
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Bioeng. Biotechnol., 13 October 2022
Sec. Tissue Engineering and Regenerative Medicine
Volume 10 - 2022 | https://doi.org/10.3389/fbioe.2022.958669
This article is part of the Research TopicPerinatal Derivatives and the Road to Clinical Translation, Volume IIView all 14 articles
 Antonietta R. Silini1†
Antonietta R. Silini1† Taja Železnik Ramuta2†
Taja Železnik Ramuta2† Ana Salomé Pires3,4,5
Ana Salomé Pires3,4,5 Asmita Banerjee6
Asmita Banerjee6 Marie Dubus7
Marie Dubus7 Florelle Gindraux8
Florelle Gindraux8 Halima Kerdjoudj7
Halima Kerdjoudj7 Justinas Maciulatis9
Justinas Maciulatis9 Adelheid Weidinger6
Adelheid Weidinger6 Susanne Wolbank6
Susanne Wolbank6 Günther Eissner10
Günther Eissner10 Bernd Giebel11
Bernd Giebel11 Michela Pozzobon12
Michela Pozzobon12 Ornella Parolini13,14
Ornella Parolini13,14 Mateja Erdani Kreft2*
Mateja Erdani Kreft2*Perinatal derivatives or PnDs refer to tissues, cells and secretomes from perinatal, or birth-associated tissues. In the past 2 decades PnDs have been highly investigated for their multimodal mechanisms of action that have been exploited in various disease settings, including in different cancers and infections. Indeed, there is growing evidence that PnDs possess anticancer and antimicrobial activities, but an urgent issue that needs to be addressed is the reproducible evaluation of efficacy, both in vitro and in vivo. Herein we present the most commonly used functional assays for the assessment of antitumor and antimicrobial properties of PnDs, and we discuss their advantages and disadvantages in assessing the functionality. This review is part of a quadrinomial series on functional assays for the validation of PnDs spanning biological functions such as immunomodulation, anticancer and antimicrobial, wound healing, and regeneration.
Perinatal derivatives or PnDs (that include tissues and cells and secretomes from perinatal, or birth-associated tissues), and especially mesenchymal stromal cells (MSC) from perinatal tissues, have been mostly exploited for their applications in regenerative medicine, substantiated by their ability to modulate immune responses and/or to act in a pro-regenerative manner by promoting progenitor/stem cell differentiation (Silini et al., 2017; Silini et al., 2019). What has been largely unexplored is the applications of PnDs in other sectors, such as in oncology.
As a matter of fact, PnDs have only recently been investigated in vitro and in vivo for their potential as antitumor therapies. In this context, PnDs have been shown to act multimodally through complex mechanisms of action (MoA), and to target various hallmarks of cancer that capture essential features of tumorigenesis, such as sustaining proliferative signaling, evading growth suppressors, resisting cell death, enabling replicative immortality, inducing angiogenesis, activating invasion and metastasis, reprogramming cellular metabolism, avoiding immune destruction, tumor-promoting inflammation, genome instability, and deregulating cellular metabolism (Hanahan and Weinberg 2011). Treatments that shift the focus to the systemic level and that target the hallmarks of cancer are urgently needed and highly relevant.
In the context of cancer, PnDs have been reported to play opposing roles. Herein we will discuss these studies along with the in vitro and in vivo assays implemented to obtain a clearer understanding of the potential applications of PnDs as an antitumor treatment.
In addition to the potential applications of PnDs in oncology, several PnDs have been shown to possess antimicrobial properties (King et al., 2007a; Klaffenbach et al., 2011; Ramuta et al., 2021a), which is highly relevant in the field of oncology. As a matter of fact, in the last decade, growing evidence has emerged demonstrating that microbiota and microbial pathogens have immense effect on cancer development and treatment (Bhatt et al., 2017; Raza et al., 2019; Jain et al., 2021). While in 15–20% of cancer cases microbial pathogens drive tumorigenesis, even a larger part of malignancies is associated with the altered composition of commensal microbiota (Bhatt et al., 2017). Furthermore, bacteria are also able to affect the efficacy of chemotherapeutic drugs, either by inhibiting or enhancing their effect (Lehouritis et al., 2015; Roy and Trinchieri 2017). Namely, the microbiota and pathogens may affect the pharmacokinetics, antitumor activity and drug toxicity of chemotherapeutic agents by 1) changing the chemical structure of the drug (biotransformation) (Haiser and Turnbaugh 2012; Wilson and Nicholson 2017), 2) decreasing the absorption of certain drugs (Carmody and Turnbaugh 2014), or even indirectly by 3) affecting the host’s gene expression and physiology (e.g., of the local mucosal barrier), which results in altered metabolism of drugs (Björkholm et al., 2009; Selwyn et al., 2015; Selwyn et al., 2016; Roy and Trinchieri 2017).
Thus, herein both the anticancer and antimicrobial properties will be discussed alongside one another to provide a better understanding of the potential applications of PnDs as therapeutic strategies in these fields. We briefly summarize the results obtained using PnDs and then focus on the most frequently used functional assays for analyzing the antitumor and antimicrobial effects of PnDs in order to potentially provide insight into the future development of new functional and/or potency assays.
This review is part of a series of contributions from the COST Action (CA17116) entitled “International Network for Translating Research on Perinatal Derivatives into Therapeutic Approaches-SPRINT”. This Action is broadly aimed at establishing consensus for different aspects of PnDs research. The aim of this review is to provide inputs for the development of functional and potency assays that can be used to test PnDs before their oncological and antimicrobial application.
In this section we summarize the most frequently used in vitro and in vivo assays for analyzing the effects of PnDs on several hallmarks of cancer such as cell proliferation and metabolism, cell death and apoptosis, cell migration and invasion, angiogenesis, and metastasis.
One of the most widely analyzed hallmark of tumor cells is proliferation. Indeed, in vitro PnDs have been shown to mostly exert antiproliferative effects on tumor cells (Gauthaman et al., 2012; Magatti et al., 2012; Liu et al., 2013; Hendijani et al., 2015; Kalamegam et al., 2019; Ramuta et al., 2020a), but it has also been shown that they may induce tumor cell proliferation (Kim et al., 2015; Li et al., 2015).
Several assays are used to assess the effect of PnDs on tumor cell proliferation. The investigation of DNA synthesis analyzes the effects of PnDs on tumor cell proliferation. Incorporation of radiolabeled DNA precursor 3H-thymidine (Ayuzawa et al., 2009; Magatti et al., 2012; Marleau et al., 2012; Yuan et al., 2013; Riedel et al., 2019; Ramuta et al., 2020b) into new strands of chromosomal DNA or more recent analogs, BrdU (bromoeoxyuridin) (Tian et al., 2010; Gauthaman et al., 2012; Han et al., 2014; Hendijani et al., 2015) and EdU (5-ethynyl-2 deoxyuridine) (Wang M et al., 2015; Janev et al., 2021), have been widely used to assess de novo DNA synthesis. 3H-thymidine detection requires radioactive labeling followed by detection with a scintillation beta-counter, while BrdU can be easily detected by antibodies followed by flow cytometry or by immunohistochemistry. BrdU is mutagenic and can alter the cell cycle, thus adequate controls should be implemented. In contrast to BrdU, the newer analog EdU does not require DNA denaturation by exposing cells to HCl, heat or DNAse, and its detection is rapid and highly sensitive. BrdU and EdU are considered reliable assays for a direct index of proliferation. Other analyses implemented for the analysis of tumor cell proliferation are Ki67 cell immunolabelling (Lin et al., 2014; Riedel et al., 2019), and other assays aimed at investigating genes or proteins involved in cell cycle progression, such as cyclins (Magatti, De Munari et al., 2012; Riedel, Pérez-Pérez et al., 2019).
The colony forming unit (CFU) assay analyzes the consequences of external stress signals on the cell’s ability to proliferate and form a colony. This assay has been widely used to test the effect of PnDs (Ayuzawa et al., 2009; Liu et al., 2013; Ciavarella et al., 2015; Li et al., 2015; Wang M et al., 2015; Wang W et al., 2015) and is especially useful to assess long-term effects. This method is however time consuming with extended incubation times, plus colony formation ability differs between cells. The traditional assay requires weeks for completion and usually does not allow cell retrieval; however, this can be circumvented by the use of fluorescent dyes allowing quantitative, high throughput colony counting and by the use of specialized agars that allow cell suspension and growth. The assay can also be used to predict tumorigenicity in vivo.
In vivo objective assessment of tumor growth is a crucial tool for the advancement of cancer therapies. In vivo, PnDs have been shown to both inhibit (Ayuzawa et al., 2009; Wu et al., 2013) and induce (Yang C. et al., 2014; Wang M et al., 2015; Svitina et al., 2018) tumor growth. These contradictory results could be due to several variations, either given by the PnDs used (e.g., tissues, cells, secretomes, homogenates), by the tumor model (e.g., tumor cells, rodent model and strain), by the different treatment regimens (e.g., administration route, dosage), or by the method used to monitor tumor progression (e.g., caliper measurements, imaging).
Monitoring tumor progression after PnDs treatment has been performed through the measurement of tumor diameters (Du et al., 2014; Ma et al., 2015; Bu et al., 2017; Yuan L et al., 2019) (or volumes deduced from diameters) or tumor weight, but also through more sophisticated in vivo optical imaging techniques (Di et al., 2014; Leng et al., 2014; Ciavarella et al., 2015; Cafforio et al., 2017; Zhang et al., 2017; Yuan Z et al., 2019; Chetty et al., 2020), which include fluorescence and bioluminescence studies. Although traditional caliper measurement is a simple and low-cost method, its major disadvantages include variability of tumor size measurements and tumoral heterogeneity. In contrast, optical imaging is a more accurate, sensitive and specific technique for tumor imaging, allowing the detection of microscopic tumors. The main limitation of optical imaging is the need of tumor cells to express a reporter gene (Puaux et al., 2011). Cell proliferation is often used as a measure of tumor response, with immunohistochemistry being the most widely used in vivo technique.
Altered cell metabolism is another feature of tumor cells, and proliferating tumor cells hijack their metabolism to fuel continuous growth. Cell metabolism is commonly measured through the reduction of substrates to a final product by intracellular enzyme activity in living cells can be assessed by colorimetric assays. The degree of color change is not directly proportional to the number of viable cells, but rather to enzyme activity. This method permits only a moderately robust measurement of viability; however, the ease of use and potential for high throughput analysis in multiwell plates has made it very popular. The most frequently used tetrazolium compound (MTT) is reduced to formazan (Li et al., 2011; Chao et al., 2012; Gauthaman et al., 2012; Kang et al., 2012; Liu et al., 2013; Lin et al., 2014; Niknejad et al., 2014; Rolfo et al., 2014; Bonomi et al., 2015; Lang et al., 2015; Li et al., 2015; Kamalabadi-Farahani et al., 2018; Mandal et al., 2019; Riedel et al., 2019). A less toxic alternative, Cell Counting Kit-8 (CCK-8) (Wu et al., 2013; Yan et al., 2013; Yang C. et al., 2014; Kim et al., 2015; Di Germanio et al., 2016), has also been used, with a detection sensitivity higher than tetrazolium salts such as MTT or MTS(Wang W et al., 2015). Tumor cell oxidative stress, using superoxide dismutase, intracellular accumulation of reactive oxygen species (ROS), glutathione peroxidase, hydrogen peroxide and lipid peroxidation assays have also been investigated after PnDs treatment (Lin et al., 2014).
Similar to their effects on tumor cell proliferation PnDs have been shown to have dual effects on tumor cell death and apoptosis, by either promoting (Chen et al., 2012; Jiao et al., 2012; Del Fattore et al., 2015; Kalamegam et al., 2018) or inhibiting (Niknejad et al., 2014) these processes.
There are several assays used to assess the effect of PnDs on tumor cell death. For example, annexin V/PI assay and flow cytometry are the most popular approaches for detection of apoptosis in tumor cells after PnDs treatment (Gauthaman et al., 2012; Wu et al., 2013; Yang X. et al., 2014; Niknejad et al., 2014; Mamede et al., 2015; Lin et al., 2016; Paris et al., 2016; Shen et al., 2016; Lin H et al., 2017; Chai et al., 2018; Jiao et al., 2018; Yuan et al., 2018; Khalil et al., 2019; Rezaei-Tazangi et al., 2020; Silva et al., 2020). The TUNEL assay (Wu et al., 2013; Niknejad et al., 2014) detects DNA fragmentation in apoptotic cells in vitro. After staining, cells can be analyzed by light or fluorescent microscopy. TUNEL staining is fast, accurate and sensitive but fails in discriminating the different types of cell death. The assay can also be used to assess the effect of PnDs on tumor cell death ex vivo. Detection of apoptosis regulators, such as caspases (Wu et al., 2013; Yang C. et al., 2014; Niknejad et al., 2014; Mamede et al., 2015; Lin et al., 2016; Kalamegam et al., 2018; Mandal et al., 2019; Rezaei-Tazangi et al., 2020), cytochrome c, Bcl-2, Bax, Fas, FasL, Danger Associated Molecular Proteins (DAMPs), CRT, Hsp90 and Hsp70, etc. Can be detected at the protein and/or mRNA level using flow cytometry, multi-detection plate reader (Mamede et al., 2015), immunoblot (Dzobo et al., 2016; Shen et al., 2016; Yuan et al., 2018), immunohistochemistry, immunofluorescence (Dzobo et al., 2016; Lin H et al., 2017), and RT-PCR (Yang X. et al., 2014). As further progress is made in understanding the mechanisms of cell death, more accurate and precise interpretations of the results of these tests will be possible.
Cell death induced by PnDs in animal models has been mainly analyzed by in situ detection of apoptosis as mentioned previously regarding the in vitro assays (Dong et al., 2018; Kamalabadi-Farahani et al., 2018; Chen et al., 2019; Fan et al., 2020). An alternative approach is the in vivo imaging of apoptosis, using radiolabeled forms of annexin V for positron emission tomography (PET) and single photon emission computed tomography (SPECT) (Iravani and Hicks 2020).
Mitochondria play a key role in response to cellular stress and injury. For this reason, the evaluation of mitochondrial membrane potential (MMP) (Mamede et al., 2015; Lin et al., 2016) could also be used as the marker of cell death. However, the interpretation should be carefully and critically performed. Measurement of mitochondrial activity could not only serve as an alternative to cell viability assays, but also provide distinct information on the metabolic state, and therefore the quality of a cell. For the application of therapeutic cells, knowledge on the mode of energy production can be important. Mitochondrial activity can be determined by quantification of the activity of single complexes such as complex I by measuring the light absorbance of nicotinamide adenine dinucleotide (NADH), the electron donor for complex I, at 340 nm (Nelson and Cox 2004). The downside of this method is that it does not provide any information on the coupling state of the electron transfer system, and therefore, also not on the production level of adenosine triphosphate (ATP). Activity of the entire electron transfer system can be determined, for example, with a Clark electrode-based measurement (Hütter et al., 2006). With this method, the oxygen concentration of a solution is measured as oxygen is reduced at the cathode. The resulting current is directly proportional to the oxygen concentration of the solution (Gnaiger 2008). The advantage of this method is that distinct respiration states can be determined (Gnaiger 2008). By addition of specific substrates and inhibitors, total oxygen consumption (routine respiration) can be distinguished from oxygen consumption with ATP production (oxidative phosphorylation) and oxygen consumption without ATP production (LEAK) (Gnaiger 2008). With this method, mitochondrial respiration can be measured in tissue, tissue homogenate, cells and isolated mitochondria. To our knowledge, there are no studies that have evaluated the effects of PnD on tumor MMP, however, such measurements have successfully been performed in human amniotic membrane tissue (Banerjee et al., 2015; Poženel et al., 2019), isolated human amniotic membrane epithelial cells (Banerjee et al., 2018a), and human amniotic membrane derived MSC (Banerjee et al., 2018a; Banerjee et al., 2018b). In addition to the mitochondrial respiration assay, mitochondrial status can also be monitored with the membrane permeable dye JC-1. JC-1 is a fluorescent cationic carbocyanine dye that exhibits potential-dependent accumulation in mitochondria, forming J-aggregates and diffuses across mitochondria upon depolarisation to form a monomeric state (Sivandzade et al., 2019). Currently, to our knowledge, there is only one study that has investigated the function of amniotic membrane proteins (AMPs) extracted from hAM against hypoxia-induced H9c2 cardiomyoblast cells (Faridvand et al., 2019). AMPs have potent cardioprotective effects in H9c2 cells by inhibiting the Ca2+ overload and the mitochondrial membrane potential dysfunction during hypoxia (Faridvand et al., 2019). Furthermore, there are few studies that evaluated mitochondrial membrane potential in placental trophoblast cells from patients with preeclampsia (Zhang et al., 2022). The anticancer effect of PnDs have not yet been evaluated with the mitochondrial dye JC-1.
In this section, we presented an overview of the most common functional assays that have been used for evaluating the pro-cell death/proapoptotic activity of PnDs. Each assay has its advantages and an understanding of strengths and limitations can allow for the selection of the optimal assay based on a specific need. No matter how appropriate and well accepted the assay is, it is recommended that a second assay using a different principle should be used to confirm the detection of cell-death. In the future, we should also have in mind that intact efferocytosis, i.e. the clearance of apoptotic cells, can promote cancer disease (Vaught et al., 2015). It is therefore important that both processes, apoptosis and efferocytosis are fine-tuned in specific tumor microenvironments. Hence, the novel functional assays for analyzing the influence of PnD on efferocytosis-mediated regulation of the tumor microenvironment needs to be developed/used.
PnDs have been reported to both inhibit (Fong et al., 2011; Gauthaman et al., 2012; Kalamegam et al., 2018; Li et al., 2019) and induce (Kim et al., 2015; Li et al., 2015) tumor cell migration.
The spread of neoplastic disease has been described as a sequential multi-step process, termed the invasion-metastatic cascade (Martin and Jiang 2009). During migration and invasion, cells squeeze through tight interstitial spaces, which includes cellular and nuclear deformation caused by the confining microenvironment (Kramer et al., 2013). Overall, during the metastatic cascade, changes in cell-cell and cell-matrix adhesion are of paramount importance and lead to the formation of secondary tumors in distant organs and are largely responsible for the mortality and morbidity of cancer (Martin et al., 2013). There are several commonly used in vitro assays to investigate the effects of PnDs on tumor cell migration and invasion potential. Transwell migration and invasion assay (Boyden chamber) is the most frequently used approach. In this assay, a double chamber is filled with two media, one with an attractant (like FBS) to trigger chemotaxis. Cells are seeded in the upper well and migrate vertically between the chambers through a porous membrane (Menyhárt et al., 2016). Migrated cells can be visualized by cytological dyes or stained fluorescent and then assessed by flow cytometry, light or fluorescence microscopy, or lysed and assessed by a plate reader, usually following treatment with MTT reagent (Gauthaman et al., 2012; Kim et al., 2015; Li et al., 2015; Kalamegam et al., 2018; Yuan et al., 2018; Mandal et al., 2019; Silva et al., 2020). By coating the porous filter with ECM components like type I collagen or a basement membrane-like matrix (Matrigel (So et al., 2015; Bu et al., 2017; Meng et al., 2019)) or reduced growth factor matrix (Touboul et al., 2013), invasive cells can be detected by their ability to degrade the matrix and move through the membrane to the bottom well. Parallel measurements with ECM-coated and non-coated assays allow one to calculate an “invasive index”: the rate of invasiveness versus migration (Marshall 2011), however this approach is currently missing in vitro assays investigating migration and invasion potential of PnDs. The most frequently used method due to its easy setup is the transwell migration assay. More sophisticated migration assays using microfluidic migration devices overcome the limitations of traditional migration assays and promote a stable diffusion-generated concentration gradient that is consistently linear and lasts for more than 48 h. These devices are usually plastic with high optical qualities similar to those of glass, and are specially designed for video microscopy assays. At specific time intervals, images of the observation area can be acquired, allowing real-time monitoring and quantitative measurements of cell migration and thus could also be used in investigating PnDs actions on cancer cell migration and invasion.
Investigation into the effect of PnDs on the invasive potential of cancer cells has been also performed using intact or decellularized human amniotic membrane or amniochorionic membrane. These were used in some studies as a natural 3D scaffold to evaluate tumor cell metastatic and invasion potential in 3D conditions (Ganjibakhsh et al., 2019) or direct influence of PnDs on metastatic and invasion behavior of tumor cells (Touboul et al., 2013; Ramuta T. Z. et al., 2020).
PnDs have been widely reported to produce angiogenic factors and induce angiogenesis (Bajetto et al., 2017; Dabrowski et al., 2017; Komaki et al., 2017; Wu et al., 2022), yet some studies have described antiangiogenic effects (Faraj, Stewart et al., 2015), and PnD preparation seems to be a critical point that can influence this feature (Wolbank et al., 2009).
Quantitative real time polymerase chain reaction is often performed to detect and quantify the relative expression levels of angiogenic genes, such as VEGF (Lin D et al., 2017; Mandal et al., 2019), ANG (Mandal et al., 2019; Yuan L et al., 2019), PDGF (Yuan Z et al., 2019), FGF-2 (Subramanian et al., 2012), etc. However, this method is time and resource consuming, requires subsequent post-PCR analysis and may provide only limited information on gene expression that must be followed with immunoblot or ELISA. Indeed, the latter have been widely used to detect known angiogenesis activators by immunoblot or ELISA (Table 1). However, Western blot can produce false-positive/negative results in the sample of interest and requires a larger amount of starting material. Immunophenotypic analysis to measure the expression of major angiogenic proteins, such as VEGF (Borghesi et al., 2020) can be performed with flow cytometry. Despite its ability to identify small populations and quantify the intensity of fluorescence, it still requires complex instrumentation, and highly trained technical staff to manage microfluidics, laser calibration and cleaning, as well as ample experience with the relevant software. In addition, direct and indirect immunocytochemistry (ICCH) can visualize and localize the target VEGF protein expression at a cell compartment level (Subramanian et al., 2012). Indirect ICCH can, however, be more laborious and time-consuming, with the additional risk of non-specific binding of the secondary antibody.
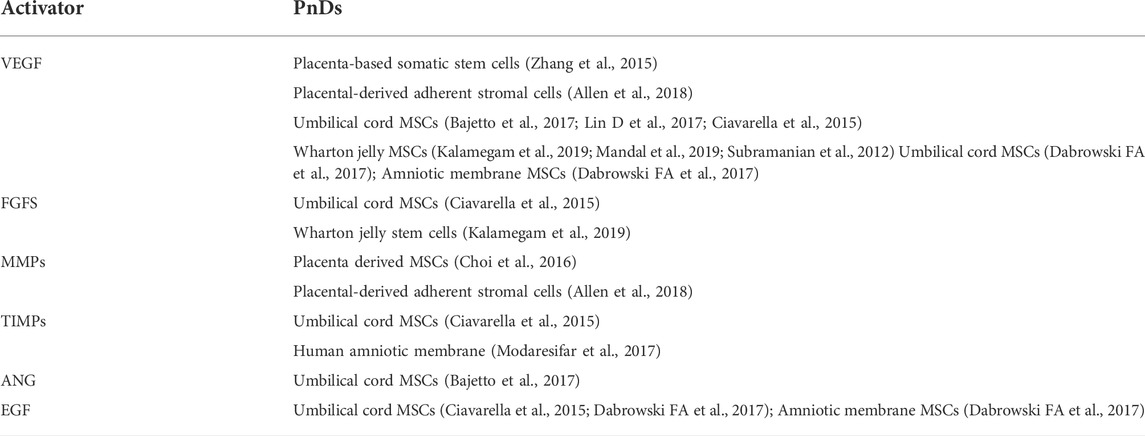
TABLE 1. Angiogenesis activators produced by PnDs.
Monitoring the proliferation of human umbilical vein endothelial cells (HUVECs) is often used to evaluate the developing tumor vasculature. These assays have been used to estimate antiangiogenic properties of PnDs in the context of early tumor pre-vasculature. CCK8 (Yuan L et al., 2019) assay offer reproducible and easy setups and provide quantifiable data on the inhibition of endothelial cell proliferation. Alternative methods, such as Trypan blue analysis (Chen et al., 2012) might be more difficult to reproduce and validate. Monitoring of electrical impedance changes (Grzywocz et al., 2018) caused by the proliferation of HUVECs could be enabled by real-time systems. In addition, the digital endothelial tube formation assay-derived images could potentially enable the calculation of digital angiogenic indices (Alshareeda et al., 2018), covering the numerical values of tube morphometry, such as extremities, number of segments, branches and the length of tubes.
To evaluate angiogenesis in vitro, the rat aorta ring assay (Modaresifar et al., 2017) can be performed with PnDs. In this assay, aorta rings are dissected from the descending thoracic aorta, rinsed, and cut into circular sections of several millimeters thick. These sections are put on cultured cells and the angiogenic potential is determined by microscopically visualizing endothelial cell sprouting, polarization, and outgrowth to the periphery. This test enables evaluation of angiogenic and antiangiogenic effects of PnDs and is more representative of in vivo angiogenesis than two dimensional assays.
One of the most intriguing characteristics of PnDs is their direct and indirect antimicrobial properties, which have therapeutic potential. PnDs have been shown to possess antimicrobial activity against various microorganisms in planktonic form and also in complex microenvironments. While the precise nature of antimicrobial action of PnDs is not well understood, it is clear that more than one mechanism working simultaneously to inhibit microbe growth and endotoxin activity, contributes to this activity (Magatti, Vertua et al., 2017). Antimicrobial properties of PnDs have been investigated in bacteria (Talmi et al., 1991; Mao et al., 2016; Mao et al., 2017; Tehrani et al., 2017; Mao et al., 2018; Ashraf et al., 2019; Palanker et al., 2019; Šket et al., 2019; Ramuta et al., 2020a), fungi (Wang, Xie et al., 2012), bacteria-infected cell cultures (Ramuta et al., 2021b) and rat in vivo models (Robson and Krizek 1973; Yadav et al., 2017). Antimicrobial peptides, such as α and β defensins, human cathelicidin LL37, lipocalin, elafin and secretory leukocyte protease inhibitor (SLPI), have been identified in various PnDs (King et al., 2007a; King et al., 2007b; Ramuta T.Ž. et al., 2021; Dubus et al., 2022). Furthermore, it was shown that histones H2A and H2B could also exert an antimicrobial action as a endotoxin-neutralizing barrier (Kim et al., 2002). Moreover, it was reported that hemoglobin-derived peptides purified from a human placenta exhibited antimicrobial activity. These peptides inhibited the growth of Gram-positive and Gram-negative bacteria and yeasts in micromolar concentrations, as well they reduced endotoxin activity by binding to LPS (Liepke et al., 2003; Dubus et al., 2022). In case of decellularized Wharton’s jelly tissue, the mass spectrometry analysis showed the release of antimicrobial molecules involved in the innate immune response but also some molecules involved in bacterial agglutination such as fibrinogen beta chain and Fibulin 1 (Dubus et al., 2022). These molecules are thought to exert a bacteriostatic effect on both Gram-positive and Gram-negative strain. In this section, we offer an overview of the most common in vitro and in vivo functional assays that have been used for evaluating the antimicrobial activity of PnDs against bacteria and fungi.
The gold standards for antimicrobial susceptibility testing have been set by the Clinical and Laboratory Standards Institute, however, due to the versatility of PnDs, the following assays do not strictly follow the standard protocols, as they had to be adapted to enable the analysis of various PnDs-derived preparations in bacterial suspensions.
A broth (micro)dilution assay is a simple and inexpensive method which is consequently often used to test the susceptibility of bacterial isolates to various antimicrobials. PnDs (often at various dilutions) are added to liquid broth media, which are then inoculated with bacterial suspensions. Following incubation (the length of which can vary from a couple of hours to several days), bacterial growth is evaluated based on turbidity by using visual or spectrophotometric methods (Jorgensen and Ferraro 2009; Sung et al., 2016; Šket et al., 2019; Dubus et al., 2020; El-Mahdy et al., 2021). Furthermore, bacterial growth can be also quantified by plating serial dilutions of bacterial suspensions incubated with the antimicrobial agent and counting the colony forming units (CFU) (Thadepalli et al., 1977; Mao et al., 2017; Šket et al., 2019). The broth microdilution method is often used to determine the minimum inhibitory concentration (Wiegand et al., 2008; Kim et al., 2012; Yadav et al., 2017) of an antimicrobial agent, which is the lowest concentration that will inhibit the visible growth of a microorganism after overnight incubation. The advantages of using the broth (micro)dilution test are the generation of a quantitative result and high reproducibility, while the main shortcoming is that it is less sensitive and more time-consuming (Jorgensen and Ferraro 2009) than some of the other functional assays for determination of the antimicrobial effects of PnDs. Furthermore, the presence of dead bacteria in the presence of PnDs cannot be determined.
A similar method is a disk diffusion assay. Namely, a bacterial inoculum is plated onto the surface of the agar plate and subsequently PnDs (often at various dilutions) are applied to the inoculated agar surface. After incubation of the agar plates the inhibition zones around the site of application of the PnDs antimicrobials are measured (Talmi et al., 1991; Kjaergaard et al., 2001; Jorgensen and Ferraro 2009; Tehrani et al., 2013; Tehrani et al., 2017; Šket et al., 2019; Ramuta et al., 2020b; Ramuta T.Ž. et al., 2021). The shortcoming of this method is its inability to precisely determine the minimum inhibitory concentration of the antimicrobial agent.
The antimicrobial properties of PnDs have been evaluated in complex (micro)environments, such as biofilms and bacteria-infected epithelia. The effect of PnDs on biofilms has been evaluated by the biofilm formation assay (Dubus et al., 2020; El-Mahdy et al., 2021). The antiadhesive and antifouling properties of PnDs such as Wharton’s jelly were mainly attributed the presence of hyaluronic acid and its composites (Drago et al., 2014; Marcuzzo et al., 2017). Biofilm is defined as a bacterial community which is metabolically heterogeneous and embedded in a self-produced extracellular matrix, causing a critical virulence factor responsible for treatment failure and chronicity in medical device-related infections. The (micro)plates are inoculated with fresh bacterial cell suspensions and subsequently the PnDs are added. The biofilms are grown for several hours to days. After staining with crystal violet, the effect of the PnDs on biofilm is quantified spectrophotometrically. This is an inexpensive and easy method which can directly evaluate the biofilm formation on several surfaces (i.e., titanium alloys, calcium phosphate, polymers, etc). However, there are a few shortcomings of the biofilm formation assay. Firstly, it is not possible to distinguish whether the antimicrobial effect can be attributed to killing of planktonic bacteria before the biofilm forms or to the specific antibiofilm effects, because the antimicrobial agent is added to the bacterial suspension before the biofilm is formed. Furthermore, the crystal violet stains the whole biomass (bacteria and exopolymers) and not only the living bacteria, hence it is not possible to determine the ratio of alive vs. dead bacteria in the biofilm (Haney et al., 2018). To get comprehensive insight into how PnDs affect biofilms, additional experiments should be performed also on pre-formed biofilms (Segev-Zarko and Shai 2017). To determine the metabolically active cells in the biofilm, resazurin staining (Yadav et al., 2017) is used. To evaluate the viability of the cells, forming the biofilm, the live/dead bacterial viability kit (El-Mahdy et al., 2021; Dubus et al., 2022) is used. To evaluate the morphology of the biofilm, most frequently confocal microscopy or scanning electron microscopy are used (Yadav et al., 2017; Mao et al., 2018; El-Mahdy et al., 2021).
To evaluate the antimicrobial effect of PnDs in bacteria-infected epithelia, the following cellular in vitro models have been established. Cells are grown to confluence using antibiotic-free medium and then inoculated with bacteria. The number of viable bacteria in culture medium are quantified by plating serial dilutions of bacterial suspensions and counting the CFU (Josse et al., 2014; Sung et al., 2016; Dubus et al., 2020; Ramuta et al., 2021a; El-Mahdy et al., 2021). Next, the permeabilization agent is used to release the intracellular bacteria, which are then quantified by plating serial dilutions of bacterial suspensions and counting the CFU. (Josse et al., 2014; Dubus et al., 2020; El-Mahdy et al., 2021). Moreover, the effect of bacteria and PnDs on eukaryotic cells are evaluated by quantifying the number and viability of eukaryotic cells and also the intracellular localization of bacteria is assessed by using various methods of light, confocal and electron microscopy (Josse et al., 2014; Ramuta et al., 2021b; El-Mahdy et al., 2021). To gain better understanding of the effect of PnDs in bacteria-infected epithelia, special attention must be given to establishment of physiologically-relevant in vitro models. For example, a multilayered biomimetic porcine urothelial model that has been shown to react differently to pathogenic vs. non-pathogenic E. coli strains has been used for evaluating the antimicrobial properties of PnDs (Ramuta T.Ž. et al., 2021; Predojević et al., 2022). Similar complex in vitro models have been established to study the host-pathogen interactions in the airway mucosa (Marrazzo et al., 2016; Hasan et al., 2018), intestine (Pearce et al., 2018; García-Díaz et al., 2022) and skin (Bolle et al., 2020), but have not yet been used for evaluation of PnDs.
The antimicrobial properties of PnDs have also been evaluated in vivo using several different methods. These studies have assessed the antimicrobial activity by quantifying bacteria in the spleen or blood or indirectly by measuring antimicrobial cytokines. For example, PnDs have been shown to protect against experimental sepsis (murine cecal ligation and puncture model of sepsis) (Parolini et al., 2014; Laroye et al., 2019) and E. coli-induced acute lung injury (Sung et al., 2016).
PnDs have demonstrated contradictory effects in the field of oncology. Various factors that include the specific PnD tissue of origin, the type and size of tumor, the PnD injection route, the treatment regimen and interactions with the host appear to play a role in determining whether PnD exert pro-tumorigenic or antitumorigenic properties. To facilitate the translation of PnDs towards the clinic, it is crucial to standardize procedures for evaluating the properties of PnDs and to define the criteria that distinguish each PnDs as suitable for clinical use. PnDs-derived preparations are a very versatile group, ranging from cells and their conditioned media to tissue-derived scaffolds. This must be taken into account when selecting methods and defining criteria for validating the multimodal functions of PnDs to be used in oncological and antimicrobial applications. Another challenge is the development of assays that can efficiently and reproducibly measure the anticancer and antimicrobial properties of PnDs in vitro and in vivo models of cancer and infection.
In summary, there is a growing awareness that PnDs possess precisely tuned anticancer and antimicrobial activities. In this review, we therefore present the most commonly used functional assays (Table 2) with their advantages and disadvantages in assessing the anticancer and antimicrobial functionality of PnDs. This must be considered in future research and in the development of more effective PnDs therapies.

TABLE 2. Summary of assays used to detect antitumor and antimicrobial effects of PnDs.
All authors contributed to manuscript preparation and revision. AS, TZR, and MEK coordinated the work and compiled the manuscript.
This work was supported by: COST Action CA17116 International Network for Translating Research on Perinatal Derivatives into Therapeutic Approaches (SPRINT), supported by COST (European Cooperation in Science and Technology), the French Ministry of Health (PHRCI-2020), the Slovenian Research Agency (J7-2594, P3-0108), Italian Ministry of Research and University (MIUR, 5x1000) and the “Fondazione Alessandra Bono Onlus”.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Allen, H., Shraga-Heled, N., Blumenfeld, M., Dego-Ashto, T., Fuchs-Telem, D., Gilert, A., et al. (2018). Human placental-derived adherent stromal cells Co-induced with TNF-alpha and IFN-gamma inhibit triple-negative breast cancer in nude mouse xenograft models. Sci. Rep. 8 (1), 670. doi:10.1038/s41598-017-18428-1
Alshareeda, A. T., Rakha, E., Alghwainem, A., Alrfaei, B., Alsowayan, B., Albugami, A., et al. (2018). The effect of human placental chorionic villi derived mesenchymal stem cell on triple-negative breast cancer hallmarks. PLoS One 13 (11), e0207593. doi:10.1371/journal.pone.0207593
Ashraf, H., Font, K., Powell, C., and Schurr, M. (2019). Antimicrobial activity of an amnion-chorion membrane to oral microbes. Int. J. Dent. 2019, 1269534. doi:10.1155/2019/1269534
Ayuzawa, R., Doi, C., Rachakatla, R. S., Pyle, M. M., Maurya, D. K., Troyer, D., et al. (2009). Naive human umbilical cord matrix derived stem cells significantly attenuate growth of human breast cancer cells in vitro and in vivo. Cancer Lett. 280 (1), 31–37. doi:10.1016/j.canlet.2009.02.011
Bajetto, A., Pattarozzi, A., Corsaro, A., Barbieri, F., Daga, A., Bosio, A., et al. (2017). Different effects of human umbilical cord mesenchymal stem cells on glioblastoma stem cells by direct cell interaction or via released soluble factors. Front. Cell. Neurosci. 11, 312. doi:10.3389/fncel.2017.00312
Banerjee, A., Lindenmair, A., Hennerbichler, S., Steindorf, P., Steinborn, R., Kozlov, A. V., et al. (2018a). Cellular and site-specific mitochondrial characterization of vital human amniotic membrane. Cell Transpl. 27 (1), 3–11. doi:10.1177/0963689717735332
Banerjee, A., Lindenmair, A., Steinborn, R., Dumitrescu, S. D., Hennerbichler, S., Kozlov, A. V., et al. (2018b). Oxygen tension strongly influences metabolic parameters and the release of interleukin-6 of human amniotic mesenchymal stromal cellsin vitro. Stem Cells Int. 2018, 11. doi:10.1155/2018/9502451
Banerjee, A., Weidinger, A., Hofer, M., Steinborn, R., Lindenmair, A., Hennerbichler-Lugscheider, S., et al. (2015). Different metabolic activity in placental and reflected regions of the human amniotic membrane. Placenta 36 (11), 1329–1332. doi:10.1016/j.placenta.2015.08.015
Bhatt, A. P., Redinbo, M. R., and Bultman, S. J. (2017). The role of the microbiome in cancer development and therapy. CA A Cancer J. Clin. 67 (4), 326–344. doi:10.3322/caac.21398
Björkholm, B., Bok, C. M., Lundin, A., Rafter, J., Hibberd, M. L., and Pettersson, S. (2009). Intestinal microbiota regulate xenobiotic metabolism in the liver. PloS one 4 (9), e6958. doi:10.1371/journal.pone.0006958
Bolle, E. C. L., Verderosa, A. D., Dhouib, R., Parker, T. J., Fraser, J. F., Dargaville, T. R., et al. (2020). An in vitro reconstructed human skin equivalent model to study the role of skin integration around percutaneous devices against bacterial infection. Front. Microbiol. 11, 670. doi:10.3389/fmicb.2020.00670
Bonomi, A., Silini, A., Vertua, E., Signoroni, P. B., Cocce, V., Cavicchini, L., et al. (2015). Human amniotic mesenchymal stromal cells (hAMSCs) as potential vehicles for drug delivery in cancer therapy: An in vitro study. Stem Cell Res. Ther. 6 (1), 155. doi:10.1186/s13287-015-0140-z
Borghesi, J., Caceres, S., Mario, L. C., Alonso-Diez, A., Silveira Rabelo, A. C., Illera, M. J., et al. (2020). Effects of doxorubicin associated with amniotic membrane stem cells in the treatment of canine inflammatory breast carcinoma (IPC-366) cells. BMC Vet. Res. 16 (1), 353. doi:10.1186/s12917-020-02576-0
Bu, S., Zhang, Q., Wang, Q., and Lai, D. (2017). Human amniotic epithelial cells inhibit growth of epithelial ovarian cancer cells via TGF-β1-mediated cell cycle arrest. Int. J. Oncol. 51 (5), 1405–1414. doi:10.3892/ijo.2017.4123
Cafforio, P., Viggiano, L., Mannavola, F., Pellè, E., Caporusso, C., Maiorano, E., et al. (2017). pIL6-TRAIL-engineered umbilical cord mesenchymal/stromal stem cells are highly cytotoxic for myeloma cells both in vitro and in vivo. Stem Cell Res. Ther. 8 (1), 206. doi:10.1186/s13287-017-0655-6
Carmody, R. N., and Turnbaugh, P. J. (2014). Host-microbial interactions in the metabolism of therapeutic and diet-derived xenobiotics. J. Clin. Invest. 124 (10), 4173–4181. doi:10.1172/jci72335
Chai, L., Bai, L., Li, L., Chen, F., and Zhang, J. (2018). Biological functions of lung cancer cells are suppressed in co-culture with mesenchymal stem cells isolated from umbilical cord. Exp. Ther. Med. 15 (1), 1076–1080. doi:10.3892/etm.2017.5456
Chao, K. C., Yang, H. T., and Chen, M. W. (2012). Human umbilical cord mesenchymal stem cells suppress breast cancer tumourigenesis through direct cell-cell contact and internalization. J. Cell. Mol. Med. 16 (8), 1803–1815. doi:10.1111/j.1582-4934.2011.01459.x
Chen, Q., Cheng, P., Song, N., Yin, T., He, H., Yang, L., et al. (2012). Antitumor activity of placenta-derived mesenchymal stem cells producing pigment epithelium-derived factor in a mouse melanoma model. Oncol. Lett. 4 (3), 413–418. doi:10.3892/ol.2012.772
Chen, X., Wang, K., Chen, S., and Chen, Y. (2019). Effects of mesenchymal stem cells harboring the Interferon-β gene on A549 lung cancer in nude mice. Pathology - Res. Pract. 215 (3), 586–593. doi:10.1016/j.prp.2019.01.013
Chetty, S. S., Praneetha, S., Vadivel Murugan, A., Govarthanan, K., and Verma, R. S. (2020). Human umbilical cord wharton's jelly-derived mesenchymal stem cells labeled with Mn(2+) and Gd(3+) Co-doped CuInS(2)-ZnS nanocrystals for multimodality imaging in a tumor mice model. ACS Appl. Mat. Interfaces 12 (3), 3415–3429. doi:10.1021/acsami.9b19054
Choi, J. H., Lim, S. M., Yoo, Y. I., Jung, J., Park, J. W., and Kim, G. J. (2016). Microenvironmental interaction between hypoxia and endothelial cells controls the migration ability of placenta-derived mesenchymal stem cells via α4 integrin and rho signaling. J. Cell. Biochem. 117 (5), 1145–1157. doi:10.1002/jcb.25398
Ciavarella, S., Caselli, A., Tamma, A. V., Savonarola, A., Loverro, G., Paganelli, R., et al. (2015). A peculiar molecular profile of umbilical cord-mesenchymal stromal cells drives their inhibitory effects on multiple myeloma cell growth and tumor progression. Stem Cells Dev. 24 (12), 1457–1470. doi:10.1089/scd.2014.0254
Dabrowski, F. A., Burdzinska, A., Kulesza, A., Sladowska, A., Zolocinska, A., Gala, K., et al. (2017). Comparison of the paracrine activity of mesenchymal stem cells derived from human umbilical cord, amniotic membrane and adipose tissue. J. Obstet. Gynaecol. Res. 43 (11), 1758–1768. doi:10.1111/jog.13432
Del Fattore, A., Luciano, R., Saracino, R., Battafarano, G., Rizzo, C., Pascucci, L., et al. (2015). Differential effects of extracellular vesicles secreted by mesenchymal stem cells from different sources on glioblastoma cells. Expert Opin. Biol. Ther. 15 (4), 495–504. doi:10.1517/14712598.2015.997706
Di Germanio, C., Bernier, M., Petr, M., Mattioli, M., Barboni, B., and de Cabo, R. (2016). Conditioned medium derived from rat amniotic epithelial cells confers protection against inflammation, cancer, and senescence. Oncotarget 7, 39051–39064. doi:10.18632/oncotarget.9694
Di, G. H., Liu, Y., Lu, Y., Liu, J., Wu, C., and Duan, H. F. (2014). IL-6 secreted from senescent mesenchymal stem cells promotes proliferation and migration of breast cancer cells. PLoS One 9 (11), e113572. doi:10.1371/journal.pone.0113572
Dong, L., Pu, Y., Zhang, L., Qi, Q., Xu, L., Li, W., et al. (2018). Human umbilical cord mesenchymal stem cell-derived extracellular vesicles promote lung adenocarcinoma growth by transferring miR-410. Cell Death Dis. 9 (2), 218. doi:10.1038/s41419-018-0323-5
Drago, L., Cappelletti, L., De Vecchi, E., Pignataro, L., Torretta, S., and Mattina, R. (2014). Antiadhesive and antibiofilm activity of hyaluronic acid against bacteria responsible for respiratory tract infections. Apmis 122 (10), 1013–1019. doi:10.1111/apm.12254
Du, T., Ju, G., Wu, S., Cheng, Z., Cheng, J., Zou, X., et al. (2014). Microvesicles derived from human Wharton's jelly mesenchymal stem cells promote human renal cancer cell growth and aggressiveness through induction of hepatocyte growth factor. PLoS One 9 (5), e96836. doi:10.1371/journal.pone.0096836
Dubus, M., Scomazzon, L., Chevrier, J., Ledouble, C., Baldit, A., Braux, J., et al. (2022). Antibacterial and immunomodulatory properties of acellular wharton's jelly matrix. Biomedicines 10 (2), 227. doi:10.3390/biomedicines10020227
Dubus, M., Varin-Simon, J., Prada, P., Scomazzon, L., Reffuveille, F., Alem, H., et al. (2020). Biopolymers-calcium phosphate antibacterial coating reduces the pathogenicity of internalized bacteria by mesenchymal stromal cells. Biomater. Sci. 8 (20), 5763–5773. doi:10.1039/d0bm00962h
Dzobo, K., Vogelsang, M., Thomford, N. E., Dandara, C., Kallmeyer, K., Pepper, M. S., et al. (2016). Wharton’s jelly-derived mesenchymal stromal cells and fibroblast-derived extracellular matrix synergistically activate apoptosis in a p21-dependent mechanism in WHCO1 and MDA MB 231 cancer cellsin vitro. Stem Cells Int. 2016, 1–17. doi:10.1155/2016/4842134
El-Mahdy, T. S., Mongaret, C., Varin-Simon, J., Lamret, F., Vernet-Garnier, V., Rammal, H., et al. (2021). Interaction of implant infection-related commensal bacteria with mesenchymal stem cells: A comparison between cutibacterium acnes and Staphylococcus aureus. FEMS Microbiol. Lett. 368 (4), fnab014. doi:10.1093/femsle/fnab014
Fan, S., Gao, H., Ji, W., Zhu, F., Sun, L., Liu, Y., et al. (2020). Umbilical cord-derived mesenchymal stromal/stem cells expressing IL-24 induce apoptosis in gliomas. J. Cell. Physiol. 235 (2), 1769–1779. doi:10.1002/jcp.29095
Faraj, L., Stewart, E., Albert, R., Allen, C., Petrovski, G., Dua, H., et al. (2015). In-vitro anti angiogenic effects of cryo-preserved amniotic membrane. Acta Ophthalmol. 93 (S255). doi:10.1111/j.1755-3768.2015.0635
Faridvand, Y., Nozari, S., Vahedian, V., Safaie, N., Pezeshkian, M., Haddadi, P., et al. (2019). Nrf2 activation and down-regulation of HMGB1 and MyD88 expression by amnion membrane extracts in response to the hypoxia-induced injury in cardiac H9c2 cells. Biomed. Pharmacother. 109, 360–368. doi:10.1016/j.biopha.2018.10.035
Fong, C. Y., Chak, L. L., Biswas, A., Tan, J. H., Gauthaman, K., Chan, W. K., et al. (2011). Human Wharton's jelly stem cells have unique transcriptome profiles compared to human embryonic stem cells and other mesenchymal stem cells. Stem Cell Rev. Rep. 7 (1), 1–16. doi:10.1007/s12015-010-9166-x
Ganjibakhsh, M., Mehraein, F., Koruji, M., Aflatoonian, R., and Farzaneh, P. (2019). Three-dimensional decellularized amnion membrane scaffold as a novel tool for cancer research; cell behavior, drug resistance and cancer stem cell content. Mater. Sci. Eng. C 100, 330–340. doi:10.1016/j.msec.2019.02.090
García-Díaz, M., Cendra, M. D. M., Alonso-Roman, R., Urdániz, M., Torrents, E., and Martínez, E. (2022). Mimicking the intestinal host-pathogen interactions in a 3D in vitro model: The role of the mucus layer. Pharmaceutics 14 (8), 1552. doi:10.3390/pharmaceutics14081552
Gauthaman, K., Yee, F. C., Cheyyatraivendran, S., Biswas, A., Choolani, M., and Bongso, A. (2012). Human umbilical cord Wharton's jelly stem cell (hWJSC) extracts inhibit cancer cell growth in vitro. J. Cell. Biochem. 113 (6), 2027–2039. doi:10.1002/jcb.24073
Gnaiger, E. (2008). Polarographic oxygen sensors, the oxygraph, and high-resolution respirometry to assess mitochondrial function. in Drug-induced mitochondrial dysfunction. New Jersey, USA: John Wiley & Sons (Wiley), 325–352.
Grzywocz, Z., Hoser, G., Sabalinska, S., Ladyzynski, P., Czubak, J., Dworczynska, M., et al. (2018). Response of human normal and leukemia cells to factors released by amnion fragments in vitro. PLOS ONE 13 (3), e0195035. doi:10.1371/journal.pone.0195035
Haiser, H. J., and Turnbaugh, P. J. (2012). Is it time for a metagenomic basis of therapeutics? Science 336 (6086), 1253–1255. doi:10.1126/science.1224396
Han, I., Yun, M., Kim, E. O., Kim, B., Jung, M. H., and Kim, S. H. (2014). Retracted article: Umbilical cord tissue-derived mesenchymal stem cells induce apoptosis in PC-3 prostate cancer cells through activation of JNK and downregulation of PI3K/AKT signaling. Stem Cell Res. Ther. 5 (2), 54. doi:10.1186/scrt443
Hanahan, D., and Weinberg, R. A. (2011). Hallmarks of cancer: The next generation. Cell 144 (5), 646–674. doi:10.1016/j.cell.2011.02.013
Haney, E. F., Trimble, M. J., Cheng, J. T., Vallé, Q., and Hancock, R. E. W. (2018). Critical assessment of methods to quantify biofilm growth and evaluate antibiofilm activity of host defence peptides. Biomolecules 8 (2), 29. doi:10.3390/biom8020029
Hasan, S., Sebo, P., and Osicka, R. (2018). A guide to polarized airway epithelial models for studies of host-pathogen interactions. FEBS J. 285 (23), 4343–4358. doi:10.1111/febs.14582
Hendijani, F., Javanmard, S. H., and Sadeghi-aliabadi, H. (2015). Human Wharton's jelly mesenchymal stem cell secretome display antiproliferative effect on leukemia cell line and produce additive cytotoxic effect in combination with doxorubicin. Tissue Cell 47 (3), 229–234. doi:10.1016/j.tice.2015.01.005
Hütter, E., Unterluggauer, H., Garedew, A., Jansen-Dürr, P., and Gnaiger, E. (2006). High-resolution respirometry--a modern tool in aging research. Exp. Gerontol. 41 (1), 103–109. doi:10.1016/j.exger.2005.09.011
Iravani, A., and Hicks, R. J. (2020). Imaging the cancer immune environment and its response to pharmacologic intervention, Part 2: The role of novel PET agents. J. Nucl. Med. 61 (11), 1553–1559. doi:10.2967/jnumed.120.248823
Jain, T., Sharma, P., Are, A. C., Vickers, S. M., and Dudeja, V. (2021). New insights into the cancer-microbiome-immune Axis: Decrypting a decade of discoveries. Front. Immunol. 12, 622064. doi:10.3389/fimmu.2021.622064
Janev, A., Ramuta, T. Ž., Tratnjek, L., Sardoč, Ž., Obradović, H., Mojsilović, S., et al. (2021). Detrimental effect of various preparations of the human amniotic membrane homogenate on the 2D and 3D bladder cancer in vitro models. Front. Bioeng. Biotechnol. 9, 690358. doi:10.3389/fbioe.2021.690358
Jiao, H., Guan, F., Yang, B., Li, J., Song, L., Hu, X., et al. (2012). Human amniotic membrane derived-mesenchymal stem cells induce C6 glioma apoptosis in vivo through the Bcl-2/caspase pathways. Mol. Biol. Rep. 39 (1), 467–473. doi:10.1007/s11033-011-0760-z
Jiao, Y., Zhao, H., Chen, G., Sang, X., Yang, L., Hou, Z., et al. (2018). Pyroptosis of MCF7 cells induced by the secreted factors of hUCMSCs. Stem Cells Int. 2018, 1–12. doi:10.1155/2018/5912194
Jorgensen, J. H., and Ferraro, M. J. (2009). Antimicrobial susceptibility testing: A review of general principles and contemporary practices. Clin. Infect. Dis. 49 (11), 1749–1755. doi:10.1086/647952
Josse, J., Velard, F., Mechiche Alami, S., Brun, V., Guillaume, C., Kerdjoudj, H., et al. (2014). Increased internalization of Staphylococcus aureus and cytokine expression in human Wharton's jelly mesenchymal stem cells. Biomed. Mat. Eng. 24 (1), 27–35. doi:10.3233/bme-140971
Kalamegam, G., Sait, K. H. W., Ahmed, F., Kadam, R., Pushparaj, P. N., Anfinan, N., et al. (2018). Human wharton's jelly stem cell (hWJSC) extracts inhibit ovarian cancer cell lines OVCAR3 and SKOV3 in vitro by inducing cell cycle arrest and apoptosis. Front. Oncol. 8, 592. doi:10.3389/fonc.2018.00592
Kalamegam, G., Sait, K. H. W., Anfinan, N., Kadam, R., Ahmed, F., Rasool, M., et al. (2019). Cytokines secreted by human Wharton's jelly stem cells inhibit the proliferation of ovarian cancer (OVCAR3) cells in vitro. Oncol. Lett. 17 (5), 4521–4531. doi:10.3892/ol.2019.10094
Kamalabadi-Farahani, M., Vasei, M., Ahmadbeigi, N., Ebrahimi-Barough, S., Soleimani, M., and Roozafzoon, R. (2018). Anti-tumour effects of TRAIL-expressing human placental derived mesenchymal stem cells with curcumin-loaded chitosan nanoparticles in a mice model of triple negative breast cancer. Artif. Cells Nanomed. Biotechnol. 46 (3), S1011–s1021. doi:10.1080/21691401.2018.1527345
Kang, N. H., Yi, B. R., Lim, S. Y., Hwang, K. A., Baek, Y. S., Kang, K. S., et al. (2012). Human amniotic membrane-derived epithelial stem cells display anticancer activity in BALB/c female nude mice bearing disseminated breast cancer xenografts. Int. J. Oncol. 40 (6), 2022–2028. doi:10.3892/ijo.2012.1372
Khalil, C., Moussa, M., Azar, A., Tawk, J., Habbouche, J., Salameh, R., et al. (2019). Anti-proliferative effects of mesenchymal stem cells (MSCs) derived from multiple sources on ovarian cancer cell lines: An in-vitro experimental study. J. Ovarian Res. 12 (1), 70. doi:10.1186/s13048-019-0546-9
Kim, H. S., Cho, J. H., Park, H. W., Yoon, H., Kim, M. S., and Kim, S. C. (2002). Endotoxin-neutralizing antimicrobial proteins of the human placenta. J. Immunol. 168 (5), 2356–2364. doi:10.4049/jimmunol.168.5.2356
Kim, J.-Y., Park, S.-C., Lee, J.-K., Choi, S. J., Hahm, K.-S., and Park, Y. (2012). Novel antibacterial activity of β2-microglobulin in human amniotic fluid. PLOS ONE 7 (11), e47642. doi:10.1371/journal.pone.0047642
Kim, S.-H., Bang, S. H., Kang, S. Y., Park, K. D., Eom, J. H., Oh, I. U., et al. (2015). Human amniotic membrane-derived stromal cells (hAMSC) interact depending on breast cancer cell type through secreted molecules. Tissue Cell 47 (1), 10–16. doi:10.1016/j.tice.2014.10.003
King, A. E., Kelly, R. W., Sallenave, J. M., Bocking, A. D., and Challis, J. R. (2007a). Innate immune defences in the human uterus during pregnancy. Placenta 28 (11-12), 1099–1106. doi:10.1016/j.placenta.2007.06.002
King, A. E., Paltoo, A., Kelly, R. W., Sallenave, J. M., Bocking, A. D., and Challis, J. R. (2007b). Expression of natural antimicrobials by human placenta and fetal membranes. Placenta 28 (2-3), 161–169. doi:10.1016/j.placenta.2006.01.006
Kjaergaard, N., Hein, M., Hyttel, L., Helmig, R. B., Schønheyder, H. C., Uldbjerg, N., et al. (2001). Antibacterial properties of human amnion and chorion in vitro. Eur. J. Obstet. Gynecol. Reprod. Biol. X. 94 (2), 224–229. doi:10.1016/s0301-2115(00)00345-6
Klaffenbach, D., Friedrich, D., Strick, R., Strissel, P. L., Beckmann, M. W., Rascher, W., et al. (2011). Contribution of different placental cells to the expression and stimulation of antimicrobial proteins (AMPs). Placenta 32 (11), 830–837. doi:10.1016/j.placenta.2011.08.004
Komaki, M., Numata, Y., Morioka, C., Honda, I., Tooi, M., Yokoyama, N., et al. (2017). Exosomes of human placenta-derived mesenchymal stem cells stimulate angiogenesis. Stem Cell Res. Ther. 8 (1), 219. doi:10.1186/s13287-017-0660-9
Kramer, N., Walzl, A., Unger, C., Rosner, M., Krupitza, G., Hengstschläger, M., et al. (2013). In vitro cell migration and invasion assays. Mutat. Research/Reviews Mutat. Res. 752 (1), 10–24. doi:10.1016/j.mrrev.2012.08.001
Lang, D. S., Marwitz, S., Zeiser, T., Seehase, S., Watermann, I., Vollmer, E., et al. (2015). Placenta-derived conditioned medium with anti-tumor properties on human NSCLC. Eur. Respir. J. 46 (59), OA4983. doi:10.1183/13993003
Laroye, C., Boufenzer, A., Jolly, L., Cunat, L., Alauzet, C., Merlin, J.-L., et al. (2019). Bone marrow vs Wharton’s jelly mesenchymal stem cells in experimental sepsis: A comparative study. Stem Cell Res. Ther. 10 (1), 192. doi:10.1186/s13287-019-1295-9
Lehouritis, P., Cummins, J., Stanton, M., Murphy, C. T., McCarthy, F. O., Reid, G., et al. (2015). Local bacteria affect the efficacy of chemotherapeutic drugs. Sci. Rep. 5, 14554. doi:10.1038/srep14554
Leng, L., Wang, Y., He, N., Wang, D., Zhao, Q., Feng, G., et al. (2014). Molecular imaging for assessment of mesenchymal stem cells mediated breast cancer therapy. Biomaterials 35 (19), 5162–5170. doi:10.1016/j.biomaterials.2014.03.014
Li, T., Zhang, C., Ding, Y., Zhai, W., Liu, K., Bu, F., et al. (2015). Umbilical cord-derived mesenchymal stem cells promote proliferation and migration in MCF-7 and MDA-MB-231 breast cancer cells through activation of the ERK pathway. Oncol. Rep. 34 (3), 1469–1477. doi:10.3892/or.2015.4109
Li, X., Ling, W., Pennisi, A., Wang, Y., Khan, S., Heidaran, M., et al. (2011). Human placenta-derived adherent cells prevent bone loss, stimulate bone formation, and suppress growth of multiple myeloma in bone. Stem Cells 29 (2), 263–273. doi:10.1002/stem.572
Li, X., Liu, L. l., Yao, J. l., Wang, K., and Ai, H. (2019). Human umbilical cord mesenchymal stem cell-derived extracellular vesicles inhibit endometrial cancer cell proliferation and migration through delivery of exogenous miR-302a. Stem Cells Int. 2019, 1–11. doi:10.1155/2019/8108576
Liepke, C., Baxmann, S., Heine, C., Breithaupt, N., Ständker, L., and Forssmann, W. G. (2003). Human hemoglobin-derived peptides exhibit antimicrobial activity: A class of host defense peptides. J. Chromatogr. B 791 (1-2), 345–356. doi:10.1016/s1570-0232(03)00245-9
Lin D, D. H., Biswas, A., Choolani, M., Fong, C. Y., and Bongso, A. (2017). Induction of immunogenic cell death in lymphoma cells by wharton's jelly mesenchymal stem cell conditioned medium. Stem Cell Rev. Rep. 13 (6), 801–816. doi:10.1007/s12015-017-9767-8
Lin H, H., Fang, Z., Su, Y., Li, P., Wang, J., Liao, H., et al. (2017). DHX32 promotes angiogenesis in colorectal cancer through augmenting β-catenin signaling to induce expression of VEGFA. EBioMedicine 18, 62–72. doi:10.1016/j.ebiom.2017.03.012
Lin, H. D., Fong, C. Y., Biswas, A., Choolani, M., and Bongso, A. (2016). Human umbilical cord wharton's jelly stem cell conditioned medium induces tumoricidal effects on lymphoma cells through hydrogen peroxide mediation. J. Cell. Biochem. 117 (9), 2045–2055. doi:10.1002/jcb.25501
Lin, H. D., Fong, C. Y., Biswas, A., Choolani, M., and Bongso, A. (2014). Human Wharton's jelly stem cells, its conditioned medium and cell-free lysate inhibit the growth of human lymphoma cells. Stem Cell Rev. Rep. 10 (4), 573–586. doi:10.1007/s12015-014-9514-3
Liu, J., Han, G., Liu, H., and Qin, C. (2013). Suppression of cholangiocarcinoma cell growth by human umbilical cord mesenchymal stem cells: A possible role of wnt and akt signaling. PLoS ONE 8 (4), e62844. doi:10.1371/journal.pone.0062844
Ma, F., Chen, D., Chen, F., Chi, Y., Han, Z., Feng, X., et al. (2015). Human umbilical cord mesenchymal stem cells promote breast cancer metastasis by interleukin-8- and interleukin-6-dependent induction of CD44(+)/CD24(-) cells. Cell Transpl. 24 (12), 2585–2599. doi:10.3727/096368915x687462
Magatti, M., De Munari, S., Vertua, E., and Parolini, O. (2012). Amniotic membrane-derived cells inhibit proliferation of cancer cell lines by inducing cell cycle arrest. J. Cell. Mol. Med. 16 (9), 2208–2218. doi:10.1111/j.1582-4934.2012.01531.x
Magatti, M., Vertua, E., De Munari, S., Caro, M., Caruso, M., Silini, A., et al. (2017). Human amnion favours tissue repair by inducing the M1-to-M2 switch and enhancing M2 macrophage features. J. Tissue Eng. Regen. Med. 11 (10), 2895–2911. doi:10.1002/term.2193
Mamede, A. C., Guerra, S., Laranjo, M., Carvalho, M. J., Oliveira, R. C., Goncalves, A. C., et al. (2015). Selective cytotoxicity and cell death induced by human amniotic membrane in hepatocellular carcinoma. Med. Oncol. 32 (12), 257. doi:10.1007/s12032-015-0702-z
Mandal, S., Arfuso, F., Sethi, G., Dharmarajan, A., and Warrier, S. (2019). Encapsulated human mesenchymal stem cells (eMSCs) as a novel anti-cancer agent targeting breast cancer stem cells: Development of 3D primed therapeutic MSCs. Int. J. Biochem. Cell Biol. 110, 59–69. doi:10.1016/j.biocel.2019.02.001
Mao, Y., Hoffman, T., Johnson, A., Duan-Arnold, Y., Danilkovitch, A., and Kohn, J. (2016). Human cryopreserved viable amniotic membrane inhibits the growth of bacteria associated with chronic wounds. J. Diabet. Foot Complicat. 8 (2), 8.
Mao, Y., Hoffman, T., Singh-Varma, A., Duan-Arnold, Y., Moorman, M., Danilkovitch, A., et al. (2017). Antimicrobial peptides secreted from human cryopreserved viable amniotic membrane contribute to its antibacterial activity. Sci. Rep. 7 (1), 13722. doi:10.1038/s41598-017-13310-6
Mao, Y., Singh-Varma, A., Hoffman, T., Dhall, S., Danilkovitch, A., and Kohn, J. (2018). The effect of cryopreserved human placental tissues on biofilm formation of wound-associated pathogens. J. Funct. Biomater. 9 (1), 3. doi:10.3390/jfb9010003
Marcuzzo, A. V., Tofanelli, M., Boscolo Nata, F., Gatto, A., and Tirelli, G. (2017). Hyaluronate effect on bacterial biofilm in ENT district infections: A review. Apmis 125 (9), 763–772. doi:10.1111/apm.12728
Marleau, A. M., McDonald, G., Koropatnick, J., Chen, C.-S., and Koos, D. (2012). Reduction of tumorigenicity by placental extracts. Anticancer Res. 32 (4), 1153–1161.
Marrazzo, P., Maccari, S., Taddei, A., Bevan, L., Telford, J., Soriani, M., et al. (2016). 3D reconstruction of the human airway mucosa in vitro as an experimental model to study NTHi infections. PLoS One 11 (4), e0153985. doi:10.1371/journal.pone.0153985
Marshall, J. (2011). Transwell(®) invasion assays. Methods Mol. Biol. 769, 97–110. doi:10.1007/978-1-61779-207-6_8
Martin, T. A., and Jiang, W. G. (2009). Loss of tight junction barrier function and its role in cancer metastasis. Biochimica Biophysica Acta - Biomembr. 1788 (4), 872–891. doi:10.1016/j.bbamem.2008.11.005
Martin, T. A., Ye, L., Sanders, A. J., Lane, J., and Jiang, W. G. (2013). Cancer invasion and metastasis: Molecular and cellular perspective. Austin (TX): Landes Bioscience.
Meng, M. Y., Li, L., Wang, W. J., Liu, F. F., Song, J., Yang, S. L., et al. (2019). Assessment of tumor promoting effects of amniotic and umbilical cord mesenchymal stem cells in vitro and in vivo. J. Cancer Res. Clin. Oncol. 145 (5), 1133–1146. doi:10.1007/s00432-019-02859-6
Menyhárt, O., Harami-Papp, H., Sukumar, S., Schäfer, R., Magnani, L., de Barrios, O., et al. (2016). Guidelines for the selection of functional assays to evaluate the hallmarks of cancer. Biochimica Biophysica Acta (BBA) - Rev. Cancer 1866 (2), 300–319. doi:10.1016/j.bbcan.2016.10.002
Modaresifar, K., Azizian, S., Zolghadr, M., Moravvej, H., Ahmadiani, A., and Niknejad, H. (2017). The effect of cryopreservation on anti-cancer activity of human amniotic membrane. Cryobiology 74, 61–67. doi:10.1016/j.cryobiol.2016.12.001
Nelson, D. L., and Cox, M. M. (2004). Lehninger principles of biochemistry. New York, NY: W. H. Freeman.
Niknejad, H., Khayat-Khoei, M., Peirovi, H., and Abolghasemi, H. (2014). Human amniotic epithelial cells induce apoptosis of cancer cells: A new anti-tumor therapeutic strategy. Cytotherapy 16 (1), 33–40. doi:10.1016/j.jcyt.2013.07.005
Palanker, N. D., Lee, C. T., Weltman, R. L., Tribble, G. D., van der Hoeven, R., Hong, J., et al. (2019). Antimicrobial efficacy assessment of human derived composite amnion-chorion membrane. Sci. Rep. 9 (1), 15600. doi:10.1038/s41598-019-52150-4
Paris, J. L., de la Torre, P., Manzano, M., Cabañas, M. V., Flores, A. I., and Vallet-Regí, M. (2016). Decidua-derived mesenchymal stem cells as carriers of mesoporous silica nanoparticles. in vitro and in vivo evaluation on mammary tumors. Acta Biomater. 33, 275–282. doi:10.1016/j.actbio.2016.01.017
Parolini, O., Souza-Moreira, L., O'Valle, F., Magatti, M., Hernandez-Cortes, P., Gonzalez-Rey, E., et al. (2014). Therapeutic effect of human amniotic membrane-derived cells on experimental arthritis and other inflammatory disorders. Arthritis Rheumatol. 66 (2), 327–339. doi:10.1002/art.38206
Pearce, S. C., Coia, H. G., Karl, J. P., Pantoja-Feliciano, I. G., Zachos, N. C., and Racicot, K. (2018). Intestinal in vitro and ex vivo models to study host-microbiome interactions and acute stressors. Front. Physiol. 9, 1584. doi:10.3389/fphys.2018.01584
Poženel, L., Lindenmair, A., Schmidt, K., Kozlov, A. V., Grillari, J., Wolbank, S., et al. (2019). Critical impact of human amniotic membrane tension on mitochondrial function and cell viability in vitro. Cells 8 (12), 1641. doi:10.3390/cells8121641
Predojević, L., Keše, D., Žgur Bertok, D., Železnik Ramuta, T., Veranič, P., Erdani Kreft, M., et al. (2022). A biomimetic porcine urothelial model for assessing Escherichia coli pathogenicity. Microorganisms 10 (4), 783. doi:10.3390/microorganisms10040783
Puaux, A. L., Ong, L. C., Jin, Y., Teh, I., Hong, M., Chow, P. K., et al. (2011). A comparison of imaging techniques to monitor tumor growth and cancer progression in living animals. Int. J. Mol. Imaging 2011, 321538. doi:10.1155/2011/321538
Ramuta, T., Jerman, U. D., Tratnjek, L., Janev, A., Magatti, M., Vertua, E., et al. (2020a). The cells and extracellular matrix of human amniotic membrane hinder the growth and invasive potential of bladder urothelial cancer cells. Front. Bioeng. Biotechnol. 8, 554530. doi:10.3389/fbioe.2020.554530
Ramuta, T., Šket, T., Starčič Erjavec, M., and Kreft, M. E. (2021a). Antimicrobial activity of human fetal membranes: From biological function to clinical use. Front. Bioeng. Biotechnol. 9, 691522. doi:10.3389/fbioe.2021.691522
Ramuta, T., Starčič Erjavec, M., and Kreft, M. E. (2020b). Amniotic membrane preparation crucially affects its broad-spectrum activity against uropathogenic bacteria. Front. Microbiol. 11, 469. doi:10.3389/fmicb.2020.00469
Ramuta, T., Tratnjek, L., Janev, A., Seme, K., Starčič Erjavec, M., and Kreft, M. E. (2021b). The antibacterial activity of human amniotic membrane against multidrug-resistant bacteria associated with urinary tract infections: New insights from normal and cancerous urothelial models. Biomedicines 9 (2), 218. doi:10.3390/biomedicines9020218
Ramuta, T. Z., Starčič Erjavec, M., and Kreft, M. E. (2020c). Amniotic membrane preparation crucially affects its broad-spectrum activity against uropathogenic bacteria. Front. Microbiol. 11, 469. doi:10.3389/fmicb.2020.00469
Ramuta, T. Ž., Tratnjek, L., Janev, A., Seme, K., Starčič Erjavec, M., and Kreft, M. E. (2021c). The antibacterial activity of human amniotic membrane against multidrug-resistant bacteria associated with urinary tract infections: New insights from normal and cancerous urothelial models. Biomedicines 9 (2), 218. doi:10.3390/biomedicines9020218
Raza, M. H., Gul, K., Arshad, A., Riaz, N., Waheed, U., Rauf, A., et al. (2019). Microbiota in cancer development and treatment. J. Cancer Res. Clin. Oncol. 145 (1), 49–63. doi:10.1007/s00432-018-2816-0
Rezaei-Tazangi, F., Alidadi, H., Samimi, A., Karimi, S., and Kahorsandi, L. (2020). Effects of Wharton's jelly mesenchymal stem cells-derived secretome on colon carcinoma HT-29 cells. Tissue Cell 67, 101413. doi:10.1016/j.tice.2020.101413
Riedel, R., Pérez-Pérez, A., Carmona-Fernández, A., Jaime, M., Casale, R., Dueñas, J. L., et al. (2019). Human amniotic membrane conditioned medium inhibits proliferation and modulates related microRNAs expression in hepatocarcinoma cells. Sci. Rep. 9 (1), 14193. doi:10.1038/s41598-019-50648-5
Robson, M. C., and Krizek, T. J. (1973). The effect of human amniotic membranes on the bacterial population of infected rat burns. Ann. Surg. 177 (2), 144–149. doi:10.1097/00000658-197302000-00003
Rolfo, A., Giuffrida, D., Giuffrida, M. C., Todros, T., and Calogero, A. E. (2014). New perspectives for prostate cancer treatment: In vitro inhibition of LNCaP and PC3 cell proliferation by amnion-derived mesenchymal stromal cells conditioned media. Aging Male 17 (2), 94–101. doi:10.3109/13685538.2014.896894
Roy, S., and Trinchieri, G. (2017). Microbiota: A key orchestrator of cancer therapy. Nat. Rev. Cancer 17 (5), 271–285. doi:10.1038/nrc.2017.13
Segev-Zarko, L. A., and Shai, Y. (2017). Methods for investigating biofilm inhibition and degradation by antimicrobial peptides. Methods Mol. Biol. 1548, 309–322. doi:10.1007/978-1-4939-6737-7_22
Selwyn, F. P., Cheng, S. L., Bammler, T. K., Prasad, B., Vrana, M., Klaassen, C., et al. (2015). Developmental regulation of drug-processing genes in livers of germ-free mice. Toxicol. Sci. 147 (1), 84–103. doi:10.1093/toxsci/kfv110
Selwyn, F. P., Cheng, S. L., Klaassen, C. D., and Cui, J. Y. (2016). Regulation of hepatic drug-metabolizing enzymes in germ-free mice by conventionalization and probiotics. Drug Metab. Dispos. 44 (2), 262–274. doi:10.1124/dmd.115.067504
Shen, C. J., Chan, T. F., Chen, C. C., Hsu, Y. C., Long, C. Y., and Lai, C. S. (2016). Human umbilical cord matrix-derived stem cells expressing interferon-β gene inhibit breast cancer cells via apoptosis. Oncotarget 7 (23), 34172–34179. doi:10.18632/oncotarget.8997
Silini, A. R., Magatti, M., Cargnoni, A., and Parolini, O. (2017). Is immune modulation the mechanism underlying the beneficial effects of amniotic cells and their derivatives in regenerative medicine? Cell Transpl. 26 (4), 531–539. doi:10.3727/096368916x693699
Silini, A. R., Masserdotti, A., Papait, A., and Parolini, O. (2019). Shaping the future of perinatal cells: Lessons from the past and interpretations of the present. Front. Bioeng. Biotechnol. 7, 75. doi:10.3389/fbioe.2019.00075
Silva, M., Monteiro, G. A., Fialho, A. M., Bernardes, N., and da Silva, C. L. (2020). Conditioned medium from azurin-expressing human mesenchymal stromal cells demonstrates antitumor activity against breast and lung cancer cell lines. Front. Cell Dev. Biol. 8, 471. doi:10.3389/fcell.2020.00471
Sivandzade, F., Bhalerao, A., and Cucullo, L. (2019). Analysis of the mitochondrial membrane potential using the cationic JC-1 dye as a sensitive fluorescent probe. Bio. Protoc. 9 (1), e3128. doi:10.21769/bioprotoc.3128
Šket, T., Ramuta, T., Starčič Erjavec, M., and Kreft, M. E. (2019). Different effects of amniotic membrane homogenate on the growth of uropathogenic Escherichia coli, Staphylococcus aureus and Serratia marcescens. Infect. Drug Resist. 12, 3365–3375. doi:10.2147/IDR.S215006
So, K. A., Min, K. J., Hong, J. H., and Lee, J. K. (2015). Interleukin-6 expression by interactions between gynecologic cancer cells and human mesenchymal stem cells promotes epithelial-mesenchymal transition. Int. J. Oncol. 47 (4), 1451–1459. doi:10.3892/ijo.2015.3122
Subramanian, A., Shu-Uin, G., Kae-Siang, N., Gauthaman, K., Biswas, A., Choolani, M., et al. (2012). Human umbilical cord Wharton's jelly mesenchymal stem cells do not transform to tumor-associated fibroblasts in the presence of breast and ovarian cancer cells unlike bone marrow mesenchymal stem cells. J. Cell. Biochem. 113 (6), 1886–1895. doi:10.1002/jcb.24057
Sung, D. K., Chang, Y. S., Sung, S. I., Yoo, H. S., Ahn, S. Y., and Park, W. S. (2016). Antibacterial effect of mesenchymal stem cells against Escherichia coli is mediated by secretion of beta- defensin- 2 via toll- like receptor 4 signalling. Cell. Microbiol. 18 (3), 424–436. doi:10.1111/cmi.12522
Svitina, H., Skrypkina, I., Areshkov, P., Kyryk, V., Bukreieva, T., Klymenko, P., et al. (2018). Transplantation of placenta-derived multipotent cells in rats with dimethylhydrazine-induced colon cancer decreases survival rate. Oncol. Lett. 15 (4), 5034–5042. doi:10.3892/ol.2018.7996
Talmi, Y. P., Sigler, L., Inge, E., Finkelstein, Y., and Zohar, Y. (1991). Antibacterial properties of human amniotic membranes. Placenta 12 (3), 285–288. doi:10.1016/0143-4004(91)90010-d
Tehrani, F. A., Ahmadiani, A., and Niknejad, H. (2013). The effects of preservation procedures on antibacterial property of amniotic membrane. Cryobiology 67 (3), 293–298. doi:10.1016/j.cryobiol.2013.08.010
Tehrani, F. A., Modaresifar, K., Azizian, S., and Niknejad, H. (2017). Induction of antimicrobial peptides secretion by IL-1β enhances human amniotic membrane for regenerative medicine. Sci. Rep. 7 (1), 17022. doi:10.1038/s41598-017-17210-7
Thadepalli, H., Appleman, M. D., Maidman, J. E., Arce, J. J., and Davidson, E. C. (1977). Antimicrobial effect of amniotic fluid against anaerobic bacteria. Am. J. Obstet. Gynecol. 127 (3), 250–254. doi:10.1016/0002-9378(77)90463-x
Tian, K., Yang, S., Ren, Q., Han, Z., Lu, S., Ma, F., et al. (2010). p38 MAPK contributes to the growth inhibition of leukemic tumor cells mediated by human umbilical cord mesenchymal stem cells. Cell. Physiol. biochem. 26 (6), 799–808. doi:10.1159/000323973
Touboul, C., Lis, R., Al Farsi, H., Raynaud, C. M., Warfa, M., Althawadi, H., et al. (2013). Mesenchymal stem cells enhance ovarian cancer cell infiltration through IL6 secretion in an amniochorionic membrane based 3D model. J. Transl. Med. 11 (1), 28. doi:10.1186/1479-5876-11-28
Vaught, D. B., Stanford, J. C., and Cook, R. S. (2015). Efferocytosis creates a tumor microenvironment supportive of tumor survival and metastasis. Cancer Cell Microenviron. 2 (1), e666. doi:10.14800/ccm.666
Wang M, M., Cai, J., Huang, F., Zhu, M., Zhang, Q., Yang, T., et al. (2015). Pre-treatment of human umbilical cord-derived mesenchymal stem cells with interleukin-6 abolishes their growth-promoting effect on gastric cancer cells. Int. J. Mol. Med. 35 (2), 367–375. doi:10.3892/ijmm.2014.2019
Wang W, W., Zhong, W., Yuan, J., Yan, C., Hu, S., Tong, Y., et al. (2015). Involvement of Wnt/β-catenin signaling in the mesenchymal stem cells promote metastatic growth and chemoresistance of cholangiocarcinoma. Oncotarget 6 (39), 42276–42289. doi:10.18632/oncotarget.5514
Wang, X., Xie, J., Tan, L., Huo, J., and Xie, H. (2012). A new vision of definition, commentary, and understanding in clinical and translational medicine. Clin. Transl. Med. 6 (21), 5. doi:10.1186/2001-1326-1-5
Wiegand, I., Hilpert, K., and Hancock, R. E. (2008). Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 3 (2), 163–175. doi:10.1038/nprot.2007.521
Wilson, I. D., and Nicholson, J. K. (2017). Gut microbiome interactions with drug metabolism, efficacy, and toxicity. Transl. Res. 179, 204–222. doi:10.1016/j.trsl.2016.08.002
Wolbank, S., Hildner, F., Redl, H., van Griensven, M., Gabriel, C., and Hennerbichler, S. (2009). Impact of human amniotic membrane preparation on release of angiogenic factors. J. Tissue Eng. Regen. Med. 3 (8), 651–654. doi:10.1002/term.207
Wu, M., Yu, Z., Matar, D. Y., Karvar, M., Chen, Z., Ng, B., et al. (2022). Human amniotic membrane promotes angiogenesis in an oxidative stress chronic diabetic murine wound model. Adv. Wound Care (New Rochelle) [Epub ahead of print]. doi:10.1089/wound.2022.0005
Wu, S., Ju, G. Q., Du, T., Zhu, Y. J., and Liu, G. H. (2013). Microvesicles derived from human umbilical cord Wharton's jelly mesenchymal stem cells attenuate bladder tumor cell growth in vitro and in vivo. PLoS One 8 (4), e61366. doi:10.1371/journal.pone.0061366
Yadav, M. K., Go, Y. Y., Kim, S. H., Chae, S.-W., and Song, J.-J. (2017). Antimicrobial and antibiofilm effects of human amniotic/chorionic membrane extract on Streptococcus pneumoniae. Front. Microbiol. 8, 1948. doi:10.3389/fmicb.2017.01948
Yan, C., Li, S., Li, Z., Peng, H., Yuan, X., Jiang, L., et al. (2013). Human umbilical cord mesenchymal stem cells as vehicles of CD20-specific TRAIL fusion protein delivery: A double-target therapy against non-hodgkin's lymphoma. Mol. Pharm. 10 (1), 142–151. doi:10.1021/mp300261e
Yang, C., Lei, D., Ouyang, W., Ren, J., Li, H., Hu, J., et al. (2014a). Conditioned media from human adipose tissue-derived mesenchymal stem cells and umbilical cord-derived mesenchymal stem cells efficiently induced the apoptosis and differentiation in human glioma cell lines in vitro. Biomed. Res. Int. 2014, 109389. doi:10.1155/2014/109389
Yang, X., Li, Z., Ma, Y., Gao, J., Liu, S., Gao, Y., et al. (2014b). Human umbilical cord mesenchymal stem cells promote carcinoma growth and lymph node metastasis when co-injected with esophageal carcinoma cells in nude mice. Cancer Cell Int. 14 (1), 93. doi:10.1186/s12935-014-0093-9
Yuan L, L., Liu, Y., Qu, Y., Liu, L., and Li, H. (2019). Exosomes derived from MicroRNA-148b-3p-overexpressing human umbilical cord mesenchymal stem cells restrain breast cancer progression. Front. Oncol. 9, 1076. doi:10.3389/fonc.2019.01076
Yuan, Y., Lu, X., Tao, C. L., Chen, X., Shao, H. W., and Huang, S. L. (2013). Forced expression of indoleamine-2, 3-dioxygenase in human umbilical cord-derived mesenchymal stem cells abolishes their anti-apoptotic effect on leukemia cell lines in vitro. Vitro Cell. Dev. Biol. -Animal. 49 (10), 752–758. doi:10.1007/s11626-013-9667-4
Yuan, Y., Zhou, C., Chen, X., Tao, C., Cheng, H., and Lu, X. (2018). Suppression of tumor cell proliferation and migration by human umbilical cord mesenchymal stem cells: A possible role for apoptosis and wnt signaling. Oncol. Lett. 15 (6), 8536–8544. doi:10.3892/ol.2018.8368
Yuan Z, Z., Bian, Y., Ma, X., Tang, Z., Chen, N., and Shen, M. (2019). LncRNA H19 knockdown in human amniotic mesenchymal stem cells suppresses angiogenesis by associating with EZH2 and activating vasohibin-1. Stem Cells Dev. 28 (12), 781–790. doi:10.1089/scd.2019.0014
Zhang, J., Huang, J., Lin, X., Fei, K., Xie, Y., Peng, Q., et al. (2022). Phosphoglycerate mutase 5 promotes necroptosis in trophoblast cells through activation of dynamin-related protein 1 in early-onset preeclampsia. Am. J. Reprod. Immunol. 87 (6), e13539. doi:10.1111/aji.13539
Zhang, W., Yang, H., Zhang, Y., Lu, Y., Zhou, T., Li, M., et al. (2015). A biotherapy based on PSCs-in-3D spheroid-ameliorated biologics depletes in vivo cancer-sustaining stem cells. Oncotarget 6, 40762–40774. doi:10.18632/oncotarget.5691
Zhang, X., Yang, Y., Zhang, L., Lu, Y., Zhang, Q., Fan, D., et al. (2017). Mesenchymal stromal cells as vehicles of tetravalent bispecific Tandab (CD3/CD19) for the treatment of B cell lymphoma combined with Ido pathway inhibitor D-1-methyl-tryptophan. J. Hematol. Oncol. 10 (1), 56. doi:10.1186/s13045-017-0397-z
Keywords: perinatal derivatives, biological assays, functional assays, potency assays, mechanisms of action, pharmacologic actions, cancer, infections
Citation: Silini AR, Ramuta TŽ, Pires AS, Banerjee A, Dubus M, Gindraux F, Kerdjoudj H, Maciulatis J, Weidinger A, Wolbank S, Eissner G, Giebel B, Pozzobon M, Parolini O and Kreft ME (2022) Methods and criteria for validating the multimodal functions of perinatal derivatives when used in oncological and antimicrobial applications. Front. Bioeng. Biotechnol. 10:958669. doi: 10.3389/fbioe.2022.958669
Received: 31 May 2022; Accepted: 26 September 2022;
Published: 13 October 2022.
Edited by:
Karthikeyan Narayanan, Rensselaer Polytechnic Institute, United StatesReviewed by:
Amudha Ganapathy, University of Illinois at Chicago, United StatesCopyright © 2022 Silini, Ramuta, Pires, Banerjee, Dubus, Gindraux, Kerdjoudj, Maciulatis, Weidinger, Wolbank, Eissner, Giebel, Pozzobon, Parolini and Kreft. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mateja Erdani Kreft, bWF0ZWphLmVyZGFuaUBtZi51bmktbGouc2k=
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.