 Susana González
Susana González- 1Instituto de Investigaciones Biológicas Clemente Estable, Ministerio de Educación y Cultura (IIBCE-MEC), Montevideo, Uruguay
- 2Deer Specialist Group – International Union for Conservation of Nature Species Survival Commission (IUCN), Gland, Switzerland
- 3Facultad de Humanidades y Ciencias de la Educación, Universidad de la República, Montevideo, Uruguay
Introduction
The Neotropical biogeographical region stands as one of the richest in deer diversity, with the marsh deer (Blastocerus dichotomus, Illiger, 1815) emerging as the largest contemporary species (González and Duarte, 2020). Historically, this species roamed across various wetland habitats throughout Argentina, Bolivia, Brazil, Paraguay, Peru, and Uruguay (Weber and González, 2003). However, extensive habitat loss has led to a drastic reduction in its distribution, with current populations being small and isolated (Pinder and Grosse, 1991; Tomas et al., 1997). Although marsh deer once inhabited northeastern Brazil during the Pleistocene (Pinder and Grosse, 1991; Magalhaes et al., 1992), no paleontological or archaeological evidence exists for their presence in the Holocene north of latitude 10° S. Today, relictual populations persist in key areas such as the Pantanal in Brazil, and the Iberá and Paraná Delta in Argentina (Pinder and Grosse, 1991; Beccaceci, 1994; Mauro et al., 1998; Varela, 2003).
Currently, in Uruguay, the marsh deer is considered extinct in the wild, marking the southernmost limit of its range and the distribution boundary for several species of tropical flora and fauna, fostering the assembly of diverse ecotypes (González and Lessa, 2014). Historical records place the species in the departments of Rocha and Treinta y Tres within the wetlands of the “Bañados del Este,” and along the Uruguay River to the west (González, 1994). The last documented sighting dates to the capture of a specimen in 1958 in the marshy area known as “Los Indios” (Arredondo, 1959). Additionally, in 1991, an individual was poached in the Department of Soriano, though it likely originated from the Tigre Delta area in Argentina rather than representing a stable population in Uruguay (González, 1994). Evidence suggests that populations existing north of the Cuareim River in Brazil, near the Uruguayan border, may extend into the Artigas Department (Autuori, 1972).
In Argentina several contributions have recognized the presence of Blastocerus during the Platan Stage/Age, spanning from 8000 years BP to the present, as documented by Menegaz (2000); Alcaraz (2010), and Cione and Tonni (1999), Cione and Tonni (2005). Additionally, references to cf. Blastocerus have been made in the Argentine Pampean region during the Late Pleistocene (Chimento et al., 2019; Vezzosi et al., 2019). Concerning the archaeological record, numerous studies have documented the presence of marsh deer in Argentina, contributing significantly to our understanding of the species’ historical distribution within the region (Politis et al., 2011; Rodriguez and Bonomo, 2021).
However in Uruguay the presence was cited in archaeological sites (Moreno, 2016). During a drought in 1995, the retraction of the edge of Negra Lagoon (33° 95’ 83” S, 53° 38’ 41” W) led to the recovery of two partial male skeletons, though the fossilization process precluded material isolation for dating and genetic analysis (Roibal et al., 1996). In this report, we document the discovery of a complete male skull recovered in 2010 from the coastline of the Solís Grande stream, situated on the border of the Canelones and Maldonado departments in Uruguay (Figure 1). Our objective was to estimate the specimen’s age through radiocarbon dating, extract DNA, and analyze with mitochondrial marker to compare them with haplotypes described in Argentina and Brazil.
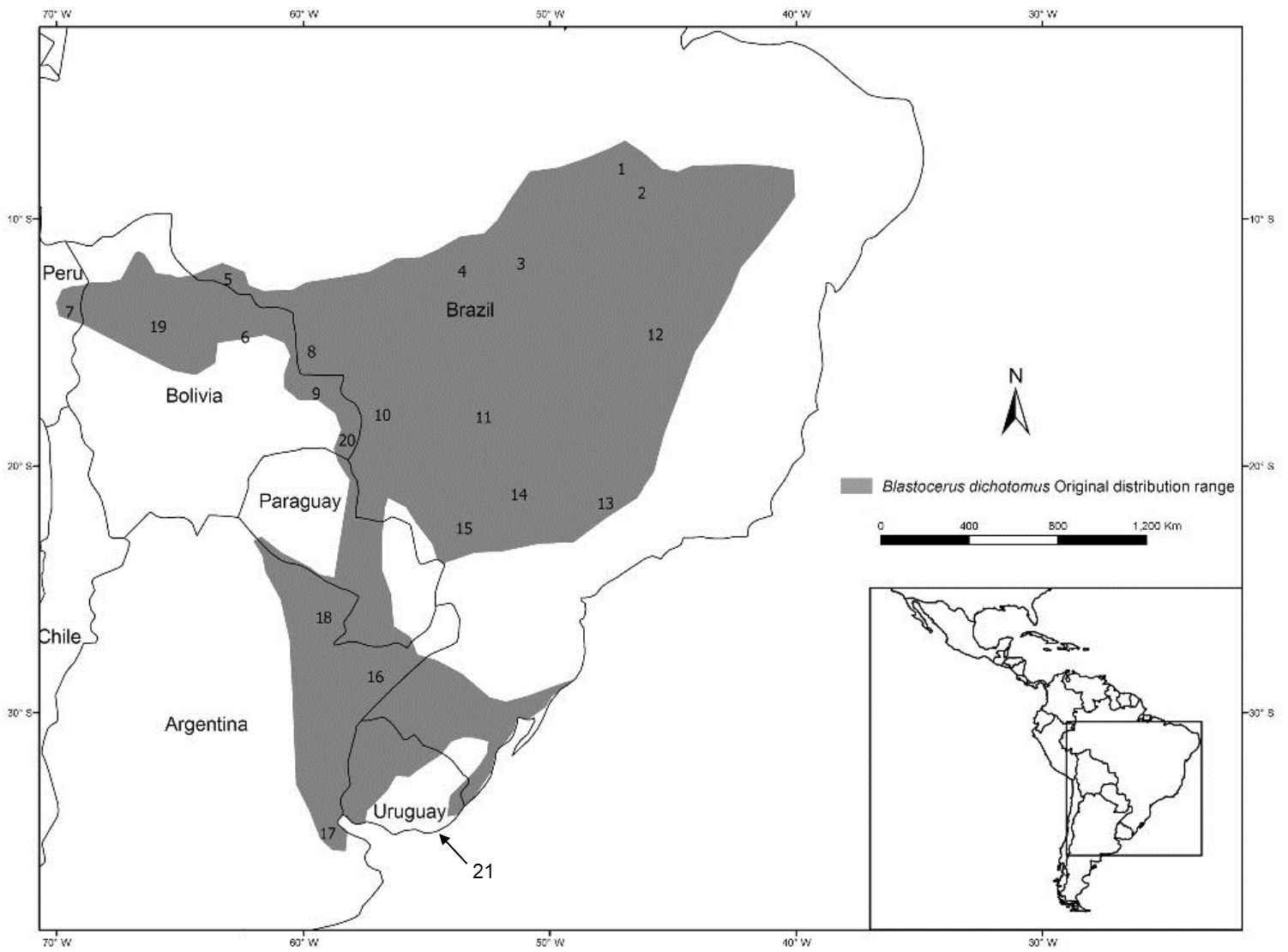
Figure 1 Geographic range of Blastocerus dichotomus and current known populations. 1 – Krahô Indian Land, Brazil; 2 – Southern Maranhão state, Brazil; 3 – floodplains of the Araguaia and Mortes rivers, Brazil; 4 – Xingu National Park, Brazil; 5 – Guaporé river floodplains in Bolivia and Brazil; 6 – Park Noel Kempff, Bolivia; 7 – Pampas del Heath floodplains, Peru; 8 – upper Guaporé river floodplains, Brazil; 9 – San Matias region, Bolivia; 10 - Pantanal wetland, Brazil; 11 – Emas National Park and Correntes river, Brazil; 12 – Grande Sertão Veredas National Park and headwaters of tributaries of São Francisco River near to the Serra Geral de Goiás/Chapadão and Eastern Bahia, Brazil; 13 – Jataí Ecological Station, Brazil; 14 – tributaries of the Porto Primavera Dam, Brazil; 15 – Floodplains of the Paraná river, Brazil; 16 – Esteros de Iberá/Esteros Santa Lucía, Riachuelo, Miriñay y Batel/Cuenca del Aguapey, Corrientes, Argentina; 17 – Delta del Paraná, Argentina; 18 – Wetlands of Eastern Formosa and Chaco, Argentina; 19 – Department of Beni, Bolivia; 20 – Otuquis National Park, Bolivia; 21 New record Department of Maldonado, Uruguay. Map based on Piovezan et al. (2010).
Methods
Sample collection
In the summer of 2010, during a drought along the coastline of the Solís Grande stream (34° 44’ 14.33” S; 55° 27’ 3.68” W), bordering the departments of Canelones and Maldonado in Uruguay, a marsh deer skull was observed and manually extracted. The sedimentary deposits are from the Quaternary period, composed of very fine sand and Holocene mud, specifically located on the river margin with variable depth (Spoturno et al., 2012). The substrate is characterized by elongated bays, each with moderate concavity and asymmetry towards the east, filled by beach deposits and wind accumulations. The skull was photographed and measured, confirming it as an adult male marsh deer (see Supplementary Figures 1–3; Supplementary Tables 1–3).
Radiocarbon dating
Approximately 1 g of turbinate bone was extracted and sent to the Accelerator Mass Spectrometry Laboratory at the University of Arizona for radiocarbon dating. Radiocarbon ages were calculated as weighted averages of combined machine runs, with a small sample correction applied to samples with a carbon mass less than 0.50 mg.
DNA extraction and PCR amplification
Genomic DNA was extracted from tissue samples (50 mg) following González et al. (1998) protocol. We used two set of primers to amplify a partial fragment of the D-loop 364 bp. DNA was amplified using two primers designer with Primer 3 software specifically for marsh deer (BD1F Primer 1 → 5’ACCAATCTCCTAAGACTCAAGG -3’ and BD1R Primer 2 → 5’TGGCTATTGAGTACAAAACTGT-3’ and DLFHB F 5’GCGGCATGGTAATTAAGCTC 3’, DLRHB R 5’CATTACATTATATGCCCCATGC 3’).
The PCR reactions were set up in a type II A laminar flow chamber with UV light in a room separate from that with the real time PCR thermal using negative controls.
PCRs were performed in 10 μL reactions with 1× Sensimix Evagreen PCR Buffer containing a hot-start DNA polymerase (Bioline) for high specificity and sensitivity PCR, 0.3 pM of each primer, and 0.7–1.5 μL of DNA extract. Two replicates were prepared for each sample and PCR negative controls were included for each set of reactions. PCR was done in a Corbett 6000™ (Rotor Gene) thermal cycler using the following conditions: an initial denaturation at 95°C for 10 min, followed by 45 cycles of 15 s at 95°C, 15 s at 55°C and 15 s at 72°C. The final extension was 10 min at 72°C. The HRM step included a 65–95°C ramp increasing at 0.1°C per step with a 2 s pause at each step.
PCR products were purified with DNAClean and Concentrator™ (Zymo Research™) kit and diluted to an equal final concentration using a Nanodrop 1000™ Spectrophotometer (Thermo Fisher Scientific). The amplicons were sent to sequence by the Sanger method to Macrogen service System Sequencer (MACROGEN Inc., Korea).
Bioinformatic analysis
Sequences were aligned and edited using MEGA11 (Tamura et al., 2021) and compared with the nucleotide database available in the National Centre for Biotechnology Information (NCBI) using the BLAST utility. Diversity indices such as haplotype and nucleotide diversities were calculated, and the number of polymorphic sites determined. Evolutionary genetic distances among haplotypes were determined using the Kimura 2-Parameter distance, with nucleotide sequence data analyzed using Maximum Parsimony (MP) and Neighbor Joining (NJ) methods (Kimura, 1980; Bandelt et al., 1999; Rozas et al., 2017).
Haplotype networks
To evaluate the evolutionary relationships among haplotypes and their geographic distribution, a haplotype network of D-loop sequences was constructed using the median-joining network approach implemented on PopART 1.7 (Márquez et al., 2006; Leigh and Bryant, 2015). We downloaded all the available D-loop sequences deposited in GenBank for the marsh deer, representing 17 haplotypes (AY326235–AY326251).
Preliminary analysis and conservation implications
Radiocarbon dating yielded a calibrated age range of 294 years to the present with a 95% probability, confirming the presence of marsh deer in Uruguay over the last three centuries and extending the species’ geographic range, providing valuable insights into its historical distribution.
Furthermore, the analysis of a 364 bp D-loop fragment and comparison with haplotypes from six South American localities including Argentina, Bolivia, and Brazil revealed significant genetic diversity and past population dynamics of marsh deer in the region (Figure 2). The R haplotype displayed a clear connection to Brazilian haplotypes, indicating a population expansion from the ancestral A haplotype, which underwent four mutations in the haplotype network.
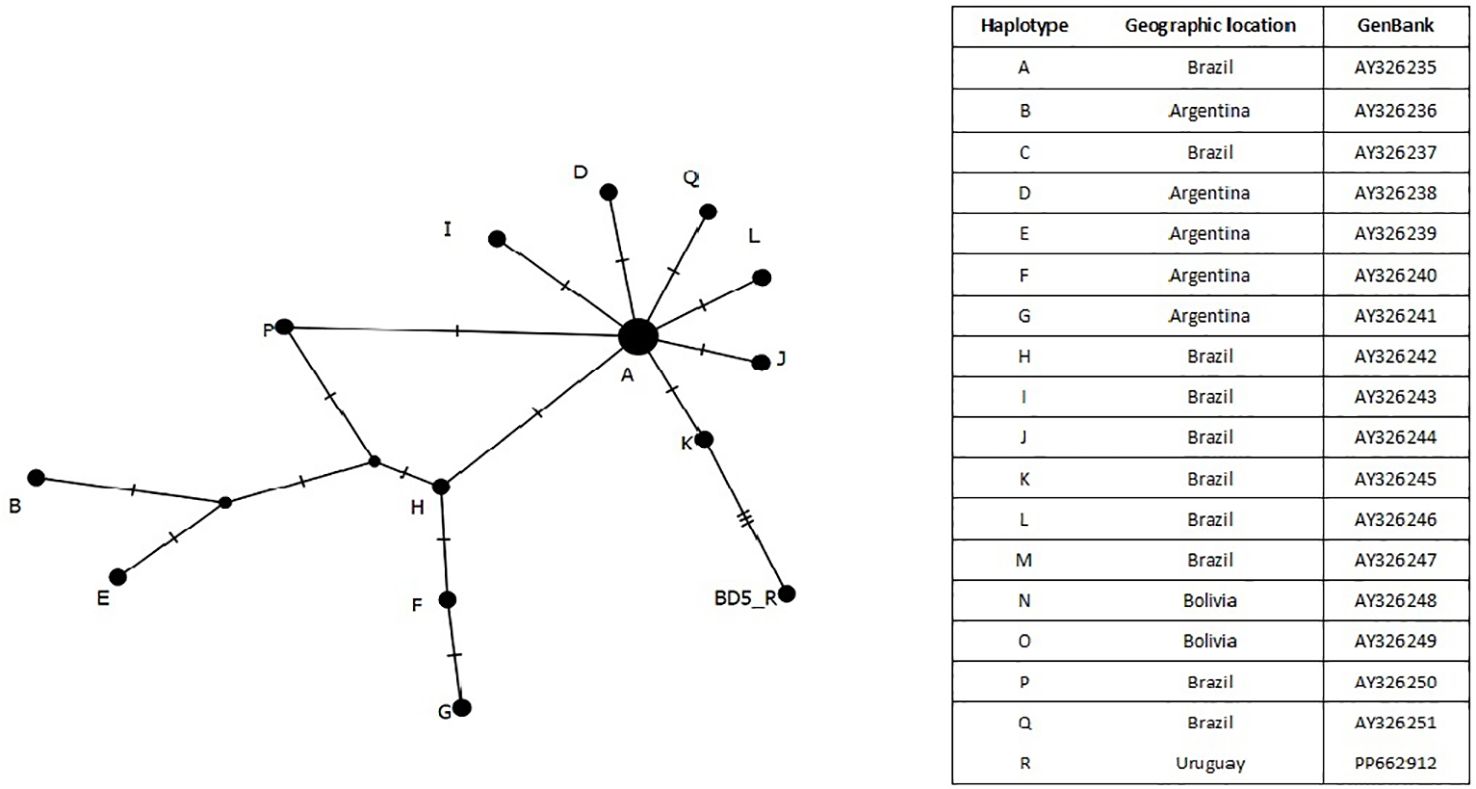
Figure 2 Minimum Spanning Network D-loop haplotypes connection using the median-joining network approach implemented on PopART 1.7 (Leigh and Bryant, 2015). The new record is BD5 belonging to haplotype R submitted to GenBank, PP662912.
Identification of a new haplotype in this skull sample provides compelling evidence for the presence of marsh deer populations in southeast Uruguay. These findings underscore the critical need for habitat restoration as a clear mandate for planning future rewilding strategies.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author contributions
SG: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. MA: Methodology, Writing – original draft. FM: Conceptualization, Formal analysis, Methodology, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. The research was supported by Lóreal-Unesco from Uruguay Women in Science, Sistema Nacional de Investigadores-Agencia Nacional de Investigación e Innovación (SNI-ANII) Comisión Sectorial de Investigación Científica de Universidad de la República CSIC-Udelar I+D 2010 C024, and Programa de Desarrollo de Ciencias Básicas (PEDECIBA).
Acknowledgments
We acknowledge to Alicia Gutiérrez and Jorge Gutiérrez from the ranch La Cambicha who recovered, collected, and donated the skull for research in our laboratory during 2015.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2024.1424867/full#supplementary-material
References
Alcaraz M. A. (2010). Sistemática de los cérvidos (Mammalia, Artiodactyla) del Pleistoceno de las áreas extraandinas de Argentina (La Plata (Ar: Universidad Nacional de la Plata).
Autuori M. P. (1972). Biological data and growth of the first horns of the marsh deer, Blastocerus dichotomus (Illiger 1811) (Cervidae). Zool. Garten 42, 225–235.
Bandelt H. J., Forster P., Röhl A. (1999). Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 16, 37–48. doi: 10.1093/oxfordjournals.molbev.a026036
Beccaceci M. D. (1994). A census of marsh deer in Iberá Natural Reserve, its Argentine stronghold. Oryx 29, 131–134. doi: 10.1017/S0030605300028441
Chimento N., Zuccari J. I., Machetto J. M., Berbach L. (2019). Nuevos restos de ciervos (Cervidae, Mammalia) para el Pleistoceno de la Región Pampeana (Argentina): consideraciones paleobiogeográficas y paleoecológicas. Rev. Bras. Paleontologia 22, 67–85. doi: 10.4072/rbp
Cione A. L., Tonni E. P. (1999). “Biostratigraphy and chronological scale of uppermost Cenozoic in the Pampean Area, Argentina,” in Quaternary Vertebrate Paleontology in South America. Quaternary of South America & Antarctic Peninsula 12. Eds. Tonni E. P., Cione A. L. (A. A. Balkema Publishers, Rotterdam), 23–51.
Cione A. L., Tonni E. P. (2005). Bioestratigrafía basada en mamíferos del Cenozoico superior de la provincia de Buenos Aires, Argentina. Geología y recursos minerales la provincia Buenos Aires 11, 183–200.
González S. (1994). Marsh deer in Uruguay. Population and Habitat Viability Assessment, in Workshop for the Marsh Deer (Blastocerus dichotomus) Vol. Section 4. Ed. IUCN/CBSG (Apple Valley, Minnesota: CBSG publication), 1–6.
González S., Duarte J. M. B. (2020). Speciation, evolutionary history, and conservation trends of Neotropical deer. Mastozoologia Neotropical 27, 37–47. doi: 10.31687/saremMN_SI.20.27.1.05
González E. M., Lessa E. P. (2014). Historia de la mastozoología en Uruguay, in Historia de la mastozoología en Latinoamérica, las Guayanas y el Caribe. Eds. Ortega J., Martínez J. L., Tirira D. G. (Quito y México DF: Editorial Murciélago Blanco y Asociación Ecuatoriana de Mastozoología), 381–404.
González S., Maldonado J. E., Leonard J. A., Vilà C., Barbanti Duarte J. M., Merino M., et al. (1998). Conservation genetics of the endangered Pampas deer (Ozotoceros bezoarticus). Mol. Ecol. 7, 47–56. doi: 10.1046/j.1365-294x.1998.00303.x
Kimura M. (1980). A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 16, 111–120. doi: 10.1007/BF01731581
Leigh J. W., Bryant D. (2015). POPART: full-feature software for haplotype network construction. Methods Ecol. Evol. 6, 1110–1116. doi: 10.1111/2041-210X.12410
Magalhaes R. M., Mello M. G., Bergvist L. P. (1992). Pleistocene cervid from the northeastern region of Brazil. Anais da Academia Bras. Cienc. 64, 149–168.
Márquez A., Maldonado J. E., González S., Beccaceci M. D., Garcia J. E., Duarte J. M. B. (2006). Phylogeography and Pleistocene demographic history of the endangered marsh deer (Blastocerus dichotomus) from the Río de la Plata Basin. Conserv. Genet. 7, 563–575. doi: 10.1007/s10592-005-9067-8
Mauro R. A., Mourão G. M., Coutinho M. E., Silva M. P., Magnusson W. E. (1998). Abundance and distribution of marsh deer Blastocerus dichotomus (Artyodactila: Cervidae) in the Pantanal, Brazil. Rev. Ecología Latino Americana 5, 13–20.
Menegaz A. N. (2000). Los Camélidos y Cérvidos del Cuaternario del Sector Bonaerense de la Región Pampeana. [dissertation thesis] (La Plata (Ar: Universidad Nacional de la Plata).
Moreno F. (2016). La gestión animal en la prehistoria del este de Uruguay: de la economía de amplio espectro al control de animales salvajes. Tessituras 4, 161–187.
Pinder L., Grosse A. P. (1991). Blastocerus dichotomus. Mamm. Species 380, 1–4. doi: 10.2307/3504311
Piovezan U., Tiepolo L. M., Tomas W. M., Duarte J. B., Varela D., Marinho-Filho J. S. (2010). “Marsh deer Blastocerus dichotomus (Illiger, 1815)", in Neotropical Cervidology: biology and medicine of Latin American deer, ed. Duarte J. B., González S. (Jaboticabal: FUNEP/IUCN), 66–76.
Politis G. G., Prates L., Merino M. L., Tognelli M. F. (2011). Distribution parameters of guanaco (Lama guanicoe), pampas deer (Ozotoceros bezoarticus) and marsh deer (Blastocerus dichotomus) in Central Argentina: Archaeological and paleoenvironmental implications. J. Archaeological Sci. 38, 1405–1416. doi: 10.1016/j.jas.2011.01.013
Rodriguez J. M., Bonomo M. (2021). Nuevos análisis zooarqueológicos y tafonómicos del sitio Cerro Tapera Zázquez (Parque Nacional Pre-delta, Argentina). COMECHINGONIA 25, 81–100. doi: 10.37603/2250.7728.v25.n1.30411
Roibal D., González S., Ubilla M. (1996). “Restos de ciervo de los pantanos en El Potrerillo de Santa Teresa,” in IV Jornadas de Zoología del Uruguay (Sociedad Zoológica del Uruguay, Montevideo), 35.
Rozas J., Ferrer-Mata A., Sánchez-Del Barrio J. C., Guirao-Rico S., Librado P., Ramos-Onsins S. E., et al. (2017). DnaSP 6: DNA sequence polymorphism analysis of large datasets mol. Biol. Evol. 34, 3299–3302. doi: 10.1093/molbev/msx248
Spoturno J. J., Oyhantçabal P. B., Loureiro J. (2012). Mapa Geológico del Departamento de Maldonado escala 1/100 000 (Uruguay: Informe Ministerio de Industria y Energia).
Tamura K., Stecher G., Kumar S. (2021). MEGA11: molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 38, 3022–3027. doi: 10.1093/molbev/msab120
Tomas W. M., Beccaceci M. D., Pinder L. (1997). “Cervo-do-pantanal (Blastocerus dichotomus),” in Biologia e conservação de cervídeos sul-americanos. Ed. Duarte J. M. B. (FUNEP, Jaboticabal), 24–40.
Varela D. (2003). Distribución, abundancia y conservación del ciervo de los pantanos (Blastocerus dichotomus) en el Bajo Delta del río Paraná, Provincia de Buenos Aires, Argentina. Licentiate thesis (Buenos Aires: University of Buenos Aires).
Vezzosi R. I., Brunetto E., Zalazar M. C. (2019). El registro fósil de vertebrados del Cuaternario de Santa Fe (Argentina): Primera Actualización. Opera Lillioana 52, 638–670.
Keywords: neotropical deer, mitochondrial DNA, radiocarbon dating, Cervidae, haplotype
Citation: González S, Aristimuño MP and Moreno F (2024) New record in Uruguay of the marsh deer (Blastocerus dichotomus Illiger, 1815) redefines its southern geographic distribution area. Front. Ecol. Evol. 12:1424867. doi: 10.3389/fevo.2024.1424867
Received: 28 April 2024; Accepted: 28 June 2024;
Published: 16 July 2024.
Edited by:
Patricia Amavet, Facultad de Humanidades y Ciencias (UNL), CONICET, ArgentinaReviewed by:
Brenda Ferrero, National Scientific and Technical Research Council (CONICET), ArgentinaSolana Tabeni, CONICET Mendoza, Argentina
Copyright © 2024 González, Aristimuño and Moreno. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Susana González, sgonzalez@iibce.edu.uy
†ORCID: Susana González, orcid.org/0000-0001-6470-6182