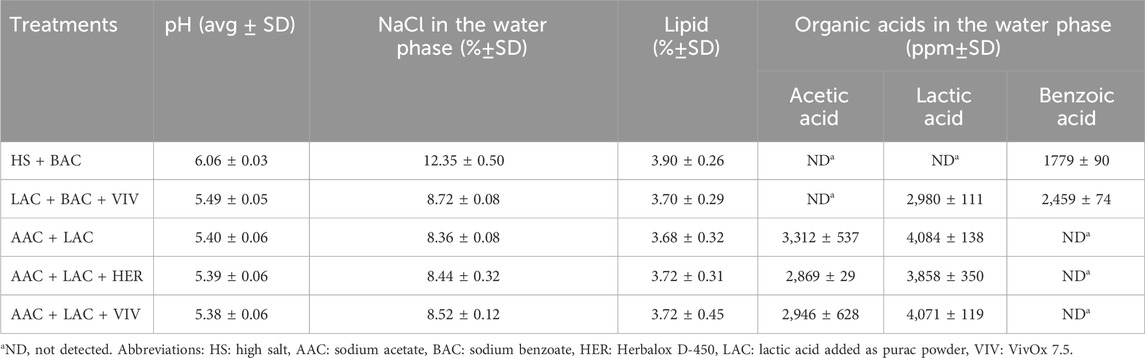
Table 2. Characteristics of lumpfish roe from the five studied treatments.
3.2 Microbial changes and identification of microbiota
AVC increased from 5.5-6 log CFU/g at day 50 to between 6.1–6.5 log CFU/g after 120 days of storage and remained fairly stable through the remaining storage period for all treatments (Figure 2A). Significant (p < 0.05) differences between concentrations of AVC and yeast and moulds were only seen on day 50 for the HS + BAC treatment and on days 288 and 353 for the AAC + LAC + HER treatment with higher concentrations of AVC, indicating that the microbiota was dominated by yeast and moulds (Figures 2A,B). After 210 days of storage, significantly (p < 0.05) higher concentrations of yeast and moulds were seen in HS + BAC compared to roe samples from treatments with acetic and lactic acids (AAC + LAC, AAC + LAC + HER and AAC + LAC + VIV). No significant (p > 0.05) increase was seen in growth on CFC media for any of the five treatments and the average cell concentration was 4.76 ± 0.68 log CFU/g (Figure 2C). Day 50 concentrations of lactic acid bacteria were significantly (p < 0.05) higher in roe from treatments without benzoic acid (4.4–5.4 log CFU/g, AAC + LAC, AAC + LAC + HER and AAC + LAC + VIV), than in roe from treatments with benzoic acid (1-2 log CFU/g, HS + BAC and LAC + BAC + VIV). Between day 50 and 353, significant (p < 0.05) growth of lactic acid bacteria was exclusively seen in roe from treatment HS + BAC with an increase of 1.17 log CFU/g (Figure 2D).
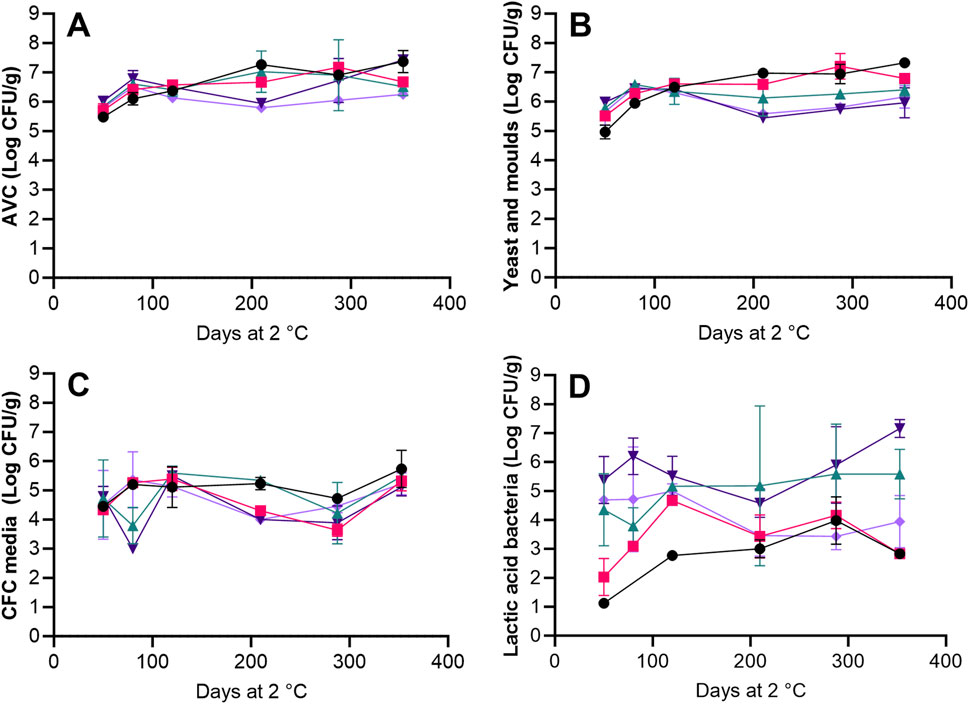
Figure 2. Microbiological concentrations of the roe samples during refrigerated storage. AVC (A) determined on Long and Hammer agar, concentrations of yeast and moulds (B) determined on OYGE agar, concentrations of presumptive Pseudomonas spp. on CFC agar (C) and concentrations for lactic acid bacteria (D) determined on NAP agar, for the HS + BAC (●) LAC + BAC + VIV (




For all treatments, the roe microbiota observed on day 50, as identified by MALDI-TOF, were dominated by yeast (Debaryomyces hansenii and Candida zeylanoides) and by unidentified microorganisms, as exemplified by the HS + BAC treatment, where 75% of the day 50 microbiota was unidentified and the remaining 25% consisted of D. hansenii (Table 3). After 353 days of storage, the microbiota of the treatments HS + BAC and LAC + BAC + VIV were dominated by D. hansenii (>88% of the microbiota), whereas AAC + LAC and AAC + LAC + VIV had a more diverse microbiota including species of Psychrobacter and Latilactobacillus. Roe from the AAC + LAC + HER treatment was dominated by D. hansenii, but also contained C. zeylanoides and Psychrobacter (Table 3).
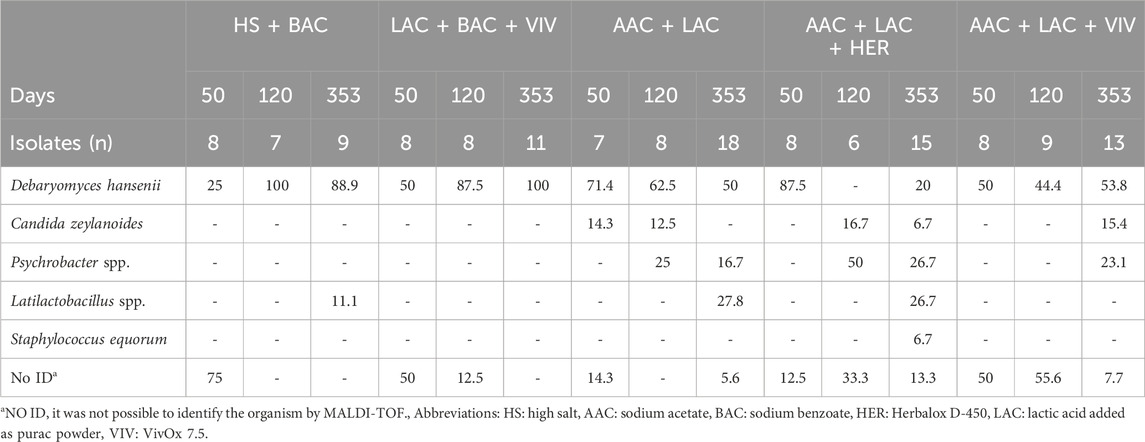
Table 3. Identification of isolates by MALDI-TOF reported as % of the total isolates investigated.
Similar results were obtained by metagenomic sequencing, however with even more pronounced domination of D. hansenii across all treatments during storage (Figure 3). In general, a higher diversity with different Ascomycota spp. was seen on day 50, while D. hansenii made up 96% ± 3% of the metagenomic abundance in samples on day 353 (Supplementary Table S1). Bacteria were generally found in low abundance making up only 1.5% ± 1.7% of the metagenome. Only two bacterial families were found in relative abundances >1% with Catellicoccaceae exclusively found in roe from the HS + BAC treatment after 50 days and with Lactobacillaceae having the highest abundance (2%–5%) in roe samples from the AAC + LAC, and AAC + LAC + HER treatments after 353 days (Figure 3). While not mirroring the abundances detected by the MALDI-TOF identification, Psychrobacter spp. Sequences were only found in roe samples preserved with AAC containing treatments after 353 days (data not shown). Similarly, C. zeylanoides, belonging to the class Saccharomycetes, were found in all metagenomic samples of day 50 and 353, though at abundancies below 0.01% (data not shown).
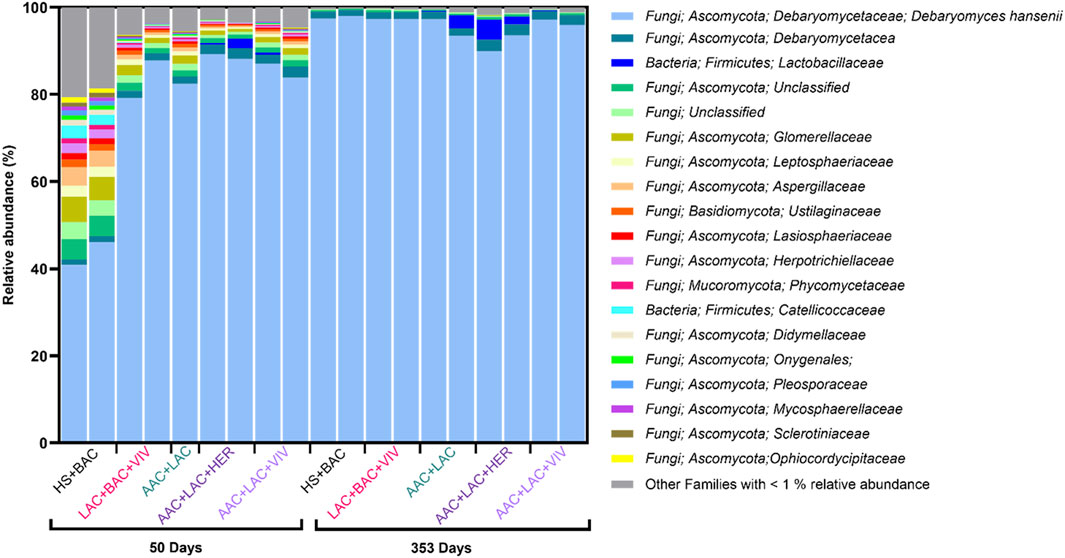
Figure 3. Microbiome composition of brined treated lumpfish roe as determined by metagenomic sequencing of roe samples (n = 2 for each of the five treatments) after 50 and 353 days of storage. One roe sample (day 50, AAC + LAC) failed to be sequenced. Exclusively, taxa present at a relative abundance of at least 1% in one sample are shown in the legend.
3.3 Chemical changes
The content of free fatty acids (FFAs) increased by 8%–10% during storage of roe from all treatments, though a lower content of FFA was seen in the roe from the HS + BAC treatment throughout the storage period (Figure 4).
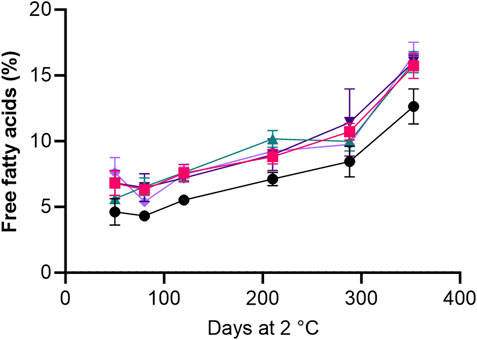
Figure 4. Effect of refrigerated storage on the content of % free fatty acids (FFA) in roe samples of the HS + BAC (●) LAC + BAC + VIV (




Tocopherols possess antioxidative properties and acts either as scavengers of radicals or by reacting with singlet oxygen, hence a reduction in tocopherols is often seen in oxidized products (Pazos et al., 2005). Of the four analysed tocopherol homologues, α-tocopherol was the only one detected and higher concentrations were found in roe from treatments with added antioxidants (∼12.7 μg/g) compared to treatments without antioxidants (∼7.2 μg/g). No significant (p > 0.05) consumption of α-tocopherol was observed during storage for any of the five treatments (data not shown).
Similar contents of carnosic acid and carnosol were found in roe samples treated with antioxidants. The content of carnosic acid decreased, though not significantly (p > 0.05) for LAC + BAC + VIV and AAC + LAC + HER during storage (0.65 μg/g reduction). A significant (p < 0.05) decrease of 1.29 μg/g was seen for AAC + LAC + VIV (Figure 5A). During storage, the initial content of carnosol in LAC + BAC + VIV was reduced to undetectable levels, whereas the initial contents in AAC + LAC + HER and AAC + LAC + VIV were significantly (p < 0.05) reduced during storage by 2.83 and 3.97 μg/g, respectively (Figure 5B). As expected, neither carnosic acid nor carnosol was detected in roe from the HS + BAC or AAC + LAC treatments (data not shown).
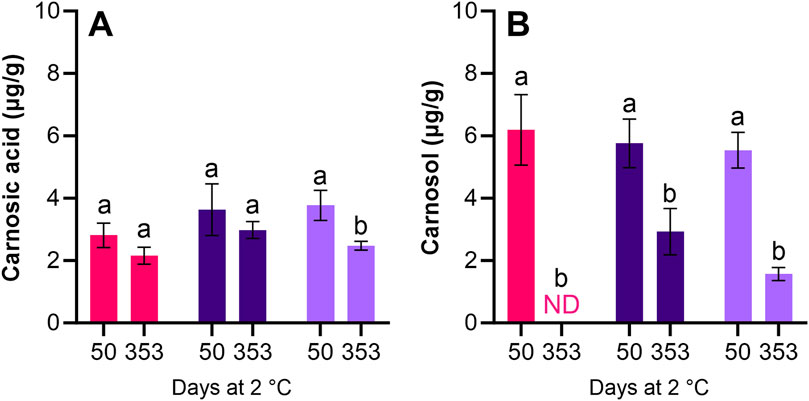
Figure 5. Content of carnosic acid (A) and carnosol (B) in the roe samples of the LAC + BAC + VIV (



Primary oxidation products can be measured by determinations of PV, which measures the amount of lipid hydroperoxides in the sample. Treatments with antioxidants (LAC + BAC + VIV, AAC + LAC + HER and AAC + LAC + VIV) showed lower initial PV (<2.04 meq. ROOH/kg oil) in roe samples compared to treatments without antioxidants added (>4 meq. ROOH/kg oil, HS + BAC and AAC + LAC; Figure 6A). Significant (p < 0.05) increases were seen during 288 days of storage of roe from all treatments except for AAC + LAC, which is due to the higher PV levels at days 80 and 120. Importantly, the lowest increase over time was observed in roe from the AAC + LAC + HER treatment, which only increased from 0.99 meq. ROOH/kg oil to 2.79 meq. ROOH/kg oil after 210 days of storage (Figure 6A). The PVs of the remaining four treatments increased to 7–8 meq ROOH/kg oil after 210 days of storage. This indicates that the addition of Herbalox was more effective in suppressing hydroperoxide formation compared to VivOx 7.5. Significant (p < 0.05) decreases in PV were observed after 288 days of storage for samples containing acetic acid and lactic acid (AAC + LAC, AAC + LAC + VIV and AAC + LAC + HER), with AAC + LAC showing the largest decrease of 6.3 meq. ROOH/kg oil.

Figure 6. PV (A) and TBARS (B) in roe samples of the HS + BAC (●) LAC + BAC + VIV (




Secondary oxidation products can be measured by TBARS. A significant (p < 0.05) increase was observed in TBARS over time in roe from all treatments. TBARS were significantly (p < 0.05) higher for AAC + LAC throughout storage compared to the other treatments, though TBARS in roe samples of HS + BAC also showed a tendency to be higher than TBARS of samples of roe from treatments with antioxidants. Significantly (p < 0.05) lower TBARS was seen after 220 days of storage for roe samples with added antioxidants (LAC + BAC + VIV, AAC + LAC + HER and AAC + LAC + VIV) compared to roe samples without added antioxidants (HS + BAC and AAC + LAC; Figure 6B).
Secondary oxidation products can also be measured by analysis of volatile oxidation products. The quantification of volatile compounds by GC-MS gives a more nuanced understanding of the secondary oxidation occurring during storage compared to TBARS, as the formation of specific volatile compounds can often be related to the oxidation of their precursors, mainly fatty acids. A total of 15 volatile compounds were quantified, but only 2-ethyl-furan, 2-penten-2-ol, 1-penten-3-ol, 3-methyl-butanol, 2-methyl-butanol, pentanal and nonanal had concentrations above the LOQ (approx. 10 ng/g) of the method or increased to above LOQ within the storage period.
The content of pentanal (Figure 7A) in roe increased to above the LOQ for LAC + BAC + VIV and AAC + LAC + VIV treatments after 288 days of storage, whereas contents in roe from HS + BAC, AAC + LAC and AAC + LAC + HER treatment remained below the LOQ throughout storage. The content of 1-penten-3-ol (Figure 7B) increased to more than 400 ng/g in roe from the HS + BAC treatment, and to more than 200 ng/g in roe from AAC + LAC, whereas the content in roe samples with antioxidants (LAC + BAC + VIV, AAC + LAC + HER and AAC + LAC + VIV) remained around or below 100 ng/g throughout the storage period, with the highest final content seen in LAC + BAC + VIV (123.4 ± 8.4 ng/g) and the lowest in AAC + LAC + HER (57 ± 6.7 ng/g). The contents of 2-methyl-butanol (Figure 7C) and 3-methyl-butanol (Figure 7D) increased during storage for roe samples prepared in brines containing both acetic and lactic acids (AAC + LAC, AAC + LAC + HER and AAC + LAC + VIV), whereas roe samples preserved with benzoic acid (HS + BAC and LAC + BAC + VIV) showed significantly (p < 0.05) lower increases during storage. Of the remaining volatile compounds with concentrations above the LOQ, the content of 2-ethyl-furan increased to ∼95 ng/g in roe from AAC + LAC, and to 38 ng/g in HS + BAC, whereas roe samples with antioxidants remained below 25 ng/g throughout storage, yet again showing the effect of the added antioxidants and of the higher salt content. 2-penten-2-ol showed the highest increase during storage of roe in HS + BAC with a final content of ∼18 ng/g, AAC + LAC increased to ∼11 ng/g, whereas roe samples prepared with antioxidants remained around 5–7 ng/g. The content of nonanal showed no consistent trend, fluctuating with both increases and decreases in roe samples throughout the storage period. The decrease in PV in roe samples after 288 days of storage, as seen in Figure 6A, could not be explained by changes in secondary oxidation products as determined by changes in TBARS (Figure 6B) or volatile compounds (Figures 7A–D).
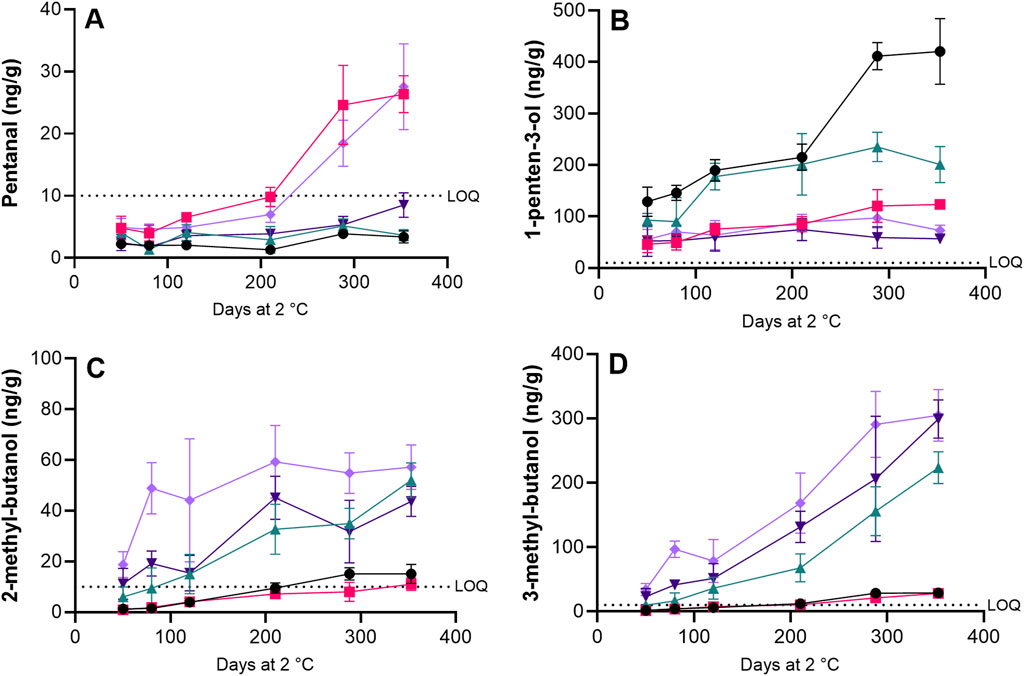
Figure 7. Development of selected secondary volatile oxidation products during refrigerated storage of roe samples: pentanal (A), 1-penten-3-ol (B), 2-methyl-butanol (C) and 3-methyl-butanol (D) for the HS + BAC (●) LAC + BAC + VIV (




The PCA plot, as shown in Figure 8, clearly demonstrates how different treatments affect the roe samples and highlights which of the analysed chemical and/or microbiological quality attributes are important for the variation in each treatment. This analysis allows for the evaluation of correlations (positive or negative) between treatments and the measured parameters in the roe.
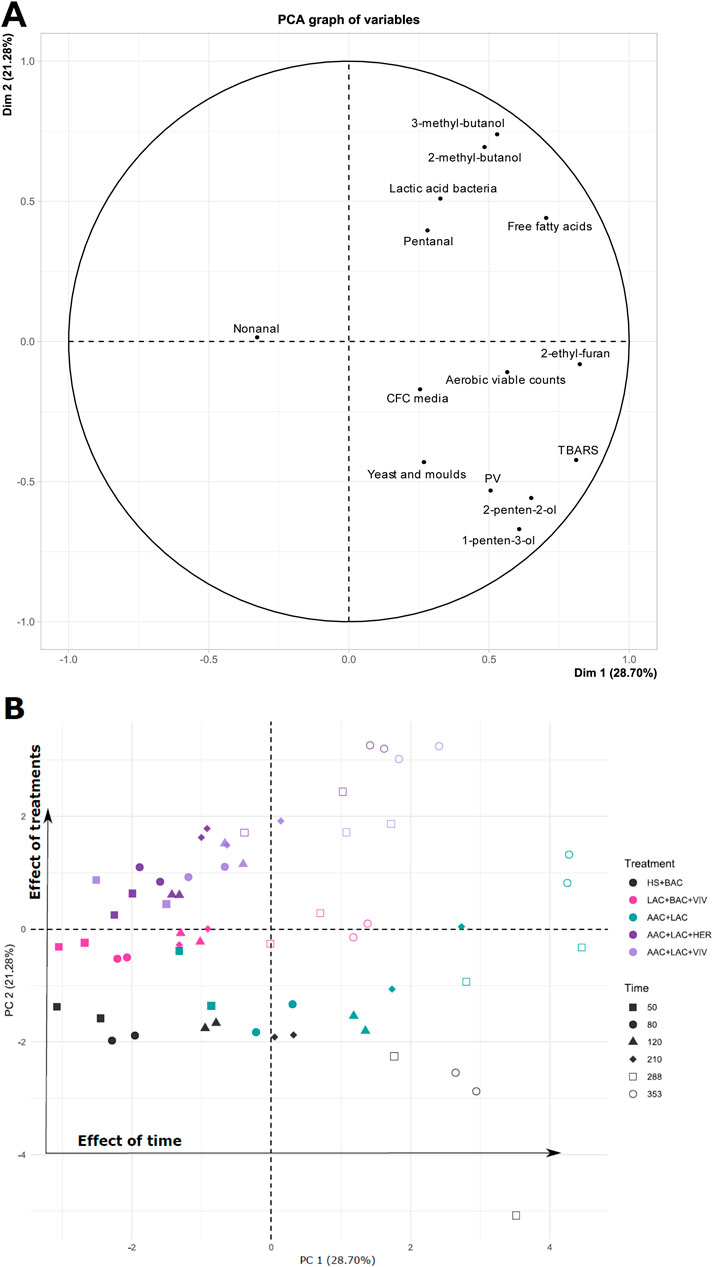
Figure 8. Principal Component Analysis (PCA) based on roe samples’ microbial concentrations, FFA, PV, TBARS and volatile compounds: Loadings (A) and Scores (B). Roe sample treatments: HS + BAC (black) LAC + BAC + VIV (
Principal component 1 (PC1) and PC2 explain 30.64% and 21.24% of the variation in the data, respectively. The plots show a clear time-related tendency along PC1, whereas PC2 appears to explain variance between treatments (Figure 8B). Based on the locations of scores along the PC1-axis, roe samples preserved with antioxidants appear to vary less with time compared to samples without antioxidants. Roe samples from days 288 and 353 of all treatments, except HS + BAC, are in the 1st quadrant (Figure 8B) and correlates with both free fatty acids, lactic acid bacteria and the three volatile compounds pentanal, 2-methyl-butanol and 3-methyl-butanol, i.e., higher content (Figure 8A). This corresponds well with the observed higher contents of FFA (Figure 4), 2-methyl-butanol, 3-methyl-butanol and pentanal in roe samples from the AAC + LAC + VIV and LAC + BAC + VIV treatments (Figure 7). The position of AAC + LAC + HER treated roe can be explained by changes in concentrations of FFA, 2-methyl-butanol and 3-methyl-butanol. Samples of HS + BAC from day 288 and 353 appear to correlate with PV, yeast and moulds and the two volatile alcohols 2-penten-2-ol and 1-penten-3-ol. The higher TBARS, PV and 1-penten-3-ol explain the location of roe samples of the HS + BAC and AAC + LAC treatments. Additionally, the general increases in the values of most parameters during the storage period explain the variance along PC1.
4 Discussion
4.1 Microbial changes
AVC showed levels of 6.1–7.4 log CFU/g during the final storage period (>120 days) in roe from all treatments. The high AVC levels are similar to values reported in literature for storage of other roe products stabilized with markedly lower salt concentrations of 3%–5% salt with and without organic acids (Basby et al., 1998c; Panjaitan et al., 2020; Lopez et al., 2021). The lack of effect on AVC of the addition of organic acids to the roe was unexpected, since previous research has shown its ability to reduce microbial growth in roe products (Lopez et al., 2021), in brined shrimp (Mejlholm et al., 2008) and for white sturgeon caviar during long term storage at −2°C (Lopez et al., 2021). In contrast to studies of other fishery products (Campo et al., 2000; Feng et al., 2022), no apparent antimicrobial effect of rosemary extract was observed in our study, potentially due to composition of the microbiota, low product pH or high salt content of the roe.
Over the course of the storage period, the microbiota of all samples became dominated by the yeast D. hansenii (Figures 2, 3; Table 3). This was unexpected for roe samples treated with a brine containing benzoic acid, as this organic acid typically inhibits yeast (Chipley, 2020). However, the effect of organic acids depends on the pH and product type. For instance, the minimum inhibitory concentration (MIC) of benzoic acid against D. hansenii varies with pH: 100–200 ppm at pH 3.5, 250–500 ppm at pH 5, and 1,500 ppm at pH 7 (Fleet, 2011; Chipley, 2020). Despite >1,500 ppm benzoic acid (Table 2) growth of D. hansenii was not prevented during storage of the studied SIP treatments. Concentrations of AVC and yeast in freshly produced SIP from Paamiut were, respectively, 3.2 ± 0.5 log CFU/g and 2.1 ± 0.2 log CFU/g during the 2022 season (n = 36; results not shown). The yeast, present after processing, grew in SIP as observed in the present study. The 50-day microbiota for HS + BAC was less dominated by yeast than other treatments, which could result from >12% water phase salt, to some extent, reducing growth of D. hansenii. Bagge-Ravn et al. (2003) found yeast to account for 21% of the microbiota recovered from equipment used when SIP is further processed into retail lumpfish roe products. This may suggest yeast from SIP have contaminated the processing equipment. Higher day-50 concentrations of lactic acid bacteria were seen in roe samples stabilized in brines without benzoic acid (Figure 2C), indicating an initial inhibiting effect of the benzoic acid on the lactic acid bacteria.
During storage, the microbiota became less diverse, as the microorganisms, which were best adapted to the product environment, came to dominate the roe samples. This is seen by fewer taxa being present in the roe at the end of storage (Figure 3). The exclusion of benzoic acid in the recipes of AAC + LAC, AAC + LAC + HER and AAC + LAC + VIV brines also seemed to impact the diversity of the microbiota as several bacterial species were detected and identified by MALDI-TOF in the roe samples without benzoic acid. However, in the metagenomic analysis, Lactobacillales were the only bacterial family with a relative abundance >1% in the roe. Lactic acid bacteria have previously been shown to spoil lightly salted lumpfish roe (Basby et al., 1998a; Basby et al., 1998c) and have also been isolated from seafood processing equipment (Bagge-Ravn et al., 2003). Specifically, Latilactobacillus sakei has previously been isolated from seafood in brine from arctic waters (Mejlholm et al., 2008), which may explain its presence in the roe samples. Lactic acid bacteria have previously been shown to be able to spoil seafood products by producing sulphurous and acidic odours (Leroi, 2010). D. hansenii was the dominant yeast detected in all samples by both culture-dependent (MALDI-TOF) and culture-independent (metagenomics) determinations and was especially dominant after 353 days of storage. It was initially isolated from seawater (Norkrans, 1966), which could lead to the lumpfish being naturally contaminated prior to harvest and hence its presence in the roe samples. The yeast has repeatedly been isolated from foods stored in salt brines, such as cheese and cured meat products (Haastrup et al., 2018; Hernández et al., 2018). The yeast is known to be halotolerant, able to grow at low pH and in the presence of organic acids (Breuer and Harms, 2006), probably causing it to become dominant during storage. Literature both reports the yeast acting as a spoiler and as a flavour-enhancer (Breuer and Harms, 2006; Ashaolu et al., 2023), likely depending on the food matrix in which it grows and on the microbial concentrations. Studies to elucidate its role in spoilage of lumpfish roe should be performed in the future. The identification of a single colony of Staphylococcus equorum in roe of the AAC + LAC + HER treatment on day 353 of storage is interesting as this bacterium is generally not found to grow at temperatures below 6°C (Vos et al., 2009). The metagenomic analysis also detected DNA from S. equorum, but at an abundance below 0.001%, showing its presence albeit in very low amounts. Staphylococcus spp. have previously been found to be present on processing equipment for lumpfish roe (Bagge-Ravn et al., 2003) and have been isolated from marine salts (Cordero and Zumalacárregui, 2000) suggesting that the genus can be introduced through the addition of salt to the products, or by the surrounding marine or processing environment.
The rather large number of unidentified isolates shows the limitations of using MALDI-TOF for investigation of the microbial community of SIP. However, this can most likely be attributed to the size and type of library applied for identification of the isolates. The library used for this study consists of approximately 4,000 species and the library generally targets clinical and terrestrial isolates (Han et al., 2021). A limitation of culture-independent metagenomic based analysis is the indiscriminate sequencing of DNA from all viable, non-culturable and dead microorganisms. To counter this limitation, recent studies using propidium monoazide have, with limited efficiency, tried to reduce the bias even in complex communities (Yap et al., 2022; Liu et al., 2023; Yang et al., 2023). Despite the limitations of both culture-dependent and independent techniques, this study shows that both approaches yielded the same result. An advantage of the culture-based method is that it provided us with isolates of D. hansenii, which can be used in future studies.
When storing foodstuff refrigerated for extended periods of time, the microbiological food safety must also be considered and ensured. For this purpose, predictive modelling is useful (Ross et al., 2000). Predictions from the Food Safety and Spoilage Predictor (FSSP, www.fssp.dtu.dk) forecasts that the combinations of salt, pH and organic acids in the experimental brine treatment will inhibit the growth of Listeria monocytogenes, which is an important psychrotolerant foodborne bacteria pathogen associated with lightly preserved fish products, during the storage of the salted intermediate roe products (Mejlholm and Dalgaard, 2009). Additionally, product characteristics and storage condition will efficiently prevent growth and potential toxin formation of non-proteolytic Clostridium botulinum (Koukou et al., 2021).
In summary, the microbial results show that reducing the water phase salt content from >12 to ∼8% and avoiding the use of benzoic acid in brines did not notably affect the microbial growth in the intermediate roe products. Nor was there an evident effect of adding the antioxidants on the microbial changes. This indicates that it is indeed possible to change the brine recipes to lower the salt and omit benzoic acid without affecting the microbiological quality of the intermediate product. D. hansenii dominated microbiota of the studied SIP product and yeast should be taken into account when its microbiological quality is evaluated and managed. Specifically, to limit growth of yeast it is suggested to reduce product pH as a change from 5.5 to 5.2 will have the same stabilizing effect as doubling concentrations of organic acids.
4.2 Chemical changes
FFA in seafood are produced by lipolysis of triglycerides, usually by lipolytic enzymes originating from products or from the microbiota, but heat-induced lipolysis can also occur. FFA are more prone to oxidation during storage than fatty acids bound to triacylglycerides or phospholipids. The FFA content increased similarly in all roe samples during storage, regardless of the addition of antioxidants to brines indicating that the microbial or endogenous enzymes were likely responsible for the formation of FFA. This finding aligns with a study by Ozogul et al. (2010), which found that adding rosemary extract to vacuum-packed sardines stored at 4°C had no considerable effect on reducing the FFA formation compared to a control without rosemary extract. A lower FFA content at day 50 was determined in roe from the HS + BAC treatment, which could be related to the higher salt concentration compared to the other treatments (Figure 4). This is likely due to an inhibitory effect of salt on lipases (Albarracín et al., 2011). An increase in FFA has previously been linked to the rancid odour characteristics of seafood (Refsgaard et al., 1998; Refsgaard et al., 2000), but more studies should be performed to determine their sensory impact in salted lumpfish roe.
The addition of antioxidants inhibited the formation of hydroperoxides compared to samples without antioxidants. Moreover, Herbalox D-450 seemed to be more effective than VivOx 7.5. The reason for this difference between the two rosemary-based antioxidants is unclear, as the only difference between them is the addition of maltodextrin to VivOx 7.5. Further studies are needed to explore this difference. At the final sampling point after 353 days, PV decreased for all samples (Figure 6A), suggesting that hydroperoxides decomposed into secondary oxidation products at a faster rate than they were formed. This decrease in PV during storage has also been observed in other fishery products (Larsson et al., 2007; Elavarasan and Shamasundar, 2022; Jensen et al., 2024).
Lower increases in TBARS values during storage were observed in roe samples preserved with antioxidants regardless of differences in organic acid composition (Figure 6B), showing the beneficial effect of adding the antioxidants to maintain the oxidative stability of the roe. A similar positive effect has been shown for the addition of rosemary extract to other fish products, such as chilled, vacuum-packed sardines stored at 4°C (Ozogul et al., 2010) and chilled sea bass (Feng et al., 2022). The results indicate that the high salt content in HS + BAC (∼12% NaCl) compared to AAC + LAC (∼8% NaCl) could reduce the formation of TBARS. A similar effect of high salt contents in reducing TBARS was also reported by İnanli et al. (2010) and Alak et al. (2021) for salted rainbow trout (Oncorhynchus mykiss) roe stored at 4°C. The latter study suggested that the reduced formation of TBARS, was due to dehydration of the caviar caused by salt addition, which changed the relative concentrations of lipids and proteins in the samples. TBARS have previously been reported to correlate with sensory changes of fish products (Larsson et al., 2007). However, due to the complexity of lipid oxidation reactions, finding a universal level of TBARS to determine shelf-life and/or sensory changes is not simple.
The aldehyde pentanal is linked to the oxidation of linoleic acid (Gorji et al., 2019). The formation of pentanal appears to be related to the presence of the antioxidant VivOx 7.5, as increases to above LOQ were only seen in roe samples preserved with this antioxidant i.e., LAC + BAC + VIV and AAC + LAC + VIV 5 (Figure 7A). This could be related to the content of maltodextrin in VivOx 7.5, which is not present in Herbalox D-450. More studies should be performed to investigate the potential for maltodextrin to affect the formation of pentanal. 1-penten-3-ol can be formed from the oxidation of n-3 polyunsaturated fatty acids (e.g., docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA)) (Lee et al., 2003). The addition of antioxidants caused the formation of this volatile to be markedly reduced compared to roe samples without antioxidants (Figure 7B). Furthermore, a 2-fold increase was observed for HS + BAC roe samples between 210 and 288 days of storage and the concentration was 2-fold higher than in roe from the AAC + LAC after 353 days of storage. We have no clear explanation for the sudden and notable increase in this volatile compound in the HS + BAC samples compared to the AAC + LAC samples. 2-methyl-butanol and 3-methyl-butanol are likely derived from the amino acids isoleucine and leucine, respectively. An intermediate step in this metabolic pathway could involve the enzymatic degradation of 2-methyl-butanal and 3-methyl-butanal, by alcohol dehydrogenases (ADH) originating from the microorganisms present in the product (Zhuang et al., 2021). From the results, it appears that the presence of benzoic acid limited the formation of these two volatile compounds in roe (Figures 7C,D), indicating an effect of the organic acid on the microorganisms that produce these metabolites during storage. Looking at the microbial identification, it seems likely that members of Lactobacillales could be contributing to the formation of these two volatile compounds, as these were found in higher abundance in roe samples without benzoic acid throughout storage. Also, D. hansenii and Psychrobacter spp. have been reported to produce 2-methyl-butanol and/or 3-methyl-butanol in foods and model systems (Freiding et al., 2011), supporting the likelihood that the microorganisms of the roe contributed to the formation of these two volatile compounds during storage. Similar to results of this study, the contents of 1-penten-3-ol, and 3-methyl-butanol were also shown to increase in salted white sturgeon caviar during storage at −2°C (Lopez et al., 2021). Other authors have described 3-methyl-butanol to be an important contributor to the odour of over-ripened cod roe, and this compound was suggested to be a potential indicator of spoilage (Jonsdottir et al., 2004), but further investigations must be performed to determine the importance of volatiles for the sensory deterioration of lumpfish roe.
All in all, the results of PV, TBARS and volatile compounds show the value of adding antioxidants for reducing the oxidative changes in the intermediate roe product during chilled storage. Especially the addition of Herbalox D-450 was found to be favourable in reducing both the formation of primary and secondary oxidation products compared to the use of VivOx 7.5 (Figures 6, 7). As similar concentrations of the active compounds; carnosic acid and carnosol, were added to the brines (Figure 4), the difference might be related to the addition of maltodextrin in VivOx 7.5, but maltodextrin has previously been shown to improve oxidative stability rather than promoting oxidation. Thus, causes for the differences in effect of the two antioxidants should be further investigated.
Differences in the colour of roe from the treatments were noted during the storage period; however, the changes were not quantified. Further work should be undertaken to characterize the sensory characteristics of the roe during storage, as well as to determine if the applied antioxidants affect the sensory properties of the lumpfish roe.
5 Conclusion
The present study is to the best of our knowledge the first to evaluate microbial and oxidative changes for the brined intermediate lumpfish roe product. The results of this study show how beneficial it is to investigate both microbial and chemical changes of products to be able to understand the changes, potential causes of these and the effect of different treatments on a product’s storage stability. For the microbial stability, no evident effect of the different combination of organic acids in the brines on microbial concentrations was observed, as all treatments reached AVC levels of 6.1–7.4 Log CFU/g after 120 days of storage. The formation of the two volatile compounds 2-methyl-butanol and 3-methyl-butanol was likely related to the microbiota of the intermediate roe including D. hansenii and lactic acid bacteria, which occurred in higher concentrations in treatments without benzoic acid. Additionally, no clear antimicrobial effect of the rosemary extracts was seen. The yeast D. hansenii, identified by MALDI-TOF and by metagenome sequencing, grew to high concentrations both in the high salt treatment (10% NaCl, pH 6.1 and with benzoic acid) and in the studied alternative treatments with reduced salt (8%), pH ∼5.4 and where benzoic acid was replaced with a combination of acetic and lactic acids. To be able to reduce salt and benzoic acid in the salted intermediate roe product and still be able to control the microbial growth, this study suggests that pH must be lowered further than the studied level of ∼5.4 as this will increase the inhibitory effect of the added organic acids. Looking at the oxidative stability, the addition of antioxidants reduced the oxidative reactions in terms of the formation of hydroperoxides, TBARS and volatile compounds compared to samples without added antioxidants. No clear effect of the different organic acids was observed on the oxidative changes. Herbalox D-450 was better than VivOx 7.5 at inhibiting the formation of hydroperoxides, TBARS and the two volatile compounds pentanal and 1-penten-3-ol. In conclusion, it is evident from the results that it is possible to reduce the salt content and use alternatives to the potential food allergen, benzoic acid, without compromising the product quality.
Data availability statement
The data presented in this study are available on request from the corresponding authors. The raw metagenomic data used can be found in online repository. The name of the repository and accession number can be found below: https://www.ebi.ac.uk/ena, PRJEB78984.
Author contributions
HJ: Formal Analysis, Visualization, Writing–original draft, Writing–review and editing. LP: Conceptualization, Investigation, Methodology, Writing–review and editing. RH: Investigation, Writing–review and editing. PA: Investigation, Writing–review and editing. NB: Conceptualization, Funding acquisition, Investigation, Methodology, Resources, Supervision, Writing–review and editing. MK: Formal Analysis, Visualization, Writing–review and editing. LH: Conceptualization, Funding acquisition, Methodology, Supervision, Writing–review and editing. OM: Conceptualization, Funding acquisition, Methodology, Resources, Supervision, Writing–review and editing. CJ: Conceptualization, Funding acquisition, Methodology, Supervision, Writing–review and editing. PD: Conceptualization, Funding acquisition, Methodology, Supervision, Writing–review and editing. A-DS: Conceptualization, Funding acquisition, Methodology, Project administration, Supervision, Writing–review and editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. The article is part of the project PROFIUS (Preservation of underutilized fish biomasses for improved quality, stability, and utilization). This project has received funding from the European Union’s Horizon 2020 research and innovation programme under grant agreement 817992 and Innovation Fund Denmark.
Acknowledgments
We would like to thank the fishermen in Paamiut, Greenland for help with catching the Lumpfish and the workers at the factory in Paamiut, Greenland for technical assistance. Kalsec Inc. (Kalamazoo, MI, USA). is thanked for supplying the Herbalox D-450. Laboratory technician Inge Holmberg is thanked for assistance with analytical work.
Conflict of interest
The authors NB, PA, and OM were employed by Royal Greenland during the experiment.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/frfst.2024.1498035/full#supplementary-material
References
Alak, G., Kaynar, Ö., and Atamanalp, M. (2021). The impact of salt concentrations on the physicochemical and microbiological changes of rainbow trout caviar. Food Biosci. 41, 100976. doi:10.1016/j.fbio.2021.100976
Albarracín, W., Sánchez, I. C., Grau, R., and Barat, J. M. (2011). Salt in food processing; usage and reduction: a review. Int J Food Sci Tech 46, 1329–1336. doi:10.1111/j.1365-2621.2010.02492.x
AOAC (1976). Sodium chloride in canned vegetables. Potentiometric method. Arlington, VA, USA: AOAC. Official method 971.27.
AOAC (1996). Fish and marine products. Treatment and preparation of sample procedure. Arlington, VA, USA: AOAC. Official method 937.07.
AOAC (2000). Salt (chlorine as sodium chloride) in seafood. Potentiometric method. Arlington, VA, USA: AOAC. Official method 976.18.
AOCS (1998). Tocopherols and tocotrienols in vegetable oils and fats by HPLC. IL, USA: AOCS: Campaign. Official method Ce 8-89.
AOCS (2004). Free fatty acids in crude and refined fats and oils. IL, USA: AOCS: Campaign. Official method Ca 5a-40.
Ashaolu, T. J., Khalifa, I., Mesak, M. A., Lorenzo, J. M., and Farag, M. A. (2023). A comprehensive review of the role of microorganisms on texture change, flavor and biogenic amines formation in fermented meat with their action mechanisms and safety. Crit. Rev. Food Sci. Nutr. 63, 3538–3555. doi:10.1080/10408398.2021.1929059
Bagge-Ravn, D., Ng, Y., Hjelm, M., Christiansen, J. N., Johansen, C., and Gram, L. (2003). The microbial ecology of processing equipment in different fish industries - analysis of the microflora during processing and following cleaning and disinfection. Int. J. Food Microbiol. 87, 239–250. doi:10.1016/S0168-1605(03)00067-9
Basby, M., Jeppesen, V. F., and Huss, H. H. (1998a). Spoilage of lightly salted Lumpfish (Cyclopterus lumpus) roe at 5 °C. J. Aquat. Food Prod. 7, 23–34. doi:10.1300/J030v07n04_04
Basby, M., Jeppesen, V. F., and Huss, H. H. (1998b). Chemical composition of fresh and salted Lumpfish (Cyclopterus lumpus) roe. J. Aquat. Food Prod. 7, 7–21. doi:10.1300/J030v07n04_03
Basby, M., Jeppesen, V. F., and Huss, H. H. (1998c). Characterization of the microflora of lightly salted Lumpfish (Cyclopterus lumpus) roe stored at 5 °C. J. Aquat. Food Prod. 7, 35–51. doi:10.1300/J030v07n04_05
Bizzini, A., Durussel, C., Bille, J., Greub, G., and Prod’hom, G. (2010). Performance of matrix-assisted laser desorption ionization-time of flight mass spectrometry for identification of bacterial strains routinely isolated in a clinical microbiology laboratory. J. Clin. Microbiol. 48, 1549–1554. doi:10.1128/JCM.01794-09
Bledsoe, G. E., Bledsoe, C. D., and Rasco, B. (2003). Caviars and fish roe products. Crit. Rev. Food Sci. Nutr. 43, 317–356. doi:10.1080/10408690390826545
Bligh, E. G., and Dyer, W. J. (1959). A rapid method of total lipid extraction and purification. J. Biochem. Physiol. 37, 911–917. doi:10.1139/o59-099
Breuer, U., and Harms, H. (2006). Debaryomyces hansenii — an extremophilic yeast with biotechnological potential. Yeast 23, 415–437. doi:10.1002/yea.1374
Campo, J. D., Amiot, M.-J., and Nguyen-The, C. (2000). Antimicrobial effect of rosemary extracts. J. Food Prot. 63, 1359–1368. doi:10.4315/0362-028X-63.10.1359
Chipley, J. R. (2020). “Sodium benzoate and benzoic acid,” in Antimicrobials in food. Editors P. M. Davidson, T. M. Taylor, and J. R. D. David Fourth edition (Boca Raton: CRC Press), 41–88. doi:10.1201/9780429058196-3
Cordero, M. R., and Zumalacárregui, J. M. (2000). Characterization of Micrococcaceae isolated from salt used for Spanish dry-cured ham. Lett. Appl. Microbiol. 31, 303–306. doi:10.1046/j.1472-765x.2000.00818.x
Dalgaard, P., Gram, L., and Huss, H. H. (1993). Spoilage and shelf-life of cod fillets packed in vacuum or modified atmospheres. Int. J. Food Microbiol. 19, 283–294. doi:10.1016/0168-1605(93)90020-H
Dalgaard, P., and Jørgensen, L. V. (2000). Cooked and brined shrimps packed in a modified atmosphere have a shelf-life of >7 months at 0°C, but spoil in 4–6 days at 25°C. Int. J. Food Sci. Technol. 35, 431–442. doi:10.1046/j.1365-2621.2000.00402.x
Davidson, C. M., and Cronin, F. (1973). Medium for the selective enumeration of lactic acid bacteria from foods. Appl. Microbiol. 26, 439–440. doi:10.1128/am.26.3.439-440.1973
Elavarasan, K., and Shamasundar, B. A. (2022). Antioxidant properties of papain mediated protein hydrolysates from fresh water carps (Catla catla, Labeo rohita and Cirrhinus mrigala) and its application on inhibition of lipid oxidation in oil sardine mince during ice storage. J. Food Sci. Technol. 59, 636–645. doi:10.1007/s13197-021-05053-0
Feng, H., Liu, J., Timira, V., Lin, H., Wang, H., and Li, Z. (2022). Effects of Plant extracts (Clove, Fennel, Rosemary, Cinnamon, and Tea Polyphenols) on microbiological, chemical, and sensory quality of smoke-flavored sea bass during chilled storage. ACS Food Sci. Technol. 2, 1692–1700. doi:10.1021/acsfoodscitech.2c00131
Fleet, G. H. (2011). “Yeast spoilage of foods and beverages,” in The yeasts (Elsevier), 53–63. doi:10.1016/B978-0-444-52149-1.00005-7
Freiding, S., Gutsche, K. A., Ehrmann, M. A., and Vogel, R. F. (2011). Genetic screening of Lactobacillus sakei and Lactobacillus curvatus strains for their peptidolytic system and amino acid metabolism, and comparison of their volatilomes in a model system. Syst. Appl. Microbiol. 34, 311–320. doi:10.1016/j.syapm.2010.12.006
Gorji, S. G., Calingacion, M., Smyth, H. E., and Fitzgerald, M. (2019). Comprehensive profiling of lipid oxidation volatile compounds during storage of mayonnaise. J. Food Sci. Technol. 56, 4076–4090. doi:10.1007/s13197-019-03876-6
Haastrup, M. K., Johansen, P., Malskær, A. H., Castro-Mejía, J. L., Kot, W., Krych, L., et al. (2018). Cheese brines from Danish dairies reveal a complex microbiota comprising several halotolerant bacteria and yeasts. Int. J. Food Microbiol. 285, 173–187. doi:10.1016/j.ijfoodmicro.2018.08.015
Han, S.-S., Jeong, Y.-S., and Choi, S.-K. (2021). Current scenario and challenges in the direct identification of microorganisms using MALDI TOF MS. Microorganisms 9, 1917. doi:10.3390/microorganisms9091917
Hernández, A., Pérez-Nevado, F., Ruiz-Moyano, S., Serradilla, M. J., Villalobos, M. C., Martín, A., et al. (2018). Spoilage yeasts: what are the sources of contamination of foods and beverages? Int. J. Food Microbiol. 286, 98–110. doi:10.1016/j.ijfoodmicro.2018.07.031
İnanli, A. G., Coban, Ö. E., and Dartay, M. (2010). The chemical and sensorial changes in rainbow trout caviar salted in different ratios during storage. Fish. Sci. 76, 879–883. doi:10.1007/s12562-010-0279-6
Iverson, S. J., Lang, S. L. C., and Cooper, M. H. (2001). Comparison of the Bligh and Dyer and Folch methods for total lipid determination in a broad range of marine tissue. Lipids 36, 1283–1287. doi:10.1007/s11745-001-0843-0
Jensen, H. A., Sørensen, A.-D. M., Mejlholm, O., Bøknæs, N., and Jacobsen, C. (2024). Effect of different antioxidants and application methods on the shelf-life of whole, cooked, frozen Northern shrimp (Pandalus borealis). Food Chem. Adv. 5, 100764. doi:10.1016/j.focha.2024.100764
Johannesson, J. (2006). Lumpfish caviar - from vessel to consumer. Rome: FAO, 60. FAO Fisheries Technical Paper. No. 485.
Jonsdottir, R., Olafsdottir, G., Martinsdottir, E., and Stefansson, G. (2004). Flavor characterization of ripened cod roe by gas chromatography, sensory analysis, and electronic nose. J. Agric. Food Chem. 52, 6250–6256. doi:10.1021/jf049456g
Kennedy, J., Durif, C. M. F., Florin, A.-B., Fréchet, A., Gauthier, J., Hüssy, K., et al. (2019). A brief history of lumpfishing, assessment, and management across the North Atlantic. ICES J. Mar. Sci. 76, 181–191. doi:10.1093/icesjms/fsy146
Koukou, I., Mejlholm, O., and Dalgaard, P. (2021). Cardinal parameter growth and growth boundary model for non-proteolytic Clostridium botulinum – effect of eight environmental factors. Int. J. Food Microbiol. 346, 109162. doi:10.1016/j.ijfoodmicro.2021.109162
Larsson, K., Almgren, A., and Undeland, I. (2007). Hemoglobin-mediated lipid oxidation and compositional characteristics of washed fish mince model systems made from cod (Gadus morhua), herring (Clupea harengus), and salmon (Salmo salar) muscle. J. Agric. Food Chem. 55, 9027–9035. doi:10.1021/jf070522z
Lee, H., Kizito, S. A., Weese, S. J., Craig-Schmidt, M. C., Lee, Y., Wei, C.-I., et al. (2003). Analysis of headspace volatile and oxidized volatile compounds in DHA-enriched fish oil on accelerated oxidative storage. J. Food Sci. 68, 2169–2177. doi:10.1111/j.1365-2621.2003.tb05742.x
Leroi, F. (2010). Occurrence and role of lactic acid bacteria in seafood products. Food Microbiol. 27, 698–709. doi:10.1016/j.fm.2010.05.016
Liu, F., Lu, H., Dong, B., Huang, X., Cheng, H., Qu, R., et al. (2023). Systematic evaluation of the viable microbiome in the human oral and gut samples with spike-in Gram+/– bacteria. mSystems 8, e0073822–22. doi:10.1128/msystems.00738-22
Lopez, A., Bellagamba, F., Tirloni, E., Vasconi, M., Stella, S., Bernardi, C., et al. (2021). Evolution of food safety features and volatile profile in white sturgeon caviar treated with different formulations of salt and preservatives during a long-term storage time. Foods 10, 850. doi:10.3390/foods10040850
Lu, F. S. H., Bruheim, I., Ale, M. T., and Jacobsen, C. (2015). The effect of thermal treatment on the quality changes of Antartic krill meal during the manufacturing process: high processing temperatures decrease product quality. Eur. J. Lipid Sci. Technol. 117, 411–420. doi:10.1002/ejlt.201400280
Mejlholm, O., and Dalgaard, P. (2009). Development and validation of an extensive growth and growth boundary model for Listeria monocytogenes in lightly preserved and ready-to-eat shrimp. J. Food Prot. 72, 2132–2143. doi:10.4315/0362-028X-72.10.2132
Mejlholm, O., Kjeldgaard, J., Modberg, A., Vest, M. B., Bøknæs, N., Koort, J., et al. (2008). Microbial changes and growth of Listeria monocytogenes during chilled storage of brined shrimp (Pandalus borealis). Int. J. Food Microbiol. 124, 250–259. doi:10.1016/j.ijfoodmicro.2008.03.022
NMKL (2006). Aerobic count and specific spoilage organisms in fish and fish products No. 184. Bergen, Norway: Nordic Committee on Food Analysis, 2–7.
Nonnemann, B., Lyhs, U., Svennesen, L., Kristensen, K. A., Klaas, I. C., and Pedersen, K. (2019). Bovine mastitis bacteria resolved by MALDI-TOF mass spectrometry. J. Dairy Sci. 102, 2515–2524. doi:10.3168/jds.2018-15424
Norkrans, B. (1966). Studies on marine occurring yeasts: growth related to pH, NaCl concentration and temperature. Arch. Mikrobiol. 54, 374–392. doi:10.1007/BF00406719
Ozogul, Y., Ayas, D., Yazgan, H., Ozogul, F., Boga, E. K., and Ozyurt, G. (2010). The capability of rosemary extract in preventing oxidation of fish lipid. Int. J. Food. Sci. Tech. 45, 1717–1723. doi:10.1111/j.1365-2621.2010.02326.x
Panjaitan, F. C. A., Yeh, N., and Chai, H. (2020). Effect of different processing on quality changes of barramundi (Lates calcarifer) caviar during chilled storage (4°C). J. Food Process. Preserv. 44. doi:10.1111/jfpp.14790
Pazos, M., Sánchez, L., and Medina, I. (2005). Alpha-tocopherol oxidation in fish muscle during chilling and frozen storage. J. Agric. Food Chem. 53, 4000–4005. doi:10.1021/jf047843k
Pountney, S. M., Lein, I., Selly, S.-L. C., Migaud, H., and Davie, A. (2022). Comparative proximate analysis of wild and captive lumpfish (Cyclopterus lumpus) eggs show deficiencies in captive eggs and possible egg quality determinants. Aquaculture 557, 738356. doi:10.1016/j.aquaculture.2022.738356
Refsgaard, H. H. F., Brockhoff, P. B., and Jensen, B. (1998). Sensory and chemical changes in farmed Atlantic salmon (Salmo salar) during frozen storage. J. Agric. Food Chem. 46, 3473–3479. doi:10.1021/jf980309q
Refsgaard, H. H. F., Brockhoff, P. M. B., and Jensen, B. (2000). Free polyunsaturated fatty acids cause taste deterioration of Salmon during frozen storage. J. Agric. Food Chem. 48, 3280–3285. doi:10.1021/jf000021c
Ross, T., Dalgaard, P., and Tienungoon, S. (2000). Predictive modelling of the growth and survival of Listeria in fishery products. Int. J. Food Microbiol. 62, 231–245. doi:10.1016/S0168-1605(00)00340-8
Shantha, N. C., and Decker, E. A. (1994). Rapid, sensitive, iron-based spectrophotometric methods for determination of peroxide values of food lipids. J. AOAC Int. 77, 421–424. doi:10.1093/jaoac/77.2.421
Vasconi, M., Tirloni, E., Stella, S., Coppola, C., Lopez, A., Bellagamba, F., et al. (2020). Comparison of chemical composition and safety issues in fish roe products: application of chemometrics to chemical data. Foods 9, 540. doi:10.3390/foods9050540
P. Vos, G. M. Garrity, D. Jones, N. R. Krieg, W. Ludwig, F. A. Raineyet al. (2009). Bergey’s manual of systematic bacteriology. Volume 3: the firmicutes. 2nd Edn. (NY: Springer New York). doi:10.1007/978-0-387-68489-5
Vyncke, W. (1970). Direct determination of the thiobarbituric acid value in trichloracetic acid extracts of fish as a measure of oxidative rancidity. Fette, Seifen, Anstrichm. 72, 1084–1087. doi:10.1002/lipi.19700721218
Yang, Y., Deng, Y., Shi, X., Liu, L., Yin, X., Zhao, W., et al. (2023). QMRA of beach water by Nanopore sequencing-based viability-metagenomics absolute quantification. Water Res. 235, 119858. doi:10.1016/j.watres.2023.119858
Yap, M., O’Sullivan, O., O’Toole, P. W., and Cotter, P. D. (2022). Development of sequencing-based methodologies to distinguish viable from non-viable cells in a bovine milk matrix: a pilot study. Front. Microbiol. 13, 1036643. doi:10.3389/fmicb.2022.1036643
Keywords: microbiota, free fatty acids, preservation, roe quality, lipid oxidation, rosemary extract
Citation: Jensen HA, Plaza LG, Houmann RH, Andreasen PB, Bøknæs N, Kragh ML, Hansen LT, Mejlholm O, Jacobsen C, Dalgaard P and Sørensen A-DM (2024) Microbial and chemical stability of lumpfish (Cyclopterus lumpus) roe during refrigerated storage in acid-salt brines with or without antioxidants. Front. Food. Sci. Technol. 4:1498035. doi: 10.3389/frfst.2024.1498035
Received: 18 September 2024; Accepted: 31 October 2024;
Published: 05 December 2024.
Edited by:
Aris E. Giannakas, University of Patras, GreeceReviewed by:
Nikolaos D. Andritsos, University of Patras, GreeceChunlu Qian, Yangzhou University, China
Copyright © 2024 Jensen, Plaza, Houmann, Andreasen, Bøknæs, Kragh, Hansen, Mejlholm, Jacobsen, Dalgaard and Sørensen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hanne Aarslev Jensen, aGFhakBmb29kLmR0dS5kaw==; Ann-Dorit Moltke Sørensen, YWRtc0Bmb29kLmR0dS5kaw==
†Present address: DTU Bioengineering, Kgs. Lyngby, Denmark
‡Deceased as of 5th January 2024