
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Allergy, 29 April 2022
Sec. Asthma
Volume 3 - 2022 | https://doi.org/10.3389/falgy.2022.875295
This article is part of the Research TopicAirway Remodeling in Asthma - What is new?View all 7 articles
 Carolyn Damilola Ekpruke1
Carolyn Damilola Ekpruke1 Patricia Silveyra1,2*
Patricia Silveyra1,2*Asthma is characterized by an increase in the contraction and inflammation of airway muscles, resulting in airflow obstruction. The prevalence of asthma is lower in females than in males until the start of puberty, and higher in adult women than men. This sex disparity and switch at the onset of puberty has been an object of debate among many researchers. Hence, in this review, we have summarized these observations to pinpoint areas needing more research work and to provide better sex-specific diagnosis and management of asthma. While some researchers have attributed it to the anatomical and physiological differences in the male and female respiratory systems, the influences of hormonal interplay after puberty have also been stressed. Other hormones such as leptin have been linked to the sex differences in asthma in both obese and non-obese patients. Recently, many scientists have also demonstrated the influence of the sex-specific genomic framework as a key player, and others have linked it to environmental, social lifestyle, and occupational exposures. The majority of studies concluded that adult men are less susceptible to developing asthma than women and that women display more severe forms of the disease. Therefore, the understanding of the roles played by sex- and gender-specific factors, and the biological mechanisms involved will help develop novel and more accurate diagnostic and therapeutic plans for sex-specific asthma management.
According to the most recent National Health Interview survey data, current asthma patients represent about 8% of the United States population (1). Asthma is an inflammatory lung disease characterized by an increase in the contraction and inflammation of the airway and related muscles, resulting in airflow obstruction. In adults, the prevalence of asthma in females is higher than that of males (10.7 vs. 6.5% globally, and 9.8 vs. 6.1% in the US, respectively). While asthma prevalence is greater in male children than in females, it reverses at age 13 (which is mostly the onset of puberty) and continues to about age 65–70 years of adulthood (2). In 2019, the global prevalence of asthma was 136 per million in females, and 127 per million in males (3). In addition, females have been consistently shown to have a higher death rate due to asthma than males (1, 4, 5).
Airway remodeling and hyperresponsiveness in asthma have been documented long ago by many researchers to be the anatomical and physiological alterations that occur in the airway of an asthmatic individual. Some of the pro-inflammatory responses that trigger airway remodeling include the infiltration of eosinophils into the airway, recruitment of inflammatory cells, increase in the secretion of interleukins, increase in immunoglobulin E, shedding of epithelial layers, thickening of the subepithelial layer, increase in smooth muscle mass, increased secretion and sizes of mucous secreting cells, changes in blood vessels associated with the airway, and wearing-off of the soft bones of the airway (6–11). In general, males are known to be less prone to certain immunological illnesses compared to females and the role of sex hormones has been highlighted extensively (12–15). In both human and animal studies, clear sex distinctions have been documented in airway remodeling in asthma. The reasons for these differences have not been fully elucidated, but many studies have implicated the role of hormonal, immunological, occupational, and environmental factors (13, 16, 17). Hence, the purpose of this review is to discuss both sex and gender differences in airway remodeling in asthma, using data available from clinical and animal studies. Understanding the mechanisms underlying these disparities will guide the development of novel sex- and gender-specific diagnosis and therapeutic options for the management of asthma.
In clinical studies, researchers have attributed the sex differences in airway remodeling to anatomical and physiological differences in the structure and function of the respiratory system (Table 1). Others argue that the observed differences are majorly due to contributions of sex hormones and other sex-specific biochemical processes. Some of the anatomical observations reported are in the nasal cavity and floor, which is lower in females compared to males (18), and cranial airways which seem smaller in females than males (19). It has also been shown that the upper airway compliance during non-rapid eye movement sleep is lower in females than males though it was observed that the neck circumference and surface area of the body are also involved (20). Some earlier studies also reported observations in the pharynx, which is smaller in size and cross-sectional area and has low resistance in females than in males (21–23). These are areas of the respiratory system that are known to humidify and warm the inspired air, as most of them are lined with columnar epithelial cells that secrete mucus. They also participate in the mucocilliary clearance process when aerosol particles are inhaled, which makes them very important in allergic airway diseases. Overall, while the lung of adult males is bigger than that of females, it is also said to be age and stature-dependent (25). The size of the lungs influences the total lung capacity (TLC) of an individual, as TLC is directly proportional to the size of the lung. TLC also reflects the amount of air that goes in and out of the lungs, which is affected in asthma. Additionally, the number of alveoli is higher in adult males than females. Since the alveoli are the major sites where the exchange of gases occurs in the respiratory system, males having a larger surface area for gaseous exchange when compared to females, can influence asthma risk (25, 30, 31).
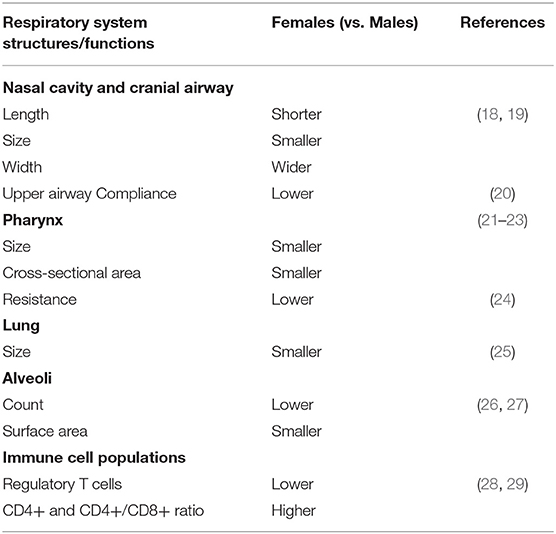
Table 1. Sex differences in the respiratory system structure and functions.
Sex dimorphism in immune response has been reported by many researchers both in animal and human studies. Innate and adaptive immune responses were said to be lower in males than in females. Many scientists have attributed the observed difference as being influenced by sex hormones. For instance, in animal studies, the activities of macrophages were lower in males compared to that of females attributing this to the protective role of male sex hormone (32). This report was substantiated with that the finding that antibody and cell-mediated responses were low in males than in females only when their testosterone level was at its peak (33, 34). In animal studies, regulatory T cells have been shown to play vital roles in mechanisms of inflammation in allergy, by preventing the production of cytokines such as TGF-beta and interleukin (IL)-10 (35), and proliferation of T cells (36), though the mechanisms involved are not clearly understood. These cells are very few in the lungs of female mice compared with that of the males (26, 27). Moreover, the subsets of CD4+ and CD8+ T cells were found to be numerous in the peripheral blood and lung tissue of asthmatic patients (37, 38). Inflammation mediated by allergens is said to be dependent on CD4 and CD8 cells rather than the previous belief that solely implicates immunoglobulin E and B cells since the lungs of knockout mice for immunoglobulin E and B still produce an allergic-mediated inflammation with house dust mite sensitization (39). CD8+ T cells are known for the role they play in immune tolerance, and they are a good source of proinflammatory cytokines in asthma (40) CD4+ cells and the ratio of CD4+/CD8+ cells (a marker of chronic lung disease) (41) are lower in males than females throughout adulthood (28, 29). Lamson et al. concluded that female mice express genes associated with adaptive immune response, while male mice express genes associated with innate immunity (42). Combined, these features may contribute to the observed sex differences in airway remodeling in asthma.
In clinical studies, a strong link has been established between the hypothalamic-gonadal-pituitary axis and the lungs (43). Many physiological functions of the lungs have been linked to the influence of different hormones throughout the life span. For instance, the influence of hormones on physiological surfactant secretion, lung development, and inflammatory markers production and functions have been documented by many researchers (28, 44–49). Estrogen has been named a key player in the quick maturation of surfactant-producing cells; this explains the faster development of female lungs compared with that of males (50–52). The production of surfactants also decreases as estrogen levels decline in females with an increase in age. On the other hand, androgens have an inhibitory effect on the production of surfactants at a young age (53). The role of surfactants has also been suggested in the recruitment of inflammatory cells in asthma (54). Similarly, a strong correlation between the production of surfactant and eosinophil counts has been shown, suggesting that it may serve as an immunomodulator (54).
Estrogen and its receptors (alpha and beta) have been demonstrated in human studies to play a vital role in the regulation of anatomical and physiological functions of the airway (55–59), as reviewed by us in (60). However, the role of estrogen and its receptors in airway remodeling in asthma has been controversial (61). Earlier animal studies reported suppression of the immune system with the increase in estrogen levels in the circulatory system (62). Hormonal fluctuations in the mouse estrous cycle influenced the expression of inflammatory genes in ozone-challenged female mice (63). Estrogen was also found to increase airway inflammation by enhancing the activities of the T-helper cells type 2 in asthma (14). Thus, it has been suggested that the role of estrogen can be both destructive and advantageous depending on whether the alpha or the beta estrogen receptor signaling pathway is in use.
The nuclear estrogen receptors alpha and beta are both found in the respiratory system but different proportions (62). One study reported that the activation of the beta estrogen receptor showed a reduction of extracellular matrix in asthmatic humans by suppressing the activity of the NF-kB pathway (58). Another study showed that beta estrogen receptor activation led to the proliferation of airway smooth muscle cells by inhibiting the activities of the platelet cells (55). The same study showed that estrogen inhibits smooth muscle contraction by reducing calcium ion influx during inflammation in asthmatic conditions (57).
Another female hormone, progesterone, and its receptors (alpha and beta receptors) have been implicated in sex differences in asthma (16). This hormone is present in both males and females, but the level is higher in females than in males (64). Many decades ago, progesterone was found to enhance the dilation of bronchi (65–67). However, there is little information available for the role it plays in the allergic immune response. Some researchers have documented that progesterone contributes to the effects of other sex hormones. For instance, it is known to have a high affinity for the enzyme 5-alpha reductase that helps in the conversion of testosterone to active 5-dihydrotestosterone (64). Other studies have shown that testosterone is inactive in the presence of progesterone (68). In postmenopausal women undergoing hormonal therapy, the combination of progesterone and estrogen increases their risk of developing asthma, though the associated mechanisms have not been studied (69). A positive correlation of serum progesterone and peak flow rate in different menstrual phases was also found (70). Also, progesterone was said to regulate the production and activities of IL-17 which was enhanced in women with severe asthma (71). Similarly, human airway epithelial cells treated with progesterone display a reduced frequency of cilia movement, indicating that progesterone negatively affects the functions of the micro ciliary apparatus (72). Since the levels of this hormone are higher in females than in males, it is possible that progesterone fluctuations in females contribute to asthma susceptibility.
Regarding male hormones, testosterone and its associated metabolites also play a significant role in modulating T cell activity, which helps provide an equilibrium between hypersensitivity reactions and the body's defense system. Testosterone is classified as an immune-protective hormone, along with 5-alpha dihydrotestosterone (5-alpha DHT). Both help reduce airway inflammation in asthma by reducing the response of the innate and adaptive immunity (14).
Adipose tissue is known for its ability to store energy in form of triglycerides, recently, it becomes a recognized endocrine organ. It is known to secrete hormones such as adiponectin, C1a-TNF related protein 9, retinol-binding protein 4, leptin, and omentin (73, 74). This group of hormones is referred to as the adipokines in addition to the proinflammatory and anti-inflammatory cytokines secreted by the same tissue. All these go into the circulatory system where they mediate the activities between adipose tissue and other tissues/ organs of the body (75). The ability of the adiponectin to produce more than one effect makes it an interesting adipokine to study among researchers. It is a general belief that it has an anti-inflammatory property. This was demonstrated in the research of (76), though it was not in lung disease cardiovascular system disease. Adiponectin is known to carry out its anti-inflammatory effect by acting on the macrophages through the prevention of progenitor myeloid cells differentiation (77, 78). It also possesses the ability to alter the activities of the macrophages and toll-like receptor 4 (79). The role of some adipokines in pulmonary diseases has been reviewed by (80).
Leptin is a hormone of great interest in airway remodeling in asthma, as its levels are known to be enhanced during allergic reactions (81). A few studies tried to implicate this hormone as a key player in the strong association existing between obesity and asthma. However, there are very few studies on the roles of this hormone and its mechanisms of action. There is a great expression of leptin receptors (alpha and beta) in bronchoalveolar epithelial cells and alveolar macrophages (82–84), as well as in other immune cells (85, 86). Decades ago, leptin was shown to stimulate the release of IL-6 and tumor necrosis factors from adipose tissue cells (87, 88). Interleukin 6 is known for its role in the release of the inflammatory marker, C-reactive protein, from the liver in inflammatory conditions (89, 90), as well as in mediating interferon production in T helper 1 cells (91, 92). In mice, the level of macrophage inflammatory protein-2 was directly proportional to the serum level of leptin in ozone-induced airway inflammation (93). Leptin has also been found associated with lung injury and asthma (94). The levels of leptin were found to be higher in children with asthma compared with healthy controls (95). In the same study, the non-asthma group displayed sex differences in leptin levels, whereas no sex differences were observed in the asthma group. In mouse studies, leptin administered exogenously led to an increase in secretions of tumor necrosis factor, IL-6, and IL-12 (95).
Multiple researchers have demonstrated the role of hormones and their various receptors in airway remodeling in asthma using experimental animals. Of great interest, the role of estrogen and its receptors, alpha, and beta, have been documented. The importance of both receptors was seen in lung maturation and the size of alveoli, but the beta estrogen receptor was found to contribute to a larger extent than the alpha in lung elasticity. The effect was more pronounced in male vs. female mice knockout for the beta estrogen receptor (49). In addition, gonadectomized ovalbumin-asthma-induced mice showed a significant increase in infiltration of eosinophils, lymphocytes, and some interleukins in their airway compared to control mice. Dimitropolou et al. also demonstrated the role of estrogen in mouse isolated trachea rings sensitized with serum from asthma and healthy patient (96). Contraction occurred in the trachea ring sensitized by asthmatic serum when carbachol was added but when the same sample was pretreated with estrogen, the contraction was abolished. The authors concluded that the estrogen beta receptor was activated, decreasing the contraction through the stimulation of potassium channels. Other researchers reported sex differences in airway remodeling and attributed the effects to sex hormones. In one study by Riffo-Vasquez et al. female mice that have undergone ovariectomy before being sensitized with ovalbumin showed a reduction in IL-5 levels, eosinophil infiltration, and hyperresponsiveness to methacholine compared to control mice (97). Similarly, Takeda et al. reported a reduction in eosinophil counts, production of serum IgE and hypersensitivity of the airway in ovariectomized mice challenged with ovalbumin or house dust mites (98). Also, in ovalbumin-sensitized females, an increase in airway hyperresponsiveness was observed with no effect on inflammation of the airway after the activation of the alpha estrogen receptor (99).
The role of progesterone and its receptors on airway remodeling in asthma has not been widely studied in experimental animals. Administration of progesterone contraceptives to females in an influenza A mouse model helped to enhance lung function and the repair of the damaged epithelium caused by the infection and inflammation (100). In contrast, Hellings et al. rather reported a worsened airway disease in mice following the exogenous administration of progesterone as it led to an increase in infiltration of eosinophils to the airway and hyperresponsiveness of the airway (101). In type 2 helper cell-mediated immune responses, progesterone stimulates the production of some proinflammatory cytokines (102). This study also confirmed that the female sex hormones play important roles in airway remodeling in asthma.
Male sex hormones have been also documented to play a role in inflammation of the airway mediated by type 2 cells in animal studies. A study by Yu et al. investigated the role of male sex hormones and their derivatives on airway remodeling in asthma (103). By adding DHEA (dehydroepiandrosterone) to the diet of house dust mite sensitized mice, they observed a significant increase in the resulting airway inflammation and infiltration of eosinophils and interleukins into the airway compared to mice eating normal chow, though there was no change in the immunoglobulin E level of both groups (103). The study did not consider sex as a biological variable.
Sex differences in biomarkers of type 2 inflammation expressed in asthma have been widely investigated (Table 2). Clinical studies have reported sex differences in asthma control by measuring such inflammatory biomarkers, blood eosinophils, exhaled nitric oxide, and serum E levels, and indicated a significantly high symptom control in males compared to the females in the same age group (104, 110). Other researchers found no differences in C-reactive protein in asthma (108). In animal studies of ovalbumin-induced asthma, infiltration of eosinophils, as well as the concentration of serum immunoglobulin E (IgE) and IL-3., were found to be increased in the lungs of female mice when compared to males (98, 111). Similarly, in studies where sex differences in airway-remodeling were hypothesized, the serum concentration of IgE was found to be increased in the female vs. male lungs in two different asthma models used (ovalbumin challenge and house mite dust exposure) (109). The responsiveness of the airway to methacholine was also higher in females than in males. Card et al. also observed that male mice displayed higher airway hyperresponsiveness during methacholine challenge than females (112). This study is one of very few whose observations reflect that male lungs are more affected by asthma than females. The studies of Melgert et al. (26) and Okuyama et al. (113) support the fact that females are more susceptible to airway inflammation caused by ovalbumin challenge than males (26, 113). Both studies observed an increase in airway hyperresponsiveness, eosinophil, T, and B cell counts, and level of cytokines in female vs. male mice challenged with ovalbumin. Moreover, Treg cells (known for their vital role in the prevention of inflammation in allergy) were found lower levels in the lungs of female mice compared to males (26).
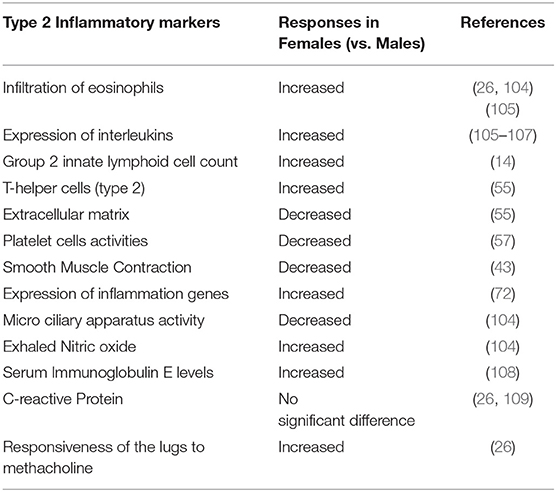
Table 2. Sex-difference in biomarkers of type 2 inflammation: comparing male and female responses.
The recently discovered type 2 innate lymphoid cells (ILCs) have been reported to increase in peripheral blood (114, 115) and sputum (116) of patients with asthma. They are known to mediate allergy responses in the lungs (117–119). This was further substantiated by a study reporting an increase in circulating type 2 ILCs counts in women compared with men in a population with moderate to severe asthma (105), where 5-alpha DHT decreased these cell counts and expression of IL-5 and IL-13, both involved airway inflammation and hyperresponsiveness (117). These type 2 ILCs have an inflammatory subset that is found in the lungs and can move in the different mucous-producing sites during airway remodeling in asthma (120). One of the major roles of these cells is the expression of GATA binding protein 3 and inflammatory cytokines. In the lungs, the transformation of precursor ILCs to type 2 ILCs depends on the GATA binding protein 3. Interestingly, these cells are found to be more numerous in females than males (121). The inflammatory subset of type 2 ILCs activates the lectin receptor G1 (116), which is also higher in females. This special subset of type 2 ILCs is known for its role in cytokine production, and it also increases in number with age. To explain the sex difference observed, studies in gonadectomized and estrogen alpha knock-out mice showed that male sex hormones, but not female hormones, regulate the proliferation and function of type 2 ILCs (121). This agrees with the findings of Warren et al. who observed an increase in IL-33 production by type 2 ILCs in female vs. male mice sensitized with ovalbumin (106). Moreover, Laffont et al. showed that the androgen receptor signaling reduced type 2 ILCs proliferation (122). They also observed a sex disparity in the lymphoid cell counts, which was abolished, and type 2 mediated inflammation restored, after orchiectomy.
Several epidemiological studies have attributed the sex differences in airway remodeling in asthma to environmental, occupational, and social factors. Hence, it is important to distinguish sex as a biological factor vs. gender as a social construct in these analyses (123). Female gender and tobacco smoke have been greatly associated with severe refractory asthma (124).
Occupational factors have also been implicated in the observed sex differences of airway remodeling in asthma. Gendered roles and changes in occupations traditionally performed by men or women can influence asthma development (125). Recently, females were reported to work in highly polluted places like hospitals, homes, and schools, and thus display a higher frequency of work-related asthma was observed in females than in males (126). Interestingly, women are also known to have more pets at home (127), and are negatively affected by secondary exposure to tobacco smoke (127) than men.
Another factor potentially contributing to sex differences in asthma is exercise (128). While a few studies have documented sex differences in immune cell counts (129–131), plasma cytokine levels (129, 131), and lymphocyte apoptosis (132) with different kinds of exercise, others reported no differences, particularly in treadmill running, bicycle, and strength training (133–135). In women, a few studies have linked immune response changes during exercise to the menstrual cycle. For instance, a study in cyclists observed an alteration in leukocytes and cytokine expression in female cyclists during the menstrual phase (134). Others found that regulation of inflammatory genes depends on the time and duration of exercise during the menstrual phase in females (136). In this context, female athletes were found to exhibit severe exercise-induced bronchoconstriction in the luteal phase of the cycle (137). A potential mechanism for this involves differential expression of pro-inflammatory genes. Northoff et al. found that proinflammatory genes are upregulated in the follicular phase, while anti-inflammatory genes are downregulated in the luteal phase in females compared with males (136). Women in the mid-luteal phase also display worsened lung function and exacerbated bronchoconstriction induced by exercise (138). These changes in the mid-luteal phase were associated with an increase in progesterone levels, although the exact mechanism remains unknown (137).
Obesity, body mass index, and serum IgE have been associated with asthma across the life span (139, 140). A study in a mixed population of smokers without any respiratory disorders showed a strong association between fat distribution and normal lung function (141). Other studies have shown sex specific associations, including a strong association between new asthma symptoms and high body mass index only in females (142, 143), and a higher prevalence of obesity in women with vs. without asthma (124, 144). In a study of 4,197 asthma patients from the 2012 National Health Interview Survey, a positive association of obesity and body mass index with asthma (overall and allergic) was found (145). The authors also reported that the association of class III obesity (BMI≥40) and asthma was stronger in women. Similarly, epidemiological studies have shown that asthma occurring with obesity (also known as “obesity-associated asthma) is more difficult to treat, more severe, and more prevalent in women (146–148). Finally, mouse models of asthma have also shown that high fat diet and the consequent weight gain contribute to the progression of allergic asthma with females displaying airway remodeling phenotypes at earlier stages than males (149, 150).
The interaction between genetics and environmental factors has also been documented as a contributing aspect in females with asthma and obesity (127, 151, 152), or males (153, 154). However, some studies have reported that the association between asthma and obesity is not gender-based (155). Though this association has been an issue of debate among many researchers (156–158) there is a need to determine the physiological parameters and hormonal influences on airway remodeling and asthma. As indicated earlier, leptin levels correlate with body mass index (92, 159), and human adipose tissue expresses inflammatory cytokines (87, 160). The mechanism through which obesity is linked to asthma has not been fully understood as very few or no animal studies have been carried out in this area.
There is a paucity of data concerning the influence of genetic framework and pathways on sex differences in airway remodeling in asthma. The genomic framework of males and females with asthma displays numerous differences as described in Figure 1. Gautam et al. identified differentially expressed genes (n = 32), as well as both male and female-specific genes (males = 439; females = 299) (161), of which five are impaired in the regulation of physiological processes during asthma. Of these genes, four were male-specific while only one was female-specific (161). They discovered that the majority of these genes were found in the airway epithelium. The four genes specific to the males included FBXL7, ITPR3, RAD51B, and ALOX15; three of them were found in the airway epithelial tissue, while only ALOX15 was found in the blood. All these genes are upregulated in asthma except RAD51B. The only gene that was specific to the female is HLA_DQA1 (downregulated) and also found in the airway epithelial tissue.

Figure 1. Sex differences in the expression of genes associated with asthma.
Some biological pathways influence the sex difference observed in asthma, and these include HIF- alpha 1 and IL-17 signaling. The HIF- alpha 1 signaling pathway has been mostly identified in male-specific differentially expressed genes (161). This pathway is known to play a role in the regulation of pro-inflammatory cytokines, chemokines in processes of infection and allergy. However, the role of this pathway in asthma, and specifically sex differences in airway remodeling has not been studied. Regarding IL-17 pathways, they have been found upregulated in females with asthma (161–163), but negatively correlated with airway hyperresponsiveness to methacholine (164). This IL-17 signaling pathway is known for its role in stimulating the cells of the epithelium to produce cytokines that help to recruit neutrophils to the inflammation site (165). Excess production of IL-17A has been linked to the risk of developing severe asthma (166–169). This was substantiated by the research of Laan et al. where it was reported that stimulation of the human epithelium with IL-17A led to greater expression of CXCL8 mRNA, which in turn enhanced neutrophils migration to the inflammation site. Meanwhile, Busse et al. documented that IL-17A does not affect the severity of asthma when patients were treated with brodalumab, a human anti-IL-17 receptor A monoclonal antibody (170).
Many studies focused on sex differences in asthma after adolescence identified that changes in DNA methylation in blood and lung tissue could play a role in the observed disparities (171–178). Furthermore, the interaction between sex-specific and sex-stratified genomes has also been found associated with childhood asthma (179). In this regard, the 17q12-21-asthma locus was highlighted to be associated with asthma in both males and females but was widely significant in females (179). Similarly, the ligand-dependent nuclear receptor co-repressor-like gene, located within the regulatory region was highlighted in males only (179). This gene peculiar to the males only is known to play a vital role in the determination of height and sperm production.
Asthma risk alleles have also been identified in a genome-wide interaction study. Four (2q23.3, 2q34, 6q27, and 17213.3) of these alleles are specific to females, and two (5q31.1 and 10q26.1) are specific to males (180). Some single nucleotide polymorphisms (SNPs) that are sex-specific and associated with asthma were discovered in beta 2 adrenergic receptors (181) and thymic stromal lymphopoietin (TSLP) (182). The beta 2 adrenergic receptor was observed to have genetic variants that are associated with the development of severe asthma (181). On other hand, TSLP is a cytokine similar to IL-17 produced by the cells of the epithelium during an allergic reaction. It is known to be associated with serum IgE in girls. It is also known to play a crucial role in the regulation of allergic responses, specifically, airway inflammation in animal models (183). Recently, this observation was substantiated in TSLP knockout mouse studies (184, 185). The specific SNPs discovered in TSLP include rs1837253, associated with the risk of asthma in males, and r2289276 in females only (182). Gauderman et al. also identified two other loci, GRIA2 and TNTRSFIIB associated with gene sex and childhood asthma using a genome-wide interaction scan in children exposed to traffic air pollution (186). The influence of sex-interact in the association between interferon-gamma gene, a protein-coding gene, and childhood asthma was also reported (187). This study showed that genotype-sex interactions on asthma were only significant at rs2069727 and rs2430561interferon gamma SNPs though the link was not additive. These genes have shown effects on interferon's response to bacterial infections and developing asthma in early childhood (187). Some researchers have argued that the genotype-sex interaction and asthma were different in each race. For instance, a gene variant known as KCNMBI was specifically identified in the African American race and is known to play a role in the contraction of bronchial smooth muscle, hence, influencing the pulmonary function (188). This gene has some other variants, of great interest is the 8,187 allele that was seen to decrease the lung function concerning low FEV1% recorded; the damaging effect of the allele was suppressed through an estrogen-mediated upregulation of the high conductance voltage calcium-potassium channel in animal experiments (188).
Epigenetics include changes in the genetic materials that not affect the DNA sequence (189). These changes are characterized by methylation of DNA, modification of histones, and microRNAs (190). Epigenetic changes can be caused by diet, air pollution exposure, tobacco smoking, and drug administration, among other factors, and can occur in any stage of life (190). The external environment is in direct contact with the lungs, hence epigenetic alterations occur in the respiratory tract. Associations of epigenetic changes, including DNA hypermethylation or hypomethylation have been identified with asthma (191–193). The hypomethylated or hypermethylated level in specific genes varies depending on the asthma phenotype. For example, hypermethylation of genes such as ARG1, ARG2 and ADAM33 in buccal cells and bronchial epithelial cells have been correlated with asthma phenotypes (194, 195). On the other hand, DNA hypomethylation has been associated with IL6 and ADAM33 expression in bronchial fibroblasts and the nasal epithelium (194, 195). As recently reported in a review by Chowdhury et al. certain CpG sites located in the interferon-related developmental regulator 1 (IFRD1) gene have been linked to sex-specific effects in asthma (140). However, there are very few studies that discussed the issue of sex differences in asthma associated with epigenetics. One of such research include the Ascaris exposure that was said to lower lung function and increase the risk of asthma development at a higher rate in males than females (196).
The major characteristics of asthma are airway remodeling with evidence of airway inflammation, increased production of mucus, increased migration of eosinophils to the airway, and airway hypersensitivity. These processes are highly regulated by the expression of inflammatory genes. MiRNAs are a class of small non-coding RNAs that play important roles in gene expression regulation (197). miRNAs are known to be associated with many respiratory disorders including asthma (198, 199) by serving as a biomarker, and mediating interactions among cells (200–202). There is a paucity of data about the sex-specific miRNA expression in asthma. However, immune responses have been known to be sex-specific and miRNAs are said to play a role in this sex bias as shown in Figure 2. Animal studies of ozone-induced lung inflammation reported sex differences in lung miRNA expression (203). In this study, nine sex-specific miRNAs were found in the ozone-induced inflammation group (203). Six of the identified miRNAs were greatly expressed (upregulated) in males (miR-338, miR-222-3p, miR-130b-3p, let 7i-5p, miR-195a-5p, miR-144-3p) while five were highly expressed (upregulated) in females (miR-301b-3p, miR-694, miR-669h-3p, miR-384-5p, miR-9-5p). Only one was downregulated in females (miR-30d5p). They also noted that there were two miRNAs (miR-712-5p, miR-106a-5p) that were expressed in both males and females exposed to ozone but with different patterns by sex. They suggested the role of hormones in the pattern of expression. In a study by another group of animals exposed to smoke in utero, three miRNAs (miR-153-2; miR-196a; miR-184) were identified in both sexes but they were differentially expressed only in males during normal lung development (204). Also, using serum from asthma patients, Kho et al. identified 22 miRNAs that were associated with lung function, nine of which were solely expressed in males (miR-139-5p; miR-156-5p, miR-186-5p, miR-342-3p; miR-374a-5p, miR-409-3p, miR-454-3p, miR-660-5p; miR-942-5p) while only three were expressed in females (miR-1290; miR-142-3p; miR-191-5p) (205). In addition, miR-126 was found to be expressed in both sexes (206). Other researchers have also identified miRNAs associated with immune system regulation (207, 208), type 2 immunity (209), regulation of allergic-induced inflammation in asthma (210), and asthma pathogenesis (211). The sex-specific expression of these miRNAs suggests their role in the sex-disparity of airway remodeling in asthma.

Figure 2. Sex-specific miRNAs associated with airway inflammation and remodeling.
Asthma is a lung inflammatory disease with clear sex differences in incidence, prevalence, and severity across the life span. In both clinical and animal studies, airway remodeling in asthma is known to be characterized by the alterations in the airway leading to obstruction of airflow. The prevalence of asthma suddenly changes at the onset of puberty. Whether this switch is also reflected in airway remodeling has been an issue of debate among researchers over many decades. Many scientists have attributed it to the differences in the structure and functions of the respiratory system, including the nasal cavity, upper airway compliance, lung size, alveoli, and the population of immune system cells in males and females. In great curiosity, many studies linked these sex disparities to changes in sex hormones at puberty, since there is usually a great alteration in the level of these hormones majorly at the onset of puberty. The role of estrogen, testosterone, and progesterone alongside their receptors has been well documented to date. Another hormone implicated in this observation is leptin, although there is still a paucity of data concerning the mechanisms involved and the role it plays in airway remodeling in asthma. The sex differences in biomarkers of type-2-inflammation have also been reported, many of which were suggested to be protective in the male respiratory system, suggesting that females are more susceptible to asthma than males. More recently, researchers have tried to link the observed sex differences in asthma to the genomic frameworks of males and females. This led to the discovery and identification of many sex-specific genes, gene variants, and miRNAs that are directly linked to lung function, lung inflammation, and asthma in general, though much research is still ongoing in this area. Another major concern is the strong link between asthma and each of occupational, environmental, and lifestyle factors that are stronger in females than in males. Many of these observations were from epidemiological studies, thus more experimental studies in this area are highly needed to identify sex-specific mechanisms and pathways involved. However, it is very important to review lessons learned from both clinical and animal studies as this helped us identify the gaps that are needed to be filled to justify the sex difference of airway remodeling in asthma. This will help to intensify efforts in such areas to identify proper sex-specific diagnosis and therapeutic pathways in the management of asthma.
CE and PS contributed to the conception of the study and wrote sections of the manuscript. CE organized the database, extracted information, and wrote the first draft of the manuscript. All authors contributed to manuscript revision, read, and approved the submitted version.
This study was supported by the National Institutes of Health, R01HL159764 and R03HL141618 (PS).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Pate CA, Zahran HS, Qin X, Johnson C, Hummelman E, Malilay J. Asthma surveillance - United States, 2006-2018. MMWR Surveill Summ. (2021) 70:1–32. doi: 10.15585/mmwr.ss7005a1
2. Mandhane PJ, Greene JM, Cowan JO, Taylor DR, Sears MR. Sex differences in factors associated with childhood-and adolescent-onset wheeze. Am J Respir Crit Care Med. (2005) 172:45–54. doi: 10.1164/rccm.200412-1738OC
3. Safiri S, Carson-Chahhoud K, Karamzad N, Sullman MJM, Nejadghaderi SA, Taghizadieh A, et al. Prevalence, deaths, and disability-adjusted life-years due to asthma and its attributable risk factors in 204 countries and territories, 1990-2019. Chest. (2022) 161:318–29. doi: 10.1016/j.chest.2021.09.042
4. Zein JG, Udeh BL, Teague WG, Koroukian SM, Schlitz NK, Bleecker ER, et al. Impact of age and sex on outcomes and hospital cost of acute asthma in the United States, 2011-2012. PLoS One. (2016) 11:e0157301. doi: 10.1371/journal.pone.0157301
5. Busse WW, Kraft M. Current unmet needs and potential solutions to uncontrolled asthma. Eur Respir Rev. (2022) 31:210176. doi: 10.1183/16000617.0176-2021
6. Haraguchi M, Shimura S, Shirato K. Morphometric analysis of bronchial cartilage in chronic obstructive pulmonary disease and bronchial asthma. Am J Respir Crit Care Med. (1999) 159:1005–13. doi: 10.1164/ajrccm.159.3.9712144
7. Hammad H, Lambrecht BN. The basic immunology of asthma. Cell. (2021) 184:1469–85. doi: 10.1016/j.cell.2021.02.016
8. Metcalfe DD, Baram D, Mekori YA. Mast cells. Physiol Rev. (1997) 77:1033–79. doi: 10.1152/physrev.1997.77.4.1033
9. Carroll N, Elliot J, Morton A, James A. The structure of large and small airways in nonfatal and fatal asthma. Am Rev Respir Dis. (1993) 147:405–10. doi: 10.1164/ajrccm/147.2.405
10. Li X, Wilson JW. Increased vascularity of the bronchial mucosa in mild asthma. Am J Respir Crit Care Med. (1997) 156:229–33. doi: 10.1164/ajrccm.156.1.9607066
11. Tanaka H, Yamada G, Saikai T, Hashimoto M, Tanaka S, Suzuki K, et al. Increased airway vascularity in newly diagnosed asthma using a high-magnification bronchovideoscope. Am J Respir Crit Care Med. (2003) 168:1495–9. doi: 10.1164/rccm.200306-727OC
12. Ansar Ahmed S, Penhale WJ, Talal N. Sex hormones, immune responses, and autoimmune diseases. Mechanisms of sex hormone action. Am J Pathol. (1985) 121:531–51.
13. Yung JA, Fuseini H, Newcomb DC. Hormones, sex, and asthma. Ann Allergy Asthma Immunol. (2018) 120:488–94. doi: 10.1016/j.anai.2018.01.016
14. Fuseini H, Newcomb DC. Mechanisms driving gender differences in asthma. Curr Allergy Asthma Rep. (2017) 17:19. doi: 10.1007/s11882-017-0686-1
15. Naeem A, Silveyra P. Sex differences in paediatric and adult asthma. Eur Med J (Chelmsf). (2019) 4:27–35.
16. Silveyra P, Fuentes N, Rodriguez Bauza DE. Sex and gender differences in lung disease. Adv Exp Med Biol. (2021) 1304:227–58. doi: 10.1007/978-3-030-68748-9_14
17. Shah R, Newcomb DC. Sex bias in asthma prevalence and pathogenesis. Front Immunol. (2018) 9:2997. doi: 10.3389/fimmu.2018.02997
18. García-Martínez D, Torres-Tamayo N, Torres-Sánchez I, García-Río F, Rosas A, Bastir M. Ribcage measurements indicate greater lung capacity in Neanderthals and lower pleistocene hominins compared to modern humans. Commun Biol. (2018) 1:117. doi: 10.1038/s42003-018-0125-4
19. Bastir M, Godoy P, Rosas A. Common features of sexual dimorphism in the cranial airways of different human populations. Am J Phys Anthropol. (2011) 146:414–22. doi: 10.1002/ajpa.21596
20. Rowley JA, Sanders CS, Zahn BR, Badr MS. Gender differences in upper airway compliance during NREM sleep: role of neck circumference. J Appl Physiol. (2002) 92:2535–41. doi: 10.1152/japplphysiol.00553.2001
21. Brown I, Zamel N, Hoffstein V. Pharyngeal cross-sectional area in normal men and women. J Appl Physiol. (1986) 61:890–5. doi: 10.1152/jappl.1986.61.3.890
22. White DP, Lombard RM, Cadieux RJ, Zwillich C. Pharyngeal resistance in normal humans: influence of gender, age, and obesity. J Appl Physiol. (1985) 58:365–71. doi: 10.1152/jappl.1985.58.2.365
23. Brooks LJ, Strohl KP. Size and mechanical properties of the pharynx in healthy men and women. Am Rev Respir Dis. (1992) 146:1394–7. doi: 10.1164/ajrccm/146.6.1394
24. Bellemare F, Jeanneret A, Couture J. Sex differences in thoracic dimensions and configuration. Am J Respir Crit Care Med. (2003) 168:305–12. doi: 10.1164/rccm.200208-876OC
26. Melgert B, Postma D, Kuipers I, Geerlings M, Luinge M, Van der Strate B, et al. Female mice are more susceptible to the development of allergic airway inflammation than male mice. Clin Exp Allergy. (2005) 35:1496–503. doi: 10.1111/j.1365-2222.2005.02362.x
27. Cook MB, Dawsey SM, Freedman ND, Inskip PD, Wichner SM, Quraishi SM, et al. Sex disparities in cancer incidence by period and age. Cancer Epidemiol Biomarkers Prev. (2009) 18:1174–82. doi: 10.1158/1055-9965.EPI-08-1118
28. Uppal SS, Verma S, Dhot PS. Normal values of CD4 and CD8 lymphocyte subsets in healthy indian adults and the effects of sex, age, ethnicity, and smoking. Cytometry B Clin Cytom. (2003) 52:32–6. doi: 10.1002/cyto.b.10011
29. Rubtsov AV, Rubtsova K, Fischer A, Meehan RT, Gillis JZ, Kappler JW, et al. Toll-like receptor 7 (TLR7)–driven accumulation of a novel CD11c+ B-cell population is important for the development of autoimmunity. Blood J Am Soc Hematol. (2011) 118:1305–15. doi: 10.1182/blood-2011-01-331462
30. Cohen J, Douma WR, Ten Hacken NH, Oudkerk M, Postma DS. Physiology of the small airways: agender difference? Respir Med. (2008) 102:1264–71. doi: 10.1016/j.rmed.2008.04.007
31. Massaro GD, Mortola JP, Massaro D. Sexual dimorphism in the architecture of the lung's gas-exchange region. Proc Natl Acad Sci U S A. (1995) 92:1105–7. doi: 10.1073/pnas.92.4.1105
32. Mondal S, Rai U. Sexual dimorphism in phagocytic activity of wall lizard's splenic macrophages and its control by sex steroids. Gen Comp Endocrinol. (1999) 116:291–8. doi: 10.1006/gcen.1999.7370
33. Pap PL, Czirják GÁ, Vágási CI, Barta Z, Hasselquist D. Sexual dimorphism in immune function changes during the annual cycle in house sparrows. Naturwissenschaften. (2010) 97:891–901. doi: 10.1007/s00114-010-0706-7
34. Fargallo JA, Martínez-Padilla J, Toledano-Díaz A, Santiago-Moreno J, Davila JA. Sex and testosterone effects on growth, immunity and melanin coloration of nestling Eurasian kestrels. J Anim Ecol. (2007) 76:201–9. doi: 10.1111/j.1365-2656.2006.01193.x
35. Akbari O, Stock P, DeKruyff RH, Umetsu DT. Role of regulatory T cells in allergy and asthma. Curr Opin Immunol. (2003) 15:627–33. doi: 10.1016/j.coi.2003.09.012
36. Kondelkova K, Vokurková D, Krejsek J, Borská L, Fiala Z, Ctirad A. Regulatory T cells (TREG) and their roles in immune system with respect to immunopathological disorders. Acta Medica (Hradec Kralove). (2010) 53:73–7. doi: 10.14712/18059694.2016.63
37. Lourenço O, Fonseca AM, Taborda-Barata L. T cells in sputum of asthmatic patients are activated independently of disease severity or control. Allergol Immunopathol. (2009) 37:285–92. doi: 10.1016/j.aller.2009.05.004
38. Hirosako S, Goto E, Tsumori K, Fujii K, Hirata N, Ando M, et al. CD8 and CD103 are highly expressed in asthmatic bronchial intraepithelial lymphocytes. Int Arch Allergy Immunol. (2010) 153:157–65. doi: 10.1159/000312633
39. Raemdonck K, Baker K, Dale N, Dubuis E, Shala F, Belvisi MG, et al. CD4+ and CD8+ T cells play a central role in a HDM driven model of allergic asthma. Respir Res. (2016) 17:45. doi: 10.1186/s12931-016-0359-y
40. Hinks TSC, Hoyle RD, Gelfand EW. CD8+ Tc2 cells: underappreciated contributors to severe asthma. Eur Respir Rev. (2019) 28:190092. doi: 10.1183/16000617.0092-2019
41. Triplette M, Attia EF, Akgün KM, Soo Hoo GW, Freiberg MS, Butt AA, et al. A low peripheral blood CD4/CD8 ratio is associated with pulmonary emphysema in HIV. PLoS ONE. (2017) 12:e0170857. doi: 10.1371/journal.pone.0170857
42. Lamason R, Zhao P, Rawat R, Davis A, Hall JC, Chae JJ, et al. Sexual dimorphism in immune response genes as a function of puberty. BMC Immunol. (2006) 7:2. doi: 10.1186/1471-2172-7-2
43. Fuentes N, Silveyra P. Endocrine regulation of lung disease and inflammation. Exp Biol Med. (2018) 243:1313–22. doi: 10.1177/1535370218816653
44. Ojeda M, Gómez N, Gil E, Scardapane L, Gimenez M. Morphologic and biochemical changes in male rat lung after surgical and pharmacological castration. Braz J Med Biol Res. (2000) 33:279–85. doi: 10.1590/S0100-879X2000000300004
45. Sammohi S. Effect of progesterone, terbutaline and leptin on the function of alveolar type ii cells (MSc. thesis). Wright State University, Dayton, OH, United States (2015). Available online at: https://corescholar.libraries.wright.edu/cgi/viewcontent.cgi?article=3179&context=etd_all
46. Trotter A, Hilgendorff A, Kipp M, Beyer C, Kueppers E, Kiossis E, et al. Gender-related effects of prenatal administration of estrogen and progesterone receptor antagonists on VEGF and surfactant-proteins and on alveolarisation in the developing piglet lung. Early Hum Dev. (2009) 85:353–9. doi: 10.1016/j.earlhumdev.2008.12.013
47. Kirwin SM, Bhandari V, Dimatteo D, Barone C, Johnson L, Paul S, et al. Leptin enhances lung maturity in the fetal rat. Pediatr Res. (2006) 60:200–4. doi: 10.1203/01.pdr.0000227478.29271.52
48. Huang K, Rabold R, Abston E, Schofield B, Misra V, Galdzicka E, et al. Effects of leptin deficiency on postnatal lung development in mice. J Appl Physiol. (2008) 105:249–59. doi: 10.1152/japplphysiol.00052.2007
49. Massaro D, Massaro GD. Estrogen receptor regulation of pulmonary alveolar dimensions: alveolar sexual dimorphism in mice. Am J Physiol Lung Cell Mol Physiol. (2006) 290:L866–70. doi: 10.1152/ajplung.00396.2005
50. Nielsen HC, Torday JS. Sex differences in fetal rabbit pulmonary surfactant production. Pediatr Res. (1981) 15:1245–7. doi: 10.1203/00006450-198109000-00004
51. Torday J, Dow K. Synergistic effect of triiodothyronine and dexamethasone on male and female fetal rat lung surfactant synthesis. Dev Pharmacol Ther. (1984) 7:133–9. doi: 10.1159/000457153
52. Torday JS. The sex difference in type II cell surfactant synthesis originates in the fibroblast in vitro. Exp Lung Res. (1984) 7:187–94. doi: 10.3109/01902148409087912
53. Nielsen HC, Zinman HM, Torday JS. Dihydrotestosterone inhibits fetal rabbit pulmonary surfactant production. J Clin Invest. (1982) 69:611–6. doi: 10.1172/JCI110488
54. Choi Y, Lee DH, Lee JH, Shin YS, Kim SH, Park HS. Immunomodulatory function of surfactant protein D in eosinophilic asthma. Allergy. (2019) 74:192–5. doi: 10.1111/all.13588
55. Ambhore NS, Katragadda R, Raju Kalidhindi RS, Thompson MA, Pabelick CM, Prakash YS, et al. Estrogen receptor beta signaling inhibits PDGF induced human airway smooth muscle proliferation. Mol Cell Endocrinol. (2018) 476:37–47. doi: 10.1016/j.mce.2018.04.007
56. Aravamudan B, Goorhouse KJ, Unnikrishnan G, Thompson MA, Pabelick CM, Hawse JR, et al. Differential expression of estrogen receptor variants in response to inflammation signals in human airway smooth muscle. J Cell Physiol. (2017) 232:1754–60. doi: 10.1002/jcp.25674
57. Kalidhindi RSR, Ambhore NS, Bhallamudi S, Loganathan J, Sathish V. Role of estrogen receptors α and β in a murine model of asthma: exacerbated airway hyperresponsiveness and remodeling in ERβ knockout mice. Front Pharmacol. (2020) 10:1499. doi: 10.3389/fphar.2019.01499
58. Bhallamudi S, Connell J, Pabelick CM, Prakash YS, Sathish V. Estrogen receptors differentially regulate intracellular calcium handling in human nonasthmatic and asthmatic airway smooth muscle cells. Am J Physiol Lung Cell Mol Physiol. (2020) 318:L112–24. doi: 10.1152/ajplung.00206.2019
59. Ambhore NS, Kalidhindi RSR, Loganathan J, Sathish V. Role of differential estrogen receptor activation in airway hyperreactivity and remodeling in a murine model of asthma. Am J Respir Cell Mol Biol. (2019) 61:469–80. doi: 10.1165/rcmb.2018-0321OC
60. Fuentes N, Silveyra P. Estrogen receptor signaling mechanisms. Adv Protein Chem Struct Biol. (2019) 116:135–70. doi: 10.1016/bs.apcsb.2019.01.001
61. Bonds RS, Midoro-Horiuti T. Estrogen effects in allergy and asthma. Curr Opin Allergy Clin Immunol. (2013) 13:92–9. doi: 10.1097/ACI.0b013e32835a6dd6
62. Couse JF, Lindzey J, Grandien K, Gustafsson JA, Korach KS. Tissue distribution and quantitative analysis of estrogen receptor-α (ERα) and estrogen receptor-β (ERβ) messenger ribonucleic acid in the wild-type and ERα-knockout mouse. Endocrinology. (1997) 138:4613–21. doi: 10.1210/endo.138.11.5496
63. Fuentes N, Cabello N, Nicoleau M, Chroneos ZC, Silveyra P. Modulation of the lung inflammatory response to ozone by the estrous cycle. Physiol Rep. (2019) 7:e14026. doi: 10.14814/phy2.14026
64. Pletzer B, Steinbeisser J, van Laak L, Harris T. Beyond biological sex: interactive effects of gender role and sex hormones on spatial abilities. Front Neurosci. (2019) 13:e675. doi: 10.3389/fnins.2019.00675
65. Slatkovska L, Jensen D, Davies GA, Wolfe LA. Phasic menstrual cycle effects on the control of breathing in healthy women. Respir Physiol Neurobiol. (2006) 154:379–88. doi: 10.1016/j.resp.2006.01.011
66. Foster PS, Goldie RG, Paterson JW. Effect of steroids on beta-adrenoceptor-mediated relaxation of pig bronchus. Br J Pharmacol. (1983) 78:441–5. doi: 10.1111/j.1476-5381.1983.tb09409.x
67. Perusquia M, Hernández R, Montaño LM, Villalón CM, Campos MG. Inhibitory effect of sex steroids on guinea-pig airway smooth muscle contractions. Comp Biochem Physiol Part C Pharmacol Toxicol Endocrinol. (1997) 118:5–10. doi: 10.1016/S0742-8413(97)00029-7
68. Sitruk-Ware R. Mifepristone and misoprostol sequential regimen side effects, complications and safety. Contraception. (2006) 74:48–55. doi: 10.1016/j.contraception.2006.03.016
69. Romieu I, Fabre A, Fournier A, Kauffmann F, Varraso R, Mesrine S, et al. Postmenopausal hormone therapy and asthma onset in the E3N cohort. Thorax. (2010) 65:292–7. doi: 10.1136/thx.2009.116079
70. Mannan SR, Begum N. Correlation of serum level of progesterone with peak expiratory flow rate (PEFR) in different phases of menstrual cycle. Anwer Khan Modern Med Coll J. (2012) 3:6–9. doi: 10.3329/akmmcj.v3i1.10105
71. Newcomb DC, Cephus JY, Boswell MG, Fahrenholz JM, Langley EW, Feldman AS, et al. Estrogen and progesterone decrease let-7f microRNA expression and increase IL-23/IL-23 receptor signaling and IL-17A production in patients with severe asthma. J Allergy Clin Immunol. (2015) 136:1025.e11–34.e11. doi: 10.1016/j.jaci.2015.05.046
72. Jain R, Ray JM, Pan JH, Brody SL. Sex hormone-dependent regulation of cilia beat frequency in airway epithelium. Am J Respir Cell Mol Biol. (2012) 46:446–53. doi: 10.1165/rcmb.2011-0107OC
73. Frühbeck G, Gómez-Ambrosi J, Muruzábal FJ, Burrell MA. The adipocyte: a model for integration of endocrine and metabolic signaling in energy metabolism regulation. Am J Physiol Endocrinol Metab. (2001) 280:E827–47. doi: 10.1152/ajpendo.2001.280.6.E827
74. Trayhurn P. Endocrine and signalling role of adipose tissue: new perspectives on fat. Acta Physiol Scand. (2005) 184:285–93. doi: 10.1111/j.1365-201X.2005.01468.x
75. Wang B, Jenkins JR, Trayhurn P. Expression and secretion of inflammation-related adipokines by human adipocytes differentiated in culture: integrated response to TNF-α. Am J Physiol Endocrinol Metab. (2005) 288:E731-40. doi: 10.1152/ajpendo.00475.2004
76. Berg AH, Scherer PE. Adipose tissue, inflammation, and cardiovascular disease. Circ Res. (2005) 96:939–49. doi: 10.1161/01.RES.0000163635.62927.34
77. Gordon S, Martinez FO. Alternative activation of macrophages: mechanism and functions. Immunity. (2010) 32:593–604. doi: 10.1016/j.immuni.2010.05.007
78. Ohashi K, Shibata R, Murohara T, Ouchi N. Role of anti-inflammatory adipokines in obesity-related diseases. Trends Endocrinol Metab. (2014) 25:348–55. doi: 10.1016/j.tem.2014.03.009
79. Kadowaki T, Yamauchi T. Adiponectin and adiponectin receptors. Endocr Rev. (2005) 26:439–51. doi: 10.1210/er.2005-0005
80. Ali Assad N, Sood A. Leptin, adiponectin and pulmonary diseases. Biochimie. (2012) 94:2180–9. doi: 10.1016/j.biochi.2012.03.006
81. Shore SA, Schwartzman IN, Mellema MS, Flynt L, Imrich A, Johnston RA. Effect of leptin on allergic airway responses in mice. J Allergy Clin Immunol. (2005) 115:103–9. doi: 10.1016/j.jaci.2004.10.007
82. Bellmeyer A, Martino JM, Chandel NS, Scott Budinger GR, Dean DA, Mutlu GM. Leptin resistance protects mice from hyperoxia-induced acute lung injury. Am J Respir Crit Care Med. (2007) 175:587–94. doi: 10.1164/rccm.200603-312OC
83. Bergen HT, Cherlet TC, Manuel P, Scott JE. Identification of leptin receptors in lung and isolated fetal type II cells. Am J Respir Cell Mol Biol. (2002) 27:71–7. doi: 10.1165/ajrcmb.27.1.4540
84. Bruno A, Pace E, Chanez P, Gras D, Vachier I, Chiappara G, et al. Leptin and leptin receptor expression in asthma. J Allergy Clin Immunol. (2009) 124:230–7. doi: 10.1016/j.jaci.2009.04.032
85. La Cava A, Alviggi C, Matarese G. Unraveling the multiple roles of leptin in inflammation and autoimmunity. J Mol Med. (2004) 82:4–11. doi: 10.1007/s00109-003-0492-1
86. Loffreda S, Yang S, Lin H, Karp C, Brengman ML, Wang D, et al. Leptin regulates proinflammatory immune responses. FASEB J. (1998) 12:57–65. doi: 10.1096/fsb2fasebj.12.1.57
87. Mohamed-Ali V, Goodrick S, Rawesh A, Katz D, Miles J, Yudkin J, et al. Subcutaneous adipose tissue releases interleukin-6, but not tumor necrosis factor-α, in vivo. J Clin Endocrinol Metab. (1997) 82:4196–200. doi: 10.1210/jc.82.12.4196
88. Faggioni R, Jones-Carson J, Reed DA, Dinarello CA, Feingold KR, Grunfeld C, et al. Leptin-deficient (ob/ob) mice are protected from T cell-mediated hepatotoxicity: role of tumor necrosis factor α and IL-18. Proc Nat Acad Sci U S A. (2000) 97:2367–72. doi: 10.1073/pnas.040561297
89. Banks R, Forbes M, Storr M, Higginson J, Thompson D, Raynes J, et al. The acute phase protein response in patients receiving subcutaneous IL-6. Clin Exp Immunol. (1995) 102:217–23. doi: 10.1111/j.1365-2249.1995.tb06659.x
90. Bastard JP, Jardel C, Delattre J, Hainque B, Bruckert E, Oberlin F. Evidence for a link between adipose tissue interleukin-6 content and serum C-reactive protein concentrations in obese subjects. Circulation. (1999) 99:2221–2. doi: 10.1161/circ.99.16.2219/c
91. Palacio A, Lopez M, Perez-Bravo F, Monkeberg F, Schlesinger L. Leptin levels are associated with immune response in malnourished infants. J Clin Endocrinol Metab. (2002) 87:3040–6. doi: 10.1210/jcem.87.7.8636
92. Matarese G, La Cava A, Sanna V, Lord GM, Lechler RI, Fontana S, et al. Balancing susceptibility to infection and autoimmunity: a role for leptin? Trends Immunol. (2002) 23:182–7. doi: 10.1016/S1471-4906(02)02188-9
93. Johnston RA, Schwartzman IN, Shore SA. Macrophage inflammatory protein-2 levels are associated with changes in serum leptin concentrations following ozone-induced airway inflammation. Chest. (2003) 123:369S–70S. doi: 10.1016/S0012-3692(15)35211-9
94. Sin DD, Man SP. Why are patients with chronic obstructive pulmonary disease at increased risk of cardiovascular diseases? The potential role of systemic inflammation in chronic obstructive pulmonary disease. Circulation. (2003) 107:1514–9. doi: 10.1161/01.CIR.0000056767.69054.B3
95. Guler N, Kirerleri E, Ones U, Tamay Z, Salmayenli N, Darendeliler F. Leptin: does it have any role in childhood asthma? J Allergy Clin Immunol. (2004) 114:254–9. doi: 10.1016/j.jaci.2004.03.053
96. Dimitropoulou C, White RE, Ownby DR, Catravas JD. Estrogen reduces carbachol-induced constriction of asthmatic airways by stimulating large-conductance voltage and calcium-dependent potassium channels. Am J Respir Cell Mol Biol. (2005) 32:239–47. doi: 10.1165/rcmb.2004-0331OC
97. Riffo-Vasquez Y, Ligeiro de Oliveira AP, Page CP, Spina D, Tavares-de-Lima W. Role of sex hormones in allergic inflammation in mice. Clin Exp Allergy. (2007) 37:459–70. doi: 10.1111/j.1365-2222.2007.02670.x
98. Takeda M, Tanabe M, Ito W, Ueki S, Konnno Y, Chihara M, et al. Gender difference in allergic airway remodelling and immunoglobulin production in mouse model of asthma. Respirology. (2013) 1:797–806. doi: 10.1111/resp.12078
99. Carey MA, Card JW, Bradbury JA, Moorman MP, Haykal-Coates N, Gavett SH, et al. Spontaneous airway hyperresponsiveness in estrogen receptor-alpha-deficient mice. Am J Respir Crit Care Med. (2007) 175:126–35. doi: 10.1164/rccm.200509-1493OC
100. Hall OJ, Limjunyawong N, Vermillion MS, Robinson DP, Wohlgemuth N, Pekosz A, et al. Progesterone-based therapy protects against influenza by promoting lung repair and recovery in females. PLoS Pathog. (2016) 12:e1005840. doi: 10.1371/journal.ppat.1005840
101. Hellings PW, Vandekerckhove P, Claeys R, Billen J, Kasran A, Ceuppens JL. Progesterone increases airway eosinophilia and hyper-responsiveness in a murine model of allergic asthma. Clin Exp Allergy. (2003) 33:1457–63. doi: 10.1046/j.1365-2222.2003.01743.x
102. de Oliveira AP, Peron JP, Damazo AS, Franco AL, Domingos HV, Oliani SM, et al. Female sex hormones mediate the allergic lung reaction by regulating the release of inflammatory mediators and the expression of lung E-selectin in rats. Respir Res. (2010) 11:115. doi: 10.1186/1465-9921-11-115
103. Yu CK, Liu YH, Chen CL. Dehydroepiandrosterone attenuates allergic airway inflammation in Dermatophagoides farinae-sensitized mice. J Microbiol Immunol Infect. (2002) 35:199–202.
104. Senna G, Latorre M, Bugiani M, Caminati M, Heffler E, Morrone D, et al. Sex differences in severe asthma: results from severe asthma network in Italy-SANI. Allergy Asthma Immunol Res. (2021) 13:219. doi: 10.4168/aair.2021.13.2.219
105. Cephus JY, Stier MT, Fuseini H, Yung JA, Toki S, Bloodworth MH, et al. Testosterone attenuates group 2 innate lymphoid cell-mediated airway inflammation. Cell Rep. (2017) 21:2487–99. doi: 10.1016/j.celrep.2017.10.110
106. Warren KJ, Sweeter JM, Pavlik JA, Nelson AJ, Devasure JM, Dickinson JD, et al. Sex differences in activation of lung-related type 2 innate lymphoid cells in experimental asthma. Ann Allergy Asthma Immunol. (2017) 118:233–4. doi: 10.1016/j.anai.2016.11.011
107. Huang Y, Mao K, Chen X, Sun MA, Kawabe T, Li W, et al. S1P-dependent interorgan trafficking of group 2 innate lymphoid cells supports host defense. Science. (2018) 359:114–9. doi: 10.1126/science.aam5809
108. Deraz T, Kamel TB, El-Kerdany TA, El-Ghazoly HM. High-sensitivity C reactive protein as a biomarker for grading of childhood asthma in relation to clinical classification, induced sputum cellularity, and spirometry. Pediatr Pulmonol. (2012) 47:220–5. doi: 10.1002/ppul.21539
109. Blacquiere M, Hylkema M, Postma D, Geerlings M, Timens W, Melgert B. Airway inflammation and remodeling in two mouse models of asthma: comparison of males and females. Int Arch Allergy Immunol. (2010) 153:173–81. doi: 10.1159/000312635
110. Heffler E, Madeira LNG, Ferrando M, Puggioni F, Racca F, Malvezzi L, et al. Inhaled corticosteroids safety and adverse effects in patients with asthma. J Allergy Clin Immunol Pract. (2018) 6:776–81. doi: 10.1016/j.jaip.2018.01.025
111. Hayashi T. Adachi Y, Hasegawa K, Morimoto M. Less sensitivity for late airway inflammation in males than females in BALB/c mice. Scand J Immunol. (2003) 57:562–7. doi: 10.1046/j.1365-3083.2003.01269.x
112. Card JW, Carey MA, Bradbury JA, DeGraff LM, Morgan DL, Moorman MP, et al. Gender differences in murine airway responsiveness and lipopolysaccharide-induced inflammation. J Immunol. (2006) 177:621–30. doi: 10.4049/jimmunol.177.1.621
113. Okuyama K, Wada K, Chihara J, Takayanagi M, Ohno I. Sex-related splenocyte function in a murine model of allergic asthma. Clin Exp Allergy. (2008) 38:1212–9. doi: 10.1111/j.1365-2222.2008.03015.x
114. Bartemes KR, Kephart GM, Fox SJ, Kita H. Enhanced innate type 2 immune response in peripheral blood from patients with asthma. J Allergy Clin Immunol. (2014) 134:671.e74–678.e74. doi: 10.1016/j.jaci.2014.06.024
115. Liu T, Wu J, Zhao J, Wang J, Zhang Y, Liu L, et al. Type 2 innate lymphoid cells: a novel biomarker of eosinophilic airway inflammation in patients with mild to moderate asthma. Respir Med. (2015) 109:1391–6. doi: 10.1016/j.rmed.2015.09.016
116. Smith SG, Chen R, Kjarsgaard M, Huang C, Oliveria JP, O'Byrne PM, et al. Increased numbers of activated group 2 innate lymphoid cells in the airways of patients with severe asthma and persistent airway eosinophilia. J Allergy Clin Immunol. (2016) 137:75–86.e78. doi: 10.1016/j.jaci.2015.05.037
117. Halim TY, Steer CA, Mathä L, Gold MJ, Martinez-Gonzalez I, McNagny KM, et al. Group 2 innate lymphoid cells are critical for the initiation of adaptive T helper 2 cell-mediated allergic lung inflammation. Immunity. (2014) 40:425–35. doi: 10.1016/j.immuni.2014.01.011
118. Klein Wolterink RG, Kleinjan A, van Nimwegen M, Bergen I, de Bruijn M, Levani Y, et al. Pulmonary innate lymphoid cells are major producers of IL-5 and IL-13 in murine models of allergic asthma. Eur J Immunol. (2012) 42:1106–16. doi: 10.1002/eji.201142018
119. Zhou W, Toki S, Zhang J, Goleniewksa K, Newcomb DC, Cephus JY, et al. Prostaglandin I2 signaling and inhibition of group 2 innate lymphoid cell responses. Am J Respir Crit Care Med. (2016) 193:31–42. doi: 10.1164/rccm.201410-1793OC
120. Huang Y, Guo L, Qiu J, Chen X, Hu-Li J, Siebenlist U, et al. IL-25-responsive, lineage-negative KLRG1 hi cells are multipotential ‘inflammatory'type 2 innate lymphoid cells. Nat Immunol. (2015) 16:161–9. doi: 10.1038/ni.3078
121. Kadel S, Ainsua-Enrich E, Hatipoglu I, Turner S, Singh S, Khan S, et al. A major population of functional KLRG1(-) ILC2s in female lungs contributes to a sex bias in ILC2 numbers. Immunohorizons. (2018) 2:74–86. doi: 10.4049/immunohorizons.1800008
122. Laffont S, Blanquart E, Guéry JC. Sex differences in asthma: a key role of androgen-signaling in group 2 innate lymphoid cells. Front Immunol. (2017) 8:e1069. doi: 10.3389/fimmu.2017.01069
123. White J, Tannenbaum C, Klinge I, Schiebinger L, Clayton J. The integration of sex and gender considerations into biomedical research: lessons from international funding agencies. J Clin Endocrinol Metab. (2021) 106:3034–48. doi: 10.1210/clinem/dgab434
124. Chanez P, Wenzel SE, Anderson GP, Anto JM, Bel EH, Boulet LP, et al. Severe asthma in adults: what are the important questions? J Allergy Clin Immunol. (2007) 119:1337–48. doi: 10.1016/j.jaci.2006.11.702
125. Han MK, Arteaga-Solis E, Blenis J, Bourjeily G, Clegg DJ, DeMeo D, et al. Female sex and gender in lung/sleep health and disease. increased understanding of basic biological, pathophysiological, and behavioral mechanisms leading to better health for female patients with lung disease. Am J Respir Crit Care Med. (2018) 198:850–58. doi: 10.1164/rccm.201801-0168WS
126. Talini D, Ciberti A, Bartoli D, Del Guerra P, Iaia TE, Lemmi M, et al. Work-related asthma in a sample of subjects with established asthma. Respir Med. (2017) 130:85–91. doi: 10.1016/j.rmed.2017.07.008
127. Chen Y, Dales R, Tang M, Krewski D. Obesity may increase the incidence of asthma in women but not in men: longitudinal observations from the Canadian National Population Health Surveys. Am J Epidemiol. (2002) 155:191–7. doi: 10.1093/aje/155.3.191
128. Rodriguez Bauza D, Silveyra P. Sex differences in exercise induced bronchoconstriction in athletes: a systematic review and meta-analysis. Int J Environ Res Public Health. (2020) 17:7270. doi: 10.20944/preprints202008.0591.v1
129. Venkatraman JT, Pendergast D. Effects of the level of dietary fat intake and endurance exercise on plasma cytokines in runners. Med Sci Sports Exerc. (1998) 30:1198–204. doi: 10.1097/00005768-199808000-00003
130. Meksawan K, Venkatraman JT, Awad AB, Pendergast DR. Effect of dietary fat intake and exercise on inflammatory mediators of the immune system in sedentary men and women. J Am Coll Nutr. (2004) 23:331–40. doi: 10.1080/07315724.2004.10719376
131. Moyna N, Acker G, Fulton J, Weber K, Goss F, Robertson R, et al. Lymphocyte function and cytokine production during incremental exercise in active and sedentary males and females. Int J Sports Med. (1996) 17:585–91. doi: 10.1055/s-2007-972899
132. Navalta J, Sedlock D, Park KS. Effect of exercise intensity on exercise-induced lymphocyte apoptosis. Int J Sports Med. (2007) 28:539–42. doi: 10.1055/s-2006-955898
133. Bouman A, Moes H, Heineman MJ, de Leij LF, Faas MM. The immune response during the luteal phase of the ovarian cycle: increasing sensitivity of human monocytes to endotoxin. Fertil Steril. (2001) 76:555–9. doi: 10.1016/S0015-0282(01)01971-9
134. Stupka N, Lowther S, Chorneyko K, Bourgeois J, Hogben C, Tarnopolsky M. Gender differences in muscle inflammation after eccentric exercise. J Appl Physiol. (2000) 89:2325–32. doi: 10.1152/jappl.2000.89.6.2325
135. Timmons BW, Hamadeh MJ, Devries MC, Tarnopolsky MA. Influence of gender, menstrual phase, and oral contraceptive use on immunological changes in response to prolonged cycling. J Appl Physiol. (2005) 99:979–85. doi: 10.1152/japplphysiol.00171.2005
136. Northoff H, Symons S, Zieker D, Schaible EV, Schäfer K, Thoma S, et al. Gender-and menstrual phase dependent regulation of inflammatory gene expression in response to aerobic exercise. Exerc Immunol Rev. (2008) 14:86–103.
137. Khan NA, Lindley MR. Sex differences in athletes with asthma and exercise-induced bronchoconstriction: future directions. Future Sci. (2017) 3:FSO227. doi: 10.4155/fsoa-2017-0082
138. Stanford KI, Mickleborough TD, Ray S, Lindley MR, Koceja DM, Stager JM. Influence of menstrual cycle phase on pulmonary function in asthmatic athletes. Eur J Appl Physiol. (2006) 96:703–10. doi: 10.1007/s00421-005-0067-7
139. Hancox RJ, Milne BJ, Poulton R, Taylor DR, Greene JM, McLachlan CR, et al. Sex differences in the relation between body mass index and asthma and atopy in a birth cohort. Am J Respir Crit Care Med. (2005) 171:440–5. doi: 10.1164/rccm.200405-623OC
140. Chowdhury NU, Guntur VP, Newcomb DC, Wechsler ME. Sex and gender in asthma. Eur Respir Rev. (2021) 30:210067. doi: 10.1183/16000617.0067-2021
141. Sorlí-Aguilar M, Martín-Luján F, Flores-Mateo G, Jardí-Piñana C, Aparicio-Llopis E, Basora-Gallisà J, et al. Adiposity markers and lung function in smokers: a cross-sectional study in a Mediterranean population. BMC Pulm Med. (2016) 16:178. doi: 10.1186/s12890-016-0341-y
142. Castro-Rodríguez JA, Holberg CJ, Morgan WJ, Wright AL, Martinez FD. Increased incidence of asthmalike symptoms in girls who become overweight or obese during the school years. Am J Respir Crit Care Med. (2001) 163:1344–9. doi: 10.1164/ajrccm.163.6.2006140
143. Tantisira K, Weiss S. Complex interactions in complex traits: obesity and asthma. Thorax. (2001) 56 (suppl 2):ii64-74.
144. Gibeon D, Batuwita K, Osmond M, Heaney LG, Brightling CE, Niven R, et al. Obesity-associated severe asthma represents a distinct clinical phenotype: analysis of the British thoracic society difficult asthma registry patient cohort according to BMI. Chest. (2013) 143:406–14. doi: 10.1378/chest.12-0872
145. Wang L, Wang K, Gao X, Paul TK, Cai J, Wang Y. Sex difference in the association between obesity and asthma in U.S. adults: findings from a national study. Respir Med. (2015) 109:955–62. doi: 10.1016/j.rmed.2015.06.001
146. Miethe S, Guarino M, Alhamdan F, Simon HU, Renz H, Dufour JF, et al. Effects of obesity on asthma: immunometabolic links. Pol Arch Intern Med. (2018) 128:469–77. doi: 10.20452/pamw.4304
147. Wenzel S. Severe asthma: from characteristics to phenotypes to endotypes. Clin Exp Allergy. (2012) 42:650–8. doi: 10.1111/j.1365-2222.2011.03929.x
148. Wenzel SE. Asthma phenotypes: the evolution from clinical to molecular approaches. Nat Med. (2012) 18:716–25. doi: 10.1038/nm.2678
149. E-Lacerda RR, Anhê GF, Page CP, Riffo-Vasquez Y. Sex differences in the influence of obesity on a murine model of allergic lung inflammation. Clin Exp Allergy. (2020) 50:256–66. doi: 10.1111/cea.13541
150. Yu G, Zhu L, Li H, Shao Y, Chon L, Zhang H, et al. Influence of gender on OVA-induced airway inflammation in C57/B6J mice on a high-fat diet. (2018), letter. doi: 10.1177/2058739218760946
151. Kim S, Camargo CA Jr. Sex-race differences in the relationship between obesity and asthma: the behavioral risk factor surveillance system, 2000. Ann Epidemiol. (2003) 13:666–73. doi: 10.1016/S1047-2797(03)00054-1
152. Huovinen E, Kaprio J, Koskenvuo M. Factors associated to lifestyle and risk of adult onset asthma. Respir Med. (2003) 97:273–80. doi: 10.1053/rmed.2003.1419
153. Ford E, Mannino D, Redd S, Mokdad A, Mott J. Body mass index and asthma incidence among USA adults. Eur Respir J. (2004) 24:740–4. doi: 10.1183/09031936.04.00088003
154. Gunnbjörnsdóttir MI, Omenaas E, Gíslason T, Norrman E, Olin AC, Jogi R, et al. Obesity and nocturnal gastro-oesophageal reflux are related to onset of asthma and respiratory symptoms. Eur Respir J. (2004) 24:116–21. doi: 10.1183/09031936.04.00042603
155. Luder E, Ehrlich R, Lou W, Melnik T, Kattan M. Body mass index and the risk of asthma in adults. Respir Med. (2004) 98:29–37. doi: 10.1016/j.rmed.2003.08.004
156. Bibi H, Shoseyov D, Feigenbaum D, Genis M, Friger M, Peled R, et al. The relationship between asthma and obesity in children: is it real or a case of over diagnosis? J Asthma. (2004) 41:403–10. doi: 10.1081/JAS-120026097
157. Beuther DA, Sutherland ER. Overweight, obesity, and incident asthma: a meta-analysis of prospective epidemiologic studies. Am J Respir Crit Care Med. (2007) 175:661–6. doi: 10.1164/rccm.200611-1717OC
158. Wilson MM, Irwin RS. The association of asthma and obesity: is it real or a matter of definition, Presbyterian ministers' salaries, and earlobe creases? Arch Intern Med. (1999) 159:2513–4. doi: 10.1001/archinte.159.21.2513
159. Friedman JM, Halaas JL. Leptin and the regulation of body weight in mammals. Nature. (1998) 395:763–70. doi: 10.1038/27376
160. Visser M, Bouter LM, McQuillan GM, Wener MH, Harris TB. Elevated C-reactive protein levels in overweight and obese adults. JAMA. (1999) 282:2131–35. doi: 10.1001/jama.282.22.2131
161. Gautam Y, Afanador Y, Abebe T, López JE, Mersha TB. Genome-wide analysis revealed sex-specific gene expression in asthmatics. Hum Mol Genet. (2019) 28:2600–14. doi: 10.1093/hmg/ddz074
162. Hynes GM, Hinks TSC. The role of interleukin-17 in asthma: a protective response? ERJ Open Res. (2020) 6:00364–2019. doi: 10.1183/23120541.00364-2019
163. Molet S, Hamid Q, Davoine F, Nutku E, Taha R, Pagé N, et al. IL-17 is increased in asthmatic airways and induces human bronchial fibroblasts to produce cytokines. J Allergy Clin Immunol. (2001) 108:430–8. doi: 10.1067/mai.2001.117929
164. Barczyk A, Pierzchala W, SozaÑSka E. Interleukin-17 in sputum correlates with airway hyperresponsiveness to methacholine. Respir Med. (2003) 97:726–33. doi: 10.1053/rmed.2003.1507
165. Monin L, Gaffen SL. Interleukin 17 family cytokines: signaling mechanisms, biological activities, and therapeutic implications. Cold Spring Harb Perspect Biol. (2018) 10:a028522. doi: 10.1101/cshperspect.a028522
166. Al-Ramli W, Préfontaine D, Chouiali F, Martin JG, Olivenstein R, Lemière C, et al. T(H)17-associated cytokines (IL-17A and IL-17F) in severe asthma. J Allergy Clin Immunol. (2009) 123:1185–7. doi: 10.1016/j.jaci.2009.02.024
167. Sorbello V, Ciprandi G, Di Stefano A, Massaglia G, Favata G, Conticello S, et al. Nasal IL-17F is related to bronchial IL-17F/neutrophilia and exacerbations in stable atopic severe asthma. Allergy. (2015) 70:236–40. doi: 10.1111/all.12547
168. Bullens DM, Truyen E, Coteur L, Dilissen E, Hellings PW, Dupont LJ, et al. IL-17 mRNA in sputum of asthmatic patients: linking T cell driven inflammation and granulocytic influx. Respir Res. (2006) 7:135. doi: 10.1186/1465-9921-7-135
169. Ricciardolo FL, Sorbello V, Folino A, Gallo F, Massaglia GM, Favatà G, et al. Identification of IL-17F/frequent exacerbator endotype in asthma. J Allergy Clin Immunol. (2017) 140:395–406. doi: 10.1016/j.jaci.2016.10.034
170. Busse WW, Holgate S, Kerwin E, Chon Y, Feng J, Lin J, et al. Randomized, double-blind, placebo-controlled study of brodalumab, a human anti-IL-17 receptor monoclonal antibody, in moderate to severe asthma. Am J Respir Crit Care Med. (2013) 188:1294–302. doi: 10.1164/rccm.201212-2318OC
171. Patel R, Solatikia F, Zhang H, Wolde A, Kadalayil L, Karmaus W, et al. Sex-specific associations of asthma acquisition with changes in DNA methylation during adolescence. Clin Exp Allergy. (2021) 51:318–28. doi: 10.1111/cea.13776
172. DeVries A, Wlasiuk G, Miller SJ, Bosco A, Stern DA, Lohman IC, et al. Epigenome-wide analysis links SMAD3 methylation at birth to asthma in children of asthmatic mothers. J Allergy Clin Immunol. (2017) 140:534–42. doi: 10.1016/j.jaci.2016.10.041
173. Murphy TM, Wong CC, Arseneault L, Burrage J, Macdonald R, Hannon E, et al. Methylomic markers of persistent childhood asthma: a longitudinal study of asthma-discordant monozygotic twins. Clin Epigenetics. (2015) 7:1–13. doi: 10.1186/s13148-015-0163-4
174. Arathimos R, Suderman M, Sharp GC, Burrows K, Granell R, Tilling K, et al. Epigenome-wide association study of asthma and wheeze in childhood and adolescence. Clin Epigenetics. (2017) 9:1–16. doi: 10.1186/s13148-017-0414-7
175. Gunawardhana LP, Gibson PG, Simpson JL, Benton MC, Lea RA, Baines KJ. Characteristic DNA methylation profiles in peripheral blood monocytes are associated with inflammatory phenotypes of asthma. Epigenetics. (2014) 9:1302–16. doi: 10.4161/epi.33066
176. Prunicki M, Stell L, Dinakarpandian D, de Planell-Saguer M, Lucas RW, Hammond SK, et al. Exposure to NO 2, CO, and PM 2.5 is linked to regional DNA methylation differences in asthma. Clin Epigenetics. (2018) 10:1–1. doi: 10.1186/s13148-017-0433-4
177. Yang IV, Pedersen BS, Liu A, O'Connor GT, Teach SJ, Kattan M, et al. DNA methylation and childhood asthma in the inner city. J Allergy Clin Immunol. (2015) 136:69–80. doi: 10.1016/j.jaci.2015.01.025
178. Reese SE, Xu CJ, Herman T, Lee MK, Sikdar S, Ruiz-Arenas C, et al. Epigenome-wide meta-analysis of DNA methylation and childhood asthma. J Allergy Clin Immunol. (2019) 143:2062–74. doi: 10.1016/j.jaci.2018.11.043
179. Espuela-Ortiz A, Herrera-Luis E, Lorenzo-Díaz F, Hu D, Eng C, Villar J, et al. Role of sex on the genetic susceptibility to childhood asthma in latinos and African Americans. J Pers Med. (2021) 11:1140. doi: 10.3390/jpm11111140
180. Myers RA, Scott NM, Gauderman WJ, Qiu W, Mathias RA, Romieu I, et al. Genome-wide interaction studies reveal sex-specific asthma risk alleles. Hum Mol Genet. (2014) 23:5251–9. doi: 10.1093/hmg/ddu222
181. Santillan AA, Camargo CA Jr, Ramirez-Rivera A, Delgado-Enciso I, Rojas-Martinez A, Cantu-Diaz F, et al. Association between beta2-adrenoceptor polymorphisms and asthma diagnosis among Mexican adults. J Allergy Clin Immunol. (2003) 112:1095–100. doi: 10.1016/j.jaci.2003.09.029
182. Hunninghake GM, Soto-Quirós ME, Avila L, Kim HP, Lasky-Su J, Rafaels N, et al. TSLP polymorphisms are associated with asthma in a sex-specific fashion. Allergy. (2010) 65:1566–75. doi: 10.1111/j.1398-9995.2010.02415.x
183. Rochman Y, Leonard WJ. The role of thymic stromal lymphopoietin in CD8+ T cell homeostasis. J Immunol. (2008) 181:7699–705. doi: 10.4049/jimmunol.181.11.7699
184. Shubin NJ, Clauson M, Niino K, Kasprzak V, Tsuha A, Guga E, et al. Thymic stromal lymphopoietin protects in a model of airway damage and inflammation via regulation of caspase-1 activity and apoptosis inhibition. Mucosal Immunol. (2020) 13:584–94. doi: 10.1038/s41385-020-0271-0
185. Kabata H, Flamar AL, Mahlakõiv T, Moriyama S, Rodewald HR, Ziegler SF, et al. Targeted deletion of the TSLP receptor reveals cellular mechanisms that promote type 2 airway inflammation. Mucosal Immunol. (2020) 13:626–36. doi: 10.1038/s41385-020-0266-x
186. Gauderman WJ, Urman R, Avol E, Berhane K, McConnell R, Rappaport E, et al. Association of improved air quality with lung development in children. N Engl J Med. (2015) 372:905–13. doi: 10.1056/NEJMoa1414123
187. Loisel DA, Tan Z, Tisler CJ, Evans MD, Gangnon RE, Jackson DJ, et al. IFNG genotype and sex interact to influence the risk of childhood asthma. J Allergy Clin Immunol. (2011) 128:524–31. doi: 10.1016/j.jaci.2011.06.016
188. Seibold MA, Wang B, Eng C, Kumar G, Beckman KB, Sen S, et al. An african-specific functional polymorphism in KCNMB1 shows sex-specific association with asthma severity. Hum Mol Genet. (2008) 17:2681–90. doi: 10.1093/hmg/ddn168
189. DeVries A, Vercelli D. Epigenetic mechanisms in asthma. Ann Am Thorac Soc. (2016) 13 (Suppl 1):S48-50. doi: 10.1513/AnnalsATS.201507-420MG
190. Bae DJ, Jun JA, Chang HS, Park JS, Park CS. Epigenetic changes in asthma: role of DNA CpG methylation. Tuberc Respir Dis (Seoul). (2020) 83:1–13. doi: 10.4046/trd.2018.0088
191. Ntontsi P, Photiades A, Zervas E, Xanthou G, Samitas K. Genetics and epigenetics in asthma. Int J Mol Sci. (2021) 22:2412. doi: 10.3390/ijms22052412
192. Potaczek DP, Alashkar Alhamwe B, Miethe S, Garn H. Epigenetic mechanisms in allergy development and prevention. Handb Exp Pharmacol. (2022) 268:331–57. doi: 10.1007/164_2021_475
193. Pfefferle PI, Keber CU, Cohen RM, Garn H. The hygiene hypothesis - learning from but not living in the past. Front Immunol. (2021) 12:e635935. doi: 10.3389/fimmu.2021.635935
194. Breton CV, Byun HM, Wang X, Salam MT, Siegmund K, Gilliland FD, et al. methylation in the arginase-nitric oxide synthase pathway is associated with exhaled nitric oxide in children with asthma. Am J Respir Crit Care Med. (2011) 184:191–7. doi: 10.1164/rccm.201012-2029OC
195. Baccarelli A, Rusconi F, Bollati V, Catelan D, Accetta G, Hou L, et al. Nasal cell DNA methylation, inflammation, lung function and wheezing in children with asthma. Epigenomics. (2012) 4:91–100. doi: 10.2217/epi.11.106
196. Jõgi NO, Kitaba N, Storaas T, Schlünssen V, Triebner K, Holloway JW, et al. Ascaris exposure and its association with lung function, asthma, and DNA methylation in Northern Europe. J Allergy Clin Immunol. 14:S0091-6749(21)01797-8. (2021). doi: 10.1016/j.jaci.2021.11.013
197. Rebane A, Akdis CA. MicroRNAs: essential players in the regulation of inflammation. J Allergy Clin Immunol. (2013) 132:15–26. doi: 10.1016/j.jaci.2013.04.011
198. Tüfekci KU, Meuwissen RL, Genç S. The role of microRNAs in biological processes. Methods Mol Biol. (2014) 1107:15–31. doi: 10.1007/978-1-62703-748-8_2
199. Paul P, Chakraborty A, Sarkar D, Langthasa M, Rahman M, Bari M, et al. Interplay between miRNAs and human diseases. J Cell Physiol. (2018) 233:2007–18. doi: 10.1002/jcp.25854
200. Hayes J, Peruzzi PP, Lawler S. MicroRNAs in cancer: biomarkers, functions and therapy. Trends Mol Med. (2014) 20:460–9. doi: 10.1016/j.molmed.2014.06.005
201. Wang J, Chen J, Sen S. MicroRNA as biomarkers and diagnostics. J Cell Physiol. (2016) 231:25–30. doi: 10.1002/jcp.25056
202. Huang W. MicroRNAs: biomarkers, diagnostics, and therapeutics. Bioinform Microrna Res. (2017) 617:57–67. doi: 10.1007/978-1-4939-7046-9_4
203. Fuentes N, Roy A, Mishra V, Cabello N, Silveyra P. Sex-specific microRNA expression networks in an acute mouse model of ozone-induced lung inflammation. Biol Sex Differ. (2018) 9:18. doi: 10.1186/s13293-018-0177-7
204. Sharma S, Kho AT, Chhabra D, Haley K, Vyhlidal C, Gaedigk R, et al. Effect of intrauterine smoke exposure on microRNA-15a expression in human lung development and subsequent asthma risk. Healthcare. (2020) 8:536. doi: 10.3390/healthcare8040536
205. Kho AT, Sharma S, Davis JS, Spina J, Howard D, McEnroy K, et al. Circulating microRNAs: association with lung function in asthma. PLoS ONE. (2016) 11:e0157998. doi: 10.1371/journal.pone.0157998
206. Dehmel S, Nathan P, Bartel S, El-Merhie N, Scherb H, Milger K, et al. Intrauterine smoke exposure deregulates lung function, pulmonary transcriptomes, and in particular insulin-like growth factor (IGF)-1 in a sex-specific manner. Sci Rep. (2018) 8:7547. doi: 10.1038/s41598-018-25762-5
207. Mehta A, Baltimore D. MicroRNAs as regulatory elements in immune system logic. Nat Rev Immunol. (2016) 16:279–94. doi: 10.1038/nri.2016.40
208. Gebert LFR, MacRae IJ. Regulation of microRNA function in animals. Nat Rev Mol Cell Biol. (2019) 20:21–37. doi: 10.1038/s41580-018-0045-7
209. Johansson K, Weidner J, Rådinger M. MicroRNAs in type 2 immunity. Cancer Lett. (2018) 425:116–24. doi: 10.1016/j.canlet.2018.03.036
210. Pua HH, Ansel KM. MicroRNA regulation of allergic inflammation and asthma. Curr Opin Immunol. (2015) 36:101–8. doi: 10.1016/j.coi.2015.07.006
Keywords: asthma, airway, sex, gender, lung, hormone
Citation: Ekpruke CD and Silveyra P (2022) Sex Differences in Airway Remodeling and Inflammation: Clinical and Biological Factors. Front. Allergy 3:875295. doi: 10.3389/falgy.2022.875295
Received: 14 February 2022; Accepted: 31 March 2022;
Published: 29 April 2022.
Edited by:
Khuloud Bajbouj, University of Sharjah, United Arab EmiratesReviewed by:
Saba Al Heialy, Mohammed Bin Rashid University of Medicine and Health Sciences, United Arab EmiratesCopyright © 2022 Ekpruke and Silveyra. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Patricia Silveyra, cHNpbHZleXJAaXUuZWR1
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.