
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Allergy , 20 March 2025
Sec. Environmental & Occupational Determinants
Volume 6 - 2025 | https://doi.org/10.3389/falgy.2025.1520576
 Fikirte Debebe Zegeye1,2
Fikirte Debebe Zegeye1,2 Pål Graff3
Pål Graff3 Miriam Grgic4
Miriam Grgic4 Steen Mollerup1
Steen Mollerup1 Anani Komlavi Afanou1Berit Elisabeth Bang4,5
Anani Komlavi Afanou1Berit Elisabeth Bang4,5 Karl-Christian Nordby6
Karl-Christian Nordby6 Anne Straumfors1
Anne Straumfors1 Johanna Samulin Erdem1*
Johanna Samulin Erdem1*
Introduction: Shellfish processing workers are highly susceptible to respiratory illnesses such as allergies and asthma. This study examined respiratory symptoms and biomarkers of allergy and asthma in Norwegian shrimp processing plant workers and evaluated allergenic and irritant protein exposures in the workplace.
Material and methods: The study included 35 shrimp processing workers and 21 controls. Respiratory symptoms were assessed via questionnaire; blood samples were analysed for allergy and asthma biomarkers and specific IgE levels. Air samples were analysed for protein levels and composition.
Results: Shrimp processing workers had four to five times higher odds of reporting acute upper and chronic lower respiratory symptoms than the controls. They also had significantly higher plasma levels of IL4, CCL20, CSF2 and MMP12, with 11% of the exposed workers showing elevated levels of shrimp and crab specific IgE. Furthermore, exposed workers showed increased plasma levels of SFTPD and CHI3L1 post-shift. The median total protein exposure was 6 µg/m3, with peaks up to 66 µg/m3 in the cooking and peeling department. Total protein levels were correlated with CCL20, IL13, and basophil counts. Ninety-five shrimp proteins were identified, including seven known and eight potential allergens. Tropomyosin levels were generally high, particularly in the cooking and peeling department.
Conclusion: Shrimp workers had a higher prevalence of respiratory symptoms and biomarkers of allergy and asthma. The work environment contained tropomyosin and other allergenic proteins as well as irritants, highlighting the need for protective measures, especially in the cooking and peeling departments.
The seafood industry constitutes a large part of the Norwegian food industry, and seafood production has hit a record export volume of 2.9 million tons of seafood to a value of (€13,4 bn/£11.8 bn) in 2022 (Norwegian Seafood Council).
Globally, over 61 million individuals work in the seafood industry, and respiratory health effects such as allergies and asthma are notably prevalent among these workers (1–3). The prevalence of occupational rhinitis and occupational asthma among these workers ranges from 2% to 36% (2, 4, 5). Workers in the shellfish industry have a higher frequency and severity of respiratory symptoms and are more often sensitised than those in the fish industry (2, 6).
The seafood industry is characterised by complex exposure to chemicals and biological factors (bioaerosols). These bioaerosols include allergens, proteases, microorganisms and endotoxins (6–11). Bioaerosol exposures occur primarily through inhalation of dust, steam, vapour, and aerosolised proteins generated during butchering, thawing, cooking or boiling, peeling, and grinding (12, 13). Thermal processes such as thawing and boiling can potentially accelerate the evaporating process, leading to the formation of exposure hot spots for allergenic proteins with enhanced allergenicity (13).
Tropomyosin and arginine kinase are the two most extensively studied allergenic proteins in shellfish (14, 15). However, over the past decade, several additional allergens have been identified, including sarcoplasmic calcium-binding, myosin light chain, hemocyanin, pyruvate kinase, and enolase (7, 14–18). Exposure to these proteins may lead to occupational rhinitis and asthma, commonly through IgE-mediated allergic reactions (16, 18). Moreover, chitin, which is a biopolymer of N-acetyl-b-D-glucosamine, in the exoskeleton of shellfish, insects, and fungi has been shown to induce inflammation through the production of reactive oxygen species (ROS) in humans, and induction of chitinase in the murine lung that could lead to the development of bronchial hyperreactivity in an IgE independent manner (19–21). Further, proteases such as trypsin may trigger inflammation, respiratory tract hyperresponsiveness and possibly IgE-mediated sensitisation upon inhalation (22–24). Moreover, king crab and salmon trypsin are potent stimulators of protease-activated receptors causing pro-inflammatory responses in pulmonary epithelial cell models (23, 25–27).
Exposure to bioaerosols can trigger hypersensitivity reactions in the respiratory tract, potentially leading to decreased lung function, occupational asthma, and occupational rhinitis (28, 29). An immunologically mediated sensitisation primarily induces occupational asthma, but non-immunologic and irritant-induced cases have also been reported (28, 30). In immunologically mediated sensitisation, several chemokines and cytokines are involved, with the top three important cytokines being interleukin 4 (IL4), IL5 and IL13 (31, 32).
To date, studies of occupational exposure and respiratory effects in shrimp processing plants are scarce. However, among crustaceans, shrimp is the most common elicitor of allergic reactions. The northern shrimp (Pandalus borealis) is one of the least studied in terms of allergenic protein profiles and health impacts associated with occupational exposure during processing. Therefore, this study aimed to evaluate the prevalence of respiratory symptoms in two Norwegian shrimp processing plants and assess specific IgE and biomarker levels in the blood of exposed workers. Additionally, the study sought to characterise the protein composition of air samples collected from different work processes in the processing plants, with a focus on identifying allergenic proteins and assessing potential respiratory health effects associated with these proteins.
A cross-sectional study was conducted on workers in two Norwegian shrimp processing plants. The processing plants operate two to three shifts per day, five days a week. The inclusion criteria for the study were to be employed at one of the two selected shrimp processing plants during the study period (May and June 2022) and age above 18 years. The study was approved by the Regional Committee for Medical and Health Research Ethics (384542).
A statistical power analysis was performed for a dichotomous endpoint, aiming for 80% power at a 0.05 significance level using an occupational asthma prevalence of 36% in the shellfish industry (for the exposed group) and a current asthma prevalence of 5.4% in the general population (for unexposed control) (2, 33). The analysis concluded that the study should include at least 35 exposed workers and 18 unexposed control workers. Accordingly, all exposed workers in the shrimp processing plants (n = 65) were invited with a letter informing them about the study and its objectives. Of these, a total of 35 exposed production workers in shrimp processing plants participated in one or more parts of the study. Anonymised data from 21 administrative personnel in waste sorting plants were included as controls under ethical approval number 34312. These workers shared a similar sociodemographic background and had no prior experience in the seafood industry. Written informed consent for questionnaires and blood sampling was obtained from the study participants. Blood samples were collected from the exposed workers at two different times, pre-shift and post-shift, while a single sample was taken from the control workers. A total of 30 personal air samples and 2 stationary air samples were collected.
All employees were asked to complete a questionnaire that gathered personal information, including age, sex, height, and weight. Additionally, it inquired about their smoking or snuffing habits and whether they had been diagnosed with asthma, allergy, eczema, or experienced acute and chronic respiratory symptoms (Supplementary Data 1).
A total of 28 exposed workers and 21 control workers consented to give blood samples. Of the exposed workers, 25 workers supplied two blood samples, one before and one after shift, to examine acute health effects. The remaining exposed workers were unwilling or unable to provide blood samples after the shift. All the control workers provided one blood sample. Blood samples were collected using EDTA (BD Vacutainer K2E, BD, USA) and serum tubes (BD Vacutainer SST II Advance, BD, USA). Following the blood collection, the EDTA tubes were inverted 8–10 times and centrifuged for 10 min at 2,100 × g to separate plasma from blood cells, while the serum tubes were left to coagulate for 30 min at room temperature and then centrifuged at 2,100 × g for 5 min. Plasma samples were aliquoted and stored at – 80°C until further analysis. The whole blood and serum were sent to an accredited laboratory, Fürst Medicine Laboratory (Oslo, Norway), for differential counts of white blood cells (WBC) and analysis of specific IgE antibodies against salmon, crab, and shrimp, using the sandwich immune assay ImmunoCAPTM (Thermo Fisher Scientific, Waltham, Massachusetts, USA). The three species were selected due to their potential cross-reactivity. While fish allergens typically do not cross-react with shellfish, a previous study showed that tilapia's tropomyosin allergen shared 53.5% homology with shrimp tropomyosin (34). The standard clinical reference range for WBC analysis were total leukocytes (3.5–10 × 109/L), neutrophils (1.5–7.3 × 109/L), lymphocytes (1.1–3.3 × 109/L), monocytes (0.2–0. 8 × 109/L), eosinophil (<0.4 × 109/L) and basophil (<0.2 × 109/L). The normal reference range for the IgE levels was <0.35 KU/L. Workers who had values exceeding these ranges were reported in percentages.
Biomarkers of allergy and asthma were measured in plasma samples using a custom multiplex human Luminex discovery assay (R&D Systems, Minneapolis, MN, USA). The custom multiplex assay included C-C motif chemokine ligand 17 (CCL17), CCL20, CCL26, IL4, IL5, IL13, IL33, chitinase 3-like 1 (CHI3L1), matrix metalloproteinase 12 (MMP12), periostin (POSTN), Surfactant Protein - D (SFTPD), thymic stromal lymphopoietin (TSLP) and colony-stimulating factor 2 (CSF2) with a sensitivity ranging from 0.5 to 95.7 pg/ml for the different biomarkers. These biomarkers were selected based on their key roles in the Th2 immune response, crucial in asthma and allergy pathophysiology. Chemokines CCL17, CCL20, and CCL26 attract immune cells to inflamed sites, enhancing allergenic reactions (35, 36). Cytokines IL4, IL5, IL13, and IL33 support IgE production, eosinophil activation, and airway hyperresponsiveness (31, 32, 37, 38). CHI3L1 and MMP12 mediate airway tissue remodelling and inflammation (39–41). While POSTN exacerbates inflammation by regulating cellular interactions and supporting tissue integrity, SFTPD protects lung tissue by modulating immune responses (42, 43). TSLP initiates allergic reactions through dendritic cell activation, and CSF2 regulates inflammation by promoting granulocyte and macrophage production (32, 35, 37, 38).
The plasma samples were diluted 1:2 and analysed according to the manufacturer's instructions. All samples were analysed in parallel using the xMAP INTELLIFELEX instrument (R&D Systems). Biomarker concentrations in the plasma samples were estimated using a six-point standard curve with the Quantist data analysis software version 1.0.1.0 (R&D Systems). IL33 and TSLP levels were below the limit of detection (LOD) and were consequently excluded from further analysis.
Active air sampling was performed using a Conical Inhalable Sampler (CIS) (Casella Solutions, Kempston, UK) attached to Casella Apex2 pumps (Casella UK, Wolseley Rd, Kempston, Bedford, UK) over two consecutive days. The sampler was loaded with a 37 mm polytetrafluorethylene (PTFE) filter (1.0 µm, Millipore, Billerica, MA, USA). The airflow was set to 3.5 L/min following calibration with a digital flow meter Defender 520 M (Defender; SKC Inc., Eighty-Four, PA, USA).
Workers who consented to participate in the personal air sampling campaign carried the samplers in a small backpack, positioning the sampling cassettes in their personal breathing zone for the entire 8–9 h work shift. Sampling was done across five different work processes: (1) Thawing—thawing frozen shrimp blocks (2) Truck Driving—transporting frozen shrimp to thawing, maturation and subsequently to the cooking area. Thawers and truck drivers work in the same department but perform different tasks. (3) Technician—operating machinery for cooking and peeling the shrimp (4) Packing—conducting quality control and performing automated packing of shrimp, and (5) Flour production—drying and grinding shrimp shells to produce flour.
In addition to the personal air samplers, one stationary sampler was included in the cooking and peeling area per processing plant. The stationary samplers were equipped with PTFE filters and were placed in the cooking and peeling department. Sampling was performed for 11–23 h.
Proteins were extracted from the personal PTFE filters with 0.5% PBS-Tween 20 buffer. The samples were rotated for 4 h at 4℃ followed by centrifugation at 1,200 × g for 10 min. The supernatants were aliquoted and stored at −80℃ until further analyses of total protein, proteolytic activity, and measurements of allergen levels. Stationary samples for LC-MS/MS analysis were extracted using a 10 mM glycine/0.5% SDS buffer (pH 7.55). The samples were rotated for 16 h at 4℃ followed by centrifugation at 1,200 × g for 10 min. Supernatants were stored at −80℃ until proteomic analysis. The total protein levels were quantified using QuantiPro BCA Assay Kit (Sigma-Aldrich, St. Louis, USA) and measured with a Multiscan FC (Thermo Scientific) spectrophotometer at 560 nm according to the manufacturer's instructions.
Proteolytic activity was assessed using zymography, a sodium dodecyl sulfate-polyacrylamide electrophoresis impregnated with 0.1% gelatin substrate in a polyacrylamide gel. The gelatine degrades in the presence of protease, creating unstained regions on the gel as previously described (9). The protease activity was calculated based on a five-point standard curve (0.1–0.9 nM) from porcine trypsin with known enzyme activity. The gel image was captured using the BioDoc® Imaging system. The intensity of the degraded regions (unstained regions) was quantified using image analysis software (UVP Vision Works®, Analytik Jena GmbH + Co).
LC-MS analysis was performed on stationary samples collected in the cooking and peeling department to assess the protein composition of the bioaerosol samples. The analysis was conducted at the Proteomic and Metabolomics Core Facility (PRiME) at the Arctic University of Norway (UiT). The filter extract eluate was evaporated to dryness and resuspended in 2% SDS 50 mM TEA buffer and sonicated for 3 min in a water bath. Protein digestion was done following the S-trap micro protocol (protifi.com). The eluted peptide solution underwent evaporation and was subsequently reconstituted in 0.1% formic acid. The samples were analysed on an Easy nLC 1,200 instrument coupled to an Orbitrap Exploris 480 for LC-MS analysis.
Protein sequences were blasted in the National Center for Biotechnology Information (NCBI) protein BLAST database for identification. Only 85 proteins are available from the species Pandalus borealis in the reference database. Therefore, most identification was based on homology in the infraorder Caridea. A mass spectrometry contaminant database (MaConDa) was used to remove contaminant proteins from the samples.
Protein allergenicity prediction was performed using the AllerCatPro2.0 tool on proteins that had three or more peptides match (44). The tool identifies proteins with potential allergenicity by analysing their amino acid sequences and predicted 3D structure, comparing them against comprehensive allergen databases, and categorising them as strong, weak or no evidence for allergenicity.
Tropomyosin levels in the personal air samples were measured using a shrimp tropomyosin ELISA kit (Inbio, Charlottesville, VA, USA) with a 0.2 ng/ml limit of detection, according to the manufacturer's instructions. The tropomyosin concentration was determined using a 10-point standard curve (0.1–50 ng/ml). The absorbance was measured at 450 nm using Bio Tek Synergy Neo2 Hybrid Multimode Reader (Agilent Technologies, USA).
Arginine kinase was analysed by western blotting. Due to the low protein concentration, the samples for analysis of arginine kinase were pooled per work process. Concentrated protein extracts were separated by Any kD MP Mini protein TGX stain-free protein gradient (Biorad, Hercules, CA) SDS-PAGE and transferred to a Trans-Blot turbo PVDF membrane (Biorad). The membranes were incubated with Pierce™ protein-free blocking buffer (Thermo Fisher Scientific) for 1 h at room temperature to prevent non-specific background binding. The membranes were incubated overnight at 4°C with primary antibody against arginine kinase (Abcam, Cambridge, UK) at a concentration of 3.5 µg/ml. After washing three times with Tris-buffered saline with 0.05% Tween-20 (TBST), the membranes were incubated with a secondary antibody, HRP-conjugated anti-rabbit IgG secondary antibody (Cell Signalling Technology, MA, USA), for 1 h at room temperature. After incubation, the membranes were washed three times with TBST and incubated with Super signal west Atto ultimate sensitivity substrate (Thermo Fisher Scientific) for 5 min. Signals were captured using a CCD camera-based imager (Amersham Imager 600, UK). Densitometric analysis was performed by ImageJ 1.54 g, and the obtained values were adjusted for the total air volume per filter.
Statistical analysis was conducted using GraphPad Prism version 9 and R version 4.3.1. Power analysis was done using the pwr package by employing the pwr.2p2n.test function. Data sets that were not normally distributed, as determined by the Shapiro–Wilk test, or contained outliers identified by the Q-Q plot were log-transformed before analysis. This was followed by a sensitivity analysis when necessary. The log transformation specifics and analysis types are indicated in the footnotes of each result. Variations in age, years of employment, and body mass index (BMI) between groups were evaluated using an independent sample t-test. Differences in sex, smoking status and symptom prevalence between groups were assessed using Pearson Chi-square or Fisher's exact test. Multivariate logistic regression was used to evaluate the association between exposure status and the odds of reporting respiratory symptoms, adjusting for potential confounding variables. Multivariate linear regression was employed to examine the relationship between biomarker levels and exposure status, adjusting for potential confounding variables. The confounding variables were selected for both models based on their influence on the model using stepwise selection. The initial regression model included all potential confounders: age, sex, smoking status, BMI, length of employment, asthma, and allergy. Only variables that significantly impacted the dependent variable were retained, while others were excluded. Additionally, we assessed the associations between biomarker levels and acute and chronic respiratory symptoms using linear regression while accounting for potential confounding effects. p < 0.05 was considered significant. A paired sample t-test or Wilcoxon signed-rank test was used to compare biomarker levels in exposed production workers' blood before and after work. Comparisons of exposure levels between different workgroups were done using one-way ANOVA or the Kruskal–Wallis test. Pearson correlation was used to assess the correlation between biomarker levels, differential counts of WBC, total protein and previously reported endotoxin levels.1
The study population consisted of 56 workers, of whom 43 were men and 13 were women. Their ages ranged from 20 to 65, with a mean BMI of 26–28 (Table 1). Current and past smoking habits of the exposed workers and controls were similar. The comparison of the sociodemographic variables revealed no significant difference between the two groups (Table 1).
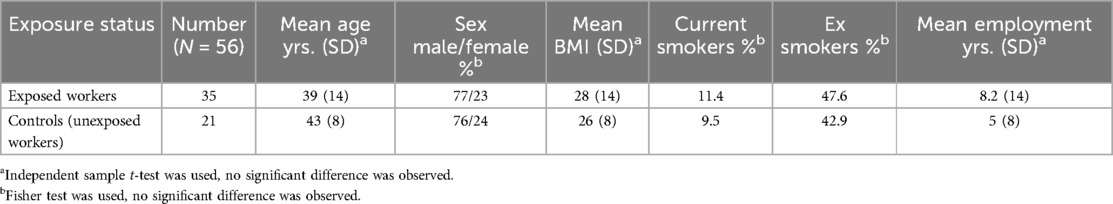
Table 1. Characteristics of the study population.
The prevalence of reported doctor-diagnosed asthma was more frequent in exposed workers (22.9%) than in controls (4.8%), but the difference was not statistically significant (Supplementary Table S1). In the past year, 2.8% of the exposed workers reported experiencing asthma attacks, and 5.7% reported using asthma medication, while no asthma attack or medicine use was reported among the control group. 11.4% of exposed workers and 4.7% of control were diagnosed with asthma as adults (Supplementary Table S1). Self-reported allergy was more frequent among controls (42.9%) than exposed workers (26.2%) (Supplementary Table S1). However, this was not statistically significant, and the reported allergies were predominantly grass or pollen allergies in both groups. In addition, 2.9% of exposed workers reported shrimp allergy (Supplementary Table S1).
All individual respiratory symptoms were relatively more frequent in exposed workers, ranging from 8.5% to 45.7%, compared to controls (0%–19%) (Table 2). Chronic lower and acute upper respiratory symptoms were significantly prevalent among the exposed workers compared to the controls (p = 0.02) (Table 2). Furthermore, the odds of having chronic lower respiratory symptoms were four times higher in exposed workers than in controls (p = 0.01) (Table 3). In addition, exposed workers had a five times higher likelihood of reporting acute upper respiratory symptoms than controls, with a significant p-value of 0.01 in a logistic model adjusted for smoking (Table 3). Although it was not statistically significant, acute lower respiratory symptoms were more than twice as frequent among exposed (37.1%) than control (14.3%) (Table 2).
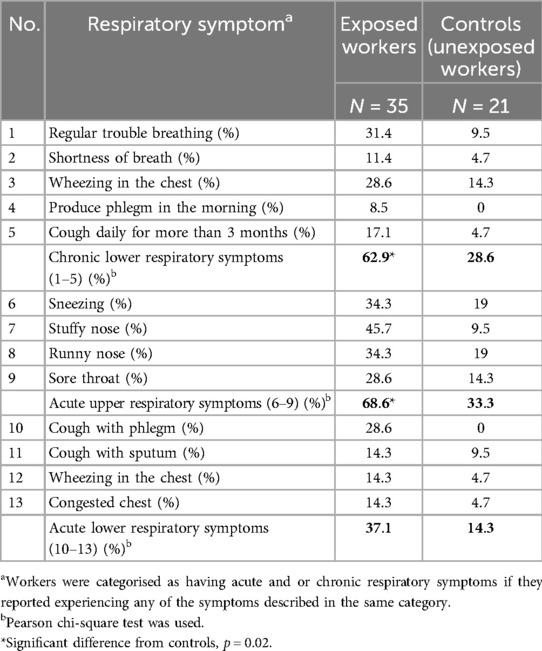
Table 2. Prevalence of self-reported respiratory symptoms.

Table 3. Association between exposure status and respiratory symptoms.
Elevated specific IgE levels against crab and shrimp allergens were detected in 11% of the exposed workers. An additional 3.7% of the exposed workers had elevated IgE against salmon (Supplementary Table S2). Furthermore, 7.1% of the exposed workers had elevated total leukocytes, lymphocytes, monocytes, and eosinophils. While the number of basophils was normal in all the production workers, 3.5% had neutrophil counts higher than the acceptable range (Supplementary Table S2).
Linear regression analysis revealed that, after adjusting for confounding factors, IL4, CCL20, MMP12 and CSF2 levels were significantly higher in exposed workers compared to the controls, whereas CCL17 levels were significantly lower than controls (Table 4). Being in the exposed group was associated with a higher level of IL4 with an estimate of 8.5 pg/ml (95% CI: 1.5–15.5), p = 0.02; CCL20 with an estimate of 2.3 pg/ml (95% CI: 1.3–4.3), p = 0.01; MMP12 with an estimate of 3.1 pg/ml (95% CI: 1.2–8.1), p = 0.02; and CSF2 with an estimate of 1.4 pg/ml (95% CI: 1.0–1.9), p = 0.03 (Table 4). On the contrary, being in the exposed group was associated with a lower level in CCL17 with an estimate of 0.7 pg/ml (95% CI: 0.6–0.9), p = 0.01 (Table 4). No effects were observed for IL5, IL13, CCL26, SFTPD, CHI3L1, and POSTN. Sex, age and self-reported allergy were identified as influential covariates for three or more biomarkers, whereas smoking status did not influence any of the biomarkers tested (Table 4). Asthma and allergies were significantly associated with CCL20 and CCL17, p < 0.03, respectively (Table 4). Raw data is available in the (Supplementary Data 2).
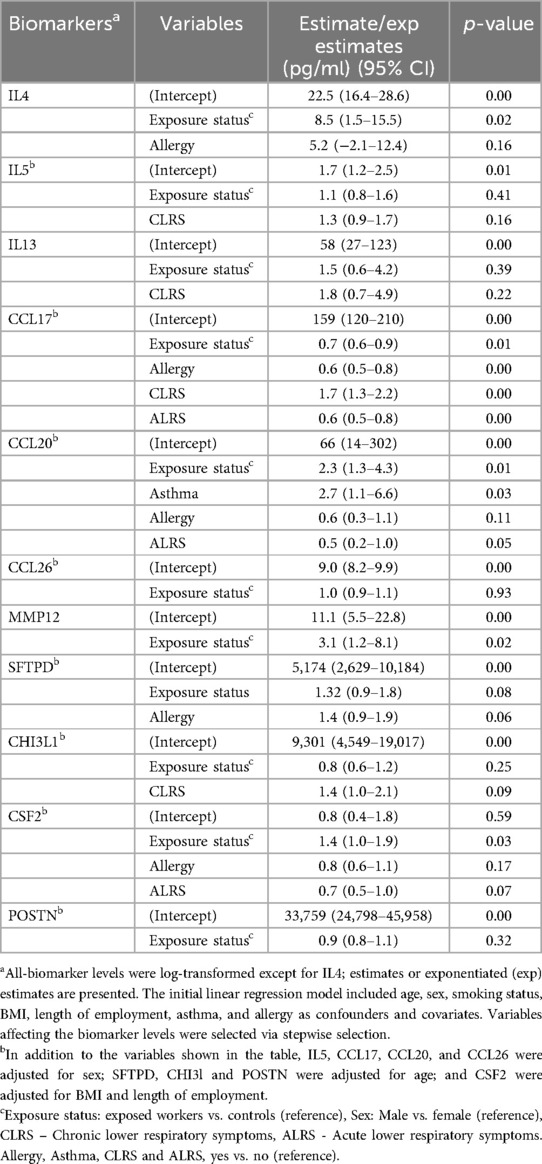
Table 4. Linear regression analysis of allergy and asthma biomarker levels in workers’ plasma.
Assessment of the associations between biomarker levels and respiratory symptoms showed that chronic lower respiratory symptoms were significantly associated with a higher level of CCL17 with an estimate of 1.7 pg/ml (95% CI: 1.3–2.2), p < 0.001. Acute lower respiratory symptoms were associated with lower CCL17 [estimate: 0.6 pg/ml, (95% CI: 0.5–0.8), p < 0.001] and a borderline significant lower level of CCL20 [estimate: 0.5 pg/ml (95% CI: 0.2–1.0), p = 0.05] (Table 4). No associations were found between acute upper respiratory symptoms and the analysed biomarkers.
A comparison of biomarker levels before and after work to assess the “acute effect” among exposed workers using a paired t-test showed a significant increase in the levels of SFTPD and CHI3L1, with a mean difference of 1.2 pg/ml (95% CI: 1.1–1.2), p < 0.001 and 1.0 pg/ml (95% CI: 1.0 to−1.1), p = 0.03 after the work shift, respectively. No effects were observed for the remaining biomarkers. Moreover, no differences were observed between workers performing different work tasks.
The total protein concentrations in the collected personal samples ranged from LOD to 66.2 µg/m3, with a median level of 6 µg/m3. The highest individual exposure was measured among the technicians, who also had overall higher protein exposure levels than the packers (p = 0.001) (Figure 1). Furthermore, the highest median exposure (23.4 µg/m3) was measured among flour production workers, while packers had the lowest median exposure (2 µg/m3) (Figure 1). Analyses of protein composition in the stationary collected aerosol samples resulted in the identification of 95 shrimp proteins. Of these, seven were known allergens: tropomyosin, arginine kinase, sarcoplasmic calcium-binding protein, troponin C, triosephosphate isomerase, myosin light chain and hemocyanin (Table 5). In addition, eight potential novel shrimp allergens were identified (Table 6). Of these, alpha-tubulin, heat shock protein 70, pyruvate kinase, troponin T, glyceraldehyde 3-phosphate dehydrogenase and myosin heavy chain showed strong evidence for allergenicity in the AllerCatPro2.0 prediction tool, whereas chitinase and vitellogenin demonstrated weak evidence of allergenicity. Detailed identification information is shown in Supplementary Table S3.
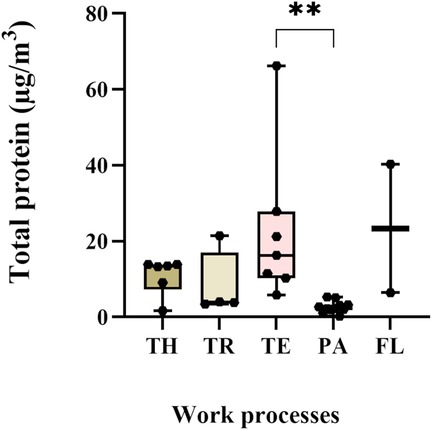
Figure 1. Exposure levels of total protein in personal samples. TH-thawers (n = 6), TR-truck drivers (n = 4), TE-technicians (n = 7), PA-packers (n = 11), and FL- flour production workers (n = 2). Group comparisons were done on log-transformed data using the Kruskal–Wallis test, followed by Dunn's multiple comparison test.
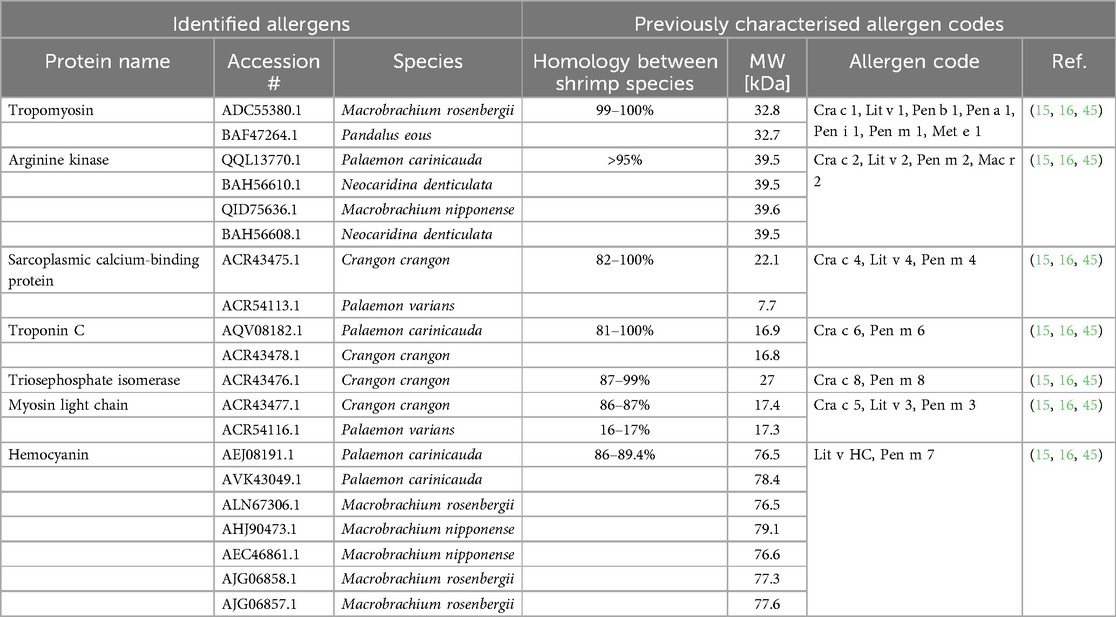
Table 5. Identified known shellfish allergens in stationary aerosol samples and their corresponding allergen codes.
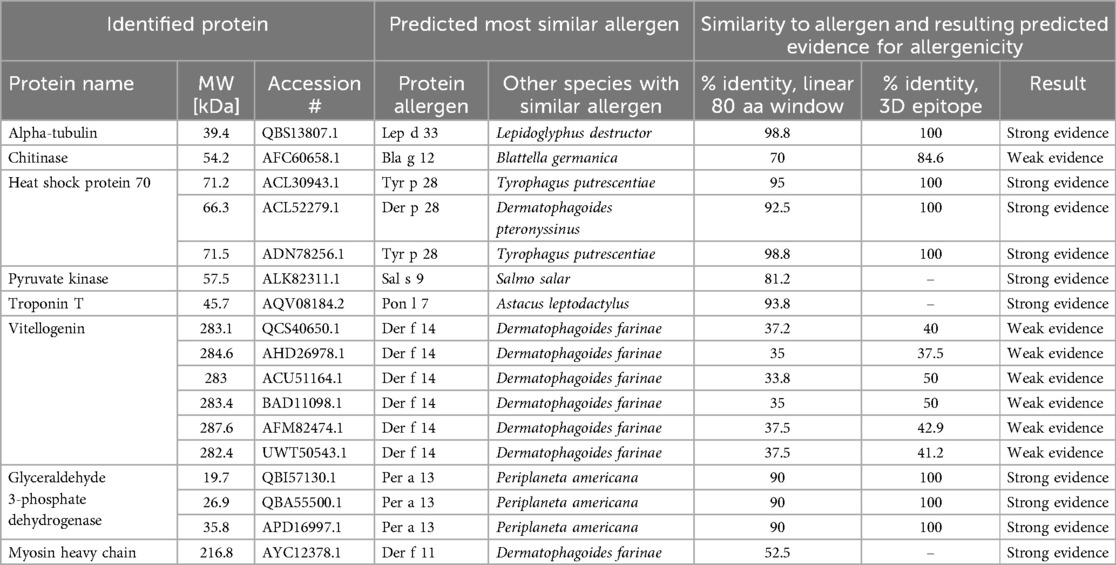
Table 6. Novel allergens identified in stationary aerosol samples collected from the cooking and peeling department using AllerCatPro2.
The major allergen in shrimp, tropomyosin, was detected in personal samples in all work processes, with a median exposure level of 300 ng/m3 with a range of 62–7,070 ng/m3 (Figure 2A). The highest median tropomyosin level (1,928 ng/m3), measured among the technicians, was significantly higher than among thawers, truck drivers, and packers. The lowest median tropomyosin level was measured in the flour production (240 ng/m3) (Figure 2A). The tropomyosin levels were moderately correlated with the measured total protein levels (r = 0.63, p < 0.001) (Figure 3). In addition to tropomyosin, levels of arginine kinase, a known shrimp allergen, were semi-quantified by western blot. The analyses identified a truncated form of arginine kinase (∼30 kDa) in all five work processes (Figure 2B). The observed truncation or partial degradation of the protein was likely due to the industrial process itself or the post-sampling treatments, resulting in a protein smaller than the estimated molecular weight of arginine kinase (39–45 kDa). The result demonstrated that arginine kinase was present in all work processes. The exposure level of arginine kinase was higher among truck drivers and technicians, showing a 15-fold and 7-fold increase compared to the thawing, respectively (Figure 2C, Supplementary Figure S1).
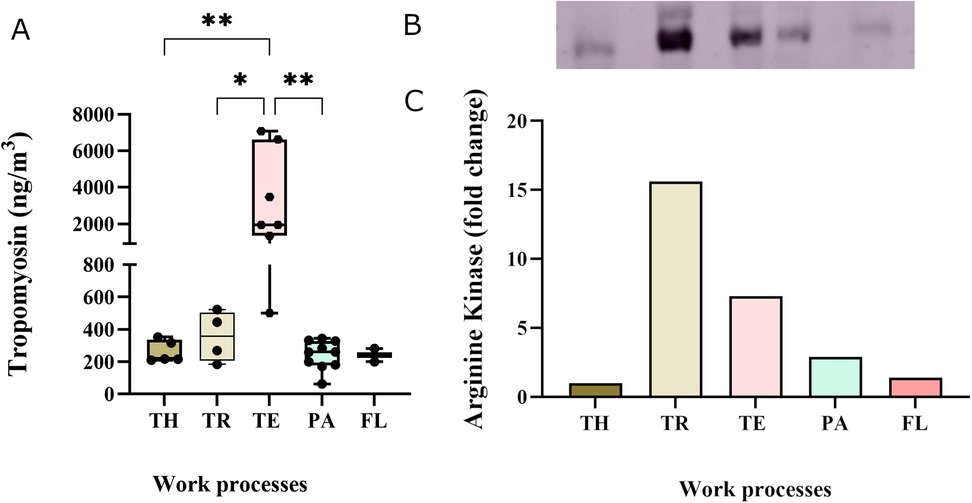
Figure 2. Exposure levels of allergenic proteins in personal aerosol samples across the five work processes: TH-thawers, TR-truck drivers, TE-technicians, PA-packers, and FL- flour production workers. (A) Tropomyosin levels TH (n = 6), TR (n = 4), TE (n = 7), PA (n = 11), and FL (n = 2). Group comparisons were done using one-way ANOVA, followed by Tukey's test. (B) Semi-quantitative analysis of arginine kinase levels by western blot, with bands representing samples pooled from each work process. (C) Graph showing the foldchange in arginine kinase concentration in relation to the thawing process.
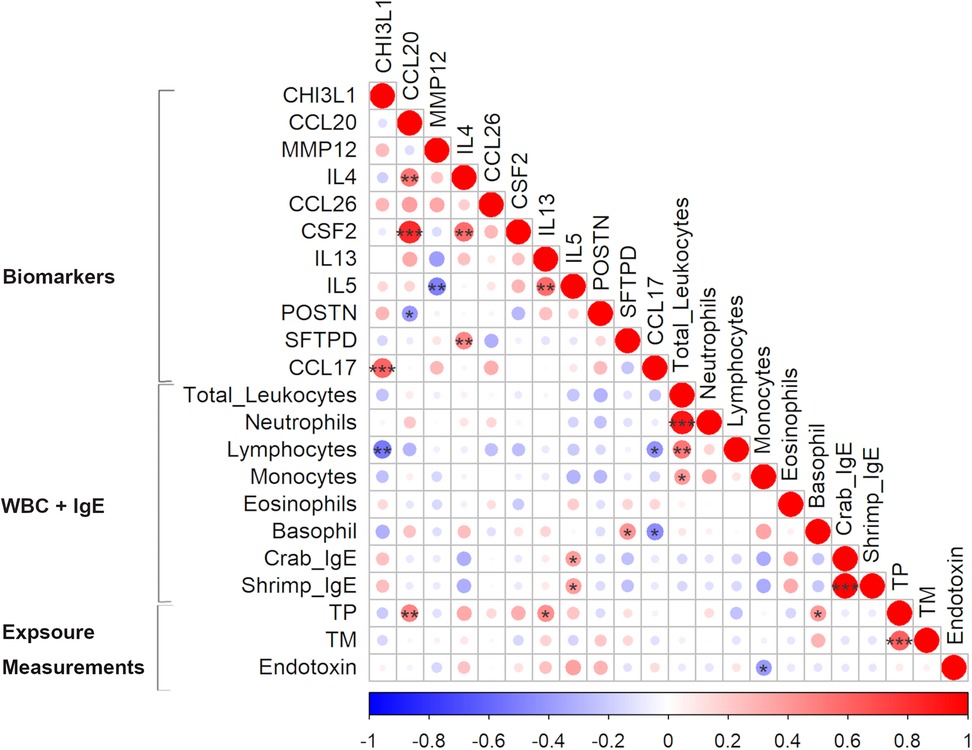
Figure 3. Pearson correlation matrix of biomarkers levels, white blood cell (WBC) count and exposure measurements from personal air samples (TP - total protein, TM—tropomyosin, and endotoxin) in exposed workers. Positive and negative correlations are indicated in red and blue, respectively. The colour intensity and size of the circles represent the strength of the correlations: larger and darker shades indicate a stronger correlation. *p < 0.05, **p < 0.01, ***p < 0.001.
Protease activity was detected in 25% of the aerosol samples, predominantly among the thawers and truck drivers. While the observed band sizes were inconsistent with trypsin (Supplementary Figure S2A), one of the detected proteases was verified to be a metalloprotease (Supplementary Figure S2B,C).
Correlation analysis showed that some exposure levels were correlated with a few of the biomarker levels. While tropomyosin levels did not correlate with any of the measured biomarkers, total protein was significantly correlated with CCL20 (r = 0.48, p = 0.01), IL13 (r = 0.44, p = 0.02) and basophil count (r = 0.39, p = 0.04) (Figure 3). In addition to the analysed protein levels, endotoxin levels are commonly measured to assess bioaerosol exposure in seafood industries. The measured endotoxin levels, ranging from LOD to 98 EU/m3, were included in the correlation analysis (see text footnote 1). A negative correlation between the measured endotoxin and monocyte count (r = −0.4, p = 0.04) was found and a positive borderline significant correlation with IL5 (r = 0.36, p = 0.05). Furthermore, the levels of crab IgE and shrimp IgE were strongly correlated (r = 0.99, p < 0.001) (Figure 3).
This study provides insight into the complex interaction between respiratory symptoms, biomarker levels in workers' blood, exposure to allergenic proteins, and irritants. The study showed that exposed workers reported more respiratory symptoms than controls. Particularly, acute upper respiratory symptoms and chronic lower respiratory symptoms were significantly prevalent among exposed workers compared to controls. In addition, exposed workers were four to five times more likely to report acute upper respiratory symptoms and chronic lower respiratory symptoms than controls. Similarly, workers in the shrimp, fish and crab industry frequently report symptoms of wheezing, shortness of breath and prolonged cough (8, 29, 46). In the present study, 22.9% of the exposed workers reported that they were diagnosed with asthma, and 11.4% of these workers were diagnosed with asthma as adults. This finding is comparable with a recent study by Laustsen and colleagues that reported 22.9% asthma incidents with 5.2% probable occupational asthma in the crab and shrimp industry (47). Occupational asthma is more frequently associated with workers in the shellfish processing industry than with the bony fish industry. Among the shellfish processing workers, the highest prevalence of occupational asthma (36%) has been reported in prawn processing plants (2). In the present study, 26.2% of the exposed workers reported having an allergy. However, most were related to pollen or grass, except for 2.9%, which reported shrimp allergy. Furthermore, 11% of the exposed workers exhibited elevated IgE levels against shrimp and crab in a specific allergy test. Furthermore, the IgE levels of shrimp and crab were strongly correlated, underscoring their cross-reactivity and potential sensitisation among the same workers (16, 48). Similarly, previous studies on shrimp processing workers in Greenland and Norway reported 10.1% and 20.3% of the workers, respectively, had elevated IgE levels against shrimp (8, 47). The improvement in the reported frequency of sensitisation in our study and the study by Laustsen et al. can likely be attributed to the continuous effort by companies to reduce exposure. In addition, individuals diagnosed with these allergies often seek alternative employment, resulting in a healthy worker effect, which may potentially lead to an underestimation of the exposure-related effects on symptom prevalence.
In addition to the respiratory symptoms reported in the present study, differential blood counts showed that 7.1% of the exposed workers exhibited higher levels of total leukocytes, lymphocytes, monocytes, and eosinophils.
To our knowledge, this study is the first to analyse the levels of allergy and asthma biomarkers in exposed workers in this industry, comparing them to controls, thereby offering new insight into the immune response in exposed workers. Our study showed that exposed workers exhibited significantly higher levels of IL4 relative to controls. This finding indicates an enhanced allergenic response, which could be caused by exposure to allergenic proteins such as tropomyosin (31, 32, 49). Additionally, the data shows substantially higher levels of CCL20, CSF2, and MMP12 among exposed workers than controls. These biomarkers are known to be involved in inflammation, asthma, and allergy pathways, further supporting the notion that exposure to allergenic proteins and irritants in shrimp processing plants may exacerbate allergenic and inflammatory reactions (35, 37, 50). CCL17 levels were lower among exposed workers and workers with allergies, and those who reported acute upper respiratory symptoms while reported chronic lower respiratory symptoms were associated with higher CCL17 levels. Similarly, increased CCL17 levels have been previously reported in the serum of patients with allergic asthma (51), while CCL17 levels were unaltered in patients with allergic rhinitis. The observed increase in CCL17 levels is likely related to the persistent inflammation that characterises chronic respiratory conditions, where elevated CCL17 levels help recruit immune cells to sustain and modulate the ongoing inflammatory response (35) Thus, while CCL17 is important in allergic inflammation, its expression in plasma may be affected by disease stage and severity. Altogether, this indicates a need for more research to evaluate the suitability of CCL17 as a biomarker of acute vs. chronic occupational respiratory symptoms.
Overall, the observed levels of these biomarkers and the increased WBC indicate that exposed workers have a higher incidence of immunological and allergic responses. Furthermore, exposed workers had significantly elevated levels of SFTPD and CHI3L1 following a work shift, showing an acute inflammatory response to the occupational exposure. These two markers have been implicated in the first line of defence and inflammatory response and tissue remodelling (39, 43, 52). Thus, the significant elevation of SFTPD and CHI3L1 levels post-shift might suggest the initiation of allergenic rhinitis following bioaerosol exposures, which may be linked to the reported acute upper respiratory symptoms (41).
While the variation in biomarker levels provides valuable insights into the physiological responses of workers, it is equally important to examine the types and levels of exposure in this work environment. Assessing these, along with the biomarkers, can help establish a potential link to respiratory symptoms. Total protein measurements are often used in the seafood industry. In the present study, up to 66.2 µg/m3 was measured in the personal air samples and the highest levels were measured in the cooking and peeling department. This finding aligns with another study conducted in a shrimp processing plant where they measured 50 µg/m3 (47). Many of these processing plants still use thermal processing, which may increase protein aerosolisation and allergenicity by altering the protein structure and creating new epitopes (13). Earlier studies in crab processing plants have reported as much as 6 mg/m3 protein (53). The total protein levels in the present study were found to be significantly correlated with the level of CCL20, IL13 and basophil count in exposed workers. The correlation suggests a collective role of these markers in the immune response to potential allergenic proteins. However, it also underscores that total protein levels provide limited information as not all proteins provoke sensitisation upon inhalation (54). Therefore, it is essential to investigate also the composition of allergenic proteins in the air. Previous studies have reported total allergen levels, based on reactivity to patient sera, of 6.3 μg/m3 in the shrimp processing industry and 75 μg/m3 in the fish industry (8, 53). However, these studies do not identify the causative allergen. In this study, we measured the levels of the major shrimp allergen tropomyosin in the aerosol samples and very high levels were observed, ranging from 62 to 7,070 ng/m3. The highest tropomyosin level, 7,070 ng/m3, was measured among the technicians in the cooking and peeling department. Moreover, all other work processes also exhibited relatively high tropomyosin levels compared to the historically reported 375 ng/m3 in shrimp processing plants (7). In addition, the peak tropomyosin level recorded in this study also surpassed the previously reported levels (5,133 ng/m3) in prawn processing plants (5). These levels significantly exceed what has been reported in the crab processing industry, which ranged from 0.15 to 138 ng/m3 (55). While we showed high levels of tropomyosin in the aerosol, the levels of tropomyosin were only moderately correlated to the total protein, indicating that additional factors contribute to the reported respiratory symptoms among exposed workers. Characterisation of the composition of the allergenic proteins in the aerosol showed that the allergenic burden in the shrimp processing industry is complex, and both other known allergens such as arginine kinase, sarcoplasmic calcium-binding protein, troponin C, triosephosphate isomerase, myosin light chain and hemocyanin, as well as potential novel allergens may contribute to the observed respiratory symptoms among exposed workers. While we reported very high levels of the major known allergen tropomyosin, the contribution of other allergens is also of interest. Although methods to quantify the other identified allergies are not yet available, we conducted a semi-quantitative analysis of arginine kinase. The results showed arginine kinase was present in all work processes, with the higher levels observed among truck drivers and technicians. Previously, Abdel Rahman and colleagues have reported an arginine kinase level of 420 ng/m3 in the shrimp processing industry using mass spectrometric analysis (7).
In addition to allergenic proteins, proteases such as trypsin may induce airway inflammation and exacerbate allergic responses through disruption of epithelial barriers and release of inflammatory mediators (23, 25, 27). In the present study, proteases were identified in personal samples collected primarily from the thawing department. While trypsin was not detected in any of the samples, metalloproteases were observed in one sample. Exposure to these proteases can stimulate the airways through the attraction of eosinophils and neutrophils, a reaction that resembles the response to allergens (22). Trypsin is predominantly located in the digestive system and is believed to be released during the gutting processes in seafood processing; thus, its absence in the shrimp industry is unsurprising. In shrimp processing, there is deveining rather than gutting, and the small size of the shrimp further contributes to the absence of trypsin. In addition, exposure to chitin has also been associated with pulmonary inflammation and asthma (19–21). High exposure levels of chitin are expected in the flour production department, where the shell is dried and ground, and could thus contribute to the reported health effects among exposed workers. Finally, while allergens are a major sensitising factor in the seafood industry, other factors, such as chemicals, endotoxins and microbes, may also contribute to respiratory ailments (8, 56). The endotoxin levels we measured in these shrimp processing plants were largely below the recommended occupational exposure levels of 90 EU/m3 (57). Despite these low endotoxin levels, significant negative correlation between the endotoxin levels and monocyte count was observed, indicating that the presence of endotoxin could also contribute to the immune response observed among exposed workers.
This study evaluated the allergenic exposure in shrimp processing plants, linking the exposure to known biomarkers of allergy and asthma. The comprehensive mapping of the level and composition of allergenic proteins in the work environment enhanced and strengthened the study. While it provides novel insight into occupational exposure and respiratory health issues among shrimp processing workers, its cross-sectional design and the relatively low sample size restrict its generalisation and limit the establishment of causality, further warranting the need for larger studies to strengthen and corroborate the findings.
In this study, respiratory symptoms were assessed as self-reported symptoms through questionnaires. Self-reporting may cause reporting bias, and additional clinical evaluations of respiratory symptoms would strengthen the study. Furthermore, it is important to note that the healthy worker effect is common in industries with a high prevalence of allergic symptoms, as workers experiencing health problems often leave the workforce or switch professions. This may result in an underestimation of the health impacts of occupational exposures (58, 59).
Finally, the study was limited by methodological challenges due to the lack of a complete sequencing of Pandalus borealis in the reference database. Consequently, protein identification was done based on homology with other species within the infraorder Caridea. This resulted in low coverage for some proteins with low homology, restricting the interpretation and highlighting the need for additional studies to verify these findings. Moreover, protocols for extracting proteins from air filter samples are not standardised. Protocol selection may potentially introduce bias in the representative composition of proteins in the samples.
This study reports a high prevalence of respiratory symptoms and respiratory sensitisation, accompanied by elevated levels of allergy and asthma biomarkers in exposed shrimp processing workers. It provides novel insight into the level and composition of allergenic exposure in the shrimp processing industry, demonstrating alarmingly high tropomyosin levels in specific work tasks. Therefore, developing suitable monitoring tools, such as high throughput standardised assays for measurement and monitoring of specific allergens, could contribute to identifying potential health-impairing exposures and help to evaluate exposure-reducing measures. Altogether, these findings underscore the importance of mitigating occupational exposure to allergenic proteins to reduce the risk of allergies and asthma among workers. This can be achieved by introducing targeted preventative initiatives in high-exposure processes, such as implementing engineering controls (including effective ventilation systems) or using personal protective equipment.
The proteomics data are available in the publicly accessible Proteomic Identification Database (PRIDE) partner repository with the dataset identifier PXD061442. Additional original data presented in this study are included in the supplementary materials. The questionnaire data presented in this article are not readily available due to ethical concerns. Requests to access these datasets should be directed to the corresponding author.
The studies involving humans were approved by Regional Committee for Medical and Health Research Ethics. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
FZ: Data curation, Formal analysis, Investigation, Methodology, Resources, Visualization, Writing – original draft, Writing – review & editing. PG: Conceptualization, Funding acquisition, Resources, Writing – review & editing. MG: Investigation, Writing – review & editing. SM: Conceptualization, Funding acquisition, Writing – review & editing. AA: Conceptualization, Funding acquisition, Writing – review & editing. BB: Resources, Writing – review & editing. KCN: Conceptualization, Funding acquisition, Writing – review & editing. AS: Conceptualization, Funding acquisition, Supervision, Writing – review & editing. JE: Conceptualization, Funding acquisition, Methodology, Project administration, Supervision, Writing – review & editing.
The author(s) declare that financial support was received for the research and/or publication of this article. This study was funded by the National Institute of Occupational Health (STAMI) under grant number 21/00139 and the Research Fund for Asthma and Allergy by the Norwegian Asthma and Allergy Association (2022).
We extend our heartfelt thanks to the employees at the participating shrimp processing plants and the engineers at the section for Occupational Psychology and Physiology, National Institute of Occupational Health, for their assistance with blood sampling. We thank Dr. Øivind Skare (National Institute of Occupational Health) for his advice and guidance in the statistical analysis.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/falgy.2025.1520576/full#supplementary-material
1. ^Zegeye FD, Straumfors A, Lei P, Graff P, Erdem JS, Afanou AK. Microbial exposure and diversity in Norwegian shrimp processing plants. J Occup Environ Hyg. (2025).
1. Fao 2024. The State of World Fisheries and Aquaculture 2024. Blue Transformation in action. Rome.
2. Jeebhay MF, Cartier A. Seafood workers and respiratory disease: an update. Curr Opin Allergy Clin Immunol. (2010) 10:104–13. doi: 10.1097/ACI.0b013e3283373bd0
3. Jeebhay MF, Robins TG, Lopata AL. World at work: fish processing workers. Occup Environ Med. (2004) 61:471–4. doi: 10.1136/oem.2002.001099
4. Fagernæs CF, Lauritzen HB, Tøndell A, Hassel E, Bang BE, Tjalvin G, et al. Occupational asthma in the Salmon processing industry: a case series. Occup Environ Med. (2024) 81:400–6. doi: 10.1136/oemed-2024-109564
5. Mason HJ, Carder M, Money A, Evans G, Seed M, Agius R, et al. Occupational asthma and its causation in the UK seafood processing industry. Ann Work Expo Health. (2020) 64:817–25. doi: 10.1093/annweh/wxaa055
6. Lopata AL, Jeebhay MF. Airborne seafood allergens as a cause of occupational allergy and asthma. Curr Allergy Asthma Rep. (2013) 13:288–97. doi: 10.1007/s11882-013-0347-y
7. Abdel Rahman AM, Kamath SD, Gagné S, Lopata AL, Helleur R. Comprehensive proteomics approach in characterizing and quantifying allergenic proteins from northern shrimp: toward better occupational asthma prevention. J Proteome Res. (2013) 12:647–56. doi: 10.1021/pr300755p
8. Bang B, Aasmoe L, Aamodt BH, Aardal L, Andorsen GS, Bolle R, et al. Exposure and airway effects of seafood industry workers in northern Norway. J Occup Environ Med. (2005) 47:482–92. doi: 10.1097/01.jom.0000161732.96555.2b
9. Bang BE, Malla N, Bhagwat SS, Aasmoe L, Winberg JO. A sensitive assay for proteases in bioaerosol samples: characterization and quantification of airborne proteases in Salmon industry work environments. Ann Work Expo Health. (2018) 62:942–52. doi: 10.1093/annweh/wxy050
10. Madsen J, Sherson D, Kjøller H, Hansen I, Rasmussen K. Occupational asthma caused by sodium disulphite in Norwegian lobster fishing. Occup Environ Med. (2004) 61:873–4. doi: 10.1136/oem.2003.011585
11. Bonlokke JH, Bang B, Aasmoe L, Rahman AMA, Syron LN, Andersson E, et al. Exposures and health effects of bioaerosols in seafood processing workers—a position statement. J Agromedicine. (2019) 24:441–8. doi: 10.1080/1059924x.2019.1646685
12. Jeebhay MF, Moscato G, Bang BE, Folletti I, Lipińska-Ojrzanowska A, Lopata AL, et al. Food processing and occupational respiratory allergy- an EAACI position paper. Allergy. (2019) 74:1852–71. doi: 10.1111/all.13807
13. Dong X, Raghavan V. Recent advances of selected novel processing techniques on shrimp allergenicity: a review. Trends Food Sci Technol. (2022) 124:334–44. doi: 10.1016/j.tifs.2022.04.024
14. Lopata AL, Kleine-Tebbe J, Kamath SD. Allergens and molecular diagnostics of shellfish allergy. Allergo J Int. (2017) 25:210–8. doi: 10.1007/s40629-016-0124-2
15. Karnaneedi S, Huerlimann R, Johnston EB, Nugraha R, Ruethers T, Taki AC, et al. Novel allergen discovery through comprehensive de novo transcriptomic analyses of five shrimp species. Int J Mol Sci. (2021) 22:32. doi: 10.3390/ijms22010032
16. Faber MA, Pascal M, El Kharbouchi O, Sabato V, Hagendorens MM, Decuyper I, et al. Shellfish allergens: tropomyosin and beyond. Allergy. (2017) 72:842–8. doi: 10.1111/all.13115
17. Lee CH, Wu CC, Tyan YC, Yu WT, Huang ES, Yu HS. Identification of pyruvate kinase as a novel allergen in whiteleg shrimp (Litopenaeus vannamei) by specific-IgE present in patients with shrimp allergy. Food Chem. (2018) 258:359–65. doi: 10.1016/j.foodchem.2018.03.088
18. Ruethers T, Taki AC, Johnston EB, Nugraha R, Le TTK, Kalic T, et al. Seafood allergy: a comprehensive review of fish and shellfish allergens. Mol Immunol. (2018) 100:28–57. doi: 10.1016/j.molimm.2018.04.008
19. Bertelsen RJ, Svanes Ø, Madsen AM, Hollund BE, Kirkeleit J, Sigsgaard T, et al. Pulmonary illness as a consequence of occupational exposure to shrimp shell powder. Environ Res. (2016) 148:491–9. doi: 10.1016/j.envres.2016.04.033
20. Brinchmann BC, Bayat M, Brøgger T, Muttuvelu DV, Tjønneland A, Sigsgaard T. A possible role of chitin in the pathogenesis of asthma and allergy. Ann Agric Environ Med. (2011) 18:7–12.21736263
21. Reese TA, Liang HE, Tager AM, Luster AD, Van Rooijen N, Voehringer D, et al. Chitin induces accumulation in tissue of innate immune cells associated with allergy. Nature. (2007) 447:92–6. doi: 10.1038/nature05746
22. Reed CE. Inflammatory effect of environmental proteases on airway mucosa. Curr Allergy Asthma Rep. (2007) 7:368–74. doi: 10.1007/s11882-007-0056-5
23. Sun S, Lopata AL. The role of shellfish proteases in allergic diseases and inflammation. Curr Aller\ Clin Immunol. (2010) 23:174–9. doi: 10.520/EJC21687
24. Zentner A, Jeep S, Wahl R, Kunkel G, Kleine-Tebbe J. Multiple IgE-mediated sensitizations to enzymes after occupational exposure: evaluation by skin prick test, RAST, and immunoblot. Allergy. (1997) 52:928–34. doi: 10.1111/j.1398-9995.1997.tb01253.x
25. Bang B, Larsen M, Larsen AK, Aasmoe L. Proteases from Salmon stimulate IL-8 in airway epithelial cells. J Toxicol Environ Health A. (2009) 72:571–3. doi: 10.1080/15287390902733499
26. Bhagwat SS, Larsen AK, Seternes OM, Bang BE. Mixed exposure to bacterial lipopolysaccharide and seafood proteases augments inflammatory signalling in an airway epithelial cell model (A549). Toxicol Ind Health. (2016) 32:1866–74. doi: 10.1177/0748233715590914
27. Larsen AK, Seternes OM, Larsen M, Aasmoe L, Bang B. Salmon Trypsin stimulates the expression of interleukin-8 via protease-activated receptor-2. Toxicol Appl Pharmacol. (2008) 230:276–82. doi: 10.1016/j.taap.2008.02.019
28. Laustsen BH, Omland Ø, Würtz ET, Sigsgaard T, Ebbehøj NE, Carstensen O, et al. Rhino conjunctivitis and asthma among seafood processing workers in Greenland. A cross-sectional study. Front Allergy. (2021) 2:747011. doi: 10.3389/falgy.2021.747011
29. Shiryaeva O, Aasmoe L, Straume B, Bang BE. Respiratory impairment in Norwegian Salmon industry workers: a cross-sectional study. J Occup Environ Med. (2010) 52:1167–72. doi: 10.1097/JOM.0b013e3181fc5e35
30. Verstraelen S, Bloemen K, Nelissen I, Witters H, Schoeters G, Van Den Heuvel R. Cell types involved in allergic asthma and their use in in vitro models to assess respiratory sensitization. Toxicol in Vitro. (2008) 22:1419–31. doi: 10.1016/j.tiv.2008.05.008
31. Kim H, Ellis AK, Fischer D, Noseworthy M, Olivenstein R, Chapman KR, et al. Asthma biomarkers in the age of biologics. Allergy Asthma Clin Immunol. (2017) 13:48. doi: 10.1186/s13223-017-0219-4
32. Lambrecht BN, Hammad H, Fahy JV. The cytokines of asthma. Immunity. (2019) 50:975–91. doi: 10.1016/j.immuni.2019.03.018
33. Song P, Adeloye D, Salim H, Dos Santos JP, Campbell H, Sheikh A, et al. Global, regional, and national prevalence of asthma in 2019: a systematic analysis and modelling study. J Glob Health. (2022) 12:04052. doi: 10.7189/jogh.12.04052
34. Liu R, Holck AL, Yang E, Liu C, Xue W. Tropomyosin from tilapia (oreochromis mossambicus) as an allergen. Clin Exp Allergy. (2013) 43:365–77. doi: 10.1111/cea.12056
35. Bao Y, Zhu X. Role of chemokines and inflammatory cells in respiratory allergy. J Asthma Allergy. (2022) 15:1805–22. doi: 10.2147/jaa.S395490
36. Lloyd C. Chemokines in allergic lung inflammation. Immunology. (2002) 105:144–54. doi: 10.1046/j.1365-2567.2002.01344.x
37. Borish LC, Steinke JW. 2. Cytokines and chemokines. J Allergy Clin Immunol. (2003) 111:S460–75. doi: 10.1067/mai.2003.108
38. Paplinska-Goryca M, Misiukiewicz-Stepien P, Proboszcz M, Nejman-Gryz P, Gorska K, Krenke R. The expressions of TSLP, IL-33, and IL-17A in monocyte derived dendritic cells from asthma and COPD patients are related to epithelial-macrophage interactions. Cells. (2020) 9:1944. doi: 10.3390/cells9091944
39. Hinks TSC, Brown T, Lau LCK, Rupani H, Barber C, Elliott S, et al. Multidimensional endotyping in patients with severe asthma reveals inflammatory heterogeneity in matrix metalloproteinases and chitinase 3-like protein 1. J Allergy Clin Immunol. (2016) 138:61–75. doi: 10.1016/j.jaci.2015.11.020
40. Komi DE, Kazemi T, Bussink AP. New insights into the relationship between chitinase-3-like-1 and asthma. Curr Allergy Asthma Rep. (2016) 16:57. doi: 10.1007/s11882-016-0637-2
41. Zeng J, Tao M, Sun H, Xu G. The emerging role of CHI3L1 as a diagnostic marker and therapeutic target in allergic diseases and respiratory comorbidities. Aller Med. (2024) 1:100001. doi: 10.1016/j.allmed.2024.100001
42. Dupin I, Contin-Bordes C, Berger P. Fibrocytes in asthma and chronic obstructive pulmonary disease: variations on the same theme. Am J Respir Cell Mol Biol. (2018) 58:288–98. doi: 10.1165/rcmb.2017-0301PS
43. Hohlfeld JM, Erpenbeck VJ, Krug N. Surfactant proteins SP-A and SP-D as modulators of the allergic inflammation in asthma. Pathobiology. (2002) 70:287–92. doi: 10.1159/000070744
44. Nguyen MN, Krutz NL, Limviphuvadh V, Lopata AL, Gerberick GF, Maurer-Stroh S. Allercatpro 2.0: a web server for predicting protein allergenicity potential. Nucleic Acids Res. (2022) 50:W36–w43. doi: 10.1093/nar/gkac446
45. Wai CY, Leung NY, Leung AS, Wong GW, Leung TF. Seafood allergy in Asia: geographical specificity and beyond. Front Aller. (2021) 2:676903. doi: 10.3389/falgy.2021.676903
46. Thomassen MR, Aasmoe L, Bang BE, Braaten T. Lung function and prevalence of respiratory symptoms in Norwegian crab processing workers. Int J Circumpolar Health. (2017) 76:1313513. doi: 10.1080/22423982.2017.1313513
47. Laustsen BH, Ebbehøj NE, Sigsgaard T, Rasmussen K, Bønløkke JH. Work environment, occupational diseases and accidents among seafood industry workers in Greenland. Dan Med J. (2022) 69(2):A05210470. Available at: https://ugeskriftet.dk/dmj/work-environment-occupational-diseases-and-accidents-among-seafood-industry-workers-greenland35088702
48. De Gier S, Verhoeckx K. Insect (food) allergy and allergens. Mol Immunol. (2018) 100:82–106. doi: 10.1016/j.molimm.2018.03.015
49. Platts-Mills TA, Woodfolk JA. Allergens and their role in the allergic immune response. Immunol Rev. (2011) 242:51–68. doi: 10.1111/j.1600-065X.2011.01021.x
50. Du L, Chen F, Xu C, Tan W, Shi J, Tang L, et al. Increased MMP12 mRNA expression in induced sputum was correlated with airway eosinophilic inflammation in asthma patients: evidence from bioinformatic analysis and experiment verification. Gene. (2021) 804:145896. doi: 10.1016/j.gene.2021.145896
51. Quoc L, Moon Q, Lee JY, Ban DH, Kim GY, & Park SH, et al. Role of Thymus and activation-regulated chemokine in allergic asthma. J Asthma Allergy. (2022) 15:157–67. doi: 10.2147/jaa.S351720
52. Principe S, Benfante A, Battaglia S, Maitland Van Der Zee AH, Scichilone N. The potential role of SP-D as an early biomarker of severity of asthma. J Breath Res. (2021) 15(4):041001. doi: 10.1088/1752-7163/ac20c1
53. Jeebhay MF. Occupational allergy and asthma in the seafood industry–emerging issues. Occu Health Southern Africa. (2011) 17:4–13.
54. Huby RD, Dearman RJ, Kimber I. Why are some proteins allergens? Toxicol Sci. (2000) 55:235–46. doi: 10.1093/toxsci/55.2.235
55. Kamath SD, Thomassen MR, Saptarshi SR, Nguyen HM, Aasmoe L, Bang BE, et al. Molecular and immunological approaches in quantifying the air-borne food allergen tropomyosin in crab processing facilities. Int J Hyg Environ Health. (2014) 217:740–50. doi: 10.1016/j.ijheh.2014.03.006
56. Thomassen MR, Kamath SD, Lopata AL, Madsen AM, Eduard W, Bang BE, et al. Occupational exposure to bioaerosols in Norwegian crab processing plants. Ann Occup Hyg. (2016) 60:781–94. doi: 10.1093/annhyg/mew030
57. Torén K. The Nordic Expert Group for Criteria Documentation of Health Risks from Chemicals and the Dutch Expert Committee on Occupational Safety. Endotoxins. Gothenburg: University of Gothenburg (2011). p. 60.
58. Le Moual N, Kauffmann F, Eisen EA, Kennedy SM. The healthy worker effect in asthma: work may cause asthma, but asthma may also influence work. Am J Respir Crit Care Med. (2008) 177:4–10. doi: 10.1164/rccm.200703-415PP
Keywords: allergy, respiratory symptoms, biomarkers, irritants, sensitisation, tropomyosin
Citation: Zegeye FD, Graff P, Grgic M, Mollerup S, Afanou AK, Bang BE, Nordby KC, Straumfors A and Erdem JS (2025) Respiratory symptoms, sensitisation and occupational exposure in the shrimp processing industry. Front. Allergy 6:1520576. doi: 10.3389/falgy.2025.1520576
Received: 31 October 2024; Accepted: 25 February 2025;
Published: 20 March 2025.
Edited by:
João Cavaleiro Rufo, University Porto, PortugalReviewed by:
Tanja Kalic Kamath, Karl Landsteiner University of Health Sciences, AustriaCopyright: © 2025 Zegeye, Graff, Grgic, Mollerup, Afanou, Bang, Nordby, Straumfors and Erdem. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Johanna Samulin Erdem, am9oYW5uYS5zYW11bGluLWVyZGVtQHN0YW1pLm5v
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.