 Mark Stephen Hamer
Mark Stephen Hamer Fabio M. V. Rossi
Fabio M. V. Rossi
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
PERSPECTIVE article
Front. Cell Dev. Biol., 22 January 2025
Sec. Stem Cell Research
Volume 12 - 2024 | https://doi.org/10.3389/fcell.2024.1526635
This article is part of the Research TopicInduced Pluripotent Stem Cells (iPSCs) for Skeletal Muscle DiseasesView all 6 articles
The generation of myogenic progenitors from iPSCs (iMPs) with therapeutic potential for in vivo tissue regeneration has long been a goal in the skeletal muscle community. Today, protocols enable the production of potent, albeit immature, iMPs that resemble Pax7+ adult muscle stem cells. While muscular dystrophies are often the primary therapeutic target for these cells, an underexplored application is their use in treating traumatic muscle injuries. Notably absent from recent reviews on iMPs is the concept of engineering these cells to perform functions post-transplantation that non-transgenic cells cannot. Here, we highlight protocols to enhance the generation, purification, and maturation of iMPs, and introduce the idea of engineering these cells to perform functions beyond their normal capacities, envisioning novel therapeutic applications.
Life-long maintenance of skeletal muscle integrity is maintained by a population of tissue resident adult muscle stem cells (MuSCs), or “satellite cells”, embedded between the extracellular matrix (ECM) and individual myofibers (Mauro, 1961). The indispensable role of MuSCs, distinguished by their tissue-specific expression of the transcription factor Pax7, has been repeatedly demonstrated (Lepper et al., 2011; Murphy et al., 2011; Relaix and Zammit, 2012; Von Maltzahn et al., 2013). Despite protocols that afford efficient isolation of primary MuSCs from both human and murine tissues, therapeutic application of these cells is severely limited by donor availability and an inability to expand these cells in vitro (Montarras et al., 2005; Yi and Rossi, 2011; Liu et al., 2015; Jensen et al., 2021). Pluripotent stem cells (PSCs) have emerged as a promising, scalable, source of myogenic progenitors (MPs). Studies into the developmental origins of MuSCs have led to a deep understanding of the biological processes underlying the emergence of these cells within skeletal muscle tissue (Chal and Pourquié, 2017). These efforts, along with those affording the generation of PSCs via reprogramming of adult somatic tissue (iPSCs), have been foundational to the concept of using autologous MPs therapeutically (Takahashi and Yamanaka, 2006; Barberi et al., 2007).
In this perspective, we present the state-of-the-art in generating and purifying therapeutically relevant iPSC-derived MPs (iMPs), focusing on workflows that enhance their similarity to MuSCs and improve engraftment for de novo muscle formation and MuSC niche repopulation (Table 1). We then explore an emerging area of cellular engineering which, despite rapid adoption in fields like immunology, remains relatively undeveloped in skeletal muscle therapeutics. Synthetic biology and the engineering of cells to perform functions beyond their natural roles presents a transformative opportunity to reshape the current paradigm of iMP-based therapies.
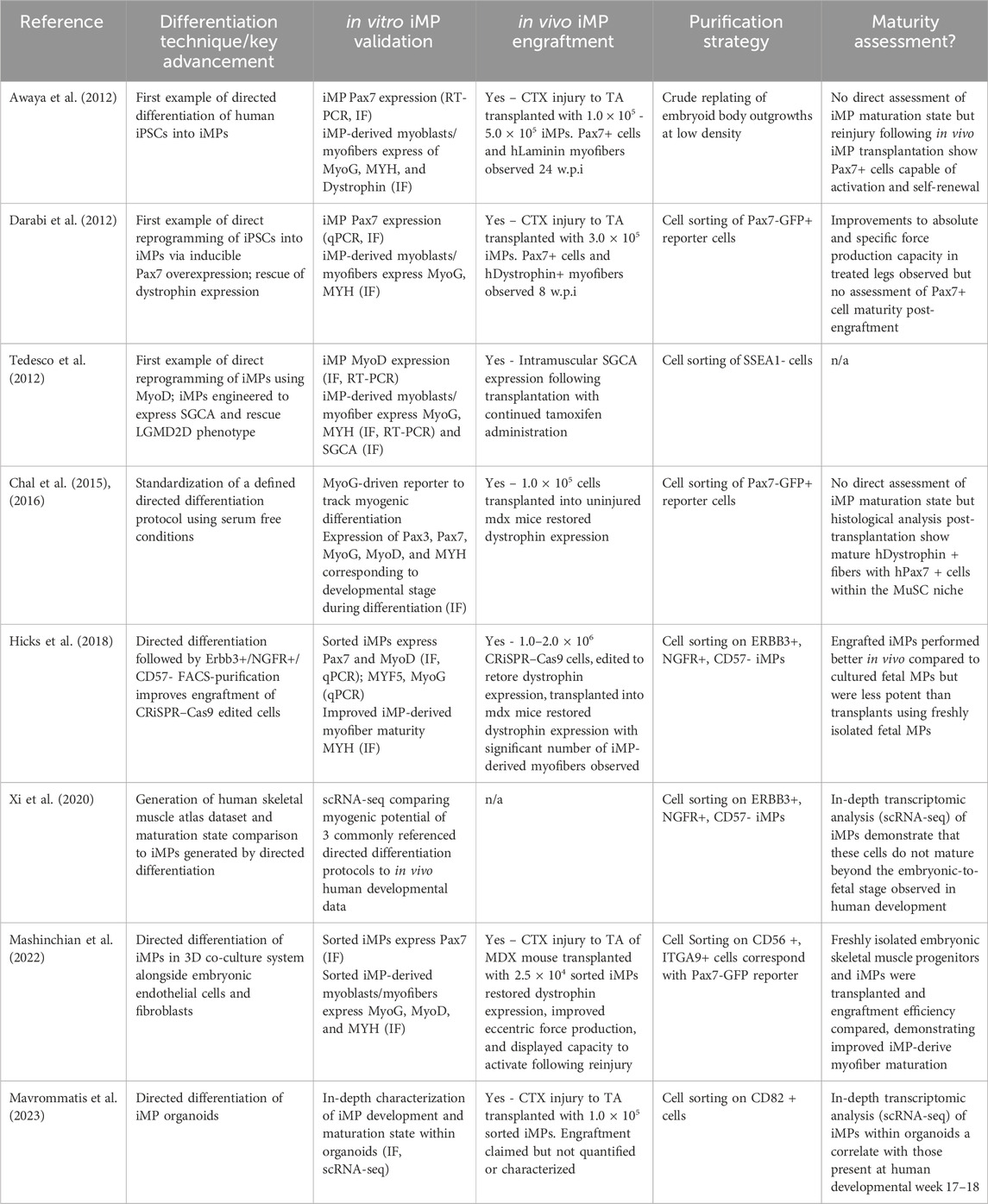
Table 1. Key findings published that have progressed the field of iMP generation to its current state. The stability and flexibility afforded by directed differentiation are highlighted along with the maturation characteristics that we believe to be critical to improving iMP therapeutic application.
Today, a number of well-established protocols for the production of iMPs exist, each falling into one of two categories (Iberite et al., 2022). Direct reprogramming relies on the overexpression of myogenic transgenes to force myogenic potential, while directed differentiation generates iMPs through meticulous recapitulation of the conditions that lead to developmental emergence of MuSCs in vivo (Hicks et al., 2018; Xi et al., 2020; Nalbandian et al., 2021). While these two methods for generating iMPs differ, both aim to produce cells that can be used directly or, preferably, cryopreserved in viable cell banks for downstream application (Figure 1).
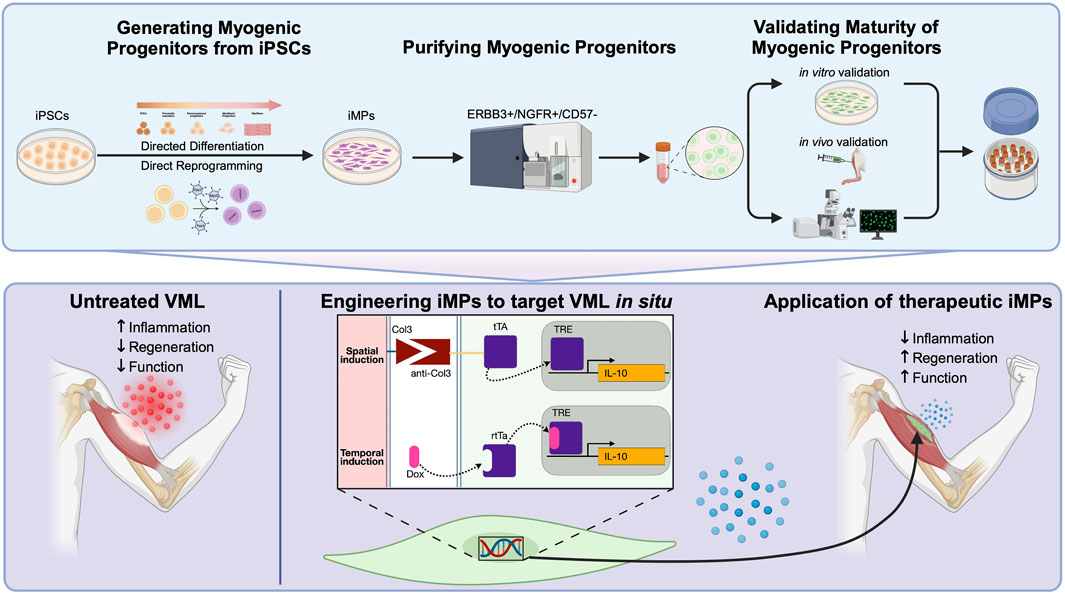
Figure 1. (Top panel) Standardized workflow for the generation and validation of iPSC-derived myogenic progenitors (iMPs) utilizing either directed differentiation or direct reprogramming approaches. Following differentiation, FACS isolation using ERBB3+/NGFR+/CD57− result in iMPs that are more mature, potent, and phenotypically comparable to MuSCs. Verification of myogenicity immediately after cell sorting include immunofluorescent staining for markers like Pax7 and MyoG and MYH and Dystrophin after secondary differentiation into myotubes. In parallel, cells should be further validated for their capacity to engraft and form de novo myofibers in vivo. Further histological assessment should confirm the capacity for some iMPs to retain Pax7 expression and populate the MuSC niche. (Bottom panel) An example of iMPs engineering to target volumetric muscle loss (VML) pathophysiology. Untreated VML is underscored by dysregulated fibrosis in the damaged muscle leading to poor regenerative potential. Engineering iMPs that express IL-10, a cytokine demonstrated to have potent pro-regenerative effects in VML, could be further enhanced by the spatio-temporal delivery afforded by this system. The improved regenerative environment should act synergistically to alleviate fibrosis and improve myogenic potential in the transplanted cells.
Overexpression of the myogenic transcription factors MyoD and Pax7 both reliably induce iPSCs toward a myogenic fate. The first examples of iMP direct reprogramming demonstrated that activation of these inducible transgenes generated cells that reliably engraft into muscle tissue, and are capable of restoring tissue specific expression of dystrophin and a-sarcoglycan in their respective murine disease models (Darabi et al., 2012; Tedesco et al., 2012). Since these initial demonstrations, direct reprogramming protocols have been used to reliably produce iMPs with similar regenerative capacity (Tanaka et al., 2013; Quattrocelli et al., 2015; Uezumi et al., 2016; Azzag et al., 2022). Despite these successes, a number of caveats specific to direct reprogramming warrant consideration. First, many direct reprogramming strategies adopt the use of small molecules and growth factors to prime iPSCs towards a mesodermal lineage prior to myogenic transgene induction, raising the question of whether introducing transgenes for the final step of myogenic specification is truly necessary (Magli et al., 2017; Rao et al., 2018; Selvaraj et al., 2019). Correspondingly, direct reprogramming inherently skips critical developmental maturation steps and little has been done to assess the relative developmental maturity of iMPs generated using this method. We will elaborate on the importance of iMP maturity in later sections; however, the lack of evidence for maturation in these cells represents one of the most critical shortcomings of direct reprogramming. Concerns over the risks associated with random transgene integration in direct reprogramming have also drawn scrutiny. Targeting transgenes to genetic safe-harbor loci and transient overexpression systems have been developed to reduce the perceived risks associated with transgene integration but are likely precautionary and immaterial to therapeutic application (Sadelain et al., 2012; Ordovás et al., 2015; Kim et al., 2017; Kim et al., 2021; Shin et al., 2020).
Despite the drawbacks associated with direct reprogramming the FDA has recently given clinical trial approval for a first-of-its-kind muscular dystrophy treatment in which allogeneic cell banks, induced to a myogenic lineage using Pax7 overexpression, will be assessed for safety, tolerability, and engraftment capacity in DMD patients (Myogenica, 2024). Successes in these early trials have the potential to transform the landscape of myopathy-targeting therapeutics and open the door to more advanced therapeutic modalities, as we discuss below.
Unlike direct reprogramming, directed differentiations use transgene-free methods of generating iMPs, relying on a holistic consideration of the developmental conditions that give rise to adult MuSCs. The first directed differentiation protocols were adapted from human embryonic stem cell (hESC) systems, and relied on undefined medias to differentiate embryoid body outgrowths into iMPs (Barberi et al., 2007; Chang et al., 2009; Mizuno et al., 2010; Awaya et al., 2012). While heterogeneity in the resulting populations was duly noted, these early studies provided evidence of myogenic potential in human iPSCs, paving way for more sophisticated protocols.
In 2016, two seminal papers established the culture conditions for generating Pax7+ myogenic progenitors from human iPSCs, optimizing an approach that induces presomitic mesoderm (PSM) specification prior to myogenic progenitor differentiation (Chal et al., 2015; Chal et al., 2016; Shelton et al., 2016). The resulting iMPs demonstrated robust myogenic capacity, but the rationale for the timing and concentration of certain growth factors remains dubious, even today. As these remain two of the most cited protocols for the generation of iMPs, a more granular screening aimed at optimizing these conditions is warranted and overdue. Data on the transition from early fetal to adult skeletal muscle in human tissue isolates are publicly available and may provide insights that would resolve this ambiguity (Xi et al., 2020).
Today, generation of iMPs using PSM to myogenic precursor differentiation has become the gold-standard in directed differentiations. However, the resulting cells are heterogeneous and there remains potential for further optimization and refinement (Bou Akar et al., 2024). Directed differentiations can generate iMPs that stably express Pax7 and are a more developmentally accurate recapitulation of MuSC-like cells than those generated via direct reprogramming. Standardization of directed differentiation protocols will be crucial to delivering therapeutic-grade iMPs. Towards this end STEMCELL technologies has released a product (STEMdiff™ Myogenic Progenitor Supplement Kit), that reliably produces a robust population of iMPs. cGMP qualification of products like this would represent a major step towards therapeutic application.
Regardless of the method used to generate iMPs, their therapeutic potential remains constrained by population heterogeneity. Direct reprogramming protocols utilize Pax7-driven fluorescent reporters to facilitate iMP purification, but neural crest progenitors, which also express Pax7, commonly contaminate these cultures (Basch et al., 2006). Non-transgenic directed differentiation strategies lack fluorescent reporters, necessitating the identification of surface markers that correlate with myogenic Pax7 expression. Cell surface markers for the purification of adult MuSCs have been identified, but are not reliably expressed in human iMPs (Wosczyna and Rando, 2018; Mierzejewski et al., 2020). Characterization of Pax7+ iMPs has led to the identification of several combinations of cell surface markers that aid in their purification. While CD54, CD10, CD82, CD56, and CDH13, and FGFR-4 all correlate with Pax7+ expression in human iMPs, the epidermal growth factor receptor ERBB3 has thus far been demonstrated the most reliable (Choi et al., 2016; Uezumi et al., 2016; Hicks et al., 2018; Sakai-Takemura et al., 2018; Wu et al., 2018; Nalbandian et al., 2021). In combination with positive selection for CD271 and negative selection of CD57 neuroectodermal contaminates, ERBB3 allows robust purification of iMPs that are phenotypically similar to the MuSCs observed developmentally during secondary myogenesis (Hicks et al., 2018; Tey et al., 2019). To validate myogenicity after cell sorting, iMPs should be plated and immediately stained for Pax7 and MyoG. Additional confirmation of myogenic potential should be confirmed by staining for myosin heavy chain (MYH) and dystrophin following subsequent differentiation into myotubes.
Despite improvements in the generation and purification of iMPs over the last decade, these cells invariably retain a late-embryonic/early-fetal phenotype, which like fetal MuSCs, show reduced engraftment capacity compared to adult MuSCs (Castiglioni et al., 2014; Xi et al., 2020). Strategies to address this lack of maturity include culturing iMPs in a 3D macroenvironment, external stimulation, and in vivo maturation (Khodabukus et al., 2019; Selvaraj et al., 2019; Kim et al., 2020; Sun et al., 2022; van der Wal et al., 2023; Crist et al., 2024). As maturation state remains the most significant identifiable cause of poor iMP engraftment, a collective effort must be made to address this problem.
3D directed differentiation of iMPs attempts to integrate the culture conditions established in 2D protocols with the spatial/environmental cues afforded by 3D organization. In one study, fully humanized multilineage embryoids containing iPSCs, growth-arrested embryonic fibroblasts and embryonic endothelial cells were grown together using a directed differentiation protocol, resulting in robust myogenic induction of the iPSCs; after only 13 days 40%–50% of cells in these embryoids were Pax7+ (Mashinchian et al., 2022). Unfortunately, the method used in this study to quantify Pax7 via RNA-FISH misrepresents the true myogenic Pax7 population, and staining with surface markers for Pax7-associated proteins significantly reduced the number of truly myogenic cells. In addition, despite acceptable engraftment and improvements to functional force production, these cells exhibited an embryonic (week 9) phenotype, indicating that they are no more mature than those produced in 2D systems.
In a more recent publication, human skeletal muscle organoids formed a largely myogenic population of cells (∼90%). Interestingly, a significant number of mesenchymal (∼4%) and neural (∼5%) cells also populated these 3D structures. While not explicitly studied, the heterogenous 3D structure was likely partially responsible for the maturation of the resulting Pax7+ iMPs, which closely resembled a fetal week 17 phenotype (Mavrommatis et al., 2023). Cell-cell communication analysis of the scRNA sequencing data generated in this work identified a number of upregulated ECM components (COL1A2, COL5A2, COL5A3, FN1) and transcription factors (FBN1, CHODL, SPRY1) that could be targeted in 2D or 3D systems to further mature iMPs.
While inducing differentiation of iMPs using 3D culture systems is relatively new, the application of differentiated iMPs within 3D systems has been widely reported (Rao et al., 2018; Selvaraj et al., 2019; van der Wal et al., 2023). It is established that 3D co-cultures can mimic the MuSC niche, enhance myofiber maturation, boost in vivo engraftment potential, and act as effective disease models (Quarta et al., 2016; Quarta et al., 20017; Bersini et al., 2018; Maffioletti et al., 2018; Rao et al., 2018; Bakooshli et al., 2019; Kim et al., 2020; Urciuolo et al., 2020; Massih et al., 2023). While recovery of mature iMPs from 3D systems may limit certain therapeutic applications (e.g., those requiring single cell suspensions), it may enhance their utility in others (e.g., traumatic skeletal muscle injury). 3D systems should be considered valuable tools for assessing iMP maturation and could help to further identify culture conditions that enhance this process.
Stimulation of myogenic cells can be achieved through either mechanical, chemical, electrical, or static tension. Early studies of C2C12-derived myofibers seeded within 3D collagen scaffolds demonstrated that electrical stimulation of these cells improved sarcomere organization and myofiber maturity (Park et al., 2008). In 2018, Rao et al. demonstrated that myofibers grown within their 3D scaffolds supported a population of self-sustaining Pax7+ iMPs (Rao et al., 2018). Despite thorough characterization of this stimulatory effect on myofiber maturity and force production, the Pax7+ iMP population was not further characterized. In a more recent study, van der Wal et al. directly compared directly differentiated iMP-derived myofibers to those of primary human myoblasts, showing that electrical stimulation of the 3D scaffolds produced comparable force production measurements (van der Wal et al., 2023). While this study provided a detailed physical and proteomic analysis of the resulting myofibers, the presence and maturity of Pax7+ cells within the constructs were not reported. In vivo, maturation of the MuSC compartment coincides with myofiber maturation following secondary myogenesis. Future studies should make use of systems like those described above to assess the effects of stimulation on the capacity of iMP-derived myofibers to support maturation of the associated iMPs.
Human MuSC development is a spatiotemporally coordinated process guided by concurrent maturation of the surrounding tissue (Chal and Pourquié, 2017). Differentiation of iPSCs to fully mature iMPs may therefore be limited by constraints inherent to culture systems. Comparing MuSCs with iMPs matured in vivo or in vitro has demonstrated that in vivo-matured iMPs more closely resembled adult MuSCs, whereas in vitro-matured iMPs retained a more fetal-like transcriptomic profile (Incitti et al., 2019). Important myogenic regulators (Stat3, Jun, Itga7, Tgfb2, Notch1, Notch3, and Jag1) and genes involved in ECM regulation were upregulated in the in vivo matured iMPs compared to those matured in vitro. Future studies aimed at targeted activation or inhibition of signaling pathways in which these factors are involved may improve maturation of iMPs in vitro. Likewise, incorporating the identified ECM modalities within engineered scaffolds may expedite the maturation of these cells when applied within certain therapeutic contexts. The key takeaway from these data is that iMP maturity in vitro may be less significant to successful therapeutic application than previously thought; instead, we may be able to rely on in vivo maturation to carry these cells towards a more MuSC-like phenotype.
Advancements in the generation, purification, and maturation of iMPs justify consideration of how to proceed with their therapeutic application. Recent reviews have highlighted the need for pre-clinical testing of the safety and efficacy of iMPs but emphasis is often placed on those capable of addressing muscular dystrophies (Iberite et al., 2022; Sun et al., 2024). In this section we will instead discuss tissue engineering strategies focused on the treatment of volumetric muscle loss (VML). We will then consider how we might employ iMPs as a therapeutic platform, focusing potential application of these cells as treatments for systemic diseases.
VML is characterized by a loss of tissue architecture that leads to chronic fibrosis and an associated loss of function (Corona et al., 2015). Current clinical treatments rely on crude muscle flap surgeries, which are limited by donor-site availability (Greising et al., 2017). Experimental therapeutics targeting VML aim to restore tissue integrity and function through the use of 3D scaffolds seeded with myoblasts or MuSCs (Corona et al., 2013; Quarta et al., 2017; Goldman et al., 2018). Despite the obvious application of iMPs as a substitute for patient-derived MuSCs, few studies have reported successful therapeutic application of these cells (Wu et al., 2021; Pinton et al., 2023). iMP maturity likely dictates survival within the scaffold and the capacity to successfully generate functional de novo muscle.
Scaffold design is critical to targeting VML pathophysiology (Wolf et al., 2015). In previous sections we have alluded to different ECM components that might help to mature iMPs in vitro. Current scaffolds can include either synthetic or natural material compositions, with natural hydrogels generally favored due to their intrinsic biocompatibility (Fischer et al., 2020; Luo et al., 2022). Hydrogels are frequently generated using proteins like collagen or fibrinogen, or polysaccharides such as hyaluronic acid or alginate. Those composed of proteins - specifically collagen - rarely consider particular isoforms, which have diverse functional characteristics (Goldman et al., 2018; Matthias et al., 2018; Pollot et al., 2018). Future studies should aim to incorporate specific collagen isotypes, like those already identified as important for iMP maturation, into scaffolds seeded with iMPs to improve maturation and survivability (Incitti et al., 2019). Additional considerations, such as the addition of growth factors and materials that promote revascularization and reinnervation must also be considered but are outside the scope of this perspective (Wu et al., 2021).
Skeletal muscle is a highly vascularized, metabolically active tissue, making it an excellent vehicle for the delivery of biological therapeutics. The engraftment of iMPs, while imperfect, is likely sufficient to deliver effective cell-based synthetic gene products in situ. A key advantage of iMPs is their capacity to be extensively engineered prior to differentiation and/or transplantation, affording flexibility in tailoring treatments. Utilizing iMPs as a delivery platform for therapeutics like growth factors, cytokines, and biologics represents a highly promising, yet largely untapped, application. This approach could revolutionize the delivery of therapeutics by directly targeting affected tissues, enhancing treatment precision and efficacy.
Following VML, a dysregulated immune response results in persistent inflammation, elevated TGFβ expression within the damaged tissue, and resulting in fibrosis (Larouche et al., 2021). Experimental treatments have successfully alleviated fibrosis through systemic administration of anti-fibrotic agents that target TGFβ, but these treatments have dangerous off-target effects (Garg et al., 2014; Greising et al., 2019; Larouche et al., 2022). More recently, work has demonstrated that local administration of the pro-regenerative cytokine IL-10 can improve the regeneration in skeletal muscle following VML (Huynh et al., 2023). Engineering iMPs to target inflammation and fibrosis - either by locally secreting anti-TGFβ antibodies or by counteracting inflammation using pro-regenerative cytokines such as IL-10 - could enhance regeneration after VML while mitigating the risks of systemic anti-TGFβ therapies (Borok et al., 2020; Narasimhulu and Singla, 2023). The capacity to generate inducible transgenes, controlled either temporally or spatially, would further improve the precision of these treatments (Figure 1).
In another example, we imagine a system in which iMPs engineered to constitutively express and secrete hormones responsible for blood glucose-homeostasis, might alleviate hyperglycemia in diabetic patients (Xie and Fussenegger, 2018). Synthetic circuits enabling glucose-mediated insulin secretion have been developed, bypassing the need for islet cells, which remain targeted by the immune system in type 1 diabetics receiving islet transplants (Xie et al., 2016). Type 2 diabetes might also be targeted using iMP-delivered synthetic gene circuits that secrete adiponectin to improve insulin sensitivity (Ye et al., 2017). Hypothetically, these synthetic gene circuits, applied to iMPs transplanted in otherwise healthy diabetic patients, could counteract hyperglycemia without the documented shortcomings of iPSC-based pancreatic beta cell therapies (Maxwell and Millman, 2021).
Thoughtful design of inducible synthetic transgenes should allow for iMPs to deliver therapeutics either locally or systemically. While the immediate application of these systems to target VML pathology are evident, the application of iMPs as a therapeutic delivery platform targeting systemic disease has yet to be discussed in the literature. The capacity for iMPs to persist long term as engrafted myofibers makes them an attractive alternative to other cell-based therapies that are often limited by poor engraftment efficiency and survivability (Levy et al., 2020; Zhou et al., 2021). We propose that future studies aiming to address disease using cell therapeutics consider the use of iMPs as a reliable delivery platform.
Temporal regulation of synthetic gene circuits is easily achieved through the use of inducible promoters, but the spatial organization of regenerating skeletal muscle may provide additional, therapeutically relevant, cues. Spatial transgene regulation has been achieved through the engineering of synthetic receptors capable of recognizing specific ligands, resulting in customizable sense/response behaviors (Roybal et al., 2016; Scheller et al., 2018). Synthetic notch (synNotch) receptors have proven a particularly powerful tool, capable of inducing the expression of myogenic transgenes in a spatially-regulated context (Morsut et al., 2016). Application of the synNotch system in vivo has demonstrated broad utility, from serving as an intercellular contact sensor capable of fate-mapping endothelial cells during development to enhancing CAR cell therapeutic efficacy (Hyrenius-Wittsten et al., 2021; Zhang et al., 2022). iMPs engineered to express customized synNotch constructs could provide spatial, context-dependent, regulation of transgene activation/deactivation. For example, pro-fibrotic collagens 3, 4 and 6 are significantly upregulated in the fibrotic lesion following VML (Hoffman et al., 2022). A self-regulating, cell-based synNotch system that uses antiCol3-tTa to control expression of anti-TGFβ antibody expression could enable targeted delivery, limiting expression to areas only where pro-fibrotic collagens are present (Figure 1).
When skeletal muscle is insufficiently innervated exogenous electrical stimulation can improve both the maturation and force production of the myofibers, aiding in the rehabilitation of muscle strength (Gordon and Mao, 1994; Crist et al., 2024). Optogenetic switches, which allow for transdermal stimulation of skeletal muscle using visible light, represents an attractive alternative to invasive patch clamps. Optogenetic switches have already proven capable of driving skeletal muscle contraction both in vitro and in vivo (Bruegmann et al., 2015; Ganji et al., 2021). Their therapeutic application as part of an iMP-based treatment should be investigated as it may allow for partial restoration of muscle function in spinal cord injury patients; or, at the very least, assist in the rehabilitation process.
The finding that myogenic differentiation is enhanced by prior PSM specification, whether it be in directed differentiation or direct reprograming protocols, has been fundamental to the generation of therapeutically relevant iMPs. Likewise, the ability to accurately purify the ERBB3+/NGFR+/CD57- myogenic Pax7+ population using cell sorting has considerably improved the purity of the resulting iMPs. Although imperfect, the current maturation capacity of these protocols allows for efficient engraftment of these cells and the generation of iMP-derived myofibers in vivo. While improvements within each of these categories are sure to further enhance the capacity for these cells to behave like adult MuSCs, using current iMPs as a therapeutic delivery platform opens the door to a wonderfully diverse and novel way of treating myopathies and systemic diseases alike. We encourage those working on cell-based therapies to consider new ways in which these cells can be employed and toward a future in which long-term iMP-based therapeutics can become a platform for therapeutics delivery.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
MH: Conceptualization, Writing–original draft, Writing–review and editing. FR: Conceptualization, Funding acquisition, Supervision, Writing–review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. MH is supported by Canadian Institutes of Health Research (FBD-181513) F.M.V.R. is supported by Canadian Institutes of Health Research (FDN-159908).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Awaya, T., Kato, T., Mizuno, Y., Chang, H., Niwa, A., Umeda, K., et al. (2012). Selective development of myogenic mesenchymal cells from human embryonic and induced pluripotent stem cells. PLoS One 7, e51638–e51639. doi:10.1371/journal.pone.0051638
Azzag, K., Bosnakovski, D., Tungtur, S., Salama, P., Kyba, M., and Perlingeiro, R. C. R. (2022). Transplantation of PSC-derived myogenic progenitors counteracts disease phenotypes in FSHD mice. NPJ Regen. Med. 7, 43–11. doi:10.1038/s41536-022-00249-0
Bakooshli, M. A., Lippmann, E. S., Mulcahy, B., Iyer, N., Nguyen, C. T., Tung, K., et al. (2019). A 3d culture model of innervated human skeletal muscle enables studies of the adult neuromuscular junction. Elife 8, 1–29. doi:10.7554/eLife.44530
Barberi, T., Bradbury, M., Dincer, Z., Panagiotakos, G., Socci, N. D., and Studer, L. (2007). Derivation of engraftable skeletal myoblasts from human embryonic stem cells. Nat. Med. 13, 642–648. doi:10.1038/nm1533
Basch, M. L., Bronner-Fraser, M., and García-Castro, M. I. (2006). Specification of the neural crest occurs during gastrulation and requires Pax7. Nature 441, 218–222. doi:10.1038/nature04684
Bersini, S., Gilardi, M., Ugolini, G. S., Sansoni, V., Talò, G., Perego, S., et al. (2018). Engineering an environment for the study of fibrosis: a 3D human muscle model with endothelium specificity and endomysium. Cell. Rep. 25, 3858–3868. doi:10.1016/j.celrep.2018.11.092
Borok, M. J., Mademtzoglou, D., and Relaix, F. (2020). Bu-M-P-ing iron: how BMP signaling regulates muscle growth and regeneration. J. Dev. Biol. 8, 4–14. doi:10.3390/JDB8010004
Bou Akar, R., Lama, C., Aubin, D., Maruotti, J., Onteniente, B., Esteves de Lima, J., et al. (2024). Generation of highly pure pluripotent stem cell-derived myogenic progenitor cells and myotubes. Stem Cell. Rep. 19, 84–99. doi:10.1016/j.stemcr.2023.11.002
Bruegmann, T., Van Bremen, T., Vogt, C. C., Send, T., Fleischmann, B. K., and Sasse, P. (2015). Optogenetic control of contractile function in skeletal muscle. Nat. Commun. 6, 7153–7158. doi:10.1038/ncomms8153
Castiglioni, A., Hettmer, S., Lynes, M. D., Rao, T. N., Tchessalova, D., Sinha, I., et al. (2014). Isolation of progenitors that exhibit myogenic/osteogenic bipotency in vitro by fluorescence-activated cell sorting from human fetal muscle. Stem Cell. Rep. 2, 92–106. doi:10.1016/j.stemcr.2013.12.006
Chal, J., Al Tanoury, Z., Hestin, M., Gobert, B., Aivio, S., Hick, A., et al. (2016). Generation of human muscle fibers and satellite-like cells from human pluripotent stem cells in vitro. Nat. Protoc. 11, 1833–1850. doi:10.1038/nprot.2016.110
Chal, J., Oginuma, M., Al Tanoury, Z., Gobert, B., Sumara, O., Hick, A., et al. (2015). Differentiation of pluripotent stem cells to muscle fiber to model Duchenne muscular dystrophy. Nat. Biotechnol. 33, 962–969. doi:10.1038/nbt.3297
Chal, J., and Pourquié, O. (2017). Making muscle: skeletal myogenesis in vivo and in vitro. Dev. Camb. 144, 2104–2122. doi:10.1242/dev.151035
Chang, H., Yoshimoto, M., Umeda, K., Iwasa, T., Mizuno, Y., Fukada, S., et al. (2009). Generation of transplantable, functional satellite-like cells from mouse embryonic stem cells. FASEB J. 23, 1907–1919. doi:10.1096/fj.08-123661
Choi, I. Y., Lim, H. T., Estrellas, K., Mula, J., Cohen, T. V., Zhang, Y., et al. (2016). Concordant but varied phenotypes among duchenne muscular dystrophy patient-specific myoblasts derived using a human iPSC-based model. Cell. Rep. 15, 2301–2312. doi:10.1016/j.celrep.2016.05.016
Corona, B. T., Rivera, J. C., Owens, J. G., Wenke, J. C., and Rathbone, C. R. (2015). Volumetric muscle loss leads to permanent disability following extremity trauma. J. Rehabil. Res. Dev. 52, 785–792. doi:10.1682/JRRD.2014.07.0165
Corona, B. T., Wu, X., Ward, C. L., McDaniel, J. S., Rathbone, C. R., and Walters, T. J. (2013). The promotion of a functional fibrosis in skeletal muscle with volumetric muscle loss injury following the transplantation of muscle-ECM. Biomaterials 34, 3324–3335. doi:10.1016/j.biomaterials.2013.01.061
Crist, S. B., Azzag, K., Kiley, J., Coleman, I., Magli, A., and Perlingeiro, R. C. R. (2024). The adult environment promotes the transcriptional maturation of human iPSC-derived muscle grafts. NPJ Regen. Med. 9, 16–11. doi:10.1038/s41536-024-00360-4
Darabi, R., Arpke, R. W., Irion, S., Dimos, J. T., Grskovic, M., Kyba, M., et al. (2012). Human ES- and iPS-derived myogenic progenitors restore DYSTROPHIN and improve contractility upon transplantation in dystrophic mice. Cell. Stem Cell. 10, 610–619. doi:10.1016/j.stem.2012.02.015
Fischer, K. M., Scott, T. E., Browe, D. P., McGaughey, T. A., Wood, C., Wolyniak, M. J., et al. (2020). Hydrogels for skeletal muscle regeneration. Regen. Eng. Transl. Med. 7, 353–361. doi:10.1007/s40883-019-00146-x
Ganji, E., Chan, C. S., Ward, C. W., and Killian, M. L. (2021). Optogenetic activation of muscle contraction in vivo. Connect. Tissue Res. 62, 15–23. doi:10.1080/03008207.2020.1798943
Garg, K., Corona, B. T., and Walters, T. J. (2014). Losartan administration reduces fibrosis but hinders functional recovery after volumetric muscle loss injury. J. Appl. Physiol. 117, 1120–1131. doi:10.1152/japplphysiol.00689.2014
Goldman, S. M., Henderson, B. E. P., Walters, T. J., and Corona, B. T. (2018). Co-delivery of a laminin-111 supplemented hyaluronic acid based hydrogel with minced muscle graft in the treatment of volumetric muscle loss injury. PLoS One 13, 01912455–e191315. doi:10.1371/journal.pone.0191245
Gordon, T., and Mao, J. (1994). Muscle atrophy and procedures for training after spinal cord injury. Phys. Ther. 74, 50–60. doi:10.1093/ptj/74.1.50
Greising, S. M., Corona, B. T., McGann, C., Frankum, J. K., and Warren, G. L. (2019). Therapeutic approaches for volumetric muscle loss injury: a systematic review and meta-analysis. Tissue Eng. Part B Rev. 25, 510–525. doi:10.1089/ten.teb.2019.0207
Greising, S. M., Rivera, J. C., Goldman, S. M., Watts, A., Aguilar, C. A., and Corona, B. T. (2017). Unwavering pathobiology of volumetric muscle loss injury. Sci. Rep. 7, 13179–13214. doi:10.1038/s41598-017-13306-2
Hicks, M. R., Hiserodt, J., Paras, K., Fujiwara, W., Eskin, A., Jan, M., et al. (2018). ERBB3 and NGFR mark a distinct skeletal muscle progenitor cell in human development and hPSCs. Nat. Cell. Biol. 20, 46–57. doi:10.1038/s41556-017-0010-2
Hoffman, D. B., Raymond-Pope, C. J., Sorensen, J. R., Corona, B. T., and Greising, S. M. (2022). Temporal changes in the muscle extracellular matrix due to volumetric muscle loss injury. Connect. Tissue Res. 63, 124–137. doi:10.1080/03008207.2021.1886285
Huynh, T., Reed, C., Blackwell, Z., Phelps, P., Herrera, L. C. P., Almodovar, J., et al. (2023). Local IL-10 delivery modulates the immune response and enhances repair of volumetric muscle loss muscle injury. Sci. Rep. 13, 1–15. doi:10.1038/s41598-023-27981-x
Hyrenius-Wittsten, A., Su, Y., Park, M., Garcia, J. M., Alavi, J., Perry, N., et al. (2021). SynNotch CAR circuits enhance solid tumor recognition and promote persistent antitumor activity in mouse models. Sci. Transl. Med. 13, eabd8836. doi:10.1126/scitranslmed.abd8836
Iberite, F., Gruppioni, E., and Ricotti, L. (2022). Skeletal muscle differentiation of human iPSCs meets bioengineering strategies: perspectives and challenges. NPJ Regen. Med. 7, 23–30. doi:10.1038/s41536-022-00216-9
Incitti, T., Magli, A., Darabi, R., Yuan, C., Lin, K., Arpke, R. W., et al. (2019). Pluripotent stem cell-derived myogenic progenitors remodel their molecular signature upon in vivo engraftment. Proc. Natl. Acad. Sci. U. S. A. 116, 4346–4351. doi:10.1073/pnas.1808303116
Jensen, J. B., Møller, A. B., Just, J., Mose, M., de Paoli, F. V., Billeskov, T. B., et al. (2021). Isolation and characterization of muscle stem cells, fibro-adipogenic progenitors, and macrophages from human skeletal muscle biopsies. Am. J. Physiol. Cell. Physiol. 321, C257–C268. doi:10.1152/ajpcell.00127.2021
Khodabukus, A., Madden, L., Prabhu, N. K., Koves, T. R., Jackman, C. P., Muoio, D. M., et al. (2019). Electrical stimulation increases hypertrophy and metabolic flux in tissue-engineered human skeletal muscle. Biomaterials 198, 259–269. doi:10.1016/j.biomaterials.2018.08.058
Kim, H., Selvaraj, S., Kiley, J., Azzag, K., Garay, B. I., and Perlingeiro, R. C. R. (2021). Genomic safe harbor expression of PAX7 for the generation of engraftable myogenic progenitors. Stem Cell. Rep. 16, 10–19. doi:10.1016/j.stemcr.2020.11.001
Kim, J., Oliveira, V. K. P., Yamamoto, A., and Perlingeiro, R. C. R. (2017). Generation of skeletal myogenic progenitors from human pluripotent stem cells using non-viral delivery of minicircle DNA. Stem Cell. Res. 23, 87–94. doi:10.1016/j.scr.2017.07.013
Kim, J. H., Kim, I., Seol, Y. J., Ko, I. K., Yoo, J. J., Atala, A., et al. (2020). Neural cell integration into 3D bioprinted skeletal muscle constructs accelerates restoration of muscle function. Nat. Commun. 11, 1025–1112. doi:10.1038/s41467-020-14930-9
Larouche, J. A., Kurpiers, S. J., Yang, B. A., Davis, C., Fraczek, P. M., Hall, M., et al. (2021). Neutrophil and natural killer cell imbalances prevent muscle stem cell mediated regeneration following murine volumetric muscle loss. bioRxiv 1, 2021. doi:10.1073/pnas.2111445119
Larouche, J. A., Wallace, E. C., Spence, B. D., Johnson, S. A., Kulkarni, M., Buras, E., et al. (2022). Spatiotemporal mapping of immune and stem cell dysregulation after volumetric muscle loss 2. doi:10.1101/2022.06.03.494707
Lepper, C., Partridge, T. A., and Fan, C. M. (2011). An absolute requirement for pax7-positive satellite cells in acute injury-induced skeletal muscle regeneration. Development 138, 3639–3646. doi:10.1242/dev.067595
Levy, O., Kuai, R., Siren, E. M. J., Bhere, D., Milton, Y., Nissar, N., et al. (2020). Shattering barriers toward clinically meaningful MSC therapies. Sci. Adv. 6, 1–18. doi:10.1126/sciadv.aba6884
Liu, L., Cheung, T. H., Charville, G. W., and Rando, T. A. (2015). Isolation of skeletal muscle stem cells by fluorescence-activated cell sorting. Nat. Protoc. 10, 1612–1624. doi:10.1038/nprot.2015.110
Luo, T., Tan, B., Zhu, L., Wang, Y., and Liao, J. (2022). A review on the design of hydrogels with different stiffness and their effects on tissue repair. Front. Bioeng. Biotechnol. 10, 817391–817418. doi:10.3389/fbioe.2022.817391
Maffioletti, S. M., Sarcar, S., Henderson, A. B. H., Mannhardt, I., Pinton, L., Moyle, L. A., et al. (2018). Three-dimensional human iPSC-derived artificial skeletal muscles model muscular dystrophies and enable multilineage tissue engineering. Cell. Rep. 23, 899–908. doi:10.1016/j.celrep.2018.03.091
Magli, A., Incitti, T., Kiley, J., Swanson, S. A., Darabi, R., Rinaldi, F., et al. (2017). PAX7 targets, CD54, integrin α9β1, and SDC2, allow isolation of human ESC/iPSC-Derived myogenic progenitors. Cell. Rep. 19, 2867–2877. doi:10.1016/j.celrep.2017.06.005
Mashinchian, O., De Franceschi, F., Nassiri, S., Michaud, J., Migliavacca, E., Aouad, P., et al. (2022). An engineered multicellular stem cell niche for the 3D derivation of human myogenic progenitors from iPSCs. EMBO J. 41, 1106555–e110717. doi:10.15252/embj.2022110655
Massih, B., Veh, A., Schenke, M., Mungwa, S., Seeger, B., Selvaraj, B. T., et al. (2023). A 3D cell culture system for bioengineering human neuromuscular junctions to model ALS. Front. Cell. Dev. Biol. 11, 996952–997011. doi:10.3389/fcell.2023.996952
Matthias, N., Hunt, S. D., Wu, J., Lo, J., Smith Callahan, L. A., Li, Y., et al. (2018). Volumetric muscle loss injury repair using in situ fibrin gel cast seeded with muscle-derived stem cells (MDSCs). Stem Cell. Res. 27, 65–73. doi:10.1016/j.scr.2018.01.008
Mauro, A. (1961). Satellite cell of skeletal muscle fibers. J. Biophys. Biochem. Cytol. 9, 493–495. doi:10.1083/jcb.9.2.493
Mavrommatis, L., Jeong, H. W., Kindler, U., Gomez-Giro, G., Kienitz, M. C., Stehling, M., et al. (2023). Human skeletal muscle organoids model fetal myogenesis and sustain uncommitted PAX7 myogenic progenitors. Elife 12, 1–26. doi:10.7554/eLife.87081
Maxwell, K. G., and Millman, J. R. (2021). Applications of iPSC-derived beta cells from patients with diabetes. Cell. Rep. Med. 2, 100238. doi:10.1016/j.xcrm.2021.100238
Mierzejewski, B., Archacka, K., Grabowska, I., Florkowska, A., Ciemerych, M. A., and Brzoska, E. (2020). Human and mouse skeletal muscle stem and progenitor cells in health and disease. Semin. Cell. Dev. Biol. 104, 93–104. doi:10.1016/j.semcdb.2020.01.004
Mizuno, Y., Chang, H., Umeda, K., Niwa, A., Iwasa, T., Awaya, T., et al. (2010). Generation of skeletal muscle stem/progenitor cells from murine induced pluripotent stem cells. FASEB J. 24, 2245–2253. doi:10.1096/fj.09-137174
Montarras, D., Morgan, J., Colins, C., Relaix, F., Zaffran, S., Cumano, A., et al. (2005). Developmental biology: Direct isolation of satellite cells for skeletal muscle regeneration. Science (1979) 309, 2064–2067. doi:10.1126/science.1114758
Morsut, L., Roybal, K. T., Xiong, X., Gordley, R. M., Coyle, S. M., Thomson, M., et al. (2016). Engineering customized cell sensing and response behaviors using synthetic notch receptors. Cell. 164, 780–791. doi:10.1016/j.cell.2016.01.012
Murphy, M. M., Lawson, J. A., Mathew, S. J., Hutcheson, D. A., and Kardon, G. (2011). Satellite cells, connective tissue fibroblasts and their interactions are crucial for muscle regeneration. Development 138, 3625–3637. doi:10.1242/dev.064162
Myogenica (2024). Available at: https://www.myogenica.com/science.
Nalbandian, M., Zhao, M., Sasaki-Honda, M., Jonouchi, T., Lucena-Cacace, A., Mizusawa, T., et al. (2021). Characterization of hiPSC-derived muscle progenitors reveals distinctive markers for myogenic cell purification toward cell therapy. Stem Cell. Rep. 16, 883–898. doi:10.1016/j.stemcr.2021.03.004
Narasimhulu, C. A., and Singla, D. K. (2023). BMP-7 attenuates sarcopenia and adverse muscle remodeling in diabetic mice via alleviation of lipids, inflammation, HMGB1, and pyroptosis. Antioxidants 12, 331. doi:10.3390/antiox12020331
Ordovás, L., Boon, R., Pistoni, M., Chen, Y., Wolfs, E., Guo, W., et al. (2015). Efficient recombinase-mediated cassette exchange in hPSCs to study the hepatocyte lineage reveals AAVS1 locus-mediated transgene inhibition. Stem Cell. Rep. 5, 918–931. doi:10.1016/j.stemcr.2015.09.004
Park, H., Bhalla, R., Saigal, R., Radisic, M., Watson, N., Langer, R., et al. (2008). Effects of electrical stimulation in C2C12 muscle constructs. J. Tissue Eng. Regen. Med. 2, 279–287. doi:10.1002/term.93
Pinton, L., Khedr, M., Lionello, V. M., Sarcar, S., Maffioletti, S. M., Dastidar, S., et al. (2023). 3D human induced pluripotent stem cell–derived bioengineered skeletal muscles for tissue, disease and therapy modeling. Nat. Protoc. 18, 1337–1376. doi:10.1038/s41596-022-00790-8
Pollot, B. E., Rathbone, C. R., Wenke, J. C., and Guda, T. (2018). Natural polymeric hydrogel evaluation for skeletal muscle tissue engineering. J. Biomed. Mater Res. B Appl. Biomater. 106, 672–679. doi:10.1002/jbm.b.33859
Quarta, M., Brett, J. O., DiMarco, R., De Morree, A., Boutet, S. C., Chacon, R., et al. (2016). An artificial niche preserves the quiescence of muscle stem cells and enhances their therapeutic efficacy. Nat. Biotechnol. 34, 752–759. doi:10.1038/nbt.3576
Quarta, M., Cromie, M., Chacon, R., Blonigan, J., Garcia, V., Akimenko, I., et al. (2017). Bioengineered constructs combined with exercise enhance stem cell-mediated treatment of volumetric muscle loss. Nat. Commun. 8, 15613–15617. doi:10.1038/ncomms15613
Quattrocelli, M., Swinnen, M., Giacomazzi, G., Camps, J., Barthélemy, I., Ceccarelli, G., et al. (2015). Mesodermal iPSC-derived progenitor cells functionally regenerate cardiac and skeletal muscle. J. Clin. Investigation 125, 4463–4482. doi:10.1172/JCI82735
Rao, L., Qian, Y., Khodabukus, A., Ribar, T., and Bursac, N. (2018). Engineering human pluripotent stem cells into a functional skeletal muscle tissue. Nat. Commun. 9, 126–212. doi:10.1038/s41467-017-02636-4
Relaix, F., and Zammit, P. S. (2012). “Satellite cells are essential for skeletal muscle regeneration: the cell on the edge returns centre stage,” Development, 139. 2845–2856. doi:10.1242/dev.069088
Roybal, K. T., Williams, J. Z., Morsut, L., Rupp, L. J., Kolinko, I., Choe, J. H., et al. (2016). Engineering T cells with customized therapeutic response programs using synthetic notch receptors. Cell. 167, 419–432. doi:10.1016/j.cell.2016.09.011
Sadelain, M., Papapetrou, E. P., and Bushman, F. D. (2012). Safe harbours for the integration of new DNA in the human genome. Nat. Rev. Cancer 12, 51–58. doi:10.1038/nrc3179
Sakai-Takemura, F., Narita, A., Masuda, S., Wakamatsu, T., Watanabe, N., Nishiyama, T., et al. (2018). Premyogenic progenitors derived from human pluripotent stem cells expand in floating culture and differentiate into transplantable myogenic progenitors. Sci. Rep. 8, 6555–6612. doi:10.1038/s41598-018-24959-y
Scheller, L., Strittmatter, T., Fuchs, D., Bojar, D., and Fussenegger, M. (2018). Generalized extracellular molecule sensor platform for programming cellular behavior. Nat. Chem. Biol. 14, 723–729. doi:10.1038/s41589-018-0046-z
Selvaraj, S., Mondragon-Gonzalez, R., Xu, B., Magli, A., Kim, H., Lainé, J., et al. (2019). Screening identifies small molecules that enhance the maturation of human pluripotent stem cell-derived myotubes. Elife 8, 479700–e48029. doi:10.7554/eLife.47970
Shelton, M., Kocharyan, A., Liu, J., Skerjanc, I. S., and Stanford, W. L. (2016). Robust generation and expansion of skeletal muscle progenitors and myocytes from human pluripotent stem cells. Methods 101, 73–84. doi:10.1016/j.ymeth.2015.09.019
Shin, S., Kim, S. H., Shin, S. W., Grav, L. M., Pedersen, L. E., Lee, J. S., et al. (2020). Comprehensive analysis of genomic safe harbors as target sites for stable expression of the heterologous gene in HEK293 cells. ACS Synth. Biol. 9, 1263–1269. doi:10.1021/acssynbio.0c00097
Sun, C., Kannan, S., Choi, I. Y., Lim, H. T., Zhang, H., Chen, G. S., et al. (2022). Human pluripotent stem cell-derived myogenic progenitors undergo maturation to quiescent satellite cells upon engraftment. Cell. Stem Cell. 29, 610–619.e5. doi:10.1016/j.stem.2022.03.004
Sun, C., Serra, C., Kalicharan, B. H., Harding, J., and Rao, M. (2024). Challenges and considerations of preclinical development for iPSC-based myogenic cell therapy. Cells 13, 596–626. doi:10.3390/cells13070596
Takahashi, K., and Yamanaka, S. (2006). Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 126, 663–676. doi:10.1016/j.cell.2006.07.024
Tanaka, A., Woltjen, K., Miyake, K., Hotta, A., Ikeya, M., Yamamoto, T., et al. (2013). Efficient and reproducible myogenic differentiation from human iPS cells: prospects for modeling miyoshi myopathy in vitro. PLoS One 8, e61540. doi:10.1371/journal.pone.0061540
Tedesco, F. S., Gerli, M. F. M., Perani, L., Benedetti, S., Ungaro, F., Cassano, M., et al. (2012). Transplantation of genetically corrected human iPSC-derived progenitors in mice with limb-girdle muscular dystrophy. Sci. Transl. Med. 4, 140ra89–13. doi:10.1126/scitranslmed.3003541
Tey, S. R., Robertson, S., Lynch, E., and Suzuki, M. (2019). Coding cell identity of human skeletal muscle progenitor cells using cell surface markers: current status and remaining challenges for characterization and isolation. Front. Cell. Dev. Biol. 7, 284. doi:10.3389/fcell.2019.00284
Uezumi, A., Nakatani, M., Ikemoto-Uezumi, M., Yamamoto, N., Morita, M., Yamaguchi, A., et al. (2016). Cell-surface protein profiling identifies distinctive markers of progenitor cells in human skeletal muscle. Stem Cell. Rep. 7, 263–278. doi:10.1016/j.stemcr.2016.07.004
Urciuolo, A., Serena, E., Ghua, R., Zatti, S., Giomo, M., Mattei, N., et al. (2020). Engineering a 3D in vitro model of human skeletal muscle at the single fiber scale. PLoS One 15, 02320811–e232117. doi:10.1371/journal.pone.0232081
van der Wal, E., Iuliano, A., t Groen, S. L. M., Bholasing, A. P., Priesmann, D., Sharma, P., et al. (2023). Highly contractile 3D tissue engineered skeletal muscles from human iPSCs reveal similarities with primary myoblast-derived tissues. Stem Cell. Rep. 18, 1954–1971. doi:10.1016/j.stemcr.2023.08.014
Von Maltzahn, J., Jones, A. E., Parks, R. J., and Rudnicki, M. A. (2013). Pax7 is critical for the normal function of satellite cells in adult skeletal muscle. Proc. Natl. Acad. Sci. U. S. A. 110, 16474–16479. doi:10.1073/pnas.1307680110
Wolf, M. T., Dearth, C. L., Sonnenberg, S. B., Loboa, E. G., and Badylak, S. F. (2015). Naturally derived and synthetic scaffolds for skeletal muscle reconstruction. Adv. Drug Deliv. Rev. 84, 208–221. doi:10.1016/j.addr.2014.08.011
Wosczyna, M. N., and Rando, T. A. (2018). A muscle stem cell support group: coordinated cellular responses in muscle regeneration. Dev. Cell. 46, 135–143. doi:10.1016/j.devcel.2018.06.018
Wu, J., Matthias, N., Bhalla, S., and Darabi, R. (2021). Evaluation of the therapeutic potential of human iPSCs in a murine model of VML. Mol. Ther. 29, 121–131. doi:10.1016/j.ymthe.2020.09.012
Wu, J., Matthias, N., Lo, J., Ortiz-Vitali, J. L., Shieh, A. W., Wang, S. H., et al. (2018). A myogenic double-reporter human pluripotent stem cell line allows prospective isolation of skeletal muscle progenitors. Cell. Rep. 25, 1966–1981. doi:10.1016/j.celrep.2018.10.067
Xi, H., Langerman, J., Sabri, S., Chien, P., Young, C. S., Younesi, S., et al. (2020). A human skeletal muscle atlas identifies the trajectories of stem and progenitor cells across development and from human pluripotent stem cells. Cell. Stem Cell. 27, 158–176. doi:10.1016/j.stem.2020.04.017
Xie, M., and Fussenegger, M. (2018). Designing cell function: assembly of synthetic gene circuits for cell biology applications. Nat. Rev. Mol. Cell. Biol. 19, 507–525. doi:10.1038/s41580-018-0024-z
Xie, M., Ye, H., Wang, H., Charpin-El Hamri, G., Lormeau, C., Saxena, P., et al. (2016). β-cell-mimetic designer cells provide closed-loop glycemic control. Science 354 (1979), 1296–1301. doi:10.1126/science.aaf4006
Ye, H., Xie, M., Xue, S., Hamri, G. C.El, Yin, J., Zulewski, H., et al. (2017). Self-adjusting synthetic gene circuit for correcting insulin resistance. Nat. Biomed. Eng. 1, 0005–0009. doi:10.1038/s41551-016-0005
Yi, L., and Rossi, F. (2011). Purification of progenitors from skeletal muscle. J. Vis. Exp. 2, 2476–2477. doi:10.3791/2476
Zhang, S., Zhao, H., Liu, Z., Liu, K., Zhu, H., Pu, W., et al. (2022). Monitoring of cell-cell communication and contact history in mammals. Science 1979, eabo5503. doi:10.1126/science.abo5503
Keywords: myogenic progenitor cells, muscle regeneration, cell therapeutic, synthetic biology (synbio), muscle stem cell (MuSC), iPSCs (induced pluripotent stem cells)
Citation: Hamer MS and Rossi FMV (2025) Multitasking muscle: engineering iPSC-derived myogenic progenitors to do more. Front. Cell Dev. Biol. 12:1526635. doi: 10.3389/fcell.2024.1526635
Received: 12 November 2024; Accepted: 23 December 2024;
Published: 22 January 2025.
Edited by:
Masatoshi Suzuki, University of Wisconsin-Madison, United StatesReviewed by:
Christopher M. Madl, University of Pennsylvania, United StatesCopyright © 2025 Hamer and Rossi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fabio M. V. Rossi, ZmFiaW9AYnJjLnViYy5jYQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.