 Yahong Chen1,2
Yahong Chen1,2 Jingjing Ji
Jingjing Ji Zhifeng Liu
Zhifeng Liu Guangli Ren
Guangli Ren
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell. Neurosci. , 08 November 2022
Sec. Cellular Neuropathology
Volume 16 - 2022 | https://doi.org/10.3389/fncel.2022.1003809
This article is part of the Research Topic ATF3: A Crucial Stress-Responsive Gene of Glia and Neurons in CNS View all 5 articles
Microglia are involved in immune responses to central nervous system (CNS) injury. Meanwhile, exosomes derived from microglia are important mediators of information and material exchange in brain, which play an important role in neuroprotective or damaging effects. Microglial exosomes contain a variety of molecular cargos, including microRNAs, soluble proteins, and lipids, which have regulatory effects on other types of cells and microenvironment in brain. In this review, we summarized microglial exosome characteristics, release patterns, pro-proliferative and pro-apoptotic effects on neurons and other glial cells, immunomodulatory effects, and regulation of the extracellular microenvironment. Understanding the relationship between microglia exosomes and brain injury can provide new targets for clinical treatment.
Microglia are derived from primitive myeloid progenitor cells in the yolk sac and are important resident innate immune cells in the central nervous system (CNS), which participated in neuroinflammation and neurodegeneration spreading (Priller and Prinz, 2019). Rahimian et al. (2018, 2019a,b,2021) have concluded that activated microglia not only secreted TNF-α, IL-1β, ROS, etc. to promote neuroinflammation in cerebral ischemia but also secreted IL-4, IL-10, and TGF-β to inhibit neuroinflammation and repair tissue. Galectin-3(Gal-3) played a key role in the activation and proliferation of microglia, which regulated angiogenesis, neurogenesis, and inflammation after stroke by activating several different signaling pathways, including IL-4, IGF-1, and Ga2+. In summary, they play a bidirectional role in neuroinflammation, as well as antigen presentation and phagocytosis along with the morphological changes in microglia. Meanwhile, they communicated with diverse cellular populations with different ways in brain injury, including cytokines, exosomes, and endocytosis. It is involved in the occurrence and development of nerve injury and repair and plays a crucial role in the survival of neurons, the establishment of neural circuits, and the maintenance of the homeostasis of the brain microenvironment.
Exosomes, a type of extracellular vesicles, are lipid bilayer-wrapped spherical structures with a diameter of 30–150 nm (Li et al., 2021). A variety of cells can secrete exosomes, including platelets, lymphocytes, adipocytes, muscle cells, tumor cells, glial cells, neurons, and stem cells (Crenshaw et al., 2019; Davis, 2016). They can also be detected in biological fluids such as semen, saliva, plasma, urine, milk, cerebrospinal fluid (CSF), amniotic fluid, and tumor effusion (Camussi et al., 2011; Davis, 2016; Crenshaw et al., 2019). Exosomal cargoes vary a lot under different stimuli (Li et al., 2021). Correspondingly, microglial exosomes can be released into microenvironment and be uptaken by neighboring cells that participate in the regulation of other cell functions and extracellular microenvironment (Osier et al., 2018; Wortzel et al., 2019). They not only promote neuronal apoptosis or proliferation but also have the functions of immunomodulation. Importantly, exosomes can cross the blood–brain barrier (BBB) and be detected in the CSF and serum, thus leading to the possibility that may be useful for the diagnosis and treatment of CNS injury.
In general, exosomes need to be classified on the limiting membrane of multivesicular endosomes (MVEs) receptors according to their contents and then target MVEs to the plasma membrane, inward budding and fission generate intraluminal vesicles (ILVs) and release into the extracellular environment to be internalized by target cells (van Niel et al., 2018). The number of exosomes increased significantly and the contents of exosomes varied under external stimuli (Figure 1). Increasing evidence shows that exosomes derived from microglia increased significantly in response to stimuli, including severe tensile injury (Zhao et al., 2021), lipopolysaccharide (LPS) (Zhao et al., 2021), ischemia and hypoxia (Xie et al., 2020), 5-HT (Glebov et al., 2015), Wnt3a (Hooper et al., 2012), Glutaminase 1 (GLS1) (Gao et al., 2020), Glutaminase C (GAC) (Gao et al., 2019), ATP (Turola et al., 2012), IFN-γ (Hou et al., 2020), and TNF-α (Van den Broek et al., 2020). Among them, Glebov et al. (2015) suggested that 5-HT receptors, including 5-HT2a, 5-HT2b, and 5-HT4Rs may activate two different pathways that converge on phospholipase C (PLC), namely the G-PLC-IP3/DAG pathway. 5-HT4R can target adenylyl cyclase (AC) directly or interact with PLC through cAMP-mediated activation of epac1/2/Rap1 in microglia. Furthermore, neurons and other glial cells in the brain can also release exosomes to activate microglia, causing them to secrete corresponding exosomes (Hou et al., 2020; Borst et al., 2021). However, TNF played a key role in the regulation of microglial exosome release. LPS stimulated microglia activation, which produced a 30-fold increase in extracellular vesicles (EVs), but a 16-fold decrease in the number of EVs released after inhibition of TNF signaling (Yang et al., 2018). Meanwhile, previous studies found that cAMP and Ca2+-dependent signaling pathways are involved in the regulation of exosome secretion. Therefore, blocking the release of microglial exosomes or clearing harmful exosomes through PLC pathway, cAMP, and Ca2+-dependent signaling pathways may effectively delay the occurrence and development of brain injury, which is beneficial to the recovery of the disease.
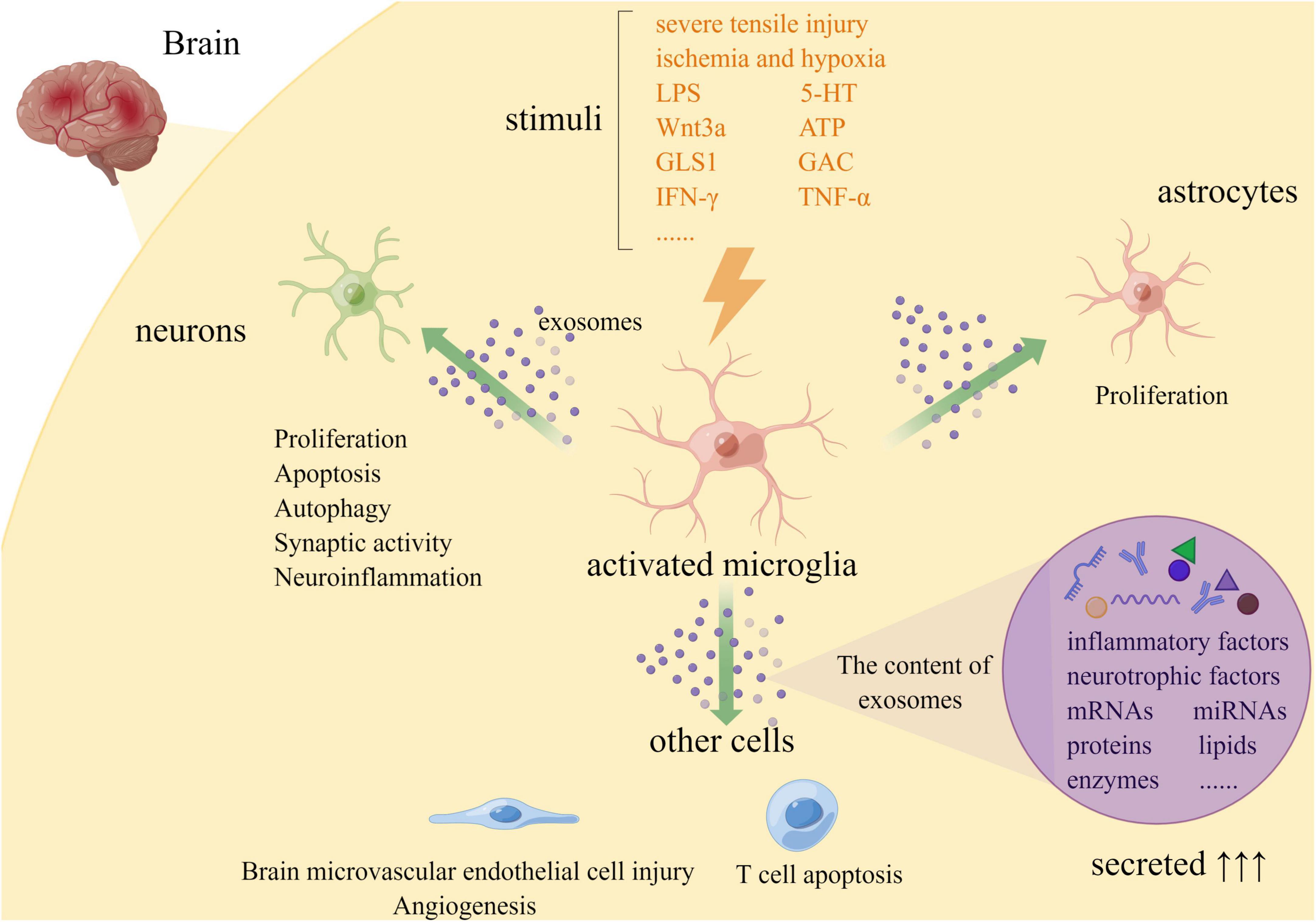
Figure 1. Release and role of microglial exosomes in brain injury (By Figdraw. ID:YAASUe13b4).
Microglia secrete exosomes containing various miRNAs and interact with neurons or other glial cells to promote their proliferation, degeneration, or apoptosis (Figure 1).
Neurons, the effector cells in the brain, can receive, process, and transmit information to target cells in the CNS through axons and synapses. It plays an important role in the formation of a neural network (Liu et al., 2017). Therefore, inhibiting the apoptosis of neurons will help reduce brain injury and promote disease recovery. Studies have shown that microglia are activated after cerebral ischemia, and upregulation of GLS1 led to an increase in pro-inflammatory exosomes, causing neuroinflammatory damage to neurons (Turola et al., 2012). Current studies have confirmed that former M2 microglia-derived exosomal miR-124 can reduce neuronal apoptosis and relieve brain injury after stroke by downregulating ubiquitin-specific protease 14 (USP14) (Song et al., 2019) and regulation of Bcl-2/Bcl-xl pathway (Yu et al., 2017). It has been reported that ischemic neurons can also take up miRNA-137-containing exosomes through the Notch1 pathway to maintain their own survival (Zhang et al., 2021). Wei et al. (2021) demonstrated that microglia activated by intracerebral hemorrhage (ICH) inhibited the expression of activating transcription factor 4 (ATF4) by secreting miR-383-3p-containing exosomes, thereby promoting neuronal necroptosis. On the other hand, neuron-derived exosomes can also promote microglial polarization to mediate neuroinflammation. Furthermore, Ge et al. (2020) demonstrated that microglial exosomes with upregulated miR-124-3p contribute to alleviating neurodegeneration and promoting neurite growth in repetitive mild traumatic brain injury (rmTBI) mice by targeting Rela/ApoE signaling pathway in hippocampal neurons.
Microglia can also affect neuronal function by regulating the proliferation of astrocytes. Ye et al. (2021) found that microglia can communicate with astrocytes through exosomes. Among them, the upregulation of microglial exosomal miRNA-145-5p can inhibit the proliferation of astrocytes in the early stage of injury by inhibiting the activity of smad3 and avoid excessive proliferation to form “glial scars,” which is conducive to the recovery of nerve function.
As a novel way, microglia communicate with other cells via exosomes in the CNS. Xie et al. demonstrated that microglial exosomal miR-424-5p upregulation under hypoxia could induce brain microvascular endothelial cell (BMEC) injury by targeting FGF2-mediated STAT3 pathway (Xie et al., 2020). Besides, some studies have indicated that under high pressure, the exosomes of microglia can transmit denaturation signals via autocrine, leading to oxidative stress and cell death (Aires et al., 2020). We also found that microglia also interacted with oligodendrocytes in diseases such as Alzheimer’s disease, multiple sclerosis, amyotrophic lateral sclerosis, schizophrenia, and depression. However, the effects of microglial exosomes on oligodendrocyte were still unclear and need more in-depth study (Rahimian et al., 2022). In conclusion, microglial exosomes with different contents provide a new view of the treatment for brain injury.
Existing studies show that microglia regulate immune response through phagocytosis and secretion of inflammatory mediators (Borst et al., 2021) or exosomes (Kalluri and LeBleu, 2020). Based on their roles in inflammation, microglia were divided into resting type, pro-inflammatory type (former M1 type), and anti-inflammatory type (former M2 type). They contacted target cells by secreting different types of inflammatory factors. However, whether these inflammatory factors were secreted directly by cells into the environment or capsuled in exosomes remains unclear. In ischemic stroke, microglia were activated to anti-inflammatory phenotype and subsequently transition to a pro-inflammatory phenotype at a later stage, achieving a balance of inflammatory responses (Jiang et al., 2020). We can distinguish former M1 and M2 subtypes by biomarkers. M1 markers include IL-1β, IL-6, IL-12 p70, TNF, IFNγ, CCL5, CCL20, CXCL1, CXCL10, GM-CSF, CD16, CD32, CD86, and iNOS. M2 markers include IL-1Ra, IL-4, IL-10, IL-4Rα, TGF-β, CCL22, CD206, CD163, Arg1, Ym1, FIZZ1, and G-CSF (Lan et al., 2017). However, with the in-depth study, more and more researches indicated that it is not sufficient to simply classify microglia into M1 and M2 phenotypes. One of the characteristic features of microglia is their transformation in response to CNS pathology. With the development of single-cell sequencing technology, the phenotype of microglia in brain injury was further revealed (Prinz et al., 2019). These studies have also furthered our understanding of the role of microglia in brain injury. Different phenotypes of microglia exosomes play different roles in brain injury due to their different contents, and we also summarized the roles of pro-inflammatory and anti-inflammatory phenotypes of microglia exosomes (Ransohoff, 2016).
Lipopolysaccharide (LPS) induces polarization of microglia into pro-inflammatory subtype through activation of TLR2/TLR4/NF-kB/inflammasome signaling cascade and release of HMGB1 (Cunha et al., 2016). Under pathological conditions, microglia transformed into pro-inflammatory phenotype through downregulation of miR-124 to promote neuroinflammation (Yang et al., 2019). These microglia aggravate brain injury by releasing pro-inflammatory and neurotoxic factors including IL-1β, TNF-α, and IFN-γ (Zhao et al., 2020). Cunha et al. (2016) found that the expression profile of inflammation-miRs in exosomes was consistent with the expression profile of inflammation-miRs found in cells. That is to say, pro-inflammatory subtype microglia can directly transport soluble inflammatory signals and miRNAs to adjacent cells through exosomes.
Huang et al. (2018) demonstrated that miR-124-3p promoted microglial transforming to anti-inflammatory subtype by targeting PDE4B to inhibit the activity of the mTOR signaling pathway and suppress neuronal inflammation in scratched neurons. Furthermore, Yu et al. (2017) found that miR-124 can also attenuate inflammatory responses via modulating transcription factor C/EBP-α. At the same time, Long et al. (2020) discovered activated astrocytes secrete exosomes rich in miR-873a-5p, which promoted transformation of anti-inflammatory microglia by inhibiting phosphorylation of ERK and NF-κB p65, thereby alleviating neurological deficits after traumatic brain injury (TBI).
Disease-associated microglia play a key role in neuroinflammation, while they can also influence multiple immune cell functions through exosomes (Prinz et al., 2019). Some studies have found that exosomal IL-10 and FASL signaling can induce T cell apoptosis, which is beneficial to the amplification of regulatory T cells and myeloid suppressor cells. Exosomes can increase the activity of natural killer (NK) cells and the survival rate of T cells, on the one hand, and inhibit the activity of NK cells, T cell proliferation, and dendritic cell (DC) differentiation, on the other hand, thus exerting a bidirectional immunomodulatory effect (El Andaloussi et al., 2013). It has been reported that microglia can also secrete NAMPT through exosomes during neuroinflammation of ischemic injury and participate in the inflammatory response (Yang et al., 2019). Several studies have shown that in brain injury such as TBI and transient middle cerebral artery occlusion (tMCAO), the treatment of microglial exosomes can reduce neuroinflammation and immune responses (Huang et al., 2018; Song et al., 2019). Taken together, microglial exosomes are involved in the immune regulation of brain injury.
Exosomes secreted by microglia can affect neurotransmitter transmission. It has been reported that microglial exosomes are involved in the homeostatic regulation of neurotransmission by enhancing spontaneous and evoked excitatory transmission through interacting with neuronal plasma membranes and neuronal sphingolipid metabolism (Turola et al., 2012). Some studies have found that exosomes secreted by microglia can inhibit presynaptic transmission by binding to receptors, which is due to the endocannabinoid N-arachidonoylethanolamine (AEA) carried on its surface. Exosomes carrying AEA can stimulate the type 1 cannabinoid (CB1) receptor expressed by GABAergic neurons, thereby inhibiting presynaptic transmission (Gabrielli et al., 2015).
Microglia can prune synapses and maintain synaptic homeostasis through phagocytosis of synapses, which play an important role in establishing neural circuits and maintaining the function of nerve cells. Exosomes derived from microglia can also transport enzymes and other substances, promote metabolism such as anaerobic glycolysis and lactate production, and supplement energy for synaptic activities (Pistono et al., 2020). In addition, studies have shown that injured microglia may affect synaptic activity by downregulating exosomal miR-5121 and transmitting signals to stretch injured neurons after TBI, inhibiting the growth of some neurites and synaptic recovery (Zhao et al., 2021).
Tian et al. (2019) found that IL-4-polarized BV2 cells may promote angiogenesis by secreting exosomes containing miR-26a. Subsequently, it was also confirmed that former M2 microglia can secrete exosomes to promote neovascularization, and HIF pathway may be involved. The delivery of oxygen, nutrients, and even immune cells to the injured site through neovascularization will facilitate the recovery of the disease. For example, the ischemic area promotes the recovery of ischemic stroke due to the entry of more Th2 or former M2 cells.
Microglia exosomes can also maintain cellular homeostasis and repair functions. They can activate autophagy through the formation of LC3B-positive autophagosomes in microglia (Van den Broek et al., 2020). For example, microglia-derived exosomes activated by exosomes from SH-SY5Y cells overexpressing α-synuclein can target PTEN through miR-19a-3p, thereby activation of the PI3K/Akt/mTOR signaling pathway ultimately inhibits neuronal autophagy. Other studies have shown that inhibition of autophagy can induce former M1 microglia polarization and the release of TNF-α, iNOS, and Cyclooxygenase 2 (Cox2), which can aggravate the injury of brain (Xia et al., 2016). Thus, microglial exosomes maintain cellular homeostasis by activating autophagy.
We have summarized the most recent evidence regarding microglia-derived exosomes playing an important role in brain injury due to changes in the type of cargoes. Microglial exosomes, as important information transfer mediators, can communicate with a variety of cells in the brain, including neurons. Meanwhile, they can affect the microenvironment, then affecting neurological function and blood–brain barrier integrity. Some studies already suggest that microglia can influence CNS function through miRNAs, neurotransmitters, and proteins in exosomes, and many types of bioactive substances in exosomes can be used as disease biomarkers of brain injury.
The authors of this review consider that analysis of microglia exosome release, delivery, and contents will provide a more comprehensive and accurate assessment of brain injury development. The information about blocking, promoting, and modifying exosomal contents will provide a safer and robust line of treatment.
YC, JZ, and JJ: drafting and refining the manuscript. ZL and GR: critical reading of the manuscript. All authors read and approved the manuscript.
This work was supported by the grants from the PLA Logistics Research Project of China (CLB20J032 and 2022-JCJQ-ZD-097-12) and Natural Science Foundation of Guangdong Province (2021A1515010170).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Aires, I. D., Ribeiro-Rodrigues, T., Boia, R., Catarino, S., Girao, H., Ambrosio, A. F., et al. (2020). Exosomes derived from microglia exposed to elevated pressure amplify the neuroinflammatory response in retinal cells. Glia 68, 2705–2724. doi: 10.1002/glia.23880
Borst, K., Dumas, A. A., and Prinz, M. (2021). Microglia: immune and non-immune functions. Immunity 54, 2194–2208. doi: 10.1016/j.immuni.2021.09.014
Camussi, G., Deregibus, M. C., Bruno, S., Grange, C., Fonsato, V., and Tetta, C. (2011). Exosome/microvesicle-mediated epigenetic reprogramming of cells. Am. J. Cancer Res. 1, 98–110.
Crenshaw, B. J., Kumar, S., Bell, C. R., Jones, L. B., Williams, S. D., Saldanha, S. N., et al. (2019). Alcohol modulates the biogenesis and composition of microglia-derived exosomes. Biology 8:25. doi: 10.3390/biology8020025
Cunha, C., Gomes, C., Vaz, A. R., and Brites, D. (2016). Exploring new inflammatory biomarkers and pathways during LPS-induced M1 polarization. Mediat. Inflamm. 2016:6986175. doi: 10.1155/2016/6986175
Davis, M. E. (2016). Exosomes: what do we love so much about them? Circ. Res. 119, 1280–1282. doi: 10.1161/CIRCRESAHA.116.309942
El Andaloussi, S., Mager, I., Breakefield, X. O., and Wood, M. J. (2013). Extracellular vesicles: biology and emerging therapeutic opportunities. Nat. Rev. Drug Discov. 12, 347–357. doi: 10.1038/nrd3978
Gabrielli, M., Battista, N., Riganti, L., Prada, I., Antonucci, F., Cantone, L., et al. (2015). Active endocannabinoids are secreted on extracellular membrane vesicles. EMBO Rep. 16, 213–220. doi: 10.15252/embr.201439668
Gao, G., Li, C., Zhu, J., Wang, Y., Huang, Y., Zhao, S., et al. (2020). Glutaminase 1 regulates neuroinflammation after cerebral ischemia through enhancing microglial activation and pro-inflammatory exosome release. Front. Immunol. 11:161. doi: 10.3389/fimmu.2020.00161
Gao, G., Zhao, S., Xia, X., Li, C., Li, C., Ji, C., et al. (2019). Glutaminase C regulates microglial activation and pro-inflammatory exosome release: relevance to the pathogenesis of Alzheimer’s disease. Front. Cell. Neurosci. 13:264. doi: 10.3389/fncel.2019.00264
Ge, X., Guo, M., Hu, T., Li, W., Huang, S., Yin, Z., et al. (2020). Increased microglial exosomal miR-124-3p alleviates neurodegeneration and improves cognitive outcome after rmTBI. Mol. Ther. 28, 503–522. doi: 10.1016/j.ymthe.2019.11.017
Glebov, K., Lochner, M., Jabs, R., Lau, T., Merkel, O., Schloss, P., et al. (2015). Serotonin stimulates secretion of exosomes from microglia cells. Glia 63, 626–634. doi: 10.1002/glia.22772
Hooper, C., Sainz-Fuertes, R., Lynham, S., Hye, A., Killick, R., Warley, A., et al. (2012). Wnt3a induces exosome secretion from primary cultured rat microglia. BMC Neurosci. 13:144. doi: 10.1186/1471-2202-13-144
Hou, B. R., Jiang, C., Wang, Z. N., and Ren, H. J. (2020). Exosome-mediated crosstalk between microglia and neural stem cells in the repair of brain injury. Neural Regen. Res. 15, 1023–1024. doi: 10.4103/1673-5374.270302
Huang, S., Ge, X., Yu, J., Han, Z., Yin, Z., Li, Y., et al. (2018). Increased miR-124-3p in microglial exosomes following traumatic brain injury inhibits neuronal inflammation and contributes to neurite outgrowth via their transfer into neurons. FASEB J. 32, 512–528. doi: 10.1096/fj.201700673r
Jiang, C. T., Wu, W. F., Deng, Y. H., and Ge, J. W. (2020). Modulators of microglia activation and polarization in ischemic stroke (Review). Mol. Med. Rep. 21, 2006–2018. doi: 10.3892/mmr.2020.11003
Kalluri, R., and LeBleu, V. S. (2020). The biology, function, and biomedical applications of exosomes. Science 367:eaau6977. doi: 10.1126/science.aau6977
Lan, X., Han, X., Li, Q., Yang, Q. W., and Wang, J. (2017). Modulators of microglial activation and polarization after intracerebral haemorrhage. Nat. Rev. Neurol. 13, 420–433. doi: 10.1038/nrneurol.2017.69
Li, Y., Liu, Z., Shi, X., Tong, H., and Su, L. (2021). Prognostic value of plasma exosomal levels of histone H3 protein in patients with heat stroke. Exp. Ther. Med. 22:922. doi: 10.3892/etm.2021.10354
Liu, F., Patterson, T. A., Zhang, J., Paule, M. G., Slikker, W. Jr., and Wang, C. (2017). Neural Cell Biology. Boca Raton, FL: CRC Press, 1–13. doi: 10.1201/9781315370491-1
Long, X., Yao, X., Jiang, Q., Yang, Y., He, X., Tian, W., et al. (2020). Astrocyte-derived exosomes enriched with miR-873a-5p inhibit neuroinflammation via microglia phenotype modulation after traumatic brain injury. J. Neuroinflamm. 17:89. doi: 10.1186/s12974-020-01761-0
Osier, N., Motamedi, V., Edwards, K., Puccio, A., Diaz-Arrastia, R., Kenney, K., et al. (2018). Exosomes in acquired neurological disorders: new insights into pathophysiology and treatment. Mol. Neurobiol. 55, 9280–9293. doi: 10.1007/s12035-018-1054-4
Pistono, C., Bister, N., Stanova, I., and Malm, T. (2020). Glia-derived extracellular vesicles: role in central nervous system communication in health and disease. Front. Cell Dev. Biol. 8:623771. doi: 10.3389/fcell.2020.623771
Priller, J., and Prinz, M. (2019). Targeting microglia in brain disorders. Science 365, 32–33. doi: 10.1126/science.aau9100
Prinz, M., Jung, S., and Priller, J. (2019). Microglia biology: one century of evolving concepts. Cell 179, 292–311. doi: 10.1016/j.cell.2019.08.053
Rahimian, R., Beland, L. C., and Kriz, J. (2018). Galectin-3: mediator of microglia responses in injured brain. Drug Discov. Today 23, 375–381. doi: 10.1016/j.drudis.2017.11.004
Rahimian, R., Beland, L. C., Sato, S., and Kriz, J. (2021). Microglia-derived galectin-3 in neuroinflammation; a bittersweet ligand? Med. Res. Rev. 41, 2582–2589. doi: 10.1002/med.21784
Rahimian, R., Cordeau, P. Jr., and Kriz, J. (2019a). Brain response to injuries: when microglia go sexist. Neuroscience 405, 14–23. doi: 10.1016/j.neuroscience.2018.02.048
Rahimian, R., Lively, S., Abdelhamid, E., Lalancette-Hebert, M., Schlichter, L., Sato, S., et al. (2019b). Delayed galectin-3-mediated reprogramming of microglia after stroke is protective. Mol. Neurobiol. 56, 6371–6385. doi: 10.1007/s12035-019-1527-0
Rahimian, R., Perlman, K., Canonne, C., and Mechawar, N. (2022). Targeting microglia-oligodendrocyte crosstalk in neurodegenerative and psychiatric disorders. Drug Discov. Today 27, 2562–2573. doi: 10.1016/j.drudis.2022.06.015
Ransohoff, R. M. (2016). A polarizing question: do M1 and M2 microglia exist? Nat. Neurosci. 19, 987–991. doi: 10.1038/nn.4338
Song, Y., Li, Z., He, T., Qu, M., Jiang, L., Li, W., et al. (2019). M2 microglia-derived exosomes protect the mouse brain from ischemia-reperfusion injury via exosomal miR-124. Theranostics 9, 2910–2923. doi: 10.7150/thno.30879
Tian, Y., Zhu, P., Liu, S., Jin, Z., Li, D., Zhao, H., et al. (2019). IL-4-polarized BV2 microglia cells promote angiogenesis by secreting exosomes. Adv. Clin. Exp. Med. 28, 421–430. doi: 10.17219/acem/91826
Turola, E., Furlan, R., Bianco, F., Matteoli, M., and Verderio, C. (2012). Microglial microvesicle secretion and intercellular signaling. Front. Physiol. 3:149. doi: 10.3389/fphys.2012.00149
Van den Broek, B., Pintelon, I., Hamad, I., Kessels, S., Haidar, M., Hellings, N., et al. (2020). Microglial derived extracellular vesicles activate autophagy and mediate multi-target signaling to maintain cellular homeostasis. J. Extracell. Vesicles 10:e12022. doi: 10.1002/jev2.12022
van Niel, G., D’Angelo, G., and Raposo, G. (2018). Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 19, 213–228. doi: 10.1038/nrm.2017.125
Wei, M., Li, C., Yan, Z., Hu, Z., Dong, L., Zhang, J., et al. (2021). Activated microglia exosomes mediated miR-383-3p promotes neuronal necroptosis through inhibiting ATF4 expression in intracerebral hemorrhage. Neurochem. Res. 46, 1337–1349. doi: 10.1007/s11064-021-03268-3
Wortzel, I., Dror, S., Kenific, C. M., and Lyden, D. (2019). Exosome-mediated metastasis: communication from a distance. Dev. Cell. 49, 347–360. doi: 10.1016/j.devcel.2019.04.011
Xia, C. Y., Zhang, S., Chu, S. F., Wang, Z. Z., Song, X. Y., Zuo, W., et al. (2016). Autophagic flux regulates microglial phenotype according to the time of oxygen-glucose deprivation/reperfusion. Int. Immunopharmacol. 39, 140–148. doi: 10.1016/j.intimp.2016.06.030
Xie, L., Zhao, H., Wang, Y., and Chen, Z. (2020). Exosomal shuttled miR-424-5p from ischemic preconditioned microglia mediates cerebral endothelial cell injury through negatively regulation of FGF2/STAT3 pathway. Exp. Neurol. 333:113411. doi: 10.1016/j.expneurol.2020.113411
Yang, Y., Boza-Serrano, A., Dunning, C. J. R., Clausen, B. H., Lambertsen, K. L., and Deierborg, T. (2018). Inflammation leads to distinct populations of extracellular vesicles from microglia. J. Neuroinflamm. 15:168. doi: 10.1186/s12974-018-1204-7
Yang, Y., Ye, Y., Kong, C., Su, X., Zhang, X., Bai, W., et al. (2019). MiR-124 enriched exosomes promoted the M2 polarization of microglia and enhanced hippocampus neurogenesis after traumatic brain injury by inhibiting TLR4 pathway. Neurochem. Res. 44, 811–828. doi: 10.1007/s11064-018-02714-z
Ye, Y., Hao, J., Hong, Z., Wu, T., Ge, X., Qian, B., et al. (2021). Downregulation of MicroRNA-145-5p in activated microglial exosomes promotes astrocyte proliferation by removal of Smad3 inhibition. Neurochem. Res. 47, 382–393. doi: 10.1007/s11064-021-03446-3
Yu, A., Zhang, T., Duan, H., Pan, Y., Zhang, X., Yang, G., et al. (2017). MiR-124 contributes to M2 polarization of microglia and confers brain inflammatory protection via the C/EBP-alpha pathway in intracerebral hemorrhage. Immunol. Lett. 182, 1–11. doi: 10.1016/j.imlet.2016.12.003
Zhang, D., Cai, G., Liu, K., Zhuang, Z., Jia, K., Pei, S., et al. (2021). Microglia exosomal miRNA-137 attenuates ischemic brain injury through targeting Notch1. Aging 13, 4079–4095. doi: 10.18632/aging.202373
Zhao, C., Deng, Y., He, Y., Huang, X., Wang, C., and Li, W. (2021). Decreased level of exosomal miR-5121 released from microglia suppresses neurite outgrowth and synapse recovery of neurons following traumatic brain injury. Neurotherapeutics 18, 1273–1294. doi: 10.1007/s13311-020-00999-z
Keywords: microglia, exosomes, brain injury, regulation, proliferation, apoptosis
Citation: Chen Y, Zhu J, Ji J, Liu Z and Ren G (2022) The role of microglial exosomes in brain injury. Front. Cell. Neurosci. 16:1003809. doi: 10.3389/fncel.2022.1003809
Received: 27 July 2022; Accepted: 17 October 2022;
Published: 08 November 2022.
Edited by:
Teresa Summavielle, Universidade do Porto, PortugalCopyright © 2022 Chen, Zhu, Ji, Liu and Ren. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guangli Ren, Z3VhbmdsaXJlbkBob3RtYWlsLmNvbQ==; Zhifeng Liu, emhpZmVuZ2xpdTc3OTdAMTYzLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.