 Jeremy DeSilva1*
Jeremy DeSilva1* Luke Fannin1,2
Luke Fannin1,2 Isabelle Cheney1
Isabelle Cheney1 Alexander Claxton3
Alexander Claxton3 Iulian Ilieş4
Iulian Ilieş4 Jessica Kittelberger1
Jessica Kittelberger1 Jeff Stibel5
Jeff Stibel5 James Traniello6
James Traniello6- 1Department of Anthropology, Dartmouth College, Hanover, NH, United States
- 2Ecology, Evolution, Environment and Society, Dartmouth College, Hanover, NH, United States
- 3Anatomy and Cell Biology, Oklahoma State University, Center for Health Sciences, Tulsa, OK, United States
- 4Healthcare Systems Engineering Institute, Department of Mechanical and Industrial Engineering, Northeastern University, Boston, MA, United States
- 5Natural History Museum, Los Angeles, CA, United States
- 6Department of Biology, Boston University, Boston, MA, United States
Human brain reduction from the Late Pleistocene/Holocene to the modern day is a longstanding anthropological observation documented with numerous lines of independent evidence. In a recent study (DeSilva et al., 2021; Front. Ecol. Evol.), we analyzed a large compilation of fossil and recent human crania and determined that this reduction was surprisingly recent, occurring rapidly within the past 5,000 to 3,000 years of human history. We attributed such a change as a consequence of population growth and cooperative intelligence and drew parallels with similar evolutionary trends in eusocial insects, such as ants. In a reply to our study, Villmoare and Grabowski (2022; Front. Ecol. Evol.) reassessed our findings using portions of our dataset and were unable to detect any reduction in brain volume during this time frame. In this paper, responding to Villmoare and Grabowski’s critique, we reaffirm recent human brain size reduction in the Holocene, and encourage our colleagues to continue to investigate both the timing and causes of brain size reduction in humans in the past 10,000 years.
Introduction
Our analysis of human brain evolution (DeSilva et al., 2021) was based on robust prior research demonstrating that human brains decreased in volume in the Late Pleistocene or Holocene. This recent reduction has been documented by numerous researchers for nearly 90 years across diverse populations globally (Figure 1; von Bonin, 1934; Weidenreich, 1946; Tobias, 1971; Schwidetzky, 1976; Wiercinski, 1979; Beals et al., 1984; Henneberg, 1988, 1998, 2004; Brown, 1992; Henneberg and Steyn, 1993, 1995; Ruff et al., 1997; Brown and Maeda, 2004; Wu et al., 2007; Bailey and Geary, 2009; Hawks, 2011; Balzeau et al., 2013; Bednarik, 2014; Liu et al., 2014; Stibel, 2021, 2023). The question we asked, then, was not whether modern human brain volume was smaller than that of Pleistocene Homo sapiens, but when this reduction occurred. Addressing this question, we could proceed to infer why an organ critical for human survival would decrease in size.
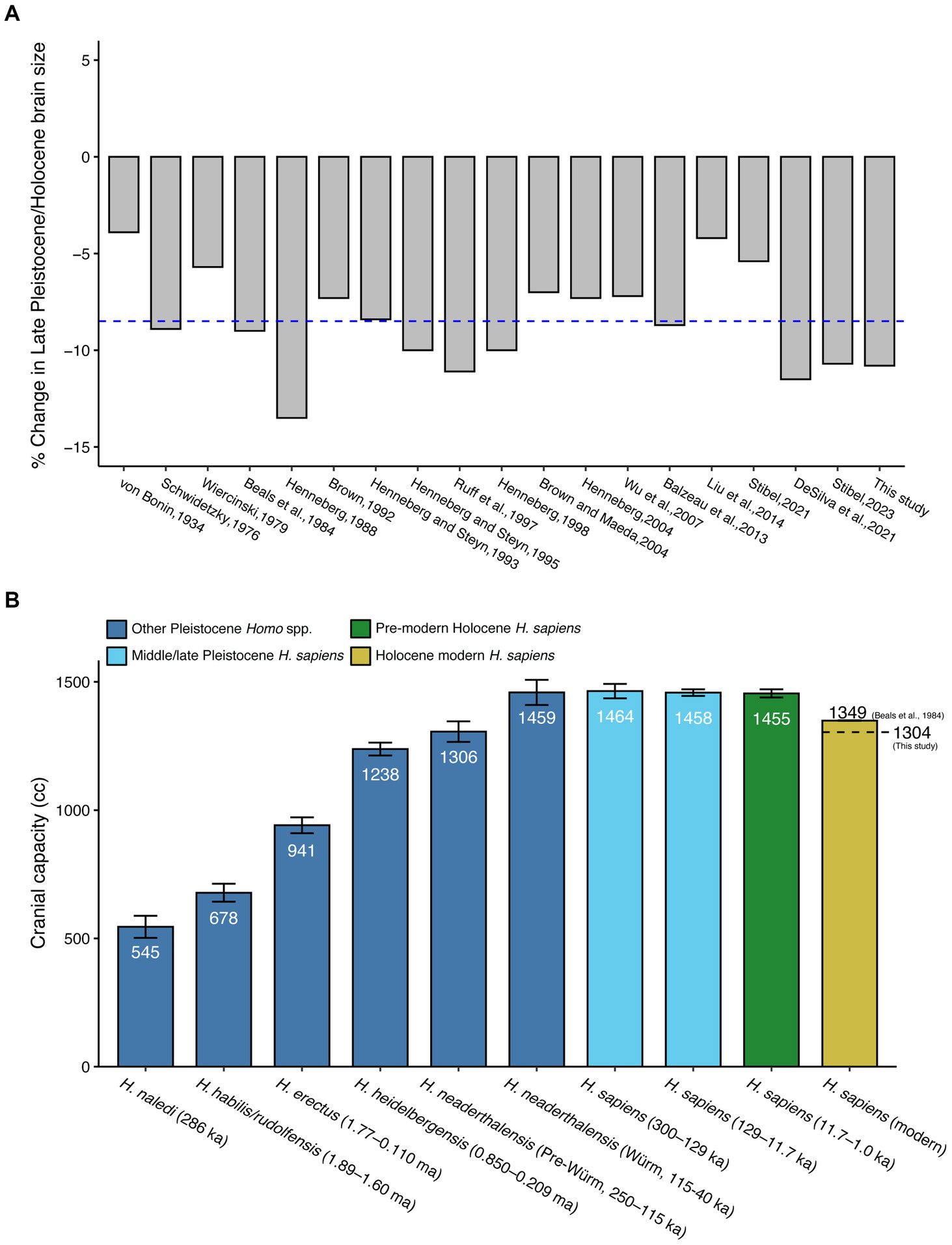
Figure 1. (A) Summary of results of published studies in the last ca. 90 years reporting a decrease in human brain size in the late Pleistocene/Holocene (N = 19). The average reported decrease across all studies is 8.5%, illustrated by the blue dotted line. Percent change was used as published or, if not available in-text, calculated from the average brain volumes published in the study that demarcated the decrease. (B) Average brain size (cranial capacity) across members of the genus Homo during the Pleistocene and Holocene epochs. Bars are means, with whiskers representing ± one standard error. The last, yellow bar includes both the global mean cranial capacity for modern H. sapiens calculated by Beals et al. (1984), and the alternative modern H. sapiens mean cranial capacity (dotted line), compiled from recent anatomical and archaeological samples in the current study.
Our original findings that brain size has reduced surprisingly recently (~5,000–3,000 years ago) is consistent with previous research and led to our hypothesis that population growth and knowledge specialization associated with cooperative intelligence led to a decrease in the volume of the brain, which is energetically expensive to develop and operate (Aiello and Wheeler, 1995; Navarrete et al., 2011; Heldstab et al., 2022). We drew parallels with patterns of brain evolution in ants, an entirely eusocial clade in which workers of different species have also undergone selection for both increased and reduced brain size in relation to higher levels of social complexity (Traniello et al., 2022). In ants, the scaling of brain size to body size and brain mosaicism vary with the behavioral and/or cognitive demands of task performance and division of labor, characteristics that are likely to impact brain evolution across diverse taxa, including humans.
Yet Villmoare and Grabowski (2022) recently argued that the dataset from which we based our findings was inadequate for the question being asked. Furthermore, they reassessed our study using portions of our dataset and were unable to detect any reduction in brain volume. Based on their analysis, they conclude that “human brain size has been remarkably stable over the last 300 ka. Thus, hypotheses of recent change are not supported by the evidence.” If these authors are correct, human brain reduction—an established fact for almost a century (Figure 1)—did not occur. In this paper, responding to Villmoare and Grabowski’s critique, we demonstrate that our revised dataset is sufficient for testing trends in brain volume through time and reaffirm recent human brain size reduction.
Recent human brain reduction: what does prior research tell us?
Recent (i.e., Late Pleistocene or Holocene) human brain reduction is not a new idea (Figure 1; Supplementary Table S1) and is not as controversial as Villmoare and Grabowski (2022) suggested. Von Bonin (1934) wrote, “there is a definite indication of a decrease at least in Europe within the last 10,000 or 20,000 years” in the human brain. Noted anthropologists Franz Weidenreich (1946) and Philip Tobias (1971) observed that modern human brain volumes are on average smaller than Pleistocene hominin crania. Schwidetzky (1976) found a decrease in the estimated number of “extraneurons” (following Jerison, 1963) since the Neolithic in parts of Europe. Wiercinski (1979) used linear measurements on 20 different populations in Europe, Africa, Asia, and Australia and reported a reduction in cranial dimensions in 17 of them, concluding that human brain reduction was a post-Aurignacian global phenomenon.
Using a dataset of 5,288 cranial capacities from 122 distinct global populations, Beals et al. (1984) detected a recent decrease in brain size and wrote, “we consider de-encephalization through the last 100,000 years as confirmed.” Henneberg (1988) evaluated primarily linear measurements taken on nearly 13,000 skulls and concluded that there had been a 10–17% decrease from the Mesolithic to modern times. Most of these data were obtained on specimens from Europe with additional skulls from northwest Africa and west Asia. Henneberg and Steyn (1993, 1995) identified a similar decrease in brain size in samples from sub-Saharan Africa and Japan. By 2004, Henneberg’s global study had exceeded 14,000 samples from 15 thousand years ago (ka) to modern day. He concluded that “Cranial capacity decreased by some 100–150 mL during the Holocene, with most of this decrease occurring during the last 3 Ka.” We unfortunately neglected to cite Henneberg (2004) in our original paper and correct the oversight here. We find it compelling that the 3 ka date is consistent with what we found using a different methodology and a different sample (DeSilva et al., 2021).
Using a sample from East Asia and Australia, Brown (1992) reported a recent 10% reduction in cranial capacity. Ruff et al. (1997) used data from Beals et al. (1984) and samples from the Pecos Pueblo (New Mexico, United States) archaeological site and found a Late Pleistocene decrease in brain volume. Brown and Maeda (2004) reported a decrease in the size of the cranium in Chinese skulls from the Neolithic to today—with an accelerating rate of change after 3,500 years before present (BP)—a finding replicated using a different dataset by Liu et al. (2014). Wu et al. (2007) took linear measurements on 718 male skulls from the Holocene of China and reported a 7.2% reduction in calculated cranial volume from the Bronze age to the present. In their study of the Cro-Magnon H. sapiens cranium, Balzeau et al. (2013) state that “a decrease in absolute endocranial size since the Upper Pleistocene is noticeable in H. sapiens.” They based this finding on 15 Pleistocene crania from 25–92 ka and 99 modern human crania from Europe, Africa, Asia, the Pacific islands, and North America. Stibel (2021) found a 5% decrease in brain volume from Pleistocene H. sapiens to modern people. In an updated paper, Stibel (2023) reported that brain size in Late Pleistocene (50–12 ka BP) H. sapiens was 10.7% larger than in Holocene humans (12 ka BP- present), a statistically significant difference (p < 0.0001, t-test).
We recognize that the history of brain science is rife with problematic studies biased by racist and sexist objectives. Furthermore, “brain size” is difficult to objectively measure and different investigators have determined brain mass and/or cranial capacity using distinct methods (see review in Tobias, 1970). Most studies report summary statistics (e.g., Ho et al., 1980) while very few report data from individuals (e.g., Bischoff, 1880). Furthermore, certain regions of the world are overrepresented (e.g., Europe) while there is little data for other human populations. Despite these limitations, independent of measurement technique, brain volume reductions have been consistently reported by researchers for over three-quarters of a century on skulls representing populations globally (Figure 1). It is difficult to accept on scientific grounds that all of these studies are in error.
How big is the average human brain?
Villmoare and Grabowski (2022) considered our average reported brain volume for recent modern humans (1,297 cc in DeSilva et al., 2021; 1,304 ± 154 cc in this study) to be lower than other reports showing roughly 1,400 cc, citing Beals et al. (1984), Henneberg (1988), Ruff et al. (1997), and De Sousa and Cunha (2012) as support. However, in the very papers they cite, modern human cranial capacities are less than 1,400 cc on average. Beals et al. (1984) sampled 5,288 crania from 122 different ethnic groups and reported a cranial capacity of 1,349 ± 78 cc. Ruff et al. (1997) supplemented the value reported in Beals et al. (1984) with the Pecos archaeological sample averaging 1,308 ± 123 cc (N = 29). Henneberg (1988) used mostly linear measurements to calculate cranial capacities. Where he used directly measured cranial capacities, the weighted average is 1,387 cc (N = 245). De Sousa and Cunha (2012) reported an average of 1,392 cc (N = 551), though these values are converted from brain weights measured in 20–30 year-olds from Dekaban and Sadowsky (1978). However, the entire Dekaban and Sadowsky (1978) adult dataset (N = 3,399) indicates an average brain size of 1,334.5 cc ± 205.9. Thus, using identical sources referenced by Villmoare and Grabowski (2022), the range never exceeds 1,400 cc and is instead 1,308–1,392 cc with a weighted average of 1,345 cc (N = 8,961). Independently, Tobias (1971) reported an identical average of 1,345 cc from “thousands” of measurements.
While it can be problematic to convert brain mass (g) to cranial capacity (cc) (see Tobias, 1970), two equations permit direct comparison. Cranial capacity can be converted from brain weight (g) using Hofman (1983)’s equation:
This equation is derived from brain volume (cc) = cranial capacity (cc) * 0.92 and the specific gravity of human brain tissue = 1.036 g/cm3. Ruff et al. (1997) established the equation:
Here, we averaged the results of the two methods which were on average only 1–2% different from one another. For those studies with an equal sex representation, brain size averages between 1,335 ± 206 cc (Dekaban and Sadowsky, 1978; N = 3,399) and 1,344 ± 137 cc (Ho et al., 1980; N = 1,261). Furthermore, Grabowski (2016) reported an average brain mass of 1,299 g (Table 2, p. 180) using data from Bischoff (1880). Converting this value to cc using the equations in Hofman (1983) and Ruff et al. (1997) yields an average of 1,350 cc.
These values, however, almost certainly overestimate the average adult human brain size, given the disproportionate representation of larger-bodied European males in the samples and the known scaling relationship between brain and body size (Ruff et al., 1997; Hawks, 2011; Grabowski, 2016). When smaller-bodied populations from East Asia, sub-Saharan Africa, and Australia are compiled (data from Henneberg and Steyn, 1993; Brown and Maeda, 2004), the weighted sample mean is 1,300 cc (N = 768). Therefore, we disagree that our original calculated value of ~1,300 cc for the average human cranial capacity is too low—being slightly larger than Albert Einstein’s (~1,291 cc converted from grams; Witelson et al., 1999), and slightly smaller than Walt Whitman’s (~1,317 cc converted from grams; Spitzka, 1907). When cranial capacity averages and standard deviations are appropriately weighted by continental populations (Source: https://www.statista.com/statistics/237584/distribution-of-the-world-population-by-continent/), we calculate an average of 1,328 ± 145 cc. Using estimated pre-colonial populations from the year 1,500, we arrive at a weighted average of 1,331 ± 153 cc. Given these data, it is unclear how the commonly reported overestimate of >1,400 cc has entered our collective knowledge.
Critical analysis of human brain volume datasets: statistical approaches
Given the literature cited above, we naturally did not explore whether brain volumes had decreased, as that had been clearly established in multiple previous studies, but estimated when. To answer this question, we employed a changepoint analysis using the segmented package in R (Muggeo, 2008; details in DeSilva et al., 2021), which led us to compile raw cranial capacities for fossil crania spanning the past 10 million years (Ma), along with a large modern human sample. Compiling these data was not difficult for Miocene and Plio-Pleistocene hominids because endocranial volumes are standard measurements reported for skulls discovered in paleoanthropological or archaeological contexts (e.g. Holloway et al., 2002). Using this dataset, we found statistically significant changes in the rates of hominin endocranial volume change at ~2 [(95% confidence interval (CI): 2.0-2.3) and ~ 1.5 (95% CI: 1.2-1.8)] Ma, findings consistent with previous work on hominin brain size evolution during these periods (Antón et al., 2014; Grabowski, 2016). We disagree with Villmoare and Grabowski (2022) that important crania from diverse taxa such as Rudapithecus, Australopithecus, and Homo erectus––which we included in our model to contextualize the temporal dynamics of hominin brain evolution before the evolution of modern humans––are not relevant in such discussions (see, for instance, Begun, 2010; Gowlett et al., 2012; Antón et al., 2014; Almécija et al., 2021).
Our use of this particular changepoint analysis was intentional, as it allowed for estimates of breakpoint times and slopes in a large data set that otherwise lacked uniform sampling from each time slice, a widespread issue for most paleoanthropological datasets. Rather, this analysis, implemented by fitting a piecewise linear regression to the data, relies primarily on standard regression assumptions, as pointed out by Villmoare and Grabowski (2022)—i.e., normality and independence of residuals, and homoscedasticity—to generate estimates of slopes and breakpoint locations. Our changepoint approach, while unconventional based on the literature cited in Villmoare and Grabowski (2022), is nevertheless common and consistent with other investigations concerned with estimating the timing of key events in the paleoanthropological record using unbinned, raw time-series data (e.g., Faith et al., 2018; Wynn et al., 2020). For our own analysis, the majority of our time series followed these a priori assumptions; in turn, Villmoare and Grabowski (2022) produced estimates for the first two breakpoints in the time series (2.1 and 1.3 Ma) that fell within the 95% CI initially reported in Table 1 of DeSilva et al. (2021).
Yet for recent humans, we were challenged to incorporate sufficient samples to accurately represent modern variation without skewing our data, a constraint we failed to address sufficiently according to Villmoare and Grabowski (2022). They rightly point out that the Holocene portion of our dataset is skewed primarily towards modern humans, an unavoidable taphonomic bias and limitation in our original model that may skew our estimate of when brain reduction occurred towards more recent periods, and, in the worst case, obscure additional, earlier change points. Yet we disagree with their proposed solution: consolidating the individual cranial data into means representing identical temporal slices of 100 years (see Figures 2 and 3 of Villmoare and Grabowski, 2022). Pooling irregularly sampled data into equal-sized time bins runs the risk of diluting trends or introducing spurious ones, depending on data density across time, and particularly on the timing of outlier measurements: e.g., a single outlier data point in a sparsely sampled period would be given the same importance as hundreds of data points from a well-sampled period. Better, less sensitive options include weighted regression models (individual points are assigned importance weights inversely proportional to data density), bootstrapping or resampling (oversampling with replacement from time periods with few data, and/or undersampling without replacement from intervals with high data density), or even log-transforming time measurements (assuming trend direction and changes therein are more important than trend type).
We are not opposed to binning the data to improve a priori statistical assumptions (cf. Figure 2 of this study), but we also find it problematic to bin data arbitrarily in such a way that is uncritical of the broader question being asked: has there been a significant change in average human brain size since the start of the Holocene? We contend that the reason Villmoare and Grabowski (2022) did not find a Holocene decrease in cranial capacity with their consolidated dataset of means is because the time-averaging process effectively removed the crucial variability in cranial capacity found in the period of interest, i.e., the last 10,000 years. Indeed, when such variability is binned more appropriately (e.g., by geological time periods defined in part by global climate changes) and incorporated into simpler statistical analyses (e.g., t-tests; see below analyses with updated data), a strong and significant decrease in modern human brain size across the Holocene boundary is detected (Figure 2), reaffirming our original conclusions (DeSilva et al., 2021).
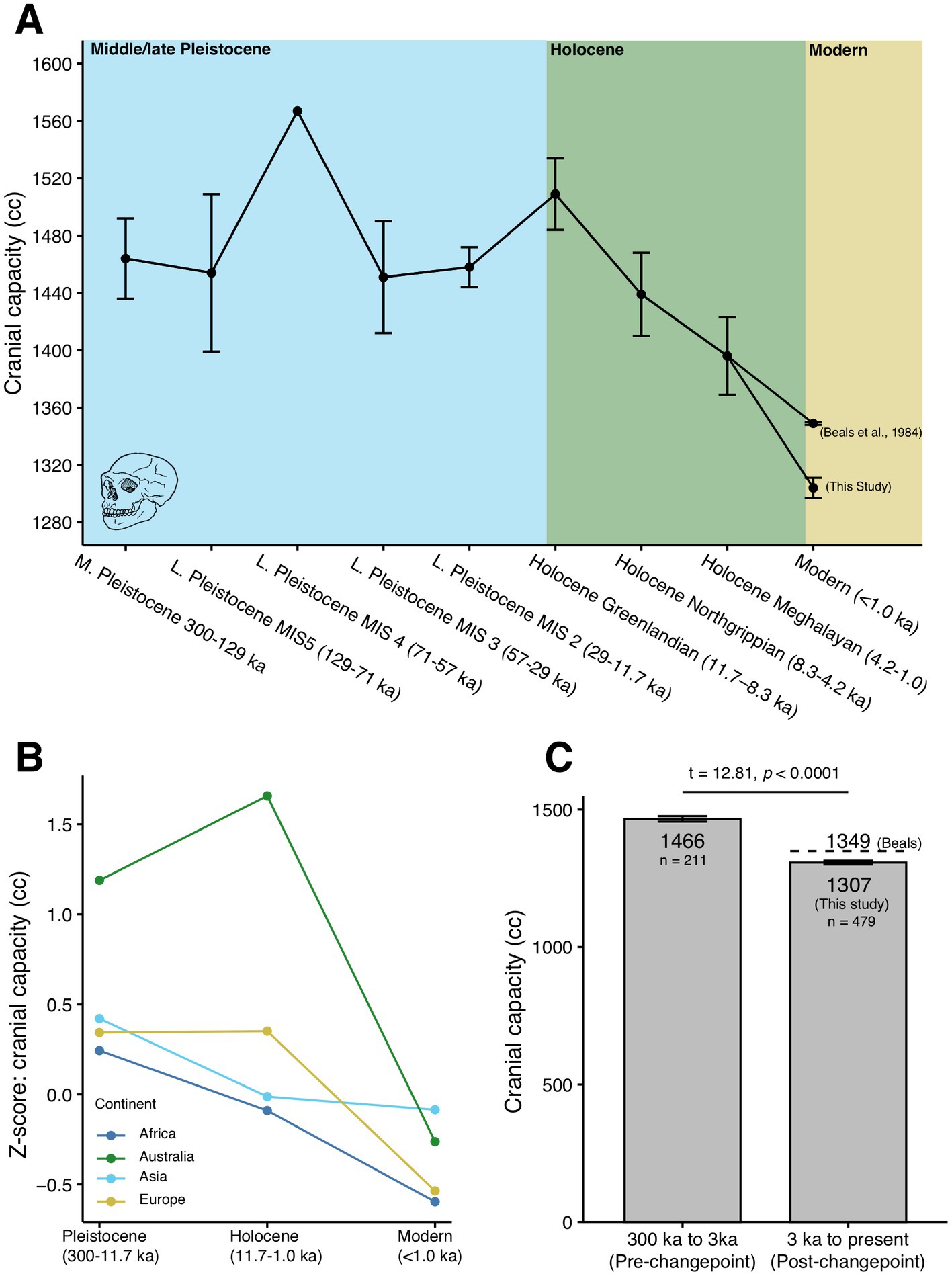
Figure 2. Dynamics of brain size reduction (cranial capacity) in H. sapiens during the Pleistocene and Holocene. (A) Changes in H. sapiens cranial capacity over the past 300,000 years, subdivided by geological epochs and climatic milestones, with recent modern samples (<1.0 ka) subdivided from the rest of the Holocene. Means represent average cranial capacity, whiskers are ± one standard error. There is only a single cranial capacity reported for MIS 4. (B) Changes in the average H. sapiens cranial capacity over the past 300,000 years, subdivided by major continental landmasses. Average cranial capacities are presented here as Z-scores (i.e., standard-deviation units). (C) Average cranial capacity in H. sapiens, before and after the original reduction date of 3,000 years proposed by DeSilva et al. (2021). An average reduction in brain size of 159 cc (using our modern estimate) or 117 cc (using the Beals et al. (1984) modern estimate) after 3,000 years is illustrated. Whiskers are ± one standard error.
A related critique by Villmoare and Grabowski (2022) was our use of questionable modern cranial samples from the collection of Samuel Morton at the Penn Museum. We agree this is a problematic dataset because it has been used to promote false and dangerous ideas of white supremacy (Morton Collection Committee, 2021; Mulligan et al., 2022). Eliminating these data from the present analysis had no appreciable impact on our reported brain volume for modern humans (Supplementary Table S2). In lieu of the Morton data, we have added modern cranial capacity data from the Terry Collection (N = 94; VanSickle et al., 2020 via lynncopes.com) and India (N = 50; Manjunath, 2002).
Villmoare and Grabowski (2022) also noted that our dataset contained few individuals from a key time-period, 1-5 ka. We added 13 individuals from this time range from Henneberg and Steyn (1993) and Stibel (2021). Additionally, we used the millet seed method to measure cranial capacities of two skulls from the South African archaeological site Byneskranskop (~3.1 ka; Sealy, 2006) and another individual from the older Plattenberg Bay site (~7 ka; Sealy, 2006). We also revised the age of the Pecos Pueblo population to 500 years BP, assuming most of the individuals derive from the Glaze V period. Finally, we removed juvenile Neanderthals (N = 4), which were inadvertently included in our dataset, and eliminated one entry of the Liujiang skull, which mistakenly appeared twice. The revised cranial volume catalog is now available as a supplementary Excel file.
Brain reduction in the Holocene
Analysis of our modified dataset shows that Holocene brain reduction remains robust (Supplementary Table S2; Figure 2). On the broadest scale, Pleistocene (300 ka-11.7 ka) H. sapiens brains average 1,458 ± 140 cc (N = 136). This is effectively identical to the average Neanderthal brain (1,459 ± 182 cc; N = 14) from the Würm period (<115 ka; DeSilva, 2018). There is no directional change in brain volume in H. sapiens throughout the Pleistocene whether the data are consolidated in consistent time intervals (Villmoare and Grabowski, 2022), or divided by geological stages of the Pleistocene epoch (Supplementary Table S2). This pattern of stasis in H. sapiens brain volume changes quite abruptly and obviously in the later part of the Holocene (Supplementary Table S2; Figures 2A,B). We agree with Villmoare and Grabowski (2022) that more samples from this time-period will be valuable for future study. However, we argue that even without any samples from the Holocene, one could identify a change in brain size by simply comparing the chronological “bookends” of the Pleistocene and today. In fact, instead of a change point analysis or data consolidation, a simple t-test can effectively evaluate if human brains today differ in volume from humans in the Pleistocene. Using Welch’s t-test, the difference between Pleistocene and Holocene human cranial capacities in our dataset is significant (t = 9.15, p < 0.0001), a result similar to that found in Stibel (2023). Even more granularly, if we were to look at the changes in H. sapiens cranial capacity before and after the originally proposed change point (3,000 years) in DeSilva et al. (2021), a t-test reveals a significant decrease in human cranial capacity post 3 ka (t = 12.81, p < 0.0001; Figure 2C).
Because Villmoare and Grabowski (2022) suggest our initial study underestimated modern human brain volumes, we repeated the analysis with brain weight data (N = 3,399) from Dekaban and Sadowsky (1978)—converted to cranial capacities—and found the same differences (t = 9.83, p < 0.0001). The Beals et al. (1984) dataset (N = 5,288), which compiles a larger global sample of cranial capacities and is therefore preferable, also reveals significant differences (t = 9.04, p < 0.0001, Welch’s t-test). Therefore, independent of the modern dataset used (e.g., Dekaban and Sadowsky, 1978; Beals et al., 1984; this study), it is clear that there has been, on average, a 100–150 cc reduction in brain volume (Figure 2C). These data are consistent with Henneberg (2004), who similarly found a 100–150 mL reduction in brain volume during the Holocene using measurements on 14,000 crania. These data further mirror a widely recognized Holocene reduction in body size (Ruff et al., 1997; Stibel, 2023) that would be difficult to reconcile with Villmoare and Grabowski’s (2022) proposed stasis in brain size.
On finer scales, similar magnitudes of Holocene brain reduction have been documented regionally and across latitudes (e.g., Henneberg and Steyn, 1993; Liu et al., 2014; Stibel, 2023). In other words, human brain volume has decreased by a standard deviation in the last 10,000 years, whether examined locally or globally (Figures 2A,B). It is probable that brain reduction occurred at different rates in different areas during the Holocene—a point also noted in the critique of our initial study. But unlike Villmoare and Grabowski (2022), we view these regional dynamics as integral components of an overarching global reduction in human brain size that defined the last 10,000 years. Holocene brain reduction is not a uniquely human phenomenon; rather, a widespread pattern of brain size reduction is also found in domestic and human-associated mammals during the last 10,000 years—ranging from large hooved taxa like cows, horses, llamas, and pigs to rodents like rats and guinea pigs (Balcarcel et al., 2021a,b, 2022). These findings, combined with our own analyses, speak to the profound effect that the Holocene agricultural revolution and the subsequent rise of complex societies had on the trajectory of human and, more broadly, mammalian brain evolution.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
JD, LF, and JT conceived the study and wrote the manuscript. JD, AC, IC, and JS accumulated the hominin brain dataset. JK and JD collected original data in South Africa. LF and II contributed to the statistics. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by National Science Graduate Research Fellowship to LF (no. 1840344) and National Science Foundation grant to JT (no. 1953393).
Acknowledgments
The authors are grateful to Wendy Black, Wilhelmina Seconna and the Iziko Museum advisory board for the opportunity to collect cranial capacity data on three individuals in their curatorial care in December 2016. Thanks also to M. Henneberg for insightful comments on our 2021 paper and to B. Villmoare and M. Grabowski for productive scientific discourse. This paper was improved thanks to the thoughtful suggestions of a reviewer.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1191274/full#supplementary-material
References
Aiello, L. C., and Wheeler, P. (1995). The expensive-tissue hypothesis: the brain and the digestive system in human and primate evolution. Curr. Anthropol. 36, 199–221. doi: 10.1086/204350
Almécija, S., Hammond, A. S., Thompson, N. E., Pugh, K. D., Moyà-Solà, S., and Alba, D. M. (2021). Fossil apes and human evolution. Science 372:eabb4363. doi: 10.1126/science.abb4363
Antón, S. C., Potts, R., and Aiello, L. C. (2014). Evolution of early Homo: an integrated biological perspective. Science 345:1236828. doi: 10.1126/science.1236828
Bailey, D. H., and Geary, D. C. (2009). Hominid brain evolution. Hum. Nat. 20, 67–79. doi: 10.1002/cne.900590102
Balcarcel, A. M., Geiger, M., Clauss, M., and Sánchez-Villagra, M. R. (2022). The mammalian brain under domestication: discovering patterns after a century of old and new analyses. J. Exp. Zool. B Mol. Dev. Evol. 338, 460–483. doi: 10.1002/jez.b.23105
Balcarcel, A. M., Sánchez-Villagra, M. R., Segura, V., and Evin, A. (2021a). Singular patterns of skull shape and brain size change in the domestication of south American camelids. J. Mammal. 102, 220–235. doi: 10.1093/jmammal/gyaa135
Balcarcel, A. M., Veitschegger, K., Clauss, M., and Sánchez-Villagra, M. R. (2021b). Intensive human contact correlates with smaller brains: differential brain size reduction in cattle types. Proc. R. Soc. B 288:20210813. doi: 10.1098/rspb.2021.0813
Balzeau, A., Grimaud-Hervé, D., Détroit, F., Holloway, R. L., Combès, B., and Prima, S. (2013). First description of the Cro-Magnon 1 endocast and study of brain variation and evolution in anatomically modern Homo sapiens. Bull. Mém. Soc. Anthropol. Paris 25, 1–18. doi: 10.1007/s13219-012-0069-z
Beals, K. L., Smith, C. L., and Dodd, S. M. (1984). Brain size, cranial morphology, climate, and time machines. Curr. Anthropol. 25, 301–330. doi: 10.1086/203138
Bednarik, R. G. (2014). Doing with less: hominin brain atrophy. Homo 65, 433–449. doi: 10.1016/j.jchb.2014.06.001
Begun, D. R. (2010). Miocene hominids and the origins of African apes and humans. Annu. Rev. Anthropol. 39, 67–84. doi: 10.1146/annurev.anthro.012809.105047
Brown, P. (1992). Recent human evolution in East Asia and Australasia. Philos. Trans. R. Soc. London B Biol. Sci. 337, 235–242. doi: 10.1098/rstb.1992.0101
Brown, P., and Maeda, T. (2004). Post-Pleistocene diachronic change in east Asian facial skeletons: the size, shape and volume of the orbits. Anthropol. Sci. 112, 29–40. doi: 10.1537/ase.00072
de Sousa, A., and Cunha, E. (2012). Hominins and the emergence of the modern human brain. Prog. Brain Res. 195, 293–322. doi: 10.1016/B978-0-444-53860-4.00014-3
Dekaban, A. S., and Sadowsky, D. (1978). Changes in brain weights during the span of human life: relation of brain weights to body heights and body weights. Ann. Neurol. 4, 345–356. doi: 10.1002/ana.410040410
DeSilva, J. M. (2018). Comment on “the growth pattern of Neandertals, reconstructed from a juvenile skeleton from El Sidrón (Spain)”. Science 359:eaar3611. doi: 10.1126/science.aar3611
DeSilva, J. M., Traniello, J. F. A., Claxton, A. G., and Fannin, L. D. (2021). When and why did human brains decrease in size? A new change-point analysis and insights from brain evolution in ants. Front. Ecol. Evol. 9:742639. doi: 10.3389/fevo.2021.742639
Faith, J. T., Rowan, J., Du, A., and Koch, P. L. (2018). Plio-Pleistocene decline of African megaherbivores: no evidence of ancient hominin impacts. Science 362, 938–941. doi: 10.1126/science.aau2728
Gowlett, J., Gamble, C., and Dunbar, R. (2012). Human evolution and the archaeology of the social brain. Curr. Anthropol. 53, 693–722. doi: 10.1086/667994
Grabowski, M. (2016). Bigger brains led to bigger bodies?: the correlated evolution of human brain and body size. Curr. Anthropol. 57, 174–196. doi: 10.1086/685655
Hawks, J. (2011). Selection for smaller brains in Holocene human evolution. arXIV [Epub ahead of preprint]. doi: 10.48550/arXiv.1102.5604
Heldstab, S. A., Isler, K., Graber, S. M., Schuppli, C., and van Schaik, C. P. (2022). The economics of brain size evolution in vertebrates. Curr. Biol. 32, R697–R708. doi: 10.1016/j.cub.2022.04.096
Henneberg, M. (1998). Evolution of the human brain: is bigger better? Clin. Exp. Pharmacol. Physiol. 25, 745–749. doi: 10.1111/j.1440-1681.1998.tb02289.x
Henneberg, M. (2004). The rate of human morphological microevolution and taxonomic diversity of hominids. Stud. Hist. Anthropol. 2006, 49–59.
Henneberg, M., and Steyn, M. (1993). Trends in cranial capacity and cranial index in Subsaharan Africa during the Holocene. Am. J. Hum. Biol. 5, 473–479. doi: 10.1002/ajhb.1310050411
Henneberg, M., and Steyn, M. (1995). Diachronic variation of cranial size and shape in the Holocene: a manifestation of hormonal evolution? Riv. Antropol. 73, 159–164.
Ho, K. C., Roessmann, U., Straumfjord, J. V., and Monroe, G. (1980). Analysis of brain weight. I. Adult brain weight in relation to sex, race, and age. Archives of Pathology and Laboratory Medicine 104, 635–639. doi: 10.1038/nature10629
Hofman, M. A. (1983). Encephalization in hominids: evidence for the model of punctuationalism. Brain Behav. Evol. 22, 102–117. doi: 10.1159/000121511
Holloway, R. L., Broadfield, D. C., Yuan, M. S., Schwartz, J. H., and Tattersall, I. (2002). The human fossil record, brain Endocasts--the Paleoneurological evidence (Vol. 3). Hoboken, NJ: John Wiley & Sons.
Liu, C., Tang, Y., Ge, H., Wang, F., Sun, H., Meng, H., et al. (2014). Increasing breadth of the frontal lobe but decreasing height of the human brain between two Chinese samples from a Neolithic site and from living humans. Am. J. Phys. Anthropol. 154, 94–103. doi: 10.1002/ajpa.22476
Manjunath, K. Y. (2002). Estimation of cranial volume in dissecting room cadavers. J. Anat. Soc. India 51, 168–172.
Morton Collection Committee . (2021). Report on evaluation phase. Penn Museum, https://www.penn.museum/sites/morton/MortonCommitteeReport.pdf p. 1–5
Muggeo, V. M. (2008). Segmented: an R package to fit regression models with broken-line relationships. R news 8, 20–25.
Mulligan, C. J., Boyer, D. M., Turner, T. R., Delson, E., and Leonard, W. R. (2022). Data sharing in biological anthropology. Am. J. Biol. Anthropol. 178, 26–53. doi: 10.1002/ajpa.24499
Navarrete, A., Van Schaik, C. P., and Isler, K. (2011). Energetics and the evolution of human brain size. Nature 480, 91–93. doi: 10.1038/nature10629
Ruff, C. B., Trinkaus, E., and Holliday, T. W. (1997). Body mass and encephalization in Pleistocene Homo. Nature 387, 173–176. doi: 10.1038/387173a0
Schwidetzky, I. (1976). Postpleistocene evolution of the brain? Am. J. Phys. Anthropol. 45, 605–611. doi: 10.1002/ajpa.1330450327
Sealy, J. (2006). Diet, mobility, and settlement pattern among Holocene hunter-gatherers in southernmost Africa. Curr. Anthropol. 47, 569–595. doi: 10.1086/504163
Spitzka, E. A. (1907). A study of the brains of six eminent scientists and scholars belonging to the American anthropometric society, together with a description of the skull of professor E.D. cope. Trans. Am. Philos. Soc. 21, 175–308. doi: 10.2307/1005434
Stibel, J. M. (2021). Decreases in brain size and encephalization in anatomically modern humans. Brain Behav. Evol. 96, 64–77. doi: 10.1159/000519504
Stibel, J. M. (2023). Climate change influences brain size in humans. Brain Behav. Evol. 98, 93–106. doi: 10.1159/000528710
Tobias, P. V. (1970). Brain-size, grey matter and race—fact or fiction? Am. J. Phys. Anthropol. 32, 3–25. doi: 10.1002/ajpa.1330320103
Traniello, J. F. A., Linksvayer, T. A., and Coto, Z. N. (2022). Social complexity and brain evolution: insights from ant neuroarchitecture and genomics. Curr. Opin. Insect Sci. 53:100962. doi: 10.1016/j.cois.2022.100962
VanSickle, C., Cofran, Z., and Hunt, D. (2020). Did Neandertals have large brains? Factors affecting endocranial volume comparisons. Am. J. Phys. Anthropol. 173, 768–775. doi: 10.1002/ajpa.24124
Villmoare, B., and Grabowski, M. (2022). Did the transition to complex societies in the Holocene drive a reduction in brain size? A reassessment of the DeSilva et al. (2021) hypothesis. Front. Ecol. Evol. 10:737. doi: 10.3389/fevo.2022.963568
von Bonin, G. (1934). On the size of man's brain as indicated by skull capacity. J. Comp. Neurol. 59, 1–28. doi: 10.1002/cne.900590102
Wiercinski, A. (1979). Has the brain size decreased since the upper paleolithic period?/ La taille du cerveau a-t-elle diminué depuis le Paléolithique supérieur? (en anglais). Bull. Mém. Soc. Anthropol. Paris 6, 419–427. doi: 10.3406/bmsap.1979.1979
Witelson, S. F., Kigar, D. L., and Harvey, T. (1999). The exceptional brain of Albert Einstein. Lancet 353, 2149–2153. doi: 10.1016/S0140-6736(98)10327-6
Wu, X., Liu, W., Zhang, Q., Zhu, H., and Norton, C. J. (2007). Craniofacial morphological microevolution of Holocene populations in northern China. Chin. Sci. Bull. 52, 1661–1668. doi: 10.1007/s11434-007-0227-8
Keywords: encephalization, Homo sapiens , Holocene, Pleistocene, social evolution
Citation: DeSilva J, Fannin L, Cheney I, Claxton A, Ilieş I, Kittelberger J, Stibel J and Traniello J (2023) Human brains have shrunk: the questions are when and why. Front. Ecol. Evol. 11:1191274. doi: 10.3389/fevo.2023.1191274
Edited by:
Mathieu Lihoreau, Centre National de la Recherche Scientifique (CNRS), FranceReviewed by:
Thomas Puschel, University of Oxford, United KingdomCopyright © 2023 DeSilva, Fannin, Cheney, Claxton, Ilieş, Kittelberger, Stibel and Traniello. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jeremy DeSilva, amVyZW15Lm0uZGVzaWx2YUBkYXJ0bW91dGguZWR1