 Julia Mörchen1,2*
Julia Mörchen1,2* Frances Luhn1Olivia Wassmer3Julia A. Kunz3,4Lars Kulik1Maria A. van Noordwijk3,5Carel P. van Schaik3,5Puji Rianti6,7Sri Suci Utami Atmoko8
Frances Luhn1Olivia Wassmer3Julia A. Kunz3,4Lars Kulik1Maria A. van Noordwijk3,5Carel P. van Schaik3,5Puji Rianti6,7Sri Suci Utami Atmoko8 Anja Widdig1,2†Caroline Schuppli3,9†
Anja Widdig1,2†Caroline Schuppli3,9†- 1Behavioral Ecology Research Group, Institute of Biology, University of Leipzig, Leipzig, Germany
- 2Department of Primate Behavior and Evolution, Max Planck Institute for Evolutionary Anthropology, Leipzig, Germany
- 3Department of Evolutionary Anthropology, University of Zurich, Zurich, Switzerland
- 4Institute of Evolutionary Biology of Montpellier (ISEM), University of Montpellier, Montpellier, France
- 5Comparative Socioecology, Max Planck Institute of Animal Behavior, Konstanz, Germany
- 6Primate Research Center, Institute of Research and Community Service, IPB University, Bogor, Indonesia
- 7Animal Biosystematics and Ecology Division, Department of Biology, IPB University, Bogor, Indonesia
- 8Fakultas Biologi, Universitas Nasional, Jakarta, Indonesia
- 9Development and Evolution of Cognition Research Group, Max Planck Institute of Animal Behavior, Konstanz, Germany
Dispersal has been suggested to be challenging, especially for species that heavily rely on social learning for knowledge acquisition. One of the obstacles that migrants face is learning how to cope with an unfamiliar, new habitat, which may involve learning from resident individuals. So far, only very few studies have looked at social learning in migrants after dispersal. Here we examine how migrant male orangutans use a behavior called “peering” (an indicator of observational social learning), to learn from local individuals. In total, we analyzed 4,009 daily dyadic associations with and without peering events of 77 males of the highly sociable Sumatran orangutans (Pongo abelii) at the Suaq population and 75 males of the less sociable Bornean orangutans (Pongo pygmaeus wurmbii) at the Tuanan population, covering a combined study time of 30 years. Analysis using generalized linear mixed models supported our prediction that migrant males in Suaq preferentially peered at the local adult females. However, in Tuanan, migrants peered mostly at other adult males and local immatures. Migrants’ peering rates were highest shortly after their arrival, and significantly decreased with increasing time spent in the area. Migrants in both sites peered significantly more at peering targets’ feeding on food items that are rarely eaten within the locals’ diet, than at commonly eaten ones and peered significantly more at skill-intense food items than easy-to-process ones. Further, migrants interacted significantly more with the peered-at food item after the peering event, than before, suggesting that they practice the observed behavior. Our results therefore suggest that migrant males use peering to learn new ecological knowledge after dispersal (e.g., where and what to feed on), and continue to learn complex skills even within adulthood, (e.g., how to feed on skill-intense food items). To do so, migrants selectively attend to the most knowledgeable and/or available individuals, practice the new skill afterwards and even flexibly adjust their learning, e.g., when confronted with intolerant locals or when the need for learning decreases. Together, our study provides important evidence that social learning in great apes expands towards adulthood, an ability which critically impacted also human evolution.
1 Introduction
Social learning refers to learning that is influenced by the observation of, or by associating and interacting with another individual or its products (Heyes, 1994; Heyes, 2012). Through social learning, individuals can avoid risks and costs of own exploration (Lorenz et al., 2011), acquire knowledge faster (Custance et al., 2002; van Schaik and Burkart, 2011) and benefit from the “wisdom of the crowd” (Toyokawa et al., 2019). Accordingly, social learning is a well-documented learning mode in a variety of taxa, ranging from invertebrate (Grüter and Leadbeater, 2014) to vertebrate species (Ferrari et al., 2007; Wilkinson et al., 2010; Aplin et al., 2015; Sasaki and Biro, 2017) and is especially prevalent in humans and non-human primate species (Whiten et al., 1999; van Schaik et al., 2003; Gariépy et al., 2014). Social learning is also the prerequisite for the emergence of traditions and cultures (Whiten et al., 1999; van Schaik, 2010).
Wild great apes are known to heavily rely on social learning for skill acquisition and live highly cultural lives, with chimpanzees and orangutans known to have the most sophisticated and diverse non-human cultures in nature (Whiten et al., 1999; van Schaik et al., 2003). Extensive research on wild primates suggests that the social learning strategies during lifetime development follow a three-phase model: in the first phase, infants benefit from extensive vertical learning from the primary care giver, usually their own mother (Lehmann et al., 2013; Whiten and van de Waal, 2018). This is then followed by a second phase, where juveniles gradually learn obliquely from a wider array of group members with increasing age (Whiten and van de Waal, 2018; Ehmann et al., 2021). A third phase of potential horizontal social learning would take place when individuals reach sexual maturity and disperse to new areas, where they encounter new and unfamiliar ecological and social circumstances (see below). Mathematical models support these shifts in role model choice (who to learn from) and they suggest that during these three phases also the context and content of social learning flexibly shifts, according to the requirements of the respective life stage the individual is situated in (Lehmann et al., 2013).
The context of social learning greatly depends on the knowledge, or the deficit thereof, of the learner which can vary considerably depending on an individual’s age and experience. Potential social learning contexts are foraging, mating, or migration (Brown and Laland, 2003). One pathway during context learning is to adopt a behavior of a conspecific, when e.g. being uncertain or when the own established behavior is unproductive (Laland, 2004), with an information flow going from an expert to the naïve or not knowledgeable individual, independent of learners’ age classes (Henrich and McElreath, 2003). In terms of the content of social learning, skills such as “knowing how” are dominantly shared between parents and offspring, whereas information about the current state of the environment or conspecifics (“knowing where”, or “knowing who”), as well declarative knowledge (“knowing that”, van Schaik, 2010) is shared among adults. However, it needs to be kept in mind that individuals must balance the costs and benefits of social learning. Costs can include attending to outdated information or when competing over resources with non-related conspecifics, which are reluctant to share information (Emery and Clayton, 2001; Clayton et al., 2007; Bugnyar et al., 2016).
Through these shifts in role model selection, context and contents of social learning, individuals can build up and refine their repertoires on which they will rely during adulthood (Lehmann et al., 2013). This implies that by the time individuals reach adulthood, all necessary skills and knowledge are present, representing accumulated knowledge between and within generations (Schuppli and van Schaik, 2019). It also implies that most social learners are young, and that the frequency of social learning decreases significantly with increasing age (Schuppli et al., 2016c). However, these theoretical models only apply, when the ontogeny follows a linear development within a stable environment and they do not consider the occurrence of dramatic changes in social and environmental conditions. Natal dispersal, i.e. when individuals permanently depart from their natal area, might be a disruptive phase, where individuals cannot rely on information already collected, but must – in extreme cases – start all over, learning about a new ecological and social niche. Social learning may be one way of coping with such sudden changes (Whiten, 2017; Gruber et al., 2019; Whiten, 2021). So far it has not been systematically studied how adult migrant primates make use of observational forms of social learning, to potentially expand and update their repertoire after dispersal for optimal resource exploitation.
Many primate species show sex-biased natal dispersal, which takes place when individuals reach sexual maturity (Cheney and Seyfarth, 1983; van Noordwijk and van Schaik, 1985; Suzuki et al., 1998; Pusey, 2004). The evolution of sex-biased dispersal is a result of the species’ experienced trade-off between the costs of staying e.g. risk of inbreeding and resource competition and the benefits of staying in known areas with supportive relatives (Pusey and Packer, 1987; Silk and Brown, 2004; Widdig et al., 2017). Dispersing individuals, on the other hand, face energetic, time, and opportunity costs (Ferreras et al., 2004; Bonte et al., 2012); as well as an increased risk of mortality due to predation (Cheney and Seyfarth, 1983) and starvation (Dittus, 1977). Individuals that leave their natal area when reaching sexual maturity and disperse, might experience high levels of aggression from hostile residents after arrival, whether the species exhibits male or female dispersal (van Noordwijk and van Schaik, 1985; Suzuki et al., 1998; Kahlenberg et al., 2008). In addition, migrants may lack relevant ecological knowledge in their new habitat, which is likely to be especially true for long distance dispersers (Isbell and Vuren, 1996).
Systematic research on how social learning is used to cope with challenges of dispersal under natural conditions and how learning changes over extended periods of time is so far missing. However, experimental studies in wild vervet monkeys (Chlorocebus pygerythrus) found that migrants selectively learn from philopatric females about how to open an “artificial fruit” (van de Waal et al., 2010) or a foraging box (Bono et al., 2018) and would even abandon learned food preferences in favor of the local norm (van de Waal et al., 2013). Luncz and Boesch (2014) reported that one migrant female chimpanzee (Pan troglodytes) progressively changed her own socially learned nut-cracking technique after immigrating to a new group where individuals show a different technique. Other studies also reported how migrants would adapt in different aspects of the social domain in different primates (Pan troglodytes, Nakamura and Uehara, 2004; Papio anubis, Sapolsky and Share, 2004). Furthermore, there is evidence for social learning in the social context in that migrants and locals adapt to strong intergroup aggression in chimpanzees (Wrangham and Glowacki, 2012; Wilson et al., 2014) or to highly affiliative behaviors in bonobos (Pan paniscus, Sakamaki et al., 2015; Fruth and Hohmann, 2018). However, unlike in orangutans (see below), these species live in social groups where migrants are expected to experience a certain degree of conformity in the form of peer or group pressure, which likely influences the migrants’ social learning to manage social relationships, maybe in addition to learning the local ecology.
Orangutans are especially suited to investigate social learning because of their slow development (Wich et al., 2004; van Noordwijk et al., 2018) which provides growing individuals with plenty of opportunities of social learning from mothers and others (Schuppli et al., 2016a; Ehmann et al., 2021). Wild orangutans have been shown to be highly neophobic (Forss et al., 2017), likely to avoid risks and costs of individual exploration (Forss et al., 2015). Despite their strong novelty avoidance, wild orangutans live in skill-intense feeding niches and rely on broad and difficult-to-acquire skill repertoires (Marshall et al., 2009; Schuppli et al., 2016b). Immature orangutans have to learn more than 200 different food items and skill intense foraging techniques, a process which takes around 12 years (Jaeggi et al., 2010; Schuppli et al., 2016a), while items and techniques that are rare or difficult require the highest learning effort (Schuppli et al., 2016c). During this time, immature orangutans also gain increasing independence from their mothers as evident in a decrease in time spent in physical contact, as well as behavioral changes (van Noordwijk and van Schaik, 2005; van Noordwijk et al., 2009; Mendonça et al., 2017). Results by Schuppli et al. (2016a) suggest that virtually all learning in immature orangutans happens via observational social learning in the form of peering, first from their mothers and with increasing age from other individuals (Schuppli et al., 2016c; Schuppli and van Schaik, 2019). In addition, with increasing independence from the mother, immature orangutans show sex-specific interest in their peering behavior towards specific classes of adult role models, presumably in preparation for their later dispersal patterns and adult role (Ehmann et al., 2021).
Peering is the attentive close range watching of an individual (hereafter “peering target”) with such avid attention and physical closeness, that it allows the peerer to witness the details of the peering targets activity (Figure 1, see full definition Table 1 in methods). Peering has been established as a measure of social learning in the wild: e.g. immature chimpanzees (Matsuzawa et al., 2001; Lonsdorf et al., 2004), immature capuchin monkeys (Cebus apella, Ottoni et al., 2005) adult bonobos (Idani, 1995; Péter et al., 2022), and in Sumatran (Pongo abelii) and Bornean orangutan immatures (Pongo pygmaeus wurmbii, Schuppli et al., 2016a; Ehmann et al., 2021), as well as in captivity: e.g. in adult Sumatran and Bornean orangutans (Stoinski and Whiten, 2003; Dindo et al., 2011), adult bonobos (Stevens et al., 2005) and adult chimpanzees (Yamanashi et al., 2020).

Figure 1 Migrant orangutan male (on the right side) peering at an adult local female (on the left side), feeding on termites within a dead branch, species Pongo abelii. Photo courtesy SUAQ Project, www.suaq.org.

Table 1 Definitions of the focal behaviors used as a measure of observational social learning and practice behavior, changed after Schuppli et al. (2016c).
Sumatran orangutans live in individual based fission fusion systems (van Schaik, 1999), but are highly sociable, whereas Bornean orangutans are less sociable, spending most of their time alone (van Noordwijk et al., 2012). The differences in sociability are linked to differences in food availability on the two islands. Sumatra tends to have higher forest productivity (Wich et al., 2011), which has likely led to the higher evolved levels of sociability and tolerance due to reduced competition over resources (Knott et al., 2008; Knott et al., 2010; Schuppli et al., 2017). However, in both species males disperse at the onset of reaching maturity, while females are the philopatric sex and settle close to their natal area (Arora et al., 2012; Nietlisbach et al., 2012; van Noordwijk et al., 2012). By the time males disperse for the first time, they are still in a state named “unflanged” which resemble the female appearance (Morrogh-Bernard et al., 2011). Most males later go through a secondary growth spurt and develop into so called “flanged” males with secondary sexual characteristics, including cheek pads (Utami Atmoko et al., 2002; Dunkel et al., 2013). Flanged and unflanged males may eventually settle and become residents, or remain transient and move between areas (Delgado and van Schaik, 2000). Although it is not exactly known where they disperse to, studies showed that males travel over long distances and even cross physical barriers such as rivers and mountains (Nietlisbach et al., 2012; Nater et al., 2013). Additionally, males are known to have large overlapping home ranges (van Schaik, 1999; Singleton and van Schaik, 2001). Due to their long-distance dispersal, migrants are expected to possess different knowledge compared to the local resident females (Schuppli and van Schaik, 2019). Since differences in socially learned skills are even visible between individuals living in the same study area (Jaeggi et al., 2010) and in close communities with similar habitats (Bastian et al., 2010), males that have recently arrived are expected to lack knowledge on the local area, in contrast to males who have already spent more time in the area or resident females. Accordingly, (Schuppli and van Schaik, 2019) showed that local adult females hardly peer at all.
The aim of this study is to investigate whether migrant male orangutans use peering as means to socially learn about the local ecology of their new habitat after dispersal. To do so, we investigate the peering content, peering target choice, and effects of arrival time on males’ peering frequencies. Specifically, we will test the following predictions:
1. Peering in the feeding context: If males peer to learn about the local ecology then they should mostly peer at orangutans feeding on food items which are rare, because those items are less likely to be known to the males. We do not expect an effect of the processing intensity (i.e., complexity) of the food item on peering rates, because adult migrant males should be familiar with many different processing techniques, including complex ones (prediction 1a). Furthermore, we predict a significant increase in practice rates of the peered-at behavior after a peering event (prediction 1b).
2. Peering target choice: If peering is used to learn about the local ecology, then it should be directed at role models which are most knowledgeable of the local ecology. Female orangutans spend their lives in their natal area, therefore females likely have the highest knowledge of their home range. Consequently, we predict that males peer at adult females more often than at any other age-sex class (prediction 2).
3. Peering over time: With increasing time spent in the area, males should gain more knowledge about its ecology and thus, the need to learn should decrease. Accordingly, we predict that peering rates will decrease with increasing time spent in the study area (prediction 3).
2 Methods
2.1 Data collection and study animals
The data for this study were collected from 2003 to 2018 on the population of Bornean orangutans (Pongo pygmaeus wurmbii) at the Tuanan research station, in Central Kalimantan, Borneo, Indonesia, and from 2007 to 2020 on the Sumatran Orangutan population (Pongo abelii) at the Suaq Balimbing Research Station, in South Aceh, Sumatra, Indonesia. At both sites, data were collected during nest to nest follows of focal individuals, following a standardized behavioral observation protocol and using instantaneous scan sampling at two-minute intervals (see protocol here http://www.aim.uzh.ch/de/orangutannetwork.html). A total of 157 observers contributed to data collection. All of these observers had passed through an extensive training period and achieved at least an 85% level of agreement with experienced observers. As part of the scan sampling, the activity of the focal animal as well as the distances to all other individuals in association with the focal individual (see below) were noted at two-minute intervals. Whenever the focal individual was feeding, the details were noted (see below). Additionally, behaviors of special interest to the objectives of the ongoing research at the sites, including all social interactions, were recorded at an all occurrence basis and described in detail. At both sites, all occurrence data on peering behavior (see Table 1 for definitions) were collected. For each peering event, the identities of the peerer and peering target as well as the duration of the peering event were noted. Whenever possible, the reactions of the peerer and the peering target were described in detail. This included who approached to peering distance, as well as who ended the peering event and how this was achieved (see Figures S4–S6 in Supplement A). The peered-at activities of the peering targets were divided into four categories: social, nesting, feeding and other behaviors (i.e. included activities like moving, resting, defecating, etc.). Following (Schuppli et al., 2016c), we defined nest peering distance as ranging from 0 to 5 meters between the peerer and the peering target(s) whereas for all other contexts (including feeding context) peering distance was defined as ranging from 0 to 2 meters.
For peering in the feeding context, the species of the consumed food item were noted, as well as the parts eaten by the individual (i.e., leaves, flowers, fruits, bark, pith, vegetative matter, but also insects and their products). Throughout this study, we refer to the combination of species name and the part eaten, as a “food item” (Bastian et al., 2010). Food items that were not recognized during a follow were photographed for later identification, consulting experienced staff members or the project’s extensive botanical record. In total, we had ~28,000 hours of feeding data, and a total of ~42,000 male focal follow hours available (Supplement A, Table S1). The feeding data was used to create frequency scores for food item combinations for both sites separately, by ranking each food item combination according to the time it had been eaten by the local females and their independent offspring. The frequency of particular food items being eaten varied greatly; from 2 minutes to 2500 hours throughout the record of feeding events in the populations. To control for the influence of temporal variation in food availability on social learning at both sites, we used the fruit availability index (FAI), calculated each month as the percentage of fruiting trees in established phenology plots (Marshall et al., 2009; Vogel et al., 2015).
We defined a daily dyadic association as a focal male spending time with another individual of any age-sex class, within 50 meters. Every encountered orangutan, either as an association partner of the focal (henceforth called “association member”) during a focal follow or during opportunistic encounters, was identified and recorded on a daily basis. We distinguished between four categories of association members: adult females, immatures, unflanged males and flanged males. All adult male orangutans, including 88 unflanged and 64 flanged males, were classified as adult migrants due to the species’ male-based dispersal. For each male, we estimated their arrival date in the study area based on the date he was first encountered by the research teams. We also computed a continuous record of the number of months they were encountered in the study area which represents the absolute minimum number of months they had spent in the study area (see Figure S3 in Supplement A). As locals we considered all adult females with their immature offspring, who had not reached sexual maturity (usually at 12–15 years of age, van Noordwijk et al., 2018). In total, we included 49 adult females and their 80 immature offspring from both sites into the analysis as association members. Since no peering was directed at flanged males, we excluded them as peering targets from the analysis.
In total, we compiled 4009 daily dyadic associations, with a total of 207 daily dyadic associations with 534 peering events and 3919 daily dyadic associations without peering events, of a total of 1350 dyads from both sites (see data overview in Supplement A, Table S1). To account for different durations of peering events, they were weighted according to their duration, e.g., a peering event with a duration of minimum of 5 to 120 seconds got a peer count of 1, from 120 to 240 seconds got a peer count of 2 and so on (Supplement A, Table S2). Since data availability differed for each prediction tested, the sample size varies across the different models (see Supplement A, Table S3) for amount of data used per model.
2.2 Data analysis, data sets and model structures
2.2.1 Peering in the feeding context
To test the effects of food item frequency and processing intensity on peering (prediction 1a), we calculated the number of peering events the migrants directed at peering targets feeding on different food items. To assess the level of manual and oral processing steps needed to consume each item (i.e. food item processing intensity), each food item was classified by the number of steps needed to process it before ingestion. Processing steps range from 0 (e.g., pick and eat a leaf) up to 5, which represent the most sophisticated form of food processing skills including tool use, which is regularly seen only in the Suaq population (Meulman and van Schaik, 2013; Schuppli et al., 2016c, see Supplement A, Table S4). This analysis explored patterns within male peering behavior and thus only considered days where actual peering occurred. To control for varying opportunities to peer at different food items, we included the time each food item was eaten by the peering targets while being in association with the peerer. In total, we analyzed 789 daily dyadic events with a migrant peerer associating with another orangutan of any age-sex class in 74 dyads. The structure of model 1a included the males’ peering count as response variable and the time the peering target had spent feeding on the respective food item per day as an offset term. Food item frequency and processing intensity were included as predictors and FAI as control variable. Additionally, we included date, food items and dyad as random effects and random slopes of dyad over FAI and processing intensity into the model.
To test if males practice what they peered at (prediction 1b), controlled for the time the peerer spent in a given food patch with a peering target, we calculated the migrants’ interaction rate with the peered-at item, before and after the peering event (see Table 1. for definitions). For this question only data from the Suaq population were available, since patch entry and exit times were not systematically recorded in Tuanan. In total, we here analyzed 126 dyadic events with a migrant peerer associating with another orangutan of any age-sex class of 15 different dyads, including 12 migrant males. Model 1b included the males’ number of interactions with the food item before and after peering as response variable, and the time the peerer spent in the given food patch per dyad as an offset term. We included the condition (before and after the peering event) as a categorical predictor and FAI as control variable. Additionally, we controlled for the dyad and date by including them as random effects.
2.2.2 Role model choice
To test if males learn about the local ecology by peering most at local philopatric females, who are expected to possess the most knowledge on the area (prediction 2), we used the full record of dyadic associations between migrants and individuals of the different age-sex classes during our study period. For each male, we calculated the number of peering events directed at each dyadic association partner, controlled for the time they had spent in association (i.e., between 0 and 50 meters) during a particular three-month period (hereafter “quarterly peering count”). This included all daily dyadic associations migrants spent in association with orangutans of the different age-sex classes, i.e. days with both peering absent and present. We here used 2,426 daily dyadic events with a migrant peerer associating with other party members of any age-sex class (1350 dyads). Model 2 included the males’ quarterly peering count as response variable and the time in association with the dyad partner per day as an offset term. In this model, we included an interaction between the peering targets’ age-sex class and the site due to the expected differences in social tolerance of individuals of different age-sex classes. Additionally, we controlled for male ID, dyad and year by entering them as random effects into the model.
2.2.3 Peering over time
To test if male peering decreases with increasing time spent in the area due to their increase in local knowledge (prediction 3), we included the males’ quarterly peering count as response variable and the time in association with the dyad partner per day as an offset term. We then linked this data with the number of months the migrant peerer had been recorded to be present in the area, and controlled for the respective FAI value at the specific association date. Since in 3,919 out of all 4,009 total dyadic observations (97%, Supplement A, Figure S1) the males’ peering count was zero, we here present a model excluding all dyadic associations without peering. Therefore, we only explored patterns within male peering in this analysis. Model 3 included the time a migrant had spent in the area (in months) as a continuous predictor and the FAI as continuous control variable. We further controlled for the site, dyad, year and ID by including them as random effects. Additionally, we included random slopes of ID, dyad and year over FAI and the continuous predictor present month in area. In total, we analyzed 149 daily dyadic events with a migrant peerer associating and peering at another party member of the different age-sex classes, summed up quarterly.
2.3 Statistical analyses
All analyses and graphs were done in R, version 4.2.0 (R Core Team, 2023). We used generalized linear mixed models (GLMMs) with a negative binomial family distribution (model 1a, 2) or a Poisson family distribution (model 1b, 3) as implemented in the glmmTMB package (Brooks et al., 2017). Throughout all analyses, the fruit availability index (FAI), food items frequency and processing intensity were z-standardized across sites. We ran full-null model comparison using likelihood ratio tests (LRT) by using the “anova” function, with the null model containing the random effects and the control variables only (Fox and Weisberg, 2019). If the comparison revealed that the full model fitted the data significantly better than the null model, we assessed the effect of each predictor in the full model using the “drop1” function of VGAM package (Yee, 2020). In the case that the GLMM included categorical predictors, post-hoc tests were done using Tukey pairwise comparisons implemented in the emmeans package (Searle et al., 1980). Furthermore, all models were tested for overdispersion (Mundry, 2022) and zero-inflation using the DHARMa package (Hartig, 2022). Random slopes were included in model 1a) and 3) and excluded from all other models to achieve model convergence, see Supplement B for random effects and random slopes output for each model. The dispersion parameter of all models ranged from 0.56 to 1.11, the zero-inflation ratio of observed to predicted zeros from 0.98 to 1.07 suggesting no critical cases. We assessed the overall fit of the models by calculating the conditional pseudo delta R2 using the MuMln package (Nakagawa et al., 2017; Bartoń, 2023). We validated the models’ stability using influence diagnostics of the “glmmTMB_stability” function following (Mundry, 2022), which compares model estimates from the full dataset with those that were extracted from the data excluding cases (individuals and dyads) one at a time. We detected no influential cases, since the direction of the effects were consistent in all models (Supplement B, detailed model output). Additionally, all models were checked visually to assess whether they fulfil the model assumptions (Harrell, 2015). Using the vif() function from the car package (Fox and Weisberg, 2019), we further detected no effect of multicollinearity among the fixed effects in any model, with the variance inflation factors ranging from 1.00 to 2.39. Alpha level throughout all tests was set on p = 0.05 with significant values marked with bold fond. Trends were interpreted for p values ranging between > 0.05 and 0.08 marked with bold fond and a dot.
3 Results
Descriptively, we found that in absolute numbers, most (67.84%) of the peering events by male migrants happened in the feeding context (Supplement A, Figure S2; Table S6). When analyzing the males’ peering behavior in the feeding context in more detail, we found several lines of evidence that migrant orangutan males use peering to learn about local food items (prediction 1a).
1a Peering in the feeding context: food item frequency and processing intensity
The full-null model comparison revealed that the set of predictor variables used had a significant influence on the migrants’ peering rates (full-null model comparison, LRT: X2 = 19.62, df = 4, P < 0.001). Specifically, the full model revealed that the food item processing intensity had a significant positive effect on migrants’ peering rates, with migrants peering significantly more at skill-intense food items, than at easy-to-process ones (Table 2; Figure 2). Furthermore, food item frequency had a significant negative effect on peering rates, i.e., migrants peered significantly more at rare food items than at common ones (Table 2; Figure 3).
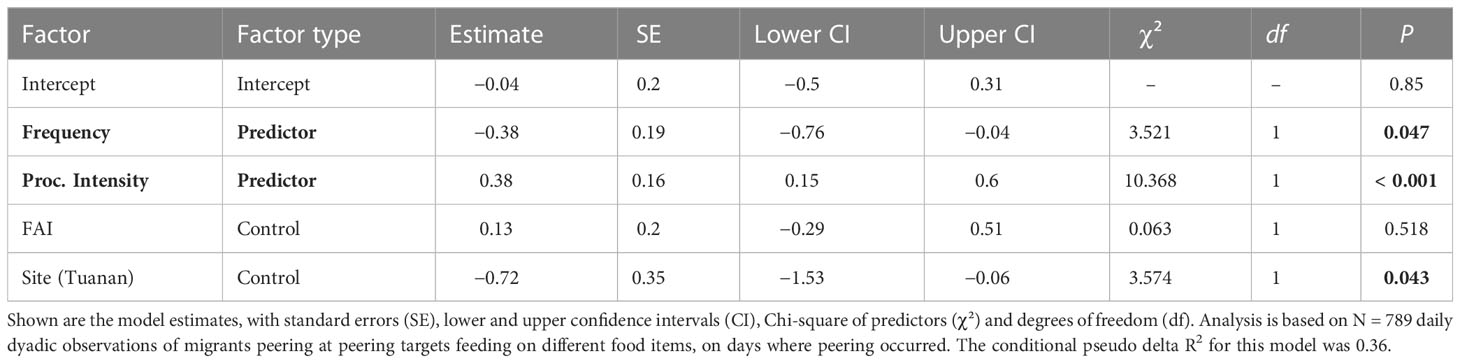
Table 2 The effect of the food items’ frequency and processing intensity on the migrants’ peering behavior analyzed using a GLMM with a negative binomial family distribution.

Figure 2 Migrants peering rates at food items with increasing food item processing intensity, ranging from 0 of being very easy to process, to 5 of being very skill-intense to process, controlled for the time in hours the peering target fed on the item. Each dot represents the average rate per individual male per complexity level and food item combination. The mean peering rate over all males is shown as a dotted line, the median is indicated by the horizontal line, the upper and lower quartile are depicted by the box and the minimum and maximum values are shown by the whiskers.
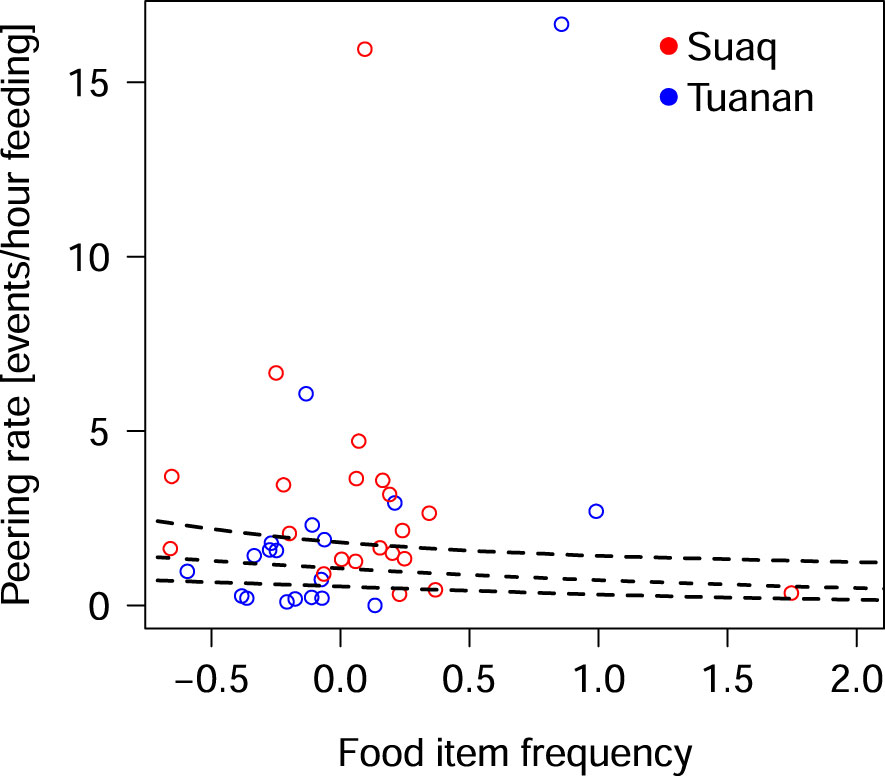
Figure 3 Migrants peering rates at different food items, depending on the items frequency in the local females’ and their independent offspring’s diet, ranging from being very rare (−0.5) to very common (1.0) controlled for the time the peering target fed on the respective food item. Each dot represents the average rate per individual male per food item combination. The dashed line depicts the fitted model, with its confidence limits for the predictors as dotted lines.
1b Peering practice behavior
The full-null model comparison revealed that the predictor variable used had a significant influence on the migrants’ interaction rate with the peered-at item (full-null model comparison, LRT: X2 = 951.403, df = 1, P < 0.001). The full model showed that migrants interact significantly more with the peered-at item after the peering event than before the peering event (Table 3; Figure 4).

Table 3 Results of the GLMM with a Poisson family distribution of the migrants’ interaction rate with the food item before and after the peering event (condition) for the Suaq population only.
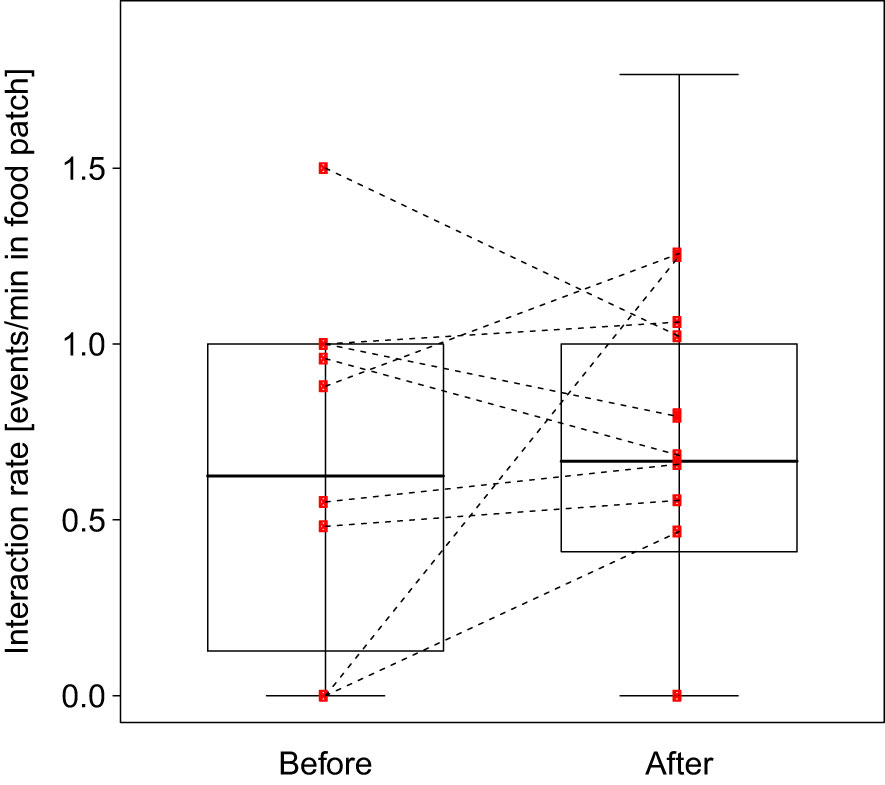
Figure 4 Migrants interaction rates per dyad with the peered-at item before and after the peering event at the Suaq population. Each dot represents the average rate per individual male depending on the condition (before and after). The median is indicated by the horizontal line, the upper and lower quartile are depicted by the box and the minimum and maximum values are shown by the whiskers.
2 Role model choice
The full-null model comparison revealed that the predictor variable used had a significant influence on the migrants’ peering rates (full-null model comparison, LRT: X2 = 49.57, df = 5, P < 0.001). The full model revealed a significant interaction between the age-sex class of the role model and study site (LRT: X2 = 15.02, df = 2, P < 0.001, Table 4; Figure 5). The interaction showed that migrants of the more sociable population Suaq peered significantly more at adult females and local immatures, compared to Tuanan, the less sociable population. Specifically, the post-hoc test showed that migrants in Suaq peered most frequently at local adult females, followed by unflanged males and local immatures (Supplement A, Table S5; Figure 5). In contrast, migrants in Tuanan peered most often at unflanged males, followed by local immatures and the least at local adult females (Supplement A, Table S5; Figure 5). At both sites, no peering was directed at flanged males.
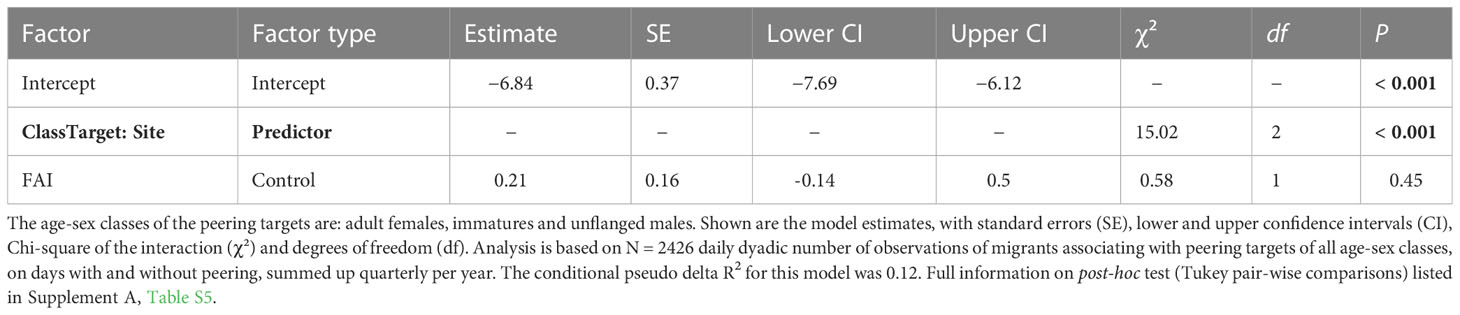
Table 4 The effect of the interaction between the role models’ age-sex classes and site, on the migrants peering behavior analyzed using a GLMM with negative binomial family distribution.
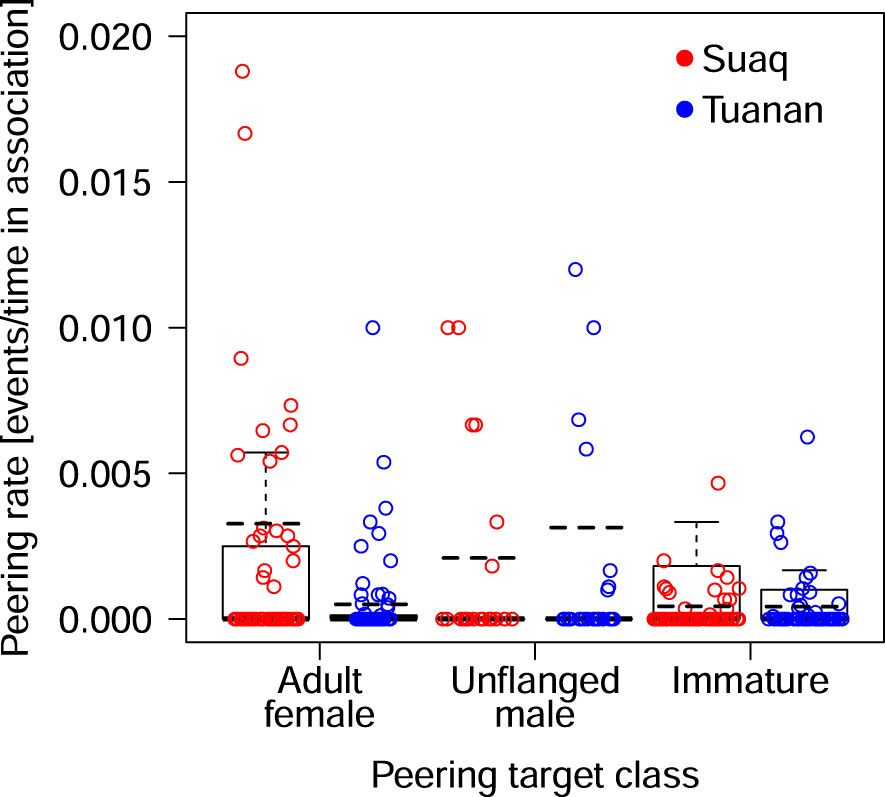
Figure 5 Migrants peering rates at specific peering target classes, controlled for the time in association. Each dot represents an average rate per individual male per peering target class, with the mean peering rate overall all males as a dotted black. The median is indicated by the horizontal line, the upper and lower quartile are depicted by the box and the minimum and maximum values are shown by the whiskers.
3 Peering over time
The full-null model comparison revealed that the predictor variable used had a significant influence on the migrants’ peering rates (full-null model comparison, LRT: X2 = 12.94, df = 2, P < 0.001). In more detail, migrants’ peering rates decreased significantly with increasing time spent in the area (Table 5; Figure 6).
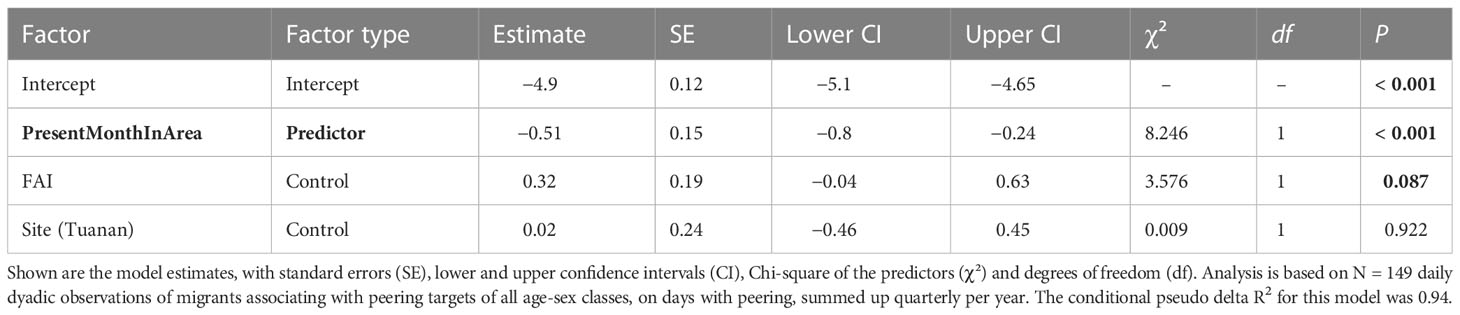
Table 5 The effect of the time spent in the area on the migrants peering behavior analyzed using a GLMM with Poisson family distribution.
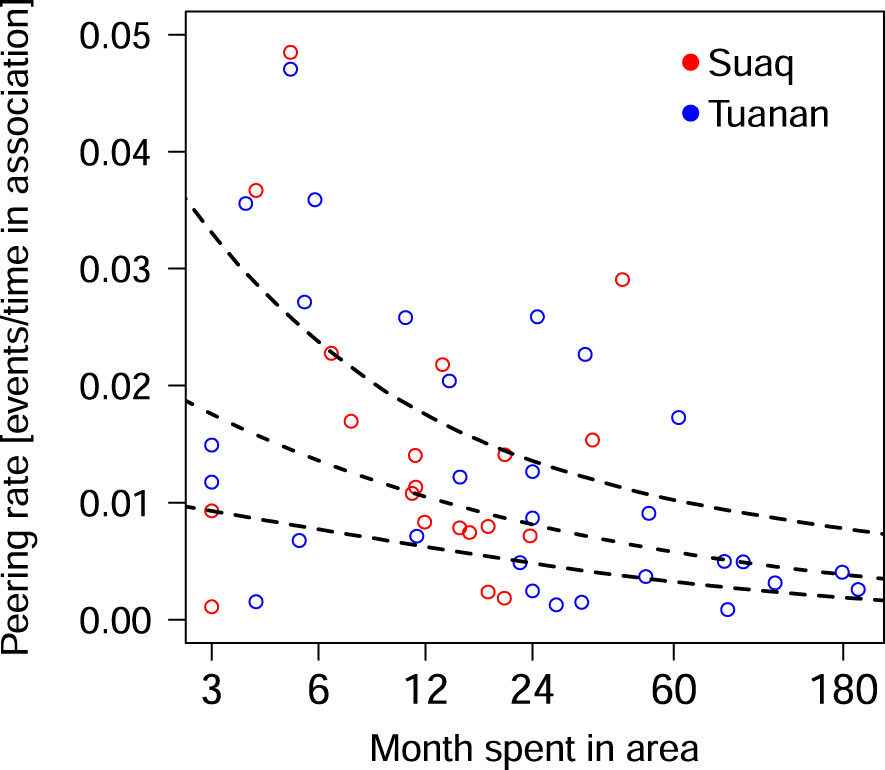
Figure 6 Migrants’ peering rates, as a function of the number of absolute months spent in the study area until the day of the peering event, controlled for the time being in association with respective peering targets. Each dot represents the average rate per individual male per time point in area. The dashed line depicts the fitted model, with its confidence limits for the predictors as dotted lines.
4 Discussion
Our study investigated whether migrant male orangutans use peering behavior to socially learn about the food items in their new habitat after dispersal. Therefore, we examined the content of migrants’ peering behavior, their role model choice and subsequent practice of the learned behavior in two different orangutan populations using data collected over 30 study years. Further, we analyzed how the migrants’ peering rates would change with increasing time spent in the area. We found several lines of evidence that male migrant orangutans indeed use peering behavior as an observational form of social learning and utilize it in a flexible and selective manner. The results suggest that migrant males use peering to acquire new ecological information after dispersal, including where and what to feed on (on rare and common items) and continue to learn complex skills even within adulthood (like how to feed on skill-intense food items). To do so, migrants selectively attend to the most knowledgeable and/or available individuals of the study areas, subsequently practice the new skill, and flexibly adjust their learning, e.g. when confronted with intolerant locals or when the need for learning decreases. Crucially, we found these effects in two orangutan species which show differences in their levels of sociability and tolerance originating from differences in food availability between Borneo and Sumatra (Wich et al., 2011).
4.1 Peering contexts and food peering
In line with our predictions we found several lines of evidence that migrant orangutan males use peering to learn about local food items (prediction 1a). Descriptively, we found that the majority of the migrants peering events happened in the feeding domain. First, and against our initial prediction, we found that migrants’ peering increases with increasing processing intensity of a food item. The highest levels of peering were shown at peering targets engaging in the most skill-intense forms of food processing skills, including tool use. It is generally assumed that in non-human primates skill learning (knowing how) is completed by the end of the developmental period (van Schaik, 2010; Schuppli et al., 2016a). In immature orangutans, adult levels of easy feeding techniques are achieved shortly after weaning, whereas the more skill intense techniques of food consumption need more time, but seem to be mostly in place before reaching adulthood (i.e., age at first reproduction) and thus before dispersal (Schuppli et al., 2016a). Furthermore, findings on humans suggest that adults are able to transfer existing skill and knowledge to solve new problems (Nisbett, 2009; van Schaik and Burkart, 2011). Our results however indicate that orangutan male migrants are lacking knowledge on local food items in the study areas and use peering to learn how to consume the easy-to-process, but especially the skill-intense local foods.
Second, we found that migrants’ peering significantly increases with increasing rarity of the respective food item in the locals’ overall diet. This result is in line with our prediction, that migrants use peering to learn what they can eat in their new habitat, as well as where and when to find it. Similar findings have been reported in immature orangutans (Schuppli et al., 2016c) and in wild capuchin monkeys, were individuals were found to pay more attention to foods, that are rare in their diet and that are difficult to process (Perry and Jimenez, 2006). It is known that with increasing geographic distance the similarities in habitat ecology, composition and culture decreases, which increases the likelihood that the migrants which are known to disperse over long distances might not know especially the rare local food items (van Schaik et al., 2003; Russon et al., 2009; Krützen et al., 2011). However, it is not known, if items that are commonly eaten at Suaq and Tuanan, are commonly available items in other areas. Furthermore, food species might be commonly eaten locally but still rarely eaten in other areas, due to diet preferences (Bastian et al., 2010). Therefore, to fully test our prediction we would have to gain knowledge about food item availability in the males’ natal populations.
Third, we found evidence that migrant males’ practice what they have learned, e.g. by interacting more frequently with the peered-at item after the peering event than before (prediction 1b). Similar selective explorative behavior has been studied in immature orangutans suggesting that peering was followed by increased rates of practice behaviors with the peered-at items (Schuppli et al., 2016c). Future studies should identify whether the learned and practiced knowledge is actually used in the migrants daily lives, and whether they also apply this knowledge when being alone without a demonstrator.
4.2 Peering target choice
In line with prediction 2, that migrants should preferably learn from those role models that have most knowledge of the local area, we found that in the highly sociable population of Suaq, migrants showed the highest peering rates at local philopatric females, followed by peering at local immatures and other adult unflanged males. Similar findings have been reported in wild male migrant vervet monkeys, that selectively learned from the philopatric females about how to open an “artificial fruit” (van de Waal et al., 2010). However, in the less sociable population of Tuanan, we found the opposite in the migrants’ role model choice, as they showed the highest peering rates at adult unflanged males, followed by peering at local immatures and the least at philopatric females. Tuanan migrant males may not have many opportunities to peer at local females because females avoid long associations with males, (Kunz et al., 2021), and thus eventually also close proximity or feeding close to males.
In Suaq and Tuanan, males overall initiate and maintain associations with females more frequently than the females themselves. However, only in Tuanan, the Bornean population with the less productive habitat, prolonged associations with males led to an increase in fecal cortisol metabolite levels in females, suggesting that associations are stressful and costly for the females (Kunz et al., 2021). Accordingly, in Tuanan, females are also avoidant of unrelated females (van Noordwijk et al., 2012). These findings are thought to stand in direct connection with the less productive habitat in Borneo that causes increased levels of resource competition (Rijksen, 1978; van Schaik, 1999; Kunz et al., 2021). This argumentation is further supported by a study on the Tuanan orangutans showing that philopatric females will decrease their low level of gregariousness and social tolerance even more, when confronted with increased levels of competition due to the long-term effects of human-induced fires (Ashbury et al., 2022). Social intolerance towards migrants in such cases therefore most likely does not originate from strong social peer pressure (conformity) as it is seen in chimpanzees (Whiten et al., 2005) or vervet monkeys (van de Waal et al., 2013), but instead from direct competition over scarce resources. This is further supported by our result that at both sites, no peering was directed at flanged males, who are known to be less tolerant towards other males, likely because they compete over ecological resources and access to females with these males (Utami Atmoko et al., 2009).
It appears that migrants at Tuanan would then choose the “second best option to learn from”, which are other adults like unflanged males or local immatures, who themselves (depending on their age, or the time already spent in the area) may have a substantial amount of local knowledge (Schuppli et al., 2016a). In general, immatures and unflanged males are the most sociable age sex classes in orangutans, which may favor relaxed associations in closer distance to each other (Mitra Setia et al., 2009). Previous studies showed that immature orangutans exhibit a growing interest in and tendency to peer at individuals other than their own mothers as they become older and more independent (Schuppli et al., 2016a). Furthermore, learning from immigrant males seems to be important for the immatures’ skill acquisition, in particular for immature males (Ehmann et al., 2021). Therefore, unlike for the adult females, the benefits of learning from adult males may outweigh the costs of close associations with these males, especially for older and male immatures. These mutual benefits of knowledge exchange between adult males and immatures may promote close associations and peering between these classes. These patterns suggest, that in contrast to the established social learning theories, under conditions like in Tuanan, migrants seem to be forced to seek information from peering targets, that are unlikely to possess complete local information: unflanged males, who originated from different areas and immatures who have not completed their learning about the local area. This horizontal and oblique learning thus may bear the risk to obtain incomplete or even wrong information (Henrich & McElreath, 2003; Laland, 2004). These findings also demonstrate the males’ selectivity and flexibility in their peering target choice. Similar flexibility in role model choice has been found in wild vervet monkeys, were migrants usually would copy local philopatric females, but do switch to copy dominant males when these males gain a much higher payoff in experimental food reward setups as females (Bono et al., 2018). This mental flexibility is generally already visible in immature orangutans, who, as they get older, increasingly learn from other individuals than the own mother (Schuppli et al., 2016c) and show sex-specific preferences for role models (Ehmann et al., 2021).
4.3 Learning over time
In line with our prediction 3, we found that the migrants peering rate significantly decreased with increasing time the male had spent in the study area. Our results show that within the first 6 months after dispersal the migrants peering rates are particularly high. At this time, they likely still lack the majority of knowledge of the new area. Our results suggest that the subsequent gradual learning process can take multiple months, which may partly be due to the fact, that many food items are not available all year around. The drop in peering rates suggests that with increasing time spent in the area, the migrants’ competency on local ecological increases and thus the need to learn decreases. These findings parallel how immature orangutans socially learn via peering, where with increasing age and competence, immature peering rates gradually decrease (Schuppli et al., 2016c). It has been estimated that over the course of their lifetime, Bornean and Sumatran orangutans peer approximately 9,000 and 38,000 times whereby the majority of all peering happens during immaturity (Schuppli and van Schaik, 2019). The results of our study suggest, that at least for the dispersing males, the “learning window” remains open over the course of their dispersal and the first years after it.
The fact that in immatures age has a strong effect on peering rates, raises the question whether age may also affect the peering rates of the males in this study. Unfortunately, because we do not know how old the males in our study are (because we have no information on their lives before their arrival at the study sites), we cannot test for age effects. However, when the males arrive in our study areas, their body sizes vary visibly (Schuppli and van Noordwijk, personal communication), which suggests that the males are of different ages. After their arrival, peering remains high for the first 6 months only, but then drops. Furthermore, in adult females, peering remains at very low levels throughout adulthood (Schuppli & van Schaik, 2019). Therefore, age effects alone are unlikely to bring about the peering patterns we found in the adult males.
5 General discussion
Our study represents a systematic analysis of adult migrant orangutans’ observational social learning (measured in peering behavior) from local residents after dispersal. By examining the actual learning process (rather than focusing on the result of learning), we were able to identify details on the content and the role model choice. This stands in contrast to existing studies that have either focused on single migrants, and single behaviors of migrants or local conspecifics only, or within conditions that were altered experimentally. Furthermore, by including data collected over a study period of 30 years, we were able to track individual migrants peering behavior as a function of the time they had spent in the study area. We were thus able to cover the process of migrants’ social learning, from their inferred arrival, followed by the chronological changes over time. The results parallel how immature orangutans socially learn during immaturity, but differ significantly in those areas (e.g. content and role model choice), that are specifically important for adult orangutans confronted with a new ecological environment after dispersal (see further down). Hence, the flexibility in wild adult orangutans’ social learning might be key to counterbalance the costs of dispersal and to successfully adapt to a new habitat.
6 Conclusions
First: Dispersal seems to be a very disruptive phase for migrant orangutans. Although when compared to immatures, migrants show very low peering rates (Schuppli et al., 2016c), our results show that migrants temporarily resume peering as an observational form of social learning. Therefore, the existing ecological knowledge of migrants seems insufficient to thrive in the new habitat without social learning. Our results support the suggested third phase of social learning during primate development, which stated that migrants make use of social learning after dispersal to adapt to the new habitat (Whiten and van de Waal, 2018).
Second: With this study we add to the growing body of evidence that peering is a means of observational social learning. Observational social learning allows migrants to learn relevant local knowledge safely and efficiently without individual re-invention while benefitting from already existing knowledge instead. Not knowing what is edible and what is dangerous entails substantial risks and can lead to fatalities. Furthermore, individuals that are in possession of critical ecological information have a survival advantage (Kurman and Ronen-Eilon, 2004; Marzec, 2020). Follow-up studies should therefore examine to what extent the male’s repertoires correlate with their fitness. Ultimately, since large bodied, long-lived and slow breeders like great apes, are especially challenged by sudden changes in their environment (next to dispersal, e.g. also due to habitat loss), relying on social learning for adaptation might be the most efficient way, since the adaptation through regular biological evolution would take too long for individuals to come up with advantageous traits just in time.
Third: In our study we found preliminary evidence that social tolerance might be the key to allow for relaxed learning situations. The local females in the less sociable species are known to avoid long associations with males (Kunz et al., 2021), which may decrease the migrants’ opportunities to socially learn from them after arrival. At this point, it is not possible to identify the causes for this diminished learning opportunities. It is possible that either females and or males will increasingly grow more intolerant in times of scarcity and will therefore decrease physical closeness to each other to avoid direct resource competition by e.g. being in the same feeding patch. Overall, the successful long-term integration of a migrant might therefore critically depend on such tolerant ties with the local females.
Fourth: Being our most distant great ape relative, the study of the orangutans may reveal shared traits at the very basis of the Hominidae lineage some 12–14 million years ago (Locke et al., 2011). The ability for migrants to use observational forms of social learning after dispersal seems to be the ancestral state of the hominin lineage. The conditions at the time of this trait’s emergence, must have selected for migrants that have the general ability to harvest the benefits of cultural knowledge through social learning from locals, but avoided the costs through competition in times of food scarcity. In the course of hominin evolution, this ability then developed further alongside the different species’ social systems and formed the respective species’ social learning biases; even eventually into the two most opposite extremes possibly, as it is seen now in modern-day chimpanzees vs. bonobos, which split approximately 2 million years ago (Prüfer et al., 2012). The human journey has always been a matter of migration and migrants are described to be exceptional people, in terms of versatility, resilience and adaptivity (Goldin et al., 2011); an ability we seem to share with our most distant great ape relative.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
This study on wild orangutans was strictly observational and non-invasive, and there was no interaction with our study animals in any way. The research protocols were approved by the Indonesian State Ministry for Research, Technology and Higher Education (RISTEK398/SIP/FRP/E5/Dit.KI/X|/2017) and complied with the legal requirements of Indonesia.
Author contributions
Design and development of models: JM, CSchu, AW, LK. Analyzing models: JM, LK. Design and conceptualizing research project: JM, AW, CSchu, CScha, MN. Data collection and preparation: JM, CSchu, MN, FL, JK, OW. Writing manuscript: JM. Review of the manuscript: all authors. Supported field work: AW, CSchu, CScha, MN, PR, SU. Supervision: AW, CSchu. All authors contributed to the article and approved the submitted version.
Funding
This research is part of a PhD project of the Leipzig School of Human Origins (IMPRS) with PhD funding provided by a University of Leipzig (DFPL grant number R_00061 (JM), a DAAD doctoral research grant (grant number 57437987 (JM), the Manfred-Hermsen-Foundation (JM), the SUAQ Foundation (JM, CSchu), the A.H. Schultz foundation (JM, CSchu), Zurich University (JM, CSchu) and the Hamburglobal Scholarship of the University of Hamburg (JM). Further funding was provided by the Volkswagen Stiftung (Freigeist Fellowship, CSchu), the Stiftung für Mensch und Tier Freiburg i.Br. (CSchu) and the Leakey foundation (Primate Research Fund, CSchu).
Acknowledgments
We thank all students, volunteers and local field assistants involved in the collection of standard behavioral data for the long-term databases of Suaq and Tuanan. We are also thankful to the local staff at the field sites and associated offices. We gratefully acknowledge the Indonesian Institute of Science (LIPI), the Indonesian State Ministry for Research and Technology (RISTEK), the Directorate General of Natural Resources and Ecosystem Conservation – Ministry of Environment & Forestry of Indonesia (KSDAE-KLHK), the Ministry of Internal affairs, the Nature Conservation Agency of Central Kalimantan (BKSDA), Universitas Nasional Jakarta, the local governments in Central Kalimantan, the Kapuas Protection Forest Management Unit (KPHL), the Bornean Orang-utan Survival Foundation (BOSF) and MAWAS in Palangkaraya, as well as the Sumatran Orangutan Conservation Programme (SOCP) and Yayasan Ekosistem Lestari (YEL) in Medan. We also thank the Fakultas Biologi Universitas Nasional (UNAS) in Jakarta and Rutgers University for their collaboration and support for the Tuanan project and in particular Dr. Tatang Mitra Setia, Dr. Anna Marzec, Dr. Brigitte Spillmann and Dr. Erin Vogel. We also thank the Department of Biology, IPB University in Bogor, for their collaboration and support. Furthermore, we also thank the Department of Biology, Bogor University (IPB) in Bogor for their collaboration and support. We thank Dr. Brigitte Schlögl for statistical advice, Richard Young for help with the databases and Dr. Roger Mundry for providing us with his functions to test for overdispersion, random slopes and model stability, as well as a function to calculate the confidence intervals in our analysis.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1158887/full#supplementary-material
References
Aplin L. M., Farine D. R., Morand-Ferron J., Cockburn A., Thornton A., Sheldon B. C. (2015). Experimentally induced innovations lead to persistent culture via conformity in wild birds. Nature 518 (7540), 538–541. doi: 10.1038/nature13998
Arora N., Noordwijk M. A.V., Ackermann C., Willems E. P., Nater A., Greminger M., et al. (2012). Parentage-based pedigree reconstruction reveals female matrilineal clusters and male-biased dispersal in nongregarious Asian great apes, the bornean orang-utans (Pongo pygmaeus). Mol. Ecol. 21 (13), 3352–3362. doi: 10.1111/j.1365-294X.2012.05608.x
Ashbury A. M., Meric de Bellefon J., Kunz J. A., Abdullah M., Marzec A. M., Fryns C., et al. (2022). After the smoke has cleared: extended low fruit productivity following forest fires decreased gregariousness and social tolerance among wild female bornean orangutans (Pongo pygmaeus wurmbii). Int. J. Primatol. 43 (2), 189–215. doi: 10.1007/s10764-021-00263-x
Bartoń K. (2023) ‘MuMIn: multi-model inference’. Available at: https://CRAN.R-project.org/package=MuMIn (Accessed 19.05.2023).
Bartoń K. (2022). MuMIn: Multi-Model Inference. R package version, 1.47.1. Available at: https://cran.r-project.org/web/packages/MuMIn/index.html.
Bastian M. L., Zweifel N., Vogel E. R., Wich S. A., van Schaik C. P. (2010). Diet traditions in wild orangutans. Am. J. Phys. Anthropol. 143 (2), 175–187. doi: 10.1002/ajpa.21304
Bono A. E. J., Whiten A., van Schaik C. P., Krützen M., Eichenberger F., Schnider A., et al. (2018). Payoff- and sex-biased social learning interact in a wild primate population. Curr. Biol. 28 (17), 2800–2805.e4. doi: 10.1016/j.cub.2018.06.015
Bonte D., Van Dyck H., Bullock J. M., Coulon A., Delgado M., Gibbs M., et al. (2012). Costs of dispersal. Biol. Rev. 87 (2), 290–312. doi: 10.1111/j.1469-185X.2011.00201.x
Brooks M., Kristensen K., van Benthem K., Magnusson A., Berg C. W., Nielsen A., et al. (2017). glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9, 378–400. doi: 10.32614/RJ-2017-066
Brown C., Laland K. N. (2003). Social learning in fishes: a review. Fish Fisheries 4 (3), 280–288. doi: 10.1046/j.1467-2979.2003.00122.x
Bugnyar T., Reber S. A., Buckner C. (2016). Ravens attribute visual access to unseen competitors. Nat. Commun. 7 (1), 10506. doi: 10.1038/ncomms10506
Cheney D. L., Seyfarth R. M. (1983). Nonrandom dispersal in free-ranging vervet monkeys: social and genetic consequences. Am. Nat. 122, 392–412. doi: 10.1086/284142
Clayton N. S., Dally J. M., Emery N. J. (2007). Social cognition by food-caching corvids. the western scrub-jay as a natural psychologist. Philos. Trans. R. Soc. B: Biol. Sci. 362 (1480), 507–522. doi: 10.1098/rstb.2006.1992
Custance D. M., Whiten A., Fredman T. (2002). Social learning and primate reintroduction. Int. J. Primatol. 23 (3), 479–499. doi: 10.1023/A:1014961415219
Delgado R. A., van Schaik C. P. (2000). The behavioral ecology and conservation of the orangutan (Pongo pygmaeus): a tale of two islands. Evolutionary Anthropology: Issues News Rev. 9 (5), 201–218. doi: 10.1002/1520-6505(2000)9:5<201::AID-EVAN2>3.0.CO;2-Y
Dindo M., Stoinski T., Whiten A. (2011). Observational learning in orangutan cultural transmission chains. Biol. Lett. 7 (2), 181–183. doi: 10.1098/rsbl.2010.0637
Dittus W. P. J. (1977). The social regulation of population density and age-sex distribution in the toque monkey. Behaviour 63 (3/4), 281–322. doi: 10.2307/4533858
Dunkel L. P., Arora N., van Noordwijk M. A., Atmoko S. S. U., Putra A. P., Krützen M., et al. (2013). Variation in developmental arrest among male orangutans: a comparison between a Sumatran and a bornean population. Front. Zool. 10 (1), 12. doi: 10.1186/1742-9994-10-12
Ehmann B., van Schaik C. P., Ashbury A. M., Mörchen J., Musdarlia H., Atmoko S. U., et al. (2021). Immature wild orangutans acquire relevant ecological knowledge through sex-specific attentional biases during social learning. PLoS Biol. 19 (5), e3001173. doi: 10.1371/journal.pbio.3001173
Emery N. J., Clayton N. S. (2001). Effects of experience and social context on prospective caching strategies by scrub jays. Nature 414 (6862), 443–446. doi: 10.1038/35106560
Ferrari M. C. O., Messier F., Chivers D. P. (2007). First documentation of cultural transmission of predator recognition by larval amphibians. Ethology 113 (6), 621–627. doi: 10.1111/j.1439-0310.2007.01362.x
Ferreras P., Delibes M., Palomares F., Fedriani J. M., Calzada J., Revilla E. (2004). Proximate and ultimate causes of dispersal in the Iberian lynx lynx pardinus. Behav. Ecol. 15 (1), 31–40. doi: 10.1093/beheco/arg097
Forss S. I. F., Schuppli C., Haiden D., Zweifel N., van Schaik C. P. (2015). Contrasting responses to novelty by wild and captive orangutans. Am. J. Primatol. 77 (10), 1109–1121. doi: 10.1002/ajp.22445
Forss S., Koski S., van Schaik C. P. (2017). Explaining the paradox of neophobic explorers: the social information hypothesis. Int. J. Primatol. 38. doi: 10.1007/s10764-017-9984-7
Fox J., Weisberg S. (2019). An {R} companion to applied regression (Thousand Oaks, California: Sage), 191320. Available at: http://socserv.socsci.mcmaster.ca/jfox/Books/Companion.
Fruth B., Hohmann G. (2018). Food sharing across borders. Hum. Nat. 29 (2), 91–103. doi: 10.1007/s12110-018-9311-9
Gariépy J. F., Watson K., Du E., Xie D., Erb J., Amasino D., et al. (2014). Social learning in humans and other animals. Front. Neurosci. 8. doi: 10.3389/fnins.2014.00058
Goldin I., Cameron G., Balarajan M. (2011). Exceptional people: how migration shaped our world and will define our future, exceptional people (United States of America: Princeton University Press). doi: 10.1515/9781400836291
Gruber T., Luncz L., Mörchen J., Schuppli C., Kendal R. L., Hockings K. (2019). Cultural change in animals: a flexible behavioural adaptation to human disturbance. Palgrave Commun. 5 (1), 9. doi: 10.1057/s41599-019-0271-4
Grüter C., Leadbeater E. (2014). Insights from insects about adaptive social information use. Trends Ecol. Evol. 29 (3), 177–184. doi: 10.1016/j.tree.2014.01.004
Harrell F. E. (2015). Regression modeling strategies. Switzerland: Springer International Publishing AG. doi: 10.1007/978-3-319-19425-7
Hartig F. (2022). DHARMa: Residual Diagnostics for Hierarchical (Multi-Level / Mixed) Regression Models. R package version 0.4.6. Available at: https://cran.r-project.org/web/packages/DHARMa/index.html.
Henrich J., McElreath R. (2003). The evolution of cultural evolution. Evolutionary Anthropology: Issues News Rev. 12 (3), 123–135. doi: 10.1002/evan.10110
Heyes C. M. (1994). Social learning in animals: categories and mechanisms. Biol. Rev. Cambridge Philos. Soc. 69 (2), 207–231. doi: 10.1111/j.1469-185x.1994.tb01506.x
Heyes C. (2012). What’s social about social learning? J. Comp. Psychol. 126 (2), 193–202. doi: 10.1037/a0025180
Idani G. (1995). Function of peering behavior among bonobos (Pan paniscus) at wamba, Zaire. Primates 36 (3), 377–383. doi: 10.1007/BF02382860
Isbell L. A., Vuren D. V. (1996). Differential costs of locational and social dispersal and their consequences for female group-living primates. Behaviour 133 (1), 1–36. doi: 10.1163/156853996X00017
Jaeggi A. V., Dunkel L. P., Van Noordwijk M. A., Wich S. A., Sura A. A. L., Van Schaik C. P. (2010). Social learning of diet and foraging skills by wild immature bornean orangutans: implications for culture. Am. J. Primatol. 72 (1), 62–71. doi: 10.1002/ajp.20752
Kahlenberg S. M., Thompson M. E., Muller M. N., Wrangham R. W. (2008). Immigration costs for female chimpanzees and male protection as an immigrant counterstrategy to intrasexual aggression. Anim. Behav. 76 (5), 1497–1509. doi: 10.1016/j.anbehav.2008.05.029
Knott C., Beaudrot L., Snaith T., White S., Tschauner H., Planansky G. (2008). Female-female competition in bornean orangutans. Int. J. Primatol. 29 (4), 975–997. doi: 10.1007/s10764-008-9278-1
Knott C. D., Emery Thompson M., Stumpf R. M., McIntyre M. H. (2010). Female reproductive strategies in orangutans, evidence for female choice and counterstrategies to infanticide in a species with frequent sexual coercion. Proc. Biol. Sci. 277 (1678), 105–113. doi: 10.1098/rspb.2009.1552
Krützen M., Willems E. P., van Schaik C. P. (2011). ‘Culture and geographic variation in orangutan behavior’. Curr. Biol. 21 (21), 1808–1812. doi: 10.1016/j.cub.2011.09.017
Kunz J. A., Duvot G. J., van Noordwijk M. A., Willems E. P., Townsend M., Mardianah N., et al. (2021). The cost of associating with males for bornean and Sumatran female orangutans: a hidden form of sexual conflict? Behav. Ecol. Sociobiol. 75 (1), 6. doi: 10.1007/s00265-020-02948-4
Kurman J., Ronen-Eilon C. (2004). Lack of knowledge of a culture’s social axioms and adaptation difficulties among immigrants. J. Cross-Cultural Psychol. 35 (2), 192–208. doi: 10.1177/0022022103262244
Laland K. N. (2004). Social learning strategies. Anim. Learn. Behav. 32 (1), 4–14. doi: 10.3758/BF03196002
Lehmann L., Wakano J. Y., Aoki K. (2013). On optimal learning schedules and the marginal value of cumulative cultural evolution. Evolution 67 (5), 1435–1445. doi: 10.1111/evo.12040
Locke D. P., Hillier L. W., Warren W. C., Worley K. C., Nazareth L. V., Muzny D. M., et al. (2011). Comparative and demographic analysis of orang-utan genomes. Nature 469 (7331), 529–533. doi: 10.1038/nature09687
Lonsdorf E. V., Eberly L. E., Pusey A. E. (2004). Sex differences in learning in chimpanzees. Nature 428 (6984), 715–716. doi: 10.1038/428715a
Lorenz J., Rauhut H., Schweitzer F., Helbing D. (2011). How social influence can undermine the wisdom of crowd effect. Proc. Natl. Acad. Sci. 108 (22), 9020–9025. doi: 10.1073/pnas.1008636108
Luncz L. V., Boesch C. (2014). Tradition over trend: neighboring chimpanzee communities maintain differences in cultural behavior despite frequent immigration of adult females. Am. J. Primatol. 76 (7), 649–657. doi: 10.1002/ajp.22259
Marshall A., Ancrenaz M., Brearley F. Q., Fredriksson G., Ghaffar N., Heydon M., et al. (2009). “The effects of forest phenology and floristics on populations of bornean and Sumatran orangutans: are Sumatran forests more productive than bornean forests?,” in Orangutans: geographic variation in behavioral ecology and conservation. Eds. Wich S. A., Utami Atmoko S. S., Mitra Setia T., van Schaik C. P. (New York: Oxford University Press), 97–117. doi: 10.1093/acprof:oso/9780199213276.003.0007
Marzec A. M. (2020). Innovations and social barriers in orangutan culture (Switzerland: University of Zurich). doi: 10.5167/UZH-197837
Matsuzawa T., Biro D., Humle T., Inoue-Nakamura N., Tonooka R., Yamakoshi G. (2001). “Emergence of culture in wild chimpanzees: education by master-apprenticeship,” in Primate origins of human cognition and behavior. Ed. Matsuzawa T. (Tokyo: Springer Japan), 557–574. doi: 10.1007/978-4-431-09423-4_28
Mendonça R. S., Kanamori T., Kuze N., Hayashi M., Bernard H., Matsuzawa T. (2017). Development and behavior of wild infant-juvenile East bornean orangutans (Pongo pygmaeus morio) in danum valley. Primates 58 (1), 211–224. doi: 10.1007/s10329-016-0567-6
Meulman E., van Schaik C. P. (2013). Orangutan tool use and the evolution of technology. In: Tool Use in Animals Cognition and Ecology. Eds. Sanz C. M., Call J., Boesch C. (Cambridge, UK: Cambridge University Press), 176–202. doi: 10.5167/uzh-92534
Mitra Setia T., Delgado R., Atmoko S., Singleton I., Schaik C. (2009). “Social organization and male-female relationships,” in Orangutans: geographic variation in behavioral ecology and conservation. Eds. Wich S. A., Utami Atmoko S. S., Mitra Setia T., van Schaik C. P. (New York: Oxford University Press), 245–253. doi: 10.1093/acprof:oso/9780199213276.003.0007
Morrogh-Bernard H. C., Morf N. V., Chivers D. J., Krützen M. (2011). Dispersal patterns of orang-utans (Pongo spp.) in a bornean peat-swamp forest. Int. J. Primatol. 32 (2), 362–376. doi: 10.1007/s10764-010-9474-7
Mundry R. (2022). Linear Models and their Application in R. (Göttingen: Leibniz Science Campus Primate Cognition). Available at: https://www.primate-cognition.eu/de/biostatistik/linear-models-in-r.html.
Nakagawa S., Johnson P. C. D., Schielzeth H. (2017). The coefficient of determination R2 and intra-class correlation coefficient from generalized linear mixed-effects models revisited and expanded. J. R. Soc. Interface 14 (134), 20170213. doi: 10.1098/rsif.2017.0213
Nakamura M., Uehara S. (2004). Proximate factors of different types of grooming hand-clasp in mahale chimpanzees: implications for chimpanzee social customs. Curr. Anthropology - Curr. ANTHROPOL 45. doi: 10.1086/381007
Nater A., Arora N., Greminger M. P., van Schaik C. P., Singleton I., Wich S. A., et al. (2013). Marked population structure and recent migration in the critically endangered Sumatran orangutan (Pongo abelii). J. Heredity 104 (1), 2–13. doi: 10.1093/jhered/ess065
Nietlisbach P., Arora N., Nater A., Goossens B., van Schaik C. P., Krützen M. (2012). Heavily male-biased long-distance dispersal of orang-utans (genus: Pongo), as revealed by y-chromosomal and mitochondrial genetic markers. Mol. Ecol. 21 (13), 3173–3186. doi: 10.1111/j.1365-294X.2012.05539.x
Nisbett R. E. (2009) Richard E. nisbett - intelligence and how to get it - why schools and cultures count-w. w. Norton & company 2009) | PDF | intelligence quotient | prefrontal cortex, scribd. Available at: https://www.scribd.com/document/401877490/Richard-E-Nisbett-Intelligence-and-How-to-Get-It-Why-Schools-and-Cultures-Count-W-W-Norton-Company-2009 (Accessed 22 November 2022).
Ottoni E. B., de Resende B. D., Izar P. (2005). Watching the best nutcrackers: what capuchin monkeys (Cebus apella) know about others’ tool-using skills. Anim. Cogn. 8 (4), 215–219. doi: 10.1007/s10071-004-0245-8
Perry S., Jimenez J. O. (2006) ‘The effects of food size, rarity, and processing complexity on white-faced capuchins visual attention to foraging conspecifics’, in. Available at: https://www.semanticscholar.org/paper/The-effects-of-food-size%2C-rarity%2C-and-processing-on-Perry-Jimenez/620df1e7e643198ee814879442978cbe8f1add89 (Accessed 26 January 2023).
Péter H., Zuberbühler K., Hobaiter C. (2022). Well-digging in a community of forest-living wild East African chimpanzees (Pan troglodytes schweinfurthii). Primates 63 (4), 355–364. doi: 10.1007/s10329-022-00992-4
Prüfer K., Munch K., Hellmann I., Akagi K., Miller J. R., Walenz B., et al. (2012). The bonobo genome compared with the chimpanzee and human genomes. Nature 486 (7404), 527–531. doi: 10.1038/nature11128
Pusey A. E. (2004). “Inbreeding avoidance in primates,” in Inbreeding, incest, and the incest taboo. Eds. Wolf A. P., Durham W. H. (Stanford: Stanford University Press), 61–75. Available at: http://books.google.de/books?id=OW1nuQxcIQgC&pg=PA61&dq=Inbreeding,+Incest,+and+the+Incest+Taboo&lr=&source=gbs_toc_r&cad=4#v=onepage&q&f=false.
Pusey A. E., Packer C. (1987). “Dispersal and philopatry,” in Primate societies. Ed. Smuts B. B., Cheney D. L., Seyfarth R. M., Wrangham R. W., Struhsaker T. T. (Chicago, IL: University of Chicago Press), 250–266.
R Core Team. (2023). R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. URL https://www.R-project.org/.
Rijksen H. D. (1978). A field study on sumatran orangutans (Pongo pygmaeus abelii lesson 1827) : ecology, behaviour and conservation (Wageningen: Veenman).
Russon A., Wich S., Ancrenaz M., Tomoko K., Knott C., Kuze N., et al. (2009). “Geographic variation in orangutan diets,” in Orangutans: geographic variation in behavioral ecology and conservation. Eds. Wich S. A., Utami Atmoko S. S., Mitra Setia T., van Schaik C. P. (New York: Oxford University Press), 135–156. doi: 10.5167/uzh-31255
Sakamaki T., Behncke I., Laporte M., Mulavwa M., Ryu H., Takemoto H., et al. (2015). “Intergroup transfer of females and social relationships between immigrants and residents in bonobo (Pan paniscus) societies,” in Dispersing primate females: life history and social strategies in Male-philopatric species primatology monographs. Eds. Furuichi T., Yamagiwa J., Aureli F. (Tokyo: Springer Japan), 127–164. doi: 10.1007/978-4-431-55480-6_6
Sapolsky R. M., Share L. J. (2004). Emergence of a peaceful culture in wild baboons. PLoS Biol. 2 (4), e106. doi: 10.1371/journal.pbio.0020124
Sasaki T., Biro D. (2017). Cumulative culture can emerge from collective intelligence in animal groups. Nat. Commun. 8 (1), 15049. doi: 10.1038/ncomms15049
Schuppli C., Forss S., Meulman E., Atmoko S. U., van Noordwijk M., van Schaik C. P. (2017). The effects of sociability on exploratory tendency and innovation repertoires in wild Sumatran and bornean orangutans. Sci. Rep. 7 (1), 15464. doi: 10.1038/s41598-017-15640-x
Schuppli C., Forss S. I. F., Meulman E. J. M., Zweifel N., Lee K. C., Rukmana E. (2016a). Development of foraging skills in two orangutan populations: needing to learn or needing to grow? Front. Zool. 13 (1), 43. doi: 10.1186/s12983-016-0178-5
Schuppli C., Graber S. M., Isler K., van Schaik C. P. (2016b). Life history, cognition and the evolution of complex foraging niches. J. Hum. Evol. 92, 91–100. doi: 10.1016/j.jhevol.2015.11.007
Schuppli C., Meulman E. J. M., Forss S. I. F., Aprilinayati F., van Noordwijk M. A., van Schaik C. P. (2016c). Observational social learning and socially induced practice of routine skills in immature wild orang-utans. Anim. Behav. 119, 87–98. doi: 10.1016/j.anbehav.2016.06.014
Schuppli C., van Schaik C. P. (2019). Animal cultures: how we’ve only seen the tip of the iceberg. Evol. Hum. Sci. 1, 1–13. doi: 10.1017/ehs.2019.1
Searle S. R., Speed F. M., Milliken G. A. (1980). Population marginal means in the linear model: an alternative to least squares means. Am. Statistician 34 (4), 216–221. doi: 10.1080/00031305.1980.10483031
Silk J. B., Brown G. R. (2004). “Sex ratios in primate groups,” in Sexual selection in primates- new and comparative perspectives Kappeler P. M., Schaik P.van C. (UK: Cambridge University Press), 253–265. doi: 10.1017/CBO9780511542459
Singleton I., van Schaik C. P. (2001). Orangutan home range size and its determinants in a Sumatran swamp forest. Int. J. Primatol. 22 (6), 877–911. doi: 10.1023/A:1012033919441
Stevens J. M. G., Vervaecke H., de Vries H., van Elsacker L. (2005). Peering is not a formal indicator of subordination in bonobos (Pan paniscus). Am. J. Primatol. 65 (3), 255–267. doi: 10.1002/ajp.20113
Stoinski T. S., Whiten A. (2003). Social learning by orangutans (Pongo abelii and Pongo pygmaeus) in a simulated food-processing task. J. Comp. Psychol. 117, 272–282. doi: 10.1037/0735-7036.117.3.272
Suzuki S., Hill D. A., Sprague D. S. (1998). Intertroop transfer and dominance rank structure of nonnatal Male Japanese macaques in yakushima, Japan. Int. J. Primatol. 19 (4), 703–722. doi: 10.1023/A:1020329010009
Toyokawa W., Whalen A., Laland K. N. (2019). Social learning strategies regulate the wisdom and madness of interactive crowds. Nat. Hum. Behav. 3 (2), 183–193. doi: 10.1038/s41562-018-0518-x
Utami Atmoko S. S., Goossens B., Bruford M. W., de Ruiter J. R., van Hooff J. A. R. A. M. (2002). Male Bimaturism and reproductive success in Sumatran orang-utans. Behav. Ecol. 13, 643–652. doi: 10.1007/s00265-015-1991-0
Utami Atmoko S. S., Singleton I., van Noordwijk M. A., van Schaik C. P., Mitra Setia T. (2009). “Male-Male relationships in orangutans,” in Orangutans: geographic variation in behavioral ecology and conservation Eds. Wich S. A., Utami Atmoko S. S., Mitra Setia T., van Schaik C. P. (New York: Oxford University Press), 225–233. doi: 10.1093/acprof:oso/9780199213276.003.0015
van de Waal E., Borgeaud C., Whiten A. (2013). Potent social learning and conformity shape a wild primate’s foraging decisions. SCIENCE 340 (6131), 483–485. doi: 10.1126/science.1232769
van de Waal E., Renevey N., Favre C. M., Bshary R. (2010). Selective attention to philopatric models causes directed social learning in wild vervet monkeys. Proc. R. Soc. B-Biological Sci. 277 (1691), 2105–2111. doi: 10.1098/rspb.2009.2260
van Noordwijk M. A., Sauren S. E. B., Nuzuar B., Abulani A., Morrogh-Bernard H. C., Utami Atmoko S. S., et al. (2009). “Development of independence: Sumatran and bornean orangutans compared,” in Orangutans: geographic variation in behavioral ecology and conservation. Eds. Wich S. A., Utami Atmoko S. S., Mitra Setia T., van Schaik C. P. (New York: Oxford University Press), 189–203. doi: 10.1093/acprof:oso/9780199213276.003.0012
van Noordwijk M. A., Arora N., Willems E. P., Dunkel L. P., Amda R. N., Mardianah N., et al. (2012). Female philopatry and its social benefits among bornean orangutans. Behav. Ecol. Sociobiol. 66 (6), 823–834. doi: 10.1007/s00265-012-1330-7
van Noordwijk M. A., Utami Atmoko S. S., Knott C. D., Kuze N., Morrogh-Bernard H. C., Oram F., et al. (2018). The slow ape: high infant survival and long interbirth intervals in wild orangutans. J. Hum. Evol. 125, 38–49. doi: 10.1016/j.jhevol.2018.09.004
van Noordwijk M. A., van Schaik C. P. (1985). Male Migration and rank acquisition in wild long-tailed macaques (Macaca fascicularis). Anim. Behav. 33 (Aug), 849–861. doi: 10.1016/S0003-3472(85)80019-1
van Noordwijk M. A., van Schaik C. P. (2005). Development of ecological competence in Sumatran orangutans. Am. J. Phys. Anthropol. 127 (1), 79–94. doi: 10.1002/ajpa.10426
van Schaik C. P. (1999). The socioecology of fission-fusion sociality in orangutans. Primates 40 (1), 69–86. doi: 10.1007/BF02557703
van Schaik C. P. (2010). “Social learning and culture in animals,” in Animal behaviour: evolution and mechanisms. Ed. Kappeler P. (Berlin, Heidelberg: Springer), 623–653. doi: 10.1007/978-3-642-02624-9_20
van Schaik C. P., Burkart J. M. (2011). Social learning and evolution: the cultural intelligence hypothesis. Philos. Trans. R. Soc. B: Biol. Sci. 366 (1567), 1008–1016. doi: 10.1098/rstb.2010.0304
van Schaik C. P., Ancrenaz M., Borgen G., Galdikas B., Knott C. D., Singleton I., et al. (2003). Orangutan cultures and the evolution of material culture. Science 299 (5603), 102–105. doi: 10.1126/science.1078004
Vogel E. R., Harrison M. E., Zulfa A., Bransford T. D., Alavi S. E., Husson S., et al. (2015). Nutritional differences between two orangutan habitats: implications for population density. PloS One 10 (10), e0138612. doi: 10.1371/journal.pone.0138612
Whiten A. (2017). Culture extends the scope of evolutionary biology in the great apes. Proc. Natl. Acad. Sci. 114 (30), 7790–7797. doi: 10.1073/pnas.1620733114
Whiten A. (2021). The burgeoning reach of animal culture. Science 372, eabe6514. doi: 10.1126/science.abe6514
Whiten A., Goodall J., McGrew W. C., Nishida T., Reynolds V., Sugiyama Y., et al. (1999). Cultures in chimpanzees. Nature 399, 682–685. doi: 10.1038/21415
Whiten A., Horner V., de Waal F. B. M. (2005). Conformity to cultural norms of tool use in chimpanzees. Nature 437 (7059), 737–740. doi: 10.1038/nature04047
Whiten A., van de Waal E. (2018). The pervasive role of social learning in primate lifetime development. Behav. Ecol. Sociobiol. 72 (5), 80. doi: 10.1007/s00265-018-2489-3
Wich S. A., Utami Atmoko S. S., Setia T. M., Rijksen H. D., Schürmann C., van Hooff J. A. R. A. M., et al. (2004). Life history of wild Sumatran orangutans (Pongo abelii). J. Hum. Evol. 47 (6), 385–398. doi: 10.1016/j.jhevol.2004.08.006
Wich S. A., Vogel E. R., Larsen M. D., Fredriksson G., Leighton M., Yeager C. P., et al. (2011). Forest fruit production is higher on Sumatra than on Borneo. PLoS One 6 (6), e21278. doi: 10.1371/journal.pone.0021278
Widdig A., Muniz L., Minkner M., Barth Y., Bley S., Ruiz-Lambides A., et al. (2017). Low incidence of inbreeding in a long-lived primate population isolated for 75 years. Behav. Ecol. Sociobiol. 71 (1), 1–15. doi: 10.1007/s00265-016-2236-6
Wilkinson A., Kuenstner K., Mueller J., Huber L. (2010). Social learning in a non-social reptile (Geochelone carbonaria). Biol. Lett. 6 (5), 614–616. doi: 10.1098/rsbl.2010.0092
Wilson M. L., Boesch C., Fruth B., Furuichi T., Gilby I. C., Hashimoto C., et al. (2014). Lethal aggression in Pan is better explained by adaptive strategies than human impacts. Nature 513 (7518), 414–417. doi: 10.1038/nature13727
Wrangham R. W., Glowacki L. (2012). Intergroup aggression in chimpanzees and war in nomadic hunter-gatherers. Hum. Nat. 23 (1), 5–29. doi: 10.1007/s12110-012-9132-1
Yamanashi Y., Bando H., Matsunaga M., Tanaka M., Nogami E., Hirata S. (2020). Development of bed-building behaviors in captive chimpanzees (Pan troglodytes): implication for critical period hypothesis and captive management. Primates 61 (5), 639–646. doi: 10.1007/s10329-020-00839-w
Keywords: observational social learning, peering, eavesdropping, dispersal, migrant male orangutans, skill learning in adults, social tolerance
Citation: Mörchen J, Luhn F, Wassmer O, Kunz JA, Kulik L, van Noordwijk MA, van Schaik CP, Rianti P, Utami Atmoko SS, Widdig A and Schuppli C (2023) Migrant orangutan males use social learning to adapt to new habitat after dispersal. Front. Ecol. Evol. 11:1158887. doi: 10.3389/fevo.2023.1158887
Received: 04 February 2023; Accepted: 02 June 2023;
Published: 05 July 2023.
Edited by:
J. Frances Kamhi, Denison University, United StatesReviewed by:
Rachna Reddy, Duke University, United StatesTetsuro Matsuzawa, California Institute of Technology, United States
Copyright © 2023 Mörchen, Luhn, Wassmer, Kunz, Kulik, van Noordwijk, van Schaik, Rianti, Utami Atmoko, Widdig and Schuppli. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Julia Mörchen, anVsaWFfbW9lcmNoZW5AZXZhLm1wZy5kZQ==
†These authors have contributed equally to this work and share last authorship