 Wencai Wang
Wencai Wang Luyao Ma†
Luyao Ma† Yongqiang Zhao
Yongqiang Zhao Xianfeng Li
Xianfeng Li
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Endocrinol., 14 September 2023
Sec. Cancer Endocrinology
Volume 14 - 2023 | https://doi.org/10.3389/fendo.2023.1216817
This article is part of the Research TopicFrom Genetic Alterations to New Molecular Targets in Pituitary DisordersView all 5 articles
Pituitary adenoma (PA) is the third most common central nervous system tumor originating from the anterior pituitary, but its pathogenesis remains unclear. The Wnt signaling pathway is a conserved pathway involved in cell proliferation, Self-renewal of stem cells, and cell differentiation. It is related to the occurrence of various tumors, including PA. This article reviews the latest developments in Wnt pathway inhibitors and pathway-targeted drugs. It discusses the possibility of combining Wnt pathway inhibitors with immunotherapy to provide a theoretical basis for the combined treatment of PA.
PAs are the most common tumor in the sellar region, accounting for 10–15% of the total intracranial tumors (1) PAs arise from the adenohypophyseal cells of the anterior pituitary lobe and are benign tumors (2). There are general symptoms associated with PAs, such as hormone overproduction and compression of the optic chiasm and pituitary gland (3, 4). According to functional classification, PAs can be divided into active and inactive PAs. Functional PAs can be classified as prolactinoma, growth hormone adenoma, adrenocorticotropic hormone adenoma, thyrotropin-secreting adenoma, gonadotropin-secreting adenoma, mixed adenoma, unclassified adenoma (5). Despite recent progress in molecular genetics, the pathogenesis of PAs still needs to be fully understood since confirmed that it originated from monoclonal cells in 1990 (6). Recent researches indicate that the pathogenesis of PAs may be influenced by cell type specificity, gene mutation, and epigenetic changes (7, 8). Also, abnormal activation of the wnt pathway has been found to play a crucial role in developing PAs.
The Wnt signaling pathway is an ancient evolutionarily conserved pathway mediated by Wnt proteins. It involves multiple events during embryonic development and tissue homeostasis, including cell multiplication, stem cell self-renewal, and cell differentiation (9, 10).
Historically, the Wnt signaling pathway has been divided into two broad categories: canonical and noncanonical (10, 11). Canonical Wnt signaling is activated when Wnt proteins increase or Wnt signaling inhibitors decrease. When Wnt proteins bind to their specific cell membrane receptors, they activate FZD and LRP5/6 (12). It then activates DVL and inhibits the multiprotein activity of GSK3β, Axin1/Axin2, APC, and CK1 (13). Blocks β-catenin phosphorylation, resulting in cytoplasmic accumulation of β-catenin. Finally, the β-catenin accumulated in the cytoplasm is transported to the nucleus, where it interacts with LEF and TC, targeting downstream target genes CyclinD1, c-myc, MMPs, Etc., triggering transcriptional activation and promoting cell proliferation or migration (14, 15).
The atypical pathway is β-catenin independent and primarily activated by Wnt5a. It can be divided into two pathways, Wnt/Ca2+ and PCP, whose primary function is to regulate cell polarity and migration (16). The Wnt5a protein binds to the FZD protein, activates DVL, and triggers PLC to release calcium ions. Calcium-sensitive enzymes such as PKC, CaMkII, and calcium-sensitive calcineurin are activated. Ultimately, the activation of NFAT leads to transcriptional activation, cytoskeletal rearrangement, and cell adhesion and migration regulation (17–21). In the PCP pathway, the Wnt5a protein binds to FZD receptors, activates DVL, sparks Rho-associated kinases and Rac/JNK signaling, and stimulates intracellular actin polymerization. It ultimately regulates cell polarity and migration (22–24).
Research has demonstrated that abnormal activation of the Wnt pathway is relevant to the progression and pathogenesis of many neoplasms, including breast carcinoma (25), melanoma (26), Barrett’s esophagus (27), and colorectal cancer (28). These two signaling pathways often form a network of overlapping signals and mutual regulation, involving the recrudesce and progression of multiple illnesses (Figure 1).
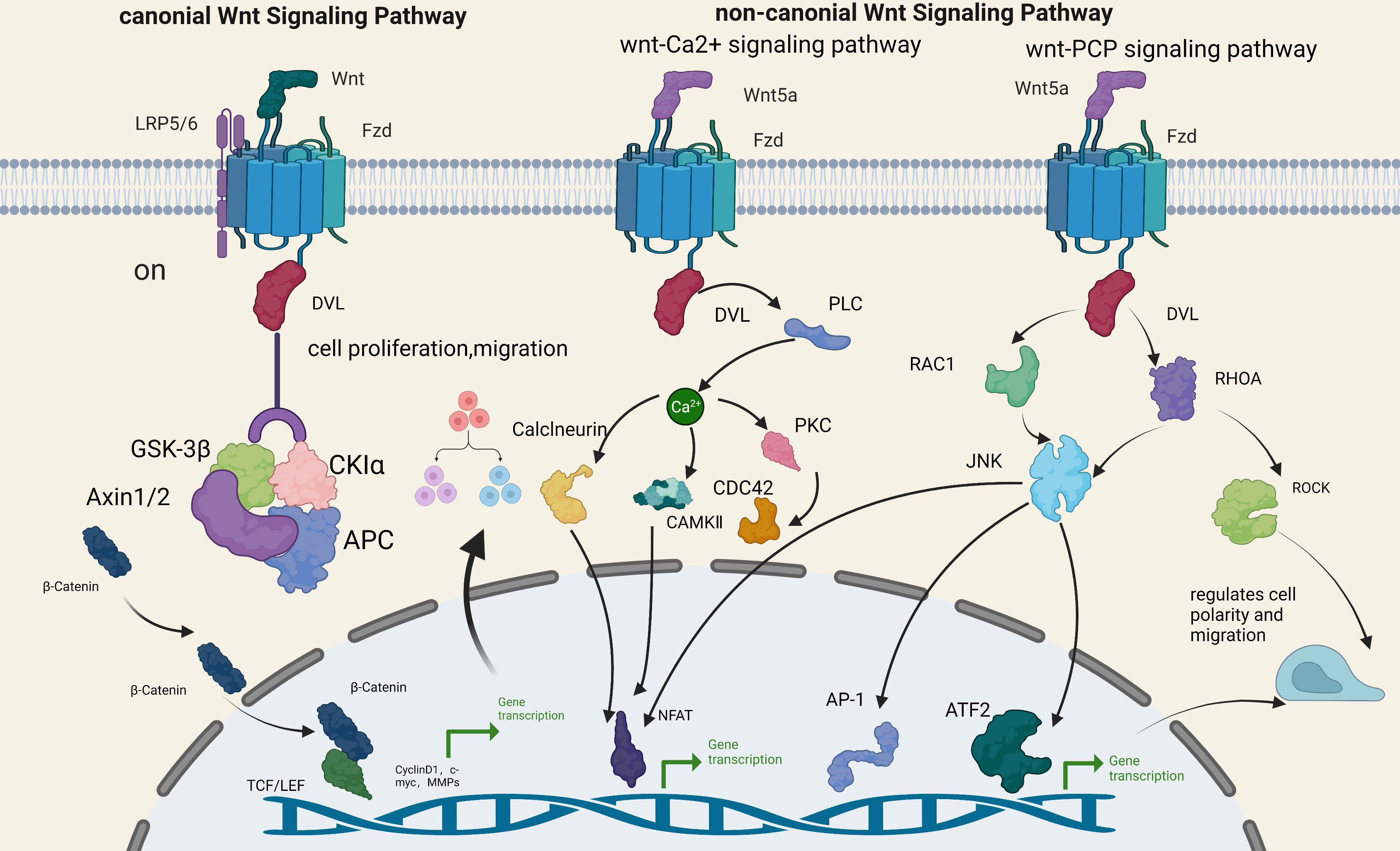
Figure 1 Wnt signaling pathway.
As early as 2005, Moreno et al. found that altered expression of SFRP1, TLE2, PITX2, NOTCH3, and DLK1 was observed in NFPA. The Wnt pathway may be crucial to the progression of NFPA (29). The Wnt4 molecule is a secreted glycoprotein involved in the multiplication and differentiation of normal and malignant cells. Although the expression of Wnt4 in various neoplasms has been extensively studied, its function in PAs remains unknown. In 2011, Gilles et al. showed that Wnt4 affects informal pituitary signals by restraining Ca2+ oscillations in GH3 cells for the first time, but the downstream effects are uncertain. It also suggests that Wnt4 is participated in adult tissue plasticity and may be related to β-catenin (30). In 2014, Weiping Li et al. discovered that the Wnt4 pathway was dysregulated in most PAs, and its overactivation could inhibit the invasion of PAs (31).
As the core protein of the canonical Wnt pathway, β-catenin is a downstream protein of Wnt4. It is crucial in promoting cellular proliferation and invasion in the PAs. In 2002, Howng et al. discovered that mutations in β-catenin, E-cadherin, and Wnt signaling pathways have participated in the progression of cerebral neoplasms (32). In 2014, Weiping Li and others found that Wnt4 and β-catenin were also overexpressed in most PAs in addition to ACTHPA (31). Furthermore, β-catenin is inversely correlated with the invasiveness of PAs. In 2015, Zhao et al. discovered that the stable β-catenin gene knockout PAs’ cell line was transfected with a plasmid. Then the cellular multiplication and invasion ability were detected by the CCK-8 kit and Transwell method. It was found that β-catenin could regulate AKT and STAT3. The expression of MMP-2/9, cyclin D1, and CDK4 boosted the multiplication and invasiveness of PAs (33).
Other downstream signaling targets, such as c-myc, have also been shown to be essential for forming PAs. C-myc may facilitate tumor growth, cell transformation, and evolution. Nevertheless, c-myc did not affect the recurrence of functional tumors (34). In 2017, Liu et al. found that β-catenin and c-myc are helpful immunohistochemical biomarkers for detecting invasive NFPA. However, they need to be validated in many samples before they can be used to predict the recurrence of NFPA (35).
In addition, pituitary homeobox 2, one of the downstream signaling targets of the Wnt pathway, also plays a significant part in pituitary formation. In 2011, Acunzo et al. found that the Wnt pathway-induced PITX2 gene may participate in the emergence of gonadotropin-secreting adenoma and NFPA through anti-apoptosis (36). In 2013, Lee et al. demonstrated for the first time that multidrug resistance p-glycoprotein, as a target gene of PITX2, can boost multidrug resistance and cellular endurance of cancer cells (37). Therefore, comprehending the function of the Wnt pathway in PAs could be further enriched by examining PITX2.
However, PAs often shares multiple signaling pathways with other tumors rather than being induced by wnt signaling alone. As the study by Hosseinkhan et al., in NFPA, Wnt, MAPK, TGF-b, Hippo, VEGF, JAK-STAT, PI3K-Akt, ErbB, TGF-b, and Ras signaling pathways are intertwined and jointly participate in the formation of tumors (38). Crosstalk between the Wnt pathway and other pathways is common in many tumors, including melanoma (39). Likewise, it was recently discovered in PAs. In 2019, SOX2 was shown to promote the cell proliferation of PAs via intermediating crosstalk between Wnt and Shh pathways (40).
Conventionally, inhibitors of Wnt signaling can be divided into two categories. The first class includes the SFRP family, WIF1, and Cerberus (41). They could theoretically block canonical or noncanonical pathways via directly binding to Wnt ligands and inhibiting their association with FZD receptor complexes (42). The second group mainly includes the DKK families. They block the canonical Wnt pathway via binding to LRP5 or LRP6 coreceptors (43, 44).
The SFRP family, located at 8q12-11.1, participates in the competitive binding of Wnt proteins through the N-terminal CRD structure (homologous to the FZD receptor). It incorporates five members (SFRP1-SFRP5). Melkonyan et al. identified SFRP1, SFRP2, and SFRP5 in mouse embryonic cells (45). By contrast, Rattner et al. discovered SFRP3 and SFRP4 (46). SFRP, an antagonist in several cancers, is a downstream target of the Wnt pathway. It plays a vital part in PA’s wnt signaling. In 2015, Wu et al. first found that SFRP4 promoter methylation was increased in invasive PAs and that SFRP4 promoter methylation decreased SFRP4 expression (47). The following year, the team found that sFRP2 inhibits tumor growth and invasion by regulating the wnt signaling pathway. The study is the first to show that sFRP2 was negatively correlated with the aggression of NFPA (48). In 2018, Ren et al. found an association between low expression of SFRP2 and features of invasive adenomas, including more prominent size and invasiveness. Overexpression of SFRP2 decreased β-catenin and suppressed the Wnt pathway in ACTH adenoma cells, thereby reducing the production of ACTH (49). Therefore, sFRP4 and sFRP2 may play a tumor suppressor role in PAs, and their expression may serve as biomarkers of aggressiveness and prognosis in PAs.
WIF1, the first secretory antagonist found in the retina, is located at 12q14.3 and is a crucial regulator of Wnt signaling (50–52). Hypermethylation of CpG promoters is downregulated. It directly relates to Wnt protein binding and inhibits the receptor signaling complex. When Wnt protein increases and WIF1 decreases, Wnt signaling is activated. Then Dsh is activated, which inhibits the activity of the APC-axin-GSK3 complex, resulting in the blocking of the phosphorylation and degradation of β-catenin. Finally, it results in β-Catenin gathering stably in the cytoplasm. β-catenin also translocates to the nucleus and plays a role in activating the transcriptional activity of T cell factor/lymphocyte enhancer, inducing and activating downstream target genes CyclinD1, c-myc, MMPs, Etc. It promotes and participates in forming tumors that can grow or migrate. In 2018, Song et al. found that the downregulation of WIF1 and sFRP4 are inversely correlated with the aggressiveness of NFPA. The results showed that miRNA-137 inhibited the invasion of NFPA by affecting the methylation of the WIF1 promoter and down-regulating WIF1 in the Wnt signaling pathway (53). One month later, Zhu et al. discovered that the expression of TGF-b1 and WIF1 in NFPA is correlated with cell multiplication and relapse (54). In recent years, studies have found that during the development of cancer cells, the transcription of SLC20A1 can absorb phosphate from interstitial fluid for cell metabolism, Signal transduction, and nucleic acid synthesis. In 2019, Li et al. found that high levels of SLC20A1 could reduce the protein levels of SFRP4 and WIF1 by activating the Wnt pathway, then facilitating the multiplication, invasiveness, and recurrence of GH adenoma. SLC20A1 is positively associated with β-catenin and negatively associated with WIF1 (55). At the same time, Cheng established a clinical prediction model from 295 NFPA tumor samples and found that three proteins, CDKN2A/p16, WIF1, TGF-β, tumor age and volume, and two clinical features were significantly associated with the recurrence of NFPA (56). Soon, Lei et al. found that miR-137 upregulated WIF1 in the Wnt pathway and inhibited the nuclear translocation of β-catenin by affecting the promoter methylation of WIF1, thereby inhibiting the growth and invasion of PRL tumors (57). Therefore, WIF1, as the primary inhibitor of the Wnt pathway, widely regulates the growth, invasion, and proliferation of PAs and can be used as a promising biomarker of PAs.
The DKK family is located at 10q11 and is mainly related to the atypical Wnt pathway by binding LRP5/6 and may simultaneously affect the canonical inhibitors of the Wnt pathway (43). DKK4 behaves like DKK1. However, the role of DKK2 in Wnt signaling remains controversial. Studies have shown that DKK2 could inhibit and activate the Wnt pathway under certain conditions (58). DKK3 (also known as DKKL1) is still considered an inhibitor of Wnt signaling, but its function must be fully comprehended (41). The clinical features of ACTH adenoma and acromegaly are usually disorders of bone metabolism, often accompanied by abnormal osteomalacia and osteoporosis. Research has confirmed that the Wnt pathway is essential in osteogenesis (59). In 2018, Belaya et al. found that acromegaly affects osteoblasts by significantly increasing the mRNA levels of DKK1 and wnt10B and altering the expression of miRNAs that participated in mesenchymal stem cellular binding (60). ACTH adenomas produce cortisol through the ACTH target gland axis, leading to osteoporosis in patients. The team also found that excess endogenous GC in humans inhibits osteogenesis by upregulating DKK1 and inactivating microRNAs involved in mesenchymal stem cell binding (61). Therefore, the Wnt pathway cannot be ignored for the secondary complications of osteoporosis and skeletal dysplasia in patients of PA. As a potent inhibitor of this signaling pathway, DKK also plays a crucial regulatory part. Therefore, further research on the Wnt pathway is significant for rationalizing treatment options and preventing subsequent complications in patients with PA.
Recently, many researchers have focused on non-coding RNAs, which also have inhibitory effects on Wnt signaling. In 2018, Song et al. miRNAs, including miRNA-137, were found to affect Wnt signaling by regulating WIF1 methylation (53). In 2019, Wang et al. showed that foxp1-induced CLRN1-AS1 ncRNA suppresses PRL tumors by blocking the miR-217/DKK1 axis to inactivate the Wnt pathway (62). In December of the same year, Lei et al. showed that miR-137 suppresses the growth and aggression of PRL tumors via inhibiting the Wnt pathway, upregulating WIF-1, and inhibiting β-catenin nuclear translocation (57). Currently, published studies have shown that many non-coding RNAs directly or indirectly inhibit the Wnt pathway, thereby inhibiting the development of PAs. In 2019, Shen et al. found that microRNA-543 downregulates Smad7, activates the Wnt pathway, boosts the invasion of PAs, and prevents apoptosis (63). In October of the same year, Zhang et al. PVT1 (lncRNA) was discovered to take a tumorigenic part in PA by activating the Wnt pathway (64). CircRNAs are one of three common ncRNAs that act as microRNA sponges to reverse the repression of target genes by disease-associated miRNAs and regulate disease-associated progression. CircRNAs involve the relapse and growth of various neoplasms, such as hepatic carcinoma (65), by promoting the Wnt signaling pathway. In 2020, Du et al. Through extensive analysis of circular RNAs, it was discovered that the involvement of hsa_circ_0001368 can noticeably promote the multiplication, aggressiveness, and GH secretion levels of GHPA cells. Database analysis found that the wnt pathway is one of the most enriched pathways for target genes (66). Therefore, circRNAs may participate in the development and pathogenesis of PAs by promoting wnt signaling. In 2022, Vetrivel et al. described the ACTH-independent expression of miR-1247-5p and miR-379-5p in CS for the first time. Then, they identified distinct adrenal miRNAs associated with CS subtypes and showed that miRNAs regulate Wnt signaling. Different genes may be individually involved in the pathology of specific subtypes of Cushing’s disease (67). Therefore, ncRNAs may not only act as inhibitors of the Wnt pathway to inhibit the tumorigenesis of PAs but also function in PAs as activators of the Wnt pathway. However, the specific mechanism of ncRNA’s dual role in PAs is unclear and needs further investigation.
According to previous studies, the abnormal expression of wnt pathway inhibitors may be related to PAs’ multiplication, aggressiveness, and typing. The research on Wnt pathway inhibitors will help to study the detailed mechanism of drugs targeting the Wnt pathway and treat PAs and their complications (Table 1).
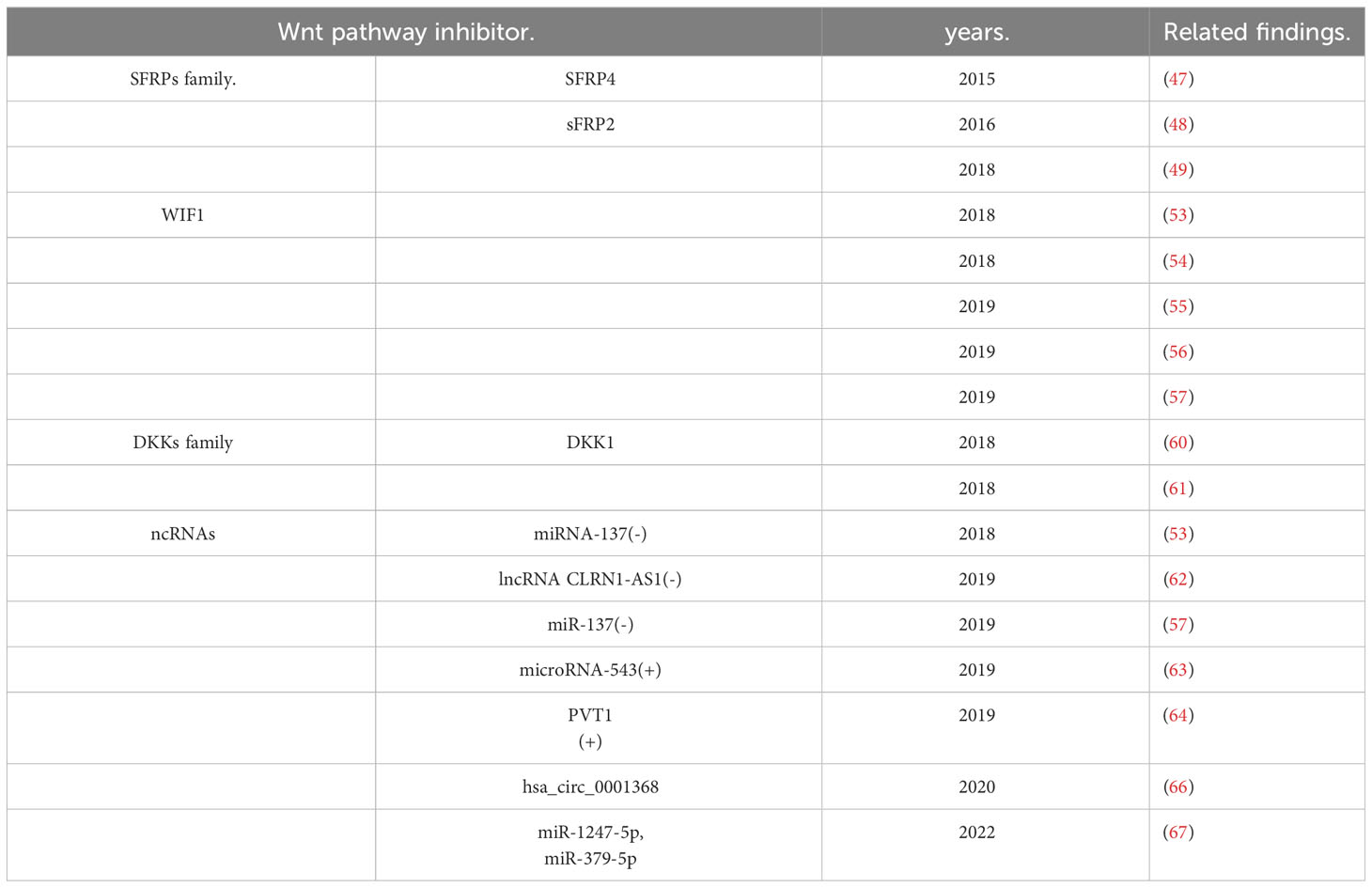
Table 1 Related findings of Wnt pathway inhibitors in PA.
Currently, prolactinomas are typically treated with drug therapy, whereas other types of PAs are often treated with surgery (68). If the symptoms can be treated or alleviated by targeting biological factors, it can bring great convenience to the clinical management of patients and reduce the need for surgery. Therefore, more research on non-surgical treatment is needed. In recent years, drug research targeting the wnt pathway has undoubtedly opened up a new direction for treating PAs.
In 2012, Bai et al. found that fulvestrant is a novel estrogen receptor antagonist that downregulates the expression of estrogen receptor α and Wnt4, upregulates the expression of WIF-1 and inhibits the cell proliferation of GH3 through estrogen receptor α and Wnt signaling pathways (69). The following year, the research team found through in vitro experiments that fulvestrant is closely related to the atypical Wnt signaling pathway. Furthermore, they also suggest that fulvestrant can be used as a therapeutic agent (70). In two 2014 papers, Cao and co-workers showed that fulvestrant could inhibit the cellular multiplication of PAs and the secretion of PRL via the Wnt pathway (71, 72). Fulvestrant’s main side effects are vasodilation and nausea (73). It is also hepatotoxic and can cause clinical symptoms such as jaundice and elevated transaminases (74). In 2019. Lei et al. found that decitabine increases WIF1 and promotes miR-137 targeting MITF. Moreover, it ultimately affects Wnt signaling, inhibiting the growth and invasion of prolactinoma (57). The most common side effects of decitabine were neutropenia, thrombocytopenia, and febrile neutropenia (75, 76).
By controlling the Wnt pathway, NEK2 can regulate the biological behavior of neoplasms, including liver cancer (77), glioblastoma (78), Etc. In 2021, Jian et al. confirmed that overexpression of NEK2 can dramatically boost the multiplication of PA cells by promoting the Wnt signaling pathway and reducing the ability of cells to have CAB sensitivity (79). Therefore, the antitumor effect of dopamine receptor agonists on PA can be enhanced by knocking down NEK2 and inhibiting wnt signaling. However, CAB should be used with particular caution in valvular heart disease and psychiatric disorders (80).
In addition, the antineoplastic drug TMZ was recently found effective against pituitary tumors. In 2022, Demarchi et al. showed that the canonical wnt pathway is involved in prolactin production and TMZ treatment in pituitary tumors. Anti-dopamine prolactinomas associated with normal pituitary undergo β-catenin relocalization. Furthermore, TMZ inhibits the tumorigenicity of prolactinomas by reducing the activation of β-catenin and the production of PRL (81). That same year, the team reported a case of an aggressive ACTH tumor in which TMZ resistance appeared relevant to β-catenin activation (82). TMZ is generally well tolerated and safe. The most common side effects are vomiting, nausea, fatigue, neutropenia, and thrombocytopenia. However, severe hematologic adverse events have also been reported, including aplastic anemia (83) and myelodysplastic syndrome (84). Drug rash, eosinophilia, systemic symptoms syndrome, and opportunistic infection (85) are the sporadic but highly challenging complications of TMZ therapy (86). The Wnt pathway is vital in treating prolactinoma, aggressive ACTHPA, and TMZ tumors, providing a new direction for curing aggressive and drug-resistant PAs.
Natural plant products have become an essential source of new anticancer drugs due to their multiple modes of action, multiple targets, safety, low toxicity, and few side effects (87). In 2017, Li et al. found that the CAG of Yunshi can induce apoptosis in cells of PAs by inhibiting the Wnt pathway. Moreover, dysregulation of the Wnt pathway may trigger ER stress in AtT-20 cells and play a vital role in apoptosis (88). In the same month, the research team found that tanshinone IIA increased β-catenin phosphorylation, inhibited β-catenin nuclear translocation, decreased β-catenin/TCF-4 complex formation, and TCF-LEF luciferase reporter gene activity, and subsequently decreased expression of cyclin D1 and MGMT. Ultimately, it can induce apoptosis in cells of PAs (89).
In addition to these drugs, it is also worth mentioning that they improve TME through the wnt pathway, thereby enhancing the efficacy of immunotherapy (90). The TME plays a constructive role in stimulating the invasiveness and persistence of PAs. It contains an extracellular matrix, CAFs, oncogenic immune cells, and other factors that can influence tumor tissue behavior. For example, TAM and TILs are related to NFPA, GH adenoma invasiveness, and tumor-aggressive behavior (91). TGFβ, FGF2, cytokines, chemokines, and growth factors released by CAFs may facilitate drug resistance, tumor fibrosis, and inflammation in PRL tumors and GH adenomas. Inhibition of Wnt signaling further suppressed the proliferation of dopamine-resistant PRL tumor cells. Invasion proteins secreted by the extracellular matrix of malignant tumors are relevant to increased angiogenesis (91–93). The TME is regulated by complex interactions of Wnt agonists, antagonists, and anti-antagonists. Therefore, improving the TME through modulation of the Wnt pathway is crucial in treating aggressive and refractory PA.
The Wnt signaling pathway has been proven to participate in various phases of regulating dendritic cells, the tumor immune cycle, T cells, and tumor cells. Although immunotherapy, especially immune checkpoint blockade therapy, has achieved good results in treating malignant tumors, some patients still have poor initial responses to immunotherapy or develop drug resistance after long-term treatment. Thus, inhibition of Wnt signaling plays a role in compensating for a deficiency in the immunotherapy of PA. Simultaneous use of Wnt pathway suppressants is supposed to improve TME and immunotherapy’s efficacy, pointing out a new direction for treating PA (Table 2; Figure 2).
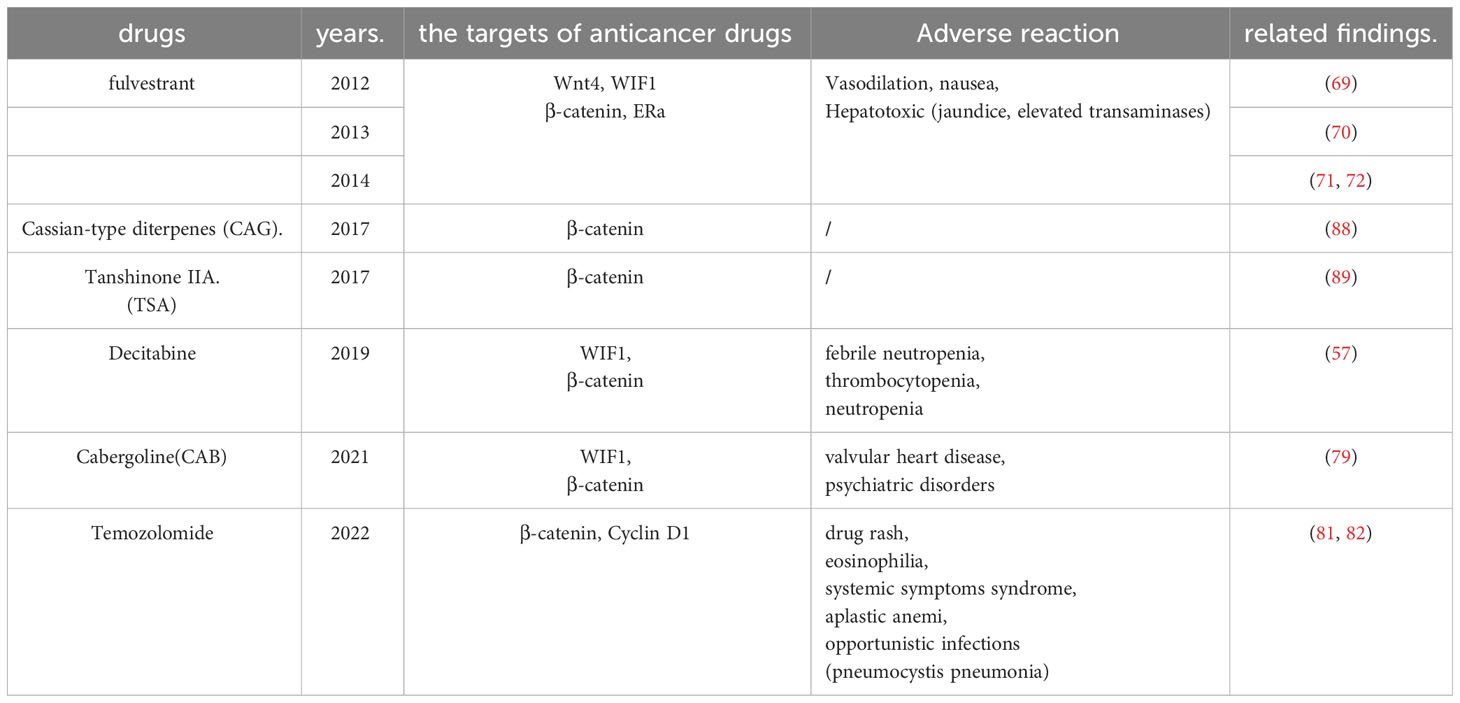
Table 2 Related research on drugs targeting WNT signaling pathway.
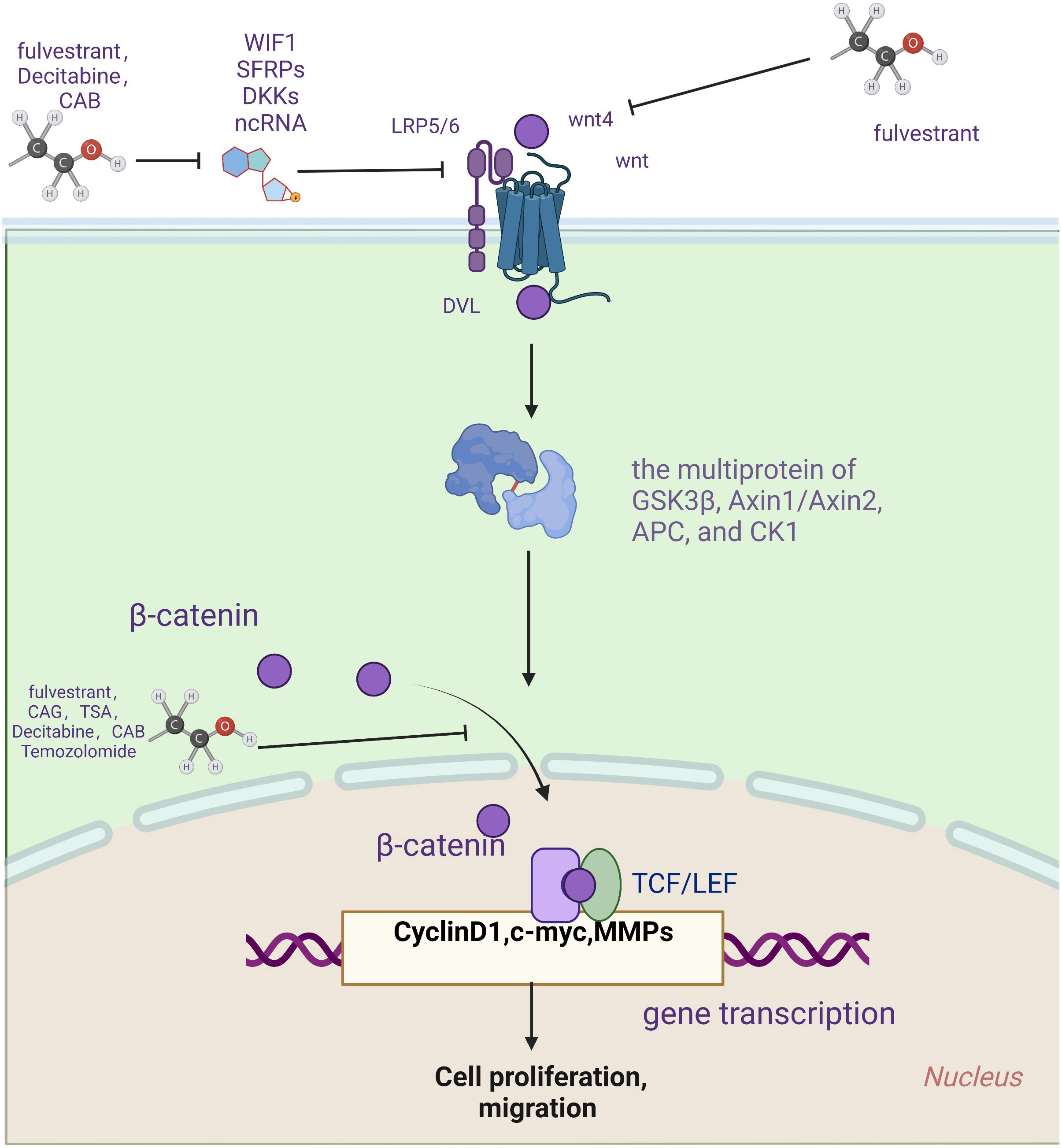
Figure 2 Drugs targeting wnt signaling pathway.
This review provides an overview of the use of Wnt signaling pathway inhibitors in treating PA. It also discusses the current understanding of the Wnt signaling pathway concerning PA and reviews studies on drugs targeting this pathway for treating PA. The Wnt pathway has traditionally been divided into canonical and atypical pathways, which form networks that communicate with other pathways and co-regulate interactions during the development of PA. Wnt signaling pathway Inhibitors include not only the SFRP family, WIF1, and DKK but also ncRNAs. ncRNAs have a dual role as inhibitors to suppress the ontogenesis of PA and as factors that spark Wnt signaling in the cells of PA.
Fulvestrant, a novel estrogen receptor antagonist, has been identified as a promising therapeutic agent for treating PA by targeting the Wnt pathway. It can treat PA by regulating the wnt pathway inhibitor. Drugs used to treat PA include fulvestrant, decitabine, TMZ, CAB, and natural compounds of plant origin, such as CAG and tanshinone IIA. They are closely related to the wnt signaling pathway. However, the specific mechanisms involved in wnt signaling remain unclear and deserve further experimental investigation. TME plays a vital role in the aggression and persistence of PA. Conventional immunotherapy cannot cover all types of PA treatment. Therefore, the combined use of Wnt modulators is expected to improve TME and immunotherapy’s efficacy, which points out a new direction for treating PA.
Future research regarding the wnt signaling pathway in PA should focus on (1) the Mechanism of the wnt signaling pathway in NFPA and prolactinomas. (2) achieving a deeper understanding of crosstalk among these pathways (e.g., Wnt, MAPK, TGF-b, Hippo, VEGF, JAK-STAT, PI3K-Akt, ErbB, TGF-b, and Ras, SOX2, Shh pathways); (3) Inhibitors of wnt signaling in PA should pay more attention to ncRNAs and to gain insight into their roles in wnt signaling in PA. (4) Targeting the wnt signaling pathway improves the TME in refractory PA. (5) Targeting the wnt pathway combined with other therapies covers more types of PAs.
In summary, the wnt signaling pathway plays a significant part in guiding the development and pathogenesis of PA. Abnormal expression of Wnt-signaling inhibitors may be related to the cell multiplication, invasion, and recurrence of PA. Currently, there are few studies on the wnt signaling pathway in PA. Furthermore, studies on these drugs have yet to go into sufficient depth into the mechanism of action. Finding new therapeutic targets and combining them with traditional treatments will reveal novel directions for the individualized treatment of PA, which needs further research.
WW conceived of the topic of the review, performed the literature searches and wrote the manuscript. LM, YZ, ML, WY, and XL contributed to intellectual development and helped to edit the final manuscript. All authors contributed to the article and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Ostrom QT, Gittleman H, Truitt G, Boscia A, Kruchko C, Barnholtz-Sloan JS. CBTRUS statistical report: primary brain and other central nervous system tumors diagnosed in the United States in 2011-2015. Neuro Oncol (2018) 20(suppl_4):iv1–iv86. doi: 10.1093/neuonc/noy131
2. Kessel KA, Diehl CD, Oechsner M, Meyer B, Gempt J, Zimmer C, et al. Patient-Reported Outcome (PRO) as an Addition to Long-Term Results after High-Precision Stereotactic Radiotherapy in Patients with Secreting and Non-Secreting Pituitary Adenomas: A Retrospective Cohort Study up to 17-Years Follow-Up. Cancers (Basel) (2019) 11(12). doi: 10.3390/cancers11121884
3. Aflorei ED, Korbonits M. Epidemiology and etiopathogenesis of pituitary adenomas. J Neurooncol (2014) 117(3):379–94. doi: 10.1007/s11060-013-1354-5
4. Gondim JA, de Almeida JP, de Albuquerque LA, Schops M, Gomes E, Ferraz T. Headache associated with pituitary tumors. J Headache Pain (2009) 10(1):15–20. doi: 10.1007/s10194-008-0084-0
5. Al-Dahmani K, Mohammad S, Imran F, Theriault C, Doucette S, Zwicker D, et al. Sellar masses: an epidemiological study. Can J Neurol Sci (2016) 43(2):291–7. doi: 10.1017/cjn.2015.301
6. Alexander JM, Biller BM, Bikkal H, Zervas NT, Arnold A, Klibanski A. Clinically nonfunctioning pituitary tumors are monoclonal in origin. J Clin Invest (1990) 86(1):336–40. doi: 10.1172/JCI114705
7. Melmed S. Mechanisms for pituitary tumorigenesis: the plastic pituitary. J Clin Invest (2003) 112(11):1603–18. doi: 10.1172/JCI20401
8. Asa SL, Ezzat S. The pathogenesis of pituitary tumors. Annu Rev Pathol (2009) 4:97–126. doi: 10.1146/annurev.pathol.4.110807.092259
9. Liu J, Xiao Q, Xiao J, Niu C, Li Y, Zhang X, et al. Wnt/β-catenin signaling: function, biological mechanisms, and therapeutic opportunities. Signal Transduct Target Ther (2022) 7(1):3. doi: 10.1038/s41392-021-00762-6
10. Cheng X, Xu X, Chen D, Zhao F, Wang W. Therapeutic potential of targeting the Wnt/β-catenin signaling pathway in colorectal cancer. BioMed Pharmacother (2019) 110:473–81. doi: 10.1016/j.biopha.2018.11.082
11. Vuong LT, Mlodzik M. Different strategies by distinct Wnt-signaling pathways in activating a nuclear transcriptional response. Curr Top Dev Biol (2022) 149:59–89. doi: 10.1016/bs.ctdb.2022.02.008
12. Eisenberg LM, Eisenberg CA. Evaluating the role of Wnt signal transduction in promoting the development of the heart. Sci World J (2007) 7:161–76. doi: 10.1100/tsw.2007.71
13. Matthijs Blankesteijn W, Hermans KC. Wnt signaling in atherosclerosis. Eur J Pharmacol (2015) 763(Pt A):122–30. doi: 10.1016/j.ejphar.2015.05.023
14. Gao Y, Chen N, Fu Z, Zhang Q. Progress of wnt signaling pathway in osteoporosis. Biomolecules (2023) 13(3). doi: 10.3390/biom13030483
15. Chen L, Song Z, Huang S, Wang R, Qin W, Guo J, et al. lncRNA DANCR suppresses odontoblast-like differentiation of human dental pulp cells by inhibiting wnt/β-catenin pathway. Cell Tissue Res (2016) 364(2):309–18. doi: 10.1007/s00441-015-2333-2
16. Cai X, Yao Z, Li L, Huang J. Role of DKK4 in tumorigenesis and tumor progression. Int J Biol Sci (2018) 14(6):616–21. doi: 10.7150/ijbs.24329
17. Okamoto M, Udagawa N, Uehara S, Maeda K, Yamashita T, Nakamichi Y, et al. Noncanonical Wnt5a enhances Wnt/beta-catenin signaling during osteoblastogenesis. Sci Rep (2014) 4:4493. doi: 10.1038/srep04493
18. Holmen SL, Salic A, Zylstra CR, Kirschner MW, Williams BO. A novel set of Wnt-Frizzled fusion proteins identifies receptor components that activate beta -catenin-dependent signaling. J Biol Chem (2002) 277(38):34727–35. doi: 10.1074/jbc.M204989200
19. Kühl M. The WNT/calcium pathway: biochemical mediators, tools and future requirements. Front Biosci (2004) 9:967–74. doi: 10.2741/1307
20. Wang HY, Malbon CC. Wnt signaling, Ca2+, and cyclic GMP: visualizing Frizzled functions. Science (2003) 300(5625):1529–30. doi: 10.1126/science.1085259
21. Veeman MT, Axelrod JD, Moon RT. A second canon. Functions and mechanisms of beta-catenin-independent Wnt signaling. Dev Cell (2003) 5(3):367–77. doi: 10.1016/S1534-5807(03)00266-1
22. Rogers S, Scholpp S. Vertebrate Wnt5a - At the crossroads of cellular signaling. Semin Cell Dev Biol (2022) 125:3–10. doi: 10.1016/j.semcdb.2021.10.002
23. Kohn AD, Moon RT. Wnt and calcium signaling: beta-catenin-independent pathways. Cell Calcium (2005) 38(3-4):439–46. doi: 10.1016/j.ceca.2005.06.022
24. Topol L, Jiang X, Choi H, Garrett-Beal L, Carolan PJ, Yang Y. Wnt-5a inhibits the canonical Wnt pathway by promoting GSK-3-independent beta-catenin degradation. J Cell Biol (2003) 162(5):899–908. doi: 10.1083/jcb.200303158
25. Zhu L, Tian Q, Gao H, Wu K, Wang B, Ge G, et al. PROX1 promotes breast cancer invasion and metastasis through WNT/β-catenin pathway via interacting with hnRNPK. Int J Biol Sci (2022) 18(5):2032–46. doi: 10.7150/ijbs.68960
26. Gajos-Michniewicz A, Czyz M. WNT signaling in melanoma. Int J Mol Sci (2020) 21(14). doi: 10.3390/ijms21144852
27. Lyros O, Lamprecht AK, Nie L, Thieme R, Götzel K, Gasparri M, et al. Dickkopf-1 (DKK1) promotes tumor growth via Akt-phosphorylation and independently of Wnt-axis in Barrett's associated esophageal adenocarcinoma. Am J Cancer Res (2019) 9(2):330–46.
28. Sarabia-Sanchez MA, Moreno-Londono AP, Castaneda-Patlan MC, Alvarado-Ortiz E, Martinez-Morales JC, Robles-Flores M. Non-canonical Wnt/Ca2+ signaling is essential to promote self-renewal and proliferation in colon cancer stem cells. Front Oncol (2023) 13:1121787. doi: 10.3389/fonc.2023.1121787
29. Moreno CS, Evans CO, Zhan X, Okor M, Desiderio DM, Oyesiku NM. Novel molecular signaling and classification of human clinically nonfunctional pituitary adenomas identified by gene expression profiling and proteomic analyses. Cancer Res (2005) 65(22):10214–22. doi: 10.1158/0008-5472.CAN-05-0884
30. Giles A, Madec F, Friedrichsen S, Featherstone K, Chambers T, Harper CV, et al. Wnt signaling in estrogen-induced lactotroph proliferation. J Cell Sci (2011) 124(Pt 4):540–7. doi: 10.1242/jcs.078642
31. Li W, Zhang Y, Zhang M, Huang G, Zhang Q. Wnt4 is overexpressed in human pituitary adenomas and is associated with tumor invasion. J Clin Neurosci (2014) 21(1):137–41. doi: 10.1016/j.jocn.2013.04.034
32. Hownga S-L, Wub C-H, Chengb T-S, Sya W-D, Lina P-CK, Wangc C, et al. Differential expression of Wnt genes, b-catenin and E-cadherin in human brain tumors. Cancer Letters (2002) 83(1):95–101. doi: 10.1016/S0304-3835(02)00085-X
33. Zhao C, Zhang M, Liu W, Wang C, Zhang Q, Li W. β-catenin knockdown inhibits pituitary adenoma cell proliferation and invasion via interfering with AKT and gelatinases expression. Int J Oncol (2015) 46(4):1643–50. doi: 10.3892/ijo.2015.2862
34. Formosa R, Gruppetta M, Falzon S, Santillo G, DeGaetano J, Xuereb-Anastasi A, et al. Expression and clinical significance of Wnt players and survivin in pituitary tumours. Endocr Pathol (2012) 23(2):123–31. doi: 10.1007/s12022-012-9197-8
35. Liu C, Wu Y, Yu S, Bai J, Li C, Wu D, et al. Increased beta−catenin and c-myc expression predict aggressive growth of non-functioning pituitary adenomas: An assessment using a tissue microarray-based approach. Mol Med Rep (2017) 15(4):1793–9. doi: 10.3892/mmr.2017.6169
36. Acunzo J, Roche C, Defilles C, Thirion S, Quentien MH, Figarella-Branger D, et al. Inactivation of PITX2 transcription factor induced apoptosis of gonadotroph tumoral cells. Endocrinology (2011) 152(10):3884–92. doi: 10.1210/en.2011-1216
37. Lee WK, Chakraborty PK, Thevenod F. Pituitary homeobox 2 (PITX2) protects renal cancer cell lines against doxorubicin toxicity by transcriptional activation of the multidrug transporter ABCB1. Int J Cancer (2013) 133(3):556–67. doi: 10.1002/ijc.28060
38. Hosseinkhan N, Honardoost M, Emami Z, Cheraghi S, Hashemi-Madani N, Khamseh ME. A systematic review of molecular alterations in invasive non-functioning pituitary adenoma. Endocrine (2022) 77(3):500–9. doi: 10.1007/s12020-022-03105-9
39. Meierjohann S. Crosstalk signaling in targeted melanoma therapy. Cancer Metastasis Rev (2017) 36(1):23–33. doi: 10.1007/s10555-017-9659-z
40. Tang J, Chen L, Wang Z, Huang G, Hu X. SOX2 mediates crosstalk between Sonic Hedgehog and the Wnt/beta-catenin signaling pathway to promote proliferation of pituitary adenoma cells. Oncol Lett (2019) 18(1):81–6. doi: 10.3892/ol.2019.10311
41. Elston MS, Clifton-Bligh RJ. Identification of Wnt family inhibitors: a pituitary tumor directed whole genome approach. Mol Cell Endocrinol (2010) 326(1-2):48–54. doi: 10.1016/j.mce.2010.02.039
42. Kim JH, Liu X, Wang J, Chen X, Zhang H, Kim SH, et al. Wnt signaling in bone formation and its therapeutic potential for bone diseases. Ther Adv Musculoskelet Dis (2013) 5(1):13–31. doi: 10.1177/1759720X12466608
43. Yu B, Kiechl S, Qi D, Wang X, Song Y, Weger S, et al. A cytokine-like protein dickkopf-related protein 3 is atheroprotective. Circulation (2017) 136(11):1022–36. doi: 10.1161/CIRCULATIONAHA.117.027690
44. Bao J, Zheng JJ, Wu D. The structural basis of DKK-mediated inhibition of Wnt/LRP signaling. Sci Signal (2012) 5(224):pe22. doi: 10.1126/scisignal.2003028
45. Melkonyan HS, Chang WC, Shapiro JP, Mahadevappa M, Fitzpatrick PA, Kiefer MC, et al. SARPs: a family of secreted apoptosis-related proteins. Proc Natl Acad Sci USA (1997) 94(25):13636–41. doi: 10.1073/pnas.94.25.13636
46. Rattner A, Hsieh JC, Smallwood PM, Gilbert DJ, Copeland NG, Jenkins NA, et al. A family of secreted proteins contains homology to the cysteine-rich ligand-binding domain of frizzled receptors. Proc Natl Acad Sci USA (1997) 94(7):2859–63. doi: 10.1073/pnas.94.7.2859
47. Wu Y, Bai J, Li Z, Wang F, Cao L, Liu C, et al. Low expression of secreted frizzled-related protein 4 in aggressive pituitary adenoma. Pituitary (2015) 18(3):335–42. doi: 10.1007/s11102-014-0579-4
48. Wu Y, Bai J, Hong L, Liu C, Yu S, Yu G, et al. Low expression of secreted frizzled-related protein 2 and nuclear accumulation of beta-catenin in aggressive nonfunctioning pituitary adenoma. Oncol Lett (2016) 12(1):199–206. doi: 10.3892/ol.2016.4560
49. Ren J, Jian F, Jiang H, Sun Y, Pan S, Gu C, et al. Decreased expression of SFRP2 promotes development of the pituitary corticotroph adenoma by upregulating Wnt signaling. Int J Oncol (2018) 52(6):1934–46. doi: 10.3892/ijo.2018.4355
50. Kerekes K, Trexler M, Bányai L, Patthy L. Wnt inhibitory factor 1 binds to and inhibits the activity of sonic hedgehog. Cells (2021) 10(12). doi: 10.3390/cells10123496
51. Urakami S, Shiina H, Enokida H, Kawakami T, Tokizane T, Ogishima T, et al. Epigenetic inactivation of Wnt inhibitory factor-1 plays an important role in bladder cancer through aberrant canonical Wnt/beta-catenin signaling pathway. Clin Cancer Res (2006) 12(2):383–91. doi: 10.1158/1078-0432.CCR-05-1344
52. Nakaya N, Lee HS, Takada Y, Tzchori I, Tomarev SI. Zebrafish olfactomedin 1 regulates retinal axon elongation in vivo and is a modulator of Wnt signaling pathway. J Neurosci (2008) 28(31):7900–10. doi: 10.1523/JNEUROSCI.0617-08.2008
53. Song W, Qian L, Jing G, Jie F, Xiaosong S, Chunhui L, et al. Aberrant expression of the sFRP and WIF1 genes in invasive non-functioning pituitary adenomas. Mol Cell Endocrinol (2018) 474:168–75. doi: 10.1016/j.mce.2018.03.005
54. Zhu H, Yao X, Wu L, Li C, Bai J, Gao H, et al. Association of TGF-beta1 and WIF1 expression with 36 paired primary/recurrent nonfunctioning pituitary adenomas: A high-throughput tissue microarrays immunohistochemical study. World Neurosurg (2018) 119:e23–31. doi: 10.1016/j.wneu.2018.06.154
55. Li J, Dong W, Li Z, Wang H, Gao H, Zhang Y. Impact of SLC20A1 on the Wnt/beta−catenin signaling pathway in somatotroph adenomas. Mol Med Rep (2019) 20(4):3276–84. doi: 10.3892/mmr.2019.10555
56. Cheng S, Wu J, Li C, Li Y, Liu C, Li G, et al. Predicting the regrowth of clinically non-functioning pituitary adenoma with a statistical model. J Transl Med (2019) 17(1):164. doi: 10.1186/s12967-019-1915-2
57. Lei C, Jing G, Jichao W, Xiaohui L, Fang Q, Hua G, et al. MiR-137's tumor suppression on prolactinomas by targeting MITF and modulating wnt signaling pathway. J Clin Endocrinol Metab (2019) 104(12):6391–402. doi: 10.1210/jc.2018-02544
58. Lee E-J, Jo M, Rho SB, Park K, Yoo Y-N, Park J, et al. Dkk3, downregulated in cervical cancer, functions as a negative regulator of β-catenin. Int J Cancer (2009) 124(2):287–97. doi: 10.1002/ijc.23913
59. Chua K, Lee VK, Chan C, Yew A, Yeo E, Virshup DM. Hematopoietic wnts modulate endochondral ossification during fracture healing. Front Endocrinol (Lausanne) (2021) 12:667480. doi: 10.3389/fendo.2021.667480
60. Belaya Z, Grebennikova T, Melnichenko G, Nikitin A, Solodovnikov A, Brovkina O, et al. Effects of active acromegaly on bone mRNA and microRNA expression patterns. Eur J Endocrinol (2018) 178(4):353–64. doi: 10.1530/EJE-17-0772
61. Belaya ZE, Grebennikova TA, Melnichenko GA, Nikitin AG, Solodovnikov AG, Brovkina OI, et al. Effects of endogenous hypercortisolism on bone mRNA and microRNA expression in humans. Osteoporos Int (2018) 29(1):211–21. doi: 10.1007/s00198-017-4241-7
62. Wang C, Tan C, Wen Y, Zhang D, Li G, Chang L, et al. FOXP1-induced lncRNA CLRN1-AS1 acts as a tumor suppressor in pituitary prolactinoma by repressing the autophagy via inactivating Wnt/β-catenin signaling pathway. Cell Death Dis (2019) 10(7). doi: 10.1038/s41419-019-1694-y
63. Shen DW, Li YL, Hou YJ, Xu ZD, Li YZ, Chang JY. MicroRNA-543 promotes cell invasion and impedes apoptosis in pituitary adenoma via activating the Wnt/beta-catenin pathway by negative regulation of Smad7. Biosci Biotechnol Biochem (2019) 83(6):1035–44. doi: 10.1080/09168451.2019.1591260
64. Zhang Y, Tan Y, Wang H, Xu M, Xu L. Long non-coding RNA plasmacytoma variant translocation 1 (PVT1) enhances proliferation, migration, and epithelial-mesenchymal transition (EMT) of pituitary adenoma cells by activating β-catenin, c-myc, and cyclin D1 expression. Med Sci Monit (2019) 25:7652–9. doi: 10.12659/MSM.917110
65. Huang G, Liang M, Liu H, Huang J, Li P, Wang C, et al. CircRNA hsa_circRNA_104348 promotes hepatocellular carcinoma progression through modulating miR-187-3p/RTKN2 axis and activating Wnt/beta-catenin pathway. Cell Death Dis (2020) 11(12):1065. doi: 10.1038/s41419-020-03276-1
66. Du Q, Zhang W, Feng Q, Hao B, Cheng C, Cheng Y, et al. Comprehensive circular RNA profiling reveals that hsa_circ_0001368 is involved in growth hormone-secreting pituitary adenoma development. Brain Res Bull (2020) 161:65–77. doi: 10.1016/j.brainresbull.2020.04.018
67. Vetrivel S, Zhang R, Engel M, Osswald A, Watts D, Chen A, et al. Characterization of adrenal miRNA-based dysregulations in cushing's syndrome. Int J Mol Sci (2022) 23(14). doi: 10.3390/ijms23147676
68. Gheorghiu ML, Negreanu F, Fleseriu M. Updates in the medical treatment of pituitary adenomas. Hormone Metab Res (2019) 52(01):8–24. doi: 10.1055/a-1066-4592
69. Bai J, Wang Y, Li C, Zhang Y. Effects of fulvestrant on biological activity and Wnt expression in rat GH3 cells. Neural Regener Res (2012) 7(4):283–9. doi: 10.3969/j.issn.1673-5374.2012.04.008
70. Bai J, Gui S, Zhang Y. Suppression of MMQ cells by fulvestrant: possible mechanism of action and potential application for bromocriptine-resistant prolactinomas. J Clin Neurosci (2013) 20(5):721–5. doi: 10.1016/j.jocn.2012.07.008
71. Cao L, Gao H, Gui S, Bai G, Lu R, Wang F, et al. Effects of the estrogen receptor antagonist fulvestrant on F344 rat prolactinoma models. J Neurooncol (2014) 116(3):523–31. doi: 10.1007/s11060-013-1351-8
72. Cao L, Gao H, Li P, Gui S, Zhang Y. The Wnt/beta-catenin signaling pathway is involved in the antitumor effect of fulvestrant on rat prolactinoma MMQ cells. Tumour Biol (2014) 35(6):5121–7. doi: 10.1007/s13277-013-1571-8
73. Jones SE. Fulvestrant: an estrogen receptor antagonist that downregulates the estrogen receptor. Semin Oncol (2003) 30(5 Suppl 16):14–20. doi: 10.1053/j.seminoncol.2003.08.003
74. Dziamski KM, Kalakonda A, Kohlitz P. A rare case of fulvestrant-associated jaundice and hepatotoxicity. Am J Ther (2019) 26(1):e122–e3. doi: 10.1097/MJT.0000000000000532
75. Garcia-Manero G, Griffiths EA, Steensma DP, Roboz GJ, Wells R, McCloskey J, et al. Oral cedazuridine/decitabine for MDS and CMML: a phase 2 pharmacokinetic/pharmacodynamic randomized crossover study. Blood (2020) 136(6):674–83. doi: 10.1182/blood.2019004143
76. DiNardo CD, Maiti A, Rausch CR, Pemmaraju N, Naqvi K, Daver NG, et al. 10-day decitabine with venetoclax for newly diagnosed intensive chemotherapy ineligible, and relapsed or refractory acute myeloid leukaemia: a single-centre, phase 2 trial. Lancet Haematol (2020) 7(10):e724–e36. doi: 10.1016/S2352-3026(20)30210-6
77. Lai XB, Nie YQ, Huang HL, Li YF, Cao CY, Yang H, et al. NIMA-related kinase 2 regulates hepatocellular carcinoma cell growth and proliferation. Oncol Lett (2017) 13(3):1587–94. doi: 10.3892/ol.2017.5618
78. Wang J, Cheng P, Pavlyukov MS, Yu H, Zhang Z, Kim SH, et al. Targeting NEK2 attenuates glioblastoma growth and radioresistance by destabilizing histone methyltransferase EZH2. J Clin Invest (2020) 130(11):6187. doi: 10.1172/JCI144618
79. Jian F, Sun Y, Sun Q, Zhang B, Bian L. NEK2 regulates cellular proliferation and cabergoline sensitivity in pituitary adenomas. J Cancer (2021) 12(7):2083–91. doi: 10.7150/jca.52937
80. Gadelha MR, Wildemberg LE, Shimon I. Pituitary acting drugs: cabergoline and pasireotide. Pituitary (2022) 25(5):722–5. doi: 10.1007/s11102-022-01238-8
81. Demarchi G, Valla S, Perrone S, Chimento A, Bonadeo N, Vitale DL, et al. beta-Catenin is reduced in membranes of human prolactinoma cells and it is inhibited by temozolomide in prolactin secreting tumor models. Tumour Biol (2022) 44(1):85–105. doi: 10.3233/TUB-211500
82. Demarchi G, Perrone S, Esper Romero G, De Bonis C, Casasco JP, Sevlever G, et al. Case Report: Progression of a Silent Corticotroph Tumor to an Aggressive Secreting Corticotroph Tumor, Treated by Temozolomide. Changes in the Clinic, the Pathology, and the beta-Catenin and alpha-SMA Expression. Front Endocrinol (Lausanne) (2022) 13:870172. doi: 10.3389/fendo.2022.870172
83. Park AK, Waheed A, Forst DA, Al-Samkari H. Characterization and prognosis of temozolomide-induced aplastic anemia in patients with central nervous system Malignancies. Neuro Oncol (2022) 24(6):964–73. doi: 10.1093/neuonc/noab240
84. Scaringi C, De Sanctis V, Minniti G, Enrici RM. Temozolomide-related hematologic toxicity. Onkologie (2013) 36(7-8):444–9. doi: 10.1159/000353752
85. Climans SA, Mason WP, Grunfeld E, Chan K. Clinical features of glioma patients who develop pneumocystis pneumonia with temozolomide chemoradiotherapy. J Neurooncol (2022) 159(3):665–74. doi: 10.1007/s11060-022-04109-1
86. Mehta H, Gendle CS, Kumaran MS, Vinay K. Temozolomide-induced drug rash with eosinophilia and systemic symptoms syndrome. Indian J Dermatol Venereol Leprol (2023) 89(1):160. doi: 10.25259/IJDVL_754_2021
87. Chen Z, Feng H, Peng C, Zhang Z, Yuan Q, Gao H, et al. Renoprotective effects of tanshinone IIA: A literature review. Molecules (2023) 28(4). doi: 10.3390/molecules28041990
88. Li ZY, Li QZ, Ma GX, Chen L, Zhang C, Chen BD, et al. Cassane-type diterpenes from Caesalpinia minax induce apoptosis in pituitary adenoma: structure-activity relationship, ER stress, and Wnt/beta-catenin pathways. J Asian Nat Prod Res (2017) 19(5):423–35. doi: 10.1080/10286020.2016.1217520
89. Li ZY, Huang GD, Chen L, Zhang C, Chen BD, Li QZ, et al. Tanshinone IIA induces apoptosis via inhibition of Wnt/beta−catenin/MGMT signaling in AtT−20 cells. Mol Med Rep (2017) 16(5):5908–14. doi: 10.3892/mmr.2017.7325
90. Lei Y, Li X, Qin D, Zhang Y, Wang Y. gC1qR: A new target for cancer immunotherapy. Front Immunol (2023) 14:1095943. doi: 10.3389/fimmu.2023.1095943
91. Ben-Shlomo A. Exploring the role of the tumor microenvironment in refractory pituitary tumor pathogenesis. Pituitary (2023). doi: 10.1007/s11102-023-01301-y
92. Ilie MD, Vasiljevic A, Bertolino P, Raverot G. Biological and therapeutic implications of the tumor microenvironment in pituitary adenomas. Endocr Rev (2023) 44(2):297–311. doi: 10.1210/endrev/bnac024
Keywords: PA, Wnt signaling pathway, Wnt pathway inhibitors, targeted drugs, non-coding RNA, TME
Citation: Wang W, Ma L, Zhao Y, Liu M, Ye W and Li X (2023) Research progress on the role of the Wnt signaling pathway in pituitary adenoma. Front. Endocrinol. 14:1216817. doi: 10.3389/fendo.2023.1216817
Received: 04 May 2023; Accepted: 28 August 2023;
Published: 14 September 2023.
Edited by:
Wang Haijun, The First Affiliated Hospital of Sun Yat-sen University, ChinaReviewed by:
Liuguan Bian, Shanghai Jiao Tong University, ChinaCopyright © 2023 Wang, Ma, Zhao, Liu, Ye and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xianfeng Li, bGl4aWFuZmVuZzIwMDBAMTYzLmNvbQ==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.