 Koen A. E. Keijzer
Koen A. E. Keijzer Erika Tsingos
Erika Tsingos Roeland M. H. Merks1,2*
Roeland M. H. Merks1,2*- 1Mathematical Institute, Faculty of Science, Leiden University, Leiden, Netherlands
- 2Institute of Biology Leiden, Faculty of Science, Leiden University, Leiden, Netherlands
Many mammalian cells, including endothelial cells and fibroblasts, align and elongate along the orientation of extracellular matrix (ECM) fibers in a gel when cultured in vitro. During cell elongation, clusters of focal adhesions (FAs) form near the poles of the elongating cells. FAs are mechanosensitive clusters of adhesions that grow under mechanical tension exerted by the cells’ pulling on the ECM and shrink when the tension is released. In this study, we use mathematical modeling to study the hypothesis that mechanical reciprocity between cells and the ECM is sufficient for directing cell shape changes and orientation. We show that FAs are preferentially stabilized along the orientation of ECM fibers, where cells can generate higher tension than in directions perpendicular to the ECM fibers. We present a hybrid computational model coupling three mathematical approaches: first, the cellular Potts model (CPM) describes an individual contractile cell; second, molecular dynamics (MD) represent the ECM as a network of cross-linked, deformable fibers; third, a set of ordinary differential equations (ODEs) describes the dynamics of the cell’s FAs, in terms of a balance between assembly and a mechanoresponsive disassembly. The resulting computational model shows that mechanical reciprocity suffices for stiffness-dependent cell spreading, local ECM remodeling, and ECM-alignment-dependent cell elongation. These combined effects are sufficient to explain how cell morphology is influenced by the local ECM structure and mechanics.
1 Introduction
The extracellular matrix (ECM) plays a crucial role in development and in disease. For example, the ECM plays a role in cancer cell migration (Najafi et al., 2019; Yamaguchi et al., 2005), wound healing (Maquart and Monboisse, 2014; Diller and Tabor, 2022), and angiogenesis (Stupack and Cheresh, 2002). The ECM is a complex collection of large fibers such as collagen, fibronectin, and other proteins (Theocharis et al., 2016). The orientation of fibers in the ECM plays an important role in tumor vascularization (Balcioglu et al., 2016), mechanical cell–cell communication (Nahum et al., 2023), and blood clot formation (Kim O. V. et al., 2017). The ECM is continuously remodeled by cells both chemically, through the synthesis and degradation of ECM fibers and associated components, and mechanically, by pulling and reorienting fibers (Theocharis et al., 2016; Winkler et al., 2020). As ECM remodeling leads to local changes in ECM properties such as stiffness, structure, density, and isotropy, to which cells respond through changes in adhesion, cell contraction, or pseudopod extension (Reinhart-King et al., 2005; Malandrino et al., 2019; Doyle et al., 2022), there is a bidirectional chemical and mechanical reciprocity between the cells and the ECM. In this work, we focus specifically on the mathematical modeling of mechanical cell–ECM reciprocity in fibrous ECM, in particular the role of ECM isotropy. For mathematical models of other forms of cell–ECM reciprocity, we refer to Daub and Merks (2013); van Oers et al. (2014); Rens and Merks (2017), Rens and Merks (2020); Chiang and Chung (2024).
The present study attempts to provide mechanistic explanations for three behaviors of cells on fibrous matrices: (1) cell spreading as a function of ECM stiffness, (2) alignment of cells to ECM fiber orientation, and (3) a hypothetical role of ECM anisotropy in mechanical cell–ECM reciprocity.
First, certain cell types, such as endothelial cells, fibroblasts, smooth muscle cells, and osteogenic cells, show a monotonic increase in spreading with substrate stiffness. These cells are relatively small on softer substrates, elongate on intermediate substrates, and achieve maximum spreading on highly stiff substrates such as coated glass (Yang et al., 2017). Other cell types, including Jurkat T cells and NIH 3T3 fibroblasts, show a biphasic response of spreading to substrate stiffness, showing maximal cell spreading at an intermediate optimal level of substrate stiffness (Oakes et al., 2018; Wahl et al., 2019; Janmey et al., 2020).
Second, cell alignment is influenced both by the mechanical properties of the fibers and by the cell adhesion properties. In Friedrichs et al. (2007), cells are cultured on a two-dimensional substrate assembled out of thin aligned collagen fibrils. Cells align along the collagen fibrils and bundle parallel fibrils together at their poles and deform the orthogonal fibrils, which creates holes in the substrate. When this experiment was repeated with fragile fibrils, the cell did not elongate, and the fibers surrounding the cell were digested. This suggests that cells require a firm ECM to adhere to and that anisotropic traction force is required to elongate and align to the fibrils. Next, they found that the cell adhesions to the fibrils influence cell alignment. In general, cells adhere to the ECM with integrins, which are membrane-piercing receptors that bind to proteins in the ECM with varying binding strengths, possibly regulated by mechanical tension (Kechagia et al., 2019). Specifically, Friedrichs et al. (2007) found that cells expressing the integrin
Finally, cells not only respond to cues in the ECM but also reorient the fibers in the ECM. Contractile breast cancer cells deform fibrous collagen and reorient the fibers to point toward themselves. Pairs of these contractile cells create aligned bridges of fibers between them (Kim J. et al., 2017).
Computational modeling is well-suited for providing insights into mechanical cell–ECM reciprocity (Crossley et al., 2024). Before introducing our own approach, we briefly review a selection of specific computational models of cell–ECM reciprocity involving mechanosensitive adhesions and a fibrous ECM. We highlight two factors that are crucial to be included in a computational model of the reciprocity between ECM fiber alignment and cell morphology, namely, (i) cell-induced changes in ECM structure and (ii) ECM-induced changes in cell shape.
We first review computational models focused on ECM mechanics in response to cell contraction (i). Using a 3D finite element (FE) representation of the ECM, Paukner et al. (2023) showed that cell contractility and force-dependent cell–ECM adhesions suffice for guiding cell migration upward stiffness gradients. This model focused on ECM deformation by the cell but could not capture cell shape change due to changes in the ECM because the cell was modeled as a point particle with an adhesive annulus. They concluded that cell contractility, combined with mechanosensitive cell–ECM adhesions, can explain several phenomena in cell migration. In a different study of cell migration, Feng et al. (2019) introduced a simple bead–spring network approximation of a deformable ECM and a migrating ellipsoidal cell. They showed that a torque balance on the mechanosensitive adhesions of the cell causes the cell to orient along fibers, after which the cell starts migrating. In Feng et al. (2019), the cell’s ability to sense the fiber orientation disappears if the fibers’ bending modulus is too high, showing that in this model, fiber orientation is sensed through mechanical interactions with the fibers. A model that links a fibrous network with breakable cross-links to a circular radial cell suggests that fiber accumulation can enhance cell–ECM adhesion by increasing the number of available binding sites for cellular adhesion (Cao et al., 2017). Altogether, these computational models have studied the potential effect of cell contractility on the ECM, but they did not include the reciprocal effects of the ECM on the cell (ii).
A number of models have considered only (ii), the effect of the ECM on cell behavior. For example, Vargas et al. (2020) showed how different cell migration modes can emerge based on adhesion maturation and stress fiber strength using a 3D finite element model of a moving cell on a non-fibrous, uniformly structured ECM. A different finite element model of cell migration showed how cell deformation and ECM porosity are of primary importance in amoeboid cell migration (Campbell and Bagchi, 2021).
Models combining (i) and (ii), thus closing the loop to full mechanical ECM reciprocity, include those by van Oers et al. (2014), Rens and Merks (2017), and Rens and Merks (2020). In these models, cell shape is modeled using the cellular Potts model (CPM) and coupled to a finite element (FEM) simulation of the ECM to form a hybrid CPM. Early CPM–ECM couplings assumed that cellular protrusions are stabilized on highly stressed substrates (van Oers et al., 2014), showing how mechanical cell–cell communication can play a role in angiogenesis. Subsequently, this coupling was extended by including a comprehensive model of mechanosensitive adhesion between the cell and the ECM, leading to emergent cell spreading, spontaneous cell elongation, and durotaxis (Rens and Merks, 2020). Although these models consider full mechanical reciprocity, their ECM is homogeneous, i.e., there are no fibers. One of the first models of mechanical cell–ECM reciprocity featuring a fibrous ECM was used to explain how bands form between two contractile cells in a fibrous ECM and how the two cells elongate toward each other by the remodeled matrix (see Reinhardt et al., 2013, reviewed in Crossley et al., 2024). Another sophisticated model of cell–ECM reciprocity is that of Kim M. C et al. (2018), where a triangulated dynamic cell is coupled to a fibrous ECM using an FEM simulation. They studied how cells can sense local stiffness in the fiber network by considering filopodia–fiber binding. In this study, we build upon our previously introduced hybrid CPM and molecular dynamics (MD) model (Tsingos et al., 2023). This model approximated the ECM by representing fibers using a beads-and-spring model, where the fibers were linked using cross-linkers. In this work, we modeled how the cell’s contractions form the ECM and how these deformations propagate far into the network. Furthermore, the cells’ contractile forces are counteracted by the ECM, leading to less contraction in a highly cross-linked, stiff ECM and high cellular contraction in a soft ECM. In this model, the cellular adhesions were static, and new adhesions could not be formed with the ECM.
To study the alignment of cells in anisotropic networks, we extend our previous hybrid CPM and molecular dynamics model (Tsingos et al., 2023) with dynamic adhesions. The choice of how to couple cellular morphology and ECM dynamics is delicate as it encodes the biological hypothesis of how cells sense and react with the ECM. In this work, we adopt the coupling between cell and ECM proposed by Rens and Merks (2020) and apply it to a hybrid CPM with discrete fibrous ECM (Tsingos et al., 2023). The coupling is made by assuming that the cell exerts cytoskeletal contraction forces through integrin-based adhesions that behave according to the two-spring model (Schwarz et al., 2006; Doyle et al., 2022). In essence, the two-spring model views adhesion as a mediator between the contractile forces of the cell and the restoring forces of the ECM. The tension on the adhesion builds up slowly on the soft ECM and quickly on the stiff ECM as the cell applies its contractile forces. Additionally, we assume that adhesions strengthen as tension increases. Adhesion strength is quantified by the number of integrin proteins bound to the adhesion. These assumptions, when combined with an isotropic material in a hybrid CPM, are sufficient to produce phenomena such as cell spreading, spontaneous elongation, and durotaxis (Rens and Merks, 2020). We implement this two-spring adhesion model in the hybrid CPM with a discrete fibrous ECM (Tsingos et al., 2023) and use the new model to investigate the reciprocity between fiber orientation and cell morphology. Specifically, with this new fibrous ECM model, we show how cell elongation on oriented gels can be considered a special case of stiffness-dependent cell spreading as fibers are easier to bend than to stretch. Furthermore, we study how cell protrusions can reorient fibers, thereby increasing tension on adhesions and stabilizing the protrusions.
2 Methods
2.1 Modeling approach
We have introduced dynamic descriptions of mechanosensitive focal adhesions (FAs) into a hybrid CPM and MD model (Tsingos et al., 2023). The CPM part dynamically describes cell shape changes, and the MD part simulates a cross-linked network of ECM fibers and its dynamical response to cellular forces. In our previous work, the CPM was connected to the MD model through static adhesion particles. In the present model, the buildup and breakdown of FAs are modeled dynamically using an ordinary differential equation model that describes FAs as clusters of integrins, with the breakdown rate assumed to be dependent on the mechanical tension within the FAs. This ordinary differential equation (ODE) model for FAs and their constituent integrins was adapted from the work by Novikova and Storm (2013), as shown in one of our previous models featuring a continuum description of the ECM (Rens and Merks, 2020). Figure 1 provides an overview of the key elements of the model.
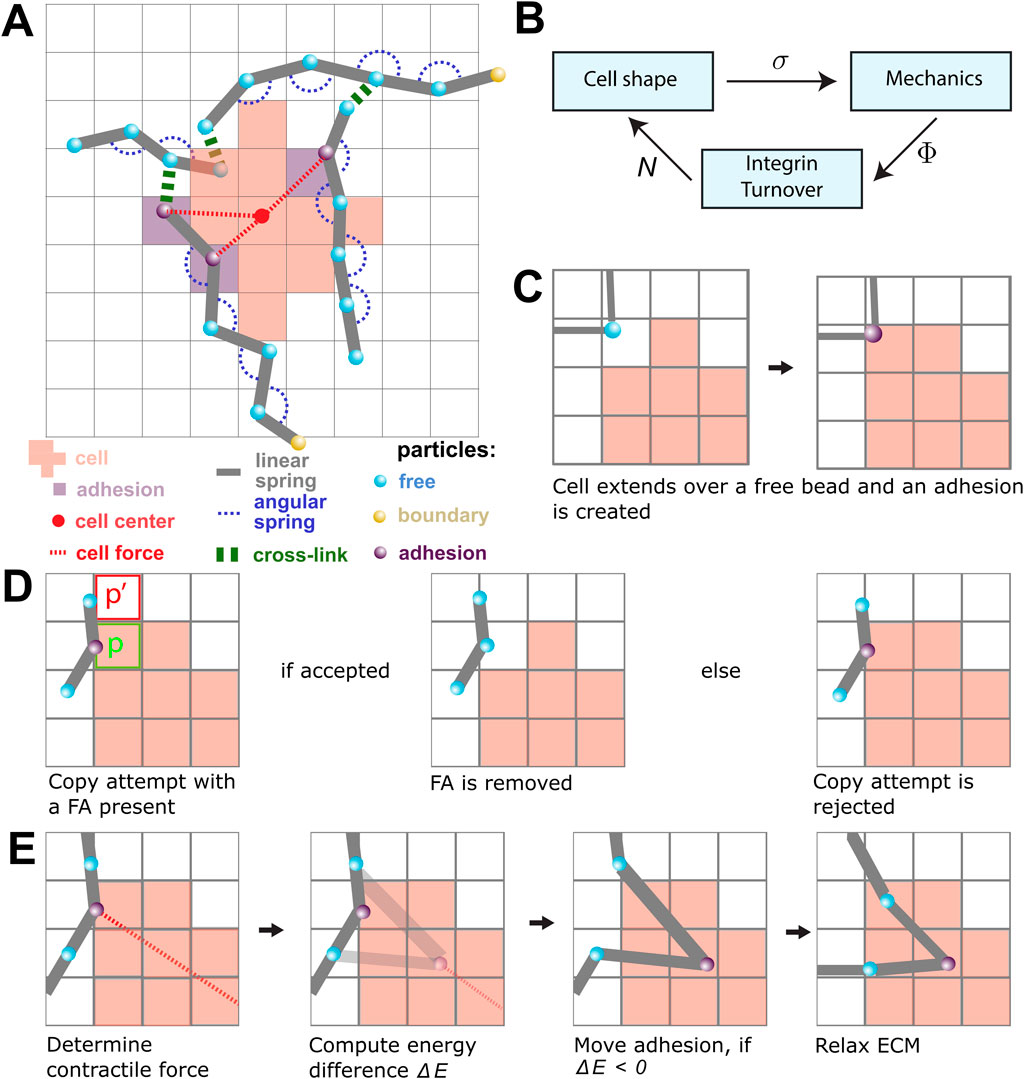
Figure 1. An overview of the model components and the model rules. (A) A schematic overview of the components in the model. The lattice-bound cellular Potts cell (red squares) is attached to the beads (cyan) and springs (gray) of the ECM through the focal adhesions (FAs; purple). The contractile force (dashed red) is visualized between FAs and the center of mass of the cell (red point). FAs connect the cell to the strands of the ECM. The strands are cross-linked (green), and particles outside the simulation domain are fixed (yellow). (B) The submodels of the model are coupled as shown in this figure. The CPM is responsible for the cell shape and sends the lattice state
The models are coupled using an operator-splitting approach. The three submodels are sequentially iterated to a steady state, where the output state of one submodel is used as the input state for the next submodel (Figure 1B). The simulations were run until a quasi-steady state was reached, i.e., until no large further changes were observed. In the remainder of this section, we will describe each of the three submodels and the coupling strategies.
2.2 Cellular Potts model
To describe cell shape changes, we employ the cellular Potts model (Graner and Glazier, 1992; Hirashima et al., 2017). The CPM is a lattice-based model in which cell shape is defined as a collection of connected lattice sites. We implemented a CPM on a square grid
The CPM evolves through a sequence of random extensions and retractions, whose probability is given by a balance of contractile and extensile forces and forces due to adhesion with the ECM. These are given by a Hamiltonian energy function.
where
The Hamiltonian is minimized through Metropolis dynamics, as previously described by Graner and Glazier (1992), thus dynamically updating the cell’s shape. In brief, we iteratively select a random lattice site
The acceptance probability of a copy attempt is determined by the energy change:
where
The term
where
2.3 Extracellular matrix model
The ECM is described as a set of fibers connected through cross-linkers, forming a fiber network that is superimposed on the CPM lattice (Figure 1A). A fiber is built out of
To create a fiber network, we followed the method introduced by Tsingos et al. (2023) with small modifications for creating networks of aligned fibers. In brief, we distributed
where
The springs connecting pairs of beads and the bending rigidity connecting triples of beads impose forces on the ECM and make the fiber network dynamic. The positions of the beads
where
The energy of a single spring with a rest length
where
where
2.4 Focal adhesions
FAs, schematically shown as purple beads in Figure 1A, are modeled as clusters of catch-slip bonds (Novikova and Storm, 2013; Rens and Merks, 2020; Schwarz et al., 2006). Each cluster is assumed to be in constant flux as integrins are added and removed from the cluster. The integrin addition rate is independent, whereas the removal rate is suppressed by mechanical tension due to the contractile force of the cell and the restoring force from the ECM. The number of integrins
where
where
The tension
2.5 Parameter values
Table 1 lists the parameter values used for the CPM and MD models. They are dimensionless and require scaling to fit to measurable quantities. We follow the previous work by Tsingos et al. (2023) for this scaling, and we briefly summarize the main points in this section. A single lattice site of the CPM is set equal to
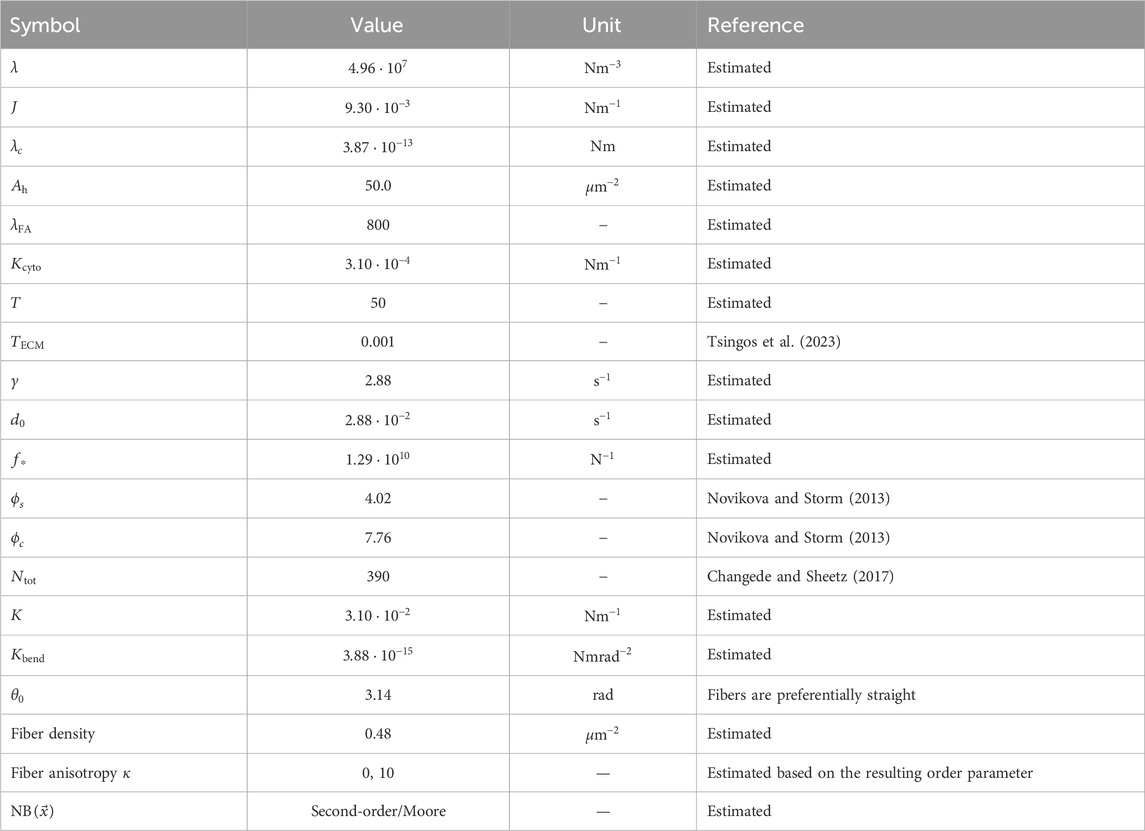
Table 1. Parameter values used in the simulations unless otherwise specified. The parameter values reported here are the scaled values, which is why they may appear as non-rounded numbers despite being chosen values.
2.6 Statistical significance
The error bars shown in Figures 2–4 denote the mean
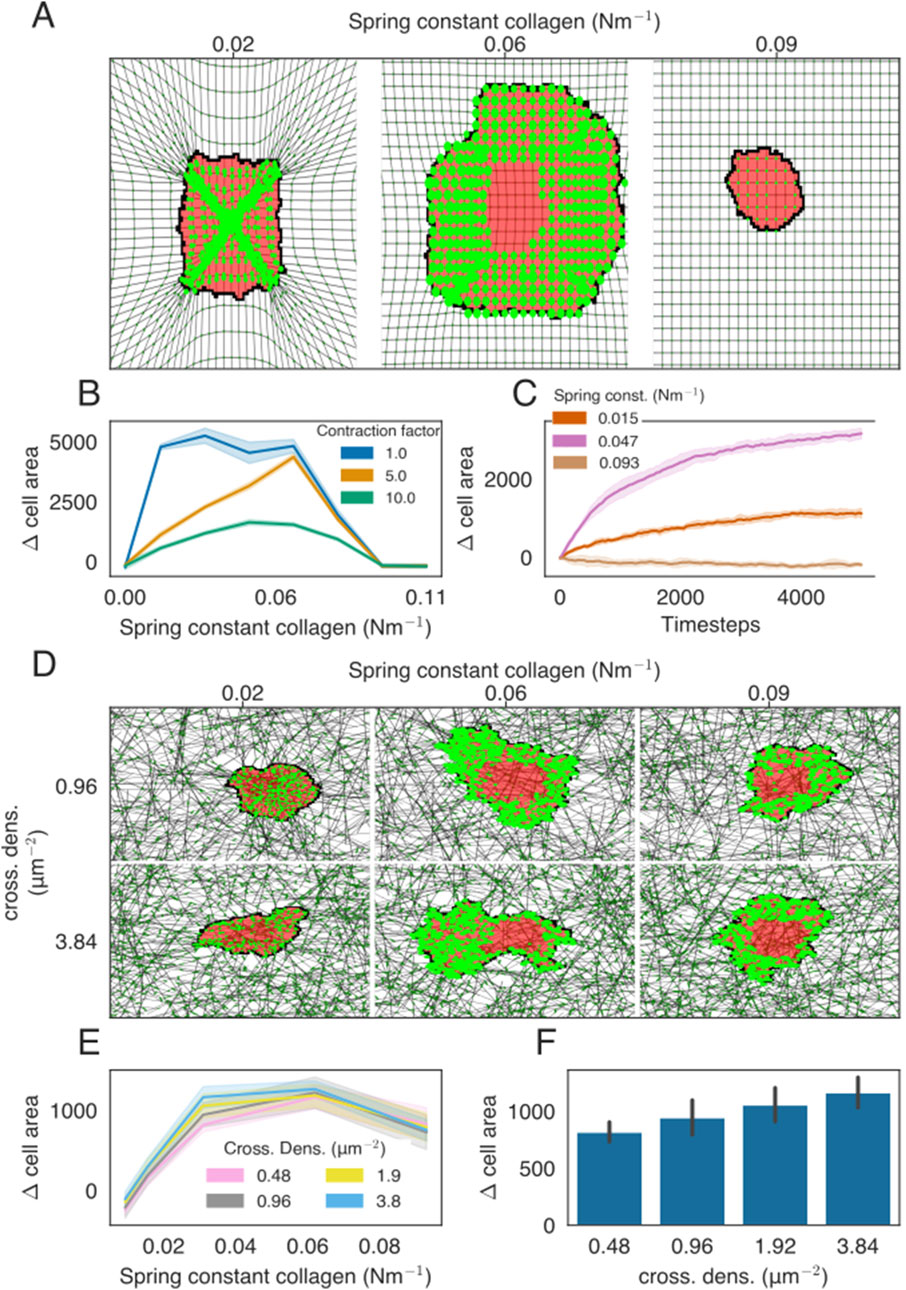
Figure 2. Cell spreading on isotropic ECMs with regular or network structures. (A) Example of cell spreading on an isotropic ECM for three different spring stiffness values. The cell is colored in red, FAs are presented as bright-green discs, ECM strands are in gray, and ECM crosslinks are shown in dark green. The radius of the bright-green discs is proportional to the size of the FAs. All FAs are assigned to only a single lattice site, even if the visualization may suggest otherwise. (B) Difference in the cell area from the starting size for the ECM of different stiffness. Colors indicate the factor by which the contraction force of the cell is multiplied. (C) Time-evolution of the cell area as a function of ECM spring stiffness. (D) Simulation snapshots of cell spreading on the isotropic ECM for a range of spring constants and cross-link densities. (E) Difference in the cell area from the starting size, at
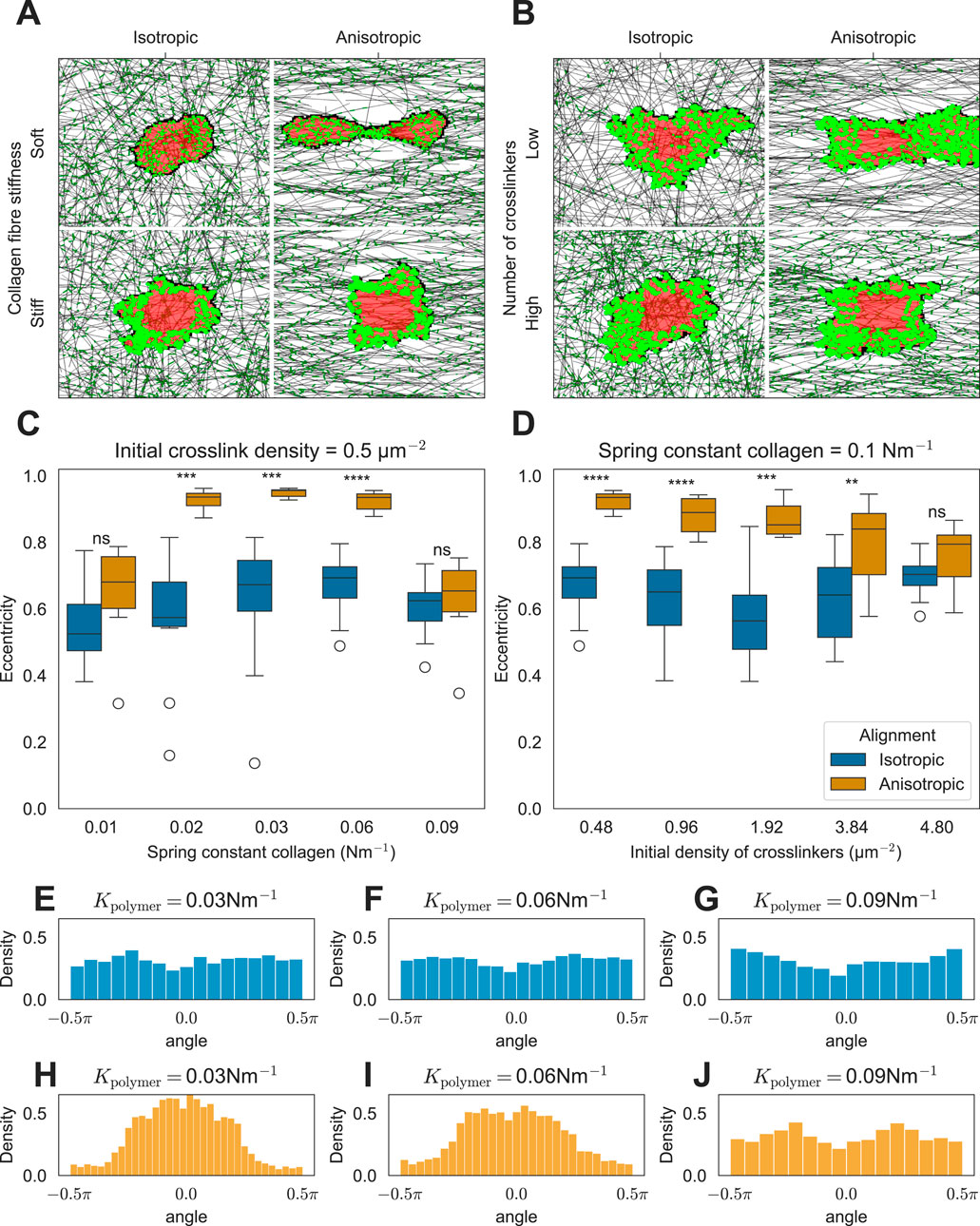
Figure 3. Effect of ECM anisotropy on the cell shape. (A) Snapshots of simulations showing the effect of matrix anisotropy on the cell shape as a function of collagen stiffness (K =
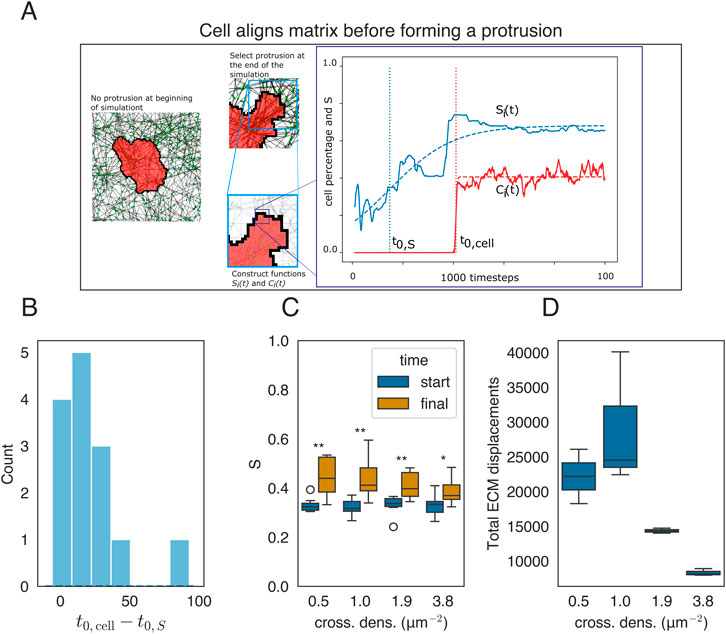
Figure 4. Mechanical reciprocity between local cell spreading and local ECM alignment. (A) Cell state at two time points of the simulation corresponding to
3 Results
3.1 Stiffness-dependent cell spreading
Cell spreading emerges in models of mechanosensitive dynamic FAs with a uniform isotropic ECM (Rens and Merks, 2020), suggesting that we should be able to capture cell spreading in the present model as well. To stay close to the simulated case of the isotropic linear elastic material used previously, we first studied cell spreading on a homogeneous matrix constructed by creating long vertical and horizontal strands and cross-linking them at the intersections. On this homogeneous matrix, the cell’s spreading area depends on matrix stiffness in a biphasic manner. Up to an optimum stiffness, the cell-spreading area increases with matrix stiffness, after which the cell spreading area decreases with matrix stiffness (Figures 2A, B; Supplementary Videos S1, S2). After this optimum stiffness, the tension within the FAs reaches the slip regime of the integrins within the FAs. This biphasic effect was not observed in the previous model with a uniform ECM (Rens and Merks, 2020) or in endothelial cells (Reinhart-King et al., 2005). However, such a biphasic response of cell spreading to matrix stiffness was observed in fibroblasts and T-cells (Oakes et al., 2018; Wahl et al., 2019). Interestingly, the biphasic effect disappears when they increase the lifetimes of the integrin–ECM bonds (Oakes et al., 2018). To test whether our model is consistent with this experimental observation, we doubled the slip parameter
In Rens and Merks (2020), a mechanism based on mechanosensitive FAs was found to explain cell spreading, cell elongation, and durotaxis on a homogeneous, regular ECM. Having implemented a conceptually similar model, we discovered the relationship between ECM stiffness and cell spreading on a homogeneous, regular ECM. Next, we studied the relationship between ECM stiffness and cell spreading on an inhomogeneous, randomized, but isotropic ECM. We constructed isotropic randomized matrices, as described in Section 2.3, by distributing elastic strands of roughly one cell length and cross-linking them together. We observed a biphasic effect on cell spreading when increasing the spring stiffness of the network (Figure 2E; Supplementary Videos S4, S5). We also modified ECM stiffness by changing the cross-link density (Figure 2F). The cell spreading area also increased with cross-linker density, which is consistent with 2D in vitro experiments (Mullen et al., 2015).
3.2 Cells elongate on the anisotropic matrix due to the local anisotropy of stiffness cues
In vitro, cells extend along fibers in the substrate (Lai et al., 2012; Friedrichs et al., 2007; Sapudom et al., 2023; Chaubaroux et al., 2015). We hypothesize that the mechanism driving such elongation is similar to the mechanism of stiffness-dependent cell spreading on a regular, isotropic ECM described in the previous section. We, therefore, asked how ECM isotropy affects cell spreading. Figures 3A, B show example simulations on inhomogeneous matrices with and without anisotropy. Cells placed on anisotropic matrices elongate along the axis of anisotropy, provided that the fibers are not excessively stiff (Figures 3A, C) and that the network is not overly cross-linked (Figures 3B, D). This phenomenon occurs because FAs stabilize rapidly under increased tension. In particular, in the ECM composed of parallel fibers, the difference in tension built up along or orthogonal to the fibers is large as fibers resist extension more effectively than bending. Consequently, FAs stabilize more rapidly in response to force along the ECM compared to force along the orthogonal orientation, resulting in the observed cell alignment. Increasing the bending modulus of ECM fibers leads to the creation of cellular protrusions orthogonal to the ECM orientation (Supplementary Figure S3A). However, these protrusions are small, and the cell still aligns with the ECM fibers (Supplementary Figure S3B).
Consistent with in vitro observations, our model simulations predict that FAs form preferentially at the poles of the cells (Figures 3A, B) (Friedrichs et al., 2007; Chaubaroux et al., 2015; Lai et al., 2012). The angles between the cell elongation axis and the FAs are roughly uniformly distributed on isotropic matrices (Figures 3E–G). By contrast, on anisotropic matrices, the FAs are centered at the poles of the elongated cell (see single peaks at approximately 0 radians in Figures 3H, I). This effect was independent of collagen fiber stiffness. For the highest stiffness tested, cells no longer elongated, and FAs become uniformly distributed around the cell (Figure 3J). Similarly, if we increased the degree of cross-linking in anisotropic ECM with intermediate fiber stiffness, the cells failed to elongate (Figures 3B, D; Supplementary Videos S8, S9). In both cases, the fibers provided more resistance to contraction forces perpendicular to the axis of anisotropy than in the low-stiffness case, either due to increased fiber stiffness or increased cross-linking. As a result, the tension in the FAs, pulling perpendicular to the axis of anisotropy, became sufficiently strong for FA maturation, leading to cell rounding. Next, we tested the behavior of the model on strongly cross-linked ECM consisting of stiff fibers. In this situation, our model predicts that cells orient perpendicularly to the axis of ECM anisotropy (Supplementary Figure S4; Supplementary Video S10) because the FAs along the direction of fiber orientation enter the slipping regime and break. Hence, the cell elongates orthogonal to the fiber orientation. Finally, we looked at how the degree of isotropy (as quantified by the order parameter
3.3 Cell remodels fibers before it starts spreading
In the previous sections, we studied how static, global anisotropies of the ECM can affect cell shape. We next looked at potential cell shape changes due to mechanical reciprocity between the cell and the local ECM, potentially driving local ECM anisotropy. We studied the time evolution of the local order parameter
The functions
Figure 4A shows two states of a model simulation, one shortly after initialization and one after
and plotted the distribution of
The cells not only remodel the ECM at the pseudopodia but also seem to remodel the ECM all around the cell. We, therefore, quantified the alignment around the cell by taking the average of order parameters in an annulus around the cell given by
where
4 Discussion
In this paper, we studied the effect of mechanical cell–matrix reciprocity across different ECMs. Assuming that tension-mediated integrin turnover drives FA maturation, we find that FA formation depends on both the angle of the ECM fibers with the cell and the cell’s contractile force. Since FAs determine where the cell adheres to the ECM, we see how ECM anisotropy influences cell morphology. In the isotropic ECM, where fibers are uniformly oriented, FAs form evenly around the cell, leading to uniform cell spreading over the fibers. On the anisotropic ECM, however, FAs preferentially form parallel to the fibers, causing the cell to elongate and align with the overall orientation of the fibers. Interestingly, this model also predicts a mechanical reciprocity between cell contractility and ECM anisotropy: the cell’s contractile forces reorient ECM fibers toward itself, enhancing the cell’s ability to adhere more strongly as it continues to spread over these newly aligned fibers. In this section, we discuss these observations and link them to existing experiments and models. We start with the similarities and differences of this model with earlier work that studies the mechanosensitivity of FAs on a homogeneous isotropic matrix.
The model proposed in this paper is not a strict improvement over the conceptually similar model proposed by Rens and Merks (2020), which uses a continuum approach to model a homogeneous isotropic ECM. Instead, the type of ECM considered in both models is different; for ECMs with small physical components, a continuous approach, as described by Rens and Merks (2020), could be better suited, whereas many other realistic ECMs require the added plastic, fibrous details that this paper develops. In the current paper, a cell situated on a fibrous and non-elastic ECM, such as collagen, is modeled, whereas Rens and Merks (2020) considered an elastic ECM, such as a polyacrylamide gel. Both models explain how cells spread less on soft substrates and more on stiffer substrates. However, in the model proposed by Rens and Merks (2020), the cell starts elongating on substrates of intermediate stiffness, whereas this spontaneous elongation is not observed in the currently discussed model. A possible reason for this discrepancy is the additional assumption made by Rens and Merks (2020), namely, that planar substrate stress strengthens FAs. To what extent FAs strengthen due to substrate stress in the context of a fibrous ECM remains unclear.
The model presented in this paper predicts a biphasic relationship between the extent of cell spreading and substrate stiffness: on an isotropic ECM, the cell spreads up to an optimum stiffness, whereas for even stiffer substrates, the cell area decreases again (Figure 2). Cell spreading on an isotropic regular ECM is best compared with in vitro spreading of cells on a hydrogel, as is found in the biphasic spreading of T cells (Oakes et al., 2018; Wahl et al., 2019). In the presented model, the biphasic relation arises from the catch-slip behavior of the integrins in the FAs, Equation 4, regulated by the slip parameter
After considering cell spreading on isotropic and homogeneous ECMs, we subsequently studied cell spreading on isotropic but inhomogeneous ECMs, which model certain types of in vitro fiber networks such as collagen or fibrin networks. The mechanics of fibrous networks are different from those of homogeneous ECMs because both respond differently to stress due to, for example, the possibility of sliding and reorientation of fibers (Storm et al., 2005). Our model predicts an increase in cell area with an increase in cross-link density, which aligns with findings that osteogenic cells are larger on highly cross-linked collagen fiber networks than on low cross-linked networks (Mullen et al., 2015). In a contrasting study, Baker et al. (2015) compared the cell spreading of NIH 3T3 fibroblasts and human MSC cells on synthetically produced fiber networks with low or high fiber stiffness. The cells spread more on softer fibers and less on stiffer fibers. Together with a computational follow-up study, a model emerged where cells on soft fibers could pull additional fibers toward the cell over which it could spread (Baker et al., 2015; Cao et al., 2017). To study this effect in our model, it can be extended to include two additional mechanisms that are included in the study by Cao et al. (2017). First, FAs should strengthen when the fiber density is higher. Second, cross-links break under stress, leading to higher fiber recruitment in soft networks.
We next studied the effect of anisotropy in the ECM by introducing a bias in the fiber orientation and found that the cell elongates in the direction of this bias and that FAs preferentially form at the poles of the cell. The model explains cell alignment to anisotropic collagen fiber networks (Lai et al., 2012; Chaubaroux et al., 2015; Sapudom et al., 2023). The model suggests that mechanosensitivity of the FAs is sufficient for the cell to sense the orientation of the fibers. This suggests that the stiffness-dependent maturation of FAs allows the cell to sense the orientation of the network. Others have suggested that a positive feedback loop between cell contractility and ECM stress drives cell elongation on anisotropic substrates, claiming that cells increase their contractile forces when sensing higher ECM stress (Alisafaei et al., 2022). This interesting explanation could be studied in further models. A conflicting observation, however, is that cells do not increase their contractile forces purely based on the stiffness of their environment (Feld et al., 2020).
After discussing the effect of pre-aligned ECM fibers on cell morphology, we studied the role of fiber orientation in mechanical cell–ECM reciprocity. Recently, it has been proposed that fiber reorientation by the cell is a two-way process: (1) cell protrusions adhere to and reorient fibers and (2) these fibers then develop anisotropic tension, which stabilizes the protrusions on aligned fibers, creating a self-reinforcing cycle (Alisafaei et al., 2022). Our model predicts a similar mechanism based on basic principles of mechanosensitive FA maturation. This mechanical reciprocity plays a role at larger multicellular scales. For example, in metastasis, the contractile forces of tumors align surrounding fibers (Balcioglu et al., 2016), and cancer cell migration is enhanced on aligned fibers (Sander, 2014; Doyle et al., 2022).
Our hybrid model predicts cell spreading, alignment, and ECM remodeling in terms of simple principles. However, we still could study to what extent it quantitatively matches experimental observations. One limitation to the model is its restriction to two dimensions. In 2D cell cultures, for example, cells extend protrusions beneath collagen fibers, wrapping around them (Friedrichs et al., 2007), a process not captured in two dimensions. Extensions capturing 3D fiber effects would require the use of a 3D or multi-layer CPM combined with additional cell-fiber behavior such as fiber repulsion. A second limitation to the ECM model is the lack of validation of its mechanical properties and network topology against real ECM structures, despite its demonstration of viscoelastic behavior (Tsingos et al., 2023).
Future work could incorporate more realistic ECM topologies, as modeled in previous studies by Davoodi Kermani et al. (2021) and Eichinger et al. (2021), which would significantly enhance the accuracy of this model. Possible other extensions include the study of cell migration along fibers by using one of the many active cell migration models implemented for the CPM such as a polarity vector (Beltman et al., 2007; Burger et al., 2022) or the Act model (Niculescu et al., 2015). An additional mechanism for FA breakdown is also needed because the current lifetimes of the FAs are unrealistically high and can even span nearly the whole simulated time (Supplementary Figure S9). The difficulty lies in the detachment of FAs at the rear of the migrating cell, which could be done, for example, by applying a model for asymmetric traction forces that would rupture the rear FAs or by introducing a chemical symmetry-breaking component (Yamaguchi and Knaut, 2022). To model cell migration in 3D, fiber exclusion should be added. In the current model, the cell interacts with the fibers only at adhesion sites, so the cell membrane could move through the fibers, which is not realistic. Another example of possible further study is that of multicellular mechanical interaction, which has been studied using CPM models (van Oers et al., 2014; Rens and Merks, 2017; Chiang and Chung, 2024). These studies applied a linear elastic continuous approach for modeling the ECM, whereas the realistic ECM has non-linear behavior such as strain stiffening. Such non-linear behavior is easily incorporated into the fibrous ECM model (Tsingos et al., 2023) and, when linked to our model of dynamic mechanosensitive FAs, could be used in modeling cell–cell mechanical communication. The present model could also be extended with the effect of proteolytic enzymes, such as MMPs, that would digest matrix fibers or cross-linkers, e.g., during angiogenesis, tissue remodeling, and tissue repair.
We demonstrated how the CPM can be used to study the mechanical reciprocity between a cell and a fibrous ECM, revealing that mechanosensitive adhesion explains the cell’s tendency to align with the dominant fiber orientation in the ECM, and their ability to reorient fibers to stabilize protrusions. This model can now be applied to systems in which these mechanisms are believed to play a key role.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
KK: conceptualization, methodology, software, visualization, writing–original draft, and writing–review and editing. ET: conceptualization, supervision, and writing–review and editing. RM: conceptualization, funding acquisition, supervision, and writing–review and editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by the NWO grant NWO/ENW-VICI 865.17.004 to RM (KK, ET, and RM) and by Prof. dr. Jan van der Hoevenstichting voor Theoretische Biologie (RM) affiliated to the Leiden University Fund (LUF). ET is currently funded by the Dutch Research Council (NWO) in the NWO Talent Programme with project number VI. Veni.222.323. The software was partly developed in context of the TissueOpt Small Scale Initiative grant NLESC.SSI.2021b.007 from the Netherlands eScience Center.
Acknowledgments
This work was performed using the compute resources from the Academic Leiden Interdisciplinary Cluster Environment (ALICE) provided by Leiden University. The authors thank Lourens Veen of the eScience Center for his contributions to software developed for this paper.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcell.2024.1462277/full#supplementary-material
References
Alisafaei, F., Shakiba, D., Iannucci, L. E., Davidson, M. D., Pryse, K. M., Chao, P.-h. G., et al. (2022). Tension anisotropy drives phenotypic transitions of cells via two-way cell-ECM feedback. doi:10.1101/2022.03.13.484154
Anderson, J. A., Glaser, J., and Glotzer, S. C. (2020). HOOMD-blue: a Python package for high-performance molecular dynamics and hard particle Monte Carlo simulations. Comput. Mater. Sci. 173, 109363. doi:10.1016/j.commatsci.2019.109363
Baker, B. M., Trappmann, B., Wang, W. Y., Sakar, M. S., Kim, I. L., Shenoy, V. B., et al. (2015). Cell-mediated fibre recruitment drives extracellular matrix mechanosensing in engineered fibrillar microenvironments. Nat. Mater. 14, 1262–1268. doi:10.1038/nmat4444
Balcioglu, H. E., Water, B. V. D., and Danen, E. H. J. (2016). Tumor-induced remote ECM network orientation steers angiogenesis. Sci. Rep. 6, 22580. doi:10.1038/srep22580
Beltman, J. B., Marée, A. F., Lynch, J. N., Miller, M. J., and De Boer, R. J. (2007). Lymph node topology dictates T cell migration behavior. J. Exp. Med. 204, 771–780. doi:10.1084/jem.20061278
Burger, G. A., Van De Water, B., Le Dévédec, S. E., and Beltman, J. B. (2022). Density-dependent migration characteristics of cancer cells driven by pseudopod interaction. Front. Cell. Dev. Biol. 10, 854721. doi:10.3389/fcell.2022.854721
Campbell, E. J., and Bagchi, P. (2021). A computational study of amoeboid motility in 3D: the role of extracellular matrix geometry, cell deformability, and cell–matrix adhesion. Biomechanics Model. Mechanobiol. 20, 167–191. doi:10.1007/s10237-020-01376-7
Cao, X., Ban, E., Baker, B. M., Lin, Y., Burdick, J. A., Chen, C. S., et al. (2017). Multiscale model predicts increasing focal adhesion size with decreasing stiffness in fibrous matrices. Proc. Natl. Acad. Sci. U. S. A., 114. E4549-E4555. doi:10.1073/pnas.1620486114
Casey, D. T., Bou Jawde, S., Herrmann, J., Mori, V., Mahoney, J. M., Suki, B., et al. (2021). Percolation of collagen stress in a random network model of the alveolar wall. Sci. Rep. 11, 16654. doi:10.1038/s41598-021-95911-w
Changede, R., and Sheetz, M. (2017). Integrin and cadherin clusters: a robust way to organize adhesions for cell mechanics. BioEssays 39, 1–12. doi:10.1002/bies.201600123
Chaubaroux, C., Perrin-Schmitt, F., Senger, B., Vidal, L., Voegel, J.-C., Schaaf, P., et al. (2015). Cell alignment driven by mechanically induced collagen fiber alignment in collagen/alginate coatings. Tissue Eng. Part C. Methods 21, 881–888. doi:10.1089/ten.tec.2014.0479
Chiang, H., and Chung, C.-A. (2024). Simulation of soluble and bound VEGF-stimulated in vitro capillary-like network formation on deformed substrate. PLOS Comput. Biol. 20, e1012281. doi:10.1371/journal.pcbi.1012281
Crossley, R. M., Johnson, S., Tsingos, E., Bell, Z., Berardi, M., Botticelli, M., et al. (2024). Modeling the extracellular matrix in cell migration and morphogenesis: a guide for the curious biologist. Front. Cell. Dev. Biol. 12, 1354132. doi:10.3389/fcell.2024.1354132
Daub, J. T., and Merks, R. M. H. (2013). A cell-based model of extracellular-matrix-guided endothelial cell migration during angiogenesis. Bull. Math. Biol. 75, 1377–1399. doi:10.1007/s11538-013-9826-5
Davoodi Kermani, I., Schmitter, M., Eichinger, J. F., Aydin, R. C., and Cyron, C. J. (2021). Computational study of the geometric properties governing the linear mechanical behavior of fiber networks. Comput. Mater. Sci. 199, 110711. doi:10.1016/j.commatsci.2021.110711
Diller, R. B., and Tabor, A. J. (2022). The role of the extracellular matrix (ecm) in wound healing: a review. Biomimetics 7, 87. doi:10.3390/biomimetics7030087
Doyle, A. D., Nazari, S. S., and Yamada, K. M. (2022). Cell–extracellular matrix dynamics. Phys. Biol. 19, 021002. doi:10.1088/1478-3975/ac4390
Eichinger, J. F., Grill, M. J., Kermani, I. D., Aydin, R. C., Wall, W. A., Humphrey, J. D., et al. (2021). A computational framework for modeling cell–matrix interactions in soft biological tissues. Biomechanics Model. Mechanobiol. 20, 1851–1870. doi:10.1007/s10237-021-01480-2
Feld, L., Kellerman, L., Mukherjee, A., Livne, A., Bouchbinder, E., and Wolfenson, H. (2020). Cellular contractile forces are nonmechanosensitive. Sci. Adv. 6, eaaz6997. doi:10.1126/sciadv.aaz6997
Feng, J., Levine, H., Mao, X., and Sander, L. M. (2019). Cell motility, contact guidance, and durotaxis. Soft Matter 15, 4856–4864. doi:10.1039/C8SM02564A
Friedrichs, J., Taubenberger, A., Franz, C. M., and Muller, D. J. (2007). Cellular remodelling of individual collagen fibrils visualized by time-lapse AFM. J. Mol. Biol. 372, 594–607. doi:10.1016/j.jmb.2007.06.078
Graner, F., and Glazier, J. A. (1992). Simulation of biological cell sorting using a two-dimensional extended Potts model. Phys. Rev. Lett. 69, 2013–2016. doi:10.1103/PhysRevLett.69.2013
Hirashima, T., Rens, E. G., and Merks, R. M. H. (2017). Cellular Potts modeling of complex multicellular behaviors in tissue morphogenesis. Dev. Growth and Differ. 59, 329–339. doi:10.1111/dgd.12358
Janmey, P. A., Fletcher, D. A., and Reinhart-King, C. A. (2020). Stiffness sensing by cells. Physiol. Rev. 100, 695–724. doi:10.1152/physrev.00013.2019
Kechagia, J. Z., Ivaska, J., and Roca-Cusachs, P. (2019). Integrins as biomechanical sensors of the microenvironment. Nat. Rev. Mol. Cell. Biol. 20, 457–473. doi:10.1038/s41580-019-0134-2
Kim, J., Feng, J., Jones, C. A. R., Mao, X., Sander, L. M., Levine, H., et al. (2017). Stress-induced plasticity of dynamic collagen networks. Nat. Commun. 8, 842. doi:10.1038/s41467-017-01011-7
Kim, M. C., Silberberg, Y. R., Abeyaratne, R., Kamm, R. D., and Asada, H. H. (2018). Computational modeling of three-dimensional ECM-rigidity sensing to guide directed cell migration. Proc. Natl. Acad. Sci. U. S. A., 115. E390-E399. doi:10.1073/pnas.1717230115
Kim, O. V., Litvinov, R. I., Alber, M. S., and Weisel, J. W. (2017b). Quantitative structural mechanobiology of platelet-driven blood clot contraction. Nat. Commun. 8, 1274. doi:10.1038/s41467-017-00885-x
Labernadie, A., and Trepat, X. (2018). Sticking, steering, squeezing and shearing: cell movements driven by heterotypic mechanical forces. Curr. Opin. Cell. Biol. 54, 57–65. doi:10.1016/j.ceb.2018.04.008
Lai, E. S., Huang, N. F., Cooke, J. P., and Fuller, G. G. (2012). Aligned nanofibrillar collagen regulates endothelial organization and migration. Regen. Med. 7, 649–661. doi:10.2217/rme.12.48
Lemmon, C. A., and Romer, L. H. (2010). A predictive model of cell traction forces based on cell geometry. Biophysical J. 99, L78–L80. doi:10.1016/j.bpj.2010.09.024
Malandrino, A., Trepat, X., Kamm, R. D., and Mak, M. (2019). Dynamic filopodial forces induce accumulation, damage, and plastic remodeling of 3D extracellular matrices. PLOS Comput. Biol. 15, e1006684. doi:10.1371/journal.pcbi.1006684
Maquart, F., and Monboisse, J. (2014). Extracellular matrix and wound healing. Pathol. Biol. 62, 91–95. doi:10.1016/j.patbio.2014.02.007
Mullen, C. A., Vaughan, T. J., Billiar, K. L., and McNamara, L. M. (2015). The effect of substrate stiffness, thickness, and cross-linking density on osteogenic cell behavior. Biophysical J. 108, 1604–1612. doi:10.1016/j.bpj.2015.02.022
Nahum, A., Koren, Y., Ergaz, B., Natan, S., Miller, G., Tamir, Y., et al. (2023). Inference of long-range cell-cell force transmission from ECM remodeling fluctuations. Commun. Biol. 6, 811. doi:10.1038/s42003-023-05179-1
Najafi, M., Farhood, B., and Mortezaee, K. (2019). Extracellular matrix (ECM) stiffness and degradation as cancer drivers. J. Cell. Biochem. 120, 2782–2790. doi:10.1002/jcb.27681
Niculescu, I., Textor, J., and De Boer, R. J. (2015). Crawling and gliding: a computational model for shape-driven cell migration. PLOS Comput. Biol. 11, e1004280. doi:10.1371/journal.pcbi.1004280
Novikova, E. A., and Storm, C. (2013). Contractile fibers and catch-bond clusters: a biological force sensor? Biophysical J. 105, 1336–1345. doi:10.1016/j.bpj.2013.07.039
Oakes, P. W., Bidone, T. C., Beckham, Y., Skeeters, A. V., Ramirez-San Juan, G. R., Winter, S. P., et al. (2018). Lamellipodium is a myosin-independent mechanosensor. Proc. Natl. Acad. Sci. 115, 2646–2651. doi:10.1073/pnas.1715869115
Paukner, D., Eichinger, J. F., and Cyron, C. J. (2023). What are the key mechanical mechanisms governing integrin-mediated cell migration in three-dimensional fiber networks? Biomechanics Model. Mechanobiol. 22, 1177–1192. doi:10.1007/s10237-023-01709-2
Reinhardt, J. W., Krakauer, D. A., and Gooch, K. J. (2013). Complex matrix remodeling and durotaxis can emerge from simple rules for cell-matrix interaction in agent-based models. J. Biomechanical Eng. 135, 071003. doi:10.1115/1.4024463
Reinhart-King, C. A., Dembo, M., and Hammer, D. A. (2005). The dynamics and mechanics of endothelial cell spreading. Biophysical J. 89, 676–689. doi:10.1529/biophysj.104.054320
Rens, E. G., and Merks, R. M. H. (2017). Cell contractility facilitates alignment of cells and tissues to static uniaxial stretch. Biophysical J. 112, 755–766. doi:10.1016/j.bpj.2016.12.012
Rens, E. G., and Merks, R. M. H. (2020). Cell shape and durotaxis explained from cell-extracellular matrix forces and focal adhesion dynamics. iScience 23, 101488. doi:10.1016/j.isci.2020.101488
Sander, L. M. (2014). Modeling contact guidance and invasion by cancer cells. Cancer Res. 74, 4588–4596. doi:10.1158/0008-5472.CAN-13-3294
Sapudom, J., Karaman, S., Quartey, B. C., Mohamed, W. K. E., Mahtani, N., Garcia-Sabaté, A., et al. (2023). Collagen fibril orientation instructs fibroblast differentiation via cell contractility. Adv. Sci. 10, 2301353. doi:10.1002/advs.202301353
Schwarz, U. S., Erdmann, T., and Bischofs, I. B. (2006). Focal adhesions as mechanosensors: the two-spring model. Biosystems 83, 225–232. doi:10.1016/j.biosystems.2005.05.019
Storm, C., Pastore, J. J., MacKintosh, F. C., Lubensky, T. C., and Janmey, P. A. (2005). Nonlinear elasticity in biological gels. Nature 435, 191–194. doi:10.1038/nature03521
Stupack, D. G., and Cheresh, D. A. (2002). ECM remodeling regulates angiogenesis: endothelial integrins look for new ligands. Science’s STKE 2002, pe7. doi:10.1126/stke.2002.119.pe7
Theocharis, A. D., Skandalis, S. S., Gialeli, C., and Karamanos, N. K. (2016). Extracellular matrix structure. Adv. Drug Deliv. Rev. 97, 4–27. doi:10.1016/j.addr.2015.11.001
Tsingos, E., Bakker, B. H., Keijzer, K. A. E., Hupkes, H. J., and Merks, R. M. H. (2023). Hybrid cellular Potts and bead-spring modeling of cells in fibrous extracellular matrix. Biophysical J., 2609–2622. doi:10.1016/j.bpj.2023.05.013
van Oers, R. F. M., Rens, E. G., LaValley, D. J., Reinhart-King, C. A., and Merks, R. M. H. (2014). Mechanical cell-matrix feedback explains pairwise and collective endothelial cell behavior in vitro. PLoS Comput. Biol. 10, e1003774. doi:10.1371/journal.pcbi.1003774
Vargas, D. A., Gonçalves, I. G., Heck, T., Smeets, B., Lafuente-Gracia, L., Ramon, H., et al. (2020). Modeling of mechanosensing mechanisms reveals distinct cell migration modes to emerge from combinations of substrate stiffness and adhesion receptor–ligand affinity. Front. Bioeng. Biotechnol. 8, 459. doi:10.3389/fbioe.2020.00459
Wahl, A., Dinet, C., Dillard, P., Nassereddine, A., Puech, P.-H., Limozin, L., et al. (2019). Biphasic mechanosensitivity of T cell receptor-mediated spreading of lymphocytes. Proc. Natl. Acad. Sci. 116, 5908–5913. doi:10.1073/pnas.1811516116
Wakatsuki, T., Kolodney, M. S., Zahalak, G. I., and Elson, E. L. (2000). Cell mechanics studied by a reconstituted model tissue. Biophysical J. 79, 2353–2368. doi:10.1016/S0006-3495(00)76481-2
Wakatsuki, T., Wysolmerski, R. B., and Elson, E. L. (2003). Mechanics of cell spreading: role of myosin II. J. Cell. Sci. 116, 1617–1625. doi:10.1242/jcs.00340
Winkler, J., Abisoye-Ogunniyan, A., Metcalf, K. J., and Werb, Z. (2020). Concepts of extracellular matrix remodelling in tumour progression and metastasis. Nat. Commun. 11, 5120. doi:10.1038/s41467-020-18794-x
Yamaguchi, H., Wyckoff, J., and Condeelis, J. (2005). Cell migration in tumors. Curr. Opin. Cell. Biol. 17, 559–564. doi:10.1016/j.ceb.2005.08.002
Yamaguchi, N., and Knaut, H. (2022). Focal adhesion-mediated cell anchoring and migration: from in vitro to in vivo. Development 149, dev200647. doi:10.1242/dev.200647
Keywords: anisotropic extracellular matrix, focal adhesions, cellular Potts model, cell spreading, cell morphology, mechanical reciprocity, mathematical biology
Citation: Keijzer KAE, Tsingos E and Merks RMH (2025) How cells align to structured collagen fibrils: a hybrid cellular Potts and molecular dynamics model with dynamic mechanosensitive focal adhesions. Front. Cell Dev. Biol. 12:1462277. doi: 10.3389/fcell.2024.1462277
Received: 09 July 2024; Accepted: 04 December 2024;
Published: 06 January 2025.
Edited by:
Karin Pfisterer, Medical University of Vienna, AustriaReviewed by:
Céline Labouesse, ETH Zürich, SwitzerlandMaria Jose Gomez-Benito, University of Zaragoza, Spain
Copyright © 2025 Keijzer, Tsingos and Merks. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Roeland M. H. Merks, bWVya3NybWhAbWF0aC5sZWlkZW51bml2Lm5s
†Present address: Erika Tsingos, Theoretical Biology, Utrecht University, Utrecht, Netherlands