 Nora M. Raschle
Nora M. Raschle Lynn V. Fehlbaum
Lynn V. Fehlbaum Willeke M. Menks
Willeke M. Menks Felix Euler1
Felix Euler1 Philipp Sterzer
Philipp Sterzer Christina Stadler
Christina Stadler
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Psychol. , 01 September 2017
Sec. Cognitive Science
Volume 8 - 2017 | https://doi.org/10.3389/fpsyg.2017.01489
The human brain has the capacity to integrate various sources of information and continuously adapts our behavior according to situational needs in order to allow a healthy functioning. Emotion–cognition interactions are a key example for such integrative processing. However, the neuronal correlates investigating the effects of emotion on cognition remain to be explored and replication studies are needed. Previous neuroimaging studies have indicated an involvement of emotion and cognition related brain structures including parietal and prefrontal cortices and limbic brain regions. Here, we employed whole brain event-related functional magnetic resonance imaging (fMRI) during an affective number Stroop task and aimed at replicating previous findings using an adaptation of an existing task design in 30 healthy young adults. The Stroop task is an indicator of cognitive control and enables the quantification of interference in relation to variations in cognitive load. By the use of emotional primes (negative/neutral) prior to Stroop task performance, an emotional variation is added as well. Behavioral in-scanner data showed that negative primes delayed and disrupted cognitive processing. Trials with high cognitive demand furthermore negatively influenced cognitive control mechanisms. Neuronally, the emotional primes consistently activated emotion-related brain regions (e.g., amygdala, insula, and prefrontal brain regions) while Stroop task performance lead to activations in cognition networks of the brain (prefrontal cortices, superior temporal lobe, and insula). When assessing the effect of emotion on cognition, increased cognitive demand led to decreases in neural activation in response to emotional stimuli (negative > neutral) within prefrontal cortex, amygdala, and insular cortex. Overall, these results suggest that emotional primes significantly impact cognitive performance and increasing cognitive demand leads to reduced neuronal activation in emotion related brain regions, and therefore support previous findings investigating emotion–cognition interaction in healthy adults. Moreover, emotion and cognition seem to be tightly related to each other, as indicated by shared neural networks involved in both of these processes. Emotion processing, cognitive control, and their interaction are crucial for healthy functioning and a lack thereof is related to psychiatric disorders such as, disruptive behavior disorders. Future studies may investigate the neural characteristics of children and adolescents with disruptive behavior disorders.
An adequate handling of emotional information is a key factor for healthy functioning within our everyday life. How a person processes and regulates emotions impacts their cognition, behavior, and well-being (Dolan, 2002; Gross, 2002; John and Gross, 2004). Thereby, emotion processing not only influences cognitive control, but cognitive control may likewise affect emotions. Research has indicated that a fine balance of the emotion and cognition networks ultimately allows appropriate functioning (Hart et al., 2010). A failure to successfully process or regulate emotions is characteristic for different mental health disorders, including disruptive behavior disorders (Sterzer et al., 2005), attention-deficit/hyperactivity disorder (ADHD; Walcott and Landau, 2004), or psychosis (Livingstone et al., 2009). Therefore, an improved understanding of the mechanism supporting successful emotion regulation skills is of utmost personal, clinical, and societal relevance (Gross, 2002).
Behavioral research studies have demonstrated that emotional stimuli can positively or negatively impact cognitive processing. For example, the presentation of emotional stimuli has shown to disrupt working memory performance (Dolcos and McCarthy, 2006) and impact reaction times during a perceptual task (Gupta and Deak, 2015). Similarly, it was demonstrated that the presence of an emotional stimulus can reduce task accuracy and reaction times during Stroop task performance, which reflects cognitive control mechanisms (Blair et al., 2007; Hart et al., 2010; Uher et al., 2014). Visually presented and/or auditory-induced emotions can also positively influence cognition, resulting in improved accuracy or shorter reaction times during tasks including conflict processing, visual attention, or decision making (Schupp et al., 2007; Kanske and Kotz, 2011; Zinchenko et al., 2015). Factors that are known to influence the interaction of cognitive and emotional processes include cognitive load, level of threat, physical stimulus properties, position of emotional distractors (left or right hemifield), individual differences, and the availability of conflict-resolving brain resources (Arnsten and Goldman-Rakic, 1998; Hartikainen et al., 2000; Pessoa, 2009; Thompson et al., 2010; Cohen and Henik, 2012; Gupta and Raymond, 2012; Kanske, 2012; Okon-Singer et al., 2013; Gupta et al., 2016). By transiently enhancing or diminishing cognitive functioning, emotional states may thus impact the control of thoughts and behavior in order to meet situational demands (Gray et al., 2002).
Neuroimaging methods, such as functional magnetic resonance imaging (fMRI), can investigate the neural networks underlying emotional and cognitive processes as well as their interaction. Brain regions responsible for simple emotion-processing tasks are the amygdala, right insula, as well as the medial and ventrolateral prefrontal cortex (Phan et al., 2002; Dolcos and McCarthy, 2006; Van Dillen et al., 2009). Thereby, the engagement of individual brain regions depends on the quality of the emotion being processed. For example fear is particularly known to elicit amygdala activation, sadness is commonly represented by subcallosal cingulate activity, and emotion processing tasks with an additional cognitive component (e.g., emotional recall) also target the insular and anterior cingulate cortex (for a review see Phan et al., 2002). Brain regions associated with simple cognitive control (e.g., during working memory, conflict resolution, inhibition, or emotion regulation tasks) include the ventromedial, right (dorso-)lateral and orbital prefrontal cortex, lateral and right superior parietal cortex, and anterior cingulate cortex (Phan et al., 2002; Ochsner et al., 2004; Ochsner and Gross, 2005; Dolcos and McCarthy, 2006; Van Dillen et al., 2009; Pitskel et al., 2011).
To date, several fMRI studies have aimed at targeting the more complex interaction between cognition and emotion. The most commonly identified neural correlates of emotion–cognition interaction sites include parietal and prefrontal cortices, as well as limbic brain regions (i.e., cingulate, amygdala, and insula; Gray et al., 2002; Etkin et al., 2006; Blair et al., 2007; Van Dillen et al., 2009; Hart et al., 2010; Melcher et al., 2011; Kellermann et al., 2012; Gu et al., 2013; Cromheeke and Mueller, 2014). For example, Etkin et al. (2006) used an emotional conflict task and found that neural activation within the amygdala, dorsomedial-, and dorsolateral prefrontal cortex represents the level of emotional conflict, while the rostral anterior cingulate may reflect emotional conflict per se (Etkin et al., 2006). Likewise, Gu et al. (2013) identified shared and distinct brain regions responsible for cognitive and emotion processing or the interaction of both (Gu et al., 2013). In particular, an interaction effect was observed within in bilateral anterior insula, somatosensory cortices, and frontoparietal regions. Using an emotional working memory task, Gray et al. (2002) pinpointed left and right lateral prefrontal cortex as the site of emotion–cognition interaction. And finally, Blair et al. (2007) as well as Hart et al. (2010) combined emotional stimuli and Stroop task performance within their designs in order to elicit areas that are dynamically modulated either by increased emotional or enhanced cognitive demands. Again, bilateral amygdala, inferior frontal/ventrolateral prefrontal, and the cingulate cortex were identified as areas of neural changes dependent on cognitive and/or emotional load (Blair et al., 2007; Hart et al., 2010).
For the present study we adapted and re-evaluated the affective number Stroop task as implemented by Hart et al. (2010). Our goal was the investigation of dynamic changes in either the emotion or cognition network elicited by both variations in emotional content (through the use of negative as opposed to neutral images), and changes in cognitive demand (using different conditions of a number Stroop task). A further motivation for this study was the characterization of the neural correlates representing the effect of emotions on cognition in young adults as a basis for future studies in children and adolescents with social disorders (e.g., disruptive behavior disorders). This is of particular interest since behavioral studies have already demonstrated altered emotion–cognition interactions in disruptive behavior disorders (Euler et al., 2014). Nevertheless, the neural correlates in these clinical populations are still unknown. Therefore, our aims were to: (I) elicit activation in emotion-related brain regions in response to the affective primes implemented within our task (e.g., amygdala, insula, and prefrontal cortex Phan et al., 2002); (II) demonstrate activation in cognitive brain regions in response to the Stroop task (e.g., prefrontal cortex, lateral, and right superior parietal cortex, anterior cingulate cortex; Laird et al., 2005); (III) investigate previously identified brain regions that are significant in relation to the emotion–cognition interaction (e.g., amygdala, prefrontal cortex, and anterior insula; Gray et al., 2002; Etkin et al., 2006; Blair et al., 2007; Hart et al., 2010; Gu et al., 2013) and assess their involvement within the task described here. Based on strong prior behavioral evidence (Homack and Riccio, 2004), we hypothesized to observe delayed reaction times and reduced task accuracy for trials with increased cognitive load (i.e., from congruent to stars to incongruent Stroop trials), and for trials following affective (negative) primes compared to neutral primes. Neurally, we expected to replicate the above mentioned findings of changes in neural activation patterns within the emotion and/or cognition network in dependence to cognitive load (Gray et al., 2002; Etkin et al., 2006; Blair et al., 2007; Van Dillen et al., 2009; Hart et al., 2010; Melcher et al., 2011; Kellermann et al., 2012; Gu et al., 2013; Cromheeke and Mueller, 2014).
Thirty healthy, German-speaking volunteers (mean age: 21.74 years; range 19–24 years; 15 males) with no prior psychological or neurological history were included in the current study. Participants took part in one testing session that included psychometric testing, one functional neuroimaging task and a T1-weighted structural image acquisition. Two participants were excluded from analysis since one of them had completely missing and the other person very low in-scanner performance (e.g., more than 20% misses in each run). All participants were further right-handed, had normal or corrected-to-normal vision, and provided written informed consent as approved by the local ethics committee (Ethikkommission der Nordwest- und Zentralschweiz).
Participants included in this study completed a battery of standardized tests comprising verbal and non-verbal IQ [German version of the Vocabulary and Matrix reasoning subtests of the WAIS-IV (Petermann, 2012), present mood (EWL; Janke, 1978)], behavioral and emotional functioning (YSR; Achenbach, 1991), psychopathic traits (YPI; Andershed et al., 2007), and handedness (EDI; Caplan and Mendoza, 2011). The YPI, YSR, and EWL were missing for one person. The resulting behavioral group characteristics are provided in Table 1.
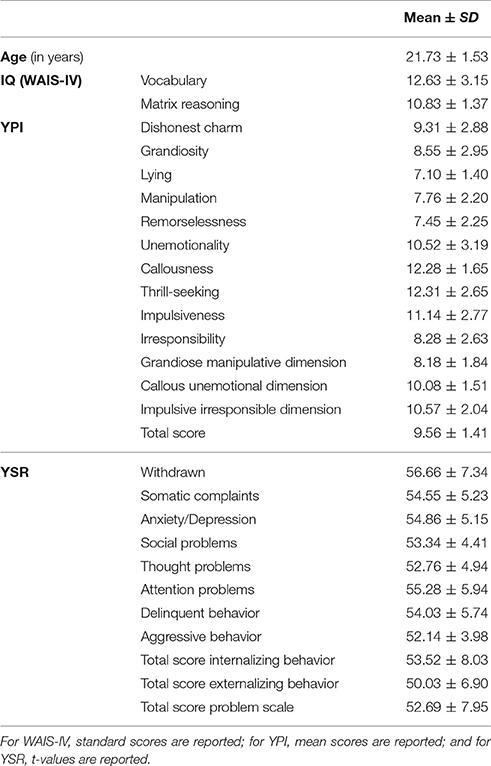
Table 1. Behavioral group characteristics.
The neuroimaging session included event-related functional neuroimaging during the performance of an emotional number Stroop task. Additionally, T1-weighted structural images were acquired for each participant. The emotional number Stroop task was adapted and modified based on a design described by Hart et al. (2010) (see trial design in Figure 1). We decided to use a number Stroop task as it has particularly been developed for use in the MR environment and has previously successfully been implemented in neuroimaging research studies (e.g., Blair et al., 2007; Hart et al., 2010). Each trial started with an emotional prime of either neutral or negative valence, presented for 150 ms. Negative (Neg) or neutral (Neu) primes were first followed by an item of the number Stroop task presented for 1,500 ms, before a short relaxation period of 350 ms ended the trial. During the number Stroop task, participants were presented with an array of 1, 2, 3, or 4 digits and were asked to indicate through button press the number of items presented. The number of items was either congruent (C) in relation to the printed digits (e.g., the digit 4 in an array of 4) or incongruent (IC) with the printed digits (e.g., digit 4 in an array of 3). Star shaped stimuli (S) were used as a control condition (no interference of digit and item number) and null trials (trials with a black screen instead of the Stroop trial) were added during the randomization process. Emotional stimuli were adapted from the Developmental Affective Photo System (DAPS; a child-appropriate picture system Cordon et al., 2013), which uses part of the IAPS (International Affective Picture System Lang et al., 2008) commonly used in adults. We implemented DAPS images because this task was designed to ultimately be employed in children and adolescents with psychiatric disorders. However, given that all images remained part of the IAPS system, the chosen stimuli were considered suitable for both adult and adolescent populations. A list of the images used is provided in Supplemental Information 1. In combination with the negative or neutral primes, the following combinations of prime and trial condition were possible: negative-congruent (Neg_C), negative-stars (Neg_S), negative-incongruent (Neg_IC), and neutral-congruent (Neu_C), neutral-stars (Neu_S), neutral-incongruent (Neu_IC). Prior to study start, our task was behaviorally tested in adults. Pilot data assessment indicated a significant emotion by cognition interaction for reaction time and accuracy measurements (see Supplemental Information 1.2).

Figure 1. fMRI task design. Three exemplary emotional Stroop trials are displayed, depicting the following conditions (from top to bottom): Negative-congruent trial, neutral-incongruent trial, and negative-stars trial.
Prior to the start of the experiments, an optimal stochastic trial order allowing for a rapid event-related design was determined using optseq2, a tool for automatically scheduling events for rapid-presentation event-related fMRI experiments (for further information see http://surfer.nmr.mgh.harvard.edu/optseq/ or experiments using similar designs Ferri et al., 2012; Kuhlmann et al., 2016). We administered a total of 300 Stroop trials (100 for each C/S/IC) and 50 null trials. All 300 Stroop trials were preceded by either neutral or negative primes (50:50). Total scan time was about 11.5 min. The complete experiment was performed over the course of 2 runs. At the end of the neuroimaging session, participants were further asked to perform valence ratings of the images presented in the scanner using a Likert scale from −2 to 2 (with −2 representing very negative valence, 0 being neutral, and 2 indicating high attractiveness of the stimulus). For each participant the mean scores of the negative images affect rating were used as a covariate of no interest within the group analysis to account for differences in valence judgments between the young adults (see also Supplemental Information 2.1 Emotional Valence Rating).
Whole brain blood oxygen level-dependent (BOLD) fMRI data and T1-weighted mprage images were acquired on a Siemens 3T MR imaging system (Siemens Prisma, Erlangen, Germany) and a 20-channel phased-array radio frequency head coil. For the fMRI task a rapid event-related stochastic design with TR = 2,000 ms, TE = 30.0 ms, FOV = 192 mm; image matrix = 64 × 64 mm; voxel size = 3 mm and number of slices = 37 was used. We further acquired a high resolution T1-weighted structural image using the following specifications: TR = 1,900.0 ms; TE = 3.42 ms; FOV = 256; image matrix = 256 × 256; voxel size = 1 mm. T1-weighted mprage structural neuroimaging data was used for co-registration and to calculate the total intracranial volume (TIV). Men and women are known to differ in overall brain size (Leonard et al., 2008; Luders et al., 2009; Giedd et al., 2012). This was also true for the present sample where TIV significantly differed between males and females [males M = 1,529.52 ± 87.22/females M = 1,360.72 ± 91.99; t(28) = 5.16, p < 0.001]. Likewise, socioeconomic status, sex, and age all correlate with TIV (Luders et al., 2009, 2014; Taki et al., 2011; Jednorog et al., 2012). This was accounted for by using a covariate of no interest in consequent random effects analyses. Therefore, TIV was extracted through the voxel-based morphometry toolbox (VBM8; http://dbm.neuro.uni-jena.de/vbm) as implemented in SPM8 and executed in MATLAB (Mathworks, Natick, MA).
All functional MRI data was analyzed using SPM8 (http://www.fil.ion.ucl.ac.uk/spm/). Preprocessing included slice timing correction, realignment, co-registration to the structural images, segmentation of the structural image, normalization to the Montreal Neurologic Institute (MNI) standard brain, and smoothing using an 8 mm full-width at half maximum Gaussian kernel. During single subject analysis, the following regressors were built: Neg_C, Neg_S, Neg_IC, Neu_C, Neu_S, Neu_IC. Contrast images were created to investigate (1) the main effect of emotion (Neg>Neu trials), (2) the main effect of cognition (IC>C or IC>S trials), and (3) the influence of emotion on cognition along with increasing cognitive demand as based on two-sample t-tests comparing Neg_C vs. Neu_C, Neg_S vs. Neu_S, and Neg_IC vs. Neu_IC trials.
Due to the challenges in capturing the intricate nature of emotion–cognition interactions, the majority of publications in this field have based their interpretation on a priori based regions of interests only. Here, we present both small volume peak-level FWE-corrected findings at p < 0.05 for the main regions of interest [i.e., amygdala, insula, and/inferior frontal junction/precentral gyrus according to previous literature (Gray et al., 2002; Etkin et al., 2006; Blair et al., 2007; Hart et al., 2010; Gu et al., 2013); defined anatomically using the automated anatomical labeling atlas (Tzourio-Mazoyer et al., 2002)] and uncorrected, exploratory, whole brain findings (p < 0.001).
In order to further characterize the effects of cognitive load on the neural basis of emotion–cognition interactions, we further extracted mean peak activation scores as based on FWE-corrected findings from the two main contrasts targeting the emotion (Neg>Neu trials) and cognition (IC>S trials) networks of the brain. More specifically the signal change at the local peak activation scores for bilateral amygdala, right insula, and bilateral precentral gyrus were extracted using the marsbar toolbox (Brett et al., 2002) and further assessed using paired-samples t-tests.
In-scanner performance was assessed by computing the mean accuracy and reaction time in response to Neg_C, Neg_S, Neg_IC, Neu_C, Neu_S, and Neu_IC Stroop trials. For both accuracy and reaction times two separate 2 (emotion: Neg, Neu) by 3 (task: C, S, IC) repeated measures ANOVAs were performed in order to investigate the main effect of task, main effect of emotion, and the influence of emotion on task.
The 2 (emotion: Neg, Neu) by 3 (task: C, S, IC) ANOVA on task accuracy (i.e., correctly answered Stroop trials) indicated a significant main effect of emotion [F(1, 29) = 7.34, p = 0.011] and a main effect of cognition [F(2, 28) = 12.38, p < 0.0001]. Bonferroni corrected post-hoc tests indicated that the main effect on emotion was constituted by lower accuracy following negative primes (compared to neutral primes) during Stroop task. Furthermore, significant differences in accuracy derived from the incongruent condition compared to the congruent (p < 0.0001) and stars condition (p = 0.002). However, the difference between congruent and stars conditions did not reach significance (p = 1.00). Finally, the emotion by cognition interaction did not reach significance [F(2, 28) = 0.33, p = 0.722].
The 2 × 3 ANOVA implementing reaction time revealed a main effect of emotion [F(1, 29) = 11.93, p = 0.002] and a main effect of cognition [F(2, 28) = 80.27, p < 0.001]. Bonferroni corrected post-hoc tests revealed significant reaction time differences for incongruent compared to congruent (p < 0.0001), congruent compared to stars (p = 0.019), and incongruent compared to stars (p < 0.0001) conditions. The emotion by cognition interaction did not reach significance [F(2, 28) = 0.54, p = 0.590]. An overview of the in-scanner performance as based on the mean accuracy and reaction time for the whole group is given in Table 2.
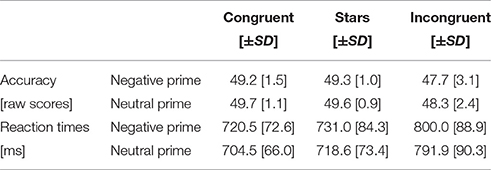
Table 2. In-scanner performance (accuracy, reaction times).
The fMRI result part is organized in line with our a priori listed main aims: (1) testing the activation of the emotion network by use of the negative prime (Neg>Neu trials); (2) assessing the activation of the cognition network by comparing an incongruent to a neutral Stroop condition (IC>S); (3) evaluating the emotion–cognition interaction dependent on increased cognitive load (Neg_C vs. Neu_C, Neg_S vs. Neu_S, and Neg_IC vs. Neu_IC trials). First, testing the (1) emotion processing network revealed that trials with a preceding negative prime compared to those with a preceding neutral prime led to significant increases in activation in known emotion processing areas of the brain (Phan et al., 2002), including insula, amygdala, and prefrontal cortices (for an overview of activated areas see Table 3, Figure 2). Secondly, testing the (2) cognition network by use of the Stroop task (IC>S) revealed activations in areas including left precentral gyrus (FWE-corrected) and uncorrected within further areas including the superior frontal brain regions, temporal cortex, and insula (Table 3, Figure 2).
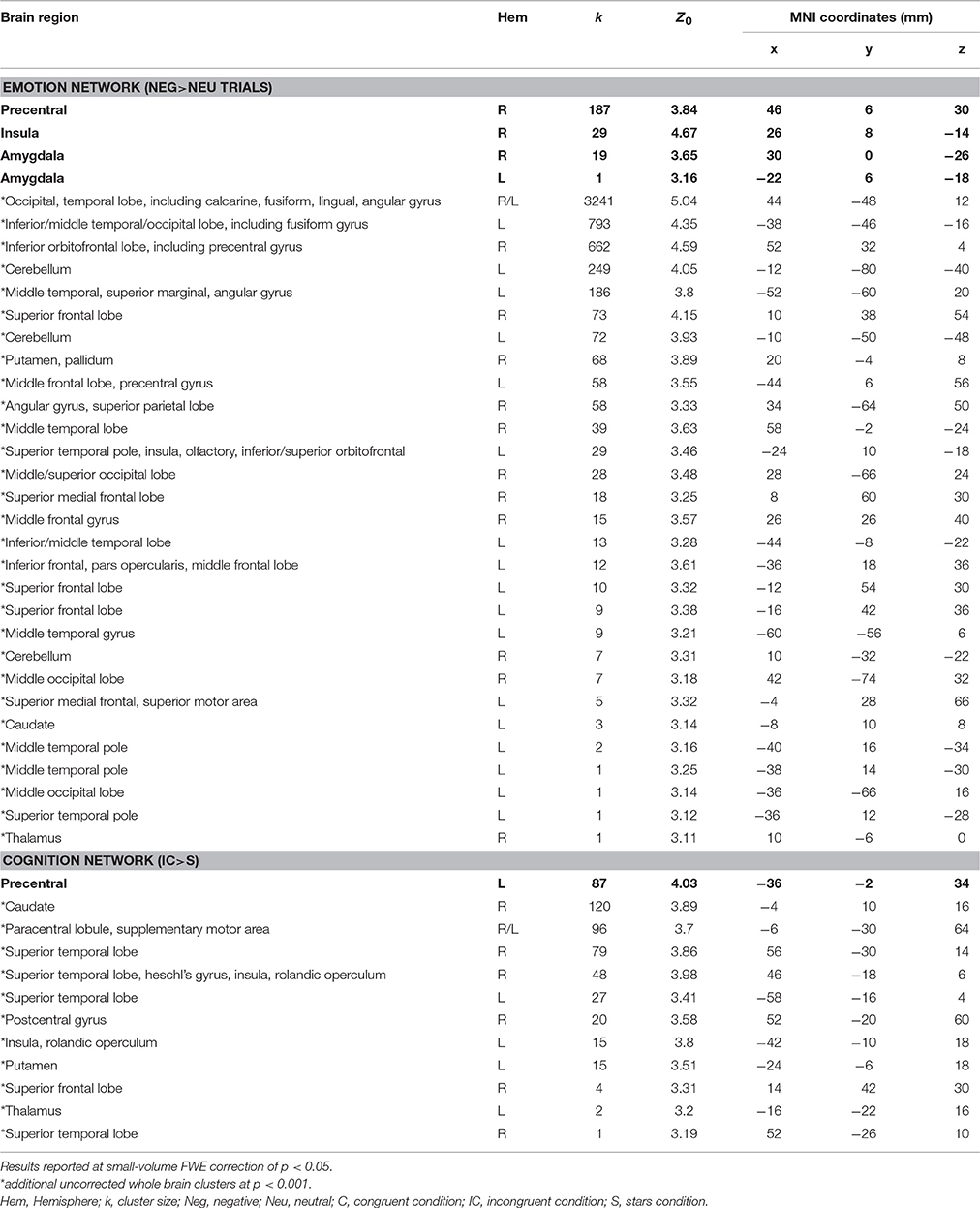
Table 3. MNI coordinates, cluster size, and Z-scores for significant FWE small-volume corrected results (indicated with bold letters) and uncorrected (p < 0.001; indicated with an asterix*) whole brain findings representing the emotion processing network (negative trials > neutral trials) and the cognition network (incongruent > stars trials) elicited by the given task.
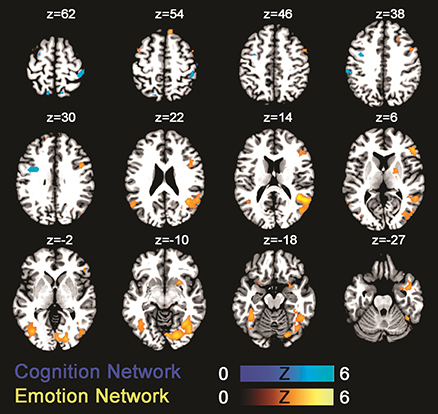
Figure 2. Statistical parametric maps showing brain activation linked to the emotion network (green-blue; negative > neutral trials) and the cognition network (gold-yellow; incongruent > congruent). Results are displayed at a p < 0.001, uncorrected threshold and neurologically displayed on axial slices using the Multi-image Analysis GUI as available at http://ric.uthscsa.edu/mango/mango.html.
Both control conditions (IC>S and IC>B) were contrasted with the IC condition. However, we decided to focus on IC>S trials for definition of the cognition network since more prefrontal activation was evoked, potentially due to different effects of the primes on congruent as opposed to stars trials. This procedure is in line with similar previous fMRI Stroop publications (for a review see Laird et al., 2005). Finally, (3) the influence of emotion on cognitive control was measured by contrasting Stroop trials with a prior negative prime to those Stroop trials following a neutral trial. Two-sample t-tests for negative vs. neutral trials were calculated for Stroop trials with increasing cognitive load (from congruent, to stars, and incongruent condition) and revealed differential activations for each of the three contrasts reflecting the influence of emotion on cognitive task performance as modulated by differential cognitive load. More specifically, for the contrast “Neg>Neu trials” during the congruent Stroop task condition, significant increases in activations were identified in bilateral amygdala, right insula, and right precentral gyrus (FWE-corrected) and on a whole-brain uncorrected level within further regions of the occipital, temporal, and inferior/middle frontal gyrus. For the opposite contrast “Neg < Neu trials” during the congruent Stroop trial, FWE-corrected activation within left precentral gyrus was observed. For the contrast “Neg>Neu trials” during the stars Stroop task condition, significant FWE-corrected findings were identified in right amygdala, while further cluster of activations were located within bilateral inferior occipital cortex (uncorrected whole brain approach). For the opposite contrast “Neg < Neu trials” significant activation was located within right precentral gyrus (FWE-corrected) and using a whole brain approach further clusters were located within occipital and superior frontal brain regions, left precuneus of the superior parietal lobe. Finally, for the contrast “Neg>Neu trials” during the incongruent Stroop task condition significant clusters of activations were based in inferior frontal and middle frontal brain regions (uncorrected findings only), while the opposite contrast of “Neg < Neu trials” during the incongruent Stroop task condition led to activity within left insula (FWE-corrected) as well as in further occipital brain regions, left pre- and postcentral gyrus, when using an uncorrected whole-brain approach (Table 4, Figure 3).
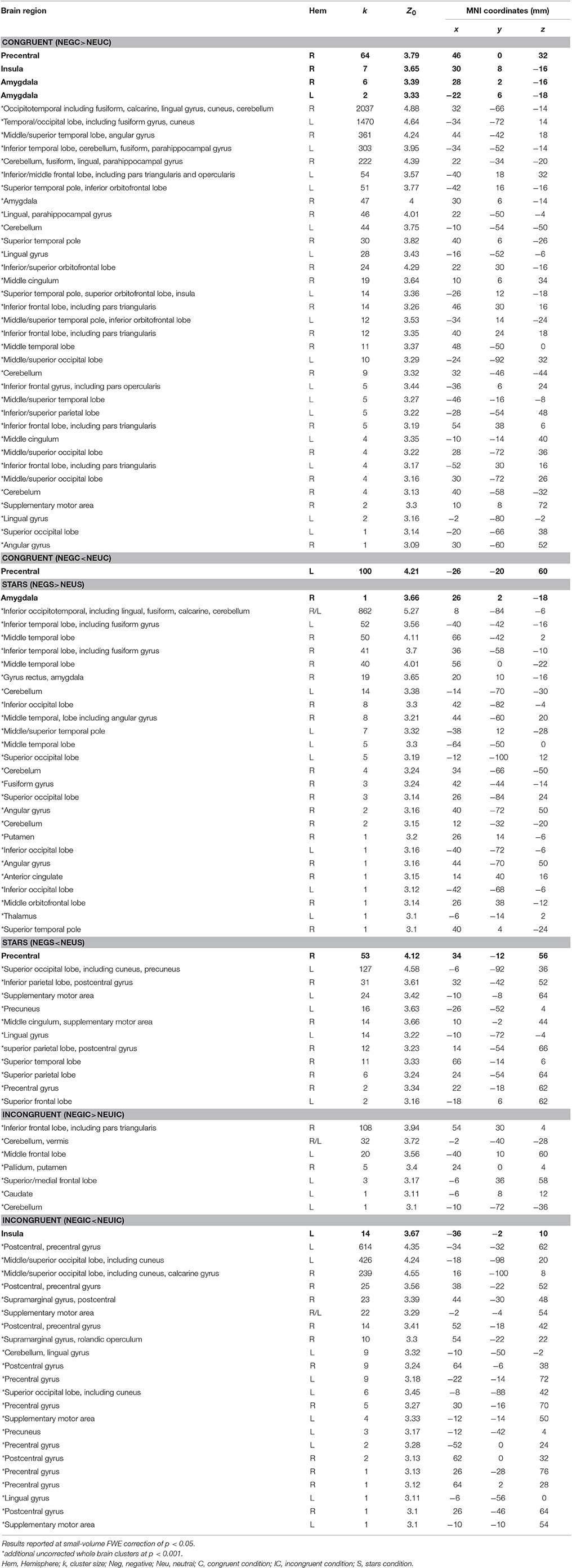
Table 4. MNI coordinates, cluster size, and Z-scores for significant FWE small-volume corrected results (indicated with bold letters) and uncorrected (p < 0.001; indicated with an asterix*) whole-brain findings representing the influence of emotion on cognitive processes dependent on cognitive load (i.e., congruent, stars, and incongruent trials).
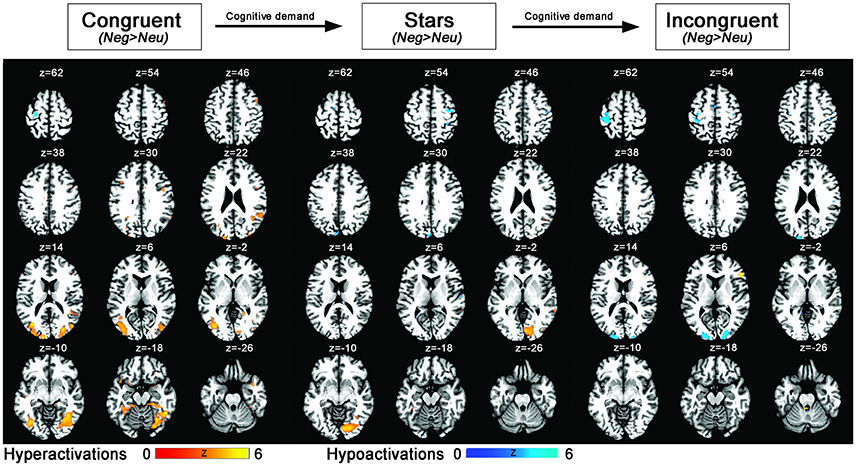
Figure 3. Statistical parametric maps showing emotion–cognition interaction related brain activation (blue: hypoactivations; red: hyperactivations) for negative > neutral emotional primes and the conditions congruent, stars, incongruent (ordered by lowest to highest cognitive load). Results are displayed at a p < 0.001, uncorrected threshold and neurologically displayed on axial slices using the Multi-image Analysis GUI as available at http://ric.uthscsa.edu/mango/mango.html.
Further investigations on the influence of emotion on cognition within peak regions of interests as based on FWE-corrected peak regions derived from the here identified emotion- (Neg>Neu trials) and cognition (IC>S) network revealed a significant trend within left amygdala, right insula and right precentral gyrus to show decreases of neural activation along for emotional primes with increasing cognitive demand. More specifically paired two samples t-tests indicated significant decreases of neural activation for (Neg>Neu) from congruent to incongruent condition within left amygdala, right insula, and right precentral gyrus (all p < 0.05). For the right insula the comparison between stars and incongruent condition likewise became significant (see Figure 4; additional graphics for the remaining regions that did not reach significance see Supplemental Information 2.2).
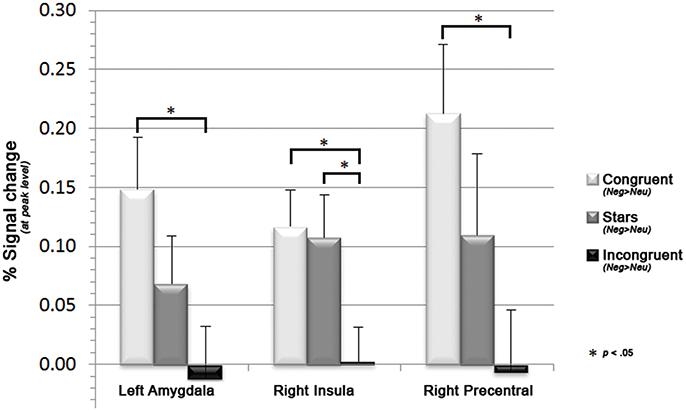
Figure 4. Bar graphs displaying decreases in the mean parameter estimates in left precentral gyrus and right amygdala along with increasing cognitive demand (i.e., for congruent, stars, and incongruent Stroop trials), as well as the associated sagittal brain slices including the statistical parametric maps (blue: hypoactivations; red: hyperactivations). Results are displayed at a p < 0.001, uncorrected threshold and neurologically displayed on axial slices using the Multi-image Analysis GUI as available at http://ric.uthscsa.edu/mango/mango.html.
In this study, we investigated the influence of emotions on cognition in each corresponding network through the use of negative or neutral primes prior to a number Stroop task with increasing levels of cognitive demand. Our main behavioral findings demonstrated increased reaction time and reduced Stroop-task accuracy following negative primes and/or increasing cognitive demand. Neurally, the emotional primes consistently activated emotion-related brain regions (including amygdala, insula, and prefrontal brain regions) while the Stroop effect was associated with activations in areas linked to cognitive processing (including left inferior frontal junction/precentral gyrus, inferior/superior parietal lobe, and insula). And finally, the neural correlates representing the influence of emotion on cognition implementing variations in cognitive demand lead to decreases in neural activation in response to emotional stimuli (negative>neutral) along with increased cognitive demand within prefrontal cortex, amygdala, and insular cortex. Additionally, in trials with no preceding emotional prime (neutral>negative), significant increases along with increasing cognitive demand where observed. Overall we conclude that neural activation during a number Stroop task performance is increasingly disrupted by preceding negative images along with increasing cognitive demand.
In line with previous work, negative compared to neutral primes prior to a Stroop task affected task performance, which resulted in decreased task accuracy and increased reaction times (Gray et al., 2002; Mitchell et al., 2006; Blair et al., 2007; Padmala et al., 2011). Furthermore, more incorrect answers were given during incongruent trials compared to congruent (Etkin et al., 2006) or stars trials, while the latter two showed similar accuracy measurements. Participants were fastest in responding to congruent trials, followed by stars trials and eventually incongruent trials, which reflects the so-called Stroop effect (Stroop, 1935; Blair et al., 2007; Hart et al., 2010). We propose that in this study reading the number was a more automatized/faster process compared to counting the actual stimuli, thus resulting in shorter reaction times for congruent compared to stars trials. Interestingly, congruent and stars trials had equivalent numbers of correctly answered trials, although participants responded faster to congruent than to stars trials. We therefore conclude that the behavioral advantage of the congruent condition only affected reaction time (increased speed), but not accuracy. However, behavioral analysis failed to observe an interaction effect between emotional priming and task difficulty in accuracy and reaction times. In line with the dual competition model by Pessoa (2009) and supported by previous studies (Hart et al., 2010; Melcher et al., 2011; Padmala et al., 2011) the interference of negative primes on task performance was expected to augment with higher cognitive task load due to competing mechanisms. However, in line with previous evidence (e.g., Blair et al., 2007; Van Dillen et al., 2009), we did not observe a significant interaction effect in the present analysis.
Here, we demonstrate that the influence of emotion on cognition is neurally reflected within brain regions including prefrontal cortex, amygdala, and insular cortex. Furthermore, activation in these brain regions shows an attenuating trend when increasing cognitive demand. Likewise, neural activation in the left inferior frontal junction/precentral gyrus, as indicative of cognitive control, is increased during neutral prime trials compared to trials following a negative prime. Prefrontal brain regions, amygdala, and insula have all consistently been identified as relevant for emotion–cognition interactions or emotional conflict resolution (Gray et al., 2002; Beer et al., 2006; Etkin et al., 2006; Blair et al., 2007; Hart et al., 2010; Gu et al., 2013; Buhle et al., 2014). Likewise, similar areas are activated during tasks requiring cognitive reappraisal (an emotion regulation strategy in which the stimulus meaning is reinterpreted to downregulate the emotional valence; Ochsner et al., 2002, 2004; Ochsner and Gross, 2005). Thus, we suggest that the here presented emotional number Stroop task activates similar areas within the neural network that are required for deliberate cognitive reappraisal.
The prefrontal cortex can functionally and cytoarchitectonically be subdivided into distinct sub-regions, several of which are of relevance to affective processing, cognition, or both (Pessoa, 2008). Overall, prefrontal brain regions are commonly linked to attention, working memory, goal-directed behavior (e.g., cognitive control or decision making (Pessoa, 2008; Stokes et al., 2013; Leech and Sharp, 2014), and affective processing (Phan et al., 2002). From an evolutionary perspective, early research has suggested that the evolution of the human prefrontal cortex, particularly its expansion in volume, may reflect the development of more complex social behavior (Dunbar and Shultz, 2007). While such an interpretation may be too simplistic, still researchers commonly agree that the distinct parts of the prefrontal cortex are recruited by different high-level cognitive demands (Eickhoff et al., 2016). Based on animal and human studies, a functional and cytoarchitectonical subdivision of the medial prefrontal cortex may at least result in areas including the orbitofrontal cortex (BA11), ventral prefrontal cortex and prefrontal pole (BA10), and dorsomedial prefrontal cortex and frontal pole (BA9; Eickhoff et al., 2016). All of these areas are strongly interconnected and associated with various other circuitries of the brain, including the limbic network (Reid et al., 2016). Finally, particularly the right inferior frontal gyrus has been suggested to be a crucial hub during inhibitory processing and may consequently be an area affected in response control disorders (Aron et al., 2014).
Emotion processing is generally assigned to medial prefrontal brain regions, whereas the amygdala, anterior cingulate cortex, or insula are thought to possess a more distinct function within emotional tasks (Phan et al., 2002). The amygdala is one of the most traditionally viewed emotion and motivation processing center. With its relatively small structure, the amygdala nevertheless comprises a multitude of anatomical connections allowing many intricate functionalities (Janak and Tye, 2015). While abundant research has linked the amygdala to affective processing and particularly fear conditioning, strong evidence points toward a more integral role of the amygdala as a key node for valence processing during different aversive states, including fear, anxiety, or reward processing (Murray et al., 2014; Janak and Tye, 2015). Here we demonstrated that the amount of amygdala activation obtained during emotion–cognition interaction was highest during Stroop trials with lowest cognitive demand and decreases with increasing task difficulty. This finding is in line with Etkin et al. (2006) as well as Blair et al. (2007) that found decreases in neural activation within the amygdala during concurrent task with increasing cognitive demand. In line with previous suggestions (Etkin et al., 2006), it could be concluded that amygdala activation mirrors the amount of emotional conflict, rather than the resolution of such. Importantly the amygdala is strongly interconnected with areas of the prefrontal/orbitofrontal cortex (Davidson et al., 2000). This bi-directional connection allows regulatory processes important for mental well-being. For example, early deprivation or life stress may lead to disruption in amygdala-prefrontal coupling and patients with symptoms including anxiety, posttraumatic stress disorder, or heightened aggression oftentimes show structural and functional impairments within these circuitries (Gee et al., 2013).
The present task closely resembles two prior fMRI study designs (Blair et al., 2007; Hart et al., 2010). With the exception of adaptations that account for the age of the participants being tested (i.e., use of only negative stimuli and age-appropriate images), it may be considered a replication study. The present manuscript used an adapted version of an affective number Stroop task as implemented by Hart et al. (2010) and resulted in comparable findings. Reproducibility of scientific studies is crucial in order to inform about the robustness of an observed phenomenon (Martin and Clarke, 2017). Comparing our findings more closely to these two prior studies confirm the following main findings: (1) In line with both studies, negative primes slowed the participants' reaction time during the number Stroop task and are thus confirmed to interrupt goal-directed processing; (2) In line with Blair et al. (2007) increasing cognitive demand led to decreases in emotion-related brain regions (e.g., amygdala, insula, prefrontal cortex). Hart et al. (2010), observed the same trend however, no neural decreases in dorsolateral prefrontal areas during incongruent trials. Therefore, the authors concluded that high cognitive demand may override the attenuation effect in the prefrontal cortex. In contrast to Hart et al. (2010), we only observed decreases in neural activation with increasing cognitive demand. It remains to be investigated whether such a difference may be due to the number and characteristics of participants tested (N = 14; 5 males in Hart et al. (2010)/N = 30; 15 males in the present study) or are potentially due to the difference in stimuli choice and/or slightly longer presentation (an additional 500 ms) of the Stroop trial in our study. It is important to note that the slightly longer Stroop presentation rate was chosen due to the aim of consequently applying this task in younger participants.
The insula is a functionally heterogeneous brain region which is situated in the depth of the Sylvian fissure and may be divided into three sections: a dorsal anterior, a ventral anterior, and a posterior part (Nieuwenhuys, 2012; Uddin et al., 2014). The anterior insular cortex is, mostly bilaterally, connected to limbic and prefrontal brain regions (e.g., the amygdala), while the posterior part is more strongly interconnected with parietal, occipital and temporal parts of the brain (Kurth et al., 2010; Nieuwenhuys, 2012). The insula has shown to be activated during a wide range of functions, including auditory processing, vestibular and somatosensory functions, the perception of pain and temperature, viscerosensation, taste, olfactory processing, somatomotor control and motor plasticity, speech production, cognitive control, bodily awareness, as well as emotion processing (Nieuwenhuys, 2012). Importantly, according to research the anterior insula has a critical role during the regulation of social behavior, since its structure and function is altered in individuals with social disorders (including disruptive behavior disorder Sterzer et al., 2007; Raschle et al., 2015). Here our results are in line with findings assigning a critical role for the insula in emotion–cognition interactions (Hart et al., 2010; Shackman et al., 2011; Gu et al., 2013).
An intact integration and healthy balance of competing emotion-cognition processing is crucial for our everyday functioning and an imbalance, as for example observed in individuals with emotion processing deficits, is linked to different mental health disorders (Monk, 2008). For example, faulty integration or regulation of emotion-cognition processes may result in heightened violence and aggression (Davidson et al., 2000). Therefore, it has been suggested that individuals with heightened aggression traits, as for example observed in children and adolescents with disruptive behavior disorders, may show impairments in these prefrontal circuitries responsible for successful emotion–cognition interaction and regulation. In fact, various structural and functional neuroimaging studies pinpoint areas of the limbic and prefrontal network to be disrupted in aggressive individuals (Raschle et al., 2015; Rogers and De Brito, 2016). Consequently, we conclude that future studies may implement the here presented design in order to further characterize aggressive youths and potentially impact individualized classification and treatment approaches in health and disease.
A potential caveat in the design of this study concerns the valence of the primes used. While we employed negative and neutral images only, positive primes have also been shown to disrupt task performance (Mitchell et al., 2006; Blair et al., 2007). Therefore, the addition of positive images should be considered in future studies aiming at characterizing the emotion–cognition interaction. However, due to practical challenges when conducting pediatric neuroimaging studies (particularly time constraints; for a discussion see for example Raschle et al., 2009) we decided that it is of importance to keep the task as short as possible and gain maximum power for the emotional condition chosen. Additionally, since the processing of negative affect is a particular problem in disruptive behavior disorders, a focus on this emotion made most sense. Secondly, we here used DAPS images as primes. This system was developed as an adaptation of the International Affective Picture System (IAPS, Lang et al., 2008) in order to be suitable for children and adolescents. The DAPS includes images from the IAPS series as well as additional stimulus material. We here only used images that were part of the IAPS and DAPS system, which can thus be considered suitable for evoking negative affect in both populations. However, IAPS images with the strongest negative affect were excluded within this process. Therefore, the images implemented here may have had a reduced impact on the young adults performing our task. Our decision to employ a child-friendly image system is due to our aim of testing our task for future use in clinical populations involving children and adolescents with disruptive behavior disorders. While effects of negative priming were observable in the young adults sample investigated here, we believe the same images may lead to stronger behavioral and neural effects in younger participants as investigated in the future. Finally, while we implemented an automatic stochastic schedule for optimal event presentation and null trials for jittering, the inter-trial intervals may gain from additional variance (i.e., variations in inter-trial intervals around for example 500–1,500 ms).
Converging evidence points toward the importance of a balanced handling of both emotional and cognitive information in our everyday life. Here we present data that validates the usefulness of the emotional number Stroop task in fMRI settings aiming to assess the neural correlates of the influence of emotion on cognition. More specifically, we show an impact in behavior and the associated neural networks depending on emotional prime and cognitive demand. The respective influence of the emotion and cognition network in the brain may therefore be seen as a dynamic process which is modulated by the executive resources available. Moreover, emotion and cognition seem to be tightly related to each other, as indicated by shared neural networks involved in both of these processes. A failure to successfully integrate emotional and cognitive demands is characteristic to many psychiatric disorders. Future studies may thus further investigate the neural characteristics of children and adolescents that fail to successfully process emotional/cognitive demand, as for example seen in disruptive behavior disorders.
This study was carried out in accordance with the recommendations of the Ethikkommission der Nordwest- und Zentralschweiz. All subjects gave written informed consent in accordance with the Declaration of Helsinki. The protocol was approved by the Ethikkommission der Nordwest- und Zentralschweiz.
Conception and design of the experiments: CS, PS, NR, FE, and WM. Data collection: LF, WM, and NR. Data analysis and interpretation: NR, CS, PS, LF, and WM. Drafting the paper: NR, CS, LF, and WM. Revision and final approval of the version to be published: NR, CS, PS, LF, WM, and FE.
This study was supported by a grant from the Swiss National Science Foundation (SNSF) [105314M_150282]. NR is a recipient of the 2017–2019 early career award of the Jacobs Center and received funding through the University Hospital Basel grant (Nachwuchsförderung).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank the families that have participated in this study. Parts of the here presented findings were also shown at: Fehlbaum et al. (2014, 2015, 2016) and Menks et al. (2015).
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpsyg.2017.01489/full#supplementary-material
Achenbach, T. M. (1991). Manual for the Youth Self-Report and 1991 Profile. Burlington, VT: University of Vermont Department of Psychiatry.
Andershed, H., Hodgins, S., and Tengström, A. (2007). Convergent validity of the Youth Psychopathic Traits Inventory (YPI): association with the Psychopathy Checklist: Youth Version (PCL:YV). Assessment 14, 144–154. doi: 10.1177/1073191106298286
Arnsten, A. F., and Goldman-Rakic, P. S. (1998). Noise stress impairs prefrontal cortical cognitive function in monkeys: evidence for a hyperdopaminergic mechanism. Arch. Gen. Psychiatry 55, 362–368. doi: 10.1001/archpsyc.55.4.362
Aron, A. R., Robbins, T. W., and Poldrack, R. A. (2014). Inhibition and the right inferior frontal cortex: one decade on. Trends Cogn. Sci. 18, 177–185. doi: 10.1016/j.tics.2013.12.003
Beer, J. S., John, O. P., Scabini, D., and Knight, R. T. (2006). Orbitofrontal cortex and social behavior: integrating self-monitoring and emotion-cognition interactions. J. Cogn. Neurosci. 18, 871–879. doi: 10.1162/jocn.2006.18.6.871
Blair, K. S., Smith, B. W., Mitchell, D. G., Morton, J., Vythilingam, M., Pessoa, L., et al. (2007). Modulation of emotion by cognition and cognition by emotion. Neuroimage 35, 430–440. doi: 10.1016/j.neuroimage.2006.11.048
Brett, M., Anton, J. L., Valabregue, R., and Poline, J.-B. (2002). “Region of interest analysis using the MarsBar toolbox for SPM 99,” in Functional Mapping of the Human Brain (Sendai: Neuroimage).
Buhle, J. T., Silvers, J. A., Wager, T. D., Lopez, R., Onyemekwu, C., Kober, H., et al. (2014). Cognitive reappraisal of emotion: a meta-analysis of human neuroimaging studies. Cereb. Cortex 24, 2981–2990. doi: 10.1093/cercor/bht154
Caplan, B., and Mendoza, J. E. (2011). “Edinburgh handedness inventory,” in Encyclopedia of Clinical Neuropsychology, eds J. S. Kreutzer, J. DeLuca, and B. Caplan (New York, NY: Springer), 928.
Cohen, N., and Henik, A. (2012). Do irrelevant emotional stimuli impair or improve executive control? Front. Integr. Neurosci. 6:33. doi: 10.3389/fnint.2012.00033
Cordon, I. M., Melinder, A. M., Goodman, G. S., and Edelstein, R. S. (2013). Children's and adults' memory for emotional pictures: examining age-related patterns using the Developmental Affective Photo System. J. Exp. Child Psychol. 114, 339–356. doi: 10.1016/j.jecp.2012.08.004
Cromheeke, S., and Mueller, S. C. (2014). Probing emotional influences on cognitive control: an ALE meta-analysis of cognition emotion interactions. Brain Struct. Funct. 219, 995–1008. doi: 10.1007/s00429-013-0549-z
Davidson, R. J., Putnam, K. M., and Larson, C. L. (2000). Dysfunction in the neural circuitry of emotion regulation - a possible prelude to violence. Science 289, 591–594. doi: 10.1126/science.289.5479.591
Dolan, R. J. (2002). Emotion, cognition, and behavior. Science 298, 1191–1194. doi: 10.1126/science.1076358
Dolcos, F., and McCarthy, G. (2006). Brain systems mediating cognitive interference by emotional distraction. J. Neurosci. 26, 2072–2079. doi: 10.1523/JNEUROSCI.5042-05.2006
Dunbar, R. I. M., and Shultz, S. (2007). Evolution in the social brain. Science 317, 1344–1347. doi: 10.1126/science.1145463
Eickhoff, S. B., Laird, A. R., Fox, P. T., Bzdok, D., and Hensel, L. (2016). Functional segregation of the human dorsomedial prefrontal cortex. Cereb. Cortex 26, 304–321. doi: 10.1093/cercor/bhu250
Etkin, A., Egner, T., Peraza, D. M., Kandel, E. R., and Hirsch, J. (2006). Resolving emotional conflict: a role for the rostral anterior cingulate cortex in modulating activity in the amygdala. Neuron 51, 871–882. doi: 10.1016/j.neuron.2006.07.029
Euler, F., Sterzer, P., and Stadler, C. (2014). Cognitive control under distressing emotional stimulation in adolescents with conduct disorder. Aggress. Behav. 40, 109–119. doi: 10.1002/ab.21508
Fehlbaum, L. V., Raschle, N. M., Menks, W. M., Euler, F., and Stadler, C. (2014). “Emotion processing and emotion regulation in youths with empathy deficits,” in Poster Abstract Accepted to the 1st Computational Psychiatry Meeting (Zurich).
Fehlbaum, L. V., Raschle, N. M., Menks, W. M., Euler, F., and Stadler, C. (2015). “Emotion processing and emotion regulation in youths,” in Poster Abstract Accepted to the Clinical Research Day of University Hospital Basel (Basel).
Fehlbaum, L. V., Raschle, N. M., Menks, W. M., Euler, F., and Stadler, C. (2016). “Neuronal basis of emotion and cognitive processing in young adults and adolescents,” in Poster Abstract Accepted to the Cognitive Neuroscience 2016 Annual Meeting (New York, NY).
Ferri, F., Frassinetti, F., Ardizzi, M., Costantini, M., and Gallese, V. (2012). A sensorimotor network for the bodily self. J. Cogn. Neurosci. 24, 1584–1595. doi: 10.1162/jocn_a_00230
Gee, D. G., Gabard-Durnam, L. J., Flannery, J., Goff, B., Humphreys, K. L., Telzer, E. H., et al. (2013). Early developmental emergence of human amygdala-prefrontal connectivity after maternal deprivation. Proc. Natl. Acad. Sci. U.S.A. 110, 15638–15643. doi: 10.1073/pnas.1307893110
Giedd, J. N., Raznahan, A., Mills, K. L., and Lenroot, R. K. (2012). Review: magnetic resonance imaging of male/female differences in human adolescent brain anatomy. Biol. Sex Differ. 3:19. doi: 10.1186/2042-6410-3-19
Gray, J. R., Braver, T. S., and Raichle, M. E. (2002). Integration of emotion and cognition in the lateral prefrontal cortex. Proc. Natl. Acad. Sci. U.S.A. 99, 4115–4120. doi: 10.1073/pnas.062381899
Gross, J. J. (2002). Emotion regulation: affective, cognitive, and social consequences. Psychophysiology 39, 281–291. doi: 10.1017/S0048577201393198
Gu, X., Liu, X., Van Dam, N. T., Hof, P. R., and Fan, J. (2013). Cognition-emotion integration in the anterior insular cortex. Cereb. Cortex 23, 20–27. doi: 10.1093/cercor/bhr367
Gupta, R., and Deak, G. O. (2015). Disarming smiles: irrelevant happy faces slow post-error responses. Cogn. Process. 16, 427–434. doi: 10.1007/s10339-015-0664-2
Gupta, R., Hur, Y. J., and Lavie, N. (2016). Distracted by pleasure: effects of positive vs. negative valence on emotional capture under load. Emotion 16, 328–337. doi: 10.1037/emo0000112
Gupta, R., and Raymond, J. E. (2012). Emotional distraction unbalances visual processing. Psychon. Bull. Rev. 19, 184–189. doi: 10.3758/s13423-011-0210-x
Hart, S. J., Green, S. R., Casp, M., and Belger, A. (2010). Emotional priming effects during Stroop task performance. Neuroimage 49, 2662–2670. doi: 10.1016/j.neuroimage.2009.10.076
Hartikainen, K. M., Ogawa, K. H., and Knight, R. T. (2000). Transient interference of right hemispheric function due to automatic emotional processing. Neuropsychologia 38, 1576–1580. doi: 10.1016/S0028-3932(00)00072-5
Homack, S., and Riccio, C. A. (2004). A meta-analysis of the sensitivity and specificity of the stroop color and word test with children. Arch. Clin. Neuropsychol. 19, 725–743. doi: 10.1016/j.acn.2003.09.003
Janak, P. H., and Tye, K. M. (2015). From circuits to behaviour in the amygdala. Nature 517, 284–292. doi: 10.1038/nature14188
Jednorog, K., Altarelli, I., Monzalvo, K., Fluss, J., Dubois, J., Billard, C., et al. (2012). The influence of socioeconomic status on children's brain structure. PLoS ONE 7:e42486. doi: 10.1371/journal.pone.0042486
John, O. P., and Gross, J. J. (2004). Healthy and unhealthy emotion regulation: personality processes, individual differences, and life span development. J. Pers. 72, 1301–1333. doi: 10.1111/j.1467-6494.2004.00298.x
Kanske, P. (2012). On the influence of emotion on conflict processing. Front. Integr. Neurosci. 6:42. doi: 10.3389/fnint.2012.00042
Kanske, P., and Kotz, S. A. (2011). Emotion speeds up conflict resolution: a new role for the ventral anterior cingulate cortex? Cereb. Cortex 21, 911–919. doi: 10.1093/cercor/bhq157
Kellermann, T. S., Sternkopf, M. A., Schneider, F., Habel, U., Turetsky, B. I., Zilles, K., et al. (2012). Modulating the processing of emotional stimuli by cognitive demand. Soc. Cogn. Affect. Neurosci. 7, 263–273. doi: 10.1093/scan/nsq104
Kuhlmann, M., Hofmann, M. J., Briesemeister, B. B., and Jacobs, A. M. (2016). Mixing positive and negative valence: affective-semantic integration of bivalent words. Sci. Rep. 6:30718. doi: 10.1038/srep30718
Kurth, F., Zilles, K., Fox, P. T., Laird, A. R., and Eickhoff, S. B. (2010). A link between the systems: functional differentiation and integration within the human insula revealed by meta-analysis. Brain Struct. Funct. 214, 519–534. doi: 10.1007/s00429-010-0255-z
Laird, A. R., McMillan, K. M., Lancaster, J. L., Kochunov, P., Turkeltaub, P. E., Pardo, J. V., et al. (2005). A comparison of label-based review and ALE meta-analysis in the stroop task. Hum. Brain Mapp. 25, 6–21. doi: 10.1002/hbm.20129
Lang, P. J., Bradley, M. M., and Cuthbert, B. N. (2008). International Affective Picture System (IAPS): Affective Ratings of Pictures and Instruction Manual. Technical Report A-8. University of Florida, Gainesville, FL.
Leech, R., and Sharp, D. J. (2014). The role of the posterior cingulate cortex in cognition and disease. Brain 137(Pt 1), 12–32. doi: 10.1093/brain/awt162
Leonard, C. M., Towler, S., Welcome, S., Halderman, L. K., Otto, R., Eckert, M. A., et al. (2008). Size matters: cerebral volume influences sex differences in neuroanatomy. Cereb. Cortex 18, 2920–2931. doi: 10.1093/cercor/bhn052
Livingstone, K., Harper, S., and Gillanders, D. (2009). An exploration of emotion regulation in psychosis. Clin. Psychol. Psychother. 16, 418–430. doi: 10.1002/cpp.635
Luders, E., Gaser, C., Narr, K. L., and Toga, A. W. (2009). Why sex matters: brain size independent differences in gray matter distributions between men and women. J. Neurosci. 29, 14265–14270. doi: 10.1523/JNEUROSCI.2261-09.2009
Luders, E., Toga, A. W., and Thompson, P. M. (2014). Why size matters: differences in brain volume account for apparent sex differences in callosal anatomy The sexual dimorphism of the corpus callosum. Neuroimage 84, 820–824. doi: 10.1016/j.neuroimage.2013.09.040
Martin, G. N., and Clarke, R. M. (2017). Are psychology journals anti-replication? A snapshot of editorial practices. Front. Psychol. 8:523. doi: 10.3389/fpsyg.2017.00523
Melcher, T., Born, C., and Gruber, O. (2011). How negative affect influences neural control processes underlying the resolution of cognitive interference: an event-related fMRI study. Neurosci. Res. 70, 415–427. doi: 10.1016/j.neures.2011.05.007
Menks, W. M., Raschle, N. M., Fehlbaum, L. V., Euler, F., and Stadler, C. (2015). “Investigating neuronal correlates of emotion regulation in young adults,” in Poster Abstract Accepted to the 16th International ESCAP Congress (Madrid).
Mitchell, D. G., Richell, R. A., Leonard, A., and Blair, R. J. (2006). Emotion at the expense of cognition: psychopathic individuals outperform controls on an operant response task. J. Abnorm. Psychol. 115, 559–566. doi: 10.1037/0021-843X.115.3.559
Monk, C. S. (2008). The development of emotion-related neural circuitry in health and psychopathology. Dev. Psychopathol. 20, 1231–1250. doi: 10.1017/S095457940800059X
Murray, R. J., Brosch, T., and Sander, D. (2014). The functional profile of the human amygdala in affective processing: insights from intracranial recordings. Cortex 60, 10–33. doi: 10.1016/j.cortex.2014.06.010
Nieuwenhuys, R. (2012). The insular cortex: a review. Prog. Brain Res. 195, 123–163. doi: 10.1016/B978-0-444-53860-4.00007-6
Ochsner, K. N., Bunge, S. A., Gross, J. J., and Gabrieli, J. D. (2002). Rethinking feelings: an FMRI study of the cognitive regulation of emotion. J. Cogn. Neurosci. 14, 1215–1229. doi: 10.1162/089892902760807212
Ochsner, K. N., and Gross, J. J. (2005). The cognitive control of emotion. Trends Cogn. Sci. 9, 242–249. doi: 10.1016/j.tics.2005.03.010
Ochsner, K. N., Ray, R. D., Cooper, J. C., Robertson, E. R., Chopra, S., Gabrieli, J. D., et al. (2004). For better or for worse: neural systems supporting the cognitive down- and up-regulation of negative emotion. Neuroimage 23, 483–499. doi: 10.1016/j.neuroimage.2004.06.030
Okon-Singer, H., Lichtenstein-Vidne, L., and Cohen, N. (2013). Dynamic modulation of emotional processing. Biol. Psychol. 92, 480–491. doi: 10.1016/j.biopsycho.2012.05.010
Padmala, S., Bauer, A., and Pessoa, L. (2011). Negative emotion impairs conflict-driven executive control. Front. Psychol. 2:192. doi: 10.3389/fpsyg.2011.00192
Pessoa, L. (2008). On the relationship between emotion and cognition. Nat. Rev. Neurosci. 9, 148–158. doi: 10.1038/nrn2317
Pessoa, L. (2009). How do emotion and motivation direct executive control? Trends Cogn. Sci. 13, 160–166. doi: 10.1016/j.tics.2009.01.006
Petermann, F. E. (2012). Wechsler Adult Intelligence Scale - Fourth Edition (WAIS-IV). Deutschsprachige Adaptation der WAIS-IV von D. Wechsler. [German Adaptation of the WAIS-IV by D. Wechsler]. Frankfurt: Pearson Assessment.
Phan, K. L., Wager, T., Taylor, S. F., and Liberzon, I. (2002). Functional neuroanatomy of emotion: a meta-analysis of emotion activation studies in PET and fMRI. Neuroimage 16, 331–348. doi: 10.1006/nimg.2002.1087
Pitskel, N. B., Bolling, D. Z., Kaiser, M. D., Crowley, M. J., and Pelphrey, K. A. (2011). How grossed out are you? The neural bases of emotion regulation from childhood to adolescence. Dev. Cogn. Neurosci. 1, 324–337. doi: 10.1016/j.dcn.2011.03.004
Raschle, N. M., Lee, M., Buechler, R., Christodoulou, J. A., Chang, M., Vakil, M., et al. (2009). Making MR imaging child's play - pediatric neuroimaging protocol, guidelines and procedure. J. Vis. Exp. 1309. doi: 10.3791/1309
Raschle, N. M., Menks, W. M., Fehlbaum, L. V., Tshomba, E., and Stadler, C. (2015). Structural and functional alterations in right dorsomedial prefrontal and left insular cortex co-localize in adolescents with aggressive behaviour: an ALE meta-analysis. PLoS ONE 10:e0136553. doi: 10.1371/journal.pone.0136553
Reid, A. T., Bzdok, D., Langner, R., Fox, P. T., Laird, A. R., Amunts, K., et al. (2016). Multimodal connectivity mapping of the human left anterior and posterior lateral prefrontal cortex. Brain Struct. Funct. 221, 2589–2605. doi: 10.1007/s00429-015-1060-5
Rogers, J. C., and De Brito, S. A. (2016). Cortical and subcortical gray matter volume in youths with conduct problems: a meta-analysis. JAMA Psychiatry 73, 64–72. doi: 10.1001/jamapsychiatry.2015.2423
Schupp, H. T., Stockburger, J., Codispoti, M., Junghofer, M., Weike, A. I., and Hamm, A. O. (2007). Selective visual attention to emotion. J. Neurosci. 27, 1082–1089. doi: 10.1523/JNEUROSCI.3223-06.2007
Shackman, A. J., Salomons, T. V., Slagter, H. A., Fox, A. S., Winter, J. J., and Davidson, R. J. (2011). The integration of negative affect, pain and cognitive control in the cingulate cortex. Nat. Rev. Neurosci. 12, 154–167. doi: 10.1038/nrn2994
Sterzer, P., Stadler, C., Krebs, A., Kleinschmidt, A., and Poustka, F. (2005). Abnormal neural responses to emotional visual stimuli in adolescents with conduct disorder. Biol. Psychiatry 57, 7–15. doi: 10.1016/j.biopsych.2004.10.008
Sterzer, P., Stadler, C., Poustka, F., and Kleinschmidt, A. (2007). A structural neural deficit in adolescents with conduct disorder and its association with lack of empathy. Neuroimage 37, 335–342. doi: 10.1016/j.neuroimage.2007.04.043
Stokes, M. G., Kusunoki, M., Sigala, N., Nili, H., Gaffan, D., and Duncan, J. (2013). Dynamic coding for cognitive control in prefrontal cortex. Neuron 78, 364–375. doi: 10.1016/j.neuron.2013.01.039
Stroop, J. R. (1935). Studies of interference in serial verbal reactions. J. Exp. Psychol. 18, 643–662. doi: 10.1037/h0054651
Taki, Y., Thyreau, B., Kinomura, S., Sato, K., Goto, R., Kawashima, R., et al. (2011). Correlations among brain gray matter volumes, age, gender, and hemisphere in healthy individuals. PLoS ONE 6:e22734. doi: 10.1371/journal.pone.0022734
Thompson, L. A., Malloy, D. M., Cone, J. M., and Hendrickson, D. L. (2010). The face-to-face light detection paradigm: a new methodology for investigating visuospatial attention across different face regions in live face-to-face communication settings. Interact. Stud. 11, 336–348. doi: 10.1075/is.11.2.22tho
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F., Etard, O., Delcroix, N., et al. (2002). Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. Neuroimage 15, 273–289. doi: 10.1006/nimg.2001.0978
Uddin, L. Q., Kinnison, J., Pessoa, L., and Anderson, M. L. (2014). Beyond the tripartite cognition-emotion-interoception model of the human insular cortex. J. Cogn. Neurosci. 26, 16–27. doi: 10.1162/jocn_a_00462
Uher, R., Brooks, S. J., Bartholdy, S., Tchanturia, K., and Campbell, I. C. (2014). Increasing cognitive load reduces interference from masked appetitive and aversive but not neutral stimuli. PLoS ONE 9:e94417. doi: 10.1371/journal.pone.0094417
Van Dillen, L. F., Heslenfeld, D. J., and Koole, S. L. (2009). Tuning down the emotional brain: an fMRI study of the effects of cognitive load on the processing of affective images. Neuroimage 45, 1212–1219. doi: 10.1016/j.neuroimage.2009.01.016
Walcott, C. M., and Landau, S. (2004). The relation between disinhibition and emotion regulation in boys with attention deficit hyperactivity disorder. J. Clin. Child Adolesc. Psychol. 33, 772–782. doi: 10.1207/s15374424jccp3304_12
Keywords: emotion processing, emotion–cognition interaction, cognition, fMRI, Stroop
Citation: Raschle NM, Fehlbaum LV, Menks WM, Euler F, Sterzer P and Stadler C (2017) Investigating the Neural Correlates of Emotion–Cognition Interaction Using an Affective Stroop Task. Front. Psychol. 8:1489. doi: 10.3389/fpsyg.2017.01489
Received: 16 December 2016; Accepted: 17 August 2017;
Published: 01 September 2017.
Edited by:
Sumitava Mukherjee, Ahmedabad University, IndiaReviewed by:
Kenneth Sung Lai Yuen, Johannes Gutenberg-Universität Mainz, GermanyCopyright © 2017 Raschle, Fehlbaum, Menks, Euler, Sterzer and Stadler. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nora M. Raschle, bm9yYS5yYXNjaGxlQHVwa2JzLmNo
†Shared first authorship.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.