 Giovanni Sgroi1†
Giovanni Sgroi1† Luigi Jacopo D’Auria1,2†
Luigi Jacopo D’Auria1,2† Maria Gabriella Lucibelli1
Maria Gabriella Lucibelli1 Andrea Mancusi1*
Andrea Mancusi1* Yolande Thérèsa Rose Proroga1
Yolande Thérèsa Rose Proroga1 Mauro Esposito1,2
Mauro Esposito1,2 Simona Rea1
Simona Rea1 Daniel Signorelli3
Daniel Signorelli3 Federica Gargano1
Federica Gargano1 Nicola D’Alessio1Ranju Ravindran Santhakumari Manoj4Peyman Khademi5
Nicola D’Alessio1Ranju Ravindran Santhakumari Manoj4Peyman Khademi5 Giuseppe Rofrano1,2
Giuseppe Rofrano1,2- 1Experimental Zooprophylactic Institute of Southern Italy, Portici, Naples, Italy
- 2Centro di Referenza Nazionale per l’analisi e lo studio delle correlazioni tra ambiente, animali e uomini, Portici, Naples, Italy
- 3Department of Cultural Heritage Sciences, University of Salerno, Salerno, Italy
- 4Department of Population Medicine and Diagnostics, Cornell University, Ithaca, NY, United States
- 5Department of Microbiology and Food Hygiene, University of Lorestan, Khorramabad, Iran
Introduction: The decline of the European/western honeybee (Apis mellifera) population is on account of a plethora of microorganisms, such as Nosema apis and Nosema ceranae, two microsporidian fungi responsible of nosemosis that affects welfare and production of the bee industry. Accordingly, this study aimed to investigate the presence of both pathogens in bees, pollen and honey from apiaries in Southwestern Italy.
Methods: From March to July 2022 and 2023, apiaries (n = 10) were selected and classified as High Impact Areas (HIAs, n = 5) and Low Impact Areas (LIAs, n = 5) according to a 5-point environmental risk index based on factors affecting bee health sand related productions. Bee, pollen and honey samples, were collected and tested for Nosema spp. DNA by specific PCR protocols targeting the 16S rRNA gene. Signs/symptoms of nosemosis were monitored and collected by the cooperation of beekeepers.
Results: Out of 10 apiaries, 6 (i.e., 60%, 95% CI: 31.3–83.2) tested positive for at least one sample to Nosema spp. DNA, being 2 positives for N. apis, 2 for N. ceranae and 2 co-infected (i.e., 20%, 5.7–51.0). Based on the biological samples, honey was positive for N. apis in one apiary, pollen for N. ceranae in two apiaries, and bees for N. apis in 3 apiaries, N. ceranae in 1 apiary, and both species in 1 apiary. In all the apiaries positive to N. apis and N. ceranae, high mortality and low honey production were observed. A higher risk of infection was observed in apiaries from HIAs (OR = 6.00). The sequences of N. apis and N. ceranae had 99.5–100% homology with those in the GenBank database. Whereas all sequences of N. apis were identical to each other, four sequences types of N. ceranae characterized by single nucleotide polymorphisms (SNPs) were identified. The computation of polymorphisms revealed high haplotype diversity (i.e., Hd = 1.000) and low nucleotide diversity (i.e., Pi = 0.00913) of N. ceranae sequence types.
Discussion: This study reveals a high circulation of N. apis and N. ceranae in Southwestern Italy, indicating the need for improved monitoring of these microsporidia to protect bee welfare and bee industry.
1 Introduction
In last years, a plethora of infectious and parasitic agents has affected the population decline of pollinating insects worldwide (1). Among these, the European honey bee (or western honey bee) Apis mellifera is commonly exposed to fungi of the genus Nosema spp. (phylum Microsporidia, family Nosematidae), obligate intracellular microsporidia that infect ventricular epithelium cells of adut bees causing nosemosis (2–5), mainly in high density apiaries (6). Nosemosis is one of the main causes of bee colony collapse and prodution loss for beekeepers (7) due to the severe dysentery caused in bees (8, 9). In addition, the oro-fecal transmission pathway of Nosema spp. and the typical food-sharing behaviour of bees (i.e., trophallaxis) (10) favour the circulation of microsporidian spores among bees, but also pollen and honey, causing organoleptic and production alterations in honey (11–15). To date, two different pathogenic species of Nosema spp. have been reported, Nosema apis (synonym Vairimorpha apis) and Nosema ceranae (synonym Vairimorpha ceranae) (16, 17). While N. apis is known from the beginning of the XX century (18), N. ceranae is an emerging species reported for the first time in 2005 (19) and now spread throughout Europe (20–26), as well as in Canada, USA, and south America (27–30).
In perspective, the fact that long cold winters, intense rain, and high relative humidity can favour the spread of N. apis and N. ceranae is of concern considering the running climatic changes and global warming (31–34), especially for N. ceranae that is considered replacing N. apis in several areas (35). As for Italy, N. ceranae has been reported in apiaries from central and Northern Italy, whereas the last report of N. apis dates back to 2010 in the North of the peninsula according to a nationwide 2 year-monitoring plan (36). Moreover, the occurrence of N. apis has been recently reported in apiaries of Southeastern Italy (15), suggesting the potential spread of this microsporidian also in other Southern areas of the country.
In order to verify this hypothesis, this study aimed to investigate the presence and distribution of Nosema spp. DNA in A. mellifera specimens and their products (i.e., pollen and honey) in selected apiaries of southwestern Italy.
2 Materials and methods
2.1 Study area and sampling
This study was approved by the Italian Ministry of Health within the project authorization no. IZS ME 08/22 RC aimed to assess the bee welfare and related productions of Southern Italy.
The study was carried out in the Campania region, Southern Italy, characterized by a typical Mediterranean temperate climate and progressively continental features of mainland and mountainous landscapes (37). In order to investigate aspects potentially correlated to presence of Nosema spp. and health status of bees, sampling areas were selected by using a risk index based on 5 environmental variables (i.e., pollution, land use, hydrographic network, air quality, bee density) and classified as high impact areas (HIAs) or low impact areas (LIAs) (38). Then, from March to July 2022–2023, bees, pollen and honey samples were collected in 10 different apiaries (5 LIAs and 5 HIAs) (Figure 1) by the staff of the Experimental Zooprophylactic Institute of Southern Italy (Portici, Italy) in collaboration with apiary owners that monitored any signs/symptoms of the hive potentially related to the infections. For each apiary, bee specimens, pollen and honey were collected using under-basket cages (39), combs inside the hives, and traps installed in front of the hives. For each apiary, a total of 50 bees, 10 g pollen and 1 g honey were collected from different hives, stored at −20°C into specific 500 mL glass jars labelled, and delivered to the Animal Health Department of the Experimental Zooprophylactic Institute of Southern Italy (Portici, Italy).
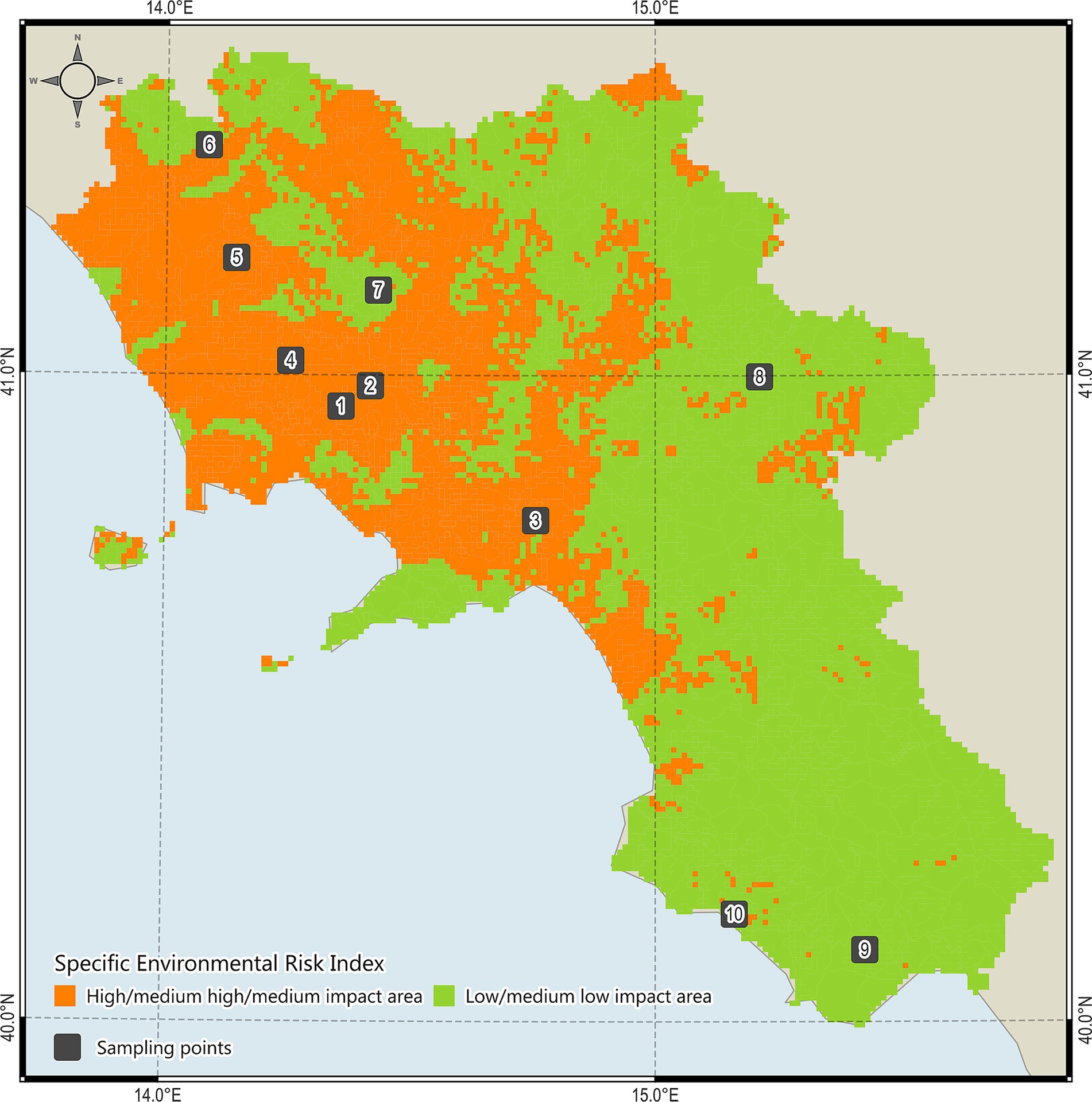
Figure 1. High impact areas (HIAs, n = 5) and low impact areas (LIAs, n = 5) investigated for Nosema and Nosema DNAs in the study area.
2.2 DNA extraction, PCR protocol, and sequencing
DNA was extracted from bee, pollen and honey samples using the QIAamp DNA mini-KIT tissue protocol (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. The obtained DNA was diluted 1:10 with Tris-glycine TGBE buffer and centrifuged at 5,000 g for 5 min. For each sample, DNA concentration was determined by BioPhotometer plus biophotometer (Eppendorf AG, Hamburg, Germany). All DNAs were screened for N. apis and N. ceranae by using two species-specific endpoint PCR protocols targeting the 16S rRNA gene, according to the National Reference Laboratory for honeybee health (40); primer details are listed in Table 1. PCRs were performed in a total volume of 25 μL containing 12.5 μL HotStarTaq Master Mix, 0.625 μL of each primer at concentration of 0.25 μM, 9.25 μL of Dnase/Rnase free water and 2 μL of DNA template. Amplification conditions included an initial denaturation/activation step at 95°C for 15 min, followed by 35 cycles of denaturation at 94°C for 15 s, primers annealing at 62°C for 30s and extension at 72°C per 30s, and a final extension step at 72°C for 7 min. Amplicons were displayed by automated capillary electrophoresis with the QIAxcel instrument (Qiagen, Hilden, Germany). Amplicons were then purified with the QIAquick PCR Purification kit (Qiagen, Hilden, Germany) and sequenced in both directions using the same primers as for PCR, with the Big Dye Terminator Cycle Sequencing Kit v1.1 in the automated sequencer 3,500 Genetic Analyzer (Thermo Fischer Scientific, United States). Consensus sequences were aligned and edited via ClustalW Multiple Alignment method in the BioEdit software (version 7.7) (41), and compared with those available in the GenBank database with the Basic Local Alignment Search Tool (BLAST).1 Edited sequences were then exported to DnaSP software (version 6) (42) for the computation of polymorphisms using Tajima’s (43) and Fu’s Fs tests (44).

Table 1. Primers for the detection of Nosema apis and Nosema ceranae used in this study.
All the sequences were submitted on GenBank database under the accession numbers PP758584 for N. apis and PP758589-PP758592 for N. ceranae, respectively.
2.3 Statistical analysis
Exact binomial 95% confidence intervals (95% CIs) by Wilson’s method were calculated for the proportions of infection herein found. The Fisher’s exact test was used for assessing any statistical differences in the frequency of infection according to the geographical origin of samples; a value of p < 0.05 was considered statistically significant. The Odds Ratio (OR) was used to verify the difference of infection risk according to the geographical origin of samples. Statistical analyses were performed by using the online software Epitools-Epidemiological Calculators (45). The distribution of Nosema spp.—positive samples in the study area was obtained with QGIS software (version 3.34.10-Prizren LTR).
3 Results
Out of 10 apiaries investigated, 6 (i.e., 60%, 95% CI: 31.3–83.2) tested positive for at least one sample to Nosema spp. DNA, being 2 positives only for N. apis, 2 only for N. ceranae and 2 co-infected (i.e., 20%, 5.7–51.0). Details of positivity to N. apis and N. ceranae, according to the different samples examined (i.e., bee specimen, pollen and honey) and apiary typology (i.e., HIAs or LIAs), are reported in Table 2. In all the apiaries tested positive for N. apis and N. ceranae, high mortality and low honey production were observed.
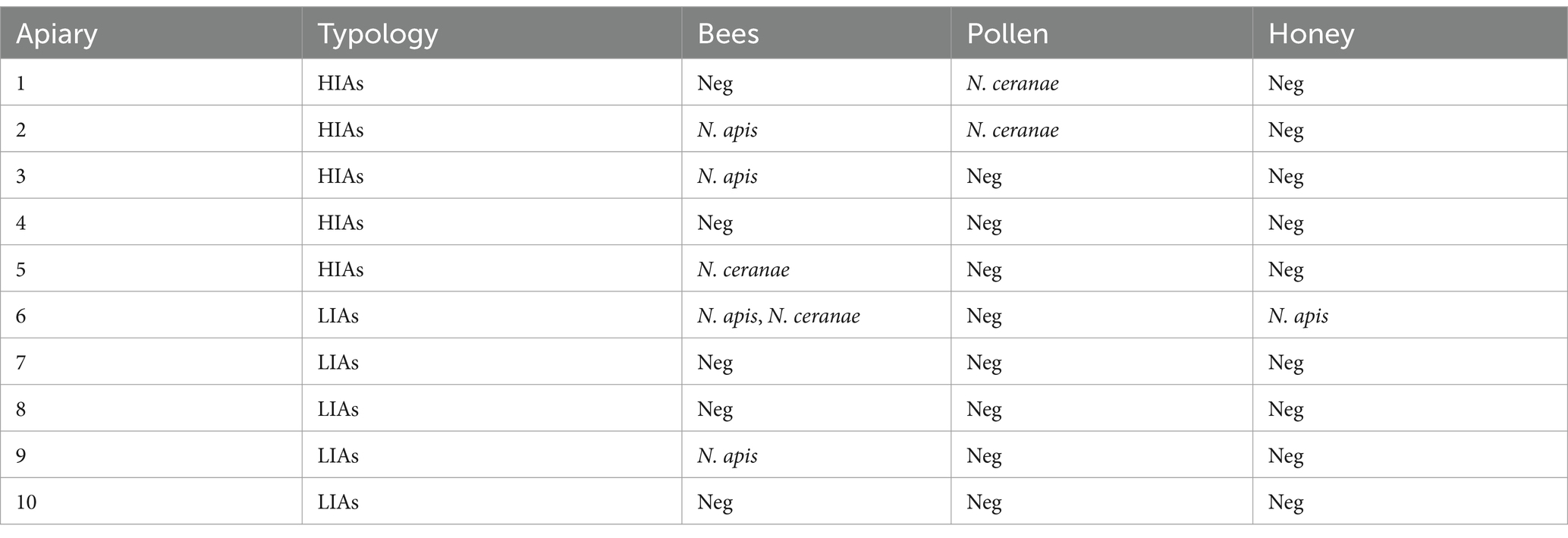
Table 2. Bee farms tested negative (Neg) and positive to Nosema spp. DNA, according to different samples (i.e., bee specimens, honey, pollen).
Although the difference of infection according to the origin of apiaries from HIAs and LIAs was not statistically significant (χ2 = 1.667, p = 0.200), a high risk of infection was observed for apiaries located in HIAs (OR = 6.00).
All the 16S rRNA partial sequences of N. apis and N. ceranae herein found had 100% query coverage and 99.5–100% nucleotide identity with those available in the GenBank database. Whereas all sequences of N. apis were identical to each other, four different sequences types of N. ceranae (i.e., ST1 in pollen, ST2 in pollen, ST3 in bee, ST4 in pollen) were identified showing single nucleotide polymorphisms (SNPs) in the positions 34, 122, 192 (Figure 2). The computation of polymorphisms revealed high haplotype diversity (i.e., Hd = 1.000) and low nucleotide diversity (i.e., Pi = 0.00913) of N. ceranae 16S rRNA sequences from this study, with not statistically significant negative values of both Tajima’s D and Fu’s Fs tests (Table 3).
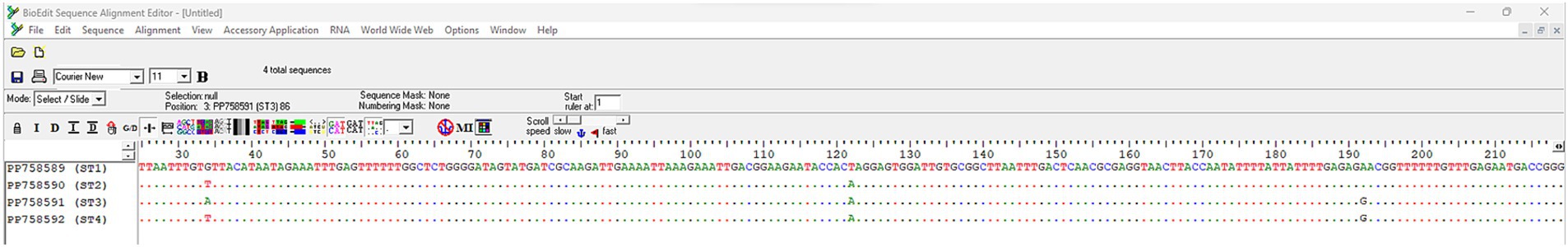
Figure 2. ClustalW multiple alignment of Nosema ceranae 16S rRNA sequence types showing single nucleotide polymorphisms in BioEdit software (version 7.7) (35).
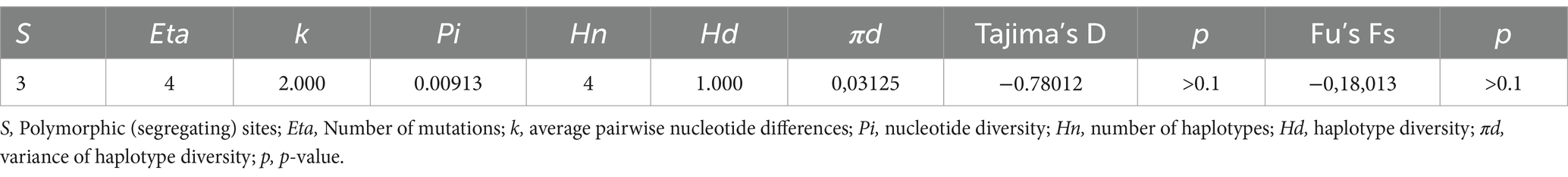
Table 3. Diversity and neutrality indices of Nosema ceranae 16S rRNA sequences (n = 4) from Southern Italy.
4 Discussion
This study reports for the first time N. apis and N. ceranae infections in A. mellifera populations, pollen and honey, from apiaries of Southwestern Italy.
The high prevalence of Nosema spp. in the apiaries herein investigated (i.e., 60%) indicates a wide spread of infection in the study area, in agreement with the national average (30–69%) (36), and with the last data available from Central (63.2%) (46), Northern (42.9–54.5%) (47, 48) and Southeastern Italy (100%) (15). The finding of N. apis is of concern due to its high pathogenic potential (49, 50) and the fact that this species has been previously reported only in 2010 in Northern Italy (36, 51), and recently in the Southeast (15). This study also confirms the occurrence of N. ceranae in the south (15, 36), as already outlined in areas of Northern (47, 48) and Central Italy (46, 52).
As well as in bees, the finding of N. apis and N. ceranae in honey and pollen samples, respectively, is not surprising given that the small size of their spores (2–6 μm) (53, 54), allow them to be transferred as wind-dispersed bioaerosols from the air to different surfaces, including pollen and honey (14, 55). Another potential route of transmission may occur via bee feces contaminated with spores, which are usually left by bees near the apiary, carried by the wind to the flowers, and then collected again by bees during pollination (56). In addition, feeding bee colonies with infected honey and pollen may represent a further risk for the transmission of Nosema spp. infections (57, 58).
The signs of infection herein observed, such as high mortality and low honey production, are in accordance with the clinical picture of nosemosis (13, 15, 52, 59).
Although no statistically significant difference between apiaries in HIAs and LIAs is observed (p = 0.200), a higher risk of infection is emerged in HIAs than LIAs (OR = 6.00), suggesting that environmental aspects of HIAs (i.e., pollution, intense land use, low air quality, high bee density) (38) are implicated in the epidemiology of Nosema spp., as well as in the overall welfare of the bee industry. This hypothesis finds support in studies indicating that environmental stressors may increase the virulence of N. ceranae (60) and influence the bee microbiota, thus altering the immune system (61) and susceptibility to pathogens (62–65). Accordingly, further studies for assessing the correlation between apiary typology and susceptibility to Nosema spp. infections are required.
The presence of SNPs in all the 16S rRNA sequences of N. ceranae from pollen (i.e., ST1, ST2, ST4) and bees (i.e., ST3) of this study, could be due to a wide genetic variability of this pathogen, in accordance with previous studies showing the presence of several sequence types of this species (51, 66–70). The high haplotype diversity and low nucleotide diversity of N. ceranae sequences herein found are indicative for a rapid demographic expansion of this microsporidian. In accordance, although not statistically significant, the negative values of Tajima’s D and Fu’S Fs suggest an excess of rare polymorphic sites which are typical features of both recent population expansion and presence of rare haplotypes compared to what is expected under neutrality, pointing to past bottleneck and/or purifying selection events (43, 44).
Finally, the fact that nosemosis is no longer a notifiable disease (71) underlines the importance of epidemiological investigations as the only way to highlight the presence of Nosema spp. in apiaries (15). Accordingly, future large-scale surveys are needed to investigate the circulation of Nosema spp. in other areas of Italy and the potential implications to the honey bee industry. At the same time, epidemiological and pathogenetic insights are required on little-investigated trypanosomatids (e.g., Lotmaria passim) circulating in bee colonies (72) and honey with a high percentage (80%) of co-occurrence with N. ceranae microsporidia (55).
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: https://www.ncbi.nlm.nih.gov/genbank/, PP758584; https://www.ncbi.nlm.nih.gov/genbank/, PP758589; https://www.ncbi.nlm.nih.gov/genbank/, PP758590; https://www.ncbi.nlm.nih.gov/genbank/, PP758591; https://www.ncbi.nlm.nih.gov/genbank/, PP758592.
Ethics statement
Ethical approval was not required for the studies involving animals in accordance with the local legislation and institutional requirements because this study was approved by the Italian Ministry of Health within the project titled “Studio degli alcaloidi pirrolizidinici e dei metaboliti secondari nei prodotti dell’alveare: verso la definizione di mappe apistiche in ottica di sicurezza alimentare” (grant number: IZS ME 08/22 RC). Written informed consent was not obtained from the owners for the participation of their animals in this study because in accordance with national legislation and institutional requirements.
Author contributions
GS: Conceptualization, Writing – original draft. LD’A: Conceptualization, Writing – review & editing. ML: Methodology, Writing – review & editing. AM: Methodology, Writing – review & editing. YP: Data curation, Formal analysis, Writing – review & editing. ME: Data curation, Formal analysis, Writing – review & editing. SR: Data curation, Formal analysis, Writing – review & editing. DS: Data curation, Formal analysis, Writing – review & editing. FG: Data curation, Writing – review & editing. ND’A: Writing – review & editing, Data curation, Formal analysis. RM: Writing – review & editing. PK: Writing – review & editing. GR: Funding acquisition, Project administration, Resources, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by the project entitled “Studio degli alcaloidi pirrolizidinici e dei metaboliti secondari nei prodotti dell’alveare: verso la definizione di mappe apistiche in ottica di sicurezza alimentare” (grant no. IZS ME 08/22 RC) funded by the Italian Ministry of Health.
Acknowledgments
The authors are grateful to Anna Granato (Istituto Zooprofilattico Sperimentale delle Venezie, Padua, Italy) for support in molecular analysis and to the apiary owners involved in this study for their kind cooperation in sampling and monitoring activities.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2024.1530169/full#supplementary-material
Footnotes
References
1. World Health Organization. Antimicrobial resistance: global report on surveillance (2014). Available at: https://www.who.int/publications/i/item/9789241564748 (Accessed April 23, 2024).
2. Crane, E. Bees and beekeeping: Science, practice and world resources. Ithaca: Cornell University Press (1990).
3. Moeller, FE. Nosema disease-its control in honey bee colonies (1978). Available at: https://ageconsearch.umn.edu/record/158100?v=pdf (Accessed April 24, 2024).
4. Morse, RA, and Nowogrodzki, R. Honey bee pests, predators and diseases. 2nd ed. Ithaca: Cornell University Press (1990).
5. Higes, M, García-Palencia, P, Urbieta, A, Nanetti, A, and Martín-Hernández, R. Nosema apis and Nosema ceranae tissue tropism in worker honey bees (Apis mellifera). Vet Pathol. (2020) 57:132–8. doi: 10.1177/0300985819864302
6. Moretto, MM, Khan, IA, and Weiss, LM. Gastrointestinal cell mediated immunity and the microsporidia. PLoS Pathog. (2012) 8:e1002775. doi: 10.1371/journal.ppat.1002775
7. Botías, C, Martín-Hernández, R, Barrios, L, Meana, A, and Higes, M. Nosema spp. infection and its negative effects on honey bees (Apis mellifera iberiensis) at the colony level. Vet Res. (2013) 44:25. doi: 10.1186/1297-9716-44-25
8. Higes, M, Nozal, M, Alvaro, A, Barrios, L, Meana, A, Martin-Hernandez, R, et al. The stability and effectiveness of fumagillin in controlling Nosema ceranae (microsporidia) infection in honey bees (Apis mellifera) under laboratory and field conditions. Apidologie. (2011) 42:364–77. doi: 10.1007/s13592-011-0003-2
9. Pizarro, R, and Montenegro, G. Las claves del Síndrome de Despoblamiento de Colmenas (2012). Available at: https://agronomia.uc.cl/component/com_sobipro/Itemid,232/pid,920/sid,1082/ (Accessed April 24, 2024).
10. Fries, I. Nosema ceranae in European honey bees (Apis mellifera). J Invertebr Pathol. (2010) 103:S73–9. doi: 10.1016/j.jip.2009.06.017
12. Giersch, T, Berg, T, Galea, F, and Hornitzky, M. Nosema ceranae infects honey bees (Apis mellifera) and contaminates honey in Australia. Apidologie. (2009) 40:117–23. doi: 10.1051/apido/2008065
13. Chen, Y, Evans, JD, Zhou, L, Boncristiani, H, Kimura, K, Xiao, T, et al. Asymmetrical coexistence of Nosema ceranae and Nosema apis in honey bees. J Invertebr Pathol. (2009) 101:204–9. doi: 10.1016/j.jip.2009.05.012
14. Sokół, R, and Michalczyk, M. Detection of Nosema spp. in worker bees, pollen and bee bread during the honey flow season. Acta Vet Brno. (2016) 85:261–6. doi: 10.2754/avb201685030261
15. Pandiscia, A, Lorusso, P, Manfredi, A, Bonerba, E, Bozzo, G, Tantillo, GM, et al. Nosema apis and Nosema ceranae in beehives of the Apulian region of Italy: how citizen science can support scientific research. Agriculture. (2024) 14:583. doi: 10.3390/agriculture14040583
16. Tokarev, YS, Huang, WF, Solter, LF, Malysh, JM, Becnel, JJ, and Vossbrinck, CR. A formal redefinition of the genera Nosema and Vairimorpha (microsporidia: Nosematidae) and reassignment of species based on molecular phylogenetics. J Invertebr Pathol. (2020) 169:107279. doi: 10.1016/j.jip.2019.107279
17. Bartolomé, C, Higes, M, Hernández, RM, Chen, YP, Evans, JD, and Huang, Q. The recent revision of the genera Nosema and Vairimorpha (microsporidia: Nosematidae) was flawed and misleads bee scientific community. J Invertebr Pathol. (2024) 206:108146. doi: 10.1016/j.jip.2024.108146
18. Paxton, RJ, Klee, J, Korpela, S, and Fries, I. Nosema ceranae has infected Apis mellifera in Europe since at least 1998 and may be more virulent than Nosema apis. Apidologie. (2007) 38:558–65. doi: 10.1051/apido:2007037
19. Higes, M, Martín, R, and Meana, A. Nosema ceranae, a new microsporidian parasite in honeybees in Europe. J Invertebr Pathol. (2006) 92:93–5. doi: 10.1016/j.jip.2006.02.005
20. Sampedro, J. El asesino de las abejas se llama Nosema (2007). http://elpais.com/diario/2007/09/09/domingo/1189309957_850215.html (Accessed April 28, 2024).
21. Villa, J, Bourgeois, A, and Danka, R. Negative evidence for effects of genetic origin of bees on Nosema ceranae, positive evidence for effects of Nosema ceranae on bees. Apidologie. (2013) 2013:511–8. doi: 10.1007/s13592-013-0201-1
22. Gajger, IT, Vugrek, O, Grilec, D, and Petrinec, Z. Prevalence and distribution of Nosema ceranae in Croatian honeybee colonies. Vet Med. (2010) 55:457–62. doi: 10.17221/2983-VETMED
23. Stevanovic, J, Stanimirovic, Z, Genersch, E, Kovacevic, SR, Ljubenkovic, J, Radakovic, M, et al. Dominance of Nosema ceranae in honey bees in the Balkan countries in the absence of symptoms of colony collapse disorder. Apidologie. (2011) 42:49–58. doi: 10.1051/apido/2010034
24. Stevanovic, J, Simeunovic, P, Gajic, B, Lakic, N, Radovic, D, Fries, I, et al. Characteristics of Nosema ceranae infection in Serbian honey bee colonies. Apidologie. (2013) 44:522–36. doi: 10.1007/s13592-013-0203-z
25. Stevanovic, J, Schwarz, RS, Vejnovic, B, Evans, JD, Irwin, RE, Glavinic, U, et al. Species-specific diagnostics of Apis mellifera trypanosomatids: a nine-year survey (2007-2015) for trypanosomatids and microsporidians in Serbian honey bees. J Invertebr Pathol. (2016) 139:6–11. doi: 10.1016/j.jip.2016.07.001
26. Hulaj, B, Granato, A, Bordin, F, Goga, I, Merovci, X, Caldon, M, et al. Emergent and known honey bee pathogens through passive surveillance in the republic of Kosovo. Appl Sci. (2024) 14:987. doi: 10.3390/app14030987
27. Williams, GR, Shafer, ABA, Rogers, REL, Shutler, D, and Stewart, DT. First detection of Nosema ceranae, a microsporidian parasite of European honey bees (Apis mellifera), in Canada and Central USA. J Invertebr Pathol. (2008) 97:189–92. doi: 10.1016/j.jip.2007.08.005
28. Castelli Norando, L. Prevalencia y distribución geográfica de Nosema apis y Nosema ceranae en Apis mellifera de Uruguay. Dissertation/master’s thesis. Montevideo, UY: University of the Republic (2012).
29. Médici, SK, Sarlo, EG, Porrini, MP, Braunstein, M, and Eguaras, MJ. Genetic variation and widespread dispersal of Nosema ceranae in Apis mellifera apiaries from Argentina. Parasitol Res. (2012) 110:859–64. doi: 10.1007/s00436-011-2566-2
30. Plischuk, S. Protistas entomopatógenos asociados a apoideos (Hymenoptera: Apoidea) polinizadores de la región Pampeana. Ecosistemas. (2013) 22:89–91. doi: 10.7818/ECOS.2013.22-1.20
31. Cornelissen, B, Neumann, P, and Schweiger, O. Global warming promotes biological invasion of a honey bee pest. Glob Change Biol. (2019) 25:3642–55. doi: 10.1111/gcb.14791
32. Urbieta-Magro, A, Higes, M, Meana, A, Gómez-Moracho, T, Rodríguez-García, C, Barrios, L, et al. The levels of natural Nosema spp. infection in Apis mellifera iberiensis brood stages. Int J Parasitol. (2019) 49:657–67. doi: 10.1016/j.ijpara.2019.04.002
33. Li, YH, Chen, YH, Chang, FM, Wu, MC, and Nai, YS. Monitoring the season-prevalence relationship of Vairimorpha ceranae in honey bees (Apis mellifera) over one year and the primary assessment of probiotic treatment in Taichung. Taiwan Insects. (2024) 15:204. doi: 10.3390/insects15030204
34. McAfee, A, Alavi-Shoushtari, N, Tran, L, Labuschagne, R, Cunningham, M, Tsvetkov, N, et al. Climatic predictors of prominent honey bee (Apis mellifera) disease agents: Varroa destructor, Melissococcus plutonius, and Vairimorpha spp. PLoS Climate. (2024) 3:e0000485. doi: 10.1371/journal.pclm.0000485
35. Martín-Hernández, R, Botías, C, Bailón, EG, Martínez-Salvador, A, Prieto, L, Meana, A, et al. Microsporidia infecting Apis mellifera: coexistence or competition. Is Nosema ceranae replacing Nosema apis? Environ Microbiol. (2012) 14:2127–38. doi: 10.1111/j.1462-2920.2011.02645.x
36. Porrini, C, Mutinelli, F, Bortolotti, L, Granato, A, Laurenson, L, Roberts, K, et al. The status of honey bee health in Italy: results from the Nationwide bee monitoring network. PLoS One. (2016) 11:e0155411. doi: 10.1371/journal.pone.0155411
37. Sgroi, G, Iatta, R, Lovreglio, P, Stufano, A, Laidoudi, Y, Mendoza-Roldan, JA, et al. Detection of endosymbiont Candidatus Midichloria mitochondrii and Tickborne pathogens in humans exposed to tick bites. Italy Emerg Infect Dis. (2022) 28:1824–32. doi: 10.3201/eid2809.220329
38. Signorelli, D, D’Auria, LJ, Di Stasio, A, Gallo, A, Siciliano, A, Esposito, M, et al. Application of a quality-specific environmental risk index for the location of hives in areas with different pollution impacts. Agriculture. (2023) 13:998. doi: 10.3390/agriculture13050998
39. Bellucci, V, Lucci, S, Bianco, P, Ubaldi, A, Felicioli, A, Porrini, C, et al. Monitoring honey bee health in five natural protected areas in Italy. Vet Ital. (2019) 55:15–25. doi: 10.12834/VetIt.1209.6739.4
40. Martín-Hernández, R, Meana, A, Prieto, L, Salvador, AM, Garrido-Bailón, E, and Higes, M. Outcome of colonization of Apis mellifera by Nosema ceranae. Appl Environ Microbiol. (2007) 73:6331–8. doi: 10.1128/AEM.00270-07
41. Hall, TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp Ser. (1999) 41:95–8.
42. Rozas, J, Ferrer-Mata, A, Sánchez-DelBarrio, JC, Guirao-Rico, S, Librado, P, Ramos-Onsins, SE, et al. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol Biol Evol. (2017) 34:3299–302. doi: 10.1093/molbev/msx248
43. Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics. (1989) 123:585–95. doi: 10.1093/genetics/123.3.585
44. Fu, YX. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics. (1997) 147:915–25. doi: 10.1093/genetics/147.2.915
45. Sergeant, ESG. Epitools: epidemiological calculators (2018). Available at: https://epitools.ausvet.com.au (Accessed June 20, 2024(.
46. Papini, R, Mancianti, F, Canovai, R, Cosci, F, Rocchigiani, G, Benelli, G, et al. Prevalence of the microsporidian Nosema ceranae in honeybee (Apis mellifera) apiaries in Central Italy. Saudi J Biol Sci. (2017) 24:979–82. doi: 10.1016/j.sjbs.2017.01.010
47. Cilia, G, Tafi, E, Zavatta, L, Caringi, V, and Nanetti, A. The epidemiological situation of the managed honey bee (Apis mellifera) colonies in the Italian region Emilia-Romagna. Vet Sci. (2022) 9:437. doi: 10.3390/vetsci9080437
48. Cilia, G, Flaminio, S, Zavatta, L, Ranalli, R, Quaranta, M, Bortolotti, L, et al. Occurrence of honey bee (Apis mellifera L.) pathogens in wild pollinators in northern Italy. Front Cell Infect Microbiol. (2022) 12:907489. doi: 10.3389/fcimb.2022.907489
49. Forsgren, E, and Fries, I. Comparative virulence of Nosema ceranae and Nosema apis in individual European honey bees. Vet Parasitol. (2010) 170:212–7. doi: 10.1016/j.vetpar.2010.02.010
50. Milbrath, MO, van Tran, T, Huang, WF, Solter, LF, Tarpy, DR, Lawrence, F, et al. Comparative virulence and competition between Nosema apis and Nosema ceranae in honey bees (Apis mellifera). J Invertebr Pathol. (2015) 125:9–15. doi: 10.1016/j.jip.2014.12.006
51. Ferroglio, E, Zanet, S, Peraldo, N, Tachis, E, Trisciuoglio, A, Laurino, D, et al. Nosema ceranae has been infecting honey bees Apis mellifera in Italy since at least 1993. J Apic Res. (2012) 52:60–1. doi: 10.3896/IBRA.1.52.2.11
52. Cilia, G, Sagona, S, Giusti, M, Jarmela Dos Santos, PE, Nanetti, A, and Felicioli, A. Nosema ceranae infection in honeybee samples from Tuscanian archipelago (Central Italy) investigated by two qPCR methods. Saudi. J Biol Sci. (2019) 26:1553–6. doi: 10.1016/j.sjbs.2018.11.017
53. Fries, I, Feng, F, da Silva, A, Slemenda, SB, and Pieniazek, NJ. Nosema ceranae n. sp. (Microspora, Nosematidae), morphological and molecular characterization of a microsporidian parasite of the Asian honey bee Apis cerana (Hymenoptera, Apidae). Eur J Protistol. (1996) 32:356–65. doi: 10.1016/S0932-4739(96)80059-9
54. Higes, M, Martín-Hernández, R, and Meana, A. Nosema ceranae in Europe: an emergent type C nosemosis. Apidologie. (2010) 41:375–92. doi: 10.1051/apido/2010019
55. Ribani, A, Utzeri, VJ, Taurisano, V, and Fontanesi, L. Honey as a source of environmental DNA for the detection and monitoring of honey bee pathogens and parasites. Vet Sci. (2020) 7:113. doi: 10.3390/vetsci7030113
56. Sulborska, A, Horecka, B, Cebrat, M, Kowalczyk, M, Skrzypek, TH, Kazimierczak, W, et al. Microsporidia Nosema spp. – obligate bee parasites are transmitted by air. Sci Rep. (2019) 9:14376. doi: 10.1038/s41598-019-50974-8
57. Salkova, D, Balkanska, R, Shumkova, R, Lazarova, S, Radoslavov, G, and Hristov, P. Molecular detection and phylogenetic relationships of honey bee-associated viruses in bee products. Vet Sci. (2024) 11:369. doi: 10.3390/vetsci11080369
58. Balkanska, R, Shumkova, R, Atsenova, N, Salkova, D, Dundarova, H, Radoslavov, G, et al. Molecular detection and phylogenetic analysis of deformed wing virus and Sacbrood virus isolated from pollen. Vet Sci. (2023) 10:140. doi: 10.3390/vetsci10020140
59. Zhang, Y, Lu, X, Huang, S, Zhang, L, Su, S, and Huang, WF. Nosema ceranae infection enhances Bifidobacterium spp. abundances in the honey bee hindgut. Apidologie. (2019) 50:353–62. doi: 10.1007/s13592-019-00644-5
60. Higes, M, Meana, A, Bartolomé, C, Botías, C, and Martín-Hernández, R. Nosema ceranae (microsporidia), a controversial 21st century honey bee pathogen. Environ Microbiol Rep. (2013) 5:17–29. doi: 10.1111/1758-2229.12024
61. Motta, EVS, Powell, JE, and Moran, NA. Glyphosate induces immune dysregulation in honey bees. Anim Microbiome. (2022) 4:16. doi: 10.1186/s42523-022-00165-0
62. Castelli, L, Branchiccela, B, Garrido, M, Invernizzi, C, Porrini, M, Romero, H, et al. Impact of nutritional stress on honeybee gut microbiota, immunity, and Nosema ceranae infection. Microb Ecol. (2020) 80:908–19. doi: 10.1007/s00248-020-01538-1
63. Alberoni, D, Di Gioia, D, and Baffoni, L. Alterations in the microbiota of caged honeybees in the presence of Nosema ceranae infection and related changes in functionality. Microb Ecol. (2023) 86:601–16. doi: 10.1007/s00248-022-02050-4
64. Braglia, C, Alberoni, D, Garrido, PM, Porrini, MP, Baffoni, L, Scott, D, et al. Vairimorpha (Nosema) ceranae can promote Serratia development in honeybee gut: an underrated threat for bees? Front Cell Infect Microbiol. (2024) 14:1323157. doi: 10.3389/fcimb.2024.1323157
65. Tiritelli, R, Flaminio, S, Zavatta, L, Ranalli, R, Giovanetti, M, Grasso, DA, et al. Ecological and social factors influence interspecific pathogens occurrence among bees. Sci Rep. (2024) 14:5136. doi: 10.1038/s41598-024-55718-x
66. Gómez-Moracho, T, Maside, X, Martín-Hernández, R, Higes, M, and Bartolomé, C. High levels of genetic diversity in Nosema ceranae within Apis mellifera colonies. Parasitology. (2014) 141:475–81. doi: 10.1017/S0031182013001790
67. Huang, Q, Wu, ZH, Li, WF, Guo, R, Xu, JS, Dang, XQ, et al. Genome and evolutionary analysis of Nosema ceranae: a microsporidian parasite of honey bees. Front Microbiol. (2021) 12:645353. doi: 10.3389/fmicb.2021.645353
68. Wei, X, Evans, JD, Chen, Y, and Huang, Q. Spillover and genome selection of the gut parasite Nosema ceranae between honey bee species. Front Cell Infect Microbiol. (2022) 12:1026154. doi: 10.3389/fcimb.2022.1026154
69. Blot, N, Clémencet, J, Jourda, C, Lefeuvre, P, Warrit, N, Esnault, O, et al. Geographic population structure of the honeybee microsporidian parasite Vairimorpha (Nosema) ceranae in the south West Indian Ocean. Sci Rep. (2023) 13:12122. doi: 10.1038/s41598-023-38905-0
70. Guo, S, Zang, H, Liu, X, Jing, X, Liu, Z, Zhang, W, et al. Full-length transcriptome construction and systematic characterization of virulence factor-associated isoforms in Vairimorpha (Nosema) Ceranae. Genes. (2024) 15:1111. doi: 10.3390/genes15091111
71. European Union. Commission delegated regulation (EU) 2018/1629 of 25 July 2018. Amending the list of diseases set out in annex ii to regulation (EU) 2016/429 of the European Parliament and of the council on transmissible animal diseases and amending and repealing certain acts in the area of animal health (‘animal health law’) (2018). https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32018R1629 (Accessed October 10, 2024).
72. Tafi, E., Capano, V., Nanetti, A., and Cilia, G. A two-year nationwide molecular survey on Trypanosomatids occurrence in Italian honey bee (Apis mellifera L.) colonies. SSRN [Preprint] (2024). Available at: https://ssrn.com/abstract=4984444 (Accessed December 9, 2024).
Keywords: bee decline, honeybee, honey, pollen, Nosema, nosemosis, Vairimorpha, Italy
Citation: Sgroi G, D’Auria LJ, Lucibelli MG, Mancusi A, Proroga YTR, Esposito M, Rea S, Signorelli D, Gargano F, D’Alessio N, Manoj RRS, Khademi P and Rofrano G (2025) Bees on the run: Nosema spp. (Microsporidia) in Apis mellifera and related products, Italy. Front. Vet. Sci. 11:1530169. doi: 10.3389/fvets.2024.1530169
Edited by:
Nicola Pugliese, University of Bari Aldo Moro, ItalyReviewed by:
Jevrosima Stevanovic, University of Belgrade, SerbiaPetar Hristov, Bulgarian Academy of Sciences, Bulgaria
Copyright © 2025 Sgroi, D’Auria, Lucibelli, Mancusi, Proroga, Esposito, Rea, Signorelli, Gargano, D’Alessio, Manoj, Khademi and Rofrano. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrea Mancusi, YW5kcmVhLm1hbmN1c2lAaXpzbXBvcnRpY2kuaXQ=
†These authors have contributed equally to this work and share first authorship