 Dustin R. Partridge
Dustin R. Partridge J. Alan Clark
J. Alan Clark- 1New York City Audubon Society, New York, NY, United States
- 2Department of Biological Sciences, Louis Calder Biological Field Station, Fordham University, Armonk, NY, United States
Global wildlife populations are in decline, in part, due to urbanization. However, in urban landscapes, green infrastructure such as green roofs are being created to provide habitat for wildlife. Green roof isolation, planting heterogeneity, and size can all influence wildlife biodiversity, as may the age of a green roof. When new habitat is created, wildlife use of these new habitats is expected to increase over time. To test this expectation for birds, we monitored bird activity prior to and after installation of small green roof plots on six buildings located within New York City parks. Contrary to expectations, bird activity and bird species richness did not increase after green roof plot installation, nor did they increase over a period of 4 years following installation. These unexpected results may reflect the relatively small size of the plots or the fact that the plots were on buildings located within urban parks. Bird activity and bird species richness varied widely between roofs, and the composition of rooftop bird species may have been more influenced by the characteristics of the surrounding landscapes than the presence of the green roof plots. These findings suggest that small urban green roofs within a larger and, potentially, higher quality habitat may not provide additional habitat for foraging birds. Urban green roofs have numerous ecological and environmental benefits, but the size and characteristics of landscapes surrounding a green roof need to be considered when installing green roofs as wildlife habitat.
Introduction
Wildlife is in decline worldwide (Hallmann et al., 2017; Lister and Garcia, 2018; Rosenberg et al., 2019; Sánchez-Bayo and Wyckhuys, 2019) due, in part, to urbanization (Guenat et al., 2019; Habel et al., 2019; Rosenberg et al., 2019; Sánchez-Bayo and Wyckhuys, 2019). Urban green spaces can help offset urbanization’s negative impacts on wildlife; therefore, understanding the ecological drivers of wildlife diversity, a measure of individual abundance and taxonomic richness, in urban green spaces is of particular importance (Oliver et al., 2011; Chiquet et al., 2013; Ferenc et al., 2013; Braaker et al., 2014; Parkins and Clark, 2015; Partridge and Clark, 2018; Forister et al., 2019; Leveau et al., 2019).
Wildlife diversity in a green space is influenced by multiple factors, including plant heterogeneity (Matteson et al., 2008; Hortal et al., 2009), green space size (Watson et al., 2005; Matteson, 2007; Leveau et al., 2019), isolation from other green spaces (Magura et al., 2001; Prugh et al., 2008), and age of the green space (Soga et al., 2014; Ma et al., 2015). Urban green spaces are generally isolated, of limited size, and either fragmented habitat relics (Soga et al., 2014) or newly built (Matteson et al., 2008; Rupprecht and Byrne, 2014; Partridge and Clark, 2018; Dromgold et al., 2020). Newly built green spaces include parks (McFrederick and LeBuhn, 2006; Nielsen et al., 2014), abandoned lots (Gardiner et al., 2013; Bonthoux et al., 2014), traffic medians (Pećarević et al., 2010), gardens (Vergnes et al., 2012; Barratt et al., 2015; Burks and Philpott, 2017; Goddard et al., 2017), and green roofs (MacIvor and Lundholm, 2011; Partridge and Clark, 2018; Partridge et al., 2020). Unlike in habitat relics, wildlife communities in newly built green spaces generally develop from colonization with installed substrates and plants (MacIvor and Ksiazek, 2015) or from immigration over time (Schrader and Böning, 2006; Fattorini et al., 2018; Perry et al., 2020), with age being an important driver of community composition (McIntyre, 2000). Consequently, the effect of age on wildlife diversity in newly built green spaces needs to be better understood.
The age of an urban green space is strongly associated with taxonomic richness, with richness usually increasing with green space age (Honnay et al., 1999; Fernández-Juricic, 2000; Ferenc et al., 2013; Nielsen et al., 2014). For example, the oldest urban green spaces, relic habitat fragments, have higher insect species richness and different species assemblages than the oldest built urban parks, while older built urban parks have higher insect species richness and abundance than newer built parks (Soga et al., 2014). Furthermore, older parks have more bird species than newer parks, with species richness increasing with increased park age (from 8 years old to over 300 years old) and plant succession (Fernández-Juricic, 2000). This relationship between green space age and taxonomic richness holds true in small urban green spaces, such as gardens that are between 4 and 48 years old (Burks and Philpott, 2017). Even unmanaged vacant lots can exhibit rapid increases in wildlife richness with increased age (i.e., within several years), though the rate of change depends on site soil conditions and the resulting plant community (Bonthoux et al., 2014).
While the relationship between green space age and wildlife diversity is strong in most urban green spaces, this relationship can be less important than other factors, such as green space size or isolation from other green spaces. For example, older relic habitat fragments had lower rodent diversity than newer fragments (Bolger et al., 1997), presumably due to a higher rate of local extinctions than recolonization as a result of isolation (Crooks et al., 2001). Butterfly and bee richness and abundance in urban gardens were more influenced by in-garden habitat conditions than garden age (Matteson and Langellotto, 2010). Furthermore, the relationship between green space age and wildlife diversity may not hold in habitats which are too small to host some species (Donnelly and Marzluff, 2004), with small (<0.5 hectare) green spaces predicted to have little value to highly mobile taxa, such as birds (Leveau et al., 2019; but see Narango et al., 2018).
Green roofs, roofs covered with an impermeable membrane, growing medium, and vegetation (Oberndorfer et al., 2007), capture stormwater (Gregoire and Clausen, 2011; Abualfaraj et al., 2018), reduce energy use (Santamouris, 2014; Alvizuri et al., 2017; Besir and Cuce, 2018), and can act as an effective tool for increasing wildlife habitat in urban landscapes (Cunningham and Liebezeit, 2015; Parkins and Clark, 2015; Ksiazek-Mikenas et al., 2018; Partridge and Clark, 2018; Dromgold et al., 2020; Partridge et al., 2020). However, most urban green roofs are small in size, isolated from other green spaces, and recently built (Stand and Peck, 2015; Treglia et al., 2018). To design urban green roofs that effectively provide habitat for wildlife, the ecological drivers of green roof wildlife communities, such as green roof size, isolation, and age, need to be better understood (Williams et al., 2014; Ksiazek-Mikenas et al., 2018).
Wildlife abundance and richness on urban green roofs can develop quickly following installation, possibly in part due to successional changes in green roof plant communities (Rowe et al., 2012; Ksiazek-Mikenas et al., 2018; Zhang et al., 2021) and substrate (Thuring and Dunnett, 2014). For example, arthropod abundance and richness increased in the 3 years following installation of a green roof in New York City, although arthropod abundance was higher in the year of installation, presumably due to arthropods being introduced to the roof with unsterilized and farm-raised Sedum mats (Partridge et al., 2019). Younger (2 – 3 years old) green roofs have more similar Collembolan community composition to each other than to older (10 – 12 years old) green roofs (Schrader and Böning, 2006). Furthermore, arthropod communities on older green roofs may, over time, become comparable to ground-level arthropod communities (Dromgold et al., 2020).
If arthropod abundance and richness increase with green roof age, green roof use by insectivorous birds (Partridge and Clark, 2018) should also increase over time. In Singapore, bird diversity increased with roof age, with older green roofs hosting more bird species than newer green roofs (Wang et al., 2017). However, as with other urban green spaces, factors such as size, isolation, and plant heterogeneity may be more important than green roof age in driving green roof wildlife community composition. For example, arthropod abundance and richness on shallow (<20 cm) green roofs in Germany did not increase with age; instead, green roof size, roof vegetation diversity, and the amount of surrounding green space were stronger predictors of arthropod richness than roof age (Ksiazek-Mikenas et al., 2018). Since urban green roofs can be used by urban avoiding, urban utilizing, and urban exploiting bird species (Blair, 1996; Johnston, 2001; Fischer et al., 2015; Partridge and Clark, 2018; Archer et al., 2019), as well as different feeding guilds (Partridge and Clark, 2018), characteristics of the roof and the landscape surrounding roofs are likely to influence bird community composition.
Documenting early successional changes in green roof bird communities is essential for predicting and assessing bird use of green roofs and for designing cities that can provide habitat for birds and other wildlife. To assess the effect of age on urban green roof biodiversity, we monitored bird communities on six small (96 m2), newly installed green roof plots in New York City over 4 years. Because the green roof plots in this study were smaller than the average-sized New York City green roof (334.5 m2) (Treglia et al., 2018), we also assessed whether small green roof plots can increase bird habitat like larger green roofs in New York City (Partridge and Clark, 2018). To understand if small green roof plots increase bird habitat, we monitored roofs before green roof plots were installed and following green roof plot installation, and we predicted that bird activity and richness would increase on roofs following green roof plot installation. To understand how bird communities changed over time, we monitored roofs for 3 of 4 years following installation and predicted that bird activity and richness would increase on roofs over time.
Understanding how wildlife communities change in the years following green roof installation is important as most urban green roofs in the Americas are relatively new. For example, as of 2014, over half of New York City’s green roof area was installed between 2010 and 2013 (Stand and Peck, 2015). Furthermore, in 2019, New York City enacted legislation that requires the roofs of most new construction and building additions to be 100% covered by green roof plantings or solar panels (Nyc Department of Buildings, 2019). Consequently, the number of newly built green roofs in New York City will soon grow rapidly. As the ecological and environmental benefits of urban green roofs become better quantified, other cities have already begun, or will soon begin, requiring installation of green roofs. Therefore, understanding how wildlife communities on newly built green roofs change over time is essential, and assessing the value of green roofs to wildlife is important (Braaker et al., 2017; Mayrand and Clergeau, 2018; Partridge and Clark, 2018; Filazzola et al., 2019; Partridge et al., 2019, 2020; Dromgold et al., 2020).
Materials and Methods
Site Description and Sampling Season
We monitored roofs on six recreation centers owned by the New York City Department of Parks and Recreation. The roofs were located across the five New York City boroughs (Figure 1). Recreation centers were two to three stories tall, and all roofs but one (Sorrentino Recreation Center) were located within a New York City park and surrounded on at least two sides by green space, which included trees (Figure 1).
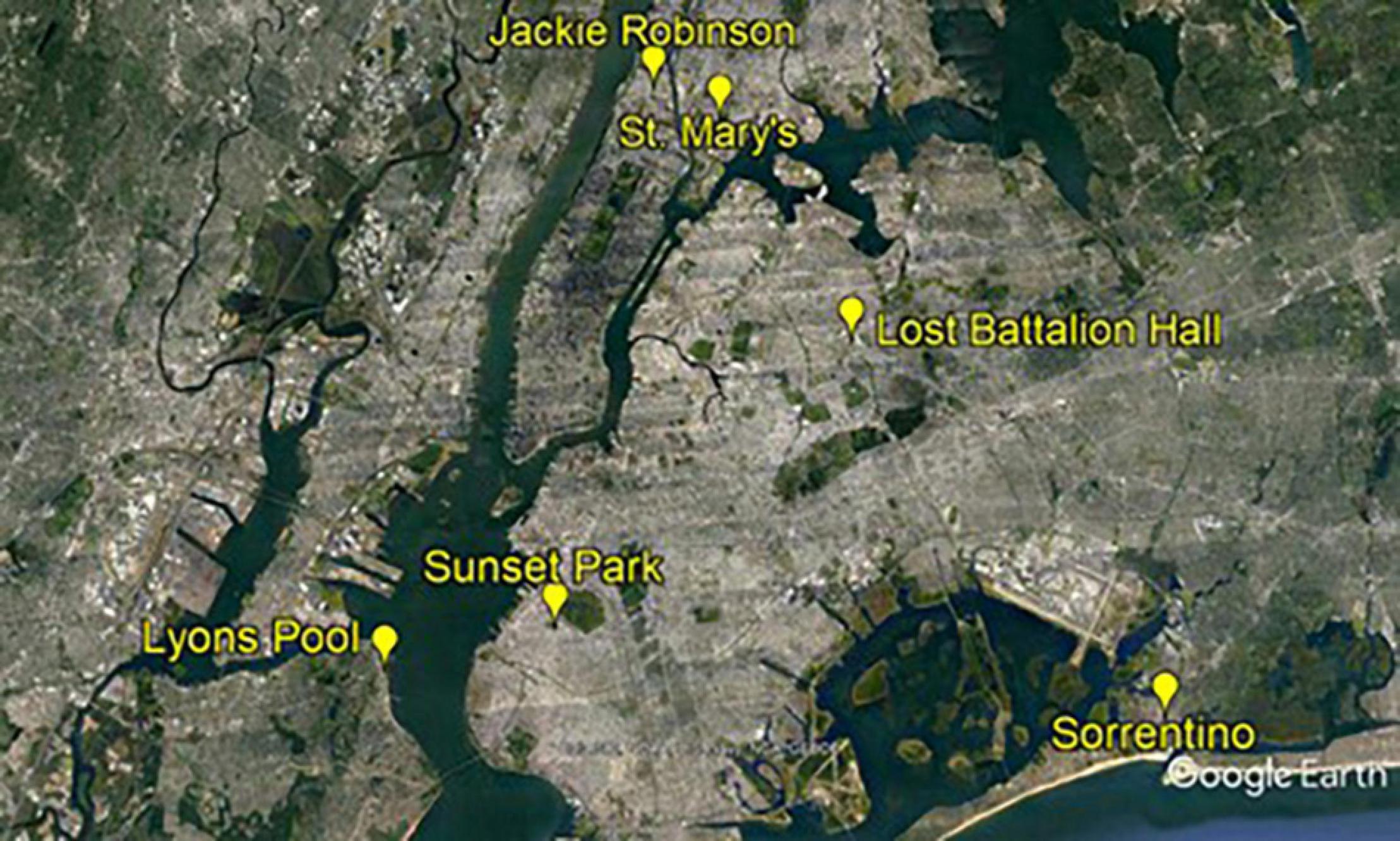
Figure 1. Location of six New York City Department of Parks and Recreation recreation centers used to survey bird activity and bird species richness before (2010) and after (2011, 2012, and 2014) installation of small, green roof plots.
We surveyed recreation center roofs during May from 2010 to 2014, excluding 2013. In the northern hemisphere, northward bird migration occurs in the spring and southward migration in the fall. In passerine (perching birds) and near passerine birds, spring migration generally occurs in a shorter amount of time (3–4 weeks) compared to fall migration which takes place over several months (Nilsson et al., 2013). To take advantage of the more compressed timeframe, we used spring migration for this study (Nilsson et al., 2013). Furthermore, using spring migration allowed for direct comparison to a previous rooftop bird study in New York City (i.e., Partridge and Clark, 2018).
Green roof plots were installed by the New York City Department of Parks and Recreation on recreation center roofs in the fall of 2010. Each green roof plot consisted of 12 planter boxes measuring 4 m x 2 m, for a total area of 96 m2 of each roof being covered by growing medium and vegetation (Figure 2). Each planter box was filled with either 10 cm or 15 cm of growing medium, divided in half, and planted with a subset of species from two native plant communities found in the New York City region (for a description of the experimental planting design see McGuire et al., 2013). We refer to these installations as “green roof plots,” as the roof area covered with plantings was substantially smaller than the entire roof area, with an average of 8.0% (ranging from 3.5 to 18.6%) of the total roof area being covered by plantings.

Figure 2. Example of green roof plots installed on New York City Department of Parks and Recreation recreation center roofs in fall of 2010, consisting of 12 planter boxes measuring 4 m x 2 m, for a total area of 96 m2. Roofs were surveyed for bird activity and bird species richness in May of 2010, 2011, 2012, and 2014.
Bird Monitoring
We used the same bird monitoring methods as Partridge and Clark (2018), which also examined bird activity on roofs in New York City. Bird activity and richness were determined by recording bird vocalizations using automated acoustic recorders (Songmeter SM2, Wildlife Acoustics, Maynard, MA, United States). Recordings were made in the first 2 h of the day (beginning one-half hour before civil sunrise) in order to record during the dawn chorus, the time when most birds peak in vocal activity (Staicer et al., 1996). Using Audacity® (1.3 Beta), we transformed sound files into a spectrogram – a visual depiction of sound in which the x-axis is time (sec) and the y-axis is frequency (kiloHertz, kHz). Once in spectrogram format, vocalizations were identified to species by visually locating bird vocalizations in the spectrogram and then identifying the vocalization to species acoustically.
To account for the continuous singing by certain species (e.g., northern mockingbirds, Mimus polyglottos), individual vocalizations were counted as those separated by at least five min. If vocalizations were simultaneous or otherwise obviously from two different individuals, both vocalizations were counted. Vocalizations of house sparrows (Passer domesticus) were not included in this analysis; their ubiquity and frequent vocalizations made them difficult to quantify acoustically (e.g., multiple overlapping vocalizations from different birds).
In addition, we included in our analyses only vocalizations that were sufficiently loud to produce strong spectrogram signatures, an indication that the bird was vocalizing near the acoustic recorder (i.e., on the roof). Vocalizations with relatively weak spectrogram signatures were not included as they may have come from birds in nearby ground-level vegetation.
We analyzed recordings from seven morning surveys each May between 2010 and 2014 (except for 2013) for all six sites with the exception of Sorrentino Recreation Center which was only surveyed in 2010 and 2011 due to acoustic recorder failure in 2012 and limited roof access in 2014. Recordings from every 5 days were analyzed. Days with rain were not included because rain produces noise interference and prohibits acoustic analysis. When rain occurred on the fifth day, the closest non-rain day was used. Bird activity was analyzed by calculating the number of vocalizations per hour for each species.
We classified bird species by habitat preference (e.g., forest, shoreline, and open woodland) based on data compiled in the Birds of North America Online (Poole, 2005). Classifying bird species by habitat preference allowed us to examine the influence of the landscape surrounding the roofs. We also classified bird species by their tolerance of urban landscapes, designating species as urban dwellers, urban utilizers, or urban avoiders (Blair, 1996; Fischer et al., 2015) based on Johnston (2001) and Archer et al. (2019). Classifying bird species by their tolerance of urban landscapes allowed us to evaluate whether green roof plots provide habitat for urban bird species or if they can provide habitat for species that might otherwise be absent in urban landscapes.
Statistical Analysis
To test for an increase in bird activity and bird richness before and after green roof plot installation we used a linear mixed effect model with year as the fixed effect and roof as the random effect with alpha set at 0.05. We also used a linear mixed effect model with year as the fixed effect and roof as the random effect to test for an increase in bird activity and richness on green roof plots over time. Results are presented ± SE.
Results
We found no changes in bird activity or richness in the season following the installation of green roof plots (F1,5 = 0.007, p = 0.94, and F1,5 = 0.07, p = 0.79, respectively), nor did we find a difference in bird activity (F1,13.1 = 1.79, p = 0.20) or bird species richness (F1,13 = 1.35, p = 0.31) on roofs in the years following green roof plot installation. In 2010, before green roof plots were installed, bird activity on the six roofs averaged 7.95 ± 1.49 vocalizations/hour (Table 1). After green roof plots were installed, average bird activity ranged from 6.51 ± 0.96 vocalizations/hour in 2012 to 8.27 ± 1.50 vocalizations/hour in 2014 (Table 1). Before green roof plots were installed, the number of bird species on the six roofs averaged 6.0 ± 0.80 species/roof. After green roof plots were installed, the average number of bird species ranged from 5.83 ± 1.44 species/roof in 2011 to 6.6 ± 0.95 species/roof in 2013 (Table 1).
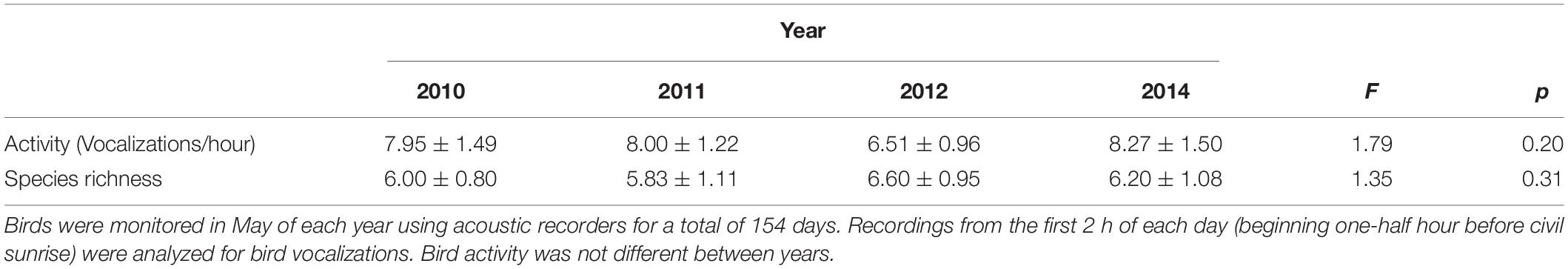
Table 1. Average bird activity (vocalizations/hour) and bird species richness on six roofs in New York City prior to, and 3 of 4 years after, green roof plot installation.
Species composition varied by roof pre- and post-installation, but urban dwellers were most common on all roofs, accounting for 38 to 85% of all activity on roofs (Figure 3). Urban utilizers accounted for 11 to 62% of activity, while urban avoiders were the least active on roofs, accounting for 0 to 4% of activity (Figure 3). European starlings (Sturnus vulgaris), an urban dweller, dominated vocalizations on all roofs except for St. Mary’s Recreation Center, both before and after green roof plot installation (Table 2). The next most common species was also an urban dweller, chimney swift (Chaetura pelagica), with the exception of Sorrentino Recreation Center, which was the only roof not located within a green space and was dominated by Larus gull species (Table 2). Species’ habitat requirements also varied by roof, but the majority of bird activity was due to species that used urban or edge/urban habitats, with the exception of Sorrentino Recreation Center, which was dominated by species which use shoreline habitats. At all sites other than Sorrentino Recreation Center, the next most active species required open woodland habitat (Figure 3).
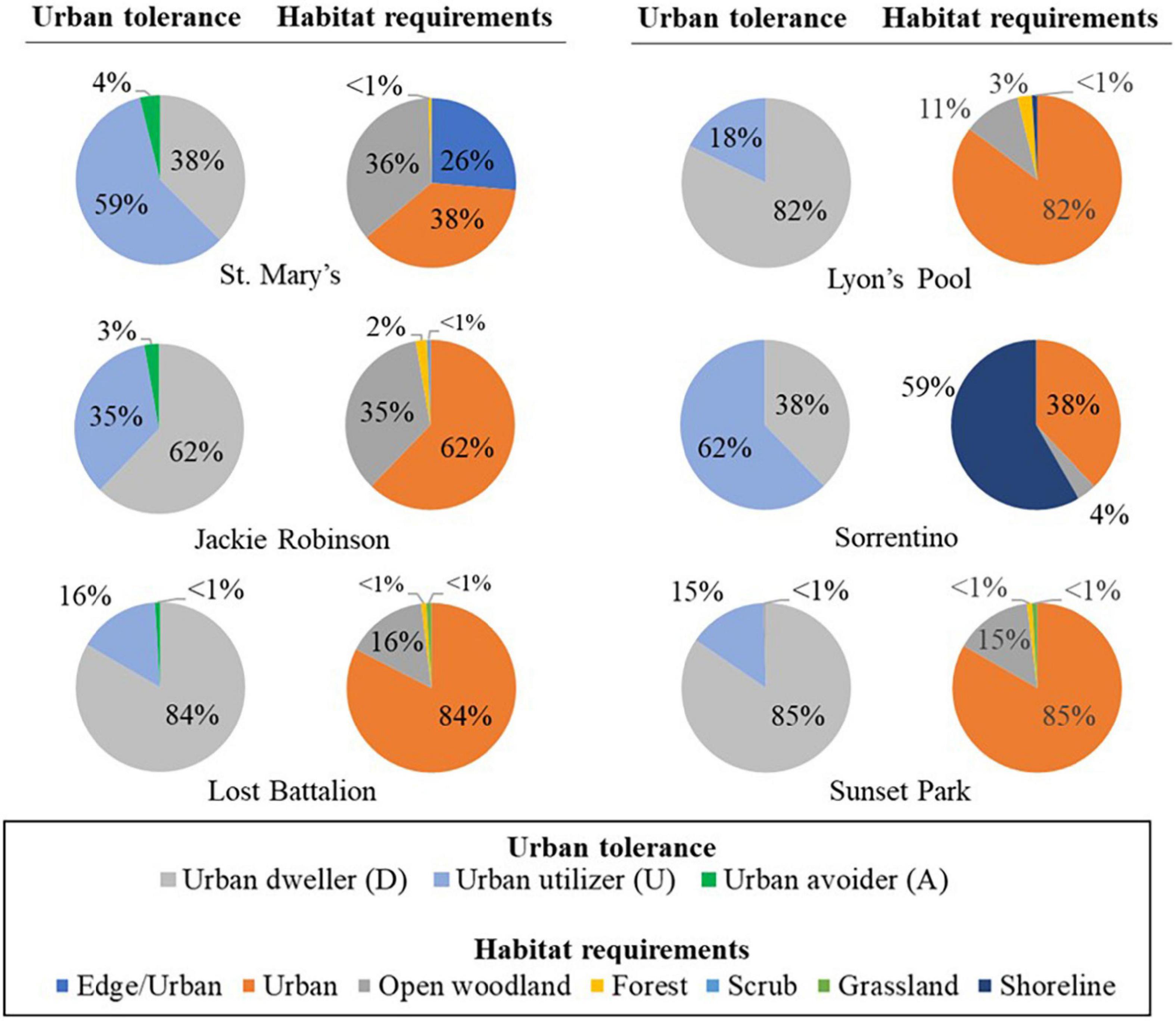
Figure 3. Composition of bird species on six New York City Department of Parks and Recreation recreation center roofs during May of 2010, 2011, 2012, and 2014 based on tolerance to urban landscape and habitat requirements. Species’ urban tolerance was determined using (Johnston, 2001; Fischer et al., 2015; Archer et al., 2019). Habitat requirements for each species was determined using the Birds of North America Online (Poole, 2005).
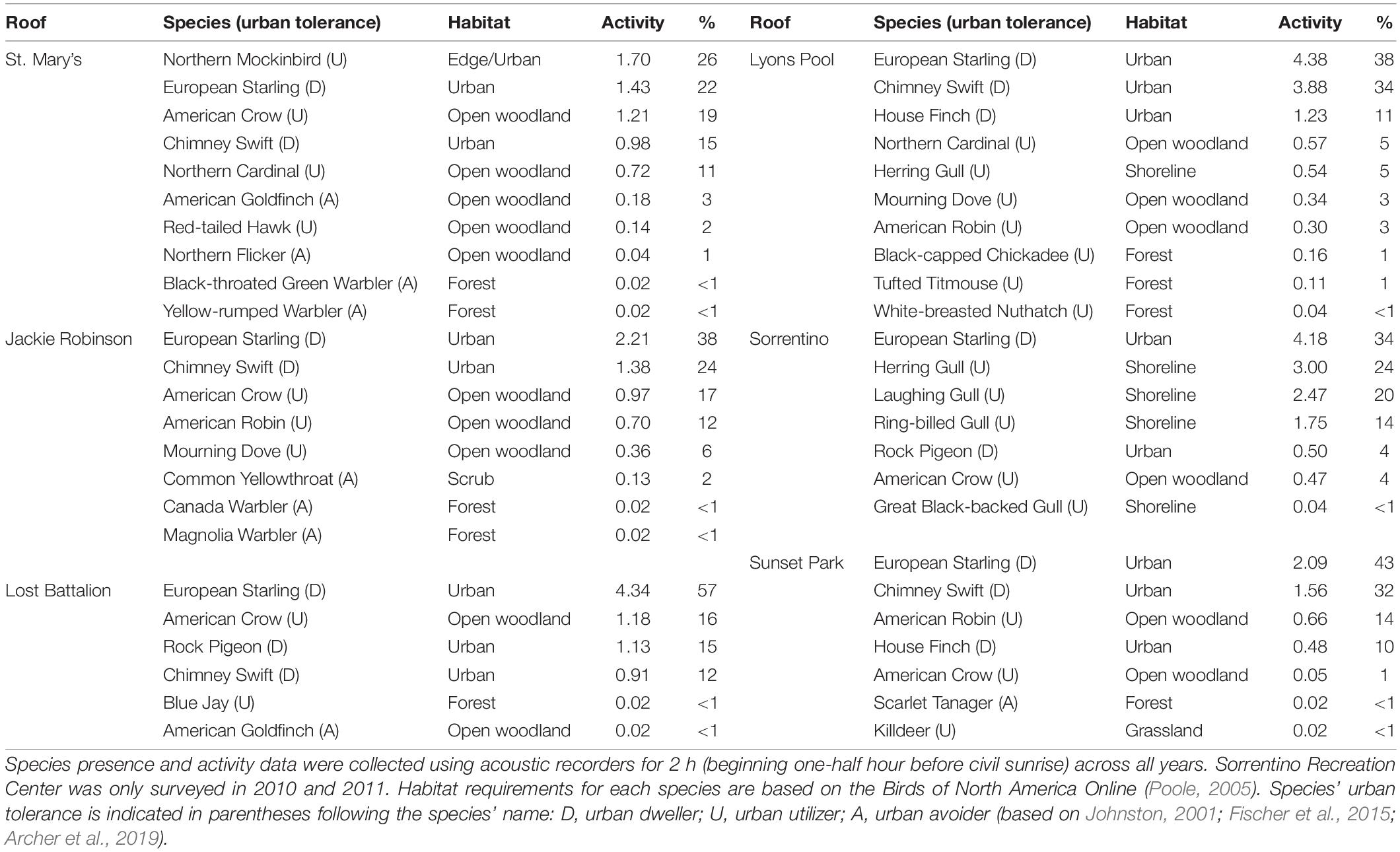
Table 2. Bird species composition, activity (vocalizations/hour), and percent activity (percent of the total activity on each roof) on six New York City Department of Parks and Recreation recreation center roofs during May of 2010, 2011, 2012, and 2014.
Saint Mary’s Recreation Center and Lyons Pool Recreation Center had the highest species richness across all years (10 species each), while Lost Battalion Hall Recreation Center had the lowest (six species) (Figure 4). Bird activity and richness varied between individual roofs resulting in roofs having a strong, but not significant, influence on bird abundance and richness (Z = 1.50, p = 0.06 and Z = 1.51, p = 0.06, respectively, Figure 4).
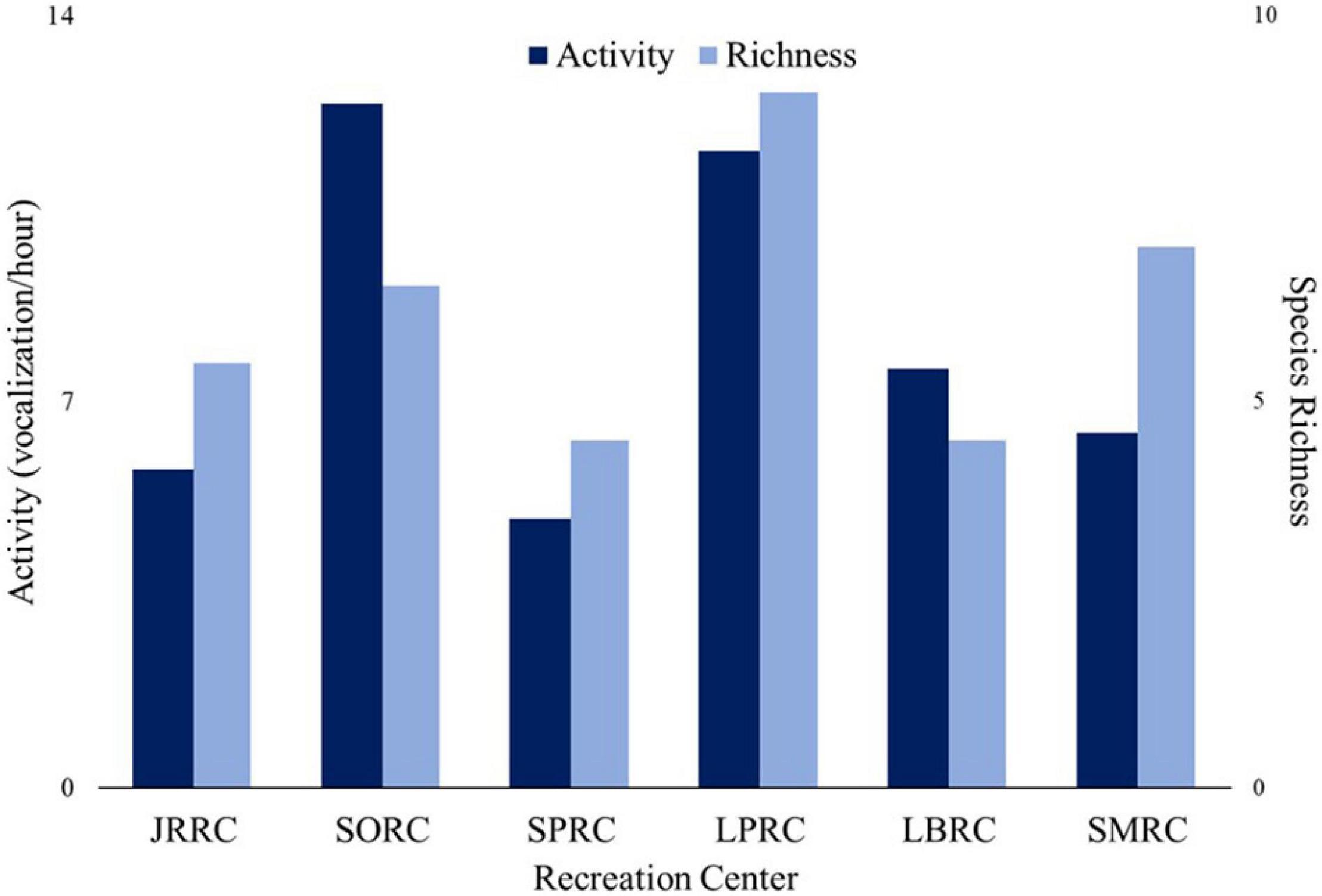
Figure 4. Average bird activity (vocalizations/hour) and bird species richness on roofs on six New York City Department of Parks and Recreation recreation center roofs. Roofs were monitored before (2010) and after (2011, 2012, and 2014) green roof plot installation. Sorrentino Recreation Center was only monitored in 2010 and 2011. JRRC, Jackie Robinson Recreation Center; SORC, Sorrentino Recreation Center; SPRC, Sunset Park Recreation Center; LPRC, Lyons Pool; LBRC, Lost Battalion Hall Recreation Center; SMRC, St. Mary’s Recreation Center.
Discussion
Contrary to our predictions, neither bird activity nor bird species richness increased following installation of green roof plots. Bird activity on the six roofs (ranging from 6.51 ± 0.96 vocalizations/hour in 2012 to 8.27 ± 1.50 vocalizations/hour in 2014) was comparable to bird activity on isolated New York City green roofs in an earlier New York City green roof study (averaging 5.77 ± 1.50 vocalizations/hour) (Partridge and Clark, 2018); however, this comparable activity was largely due to two common urban dweller species dominating roofs in this study (European starling and chimney swift), and neither species increased activity on roofs following green roof plot installation.
Unlike bird activity, bird richness following green roof plot installation in this study (ranging from 5.83 ± 1.44 to 6.60 ± 1.44 species/roof) was substantially lower than the average bird richness on other green roofs in New York City (19.50 ± 4.3 species/roof) (Partridge and Clark, 2018). Unexpectedly, bird richness on the green roof plots in this study was actually comparable to the average bird richness on conventional (non-green) roofs in New York City (7.75 ± 1.8 species/roof) (Partridge and Clark, 2018). Because bird richness did not increase following installation of green roof plots and because roofs had bird species richness similar to conventional roofs in New York City, the installation of green roof plots in this study likely did not provide a measurable habitat benefit to birds.
These results, which found that bird activity and richness did not increase with green roof plot age, are in contrast to the majority of studies that found older green roofs have higher species richness than younger roofs (Schrader and Böning, 2006; Wang et al., 2017; Dromgold et al., 2020), albiet the older roofs in these studies were much older than the 4 year old roofs examined in our study. The unexpected lack of increase over time in bird activity and richness in this study is similar to the results of a survey of arthropods on shallow-substrate urban green roofs (Ksiazek-Mikenas et al., 2018) which found that arthropod abundance and richness was not strongly influenced by green roof age but, rather, was more strongly influenced by green roof size, vegetation diversity, and surrounding green space.
The only study of which we are aware that examined the influence of green roof age on bird richness found that older green roofs (up to 28 years old) hosted more species than younger roofs (Wang et al., 2017). Perhaps surveying the green roof plots in this study for only 4 years after installation was not sufficient time to lead to a measurable increase in bird richness. However, in other urban green roof studies, bird activity and richness increased immediately after green roof installation. For example, following the installation of a large 27,316 m2 green roof in New York City, both bird activity and species richness increased in the 3 years following installation (Partridge et al., 2019).
The unexpected results of this study might also reflect the relatively small size of the green roof plots which, at 96 m2, were smaller than the average-sized (334.5 m2) New York City green roof (Treglia et al., 2018). In addition, the green roof plots in this study were only 6.4 to 19.2% of the size of green roofs used in a previous study in New York City that found green roofs provide higher quality habitat for birds than conventional roofs (Partridge and Clark, 2018). Many bird species, because of their habitat requirements, generally do not use small green spaces (Tilghman, 1987; Donnelly and Marzluff, 2004), and the green roof plots in this study may have been too small to provide additional habitat for birds (Leveau et al., 2019). Thus, our results suggest that, on small urban green roofs or green roof plots, size may also be a strong driver (or limiter) of bird activity and richness.
To further understand bird activity and richness on green roofs, roof isolation from other green spaces and characteristics of the surrounding landscape must also be considered. The isolation of an urban green space is an important driver of wildlife diversity in urban environments, with decreased isolation usually associated with increased species richness (Fernández-Juricic, 2000; Lizée et al., 2012; Chang and Lee, 2015). However, all but one of the roofs used in this study were located inside larger urban green spaces, and most roofs in this study were at low enough elevation that nearby tree canopies were taller than the roofs (Figure 2). Given these factors, and considering that green roofs are generally of lower quality habitat than ground-level green space (MacIvor and Lundholm, 2011; Braaker et al., 2014; Williams et al., 2014; Parkins et al., 2016; Ksiazek-Mikenas et al., 2018; Dromgold et al., 2020), the green roof plots in this study likely had limited habitat benefit to birds.
The results of this study are also consistent with those of Washburn et al. (2016) which found that bird activity and richness on a large green roof at Chicago O’Hare International Airport was comparable to a nearby conventional roof. The roofs in both this study and the roof in Washburn et al. (2016) were partially surrounded by green space which could have resulted in reduced bird use of the green roofs. The landscape surrounding a green space is a strong driver of wildlife in urban areas (Fernández-Juricic, 2000; Pennington and Blair, 2011), and, thus, the landscape surrounding the roofs in this study and in the Washburn et al. (2016) study was more likely the driver of local bird activity and richness. Consequently, green roof and green roof plot installations did not measurably increase local bird habitat and did not result in increased bird use of those roofs.
Green roofs that provide habitat for birds in New York City have bird communities that are more similar to other green roofs than nearby conventional roofs (Partridge and Clark, 2018). But in this study, bird species composition on roofs appears to have been strongly influenced by surrounding landscapes. The composition of bird species in this study, based on habitat requirements and their response to urban landscapes (Johnston, 2001; Fischer et al., 2015; Archer et al., 2019), likely reflected the local habitat surrounding the roof (Figure 3). For example, the Sorrentino Recreation Center is not located in a park but is less than one km from a coastal park and one km from Jamaica Bay National Wildlife Refuge, which largely consists of open water and saltmarsh; not surprisingly, the recordings from this roof were dominated by gull species. In contrast, St. Mary’s Recreation Center is set within a 35-hectare wooded park, and the recordings from this roof consisted largely of forest species (Table 2 and Figure 3). Bird communities in urban green spaces are influenced by surrounding habitats (Fernández-Juricic, 2000; Pennington and Blair, 2011), with bird community composition in small, new green spaces being driven by landscape conditions (Fernández-Juricic, 2000). Thus, bird species composition on roofs in this study was likely more strongly associated with characteristics of the green spaces in the surrounding landscape than the presence of the green roof plots.
Another factor that may have resulted in the relatively low species richness recorded on the roofs in this study could be that we analyzed recordings for only 2 h on sample days, beginning one-half hour prior to civil sunrise. Our results did not include any birds that vocalized on the roofs later in the day. As noted earlier, the dawn chorus is when the majority of passerine birds are most vocal (Staicer et al., 1996). However, while bird vocalization activity is high during the dawn chorus, movement and foraging are low, with foraging activity increasing with increasing daylight (Kacelnik, 1979; Berg et al., 2006). Birds in this study may have been vocalizing in the surrounding habitats during the dawn chorus and foraging on the roofs later in the day; however, we would not have recorded later foraging birds, and, consequently, their presence or activity would not be captured by our methods. Furthermore, our study only examined bird activity on roofs during spring, and it is possible that the small green roof plots in this study provide habitat for foraging birds during the summer, like other green roofs in New York City (Partridge and Clark, 2018).
The results of this study highlight the need for additional research to examine the influence of green roof size as well as surrounding habitat on urban green roof bird communities. Previous research demonstrated that isolated urban green roofs can be a useful tool for bird conservation (Partridge and Clark, 2018), and if the green roof plots in this study were installed in isolation, they may have been more useful to birds as the only green space available. Alternatively, if the green roof plots in this study were larger and designed to have a habitat similar to the surrounding green spaces, they may have been more heavily used by birds. Our findings suggest that, despite the numerous environmental and ecological benefits urban green roofs provide, small urban green roofs that are built in, or immediately adjacent to, larger green spaces may not provide additional habitat for birds if the green roof habitat is of lesser quality than the adjacent larger green space.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author Contributions
DP was responsible for conceptualization, formal analysis, and writing the original draft. JC was responsible for review and editing, supervision, and funding acquisition. Both authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank the New York City Department of Parks and Recreation for granting us access to their recreation center roofs and Matt Palmer of Columbia University for designing the green roof plots that were used in this study. We also thank the reviewers for their valuable comments.
References
Abualfaraj, N., Cataldo, J., Elborolosy, Y., Fagan, D., Woerdeman, S., Carson, T., et al. (2018). Monitoring and modeling the long-term rainfall-runoff response of the Jacob K. Javits Center Green Roof. Water 10, 1–23. doi: 10.3390/w10111494
Alvizuri, J., Cataldo, J., Smalls-Mantey, L. A., and Montalto, F. A. (2017). Green roof thermal buffering: insights derived from fixed and portable monitoring equipment. Energy Build. 151, 455–468. doi: 10.1016/j.enbuild.2017.06.020
Archer, J.-M. J., Hostetler, M. E., Acomb, G., and Blair, R. (2019). A systematic review of forest bird occurrence in North American forest fragments and the built environment. Landsc. Urban Plan. 185, 1–23. doi: 10.1016/j.landurbplan.2019.01.005
Barratt, B. I., Dickinson, K. J., Freeman, C., Porter, S., Johnstone, P. D., Wing, J., et al. (2015). Biodiversity of Coleoptera and other invertebrates in urban gardens: a case study in a New Zealand city. Insect Conserv. Divers. 8, 428–437. doi: 10.1111/icad.12120
Berg, K. S., Brumfield, R. T., and Apanius, V. (2006). Phylogenetic and ecological determinants of the neotropical dawn chorus. Proc. Biol. Sci. 273, 999–1005. doi: 10.1098/rspb.2005.3410
Besir, A. B., and Cuce, E. (2018). Green roofs and facades: a comprehensive review. Renew. Sustain. Energy Rev. 82, 915–939. doi: 10.1016/j.rser.2017.09.106
Blair, R. B. (1996). Land use and avian species diversity along an urban gradient. Ecol. Appl. 6, 506–519. doi: 10.2307/2269387
Bolger, D. T., Alberts, A. C., Sauvajot, R. M., Potenza, P., McCalvin, C., Tran, D., et al. (1997). Response of rodents to habitat fragmentation in coastal southern California. Ecol. Appl. 7, 552–563. doi: 10.1890/1051-0761(1997)007[0552:RORTHF]2.0.CO;2
Bonthoux, S., Brun, M., Di Pietro, F., Greulich, S., and Bouché-Pillon, S. (2014). How can wastelands promote biodiversity in cities? A review. Landsc. Urban Plan. 132, 79–88. doi: 10.1016/j.landurbplan.2014.08.010
Braaker, S., Ghazoul, J., Obrist, M. K., and Moretti, M. (2014). Habitat connectivity shapes urban arthropod communities: the key role of green roofs. Ecology 95, 1010–1021. doi: 10.1890/13-0705.1
Braaker, S., Obrist, M. K., Ghazoul, J., and Moretti, M. (2017). Habitat connectivity and local conditions shape taxonomic and functional diversity of arthropods on green roofs. J. Anim. Ecol. 86, 521–531. doi: 10.1111/1365-2656.12648
Burks, J. M., and Philpott, S. M. (2017). Local and landscape drivers of parasitoid abundance, richness, and composition in urban gardens. Environ. Entomol. 46, 201–209. doi: 10.1093/ee/nvw175
Chang, H.-Y., and Lee, Y.-F. (2015). Effects of area size, heterogeneity, isolation, and disturbances on urban park avifauna in a highly populated tropical city. Urban Ecosyst. 19, 257–274. doi: 10.1007/s11252-015-0481-5
Chiquet, C., Dover, J. W., and Mitchell, P. (2013). Birds and the urban environment: the value of green walls. Urban Ecosyst. 16, 453–462. doi: 10.1007/s11252-012-0277-9
Crooks, K. R., Suarez, A. V., Bolger, D. T., and Soulé, M. E. (2001). Extinction and colonization of birds on habitat islands. Conserv. Biol. 15, 159–172. doi: 10.1111/j.1523-1739.2001.99379.x
Cunningham, C., and Liebezeit, J. (2015). Portland’s Ecoroof Avian Monitoring Project 2012-14 Final Report. Portland, OR: Environmental Services.
Donnelly, R., and Marzluff, J. M. (2004). Importance of reserve size and landscape context to urban bird conservation. Conserv. Biol. 18, 733–745. doi: 10.1111/j.1523-1739.2004.00032.x
Dromgold, J. R., Threlfall, C. G., Norton, B. A., and Williams, N. S. (2020). Green roof and ground-level invertebrate communities are similar and are driven by building height and landscape context. J. Urban Ecol. 6:juz024. doi: 10.1093/jue/juz024
Fattorini, S., Mantoni, C., De Simoni, L., and Galassi, D. M. (2018). Island biogeography of insect conservation in urban green spaces. Environ. Conserv. 45, 1–10. doi: 10.1017/S0376892917000121
Ferenc, M., Sedláček, O., and Fuchs, R. (2013). How to improve urban greenspace for woodland birds: site and local-scale determinants of bird species richness. Urban Ecosyst. 17, 625–640. doi: 10.1007/s11252-013-0328-x
Fernández-Juricic, E. (2000). Bird community composition patterns in urban parks of Madrid: the role of age, size and isolation. Ecol. Res. 15, 373–383. doi: 10.1046/j.1440-1703.2000.00358.x
Filazzola, A., Shrestha, N., and MacIvor, J. S. (2019). The contribution of constructed green infrastructure to urban biodiversity: a synthesis and meta-analysis. J. Appl. Ecol. 56, 2131–2143. doi: 10.1111/1365-2664.13475
Fischer, J. D., Schneider, S. C., Ahlers, A. A., and Miller, J. R. (2015). Categorizing wildlife responses to urbanization and conservation implications of terminology. Conserv. Biol. 29, 1246–1248. doi: 10.1111/cobi.12451
Forister, M. L., Pelton, E. M., and Black, S. H. (2019). Declines in insect abundance and diversity: we know enough to act now. Conserv. Sci. Pract. 1:e80. doi: 10.1111/csp2.80
Gardiner, M. M., Burkman, C. E., and Prajzner, S. P. (2013). The value of urban vacant land to support arthropod biodiversity and ecosystem services. Environ. Entomol. 42, 1123–1136. doi: 10.1603/EN12275
Goddard, M. A., Ikin, K., and Lerman, S. B. (2017). “Ecological and social factors determining the diversity of birds in residential yards and gardens,” in Ecology and Conservation of Birds in Urban Environments, eds E. Murgui and M. Hedblom (Cham: Springer International Publishing).
Gregoire, B. G., and Clausen, J. C. (2011). Effect of a modular extensive green roof on stormwater runoff and water quality. Ecol. Eng. 37, 963–969. doi: 10.1016/j.ecoleng.2011.02.004
Guenat, S., Kunin, W. E., Dougill, A. J., and Dallimer, M. (2019). Effects of urbanisation and management practices on pollinators in tropical Africa. J. Appl. Ecol. 56, 214–224. doi: 10.1111/1365-2664.13270
Habel, J. C., Samways, M. J., and Schmitt, T. (2019). Mitigating the precipitous decline of terrestrial European insects: requirements for a new strategy. Biodivers. Conserv. 28, 1343–1360. doi: 10.1007/s10531-019-01741-8
Hallmann, C. A., Sorg, M., Jongejans, E., Siepel, H., Hofland, N., Schwan, H., et al. (2017). More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS One 12:e0185809. doi: 10.1371/journal.pone.0185809
Honnay, O., Hermy, M., and Coppin, P. (1999). Effects of area, age and diversity of forest patches in Belgium on plant species richness, and implications for conservation and reforestation. Biol. Conserv. 87, 73–84. doi: 10.1016/S0006-3207(98)00038-X
Hortal, J., Triantis, K. A., Meiri, S., Thébault, E., and Sfenthourakis, S. (2009). Island species richness increases with habitat diversity. Am. Nat. 174, E205–E217. doi: 10.1086/645085
Johnston, R. F. (2001). “Synanthropic birds of North America,” in Avian Ecology in an Urbanizing World, eds J. M. Marzluff, R. Bowman, and R. Donnelly (Norwell, MA: Kluwer Academic). doi: 10.1007/978-1-4615-1531-9_3
Kacelnik, A. (1979). The foraging efficiency of great tits (Parus major L.) in relation to light intensity. Anim. Behav. 27, 237–241. doi: 10.1016/0003-3472(79)90143-X
Ksiazek-Mikenas, K., Herrmann, J., Menke, S. B., and Köhler, M. (2018). If you build it, will they come? Plant and arthropod diversity on urban green roofs over time. Urban Nat. 1, 52–72.
Leveau, L. M., Ruggiero, A., Matthews, T. J., and Isabel Bellocq, M. (2019). A global consistent positive effect of urban green area size on bird richness. Avian Res. 10, 1–14. doi: 10.1186/s40657-019-0168-3
Lister, B. C., and Garcia, A. (2018). Climate-driven declines in arthropod abundance restructure a rainforest food web. Proc. Natl. Acad. Sci. U.S.A. 115, E10397–E10406. doi: 10.1073/pnas.1722477115
Lizée, M.-H., Manel, S., Mauffrey, J.-F., Tatoni, T., and Deschamps-Cottin, M. (2012). Matrix configuration and patch isolation influences override the species–area relationship for urban butterfly communities. Landsc. Ecol. 27, 159–169. doi: 10.1007/s10980-011-9651-x
Ma, L., Huang, M., Shen, Y., Cao, H., Wu, L., Ye, H., et al. (2015). Species diversity and community structure in forest fragments of Guangzhou. South China. J. Trop. For. Sci. 27, 148–157.
MacIvor, J. S., and Lundholm, J. (2011). Insect species composition and diversity on intensive green roofs and adjacent level-ground habitats. Urban Ecosyst. 14, 225–241. doi: 10.1007/s11252-010-0149-0
MacIvor, J. S., and Ksiazek, K. (2015). “Invertebrates on green roofs,” in Green Roof Ecosystems, ed. R. Sutton (New York, NY: Springer International Publishing).
Magura, T., Ködöböcz, V., and Tóthmérész, B. (2001). Effects of habitat fragmentation on carabids in forest patches. J. Biogeogr. 28, 129–138. doi: 10.1046/j.1365-2699.2001.00534.x
Matteson, K. C. (2007). Diversity And Conservation Of Insects In Urban Gardens: Theoretical And Applied Implications. Doctoral Dissertation. Bronx, NY: Fordham University.
Matteson, K. C., and Langellotto, G. A. (2010). Determinates of inner city butterfly and bee species richness. Urban Ecosyst. 13, 333–347. doi: 10.1007/s11252-010-0122-y
Matteson, K. C., Ascher, J. S., and Langellotto, G. A. (2008). Bee richness and abundance in New York City urban gardens. Ann. Entomol. Soc. Am. 101, 140–150. doi: 10.1603/0013-8746(2008)101[140:BRAAIN]2.0.CO;2
Mayrand, F., and Clergeau, P. (2018). Green roofs and green walls for biodiversity conservation: a contribution to urban connectivity? Sustainability 10:985. doi: 10.3390/su10040985
McFrederick, Q., and LeBuhn, G. (2006). Are urban parks refuges for bumble bees Bombus spp? Hymenoptera 129, 372–382. doi: 10.1016/j.biocon.2005.11.004
McGuire, K. L., Payne, S. G., Palmer, M. I., Gillikin, C. M., Keefe, D., Kim, S. J., et al. (2013). Digging the New York City skyline: soil fungal communities in green roofs and city parks. PLoS One 8:e58020. doi: 10.1371/journal.pone.0058020
McIntyre, N. E. (2000). Ecology of urban arthropods: a review and a call to action. Ann. Entomol. Soc. Am. 93, 825–835. doi: 10.1603/0013-8746(2000)093[0825:EOUAAR]2.0.CO;2
Narango, D. L., Tallamy, D. W., and Marra, P. P. (2018). Nonnative plants reduce population growth of an insectivorous bird. Proc. Natl. Acad. Sci. 45, 11549–11554. doi: 10.1073/pnas.1809259115
Nielsen, A. B., van den Bosch, M., Maruthaveeran, S., and van den Bosch, C. K. (2014). Species richness in urban parks and its drivers: a review of empirical evidence. Urban Ecosyst. 17, 305–327. doi: 10.1007/s11252-013-0316-1
Nilsson, C., Klaassen, R. H., and Alerstam, T. (2013). Differences in speed and duration of bird migration between spring and autumn. Am. Nat. 181, 837–845. doi: 10.1086/670335
Nyc Department of Buildings (2019). Buildings Bulletin 2019-010. New York City, NY: Construction Codes.
Oberndorfer, E., Lundholm, J., Bass, B., Coffman, R. R., Doshi, H., Dunnett, N., et al. (2007). Green roofs as urban ecosystems: ecological structures, functions, and services. Bioscience 57, 823–833. doi: 10.1641/B571005
Oliver, A. J., Hong-Wa, C., Devonshire, J., Olea, K. R., Rivas, G. F., and Gahl, M. K. (2011). Avifauna richness enhanced in large, isolated urban parks. Landsc. Urban Plan. 102, 215–225. doi: 10.1016/j.landurbplan.2011.04.007
Parkins, K. L., and Clark, J. A. (2015). Green roofs provide habitat for urban bats. Glob. Ecol. Conserv. 4, 349–357. doi: 10.1016/j.gecco.2015.07.011
Parkins, K. L., Mathios, M., McCann, C., and Clark, J. A. (2016). Bats in the bronx: acoustic monitoring of bats in New York City. Urban Nat. 10, 1–16.
Partridge, D. R., and Clark, J. A. (2018). Urban green roofs provide habitat for migrating and breeding birds and their arthropod prey. PLoS One 13:e0202298. doi: 10.1371/journal.pone.0202298
Partridge, D. R., Parkins, K. L., and Elbin, S. B. (2019). An Ecological Assessment of the Jacob K. Javits Center Green Roof. New York, NY: New York City Audubon Society.
Partridge, D. R., Parkins, K. L., Elbin, S. B., and Clark, J. A. (2020). Moth abundance correlates with bat activity on an urban green roof. Northeast. Nat. 27, 77–89. doi: 10.1656/045.027.0107
Pećarević, M., Danoff-Burg, J., and Dunn, R. R. (2010). Biodiversity on Broadway–enigmatic diversity of the societies of ants (Formicidae) on the streets of New York City. PLoS One 5:e13222. doi: 10.1371/journal.pone.0013222
Pennington, D. N., and Blair, R. B. (2011). Habitat selection of breeding riparian birds in an urban environment: untangling the relative importance of biophysical elements and spatial scale. Divers. Distrib. 17, 506–518. doi: 10.1111/j.1472-4642.2011.00750.x
Perry, K. I., Hoekstra, N. C., Delgado de la flor, Y. A., and Gardiner, M. M. (2020). Disentangling landscape and local drivers of ground-dwelling beetle community assembly in an urban ecosystem. Ecol. Appl. 30:e02191. doi: 10.1002/eap.2191
Prugh, L. R., Hodges, K. E., Sinclair, A. R., and Brashares, J. S. (2008). Effect of habitat area and isolation on fragmented animal populations. Proc. Natl. Acad. Sci. U.S.A. 105, 20770–20775. doi: 10.1073/pnas.0806080105
Rosenberg, K. V., Dokter, A. M., Blancher, P. J., Sauer, J. R., Smith, A. C., Smith, P. A., et al. (2019). Decline of the North American avifauna. Science 366, 120–124. doi: 10.1126/science.aaw1313
Rowe, D. B., Getter, K. L., and Durhman, A. K. (2012). Effect of green roof media depth on Crassulacean plant succession over seven years. Landsc. Urban Plan. 104, 310–319. doi: 10.1016/j.landurbplan.2011.11.010
Rupprecht, C. D., and Byrne, J. A. (2014). Informal urban green-space: comparison of quantity and characteristics in Brisbane, Australia and Sapporo, Japan. PLoS One 9:e99784. doi: 10.1371/journal.pone.0099784
Sánchez-Bayo, F., and Wyckhuys, K. A. (2019). Worldwide decline of the entomofauna: a review of its drivers. Biol. Conserv. 232, 8–27. doi: 10.1016/j.biocon.2019.01.020
Santamouris, M. (2014). Cooling the cities–a review of reflective and green roof mitigation technologies to fight heat island and improve comfort in urban environments. Solar Energy 103, 682–703. doi: 10.1016/j.solener.2012.07.003
Schrader, S., and Böning, M. (2006). Soil formation on green roofs and its contribution to urban biodiversity with emphasis on Collembolans. Pedobiologia 50, 347–356. doi: 10.1016/j.pedobi.2006.06.003
Soga, M., Yamaura, Y., Koike, S., and Gaston, K. J. (2014). Woodland remnants as an urban wildlife refuge: a cross-taxonomic assessment. Biodivers. Conserv. 23, 649–659. doi: 10.1007/s10531-014-0622-9
Staicer, C., Spector, D., and Horn, A. (1996). “The dawn chorus and other diel patterns in acoustic signaling,” in Ecology and Evolution of Acoustic Communication in Birds, eds D. E. Kroodsma and E. H. Miller (Ithaca, NY: Cornell University Press).
Stand, B., and Peck, S. (2015). 2014 Annual Green Roof Industry Survey. Toronto, Ont: Green Roofs for Healthy Cities.
Thuring, C. E., and Dunnett, N. (2014). Vegetation composition of old extensive green roofs (from 1980s Germany). Ecol. Process. 3, 1–11. doi: 10.1186/2192-1709-3-4
Tilghman, N. G. (1987). Characteristics of urban woodlands affecting breeding bird diversity and abundance. Landsc. Urban Plan. 14, 481–495. doi: 10.1016/0169-2046(87)90061-2
Treglia, M. L., McPhearson, T., Sanderson, E. W., Yetman, G., and Maxwell, E. N. (2018). Green Roofs Footprints for New York City, Assembled from Available Data and remote Sensing (Version 1.0.0) [Data set]. Zendoo. doi: 10.5281/zenodo.1469674
Vergnes, A., Viol, I. L., and Clergeau, P. (2012). Green corridors in urban landscapes affect the arthropod communities of domestic gardens. Biol. Conserv. 145, 171–178. doi: 10.1016/j.biocon.2011.11.002
Wang, J. W., Poh, C. H., Tan, C. Y. T., Lee, V. N., Jain, A., and Webb, E. L. (2017). Building biodiversity: drivers of bird and butterfly diversity on tropical urban roof gardens. Ecosphere 8:e01905. doi: 10.1002/ecs2.1905
Washburn, B. E., Swearingin, R. M., Pullins, C. K., and Rice, M. E. (2016). Composition and diversity of avian communities using a new urban habitat: green roofs. Environ. Manage. 57, 1230–1239.
Watson, J. E. M., Whittaker, R. J., and Freudenberger, D. (2005). Bird community responses to habitat fragmentation: how consistent are they across landscapes? J. Biogeogr. 32, 1353–1370. doi: 10.1111/j.1365-2699.2005.01256.x
Williams, N. S., Lundholm, J., and MacIvor, J. S. (2014). Do green roofs help urban biodiversity conservation? J. Appl. Ecol. 51, 1643–1649. doi: 10.1111/1365-2664.12333
Keywords: green roofs, birds, succession, area, biodiversity, urban, conservation
Citation: Partridge DR and Clark JA (2022) Small Urban Green Roof Plots Near Larger Green Spaces May Not Provide Additional Habitat for Birds. Front. Ecol. Evol. 10:779005. doi: 10.3389/fevo.2022.779005
Received: 17 September 2021; Accepted: 23 February 2022;
Published: 25 March 2022.
Edited by:
Maria Manso, Universidade de Lisboa, PortugalReviewed by:
Bracha Schindler, University of Haifa, IsraelOlyssa Starry, Portland State University, United States
Copyright © 2022 Partridge and Clark. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dustin R. Partridge, RHBhcnRyaWRnZUBueWNhdWR1Ym9uLm9yZw==