 Martin Bulla1,2*
Martin Bulla1,2* Clemens Küpper3
Clemens Küpper3 David B. Lank4
David B. Lank4 Jana Albrechtová5
Jana Albrechtová5 Jasmine L. Loveland3,6Katrin Martin1Kim Teltscher1Margherita Cragnolini1Michael Lierz7
Jasmine L. Loveland3,6Katrin Martin1Kim Teltscher1Margherita Cragnolini1Michael Lierz7 Tomáš Albrecht5,8
Tomáš Albrecht5,8 Wolfgang Forstmeier1Bart Kempenaers1*
Wolfgang Forstmeier1Bart Kempenaers1*- 1Department of Ornithology, Max Planck Institute for Biological Intelligence, Seewiesen, Germany
- 2Department of Ecology, Faculty of Environmental Sciences, Czech University of Life Sciences Prague, Prague, Czechia
- 3Research Group for Behavioural Genetics and Evolutionary Ecology, Max Planck Institute for Biological Intelligence, Seewiesen, Germany
- 4Department of Biological Sciences, Simon Fraser University, Burnaby, BC, Canada
- 5Institute of Vertebrate Biology, Czech Academy of Sciences, Brno, Czechia
- 6Department of Cognitive and Behavioral Biology, University of Vienna, Vienna, Austria
- 7Clinic for Birds, Reptiles, Amphibians and Fish, Justus Liebig University Giessen, Giessen, Germany
- 8Department of Zoology, Faculty of Science, Charles University, Prague, Czechia
The ruff sandpiper (Calidris pugnax) is a lekking shorebird with three male morphs that differ remarkably in behavior, ornaments, size, and endocrinology. The morphs are determined by an autosomal inversion. Aggressive Independents evolved first, female-mimicking Faeders ~4 mil year ago when a short segment of a chromosome reversed in orientation, and semi-cooperative Satellites ~70,000 years ago through a recombination of the Independent and Faeder inversion-segment genotypes. Although the genetic differences between the morphs affect numerous phenotypic traits, it is unknown whether they also affect sperm traits. Here, we use a captive-bred population of ruffs to compare ruff sperm to that of other birds and compare sperm swimming speed and morphology among the morphs. Ruff sperm resembled those of passerines, but moved differently. Faeder sperm moved the slowest and had the longest midpiece. Independents’ sperm were neither the fastest nor the least variable, but had the shortest tail and midpiece. Although the midpiece contains the energy-producing mitochondria, its length was not associated with sperm swimming speed. Instead, two of three velocity metrics weakly positively correlated with head length (absolute and relative). We conclude that there is an indication of quantitative differences in sperm between morphs, but no clear sperm polymorphism.
Introduction
In many animal species, males use different behavioral strategies to obtain fertilizations (Dougherty et al., 2022; Kustra and Alonzo, 2020; Mank, 2023). For example, dominant males may display, while other males “steal” copulations (Gross, 1996) by pretending to be a female or by positioning themselves in between the dominant male and the female that is about to copulate (Oliverira et al., 2008; Shuster and Wade, 1991). Consequently, males that use alternative mating tactics often experience different levels of sperm competition, defined as competition between the sperm from different males to fertilize one or more eggs (Parker, 1970). For example, if dominant males partially monopolize or are preferred by females, it is likely that males with alternative mating tactics experience sperm competition more often than dominant males (Dougherty et al., 2022; Kustra and Alonzo, 2020). Comparative studies have shown that species with higher levels of sperm competition typically produce sperm that swim faster, are longer (including longer sperm components), and less variable in length as a result of intense postcopulatory selection on optimal sperm length (Laskemoen et al., 2013; Lifjeld et al., 2010; Lipshutz et al., 2022; Lüpold et al., 2020; Simmons and Fitzpatrick, 2012). Within species, sperm fertilization ability has been linked to sperm viability (Gage et al., 2004; Garcia-Gonzalez and Simmons, 2005), swimming speed (Birkhead et al., 1999; Gage et al., 2004), and length (Bennison et al., 2014; Garcia-Gonzalez and Simmons, 2007; Lüpold et al., 2012). Thus, within species, selection may also have favored different sperm traits in males that consistently differ in mating behavior or levels of sperm competition. Despite general expectation that males exhibiting alternative mating tactics should differ in the investment into sperm production (potentially through differences in sperm morphology or performance), recent meta-analysis studies show little empirical evidence that males using alternative mating tactics differ substantially in investment into sperm, which prohibits broad generalizations (Dougherty et al., 2022; Kustra and Alonzo, 2020).
In the ruff (Calidris pugnax), a lekking shorebird, males occur in three distinct morphs with striking differences in reproductive behavior, ornaments, body size, and endocrinology (Hogan-Warburg, 1966; Höglund and Lundberg, 1989; Jukema and Piersma, 2006; Küpper et al., 2016; Loveland et al., 2021a; van Rhijn, 1991; Widemo, 1998). (1) Aggressive Independents make up 80-90% of all males and show a spectacular diversity of predominantly dark ornamental plumages. The Independents hold small display courts on a lek. Within Independents, dominant males obtain more matings (Vervoort and Kempenaers, 2019; Widemo, 1997). (2) Submissive and slightly smaller Satellites make up 10-20% of males, show predominantly white ornamental plumage, and co-display with court-holders on leks (Hogan-Warburg, 1966; Höglund and Lundberg, 1989; van Rhijn, 1991; Widemo, 1998). Although Satellites may “steal” some copulations, Independents may benefit from having Satellites on the lek because their presence helps attract females to the display court of that male (Hugie and Lank, 1997; Tolliver et al., 2023). (3) Faeder males are rare (~1%), mimic females in appearance (lack ornamental plumage and have smaller body size), and are thought to sneak copulations when a female solicits an ornamented displaying male (Jukema and Piersma, 2006). The morphs also differ markedly in their reproductive endocrinology. During the lekking season, Independent males have higher levels of circulating testosterone, whereas Satellites and Faeders have higher levels of androstenedione, a testosterone precursor (Küpper et al., 2016; Loveland et al., 2021a). Moreover, despite striking differences in body size, males from all three morphs have similar absolute testes sizes (Figure 1) and hence presumably can produce similar numbers of sperm.
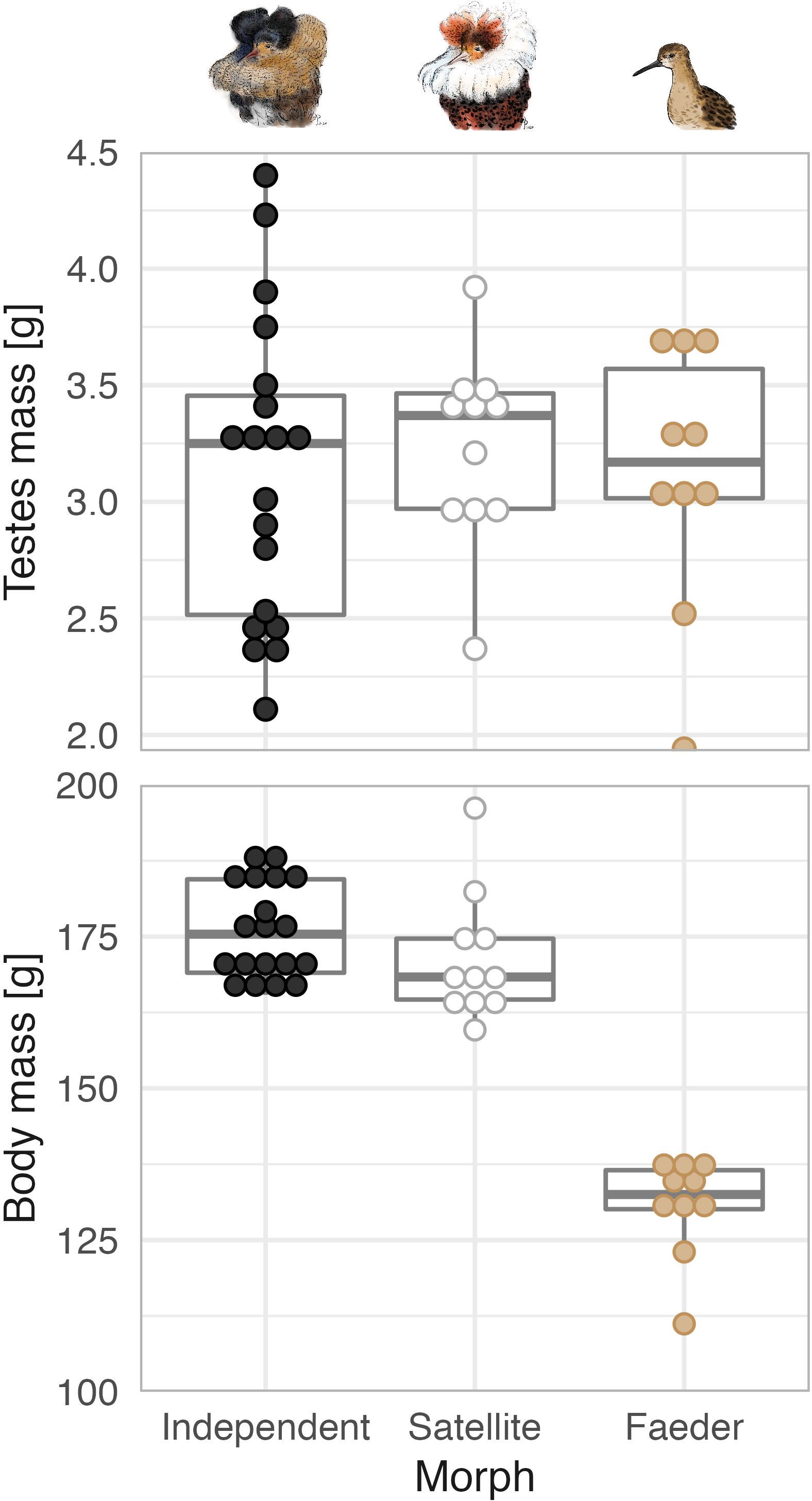
Figure 1. Testes mass and body mass in relation to morph in the ruff. Each dot represents an individual, dot color highlights the morph (black: Independent, white: Satellite, beige: Faeder). Boxplots depict the median (horizontal line inside the box), the 25th and 75th percentiles (box) and the 25th and 75th percentiles ±1.5 times the interquartile range or the minimum/maximum value, whichever is smaller (bars). Data from (Loveland et al., 2021b) and from an additional 5 males collected under the same protocol in 2022 (3 Independents, 1 Satellite and 1 Faeder; our unpublished data). Dots stacked using the ‘geom_dotplot’ ggplot2 R-function (Wickham, 2016). Ruff morph illustrations by Yifan Pei under Creative Commons Attribution (CC BY 4.0).
The phenotypic differences between the morphs are entirely genetically determined by a 4.5 Mb autosomal inversion region, which corresponds to 0.3% of the haploid genome (Küpper et al., 2016; Lamichhaney et al., 2016). Faeders and Satellites carry unique inversion haplotypes. The Faeder haplotype arose first (about 4 million years ago), whereas the Satellite haplotype originated about 70,000 years ago from a rare recombination event between the Independent and Faeder genotypes (Hill et al., 2023; Lamichhaney et al., 2016). Importantly, because one inversion breakpoint is homozygous lethal, the inverted haplotypes cannot recombine and thus only occur in hemizygous state (i.e. always combined with one Independent haplotype). Because the inversion is autosomal, females also occur as 3 morphs. Females show no strikingly different behavioral or plumage phenotypes, but ‘Faeder females’ are distinctly smaller (Lank et al., 2013).
Male ruffs are polymorphic in numerous phenotypic traits, but it is undocumented whether the morphs differ in sperm characteristics. Indeed, morph-specific sperm traits are possible for the three following reasons. (i) Two genes regulating spermatogenesis reside within the short (~125 loci) autosomal inversion that determines the Satellite and Faeder phenotypes (Küpper et al., 2016). The inversion contains the gene GAS8, which has been linked to sperm motility in mice, and the gene SPATA2L, a paralog of SPATA2, which is involved in spermatogenesis (Graziotto et al., 1999; Küpper et al., 2016; Onisto et al., 2000; Yeh et al., 2002). Moreover, expression of SPATA2L in testes of both Satellites and Faeders is biased toward the inversion allele (Loveland et al., 2021b). (ii) The inversion drastically affects the circulating levels of testosterone and androstenedione (Küpper et al., 2016; Loveland et al., 2021a), which then may cause various downstream effects such as changes in expression of genes located outside of the inversion (Maney and Küpper, 2022). For example, STAR, a gene responsible for providing the cholesterol substrate required for sex hormone synthesis, is located outside of the inversion, but is overexpressed in the testes of Satellites and Faeders (Loveland et al., 2021a). More generally, some genes or regulatory elements within the inversion region that have strong pleiotropic effects may also affect spermatogenesis. (iii) The presence of non-synonymous substitutions in the inversion alleles suggest that genes within the inversion have evolved under positive selection (Hill et al., 2023; Küpper et al., 2016; Lamichhaney et al., 2016). Sperm competition between morphs might have been one of the selective pressures.
Three observations indicate that sperm competition is intense in ruff. (1) Ruffs have the longest sperm measured among shorebirds (Johnson and Briskie, 1999), and sperm length correlates strongly with levels of multiple paternity across bird species (Briskie et al., 1997). (2) A quarter of females (or more) copulate with multiple males, and (3) more than half of all clutches are sired by multiple males (Lank et al., 2002; Thuman, 2003). Selection for morph differences, however, would require different morph-specific sperm production costs or investment into sperm. Female ruffs visit one or more leks, and copulate with one or more males, sometimes of different morphs, producing a clutch of 3 or 4 eggs two to five days later (Lank et al., 2002), using stored sperm, as in other birds (Briskie and Montgomerie, 1993). When a female visits a lek and solicits a copulation, the dominant Independent male is more likely to copulate more than once with that female than is a Satellite male who often will get to copulate just once (Lank et al., 2002). The copulation behavior of Faeders remains unquantified, but given its distinct mating tactics, its copulation behavior likely also differs from the other two morphs. In principle, one may argue that the different mating behaviors of the three morphs will translate into different mating opportunities, and may hence select for different optimal sperm traits in the three morphs. Although such adaptation is plausible, its evolution may in part be hampered by the small size of the inversion and the smaller population sizes of the inverted morphs (Giraldo-Deck et al., 2022). Importantly, the genetic background outside of the inversion has been selected to function best in the context of the most abundant morph, i.e., the Independents (Giraldo-Deck et al., 2022), thereby making it more difficult for the rare morphs to evolve special adaptations.
Here, we use sperm sampled from a captive breeding population of ruffs to examine variation in sperm traits. Specifically, our study has three objectives:
1. We assess differences in sperm swimming speed and sperm (component) length between the three morphs.
2. We explore whether variation in ruff sperm morphology predicts variation in sperm swimming speed because in birds it is unclear why species with higher levels of sperm competition have longer sperm. Although a positive within species relationship between overall sperm length, midpiece or flagellum length and swimming speed can be predicted, previous studies, including those in birds, have shown inconsistent results (reviewed in Bennison et al., 2016; Cramer et al., 2021; Fitzpatrick and Lüpold, 2014; Kim et al., 2017; Knief et al., 2017; Míčková et al., 2023; Støstad et al., 2018).
3. We describe the motility and shape of ruff sperm in comparison with that of other bird species. Sperm motility has not yet been described in scolopacids and only rarely in other species of shorebirds, and sperm shape has only been sketchily described in a few species in this group (Johnson and Briskie, 1999; Retzius, 1909). Furthermore, scolopacid sperm superficially resemble the sperm of passerines, but differ from those of all other (so far evaluated) avian lineages (Retzius, 1909).
Materials and methods
We collected and analyzed the data based on a priori designed protocols (for any deviations see Supplementary Methods S0; Bulla et al., 2024).
Captive population
A population of ~300 ruffs is housed at the Max Planck Institute for Biological Intelligence, Seewiesen, Germany (for details see Supplementary Methods S1). The population was founded in 2018 with 23 ruffs obtained from Dutch breeders, 5 ruffs from German breeders and 194 ruffs from Simon Fraser University, Vancouver, Canada. The latter were individuals from a captive-bred population founded with 110 ruffs hatched from wild eggs collected in Finland in 1985, 1989 and 1990, plus two Faeder males brought from the Netherlands in 2006 (Lank et al., 2013, 1995). Each individual of the population has been genotyped for its morph using a set of six single-nucleotide polymorphism markers located in the inversion region (Giraldo-Deck et al., 2020).
Sperm sampling
In May and June 2021, we collected sperm by abdominal massage (see a detailed protocol of sperm collection – including video - and sample preparation) or by electro‐stimulation (Lierz et al., 2013; Bulla et al., 2024). The length (15 mm) and diameter (4 mm) of the electro-stimulation probe, as well as the electric current and the number of electric impulses was adapted to the sampled individuals. The probe was inserted into the urodeum of the cloaca to stimulate the ampullae ductus deferens using three 1 s electric current impulses. The voltage was increased gradually from 0.09 V to a maximum of 1.0 V until contractions of the cloaca and the muscles of the tail were observed; each impulse was followed by a 2-3 s break. In June, we only sampled sperm using abdominal massage. The morphs were sampled in a haphazard order to avoid a potential confounding effect of sampling order. In both months, we attempted to obtain sperm by abdominal massage from all males. We obtained at least one ejaculate sample from 92 males (59 Independents, 25 Satellites and 8 Faeders).
Ejaculates (~0.5–3μl) were pipetted from the cloaca and immediately diluted and gently mixed in 50μl of preheated (40°C) Dulbecco’s Modified Eagle’s Medium (Advanced D-MEM, Invitrogen™). To record sperm swimming speed, we then pipetted an aliquot of 2.5μl onto a standard 20μm two-chamber count slide (Leja, The Netherlands) placed on a thermal plate (Tokai Hit, Tokai Hit Co., Ltd.) kept at 40°C. When sperm densities on the slide were too high, we took a new aliquot and further diluted the sample. We then recorded sperm movements. By limiting the time between ejaculate collection and recording, we avoided that changes in sperm swimming speed over time (Cramer et al., 2016) would influence the results. For morphology measurements, we pipetted an aliquot of 20μl into 50μl of a phosphate buffered saline solution (PBS; Sigma P-4417) containing 1% formalin.
Measurement of sperm swimming speed
For each sperm sample we recorded sperm swimming speed at 25 frames per second for approximately 45s in eight different fields of the Leja slide under a 100x magnification, using phase contrast and a digital camera (UI-1540-C, Olympus) mounted on a microscope (CX41, Olympus) fitted with a thermal plate (Tokai Hit, Tokai Hit Co., Ltd.) kept at a constant temperature of 40°C. We confirmed that this temperature was appropriate based on cloacal temperature measurements of five female ruffs. For each female, we inserted a high-resolution temperature-probe connected to a MSR145 data logger (MSR Electronics GmbH, https://www.msr.ch/en/) into the cloaca, and logged the temperature every 5s for 3 min, i.e., until an asymptotic value was reached. Cloacal temperatures ranged between 40.6°C and 42.3°C, and are similar to the body temperatures measured in shorebirds (Charadriiformes; range: 40.4-41.8°C, N = 10 species) and in passerines (39.2-43.5°C, N = 16 species; (McNab, 1966).
A few weeks later, a single person (JA) analyzed each recorded field using the CEROS computer-assisted sperm analysis system (Hamilton Thorne Inc.). JA visually inspected the tracked objects, excluded non-sperm objects and static spermatozoa from the analysis (Cramer et al., 2016; Laskemoen et al., 2010; Opatová et al., 2016), and noted the quality of the recording (e.g., presence of feces). We recorded a median of 192 sperm cells per sample (mean = 204, range: 5 – 562; N = 134 recordings from a total of 92 males, 46 recorded in May and 88 in June; Independent: mean = 202, N = 83; Satellite: mean = 205, N = 37; Faeder: mean = 218, N = 14), which represented a median of 78% of motile sperm in a sample (mean = 69%, range: 12 – 92%; Independent: mean = 70%; Satellite: mean = 69%; Faeder: mean = 61%). For each sample the software estimated the mean curvilinear, straight-line, and average-path velocity (for details see Supplementary Methods S2, Figures S1, S2; for within-male seasonal repeatability see Supplementary Figures S3, S4, Table S1).
Measurements of sperm morphology
Within 5 days of sample collection, we pipetted (i) 20μl of the PBS-1% formalin fixed sample into 50μl of 5% formalin for long-term storage and (ii) smeared 10μl onto a microscope slide, and let it dry at room temperature or on a heating block set at 37°C. The next day, each slide was rinsed gently with deionized water to remove salt crystals and dried at room temperature.
To each dried slide we added a drop of Bisbenzimide (Hoechst 33342; Molecular Probes) to stain the sperm nucleus, and a drop of MitoTracker™ Green FM Dye (Invitrogen™ M7514) to stain the sperm midpiece (mitochondria; Figure 2). Within 48h of staining, we inspected the microscope slides under 200x magnification with a Zeiss Axio Imager.M2 light microscope fitted with a Zeiss Axiocam 512 color camera (12 megapixel; 4250 × 2838, pixel size of 3.1μm × 3.1μm) and DAPI 465nm and green 519nm filters using Zeiss ZEN blue 3.1 imaging software. For each male we used a single sample (for 30 males from May, for 62 males from June) and photographed at least 10 intact, normal-looking spermatozoa under 400x magnification (objective size 40, ocular size 10). We then selected the 10 best, single-sperm images per male for measurements. We then randomized and renamed all pictures, such that we measured sperm blind to the morph and the identity of the individuals.

Figure 2. Stained ruff sperm. From left to right, faint greenish tail, green midpiece (mitochondria) stained by MitoTracker™ Green FM Dye, turquois nucleus stained by Hoechst 33342 and greenish acrosome.
For each sperm, we measured the length of the acrosome, the nucleus, the midpiece and the tail to the nearest 0.1μm using the open software Sperm Sizer 1.6.6 (McDiarmid et al., 2021; https://github.com/wyrli/sperm-sizer; see our Sperm Sizer protocol (Bulla et al., 2024). To assess within- and between-observer repeatability of Sperm Sizer measurements, we selected 40 single-sperm pictures and remeasured them, once by the same, once by a different person. The measurements were highly repeatable, both within- and between persons (mean = 97%; Supplementary Table S2). Nevertheless, all measurements used in the analyses were taken by the same person (KT). Each measured sperm and sperm part were numbered and referenced in the database and hence can be linked to the original picture as well as to the pictures of each part generated by Sperm Sizer, which contain the lines and measurement values (see Data in Bulla et al., 2024). This ensures transparency and allows re-measurement of the same sperm by the same or a different person.
We calculated (i) total sperm length as the sum of all parts, (ii) head length as the sum of acrosome and nucleus length, (iii) flagellum length as the sum of midpiece and tail length, (iv) relative midpiece length as midpiece length divided by total length and (v) relative flagellum length as flagellum length divided by total length. We then computed coefficients of variation within each male for each absolute trait as the standard deviation divided by the mean. Note that estimating coefficients of variation from only five sperm per male gave similar values and previous studies from different species also used ten sperm per male to estimate the coefficient of variation in sperm traits (Kleven et al., 2008; Lifjeld et al., 2010).
In the Supplement, we report correlations between sperm components (Supplementary Methods S3, Figure S5), and estimates of within-male repeatability (Supplementary Figure S3, Table S1).
Statistical analyses
All analyses were performed in R (R-Core-Team, 2022) using the ‘lm’ function to fit linear models and the ‘lmer’ function of the ‘lme4’ R-package to fit linear mixed-effects models (Gelman and Hill, 2007; Gelman and Su, 2021). We used the ‘sim’ function from the ‘arm’ R-package and a flat prior distribution (Gelman and Hill, 2007; Gelman and Su, 2021) to create a sample of 5,000 simulated values for each model parameter (i.e. posterior distribution). We report effect sizes and model predictions as medians, and the uncertainty of the estimates and predictions as Bayesian 95% credible intervals represented by the 2.5 and 97.5 percentiles (95% CI) from the posterior distribution of 5,000 simulated or predicted values. Estimates with credible intervals that include zero are statistically unclear (Dushoff et al., 2019). We graphically inspected the goodness of fit, and the distribution of the residuals. Unless stated differently, plots were created with ‘ggplot’ function from the ‘ggplo2’ R-package (Wickham, 2016).
To investigate whether sperm traits differ between the morphs we fitted linear models with each sperm trait as a response variable, and with male morph as a predictor (three-level factor). For analyses of velocity, we primarily used June values because the breeding season was at its peak in June and we obtained data from all but four males (for those four we used May values). We added the number of tracked sperm cells per sample (ln-transformed) as a covariate to the model to control for potential effects of sperm density or sample quality. For analyses of morphology, we used average male values. Alternative models using all velocity recordings, using individual sperm measurements, or controlling for sampling month or aviary gave similar results (Supplementary Figure S6). Because correlations between sperm traits and inbreeding were weak (mean r = -0.09, range: -0.26 to 0.07; Supplementary Figure S7, Methods S4), we did not include the inbreeding coefficient in the main models. Furthermore, the results were not confounded by relatedness (estimated based on microsatellite genotyping data) of the sampled individuals (see Supplementary Methods S4, Table S3, Figure S8).
To investigate whether sperm morphology explained variation in sperm swimming speed, we fitted a set of linear models with each velocity measure as the dependent variable (using predominantly June values), and with the male average of each sperm morphology trait as a predictor. All models were controlled for the number of tracked sperm (ln-transformed) and morph (three-level factor). As the relationship between sperm swimming speed and morphology might be non-linear (Bennison et al., 2016), we also fitted a set of linear models that included the quadratic term of each morphology trait (2nd order polynomial). We then tested whether the quadratic term improved the model fit, using Akaike’s Information Criterion corrected for sample size (Anderson, 2008), with the ‘AICc’ function from the ‘MuMIn’ R-package (Bartoń, 2022). Because this was not the case for any trait (Supplementary Figure S4), we report results from the simple model without the quadratic term. To investigate whether the effect of one sperm component (head, midpiece and tail length) was influenced by the other components, we also ran a multivariate model that contained the three sperm components as predictors. Correlations between the three predictors were low (Supplementary Figure S5). The effect sizes and uncertainties for head, midpiece and tail length were similar in the multivariate and the univariate models (Supplementary Figure S9) and we report the results from the univariate models in the main text.
Results and discussion
Between-morph differences in sperm traits
Sperm swimming speed in vitro in a standard medium, measured as curvilinear velocity, did not differ between the three morphs (Figure 3, Table 1; Supplementary Figure S6; Bulla et al., 2024), but measured as straight-line or average-path velocity, Faeder sperm moved slower than the sperm of both Independents and Satellites, with the effect being clearer for the straight-line velocity (Faeder relative to Independent estimate [95%CI]: –0.62 [-1.24 - -0.02], relative to Satellite: -0.78 [-1.42 - -0.13]) than for the average-path velocity (-0.36 [-0.89 - 0.16] and -0.46 [-1.03 - 0.09]; Table 1). Note, however, that the posterior distribution of the parameters indicates a 92% probability for Faeder relative to Independent and a 95% probability for Faeder relative to Satellite that the true mean difference in the average-path velocity is less than zero. Independents and Satellites did not differ in sperm velocity (Figure 3; Supplementary Figure S6, Table 1).
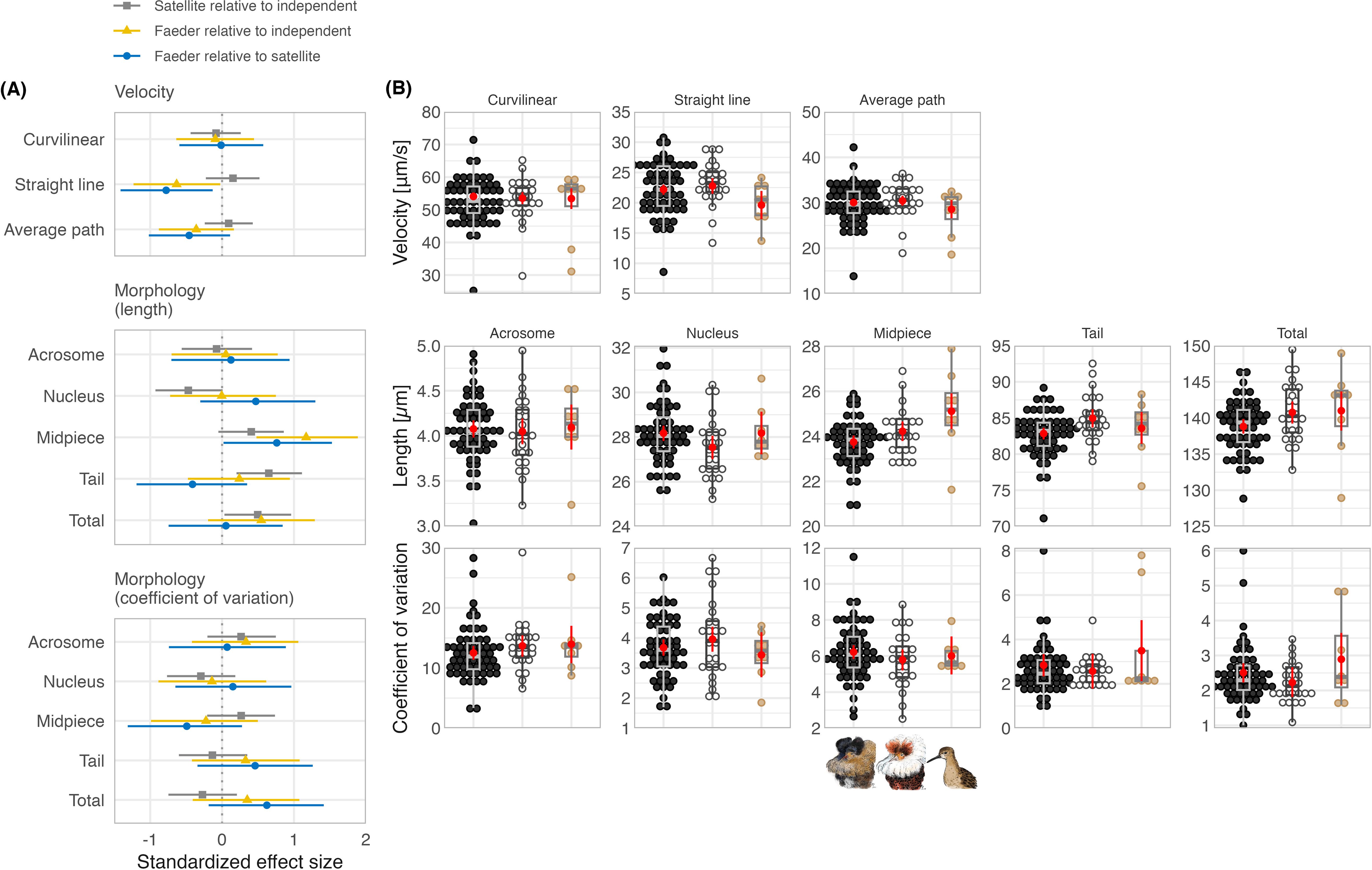
Figure 3. Differences in sperm traits between ruff morphs. (A) Model predictions of between-morph differences with their 95% CI (Table 1). (B) Morph-specific sperm traits, shown as individual data points with dot color highlighting morph (black: Independents, white: Satellites, beige: Faeders) and summarized as boxplots (see Figure 1 for definition). To aid visualization, the outlier coefficients of variation for tail (18.7) and total length (9.8) in Independents are depicted as 8 and 6, respectively. The red dots with bars show model predictions and their 95% CI (Table 1). Dots were stacked using the ‘geom_dotplot’ ggplot2 R-function (Wickham, 2016). (A, B) Velocity represents June values (with the exception of four males with May values only). Morphology represents average trait length based on 10 sperm cells per male (from a single sperm sample) and coefficients of variation are based on the same 10 sperm cells. For estimates of head, flagellum (absolute and relative) and relative midpiece length, see Supplementary Figures S6, S8. Ruff morph illustrations by Yifan Pei under Creative Commons Attribution (CC BY 4.0).
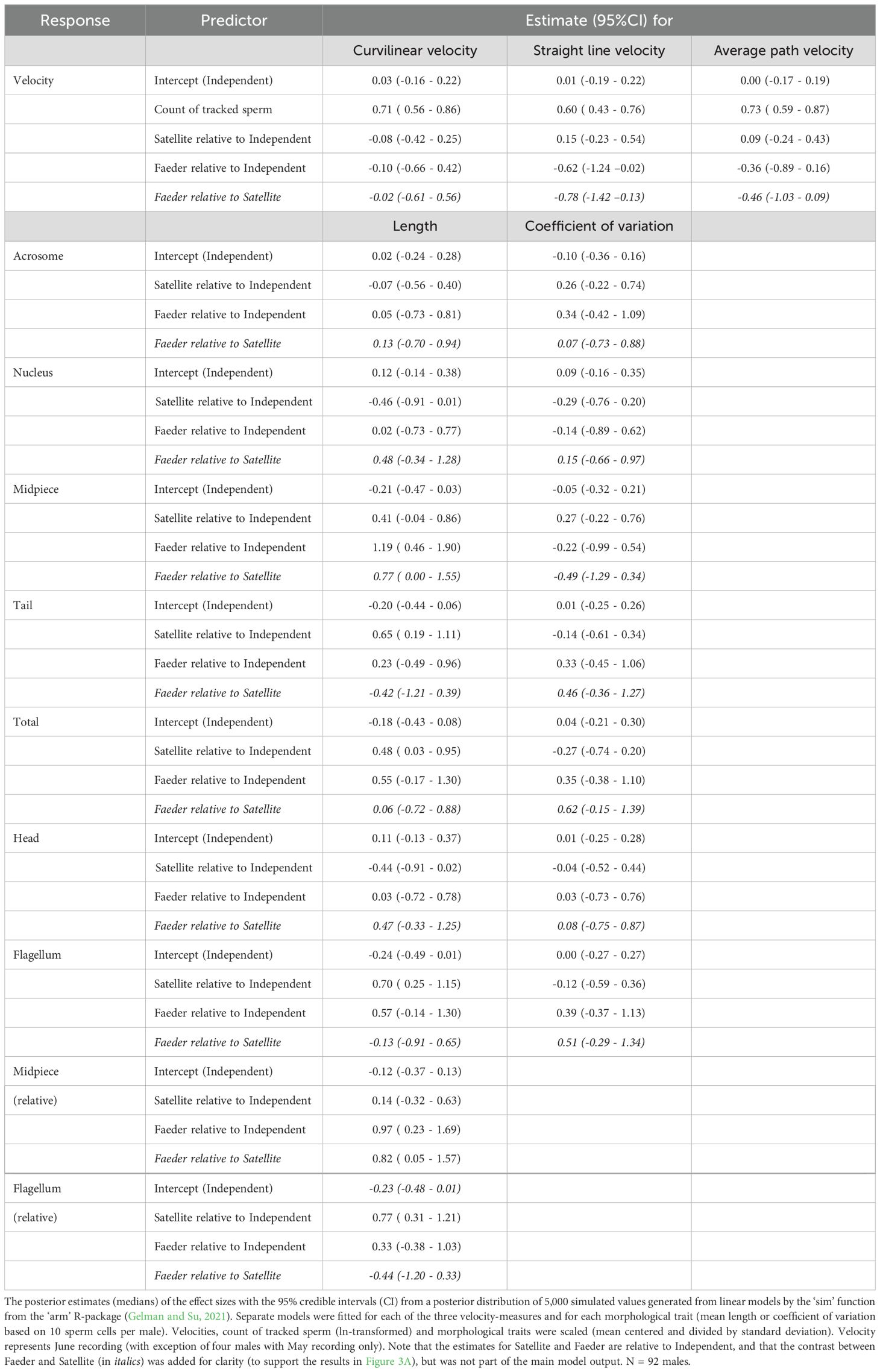
Table 1. Differences in sperm traits of ruff morphs.
Sperm morphology differed between the three morphs, with exception of midpiece, by less than one standard deviation (Figure 3; Supplementary Figure S6, Table 1). Sperm of Independents were the shortest with the shortest midpiece and tail, and hence flagellum (midpiece and tail length combined). Acrosome and nucleus length, and hence head length, as well as tail did not differ between Faeders and Independents. Satellite sperm had the shortest nucleus and head, but the longest tail. The midpiece was longest in Faeders and 1.19 standard deviations longer than the midpiece from Independents (95%CI: 0.46 - 1.88; Figure 3, Table 1). Total sperm length and flagellum length were similar for Faeders and Satellites (Figure 3; Supplementary Figure S6, Table 1). Consequently, midpiece length relative to total sperm length was largest for Faeders, but similar for Satellites and Independents; flagellum length relative to total sperm length was largest in Satellites and similar in the other two morphs (Supplementary Figure S6, Table 1). The coefficients of variation in sperm traits were noisy with unclear trends and sperm of Independents was not less variable than that of the Faeders and Satellites (Figure 3; Supplementary Figure S8, Table 1).
We thus demonstrate quantitative difference in sperm traits, but also a lack of major differences, i.e., sperm polymorphism, among the three male morphs. The two genes related to spermatogenesis that reside within the inversion thus have at most limited impact on the measured sperm traits. Such finding might be unsurprising given that tens to hundreds of genes influence spermatogenesis (Linn et al., 2021). Furthermore, the lack of major differences in sperm traits accords with previous studies showing no clear and consistent differences in sperm traits between males that use alternative mating tactics (Dougherty et al., 2022; Kustra and Alonzo, 2020).
Although one expects a steeper selection gradient for Faeder and Satellite males that have a lower probability to copulate multiple times with the same female (reviewed in Dougherty et al., 2022; Kustra and Alonzo, 2020), selection will act on all male morphs to optimize sperm traits. Additionally, males of all morphs mate with females of all morphs (Lank et al., 2002; Thuman, 2003), matching the size of sperm storage tubules in the female population may normalize selection on sperm size. Indeed, independent of the strength of sexual selection, Faeder-beneficial mutations are orders of magnitude less likely to arise than Independent-beneficial mutations, given a Faeder population size of 1% and the need for the beneficial mutation to arise within the inversion, being less than 0.5% of the genome. Moreover, outside the inversion, the genetic background has been selected to function best in the most abundant Independents, making evolution of special adaptations more difficult in the rare morphs (Giraldo-Deck et al., 2022).
Our results thus highlight that greater sperm competitiveness is an unlikely mechanism maintaining Faeders or Satellite inversions in the ruff population. The measured sperm traits seem to give the Faeder males little advantage over Independent males. However, there is a strong mating skew with only few Independents obtaining most copulations on a given lek (Tolliver et al., 2023; Vervoort and Kempenaers, 2019; Widemo and Owens, 1995), such that the average reproductive success of an Independent will be low. Thus, the rare Faeder males need moderate mating success through sneaking, facilitated by their female mimicking appearance and behavior (Jukema and Piersma, 2006) to compensate. Moreover, if Faeders copulate less frequently (no data), they may produce larger ejaculates and hence a rare copulation may be more likely to be successful everything else being equal (“more tickets in the lottery”). Possibly, Faeders do better, because morphs differ in other, non-measured, but fitness-relevant sperm traits. For example, the longer midpiece of Faeder sperm might correlate with longer sperm survival inside the female reproductive tract. However, this would need further study. To emphasize, despite the impressive work on genetics and genomics of ruff morphs (described in the introduction), we are still missing essential behavioral data from the wild, such as mating success and fitness measurements of individual males. Such lack of mating success data prohibits to make clear predictions about how differences in sperm competition strength and intensity between the three male morphs in ruffs influence their sperm traits.
Relationship between sperm swimming speed and morphology
Sperm morphology had only weak and unclear effects on variation in sperm swimming speed (Figure 4; Supplementary Figures S9, S10, Table S5). Males having longer sperm, sperm with a longer midpiece (absolute and relative) or sperm with a longer flagellum did not have sperm that swam faster (r varied between 0.05 and 0.18; Supplementary Figure S10). The lack of an association between sperm midpiece length and swimming speed may have several explanations. First, our method to measure sperm velocity might not reflect the swimming speed of the sperm within the seminal fluid or within the female reproductive tract. Note, however, that our preliminary work suggests that the way the sperm moved did not change after we placed the sperm into a solution that contained cloacal fluid of a ruff female. Also, our method is the same as the one used to measure swimming speed in passerine sperm, and we used the exact same equipment and observers as in some of the studies on passerines (e.g., Knief et al., 2017; Míčková et al., 2023; Opatová et al., 2016). Second, although the midpiece contains the energy-producing mitochondria, midpiece volume rather than length might be relevant for sperm swimming speed (Cardullo and Baltz, 1991; Mendonca et al., 2018), but see Cramer et al. (2022), or midpiece length might be associated with other sperm performance parameters such as longevity (which we did not measure). Third, it remains unclear whether oxidative phosphorylation in the midpiece is the primary source of energy for moving sperm (Turner, 2003).
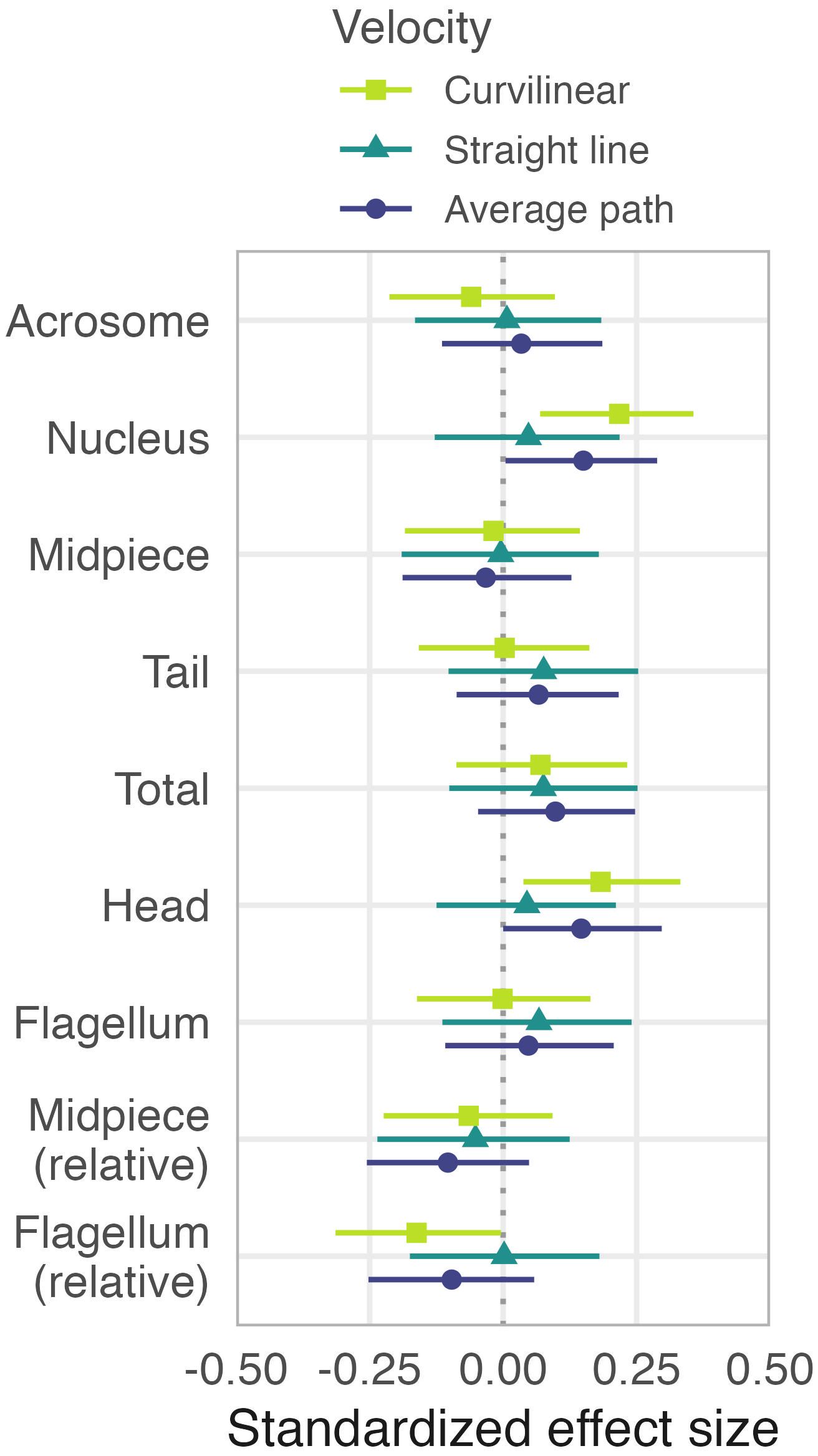
Figure 4. Effects of sperm morphology on sperm swimming speed. Shapes with horizontal bars represent estimated standardized effect sizes with their 95%CIs based on the joint posterior distribution of 5,000 simulated values generated from linear models, controlled for the number of tracked sperm (ln-transformed) and morph (Supplementary Table S5). The results are based on single June-values for velocity (with the exception of four males with May-values only) and average trait length of 10 sperm cells per male. For raw data and predicted relationships see Supplementary Figure S10.
Against the predictions (Humphries et al., 2008), sperm with longer heads (nucleus) or smaller relative flagellum length swam faster, when measured as curvilinear and average-path velocity (but not straight-line velocity), although these effects were small (< 0.25 of standard deviation, Figure 4; maximum r = 0.22, Supplementary Figure S10). Theory predicts that sperm with a shorter head relative to flagellum should swim faster because a relatively shorter head reduces drag (Humphries et al., 2008). The few existing within-species studies provide equivocal evidence for this prediction (Cramer et al., 2015, 2021; Helfenstein et al., 2009; Mossman et al., 2009), but the prediction has been supported in a between-species study in mammals (Tourmente et al., 2011) and in passerine birds (Lüpold et al., 2009). A recent study on passerines showed that sperm with a long acrosome, a short nucleus, a wide helical membrane, and a more pronounced waveform along the sperm head “core” swam faster (Støstad et al., 2018).
Sperm motility and sperm shape in ruffs compared to other taxa
We observed that ruff sperm seemed to move in a different manner to that previously described for passerine and likely also for non-passerine sperm (video examples). Specifically, ruff sperm seem to ‘vibrate’ from side to side while slowly moving forward, whereas passerine sperm typically swim straight by rapidly rotating around their longitudinal axis (Ballowitz, 1888; Humphreys, 1972; Vernon and Woolley, 1999). We can exclude that the unusual head movements of ruff sperm were caused by a lack of depth in the standard 20μm two-chamber count Leja-slide, because using a deeper Leja-slide (100μm) resulted in similar movements (not recorded). However, our observations need further verification using other methods (Vernon and Woolley, 1999). If confirmed, the difference in movement may not be due to differences in general morphology, because ruff sperm have a helical, screw-like head (Figure 2), similar to that observed in passerine sperm (Retzius, 1909). Previous studies described that non-passerine sperm moves with regular helical (dextral) waves (Ballowitz, 1888; Humphreys, 1972; Vernon and Woolley, 1999) or via counter-clockwise turning of the entire sperm body (Bird and Laguë, 1977). Given the few descriptions and the lack of video recordings of non-passerine sperm movements (e.g., Cheng et al., 2002; Denk et al., 2005; Dogliero et al., 2017; Fischer et al., 2014; Gloria et al., 2014), it remains to be seen whether the way ruff sperm propel is unique to this species, or typical for other Scolopacids, shorebirds or Charadriiformes in general. Clearly, the directional movement of ruff sperm is distinct from the smooth, snake-like, movements of sperm of non-avian taxa (e.g., human or sea-urchin Psammechinus miliaris (Cosson et al., 2015; Gillies et al., 2009; Gray, 1955; Saggiorato et al., 2017; Smith et al., 2009), but see squid sperm (Bishop, 1958)).
The screw-like head of ruff sperm is similar to the head shape reported for other Scolopaci (sandpipers), but differs from the straight heads observed in other Charadriiformes, e.g., genus Larus or Vanellus (Retzius, 1909). Our results confirm that ruff sperm is remarkably long (median and mean = 139.5 µm, range: 105.7 - 150 µm) compared to other shorebirds, with species means ranging between 57 and 100 µm, which has been interpreted as a sign that sperm competition was/is strong in ruffs (Johnson and Briskie, 1999).
Conclusions
Using sperm collected from a population of captive male-polymorphic ruffs, we found small quantitative differences between morphs in several measures of sperm swimming speed and sperm morphology, but no clear sperm polymorphism. Hence, it seems unlikely that Satellite and Faeder males have compensated for reduced mating opportunities through the production of more competitive sperm. Furthermore, we found that the sperm of ruffs move differently than sperm of passerines, despite having a similar head shape (Ballowitz, 1888; Humphreys, 1972; Retzius, 1909; Vernon and Woolley, 1999). Our results show at best weak associations between sperm swimming speed and length measurements that are inconsistent with general expectations and, thus, corroborate other comparative work showing a lack of clear and consistent differences in sperm traits of males using alternative mating tactics (Dougherty et al., 2022; Kustra and Alonzo, 2020). Our study leaves open the question of whether and how Faeder males may achieve higher reproductive success to compensate for the lower reproductive output of Faeder females (Giraldo-Deck et al., 2022), but emphasizes the low potential for the evolution of strong morph-specific sperm adaptations in this system.
Data availability statement
The original contributions presented in the study are publicly available. All data, including sperm recordings and pictures, and computer code used to generate the results and Supplement (figures, tables and documents) of this study are freely available at GitHub: https://github.com/MartinBulla/ruff_sperm_v2 (Bulla et al., 2024).
Ethics statement
The data were collected under license (311.5-5682.1/1-2014-020) of the Landratsamt Starnberg and in accordance with the local legislation and institutional requirements.
Author contributions
MB: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. CK: Conceptualization, Writing – review & editing. DL: Conceptualization, Writing – review & editing. JA: Investigation, Methodology, Writing – review & editing. JL: Conceptualization, Methodology, Writing – review & editing. KM: Investigation, Writing – review & editing. KT: Investigation, Methodology, Software, Writing – review & editing. MC: Investigation, Methodology, Software, Writing – review & editing. ML: Investigation, Methodology, Writing – review & editing. TA: Conceptualization, Funding acquisition, Investigation, Methodology, Resources, Writing – review & editing. WF: Conceptualization, Formal analysis, Methodology, Writing – review & editing. BK: Conceptualization, Funding acquisition, Methodology, Project administration, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Funding by Max Planck Society (to BK and CK), the Czech Science Foundation project 19-22538S (to TA and JA) and the Research Excellence in Environmental Sciences Project (REES 003) from the Faculty of Environmental Sciences, Czech University of Life Sciences Prague (to MB).
Acknowledgments
We thank the many people who took care of the captive ruffs, Melanie Schneider for genotyping, Callum S. McDiarmid for help with his Sperm Sizer software, Lotte Schlicht and Jochen Wolf for discussions, Esteban Botero-Delgadillo for the script on homozygosity by locus, Jarrod Hadfield for advice on a genomic relationship matrix, Mihai Valcu for advice on statistics, Yifan Pei for ruff illustrations, and seven anonymous reviewers for the comments.
Conflict of interest
The authors declare that the research was conducted in theabsence of any commercial or financial relationships that could beconstrued as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Anderson D. R. (2008). Model based inference in the life sciences: a primer on evidence (Springer), 184. Available at: https://link.springer.com/book/10.1007/978-0-387-74075-1
Ballowitz E. (1888). Untersuchungen über die Struktur der Spermatozoën, zugleich ein Beitrag zur Lehre vom feineren Bau der contraktilen Elemente. Archiv für Mikroskopische Anatomie 32, 401–473. doi: 10.1007/BF02956997
Bartoń K. (2022). MuMIn: Multi-Model Inference. R package version 1.46.0. Available online at: https://CRAN.R-project.org/package=MuMIn.
Bennison C., Hemmings N., Brookes L., Slate J., Birkhead T. (2016). Sperm morphology, adenosine triphosphate (ATP) concentration and swimming velocity: unexpected relationships in a passerine bird. Proc. Biol. Sci. 283, 20161558. doi: 10.1098/rspb.2016.1558
Bennison C., Hemmings N., Slate J., Birkhead T. (2014). Long sperm fertilize more eggs in a bird. Proc. R. Soc. B: Biol. Sci. 282, 20141897–20141897. doi: 10.1098/rspb.2014.1897
Bird D. M., Laguë P. C. (1977). Semen production of the American kestrel. Can. J. Zoology 55, 1351–1358. doi: 10.1139/z77-175
Birkhead T. R., Martínez J. G., Burke T., Froman D. P. (1999). Sperm mobility determines the outcome of sperm competition in the domestic fowl. Proc. R. Soc. B: Biol. Sci. 266, 1759–1764. doi: 10.1098/rspb.1999.0843
Bishop D. W. (1958). Motility of the sperm flagellum. Nature 182, 1638–1640. Available at: https://www.nature.com/articles/1821638a0.
Briskie J. V., Montgomerie R. (1993). Patterns of sperm storage in relation to sperm competition in passerine birds. Condor 95, 442–454. doi: 10.2307/1369366
Briskie J. V., Montgomerie R., Birkhead T. R. (1997). The evolution of sperm size in birds. Evolution 51, 937–945. doi: 10.1111/j.1558-5646.1997.tb03674.x
Bulla M., Küpper C., Lank D. B., Albrechtová J., Loveland J. L., Martin K., et al. (2024). Supporting information for '"Differences in sperm swimming speed and morphology between the three genetic morphs in the ruff sandpiper (Calidris pugnax)": GitHub. Available online at: https://github.com/MartinBulla/ruff_sperm_v2.
Cardullo R. A., Baltz J. M. (1991). Metabolic regulation in mammalian sperm: mitochondrial volume determines sperm length and Flagellar beat frequency. Cell Motil. Cytoskeleton 19, 180–188. doi: 10.1002/cm.970190306
Cheng F. P., Guo T. J., Wu J. T., Lin T. E., Ursem P. J., Colenbrander B., et al. (2002). Annual variation in semen characteristics of pigeons (Columba livia). Poultry Sci. 81, 1050–1056. doi: 10.1093/ps/81.7.1050
Cosson J. J., Prokopchuk G., Inaba K. (2015). “The flagellar mechanics of spermatozoa and its regulation,” in Flagellar Mechanics and Sperm Guidance. Ed. Cosson J. J., (Sharjah, U.A.E.: Bentham Science Publishers Ltd.) 3–134. doi: 10.2174/97816810812811150101
Cramer E. R., Alund M., McFarlane S. E., Johnsen A., Qvarnstrom A. (2016). Females discriminate against heterospecific sperm in a natural hybrid zone. Evolution 70, 1844–1855. doi: 10.1111/evo.12986
Cramer E. R. A., Garcia-Del-Rey E., Johannessen L. E., Laskemoen T., Marthinsen G., Johnsen A., et al. (2021). Longer sperm swim more slowly in the Canary Islands Chiffchaff. Cells 10. doi: 10.3390/cells10061358
Cramer E. R. A., Gronstol G., Lifjeld J. T. (2022). Flagellum tapering and midpiece volume in songbird spermatozoa. J. Morphology 283, 1577–1589. doi: 10.1002/jmor.21524
Cramer E. R., Laskemoen T., Stensrud E., Rowe M., Haas F., Lifjeld J. T., et al. (2015). Morphology-function relationships and repeatability in the sperm of Passer sparrows. J. Morphology 276, 370–377. doi: 10.1002/jmor.20346
Denk A. G., Holzmann A., Peters A., Vermeirssen E. L. M., Kempenaers B. (2005). Paternity in mallards: effects of sperm quality and female sperm selection for inbreeding avoidance. Behav. Ecol. 16, 825–833. doi: 10.1093/beheco/ari065
Dogliero A., Rota A., Lofiego R., von Degerfeld M. M., Quaranta G. (2017). Semen collection and spermatozoa characteristics in the kea parrot (Nestor notabilis). J. Avian Med. Surg. 31, 24–28. doi: 10.1647/2015-132
Dougherty L. R., Skirrow M. J. A., Jennions M. D., Simmons L. W. (2022). Male alternative reproductive tactics and sperm competition: a meta-analysis. Biol. Rev. Camb Philos. Soc. 97, 1365–1388. doi: 10.1111/brv.12846
Dushoff J., Kain M. P., Bolker B. M., O’Hara R. B. (2019). I can see clearly now: Reinterpreting statistical significance. Methods Ecol. Evol. 10, 756–759. doi: 10.1111/2041-210x.13159
Fischer D., Neumann D., Wehrend A., Lierz M. (2014). Comparison of conventional and computer-assisted semen analysis in cockatiels (Nymphicus hollandicus) and evaluation of different insemination dosages for artificial insemination. Theriogenology 82, 613–620. doi: 10.1016/j.theriogenology.2014.05.023
Fitzpatrick J. L., Lüpold S. (2014). Sexual selection and the evolution of sperm quality. Mol. Hum. Reprod. 20, 1180–1189. doi: 10.1093/molehr/gau067
Gage M. J. G., Macfarlane C. P., Yeates S., Ward R. G., Searle J. B., Parker G. A. (2004). Spermatozoal traits and sperm competition in Atlantic Salmon. Curr. Biol. 14, 44–47. doi: 10.1016/j.cub.2003.12.028
Garcia-Gonzalez F., Simmons L. W. (2005). Sperm viability matters in insect sperm competition. Curr. Biol. 15, 271–275. doi: 10.1016/j.cub.2005.01.032
Garcia-Gonzalez F., Simmons L. W. (2007). Shorter sperm confer higher competitive fertilization success. Evolution 61, 816–824. doi: 10.1111/j.1558-5646.2007.00084.x
Gelman A., Hill J. (2007). Data analysis using regression and multilevel/hierarchical models (Cambridge: Cambridge University Press).
Gelman A., Su Y.-S. (2021). arm: Data Analysis Using Regression and Multilevel/Hierarchical Models. R package version 1.12-2. (Version R package version 1.12-2.). Available online at: http://CRAN.R-project.org/package=arm.
Gillies E. A., Cannon R. M., Green R. B., Pacey A. A. (2009). Hydrodynamic propulsion of human sperm. J. Fluid Mechanics 625, 445–474. doi: 10.1017/s0022112008005685
Giraldo-Deck L. M., Goymann W., Safari I., Dawson D. A., Stocks M., Burke T., et al. (2020). Development of intraspecific size variation in black coucals, white-browed coucals and ruffs from hatching to fledging. J. Avian Biol. 51, e02440. doi: 10.1111/jav.02440
Giraldo-Deck L. M., Loveland J. L., Goymann W., Tschirren B., Burke T., Kempenaers B., et al. (2022). Intralocus conflicts associated with a supergene. Nat. Commun. 13, 1384. doi: 10.1038/s41467-022-29033-w
Gloria A., Contri A., Carluccio A., Parrillo S., Cicconi M., Robbe D. (2014). The breeding management affects fresh and cryopreserved semen characteristics in Melopsittacus undulatus. Anim. Reprod. Scis 144, 48–53. doi: 10.1016/j.anireprosci.2013.11.009
Gray J. (1955). The movement of sea-urchin spermatozoa. J. Exp. Biol. 32, 775–801. doi: 10.1242/jeb.32.4.775
Graziotto R., Foresta C., Scannapieco P., Zeilante P., Russo A., Negro A., et al. (1999). cDNA cloning and characterization of PD1: A novel human testicular protein with different expressions in various testiculopathies. Exp. Cell Res. 248, 620–626. doi: 10.1006/excr.1999.4449
Gross M. R. (1996). Alternative reproductive strategies and tactics: diversity within sexes. Trends Ecol. Evol. 11, 92–98. doi: 10.1016/0169-5347(96)81050-0
Helfenstein F., Podevin M., Richner H. (2009). Sperm morphology, swimming velocity, and longevity in the house sparrow Passer domesticus. Behav. Ecol. Sociobiology 64, 557–565. doi: 10.1007/s00265-009-0871-x
Hill J., Enbody E. D., Bi H., Lamichhaney S., Lei W., Chen J., et al. (2023). Low mutation load in a supergene underpinning alternative male mating strategies in ruff (Calidris pugnax). Mol. Biol. Evol. 40, msad224. doi: 10.1093/molbev/msad224
Hogan-Warburg A. J. (1966). Social behavior of the ruff, Philomachus pugnax (L.). Ardea 54, 109–229. doi: 10.5253/arde.v54.p109
Höglund J., Lundberg A. (1989). Plumage color correlates with body size in the ruff. Auk 106, 336–338. doi: 10.1093/auk/106.2.336
Hugie D. M., Lank D. B. (1997). The resident's dilemma: a female choice model for the evolution of alternative mating strategies in lekking male ruffs (Philomachus pugnax). Behav. Ecol. 8, 218–225. doi: 10.1093/beheco/8.2.218
Humphreys P. (1972). Brief observations on the semen and spermatozoa of certain passerine and non-passerine birds. J. Reprod. Fertility 29, 327–336. doi: 10.1530/jrf.0.0290327
Humphries S., Evans J. P., Simmons L. W. (2008). Sperm competition: linking form to function. BMC Evolutionary Biol. 8, 319. doi: 10.1186/1471-2148-8-319
Johnson D. D. P., Briskie J. V. (1999). Sperm competition and sperm length in shorebirds. Condor 101, 848–854. doi: 10.2307/1370074
Jukema J., Piersma T. (2006). Permanent female mimics in a lekking shorebird. Biol. Lett. 2, 161–164. doi: 10.1098/rsbl.2005.0416
Kim K.-W., Bennison C., Hemmings N., Brookes L., Hurley L. L., Griffith S. C., et al. (2017). A sex-linked supergene controls sperm morphology and swimming speed in a songbird. Nat. Ecol. Evol. 1, 1168–1176. doi: 10.1038/s41559-017-0235-2
Kleven O., Laskemoen T., Fossøy F., Robertson R. J., Lifjeld J. T. (2008). Intraspecific variation in sperm length is negatively related to sperm competition in passerine birds. Evolution 62, 494–499. doi: 10.1111/j.1558-5646.2007.00287.x
Knief U., Forstmeier W., Pei Y., Ihle M., Wang D., Martin K., et al. (2017). A sex-chromosome inversion causes strong overdominance for sperm traits that affect siring success. Nat. Ecol. Evol. 1, 1177–1184. doi: 10.1038/s41559-017-0236-1
Küpper C., Stocks M., Risse J. E., Dos Remedios N., Farrell L. L., McRae S. B., et al. (2016). A supergene determines highly divergent male reproductive morphs in the ruff. Nat. Genet. 48, 79–83. doi: 10.1038/ng.3443
Kustra M. C., Alonzo S. H. (2020). Sperm and alternative reproductive tactics: a review of existing theory and empirical data. Philos. Trans. R. Soc. London B Biol. Sci. 375, 20200075. doi: 10.1098/rstb.2020.0075
Lamichhaney S., Fan G., Widemo F., Gunnarsson U., Thalmann D. S., Hoeppner M. P., et al. (2016). Structural genomic changes underlie alternative reproductive strategies in the ruff (Philomachus pugnax). Nat. Genet. 48, 84–88. doi: 10.1038/ng.3430
Lank D. B., Farrell L. L., Burke T., Piersma T., McRae S. B. (2013). A dominant allele controls development into female mimic male and diminutive female ruffs. Biol. Lett. 9, 20130653. doi: 10.1098/rsbl.2013.0653
Lank D. B., Smith C. M., Hanotte O., Burke T., Cooke F. (1995). Genetic polymorphism for alternative mating behaviour in lekking male ruff Philomachus pugnax. Nature 378, 59–62. doi: 10.1038/378059a0
Lank D. B., Smith C. M., Hanotte O., Ohtonen A., Bailey S., Burke T. (2002). High frequency of polyandry in a lek mating system. Behav. Ecol. 13, 209–215. doi: 10.1093/beheco/13.2.209
Laskemoen T., Albrecht T., Bonisoli-Alquati A., Cepak J., Lope F., Hermosell I., et al. (2013). Variation in sperm morphometry and sperm competition among barn swallow (Hirundo rustica) populations. Behav. Ecol. Sociobiology 67, 1–9. doi: 10.1007/s00265-012-1450-0
Laskemoen T., Kleven O., Fossøy F., Robertson R. J., Rudolfsen G., Lifjeld J. T. (2010). Sperm quantity and quality effects on fertilization success in a highly promiscuous passerine, the tree swallow Tachycineta bicolor. Behav. Ecol. Sociobiology 64, 1–11. doi: 10.1007/s00265-010-0962-8
Lierz M., Reinschmidt M., Muller H., Wink M., Neumann D. (2013). A novel method for semen collection and artificial insemination in large parrots (Psittaciformes). Sci. Rep. 3, 2066. doi: 10.1038/srep02066
Lifjeld J. T., Laskemoen T., Kleven O., Albrecht T., Robertson R. J. (2010). Sperm length variation as a predictor of extrapair paternity in passerine birds. PloS One 5, e13456. doi: 10.1371/journal.pone.0013456
Linn E., Ghanem L., Bhakta H., Greer C., Avella M. (2021). Genes regulating spermatogenesis and sperm function associated with rare disorders. Front. Cell Dev. Biol. 9. doi: 10.3389/fcell.2021.634536
Lipshutz S. E., Torneo S. J., Rosvall K. A. (2022). How female-female competition affects male-male competition: insights on post-copulatory sexual selection from socially polyandrous species. Am. Nat. 201, 460–471. doi: 10.1086/722799
Loveland J. L., Giraldo-Deck L. M., Lank D. B., Goymann W., Gahr M., Küpper C. (2021a). Functional differences in the hypothalamic-pituitary-gonadal axis are associated with alternative reproductive tactics based on an inversion polymorphism. Hormones Behav. 127, 104877. doi: 10.1016/j.yhbeh.2020.104877
Loveland J. L., Lank D. B., Küpper C. (2021b). Gene expression modification by an autosomal inversion associated with three male mating morphs. Front. Genet. 12. doi: 10.3389/fgene.2021.641620
Lüpold S., Calhim S., Immler S., Birkhead T. R. (2009). Sperm morphology and sperm velocity in passerine birds. Proc. Biol. Sci. 276, 1175–1181. doi: 10.1098/rspb.2008.1645
Lüpold S., de Boer R. A., Evans J. P., Tomkins J. L., Fitzpatrick J. L. (2020). How sperm competition shapes the evolution of testes and sperm: a meta-analysis. Philos. Trans. R. Soc. London B Biol. Sci. 375, 20200064. doi: 10.1098/rstb.2020.0064
Lüpold S., Manier M. K., Berben K. S., Smith K. J., Daley B. D., Buckley S. H., et al. (2012). How multivariate ejaculate traits determine competitive fertilization success in Drosophila melanogaster. Curr. Biology: CB. 22, 1667–1672. doi: 10.1016/j.cub.2012.06.059
Maney D. L., Küpper C. (2022). Supergenes on steroids. Philos. Trans. R. Soc. London B Biol. Sci. 377, 20200507. doi: 10.1016/10.1098/rstb.2020.0507
Mank J. E. (2023). Sex-specific morphs: the genetics and evolution of intra-sexual variation. Nat. Rev. Genet. 24, 44–52. doi: 10.1038/s41576-022-00524-2
McDiarmid C. S., Li R., Kahrl A. F., Rowe M., Griffith S. C. (2021). Sperm Sizer: a program to semi-automate the measurement of sperm length. Behav. Ecol. Sociobiology 75, 84. doi: 10.1007/s00265-021-03013-4
McNab B. K. (1966). An analysis of the body temperatures of birds. Condor 68, 47–55. doi: 10.2307/1365174
Mendonca T., Birkhead T. R., Cadby A. J., Forstmeier W., Hemmings N. (2018). A trade-off between thickness and length in the zebra finch sperm mid-piece. Proc. Biol. Sci. 285, 20180865. doi: 10.1098/rspb.2018.0865
Míčková K., Tomášek O., Jelínek V., Šulc M., Pazdera L., Albrechtová J., et al. (2023). Age-related changes in sperm traits and evidence for aging costs of sperm production in a sexually promiscuous passerine. Front. Ecol. Evol. 11. doi: 10.3389/fevo.2023.1105596
Mossman J., Slate J., Humphries S., Birkhead T. (2009). Sperm morphology and velocity are genetically codetermined in the zebra finch. Evolution 63, 2730–2737. doi: 10.1111/j.1558-5646.2009.00753.x
Oliverira R., Taborsky M., Brockman H. (2008). Alternative Reproductive Tactics. An Integrative Approach (NY: Cambridge Univ Press).
Onisto M., Graziotto R., Scannapieco P., Marin P., Merico M., Slongo M. L., et al. (2000). A novel gene (PD1) with a potential role on rat spermatogenesis. J. Endocrinological Invest. volume 23, 605–608. doi: 10.1007/BF03343783
Opatová P., Ihle M., Albrechtová J., Tomášek O., Kempenaers B., Forstmeier W., et al. (2016). Inbreeding depression of sperm traits in the zebra finch Taeniopygia guttata. Ecol. Evol. 6, 295–304. doi: 10.1002/ece3.1868
Parker G. A. (1970). Sperm competition and its evolutionary consequences in the insects. Biol. Rev. (Cambridge) 45, 525–567. doi: 10.1111/j.1469-185X.1970.tb01176.x
R-Core-Team (2022). R: A language and environment for statistical computing (Version Version 4.2.0) (Vienna (Austria: R Foundation for Statistical Computing). Available at: http://www.R-project.org/.
Saggiorato G., Alvarez L., Jikeli J. F., Kaupp U. B., Gompper G., Elgeti J. (2017). Human sperm steer with second harmonics of the flagellar beat. Nat. Commun. 8, 1415. doi: 10.1038/s41467-017-01462-y
Shuster S. M., Wade M. J. (1991). Equal mating success among male reproductive strategies in a marine isopod. Nature 350, 608–610. doi: 10.1038/350608a0
Simmons L. W., Fitzpatrick J. L. (2012). Sperm wars and the evolution of male fertility. Reproduction 144, 519–534. doi: 10.1530/REP-12-0285
Smith D. J., Gaffney E. A., Gadelha H., Kapur N., Kirkman-Brown J. C. (2009). Bend propagation in the flagella of migrating human sperm, and its modulation by viscosity. Cell Motil. Cytoskeleton 66, 220–236. doi: 10.1002/cm.20345
Støstad H. N., Johnsen A., Lifjeld J. T., Rowe M. (2018). Sperm head morphology is associated with sperm swimming speed: A comparative study of songbirds using electron microscopy. Evolution 72, 1918–1932. doi: 10.1111/evo.13555
Thuman K. (2003). Female Reproductive Strategies in the Ruff (Philomachus pugnax). (PhD) (Uppsala: Uppsala University). Available at: https://www.diva-portal.org/smash/record.jsf?pid=diva2%3A162874&dswid=-4434.
Tolliver J. D. M., Kupán K., Lank D. B., Schindler S., Küpper C. (2023). Fitness benefits from co-display favour subdominant male–male partnerships between phenotypes. Anim. Behav. 197, 131–154. doi: 10.1016/j.anbehav.2022.12.004
Tourmente M., Gomendio M., Roldan E. R. (2011). Sperm competition and the evolution of sperm design in mammals. BMC Evolutionary Biol. 11. doi: 10.1186/1471-2148-11-12
Turner R. M. (2003). Tales from the tail: what do we really know about sperm motility? J. Androl 24, 790–803. doi: 10.1002/j.1939-4640.2003.tb03123.x
Vernon G. G., Woolley D. M. (1999). Three-dimensional motion of avian spermatozoa. Cell Motil. Cytoskeleton 42, 149–161. doi: 10.1002/(sici)1097-0169(1999)42:2%3C149::aid-cm6%3E3.0.co;2-0
Vervoort R., Kempenaers B. (2019). Variation in lek attendance and copulation success of Independent and Satellite male Ruffs Calidris pugnax. Ardea 107, 303–320. doi: 10.5253/arde.v107i3.a9
Widemo F. (1997). The social implications of traditional use of lek sites in the ruff Philomachus pugnax. Behav. Ecol. 8, 211–217. doi: 10.1093/beheco/8.2.211
Widemo F. (1998). Alternative reproductive strategies in the ruff, Philomachus pugnax: a mixed ESS? Anim. Behav. 56, 329–336. doi: 10.1006/anbe.1998.0792
Yeh S. D., Chen Y. J., Chang A. C., Ray R., She B. R., Lee W. S., et al. (2002). Isolation and properties of Gas8, a growth arrest-specific gene regulated during male gametogenesis to produce a protein associated with the sperm motility apparatus. J. Biol. Chem. 277, 6311–6317. doi: 10.1074/jbc.M106941200
Keywords: alternative mating strategies, autosomal inversion, genetic morphs, spermatozoa, sperm morphology, sperm motility, sperm swimming speed, sperm velocity
Citation: Bulla M, Küpper C, Lank DB, Albrechtová J, Loveland JL, Martin K, Teltscher K, Cragnolini M, Lierz M, Albrecht T, Forstmeier W and Kempenaers B (2024) Sperm swimming speed and morphology differ slightly among the three genetic morphs of ruff sandpiper (Calidris pugnax), but show no clear polymorphism. Front. Ecol. Evol. 12:1476254. doi: 10.3389/fevo.2024.1476254
Received: 05 August 2024; Accepted: 18 October 2024;
Published: 02 December 2024.
Edited by:
Jarle Tryti Nordeide, Nord University, NorwayReviewed by:
Pamela Espíndola Hernández, Helmholtz Association of German Research Centres (HZ), GermanyJon Paul Evans, University of Western Australia, Australia
Copyright © 2024 Bulla, Küpper, Lank, Albrechtová, Loveland, Martin, Teltscher, Cragnolini, Lierz, Albrecht, Forstmeier and Kempenaers. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Martin Bulla, YnVsbGEubWFyQGdtYWlsLmNvbQ==; Bart Kempenaers, YmFydC5rZW1wZW5hZXJzQGJpLm1wZy5kZQ==