
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Endocrinol. , 11 June 2024
Sec. Neuroendocrine Science
Volume 15 - 2024 | https://doi.org/10.3389/fendo.2024.1382861
 Rosa Fernández1,2*
Rosa Fernández1,2* Karla Ramírez1,2
Karla Ramírez1,2 Roberto Lorente-Bermúdez1
Roberto Lorente-Bermúdez1 Esther Gómez-Gil3
Esther Gómez-Gil3 Mireia Mora4
Mireia Mora4 Antonio Guillamon5
Antonio Guillamon5 Eduardo Pásaro1,2
Eduardo Pásaro1,2Introduction: Gender incongruence (GI) is characterized by a marked incongruence between an individual’s experienced/expressed gender and the assigned sex at birth. It includes strong displeasure about his or her sexual anatomy and secondary sex characteristics. In some people, this condition produces a strong distress with anxiety and depression named gender dysphoria (GD). This condition appears to be associated with genetic, epigenetics, hormonal as well as social factors. Given that L-glutamate is the major excitatory neurotransmitter in the central nervous system, also associated with male sexual behavior as well as depression, we aimed to determine whether metabotropic glutamate receptors are involved in GD.
Methods: We analyzed 74 single nucleotide polymorphisms located at the metabotropic glutamate receptors (mGluR1, mGluR3, mGluR4, mGluR5, mGluR7 and mGluR8) in 94 transgender versus 94 cisgender people. The allele and genotype frequencies were analyzed by c2 test contrasting male and female cisgender and transgender populations. The strength of the associations was measured by binary logistic regression, estimating the odds ratio (OR) for each genotype. Measurement of linkage disequilibrium, and subsequent measurement of haplotype frequencies were also performed considering three levels of significance: P ≤ 0.05, P ≤ 0.005 and P ≤ 0.0005. Furthermore, false positives were controlled with the Bonferroni correction (P ≤ 0.05/74 = 0.00067).
Results: After analysis of allele and genotypic frequencies, we found twenty-five polymorphisms with significant differences at level P ≤ 0.05, five at P ≤ 0.005 and two at P ≤ 0.0005. Furthermore, the only two polymorphisms (rs9838094 and rs1818033) that passed the Bonferroni correction were both related to the metabotropic glutamate receptor 7 (mGluR7) and showed significant differences for multiple patterns of inheritance. Moreover, the haplotype T/G [OR=0.34 (0.19–0.62); P<0.0004] had a lower representation in the transgender population than in the cisgender population, with no evidence of sex cross-interaction.
Conclusion: We provide genetic evidence that the mGluR7, and therefore glutamatergic neurotransmission, may be involved in GI and GD.
Gender identity is the consciousness of being a man or a woman. For the majority of the population this awareness is congruent with the male or female aspect of their genitals; they are cisgender men or women (CM and CW respectively). However, for some people, gender identity is not congruent with their genitalia (ICD-11; 1), 2022), they are transgender men or women (TM and TW respectively). The transgender population is a heterogeneous group: some feel they belong to the binary male-female dichotomy (binary transgender people), while others feel they are agender, bigender, gender fluid, etc., they are non-binary transgender people (2). In some transgender persons the gender incongruence could drive deep depression and anxiety, called gender dysphoria (GD) by DSM-5 (3), that require gender affirming hormone treatment (4).
Since the studies of Swaab (5) on post mortem brains of male and female cisgender and female transgender people showing that the central part of the bed nucleus of the stria terminalis was feminized in female transgender, genetic and neuroimaging in vivo studies have been carried out within the framework of sexual differentiation, and a cortical neurodevelopmental theory has been put forward (6, 7). The genetic analysis of sex steroid receptor polymorphisms (8–16) and coactivator polymorphisms (17) in transgender population support the above mentioned theory.
Within the scope of sex differences, polymorphisms of the α and β estrogen receptors as well as of the androgen receptor present particular interest (13). In transgender women, complex interaction between estrogen and androgen receptor polymorphisms was unveiled. Thus, an inverse allele interaction between rs113770630 (ERβ) and rs193922933 (AR) is characteristic of the TW population: when either of these polymorphisms is short, the other is long. rs9340799 (ERα) and rs113770630 (ERβ) are also related to the TM population although no interaction between these two polymorphisms was evidenced (13).
The ERα and ERβ receptors were initially identified as intracellular, ligand-regulated transcription factors that are expressed throughout the body, and in numerous brain regions (18). But this classical mechanism only partially explains the wide variety of effects produced by estradiol (19). Thus, recent studies suggest that the rapid effects of estrogens are the result of a novel signaling mechanism from the cell membrane (20) resulting from the coupling of classical ERs localized at the membrane (mERs) with metabotropic glutamate receptors (mGluRs) (21). In the central nervous system, the signaling cascades initiated by the mER/mGluR coupling has been shown to be involved in many physiological functions in both sexes (19).
The mGluRs are a heterogeneous group that is categorized into three subgroups based on sequence homology, pharmacology, and downstream signaling (22): Group I, consists of mGluR1 and mGluR5, group II includes mGluR2 and mGluR3, while receptors mGluR4, mGluR6, mGluR7, mGluR8 form group III. Evidence of ER interactions with the three mGluR groups has been found. Group I is primarily characterized by their postsynaptic localization and their association with Gq G proteins. Group II is primarily localized presynaptically and associate with Gi/o G proteins. While Group III is also primarily expressed presynaptically and couple to Gi/o G proteins (23). All these receptors are expressed in the brain (23).
At behavioral level, mGluRs are implicated in sexual behavior. Specifically, mGluR5 and mGluR7 receptors influence rodent male sexual behavior (24–26). Furthermore, Schwarz and McCarthy reported that antagonizing glutamate receptors during the critical period of sexual differentiation blocks estradiol-induced defeminization but not masculinization of behavior in adulthood (27).
Moreover, mGluR receptors are of particular interest for transgender people because they show higher rates of attempted and suicidal ideation compared to the overall population (28, 29) and they are nearly twice as likely to die than cisgender people (30). Interestingly, it is worth noting that glutamatergic transmission is dysregulated in suicidal individuals (28, 31, 32).
Because of the coupling of classical ERs and mGluRs, and the implications of both types of receptors in brain sex differences and behavior, in order to delve into the molecular bases of gender identity, we analyzed 74 polymorphisms located at the metabotropic glutamate receptors (mGluR1, mGluR3, mGluR4, mGluR5, mGluR7 and mGluR8) in a homogeneous male and female transgender versus to a male and female cisgender populations.
The analyzed population consisted of 47 transgender women (TW) and 47 transgender men (TM) diagnosed and recruited through the Gender Identity Unit of the Clínic Hospital of Barcelona (Spain), and 50 cisgender women (CW) and 44 cisgender men (CM) obtained from the biobank of the Regional University Hospital of Málaga (Spain).
The inclusion criterion for the entire population that participated in the study was being older than 18 years, and specifically for the transgender population, the presence of the first symptoms of GI (ICD-11) before puberty (defined as early onset).
The exclusion criteria for all participants were: presence of medical or psychiatric disorders, and previous history of alcohol and/or drug abuse. All participants were matched by geographical origin (Spanish), ethnicity (Caucasian) and the sex assigned at birth. (Supplementary Table 1).
In the case of the transgender population, genomic DNA was obtained from blood using the DNeasy Blood & Tissue Kit from Qiagen (Madrid, Spain). For the cisgender population, we obtained the DNA samples from the biobank of the Regional University Hospital of Málaga (Spain).
All analyzed polymorphisms were single nucleotide polymorphisms (SNPs) located, according to the Ensembl database (www.ensembl.org/), at the metabotropic glutamate receptors (Supplementary Table 2). Genotyping was performed by the microarray Axiom Spanish Biobank (Affymetrix). Statistical analyses were performed using the free online software SNPStats (https://www.snpstats.net/) (33). SNPStats is a simple, free, ready-to-use software which has been designed to analyze genetic-epidemiology studies of association using SNPs. Once the genotype frequencies were uploaded, and for each selected SNP, the following were calculated: allele and genotype frequencies, test for Hardy-Weinberg equilibrium, analysis of association with a response variable based on linear or logistic regression, multiple inheritance models, linkage disequilibrium statistics, haplotype frequency estimation, analysis of association of haplotypes with the response and analysis of interactions (haplotypes-covariate).
The study was approved by the ethical committees of the National University of Distance Education (UNED, Madrid). At the start of the study, written informed consent was obtained from all participants.
The genetic analyses were conducted contrasting populations by their sex assigned at birth, considering three levels of significance: P ≤ 0.05, P ≤ 0.005 and P ≤ 0.0005. Moreover, we applied the Bonferroni correction (P ≤ 0.05/74 = 0.00067) to control the overall type I error rate. The allele and genotype frequencies were analyzed by χ2 test. The strength of the associations with GI was measured by binary logistic regression, estimating the odds ratio (OR) for each genotype for multiple patterns of inheritance. Furthermore, measurement of linkage disequilibrium, and subsequent measurement of haplotype frequencies were performed using logistic regression models to determine the strength of the associations.
We analyzed the allele and genotypic frequencies, the interactions with the covariate sex assigned at birth, and the linkage disequilibrium of 74 polymorphisms at the metabotropic glutamate receptors (mGluR) (Supplementary Table 2), in a transgender population versus a cisgender population with similar geographic origin (Spanish) and ethnicity (Caucasian).
The analyses were conducted considering three levels of significance: P ≤ 0.05, P ≤ 0.005 and P ≤ 0.0005, finding significant differences at the three levels: at the P ≤ 0.05 level, we found significant differences in 25 polymorphisms distributed among the mGluR1, mGluR4, mGluR5, mGluR7, and mGluR8 receptors (Supplementary Table 2). At the P<0.005 level, five polymorphisms (Supplementary Table 2), distributed among the metabotropic receptors mGluR5 and mGluR7, reached statistical significance (rs62237207, rs62237212, rs62237216, rs62237226, rs7782149). While at the P<0.0005 level, only two polymorphisms located at the mGluR7, rs9838094 and rs1818033, reached statistical significance. These two polymorphisms that passed the Bonferroni correction, were surrounded by other polymorphisms that reached significance at 0.05 or 0.005 levels (Supplementary Table 2).
The analysis of the allele and genotype frequencies with respect to the polymorphism rs9838094, showed significant differences at level P ≤ 0.0005 and P ≤ 0.005 respectively (Table 1). The ancestral allele G and the genotype G/G were over represented in the transgender population, while genotypes T/G and T/T were more frequent in the cisgender population. The genotype T/T was absent in the transgender population (Table 1).
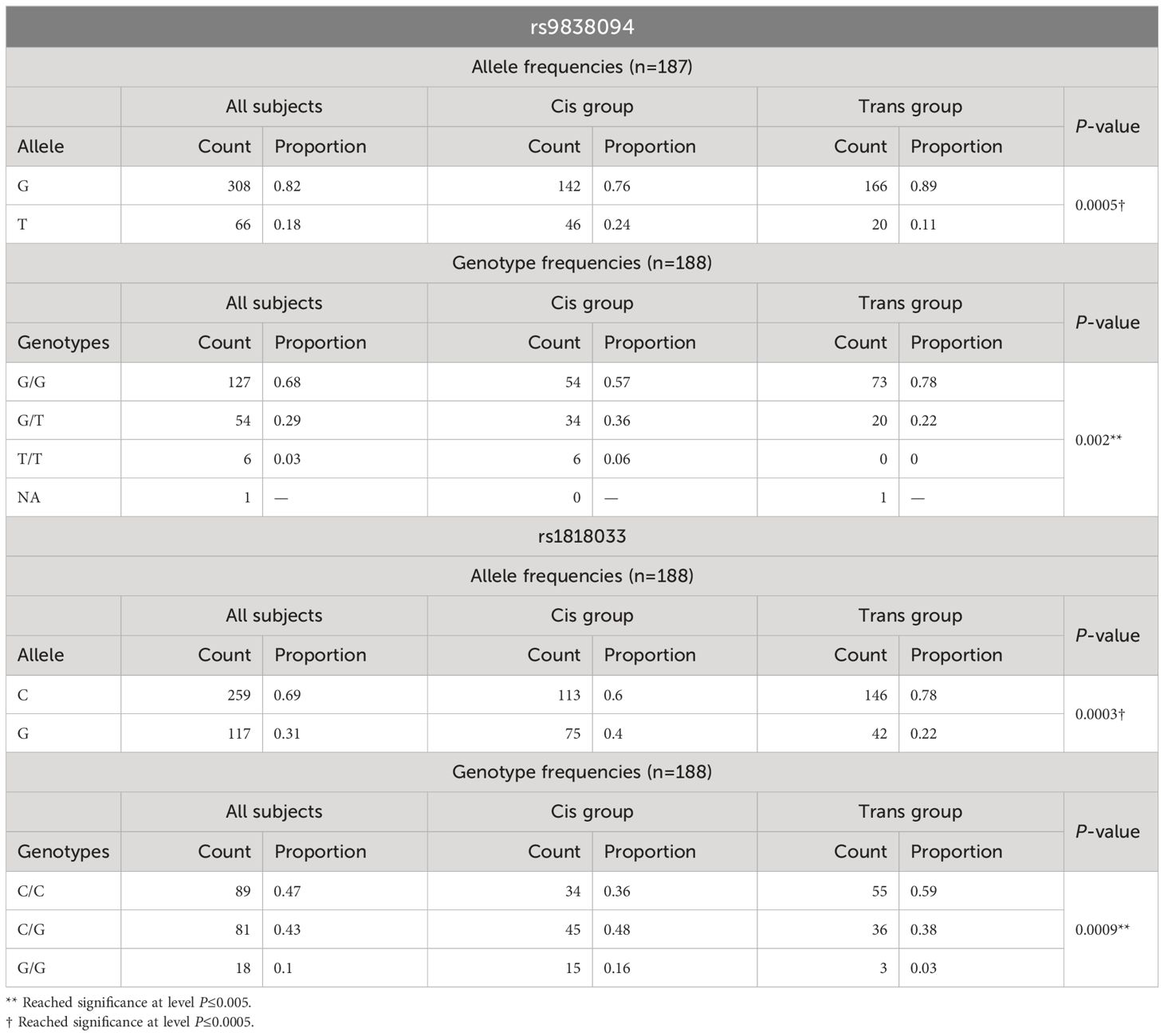
Table 1 Analysis of the allele and genotype frequencies for polymorphisms rs9838094 and rs9838094 in trans and cis populations.
The analysis of the genotype frequencies according to the different models of inheritance (Table 2) showed significant differences for all the inheritance patterns. The analysis of the covariate sex assigned at birth showed no statistical differences between males and females (Table 3).
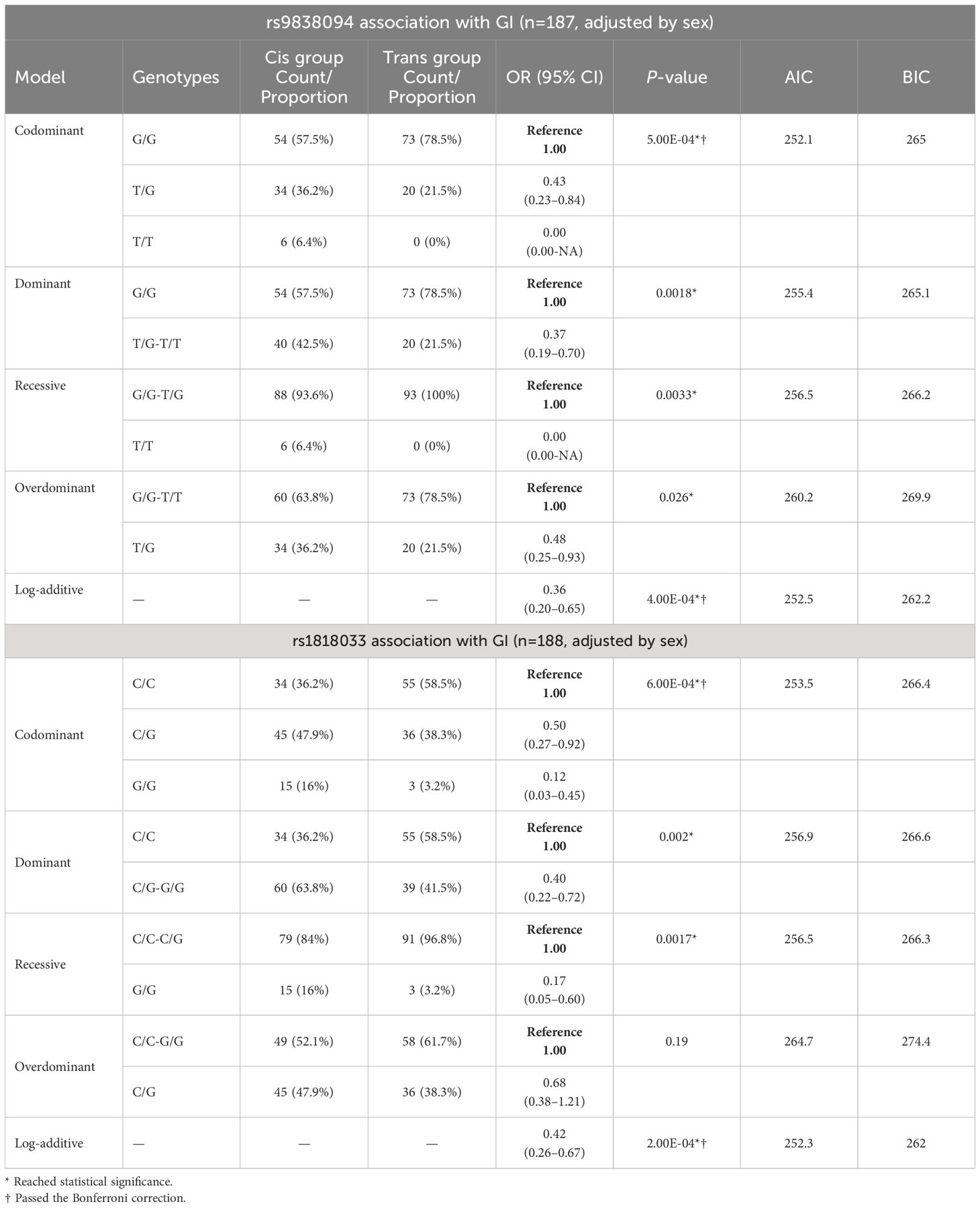
Table 2 rs9838094 and rs1818033 polymorphism association analysis with gender incongruence in trans and cis populations, in different models of inheritance.
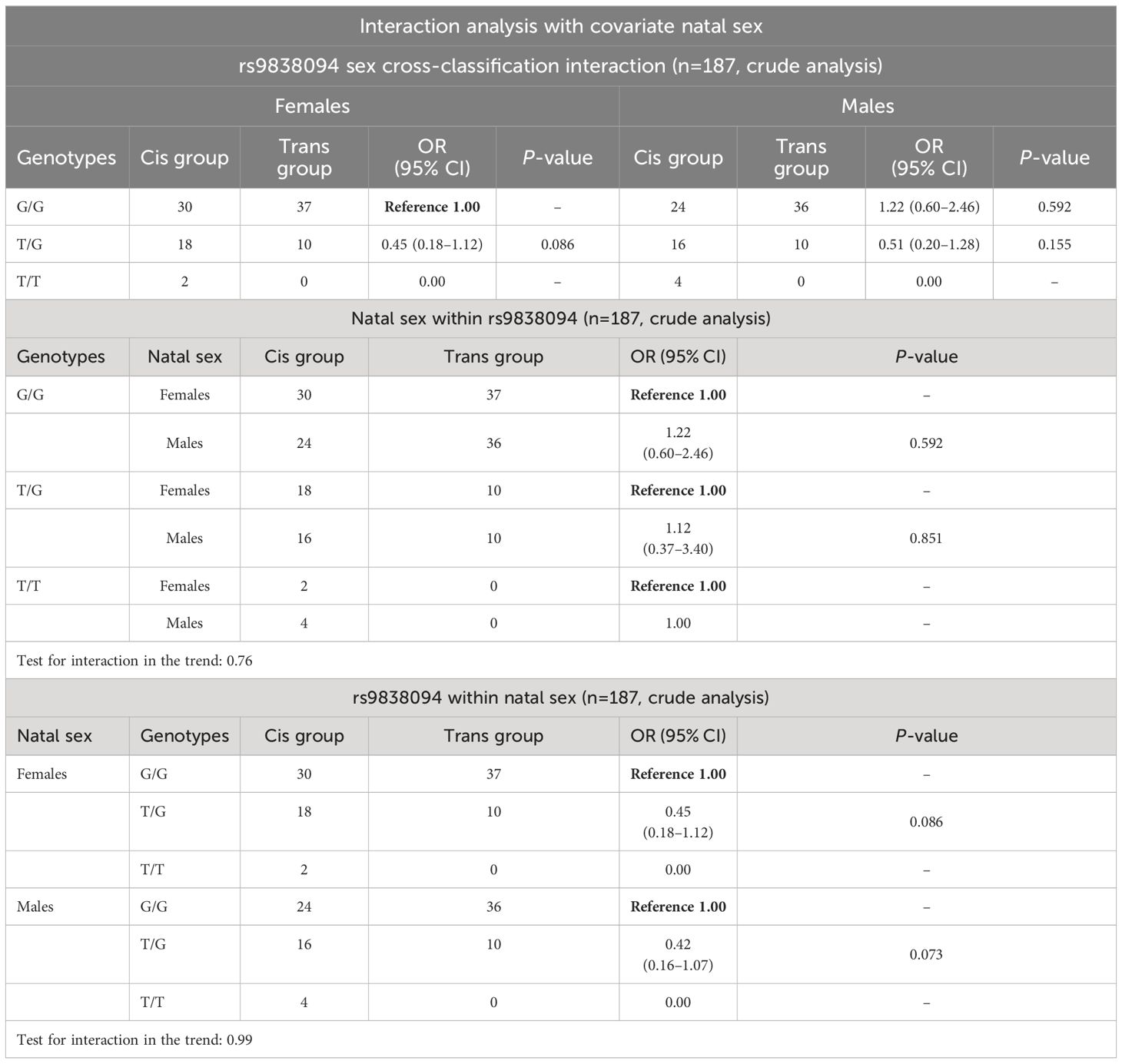
Table 3 Interaction analysis of the rs9838094 polymorphism with the covariate natal sex.
With respect to the polymorphism rs1818033, the C allele and the C/C genotype were overrepresented in the transgender population (Table 1). The association analysis with GI showed significant differences for multiple patterns of inheritance (codominant, dominant, recessive and log-additive) (Table 2). The analysis of the covariate sex assigned at birth showed no evidence of sex cross-interaction (Table 4).

Table 4 Interaction analysis of the rs1818033 polymorphism with the covariate natal sex.
We carried out the haplotype analysis of the two polymorphisms at the mGluR7 that passed the Bonferroni correction (D: 0.1215; D’: 0.9993; r: 0.6883 and P: 0). Linkage disequilibrium analysis between polymorphisms rs9838094 and rs1818033 showed that allele G from polymorphism rs9838094 was linked to allele C from polymorphism rs1818033 (haplotype 1: GC) (Table 5) and forms the most frequent haplotype (68.88%) in transgender and cisgender populations. Respect to the haplotype 2 (TG), it was more frequent in the cisgender population with an OR≤ 0.34 (0.19 - 0.62; P ≤ 0.0006; global haplotype association P ≤ 0.0004). Haplotype interaction analysis with the covariate sex assigned at birth showed that the haplotype T-G was more frequent in the cisgender population for both, the male, and the female populations, with no evidence of sex cross-interaction (Table 5).
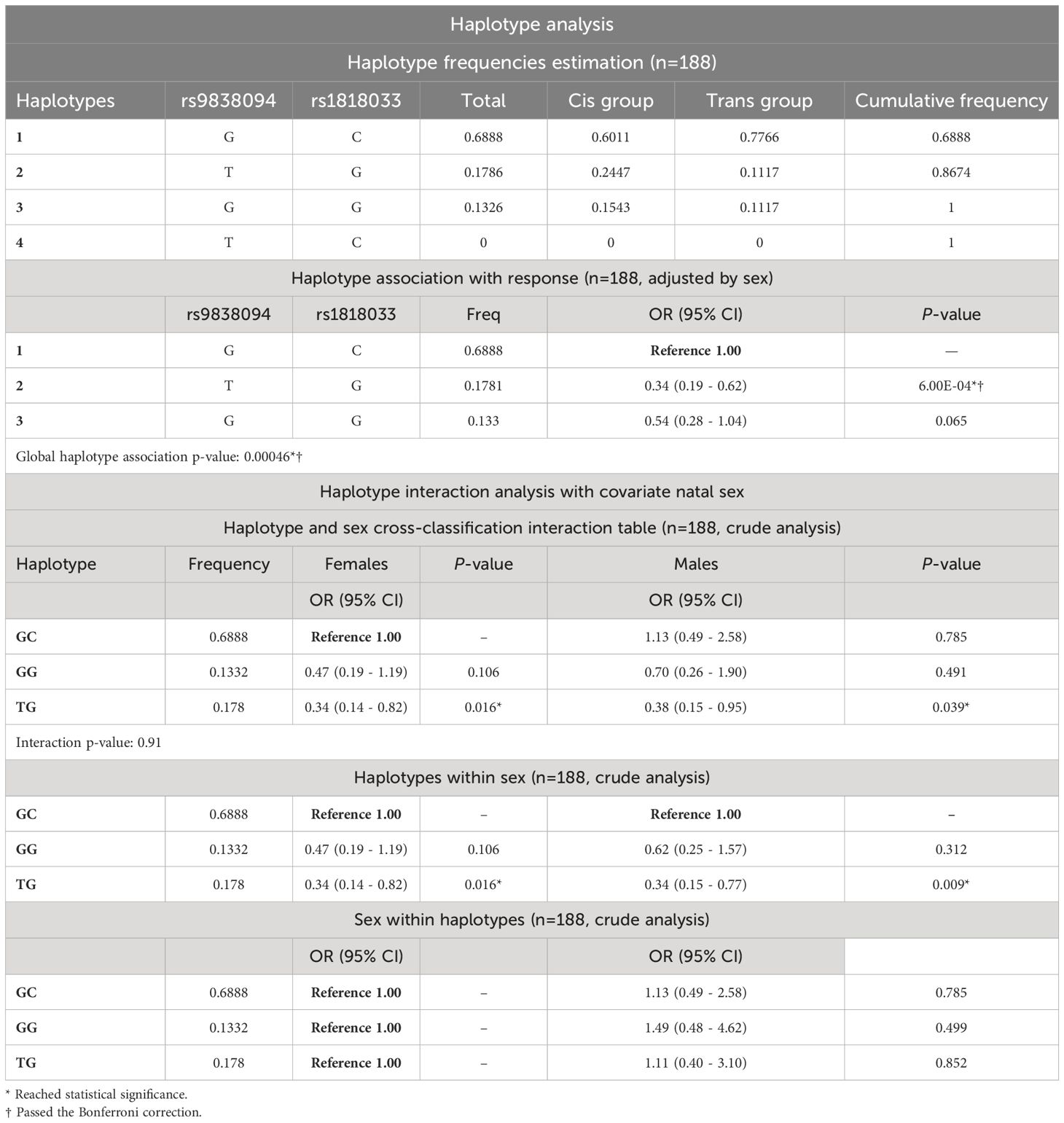
Table 5 Haplotype and Haplotype interaction analyses with the covariate natal sex. The table shows the haplotype frequencies estimation and the haplotype association with GI.
Seventy-four polymorphisms located at the metabotropic glutamate receptors (mGluRs) were analyzed at three levels of significance (P ≤ 0.05, P ≤ 0.005 and P ≤ 0.0005), founding 27 polymorphisms that reached statistical significance, two of which, located at the mGluR7, passed the Bonferroni correction. This is the first communication on the involvement of mGluR7 in gender identity.
Today, it is widely accepted that estradiol can act independently of the classical nuclear receptors, ERα and ERβ, by activating membrane-localized receptors (mER). Recent research indicates that mER signaling through mGluRs is an important and rapid mechanism by which estrogens can modulate neuronal and glial physiology, affecting various aspects of nervous system function (21, 23). There are data showing interaction between mGluRs and the ERs in several brain regions (23) that show sex differences in animals (34) and humans (35, 36). Sexual differentiation of the brain is developed by the intertwining work of gonadal hormones and neurotransmitters (37, 38). There are brain structural (6) and functional (7) differences between male and female cisgender and transgender people and a neurodevelopmental theory was proposed to explain these differences (6). The polymorphisms analyzed here show that some SNPs in the mGluR7 and mGluR5 receptors are more prevalent in transgender than in cisgender populations, suggesting the involvement of these receptors, at least mGluR7, in the development of transgender identity.
It is becoming clear that ER signaling through mGluRs is one important and rapid mechanism by which estrogens can modulate neuron and glial physiology, ultimately impacting various aspects of nervous system function (23). Studies of ERα and ERβ overexpression showed that a subpopulation of these classical receptors is trafficked to the membrane (membrane-bound estrogen receptors, mERs) activating intracellular signaling (23, 39). In addition, mERs interact with metabotropic glutamate receptors (mGluRs) (40, 41), such that mER/mGluR coupling initiates G-protein signaling cascades that rapidly affect cellular excitability and gene expression (19), influencing neuronal physiology, structure and behavior (23, 42).
Estrogens are known to be potent regulators of neuronal structure (increasing dendrite length and spine density) (43). In addition, rapid effects of estrogens have also been described for sexual behavior (44). On the basis of the above, we believe that it would be also necessary to analyze polymorphisms located in the GPER-1 (G protein-coupled estrogen receptor 1) gene, a specific membrane estrogen receptor (45, 46) that have not yet been analyzed in the transgender population.
On the other hand, the great variety of receptor pairs that are possible due to the existence of multiple subtypes of ER (α and β) and mGluRs (1–8 and subtypes), gives rise to a great diversity of molecular results that affect processes as diverse as cognition, motivation, movement or sexual behavior. For example, the mGluR5 plays an important role in the regulation of synaptic plasticity and the modulation of the neural network activity (47, 48) And both, mGluR5 and mGluR7 influence rodent male sexual behavior (24–26).
mGluR5, which is mainly located at the postsynaptic regions (49), is an important regulator of both excitatory and inhibitory pathways, and alterations in its expression are often related to a number of neurological and psychiatric conditions, including epilepsy, anxiety, and autism spectrum disorder (ASD) (50).
mGluR7, which is expressed presynaptic (51), is also widely distributed in the brain (52), and couples to G proteins. Under high glutamate concentrations, mGluR7 acts as auto-receptor to inhibit further neurotransmitter release (53). Moreover, mGluR7 also functions as a hetero receptor inhibiting GABA release (51, 54). Thus, activation of these receptors modulates glutamate release (55, 56) controlling the excitatory synapse function (57).
In addition, evidence suggests that mGluR7 exhibits a basal signaling (constitutive activity), even in the absence of its natural ligand (glutamate). When expressed in neurons, mGluR7 shows detectable basal calcium channel modulation without the need for strong receptor activation. This constitutive activity implies that mGluR7 may have a physiological role even when not fully activated by glutamate (58).
On the other hand, mGluR5 and mGluR7 are known to mediate emotional and social behavior (57, 59, 60). Thus, some studies have shown that mGluR5 protein levels in the amygdala increase concomitantly with anxiety behaviors in adolescent mice after two weeks of isolation (61). mGluR7 is also implicated in the pathogenesis of depression. Thus, some genetic polymorphisms located in this receptor are known to increase susceptibility to depression (62). Given that in the present study all transgender individuals who participated showed GD, we believe that to better understand this trait in the future, it might help to address the study of different polymorphisms located in mGluR5–7 genes.
Moreover, the GABAergic/glutamatergic system has been implicated in suicidal behaviors (63) and transgender population shows higher suicide ideation and attempts (28, 29). Post mortem studies of the dorsal prefrontal cortex show a generalized disruption of the regulation of the glutamate receptors in suicidals (32). Taking all this together, we believe that more attention should be paid to the possible link between mGluRs polymorphisms and suicide risk in transgender people. But we should not underestimate the obvious social, cultural, relational, and multiple other difficulties experienced by people with a non-conforming transgender identity. Not being fully accepted in society is an obvious non-negligible risk factor for mood disorders and suicide in transgender populations.
Our work has limitations as well as strengths. The main limitation is that the population analyzed was small. It would be necessary to replicate the data in a larger transgender population. Moreover, as with other studies, there is a lack of representativeness of cases because participants were recruited from a gender unit, this could be contributing to selection bias.
The strengths of our study are the control of the sample homogeneity. Although transgender people comprise a heterogeneous population, they can be stratified according to variables such as age of onset of dysphoria, geographic origin, and ethnicity. Our sample was rigorously controlled in all these aspects.
The original contributions presented in the study are included in the article/Supplementary material. Further inquiries can be directed to the corresponding author.
The studies involving humans were approved by Ethical committees of the National University of Distance Education (UNED, Madrid). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
RF: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. KR: Data curation, Formal analysis, Investigation, Writing – original draft, Writing – review & editing. RL: Formal analysis, Writing – original draft, Writing – review & editing, Data curation, Investigation. EG-G: Conceptualization, Investigation, Writing – original draft, Writing – review & editing. MM: Conceptualization, Investigation, Writing – original draft, Writing – review & editing. AG: Conceptualization, Investigation, Writing – original draft, Writing – review & editing, Funding acquisition, Methodology, Project administration, Resources, Supervision, Validation, Visualization. EP: Funding acquisition, Investigation, Methodology, Project administration, Supervision, Validation, Writing – original draft, Writing – review & editing, Resources, Visualization, Conceptualization, Data curation, Formal analysis.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by grants: Ministerio de Ciencia, Innovación y Universidades PGC2018–094919-B-C21 and PDI21–127547NB-C21 (AG), PGC2018–094919-B-C22 and PDI21–127547NB-C22 (RF, EP), and Xunta de Galicia ED431B 2022/16 (EP). None of these funding sources played any role in the writing of the manuscript or the decision to submit it for publication.
We thank all the people who have contributed directly or indirectly to this study, and in particular the trans and cis people who participated, without whose selfless participation this study could not have been carried out.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fendo.2024.1382861/full#supplementary-material
Supplementary Table 1 | Characteristics of the analyzed population.
1. World Health Organization. The ICD-11 classification of mental and behavioural disorders: diagnostic criteria for research. Geneva: W. H. Organization (2022).
2. Polderman TJC, Kreukels BPC, Irwig MS, Beach L, Chan Y-M, Derks EM, et al. The biological contributions to gender identity and gender diversity: bringing data to the table. Behav Genet. (2018) 48:95–108. doi: 10.1007/s10519-018-9889-z
3. American Psychiatric Association. Diagnostic and statistical manual of mental disorders. 5th ed. DSM-5. Washington, DC: American Psychiatric Publishing, Inc. (2013). doi: 10.1176/appi.books.9780890425596
4. Coleman E, Radix AE, Bouman WP, Brown GR, de Vries ALC, Deutsch MB, et al. Standards of care for the health of transgender and gender diverse people, version 8. Int J transgender Heal. (2022) 23:S1–S259. doi: 10.1080/26895269.2022.2100644
5. Zhou JN, Hofman M. a, Gooren LJ, Swaab DF. A sex difference in the human brain and its relation to transsexuality. Nature. (1995) 378:68–70. doi: 10.1038/378068a0
6. Guillamón A, Junque C, Gómez-Gil E. A review of the status of brain structure research in transsexualism. Arch Sex Behav. (2016) 45:1615–48. doi: 10.1007/s10508–016-0768–5
7. Uribe C, Junque C, Gómez-Gil E, Abos A, Mueller SC, Guillamon A. Brain network interactions in transgender individuals with gender incongruence. Neuroimage. (2020) 211:116613. doi: 10.1016/j.neuroimage.2020.116613
8. Henningsson S, Westberg L, Nilsson S, Lundstrom B, Ekselius L, Bodlund O, et al. Sex steroid-related genes and male-to-female transsexualism. Psychoneuroendocrinology. (2005) 30:657–64. doi: 10.1016/j.psyneuen.2005.02.006
9. Hare L, Bernard P, Sánchez FJJ, Baird PNN, Vilain E, Kennedy T, et al. Androgen receptor repeat length polymorphism associated with male-to-female transsexualism. Biol Psychiatry. (2009) 65:93–6. doi: 10.1016/j.biopsych.2008.08.033
10. Ujike H, Otani K, Nakatsuka M, Ishii K, Sasaki A, Oishi T, et al. Association study of gender identity disorder and sex hormone-related genes. Prog. Neuropsychopharmacol Biol Psychiatry. (2009) 33:1241–4. doi: 10.1016/j.pnpbp.2009.07.008
11. Fernández R, Esteva I, Gómez-Gil E, Rumbo T, Almaraz MC, Roda E, et al. The (CA)n polymorphism of ERβ Gene is associated with ftM transsexualism. J Sex Med. (2014) 11:720–8. doi: 10.1111/jsm.12398
12. Fernández R, Esteva I, Gómez-Gil E, Rumbo T, Almaraz MC, Roda E, et al. Association study of ERβ, AR, and CYP19A1 genes and mtF transsexualism. J Sex Med. (2014) 11:2986–94. doi: 10.1111/JSM.12673
13. Fernández R, Guillamon A, Cortés-Cortés J, Gómez-Gil E, Jácome A, Esteva I, et al. Molecular basis of gender dysphoria: androgen and estrogen receptor interaction. Psychoneuroendocrinology. (2018) 98:161–7. doi: 10.1016/j.psyneuen.2018.07.032
14. Cortés-Cortés J, Fernández R, Teijeiro N, Gómez-Gil E, Esteva I, Almaraz MC, et al. Genotypes and haplotypes of the estrogen receptor α Gene (ESR1) are associated with female-to-male gender dysphoria. J Sex Med. (2017) 14:464–72. doi: 10.1016/j.jsxm.2016.12.234
15. Foreman M, Hare L, York K, Balakrishnan K, Sánchez FJ, Harte F, et al. Genetic link between gender dysphoria and sex hormone signaling. J Clin Endocrinol Metab. (2019) 104:390–6. doi: 10.1210/jc.2018–01105
16. D’Andrea S, Pallotti F, Senofonte G, Castellini C, Paoli D, Lombardo F, et al. Polymorphic cytosine-adenine-guanine repeat length of androgen receptor gene and gender incongruence in trans women: A systematic review and meta-analysis of case-control studies. J Sex Med. (2020) 17:543–50. doi: 10.1016/j.jsxm.2019.12.010
17. Fernández R, Ramírez K, Delgado-Zayas E, Gómez-Gil E, Esteva I, Guillamon A, et al. An analysis of the role of estrogens and the receptor coactivators in the basis of gender incongruence. In: Kostoglou-Athanassiou DI, editor. Psychoneuroendocrinology. London, United Kingdom: ntechOpen (2021). p. 1–22. doi: 10.5772/intechopen.96668
18. Shughrue PJ, Komm B, Merchenthaler I. The distribution of estrogen receptor-beta mRNA in the rat hypothalamus(1996) (Accessed February 9, 2016). doi: 10.1016/S0039-128X(96)00222-X
19. Johnson CS, Micevych PE, Mermelstein PG. Membrane estrogen signaling in female reproduction and motivation. Front Endocrinol (Lausanne). (2022) 13:1009379. doi: 10.3389/fendo.2022.1009379
20. Prossnitz ER, Barton M. The G-protein-coupled estrogen receptor GPER in health and disease. Nat Rev Endocrinol. (2011) 7:715–26. doi: 10.1038/nrendo.2011.122
21. Meitzen J, Mermelstein PG. Estrogen receptors stimulate brain region specific metabotropic glutamate receptors to rapidly initiate signal transduction pathways. J Chem Neuroanat. (2011) 42:236–41. doi: 10.1016/j.jchemneu.2011.02.002
22. Niswender CM, Conn PJ. Metabotropic glutamate receptors: physiology, pharmacology, and disease. Annu Rev Pharmacol Toxicol. (2010) 50:295–322. doi: 10.1146/annurev.pharmtox.011008.145533
23. Gross KS, Mermelstein PG. Estrogen receptor signaling through metabotropic glutamate receptors. Vitam. Horm. (2020) 114:211–32. doi: 10.1016/bs.vh.2020.06.003
24. Melis MR, Succu S, Mascia MS, Cortis L, Argiolas A. Extracellular excitatory amino acids increase in the paraventricular nucleus of male rats during sexual activity: main role of N-methyl-d-aspartic acid receptors in erectile function. Eur J Neurosci. (2004) 19:2569–75. doi: 10.1111/j.0953-816X.2004.03362.x
25. Li X, Higley A, Song R, Xi Z-X. Effects of metabotropic glutamate receptor ligands on male sexual behavior in rats. Neuropharmacology. (2013) 66:373–81. doi: 10.1016/j.neuropharm.2012.08.006
26. Rodríguez-Manzo G. Glutamatergic transmission is involved in the long lasting sexual inhibition of sexually exhausted male rats. Pharmacol Biochem Behav. (2015) 131:64–70. doi: 10.1016/j.pbb.2015.02.002
27. Schwarz JM, McCarthy MM. The role of neonatal NMDA receptor activation in defeminization and masculinization of sex behavior in the rat. Horm. Behav. (2008) 54:662–8. doi: 10.1016/j.yhbeh.2008.07.004
28. Hochheimer M, Glick JL, Garrison-Desany H, Huhn AS. Transgender individuals are at higher risk for suicidal ideation and preparation than cisgender individuals in substance use treatment. Front Psychiatry. (2023) 14:1225673. doi: 10.3389/fpsyt.2023.1225673
29. Marconi E, Monti L, Marfoli A, Kotzalidis GD, Janiri D, Cianfriglia C, et al. A systematic review on gender dysphoria in adolescents and young adults: focus on suicidal and self-harming ideation and behaviours. Child Adolesc. Psychiatry Ment Health. (2023) 17:110–30. doi: 10.1186/s13034–023-00654–3
30. Hughes TL, Jackman K, Dorsen C, Arslanian-Engoren C, Ghazal L, Christenberry-Deceased T, et al. How can the nursing profession help reduce sexual and gender minority related health disparities: Recommendations from the National Nursing LGBTQ Health Summit. Nurs. Outlook. (2022) 70:513–24. doi: 10.1016/j.outlook.2022.02.005
31. Hashimoto K. Emerging role of glutamate in the pathophysiology of major depressive disorder. Brain Res Rev. (2009) 61:105–23. doi: 10.1016/j.brainresrev.2009.05.005
32. Gray AL, Hyde TM, Deep-Soboslay A, Kleinman JE, Sodhi MS. Sex differences in glutamate receptor gene expression in major depression and suicide. Mol Psychiatry. (2015) 20:1057–68. doi: 10.1038/mp.2015.114
33. Solé X, Guinó E, Valls J, Iniesta R, Moreno V. SNPStats: a web tool for the analysis of association studies. Bioinformatics. (2006) 22:1928–9. doi: 10.1093/bioinformatics/btl268
34. Segovia S, Guillamón A. Sexual dimorphism in the vomeronasal pathway and sex differences in reproductive behaviors(1993) (Accessed January 30, 2018). doi: 10.1016/0165-0173(93)90007-M
35. Garcia-Falgueras A, Junque C, Gimenez M, Caldu X, Segovia S, Guillamon A. Sex differences in the human olfactory system. Brain Res. (2006) 1116:103–11. doi: 10.1016/j.brainres.2006.07.115
36. Ruigrok ANV, Salimi-Khorshidi G, Lai M-C, Baron-Cohen S, Lombardo MV, Tait RJ, et al. A meta-analysis of sex differences in human brain structure. Neurosci Biobehav Rev. (2014) 39:34–50. doi: 10.1016/j.neubiorev.2013.12.004
37. Segovia S, Guillamón A, del Cerro MC, Ortega E, Pérez-Laso C, Rodriguez-Zafra M, et al. The development of brain sex differences: A multisignaling process. Behav Brain Res. (1999) 105:69–80. doi: 10.1016/s0166–4328(99)00083–2
38. McCarthy MM, Schwarz JM, Wright CL, Dean SL. Mechanisms mediating oestradiol modulation of the developing brain. J Neuroendocrinol. (2008) 20:777–83. doi: 10.1111/j.1365-2826.2008.01723.x
39. Razandi M, Pedram A, Greene GL, Levin ER. Cell membrane and nuclear estrogen receptors (ERs) originate from a single transcript: studies of ERalpha and ERbeta expressed in Chinese hamster ovary cells. Mol Endocrinol. (1999) 13:307–19. doi: 10.1210/mend.13.2.0239
40. Boulware MI, Weick JP, Becklund BR, Kuo SP, Groth RD, Mermelstein PG. Estradiol activates group I and II metabotropic glutamate receptor signaling, leading to opposing influences on cAMP response element-binding protein. J Neurosci Off J Soc Neurosci. (2005) 25:5066–78. doi: 10.1523/JNEUROSCI.1427–05.2005
41. Mermelstein PG. Membrane-localised oestrogen receptor alpha and beta influence neuronal activity through activation of metabotropic glutamate receptors. J Neuroendocrinol. (2009) 21:257–62. doi: 10.1111/j.1365-2826.2009.01838.x
42. Dewing P, Boulware MI, Sinchak K, Christensen A, Mermelstein PG, Micevych P. Membrane estrogen receptor-alpha interactions with metabotropic glutamate receptor 1a modulate female sexual receptivity in rats. J Neurosci Off J Soc Neurosci. (2007) 27:9294–300. doi: 10.1523/JNEUROSCI.0592–07.2007
43. Gould E, Woolley CS, Frankfurt M, McEwen BS. Gonadal steroids regulate dendritic spine density in hippocampal pyramidal cells in adulthood. J Neurosci. (1990) 10:1286–91. doi: 10.1523/JNEUROSCI.10-04-01286.1990
44. Balthazart J. Membrane-initiated actions of sex steroids and reproductive behavior: A historical account. Mol Cell Endocrinol. (2021) 538:111463. doi: 10.1016/j.mce.2021.111463
45. Filardo EJ, Quinn JA, Bland KI, Frackelton ARJ. Estrogen-induced activation of Erk-1 and Erk-2 requires the G protein-coupled receptor homolog, GPR30, and occurs via trans-activation of the epidermal growth factor receptor through release of HB-EGF. Mol Endocrinol. (2000) 14:1649–60. doi: 10.1210/mend.14.10.0532
46. Kelly MJ, Rønnekleiv OK. Minireview: neural signaling of estradiol in the hypothalamus. Mol Endocrinol. (2015) 29:645–57. doi: 10.1210/me.2014–1397
47. Minakami R, Katsuki F, Yamamoto T, Nakamura K, Sugiyama H. Molecular cloning and the functional expression of two isoforms of human metabotropic glutamate receptor subtype 5. Biochem Biophys Res Commun. (1994) 199:1136–43. doi: 10.1006/bbrc.1994.1349
48. Doré AS, Okrasa K, Patel JC, Serrano-Vega M, Bennett K, Cooke RM, et al. Structure of class C GPCR metabotropic glutamate receptor 5 transmembrane domain. Nature. (2014) 511:557–62. doi: 10.1038/nature13396
49. Palazzo E, Marabese I, de Novellis V, Rossi F, Maione S. Metabotropic glutamate receptor 7: from synaptic function to therapeutic implications. Curr Neuropharmacol. (2016) 14:504–13. doi: 10.2174/1570159x13666150716165323
50. Carey C, Singh N, Dunn JT, Sementa T, Mendez MA, Velthuis H, et al. From bench to bedside: The mGluR5 system in people with and without Autism Spectrum Disorder and animal model systems. Transl Psychiatry. (2022) 12:395. doi: 10.1038/s41398–022-02143–1
51. Dalezios Y, Luján R, Shigemoto R, Roberts JDB, Somogyi P. Enrichment of mGluR7a in the presynaptic active zones of GABAergic and non-GABAergic terminals on interneurons in the rat somatosensory cortex. Cereb Cortex. (2002) 12:961–74. doi: 10.1093/cercor/12.9.961
52. Kinzie JM, Saugstad JA, Westbrook GL, Segerson TP. Distribution of metabotropic glutamate receptor 7 messenger RNA in the developing and adult rat brain. Neuroscience. (1995) 69:167–76. doi: 10.1016/0306-4522(95)00244-D
53. Schoepp DD, Jane DE, Monn JA. Pharmacological agents acting at subtypes of metabotropic glutamate receptors. Neuropharmacology. (1999) 38:1431–76. doi: 10.1016/s0028–3908(99)00092–1
54. Somogyi P, Dalezios Y, Luján R, Roberts JDB, Watanabe M, Shigemoto R. High level of mGluR7 in the presynaptic active zones of select populations of GABAergic terminals innervating interneurons in the rat hippocampus. Eur J Neurosci. (2003) 17:2503–20. doi: 10.1046/j.1460-9568.2003.02697.x
55. Klausnitzer J, Kulla A, Manahan-Vaughan D. Role of the group III metabotropic glutamate receptor in LTP, depotentiation and LTD in dentate gyrus of freely moving rats. Neuropharmacology. (2004) 46:160–70. doi: 10.1016/j.neuropharm.2003.09.019
56. Altinbilek B, Manahan-Vaughan D. Antagonism of group III metabotropic glutamate receptors results in impairment of LTD but not LTP in the hippocampal CA1 region, and prevents long-term spatial memory. Eur J Neurosci. (2007) 26:1166–72. doi: 10.1111/j.1460-9568.2007.05742.x
57. Gee CE, Peterlik D, Neuhäuser C, Bouhelal R, Kaupmann K, Laue G, et al. Blocking metabotropic glutamate receptor subtype 7 (mGlu7) via the Venus flytrap domain (VFTD) inhibits amygdala plasticity, stress, and anxiety-related behavior. J Biol Chem. (2014) 289:10975–87. doi: 10.1074/jbc.M113.542654
58. Kammermeier PJ. Constitutive activity of metabotropic glutamate receptor 7. BMC Neurosci. (2015) 16:17. doi: 10.1186/s12868–015-0154–6
59. Nakanishi S. Metabotropic glutamate receptors: Synaptic transmission, modulation, and plasticity. Neuron. (1994) 13:1031–7. doi: 10.1016/0896–6273(94)90043–4
60. Wang X, Gao C, Zhang Y, Hu S, Qiao Y, Zhao Z, et al. Overexpression of mGluR7 in the prefrontal cortex attenuates autistic behaviors in mice. Front Cell Neurosci. (2021) 15:689611. doi: 10.3389/fncel.2021.689611
61. Lin S, Li X, Chen Y-H, Gao F, Chen H, Hu N-Y, et al. Social Isolation During Adolescence Induces Anxiety Behaviors and Enhances Firing Activity in BLA Pyramidal Neurons via mGluR5 Upregulation. Mol Neurobiol. (2018) 55:5310–20. doi: 10.1007/s12035–017-0766–1
62. Noroozi R, Taheri M, Omrani MD, Ghafouri-Fard S. Glutamate receptor metabotropic 7 (GRM7) gene polymorphisms in mood disorders and attention deficit hyperactive disorder. Neurochem Int. (2019) 129:104483–9. doi: 10.1016/j.neuint.2019.104483
Keywords: gender dysphoria (GD), gender incongruence, membrane-bound estrogen receptors, mERs, metabotropic glutamate receptors (mGluR), mGluR5, MGluR7, rapid estradiol signaling
Citation: Fernández R, Ramírez K, Lorente-Bermúdez R, Gómez-Gil E, Mora M, Guillamon A and Pásaro E (2024) Analysis of single nucleotide polymorphisms of the metabotropic glutamate receptors in a transgender population. Front. Endocrinol. 15:1382861. doi: 10.3389/fendo.2024.1382861
Received: 06 February 2024; Accepted: 21 May 2024;
Published: 11 June 2024.
Edited by:
Toshiya Funabashi, St. Marianna University School of Medicine, JapanReviewed by:
Wilson C. J. Chung, Kent State University, United StatesCopyright © 2024 Fernández, Ramírez, Lorente-Bermúdez, Gómez-Gil, Mora, Guillamon and Pásaro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rosa Fernández, cm9zYS5mZXJuYW5kZXpAdWRjLmVz
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.