 Aleksandra Podzhilkova
Aleksandra Podzhilkova Christoph Nagl
Christoph Nagl Karin Hoffmann-Sommergruber
Karin Hoffmann-Sommergruber- Institute of Pathophysiology and Allergy Research, Center of Pathophysiology, Infectiology and Immunology, Medical University of Vienna, Vienna, Austria
Allergic diseases represent a relevant global health problem, affecting adults and children and posing a significant burden for health care systems. In addition, the disease is still under-recognized and harmonized diagnostic tools and management plans for patients are still lacking. In this review the most important aspects of the diagnosis of allergic diseases are summarized and the contribution of Molecular allergology to this area is highlighted.
1. Introduction
Allergic diseases affect almost 30% of the population worldwide and induce symptoms of the respiratory system, gastrointestinal tract, skin, and the cardiovascular system (1). Allergy is a result of IgE-mediated immune responses to a foreign, usually harmless, protein (allergen). Allergens, recognized by the immune system, can induce mild or severe symptoms up to even life-threatening reactions in both, atopic children and adults (2, 3).
The immune system plays a critical role to protect the human body from viral or microbial infections. However, sometimes overstimulation of the immune system can lead to the opposite effect and induce inflammatory responses, also known as hypersensitivity reactions, Type I-IV (4, 5). Allergic reactions of Type I hypersensitivity, are defined as an immediate reaction against harmless antigens, mostly proteins, resulting in the production of specific immunoglobulin E (IgE) antibodies. This antibody-antigen interaction leads to mast cells degranulation and release of histamine and other inflammatory mediators (4) and causes vasodilation, decrease of blood pressure, bronchoconstriction and in some cases—anaphylactic shock (6).
The allergic sensitisation can develop via different ways of exposure—by inhalation, ingestion, injection, and skin contact. Aeroallergens are associated with increasing cases of respiratory disorders such as allergic rhinitis and asthma (7, 8). House dust mites (HDM), moulds, and animal fur and dander particles are frequent indoor allergen sources, while tree, weed, and grass pollens are the leading outdoor allergen sources (7, 9, 10).
Birch and grass pollens are the most common causes of allergic rhinitis and asthma in Europe (11). However, during the flowering season, pollen exposure levels can vary due to atmospheric factors such as wind, humidity, and rainfall. In addition, anthropogenic climate changes may have an impact on flowering seasons and thus pollen loads (12).
Food allergies represent another relevant health problem affecting patients of all age groups. In Westernized countries, the percentage of estimated overall food allergies are about 5% in adults and 8% in children (13). For example, allergy to cow's milk affects approximately 2.5% of the infants during the first two years of life. However, the majority of those pediatric patients resolves their milk allergy at school age (14, 15).
Allergy to tree nuts and peanuts can develop at any age. In Europe hazelnut sensitisation rates range from 1.8% (Reykjavik) to 14.35% in Zurich. However, the probable food allergy prevalence is much lower but differing across Europe, ranging from 0.06% for Athens vs. 2.57% in Zurich (16). Similar rates were observed for walnut allergy. An Australian study reported a prevalence of 2.3% of clinically relevant tree nut allergy with cashew and pistachio nuts being the most frequent allergenic tree nuts. Peanut allergy affects 1%–3% of children and persists throughout lifetime (17). The symptoms range from eczema, urticaria up to asthma and anaphylaxis. Sensitization to peanut allergens Ara h 5 and Ara h 8, belonging to the PR10 protein family and profilin, respectively usually induces symptoms belonging to the oral allergy syndrome. In contrast, Ara h 1, Ara h 2, and Ara h 3, belonging to the seed storage proteins, tend to induce more severe and systemic reactions (16, 18, 19).
Despite the great amount of allergens that are already described, the characterization of allergens, particularly in regions like Asia and Africa, are still missing due to lack of comprehensive data on allergens prevalent in local environments and food sources. Future campaigns including increased funding for joint projects, awareness campaigns and knowledge sharing initiatives can help to improve allergen characterization in Asian and African countries, leading to better allergy management and public health outcomes in these regions.
1.1. What is an allergen?
Allergens are typically proteins recognized by the immune system from a predisposed individual as foreign and potentially harmful. Upon contact with an allergen, the immune system produces specific antibodies called immunoglobulin E (IgE) antibodies (2).
Proteins, sharing similar sequences and structural elements, are assigned to protein families. Currently around 19,000 protein families are described. Out of those approximately 40 protein families contain allergenic proteins. Sequence similarity and shared structural elements of allergens are responsible for IgE cross reactivities (16).
In the recent past, bioscience has tremendously improved due to up to date technology which enables precise characterization of proteins including allergens. This approach allows to identify and produce well defined highly pure molecules for component resolved diagnosis which in turn facilitates a patient tailored diagnosis and management. Some examples of the application of “Molecular Allergology” are described below.
Depending on the allergen source, there are different types of allergens present which can cause symptoms ranging from mild to severe reactions. For example, in birch pollen several allergens have been identified. Among those, Bet v 1 is the major allergen belonging to the pathogenesis-related protein family PR-10 (20). In an Austrian study of 501 adolescents, 16.3% showed IgE reactivity to Bet v 1, and in Switzerland and Denmark, the prevalence of birch pollen sensitization was 7.9% and 13.7%, respectively (21–23). Bet v 1 can exhibit cross-reactivity with highly homologous proteins present in a range of other allergen sources including pollens and plant foods. For example, individuals allergic to Bet v 1 may experience oral allergy syndrome when consuming certain fruits and vegetables, such as apple (Mal d 1), hazelnut (Cor a 1), peach (Pru p 1), and carrot (Dau c 1) due to cross-reactivity between Bet v 1 and similar proteins in these foods (24–26).
Another important type of allergen that causes allergic reactions is the cupin superfamily, usually present in allergenic food sources, such as tree nuts, peanuts and soybeans. Cupins (11S and 7S globulins) are characterized by their barrel-shaped structure and play various biological roles in plants, such as storage of nutrients, and defence mechanisms against pathogens (27). The most abundant nut allergens are seed storage proteins: vicilins (7S trimeric globulins), and legumins (11S hexameric globulins), followed by 2S albumins, nsLTPs, profilins and hevein-related proteins (28). Cross-reactivity among cupins from different plant sources can occur (29). For example, if someone is allergic to peanut cupin, they may also experience allergic reactions when consuming other legumes or seeds that contain similar cupin proteins, such as soybeans or sesame seeds. This cross-reactivity is due to the shared structural characteristics and sequence similarities among these proteins which are recognized by specific IgE molecules.
Non-specific lipid transfer proteins (nsLTPs) have been identified from various plant sources, including fruits, vegetables, nuts, and seeds (30). These proteins form a cavity which binds and transfers lipids (fatty acids and phospholipids) across cell membranes (31). While nsLTPs serve important functions in plants, they are also recognized as allergens in certain individuals. Due to the similarities in protein structures and sequences among nsLTPs from different plant sources, individuals sensitized to a given nsLTP from one food, may also experience allergic reactions when consuming other foods that contain similar nsLTPs. For example, sensitization to peach nsLTP (Pru p 3) may result in cross-reactivity with nsLTPs from other fruits like apple (Mal d 3), cherry (Pru p 3), or hazelnut (Cor a 8) (30, 32).
Recently, novel allergens present in fruits and pollens were identified as Gibberellin-regulated proteins (GRPs). The physicochemical properties of these proteins overlap to some extent with nsLTPs. Gibberellin-regulated proteins are small cationic proteins, contain 6 conserved disulfide bridges and are expressed in pollen, peel and pulp of fruits. They are upregulated upon pathogen attack. Allergens have been reported from peach, apricot, cherry and citrus fruits and from Cupressaceae pollen (16). Based on their structure these proteins also contain a cavity. However, no specific ligands have been identified so far.
Despite the increasing knowledge of different allergenic protein families including their sequences and structural determinants, it still remains unclear why certain proteins are inducing an allergic reaction. Recent studies provided evidence that additional compounds such as lipids, carbohydrates, and fibres may contribute to the allergic sensitization and effector phase (33).
For example, Bet v 1, nsLTP, 2S albumins, secretoglobins, and lipocalins contain hydrophobic binding sites for lipid ligands which can enhance a Th2-response (33–35). In a recent study by Janssen-Weets et al. the ligand binding activity of mammalian lipocalins and secretoglobins were investigated and fatty acids, fatty alcohols, and the terpene alcohol farnesol were the most relevant ligands with strong binding affinities to the proteins. These ligands may contribute to the immunomodulating activity of allergens and thus to allergic sensitization, although further studies are needed to prove this hypothesis (7).
Tree nuts have a high lipid content, ranging from 46%–83% of fatty acids, depending on the nut source (35, 36). Recently it was shown (8, 35, 36) that lipids bound to proteins can decrease the epithelial barrier integrity thereby contributing to the allergic sensitization process. This in turn facilitates the tight interaction of lipids with several components of the innate immune system (34, 37, 38).
Also, glycan moieties can act as immunomodulating compounds. Glycosylated allergenic proteins and peptides are interacting with specific receptors on dendritic cells, such as the macrophage mannose receptor, thus inducing an (innate) immune response (21).
Furthermore, allergens usually come together with additional substances such as pollen compounds, or food matrix components. Simultaneous exposure to both allergens and potent immunogenic substances may thus facilitate the onset of an allergic sensitization process.
1.2. Diagnosis of IgE mediated allergy
The diagnostic procedure for IgE-mediated allergy usually starts with a detailed assessment of the patient's history, followed by different tests: skin prick test, basophil activation test, serum-specific IgE tests, and food challenge tests to identify the range of IgE sensitisation (Figure 1). For a restricted number of patients challenge tests are performed such as pollen exposure or food challenge tests.
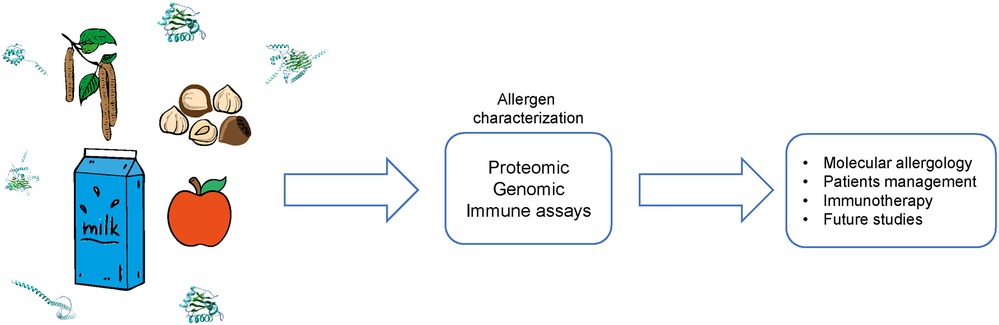
Figure 1. Overview on the critical steps of molecular allergy diagnosis.
Allergy diagnostics aim to identify the culprit allergen source and to measure the specific IgE values. In the following some of the commonly used tests are summarised:
1. Skin Prick Test (SPT): In this method, a small amount of allergen extract is applied to the skin, usually on the forearm or back. The skin is then pricked with a sterile lancet, allowing the allergen to enter the skin. If the person is allergic to a particular allergen present in the extract, they will develop a local reaction (“wheal and flare”), within 15–20 min. Positive control (histamine) and negative control (NaCl) are run in parallel. Skin prick tests identify reactions to inhalants, foods, some drugs, occupational allergens, hymenoptera venom and latex (39). Sometimes prick to prick tests are used to test a potential allergen source directly on the skin. The limitations of SPTs include:
• False positive or false negative results;
• Potential for allergic reactions: while the test is generally safe, there is a small risk of severe allergic reactions (anaphylaxis) in highly sensitive individuals. Skin prick tests should always be performed under the supervision of a qualified healthcare professional who can promptly manage any adverse reactions (40);
• In some cases, skin prick tests may produce inconclusive results. This can occur when the skin reacts to multiple allergens, making it challenging to determine the primary trigger. False negative results may be due to the fact the allergen extracts miss the culprit allergen: Additionally, individuals with certain skin conditions, such as eczema or other skin diseases, may have difficulty interpreting the test results accurately.
2. Several serum-specific IgE tests are currently used in clinical routine. Allergen extracts or single allergen batches are coupled to a solid phase and after serum incubation bound IgE antibodies are detected by labeled anti-IgE antibodies allowing a quantitative and qualitative analysis. While allergen extract based tests are regarded as a primary approach, chip based test formats containing more than hundred individual allergens are also available. The first approach provides information on the culprit allergen source and has a generally higher test sensitivity. Limitations of this approach are the batch to batch variation and the potential lack of individual allergens in some sources and the required higher volume of serum samples. The second approach also called multiplex testing provides information on IgE sensitization to individual allergens. This test has a higher specificity and requires only minute amounts of serum samples. Limitations of this test format are higher costs and less availability on a global scale. While both tests provide information on IgE sensitization to allergen sources, the multiplex assay allows to assess the allergen specific sensitization pattern, differentiating between cross-reactive structures and species specific allergens. Furthermore, information on sensitization to marker allergens allows a detailed risk assessment regarding disease progression and personalized treatment options (16).
3. The Basophil Activation Test (BAT) provides information on the presence of allergen specific IgE and cellular activation and degranulation (16). Due to the activation of basophils by allergen extracts or specific allergens, upregulation of surface marker expression can be measured and allows to assess the onset of an allergic reaction (41, 42). BAT assays can be performed using either patient-derived cells or by applying a standardized cell population from a given donor. In this case, basophils are then incubated with serum samples from patients (also called passive sensitization). Recently, Stoffersen et al. reported that the required spIgE levels for optimal cellular activation are ranging from 3,51 kUA/L (CAP class 3) up to >17,5 kUA/L (CAP class 4) using Bet v 1 as a model (43). BAT represents a promising approach for diagnosing allergic reactions (43). This functional test provides a quick and reliable way to identify specific allergens using patients' samples, paving the way for more effective treatments and personalized care. In specialized centers BAT is already included in daily diagnosis of allergies. Especially for peanut and sesame allergy a BAT assay is recommended if other test results remain inconclusive (9, 41, 42). Wheat dependent exercise-induced anaphylaxis (WDEIA) is a co-factor induced wheat allergy and considered to be a rare disease. Gabler and co-authors tested a range of gluten and non-gluten proteins from wheat in the BAT assays using samples from WDEIA patients. Using this approach they could identify gluten and additional non-gluten proteins from wheat as allergens relevant for WDEIA. This approach may improve the diagnosis of this disease and help to rule out idiopathic anaphylaxis for these patients (41). Recently, BAT tests have been shown to be helpful in another IgE mediated disease, ABPA, allergic bronchopulmonary aspergillosis. This disease is caused by the ubiquitous mould Aspergillus fumigatus and may be underdiagnosed in certain patient groups suffering from a chronic pulmonary disease. Michel and coauthors showed that BAT assays performed with samples from a cohort of patients proved useful in the diagnosis for ABPA, when using A. fumigatus extract and Asp f 1, 2, 3, 4, 6, respectively (9).
1.3. Molecular allergology and its application in prevention and management of the patient
As mentioned above molecular allergology has improved diagnosis of allergies tremendously. However, for molecular testing the quality of the analytes, that is highly pure and well characterized batches of natural or recombinant allergens, is crucial. This approach requires powerful expression platforms for recombinant proteins and purification procedures for both natural and recombinant proteins, respectively. Critical steps of the final quality assessment of allergen batches include purity, intact tertiary protein structure, and biological activity. To assess these parameters a number of physicochemical methods can be applied including mass spectrometry, CD spectroscopy, and NMR analyses. In parallel the IgE binding activity needs to be confirmed.
While for some allergens only single isoforms have been identified, other proteins are present as a range of different isoforms. To identify a representative isoform of diagnostic relevance several analytical methods need to be applied. Recently Marsh and colleagues performed a study on the range of isoforms of peanut allergens Ara h 1, 2, 3, and 6 to assess the range of isoforms present in 20 different peanut genotypes. Applying LC-MS/MS and RP/HPLC they could quantify and compare the content of Ara h 1,2,3, and 6 present in various commercially used genotypes. This approach is important for food allergy risk assessment and can be applied to obtain well defined reference material to be used for allergen detection (27).
Various physicochemical parameters such as heat treatment or pH environment can affect the structure of an allergenic protein. This in turn may impact on its immunogenic potency (44). Using profilin as a model protein family, pollen allergens Amb a 8, Art v 4, and Bet v 2 were selected and a MD simulation strategy applied to assess their structural dynamics at varying pH levels. Although these proteins share high structural and sequence similarities they showed different fold stability under different pH conditions (44).
With the application of molecular allergology the immunological effects during specific immunotherapy can also be assessed. Thörnqvist and colleagues used a peptide based approach to assess the development of specific antibody responses during grass pollen immunotherapy (45). An allergome-wide peptide microarray was developed and the IgG, IgE and IgG4 levels of grass pollen specific peptides were investigated using this high-throughput approach. The evolution of linear epitope-specific antibody responses were analysed. These data contribute to our current understanding of the development of the immune response during immunotherapy and can help to further improve immunotherapeutics.
In summary the application of molecule based allergy diagnosis provides additional information regarding allergens, helps to assess the range of potential cross reactivities, and for some allergen families also the severity of symptoms caused. This information can further help to give personal advice to patients, and train them to avoid exposure to the allergen sources such as pollens, plant foods and their presence in processed foods.
2. Conclusions
Allergy diagnostics help in accurately identifying the specific allergens that trigger an individual's allergic reactions. This knowledge is crucial for understanding the underlying cause of allergies and providing targeted and personalized treatment plans. Once the allergens are identified through diagnostic testing, healthcare professionals can develop effective treatment strategies. This may include allergen avoidance measures, medication management, or allergen-specific immunotherapy (45–47). It can significantly improve the quality of life for individuals with allergies by reducing symptoms, fine-tuning medication plans, and potentially preventing the progression of allergic symptoms.
Author contributions
AP: Writing – original draft, Writing – review & editing. CN: Writing – original draft. KH: Writing—original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article.
This work was supported by Danube Allergy Research Cluster project P07 funded by the State of Lower Austria.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Valenta R, Karaulov A, Niederberger V, Gattinger P, van Hage M, Flicker S, et al. Molecular aspects of allergens and allergy. Adv Immunol. (2018) 138:195–256. doi: 10.1016/bs.ai.2018.03.002
2. Lei DK, Grammer LC. An overview of allergens. Allergy Asthma Proc. (2019) 40:362–5. doi: 10.2500/aap.2019.40.4247
3. Seth D, Poowutikul P, Pansare M, Kamat D. Food allergy: a review. Pediatr Ann. (2020) 49:e50–8. doi: 10.3928/19382359-20191206-01
4. Abbas M, Moussa M, Akel H. Type I hypersensitivity reaction. Treasure Island, FL: StatPearls. StatPearls Publishing Copyright © 2023, StatPearls Publishing LLC (2023). 32809396 p.
5. Alsaleh NB, Brown JM. Engineered nanomaterials and type I allergic hypersensitivity reactions. Front Immunol. (2020) 11:222. doi: 10.3389/fimmu.2020.00222
6. King TC. 2—inflammation, inflammatory mediators, and immune-mediated disease. In: King TC, editor. Elsevier’s integrated pathology. Philadelphia: Mosby (2007). p. 21–57.
7. Janssen-Weets B, Kerff F, Swiontek K, Kler S, Czolk R, Revets D, et al. Mammalian derived lipocalin and secretoglobin respiratory allergens strongly bind ligands with potentially immune modulating properties. Front Allergy. (2022) 3:958711. doi: 10.3389/falgy.2022.958711
8. Martorano L, Erwin EA. Aeroallergen exposure and spread in the modern era. J Allergy Clin Immunol Pract. (2018) 6:1835–42. doi: 10.1016/j.jaip.2018.08.014
9. Michel M, Sereme Y, Mankouri F, Gouitaa M, Gautier C, Mège JL, et al. Basophil activation test with Aspergillus molecules: the case for ABPA. Front Allergy. (2022) 3:898731. doi: 10.3389/falgy.2022.898731
10. Singh M, Hays A. Indoor and outdoor allergies. Prim Care. (2016) 43:451–63. doi: 10.1016/j.pop.2016.04.013
11. Smith M, Jäger S, Berger U, Sikoparija B, Hallsdottir M, Sauliene I, et al. Geographic and temporal variations in pollen exposure across Europe. Allergy. (2014) 69:913–23. doi: 10.1111/all.12419
12. Rojo J, Oteros J, Picornell A, Maya-Manzano JM, Damialis A, Zink K, et al. Effects of future climate change on birch abundance and their pollen load. Glob Chang Biol. (2021) 27:5934–49. doi: 10.1111/gcb.15824
13. Sicherer SH, Sampson HA. Food allergy: epidemiology, pathogenesis, diagnosis, and treatment. J Allergy Clin Immunol. (2014) 133:291–307; quiz 8. doi: 10.1016/j.jaci.2013.11.020
14. Geiselhart S, Podzhilkova A, Hoffmann-Sommergruber K. Cow’s milk processing-friend or foe in food allergy? Foods. (2021) 10:572. doi: 10.3390/foods10030572
15. Iweala OI, Choudhary SK, Commins SP. Food allergy. Curr Gastroenterol Rep. (2018) 20:17. doi: 10.1007/s11894-018-0624-y
16. Dramburg S, Hilger C, Santos AF, de Las Vecillas L, Aalberse RC, Acevedo N, et al. EAACI molecular allergology user’s guide 2.0. Pediatr Allergy Immunol. (2023) 34(Suppl 28):e13854. doi: 10.1111/pai.13854
17. Sicherer SH, Sampson HA. Food allergy: a review and update on epidemiology, pathogenesis, diagnosis, prevention, and management. J Allergy Clin Immunol. (2018) 141:41–58. doi: 10.1016/j.jaci.2017.11.003
18. Chu DK. Tree nut allergy: a systematic review. Curr Opin Pediatr. (2022) 34:600–8. doi: 10.1097/MOP.0000000000001180
19. Iizuka T, Barre A, Rougé P, Charpin D, Scala E, Baudin B, et al. Gibberellin-regulated proteins: emergent allergens. Front Allergy. (2022) 3:877553. doi: 10.3389/falgy.2022.877553
20. Biedermann T, Winther L, Till SJ, Panzner P, Knulst A, Valovirta E. Birch pollen allergy in Europe. Allergy. (2019) 74:1237–48. doi: 10.1111/all.13758
21. Huby RDJ, Dearman RJ, Kimber I. Why are some proteins allergens? Toxicol Sci. (2000) 55:235–46. doi: 10.1093/toxsci/55.2.235
22. Schmitz R, Ellert U, Kalcklösch M, Dahm S, Thamm M. Patterns of sensitization to inhalant and food allergens—findings from the German health interview and examination survey for children and adolescents. Int Arch Allergy Immunol. (2013) 162:263–70. doi: 10.1159/000353344
23. Stemeseder T, Klinglmayr E, Moser S, Lueftenegger L, Lang R, Himly M, et al. Cross-sectional study on allergic sensitization of Austrian adolescents using molecule-based IgE profiling. Allergy. (2017) 72:754–63. doi: 10.1111/all.13071
24. Fahlbusch B, Rudeschko O, Müller WD, Schlenvoigt G, Vettermann S, Jäger L. Purification and characterization of the major allergen from apple and its allergenic cross-reactivity with Bet v 1. Int Arch Allergy Immunol. (1995) 108:119–26. doi: 10.1159/000237128
25. Hoffmann-Sommergruber K, O’Riordain G, Ahorn H, Ebner C, Laimer Da Camara Machado M, Pühringer H, et al. Molecular characterization of Dau c 1, the Bet v 1 homologous protein from carrot and its cross-reactivity with Bet v 1 and Api g 1. Clin Exp Allergy. (1999) 29:840–7. doi: 10.1046/j.1365-2222.1999.00529.x
26. Hofmann C, Scheurer S, Rost K, Graulich E, Jamin A, Foetisch K, et al. Cor a 1-reactive T cells and IgE are predominantly cross-reactive to Bet v 1 in patients with birch pollen-associated food allergy to hazelnut. J Allergy Clin Immunol. (2013) 131:1384–92.e6. doi: 10.1016/j.jaci.2012.10.037
27. Ramadan S, Marsh J, El-Sherbeny GA, El-Halawany EF, Luan F, Baumert JL, et al. Purification of soybean cupins and comparison of IgE binding with peanut allergens in a population of allergic subjects. Food Chem Toxicol. (2021) 147:111866. doi: 10.1016/j.fct.2020.111866
28. Roux KH, Teuber SS, Sathe SK. Tree nut allergens. Int Arch Allergy Appl Immunol. (2003) 131:234–44. doi: 10.1159/000072135
29. Bublin M, Breiteneder H. Cross-reactivities of non-homologous allergens. Allergy. (2020) 75:1019–22. doi: 10.1111/all.14120
30. Skypala IJ, Asero R, Barber D, Cecchi L, Diaz Perales A, Hoffmann-Sommergruber K, et al. Non-specific lipid-transfer proteins: allergen structure and function, cross-reactivity, sensitization, and epidemiology. Clin Transl Allergy. (2021) 1:e12010. doi: 10.1002/clt2.12010
31. Nazeer M, Waheed H, Saeed M, Ali SY, Choudhary MI, Ul-Haq Z, et al. Purification and characterization of a nonspecific lipid transfer protein 1 (nsLTP1) from Ajwain (Trachyspermum ammi) seeds. Sci Rep. (2019) 9:4148. doi: 10.1038/s41598-019-40574-x
32. Zuidmeer L, van Leeuwen WA, Budde IK, Cornelissen J, Bulder I, Rafalska I, et al. Lipid transfer proteins from fruit: cloning, expression and quantification. Int Arch Allergy Immunol. (2005) 137:273–81. doi: 10.1159/000086420
33. González Roldán N, Duda KA. Editorial: role of lipids in the dynamics of allergic airway inflammation. Front Immunol. (2020) 11:612297. doi: 10.3389/fimmu.2020.612297
34. Bublin M, Eiwegger T, Breiteneder H. Do lipids influence the allergic sensitization process? J Allergy Clin Immunol. (2014) 134:521–9. doi: 10.1016/j.jaci.2014.04.015
35. Palladino C, Ellinger I, Kalic T, Humeniuk P, Ret D, Mayr V, et al. Peanut lipids influence the response of bronchial epithelial cells to the peanut allergens Ara h 1 and Ara h 2 by decreasing barrier permeability. Front Mol Biosci. (2023) 10:1126008. doi: 10.3389/fmolb.2023.1126008
36. Dubiela P, Aina R, Polak D, Geiselhart S, Humeniuk P, Bohle B, et al. Enhanced Pru p 3 IgE-binding activity by selective free fatty acid-interaction. J Allergy Clin Immunol. (2017) 140:1728–31.e10. doi: 10.1016/j.jaci.2017.06.016
37. Eisenbarth SC, Piggott DA, Huleatt JW, Visintin I, Herrick CA, Bottomly K. Lipopolysaccharide-enhanced, toll-like receptor 4-dependent T helper cell type 2 responses to inhaled antigen. J Exp Med. (2002) 196:1645–51. doi: 10.1084/jem.20021340
38. Mogensen JE, Wimmer R, Larsen JN, Spangfort MD, Otzen DE. The major birch allergen, Bet v 1, shows affinity for a broad spectrum of physiological ligands. J Biol Chem. (2002) 277:23684–92. doi: 10.1074/jbc.M202065200
39. Frati F, Incorvaia C, Cavaliere C, Di Cara G, Marcucci F, Esposito S, et al. The skin prick test. J Biol Regul Homeost Agents. (2018) 32(1 Suppl. 1):19–24.29552869
40. Rusznak C, Davies RJ. ABC Of allergies. Diagnosing allergy. Br Med J. (1998) 316:686–9. doi: 10.1136/bmj.316.7132.686
41. Gabler AM, Gebhard J, Norwig MC, Eberlein B, Biedermann T, Brockow K, et al. Basophil activation to gluten and non-gluten proteins in wheat-dependent exercise-induced anaphylaxis. Front Allergy. (2022) 3:822554. doi: 10.3389/falgy.2022.822554
42. Santos AF, Alpan O, Hoffmann HJ. Basophil activation test: mechanisms and considerations for use in clinical trials and clinical practice. Allergy. (2021) 76:2420–32. doi: 10.1111/all.14747
43. Stoffersen P, Skov PS, Poulsen LK, Jensen BM. The allergen-specific IgE concentration is important for optimal histamine release from passively sensitized basophils. Front Allergy. (2022) 3:875119. doi: 10.3389/falgy.2022.875119
44. Hofer F, Fischer AL, Kamenik AS, Waibl F, Fernández-Quintero ML, Liedl KR. pH-dependent structural diversity of profilin allergens determines thermal stability. Front Allergy. (2022) 3:1007000. doi: 10.3389/falgy.2022.1007000
45. Thörnqvist L, Sjöberg R, Greiff L, van Hage M, Ohlin M. Linear epitope binding patterns of grass pollen-specific antibodies in allergy and in response to allergen-specific immunotherapy. Front Allergy. (2022) 3:859126. doi: 10.3389/falgy.2022.859126
46. Wong AG, Lomas JM. Allergy testing and immunotherapy. Pediatr Rev. (2019) 40:219–28. doi: 10.1542/pir.2018-0126
Keywords: allergy, allergen diagnosis, allergens, IgE-mediated allergy, molecular allergology
Citation: Podzhilkova A, Nagl C and Hoffmann-Sommergruber K (2023) Molecular allergology and its application in prevention, diagnosis and therapy. Front. Allergy 4:1260902. doi: 10.3389/falgy.2023.1260902
Received: 18 July 2023; Accepted: 28 July 2023;
Published: 7 August 2023.
Edited by:
Wayne Robert Thomas, University of Western Australia, AustraliaReviewed by:
Kyoung Yong Jeong, Yonsei University, Republic of Korea© 2023 Podzhilkova, Nagl and Hoffmann-Sommergruber. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Karin Hoffmann-Sommergruber a2FyaW4uaG9mZm1hbm4tc29tbWVyZ3J1YmVyQG1lZHVuaXdpZW4uYWMuYXQ=