 Aline Figueiredo Cardoso1†
Aline Figueiredo Cardoso1† Ediane Conceição Alves1†Sidney D. Araújo da Costa1†
Ediane Conceição Alves1†Sidney D. Araújo da Costa1† Alessandra Jackeline Guedes de Moraes1†
Alessandra Jackeline Guedes de Moraes1† Dalton Dias da Silva Júnior2†Paulo Manoel Pontes Lins3†
Dalton Dias da Silva Júnior2†Paulo Manoel Pontes Lins3† Gisele Barata da Silva1*†
Gisele Barata da Silva1*†- 1Plant Protection Laboratory, Institute of Agrarian Sciences, Federal Rural University of Amazon (UFRA), Belém, Brazil
- 2Federal University of Amazonas (UFAM), Education, Agriculture and Environment Institute, Humaitá, Brazil
- 3Sococo S.A Agroindústrias da Amazônia, Belém, Brazil
Coconut production in the Amazon requires the knowledge and development of sustainable technologies to alleviate the detrimental effects of inorganic chemical fertilizers and intensive farming practices. In this study, we investigated the effects of plant growth-promoting rhizobacteria (PGPR) isolated from coconut seedlings on nutrient use efficiency (NUE) and physiological mechanisms related to biomass accumulation of seedlings grown with reduced inorganic fertilizer levels. Of the 96 PGPR isolates tested on rice plants, the isolate Bacillus cereus (UFRABC40) was selected, as it resulted in the most significant gain in growth variables. In a commercial coconut tree nursery, we subjected seedlings to two treatments, both with seven replications: control 100% NPK chemical fertilizer (CF) and B. cereus + 50% NPK CF. The results indicated that the inoculation increased phytohormone levels [190% indole acetic acid (IAA), 31% gibberellic acid GA3, and 17% gibberellic acid GA4] and leaf gas exchange [48% by assimilation of CO2 (A), 35% stomatal conductance to water vapor (gs), 33% transpiration, and 57% instantaneous carboxylation efficiency] in leaves. Furthermore, growth parameters (shoot, root, and total dry weight, height, and diameter) and macro- and micronutrient levels (95% N, 44% P, 92% K, 103 Ca, 46% Fe, 84% B) were improved. Our results show the potential ability of strain Bacillus cereus UFRABC40 to promote the growth performance of coconut seedlings under decreased application of inorganic fertilizers. The application of microbial-based products in coconut seedling production systems improves plants’ physiological performance and the efficiency of nutrient use.
Introduction
The cultivation of coconut trees is of great economic and social importance due to the value generated by coconut production. According to FAO (2018), Indonesia is the world’s largest coconut producer, followed by the Philippines, India, Sri Lanka, and Brazil. Production in Brazil occupies an area of 216 hectares, yielding approximately 2 million tons coconuts (IBGE, 2019), 1.5 million of which are obtained from green dwarf and hybrid plants (Sindcoco, 2017). The Amazon region produces 11% of the country’s coconut yield; of this, 10% (200,000 tons) comes from the state of Pará. The coconut seedlings are the first stage affecting the productivity of the perennial plant, which has a mean production-life of 40 years. Green dwarf coconut seeds have a low germination rate, and their seedlings have low vigor and quality primarily due to the incidence of leaf spots (Rabelo et al., 2006; Vinodhini and Deshmukh, 2017).
The global demand for food has resulted in large use of CFs to attain maximum agricultural efficiency. According to Wang and Li (2019), only 50% of N from fertilizer is absorbed by crops. Moreover, it has been estimated that up to 7 million tons of P per year will be used in phosphate fertilizers by 2050 (Mogollon et al., 2018). The excessive and incorrect use of fertilizers damages the environment via leaching, runoff, and erosion (Good and Beatty, 2011; Savci, 2012; Conijn et al., 2018). It also leads to changes in the soil’s physical, chemical, and microbiological characteristics (Blanco- Blanco-Canqui and Schlegel, 2013). The low efficiency of synthetic fertilizers is related to nutrient loss via leaching and evaporation to the atmosphere (Tilman, 1998; Gyaneshwa et al., 2002). Thus, the efficient use of synthetic fertilizers is important for both productivity and environmental protection (Paungfoo-Lonhienne et al., 2019). Furthermore, technologies that decrease the adverse effects of CFs on soil microbiota while promoting crop growth and productivity should be investigated.
The use of rhizobacteria in plant production can promote growth (Gange and Gadhave, 2018), and some genera such as Pseudomonas sp. and Bacillus sp. have been shown to promote the growth of coconut seedlings (George et al., 2018). Rhizobacteria can alter anatomical characteristics and improve photosynthetic, hormonal, and nutritional performance (Glick et al., 1999; Lucy et al., 2004; Lwin et al., 2012; Samaniego-Gámez et al., 2016). They also stimulate the synthesis of phytohormones such as indole acetic acid (IAA) and gibberellins that promote root and shoot growth (Pahari and Mishra, 2017). Rhizobacteria also optimize the use of CFs and are considered a sustainable technology (Angulo et al., 2020). They include N2 fixers, phosphorus, and potassium solubilizers (Bhardwaj et al., 2014). In rice, a 50% reduction in N and P fertilization resulted in better nutrient absorption and chlorophyll content (Naher et al., 2018). Additionally, the use of Bacillus amyloliquefaciens combined with 50% CF changed the hormonal behavior of oil palm seedlings in Amazonian climatic conditions, increasing IAA levels by 66%, shoot dry matter by 110%, and root dry matter by 123% and improving macro- and micronutrient uptake (Lima et al., 2020).
Coconut production in the Amazon requires knowledge and sustainable technology development to counteract the negative impacts of CF dependency. Access to the diverse range of microorganisms associated with plants and soil in the Amazon biome may improve bioinoculant production. Bioinoculant production is a process sensitive to both biotic and abiotic factors.
Therefore, this study aimed to evaluate the effects of plant growth-promoting rhizobacteria (PGPR) inoculation on coconut seedlings growth by investigating the physiological and nutritional mechanisms in seedlings grown under low CF conditions.
Materials and Methods
The experiment was conducted in a coconut tree seedling nursery, Santa Isabel do Pará-Brazil (1°13′26″ S, 48°02′29″ W).
Isolation of Bacterial Strains
Six soil samples containing roots (100 g) were collected from an 8-year-old dwarf coconut plantation in Brazil. Each sample was divided into portions of 10 g to obtain the isolates. Each soil sample (10 g) was diluted in 50 mL of sterile distilled water and agitated for 30 min. An aliquot of 20 μL was then separated from the original suspension and diluted in 80 μL (10–3). Next, an aliquot of 50 μL was separated from the concentrated suspension and seeded into three 9-mm Petri plates containing 10 mL of culture medium (per liter: g of sucrose, 8 g of hydrolyzed acid casein, 4 g of yeast extract, 2 g of K2HPO4, 0.3 g of MgSO4, and 15 g of agar) (Kado and Heskett, 1970). Plates were subsequently incubated at 27°C ± for 12 h. After incubation, colonies with different colors, borders, and morphology in the same plate were isolated and streaked into a new plate containing culture medium (Kado and Heskett, 1970). These plates were incubated for the same time and temperature as described above, followed by bacterial isolate purification. The bacteria were collected in microtubes containing distilled and sterile water and kept at 5°C in a refrigerator.
Selection of Isolates
The selection of growth-promoting isolates was carried out on rice plants, and subsequently, their interaction with coconut seedlings was tested according to the method described by de Castro et al. (2020).
Briefly, rice seeds (10 g) were inoculated with 20 mL of bacterial suspension obtained from the culture growth in liquid medium to 108 CFU⋅mL–1 (Kado and Heskett, 1970) and kept under agitation at 114 rpm at 27°C for 24 h (Filippi et al., 2011). The experimental design consisted of 97 treatments (96 rhizobacteria isolates and a control) with three replicates each in a greenhouse. Twenty-one days after germination, plants were evaluated for root and shoot length (LR and LS, respectively) and total biomass (TDM). Analysis of variance was performed for all variables, followed by a comparison of means using the Scott–Knott test (p < 0.05). The R40 isolate (Supplementary Material) resulted in superior growth parameters compared with all other treatments; therefore, it was subjected to in vitro biochemical tests and selected for subsequent testing with green dwarf coconut seedlings from Brazil.
Identification of Bacterial Isolate
The R40 isolate was cultured in culture medium 523 (Kado and Heskett, 1970) for 24 h at 28°C. Two inoculation loops were added to a microtube containing 1 mL of extraction buffer (Tris-HCl 1x). Then, DNA extraction was performed according to the method described by Mariano and Silveira (2005). The R40 isolate was identified using the 16S rDNA region gene and 27F (5′-AGAGTTTGATCMTGGCTCAG-3′) and 1492R (5′ACCTTGTTACGACTT-3′) primers (Lane et al., 1985). The PCR amplification reaction was composed of 1x Master Mix 2x (Promega) (0.05 U μL–1 Taq DNA polymerase, 4 mM MgCl2 reaction buffer, 0.4 mM of each DNTP; Promega Corporation, Madison, WI, United States), 10 μM of each primer, and 50 ng DNA. Amplification of the 16S rDNA region was performed in a thermal cycler (MasterCycler Nexus, Eppendorf, Hamburg, Germany) with the following steps: initial denaturation at 94°C for 4 min; 25 cycles of 94°C for 1 min, 55°C for 1 min, and 72°C for 1 min; and a final extension at 72°C for 7 min. Reactions were purified using 5 μL of PCR product plus 2 μL Exo-SAP enzyme (Exonuclease). Samples were purified via a thermal cycler, performed at 37°C for 4 min, followed by an incubation period at 80°C for 1 min to inactivate both enzymes irreversibly. After the purification reaction, sequencing was carried out in an automated sequencer (ABI3730) at the Laboratory of Bioinformatics and Evolutionary Biology, Federal University of Pernambuco (LABBE-UFPE).
DNA sequence analysis and assembly of the R40 isolate contigs were performed using the Staden Package (Staden et al., 1998). The nucleotide sequence of the UFRABC40 bacteria was compared with the isolate sequences available in the National Center for Biotechnology Information (NCBI) database using the BLASTn software1. Afterward, all sequences were aligned (MEGA). Bayesian inference (IB) analysis was performed by means of Mr. Bayes v. 3.2.6 (Ronquist et al., 2012) implemented in CIPRES2 using the best nucleotide replacement model. This was selected according to Aikake’s Information Criterion (AIC) through Mr. Modeltest 2.3 (Nylander, 2004) using 1,000,000,000 generations of Markov Chain Monte Carlo (MCMC) with sampling every 1,000 and 10,000 generations. Identification of access and phylogenetic trees was obtained by comparing the selected strain with the reference strains using 29 reference accessions to identify the selected strain (R40). Identifying a bacterial isolate using 16S rRNA was used to identify the strain selected with the strain of greatest homology. Subsequent probabilities were calculated after discarding the first 25% of the generations. All trees obtained from individual genes and concatenated through the IB method were visualized through the Fig Tree 1.4.1 software3.
Biochemical Tests
Indole Acetic Acid Production
The R40 isolate was grown in a Luria Bertani (LB) medium under 100 rpm agitation and incubated at 28°C for 78 h. Subsequently, 3 mL of the suspension was centrifuged at 4°C for 10 min at 4,000 rpm (Moustaine et al., 2017). Then, 90 μL of the supernatant and 60 μL of the Salkowski reagent were placed in a microtube and incubated in the dark for 30 min to determine if a change in mean media color occurred (Gordon and Weber, 1951).
Production of Siderophores
The R40 isolate was inoculated into test tubes containing a 10 mL Tryptic Soybean Broth (TSB) (1:10 diluted) medium (3 g in 1,000 mL distilled water) and incubated at 28°C under agitation at 114 rpm for 24 h. Subsequently, tubes containing the bacterial suspension were centrifuged for 10 min at 12,000 rpm. Then, 1 mL of the supernatant was transferred into another tube containing 1 mL of the blue chrome S (BCS) solution. Fifteen minutes after mixing, if siderophores were produced, the dark blue mixture turned yellow (Schwyn and Neilands, 1987).
Phosphate Solubilization
The R40 isolate was grown in an NBRIP growth medium containing 10 g glucose, 2.5 g Ca3(PO4), 25 g MgCl2W6H2O, 0.25 g MgSO4W7H2O, 0.2 g KCl, and 0.1 g (NH4)2SO4 (Nautiyal, 1999), at a pH of 7.0, and with the addition of 1.5% agar in triplicate. The plates were incubated for 14 days at 28°C; the presence of a halo was indicative of phosphate solubilization.
Evaluating R40 Isolate Ability to Promote Growth Coconut Seedlings
Coconut Seeds Preparation
In a coconut tree nursery, the coconut seeds were sown in wooden boxes (30 cm high, 2 m wide, and 10 m long) containing coconut fiber and moistened daily for 90 days. The chemical characterization of the coconut fiber substrate (Golden Mix type 4 – AMAFIBRA®) was as follows: 0.086 g kg–1 N, 0.264 g kg–1 P, 0.580 g kg–1 K, 0.128 mg kg–1 Ca, 0.447 mg kg–1 MgO, 272.86 mg kg–1 S, 42.25 mg Na, 0.703 mg L–1 B, 0.12 g kg–1, copper Cu, 0.5 mg kg–1 Fe, 0.6 mg kg–1 Mn, 0.78 mg kg–1 Zn, and 92.43% organic matter (OM).
Coconut Seedling Preparation
Seedlings with 15 cm tall and two leaves were transplanted to polyethylene bags (40 × 40 × 40 cm) containing 7.5 kg/bag of coconut fiber (50% moist). Chemical fertilization was performed 30 days after transplanting (DAT) with 3 g urea, 40 g simple superphosphate (18% P2O5), 10 g potassium chloride (60% K2O), and 5 g magnesium oxide (30% Mg) (Lins and Viégas, 2008).
Evaluation of Coconut Seedling Growth
For the establishment of treatments, the recommendation of commercial fertilization was followed. Thus, the control treatment is characterized as 100% chemical fertilization (CF) and without the use of bioinoculant. The control treatment used standard CF (as described in the previous section) applied at 90 and 150 DAT. The treatment with rhizobacteria comprised inoculation with a suspension of R40 isolate (108 CFU) + 50% standard CF at 90 DAT. The bacterial strain was inoculated by applying 300 mL plant–1 of a suspension at 108CFU⋅mL–1 through watering at 40 and 70 DAT. Biometrics, gas exchange, hormone levels, and nutrient levels were evaluated at 160 DAT. The experimental design was completely randomized, with 10 replications and 2 treatments.
Biometrics
The following biometric variables were evaluated: shoot, root, and total dry weight, height, and diameter. Additionally, the leaf area was determined from photographs using the APS Assess software version 2.0 (Lamari, 2002).
Leaf Gas Exchange
Gas exchange parameters were estimated using the first physiologically mature, fully expanded leaf, from apex to base, at 3 months of age. The net assimilation of CO2 (A), stomatal conductance to water vapor (gs), transpiration rate (E), and instantaneous carboxylation efficiency (A/Ci) were estimated between 08:00 and 11:00 am using a portable open-flow gas-exchange system (LI6400XT, LI-COR, Lincoln, NE, United States) under an external CO2 concentration of 400 μmol mol–1 of air and artificial photosynthetically active radiation (PAR) of 900 μmol of photons m–2 s–1.
Hormone Profile
Indole acetic acid (IAA) and gibberellic acid (GA3 and GA4) hormone levels were determined according to Munné-Bosch et al. (2011). For this, 300 mg of fresh tissue from the second leaf of each plant was stored in liquid N. The tissue material was then lyophilized and macerated in liquid N. Then, 40 mg dry mass was weighed, and 400 μL of extraction solvents (methanol:isopropyl alcohol:acetic acid; 20:79:1) was added. Samples were vortexed four times for 20 s (on ice), sonicated for 5 min, placed on ice for 30 min, and then centrifuged at 13,000 rpm for 10 min at 4°C. After centrifugation, 350 μL of supernatant was removed and transferred to another microtube. Approximately 300 μL of the extract obtained in flasks was added, and 5 μL of the mixture was injected into the NuBioMol LC/MS system (Biomolecule Analysis Center, UFV, Brazil). A chromatography column (Agilent Eclipse; Agilent Technologies, Santa Clara, CA, United States) was used (RRHD, C18 column, 50 mm × 2.1 mm, 1.8 μm) with a flow rate of 0.3 mL min–1 coupled to a triple quadrupole QQQ mass spectrometer (Agilent Technologies). Mass spectra were alternately negative/positive operated according to the retention time for each hormone. The generated mass spectra were processed using the MassHunter software to obtain the extracted ion chromatograms (XIC) for each transition and area values, indicating the abundance of each hormone. A curve pattern for each hormone over a concentration range from 0.1 to 300 ng mL–1 was used to convert the XIC area values into ng g–1 of plant tissue. Molecular mass spectra analysis was conducted using the Skyline software.
Nutritional Content
Leaf samples dried in an oven with forced air circulation at 60°C were ground. The samples were submitted to sulfuric and nitroperchloric digestion. The determination of nitrogen (N) was by distillation in Microdistillator Kjeldhal, phosphorus (P) by visible ultraviolet spectrophotometry (UV-VIS), and potassium (K), calcium (Ca), and iron (Fe) by absorption spectrometry atomic, flame modality (EAA/cham). Analysis of boron (B) was undertaken after dry digestion of the samples using the method described by Azometrinah (Malavolta et al., 1997; Carmo et al., 2000). The nutrient use efficiency (NUE) was estimated from agronomic efficiency, NUE (g DW g –1) = aerial dry weight (g)/plant applied nutrient (g) (Fageria et al., 2008).
Statistical Analysis
Differences among means for treatments were evaluated using the t-test (p < 0.05). All data were analyzed using the R software (R Core Team, 2017).
Results
Isolate Selection in the Plant Model
The rice plants used as a model for selecting rhizobacteria showed that the R40 isolate was better for root and shoot length variables and total biomass compared with the other treatments (Table 1). The isolate R40 increased by 101% shoot length, 60% root length and 280% total biomass in comparation that is seedlings non-bioinoculation. In vitro biochemical tests were performed, showing that the R40 isolate was able to solubilize phosphate as proven by the halo formation around the bacterial colonies. There were also reactions indicating siderophore and IAA production when the R40 isolate was exposed to CAS solution and Salkowski’s test, respectively (Figure 1). Thus, the R40 isolate was selected to evaluate growth promotion in green dwarf coconut seedlings in Brazil under nursery conditions.
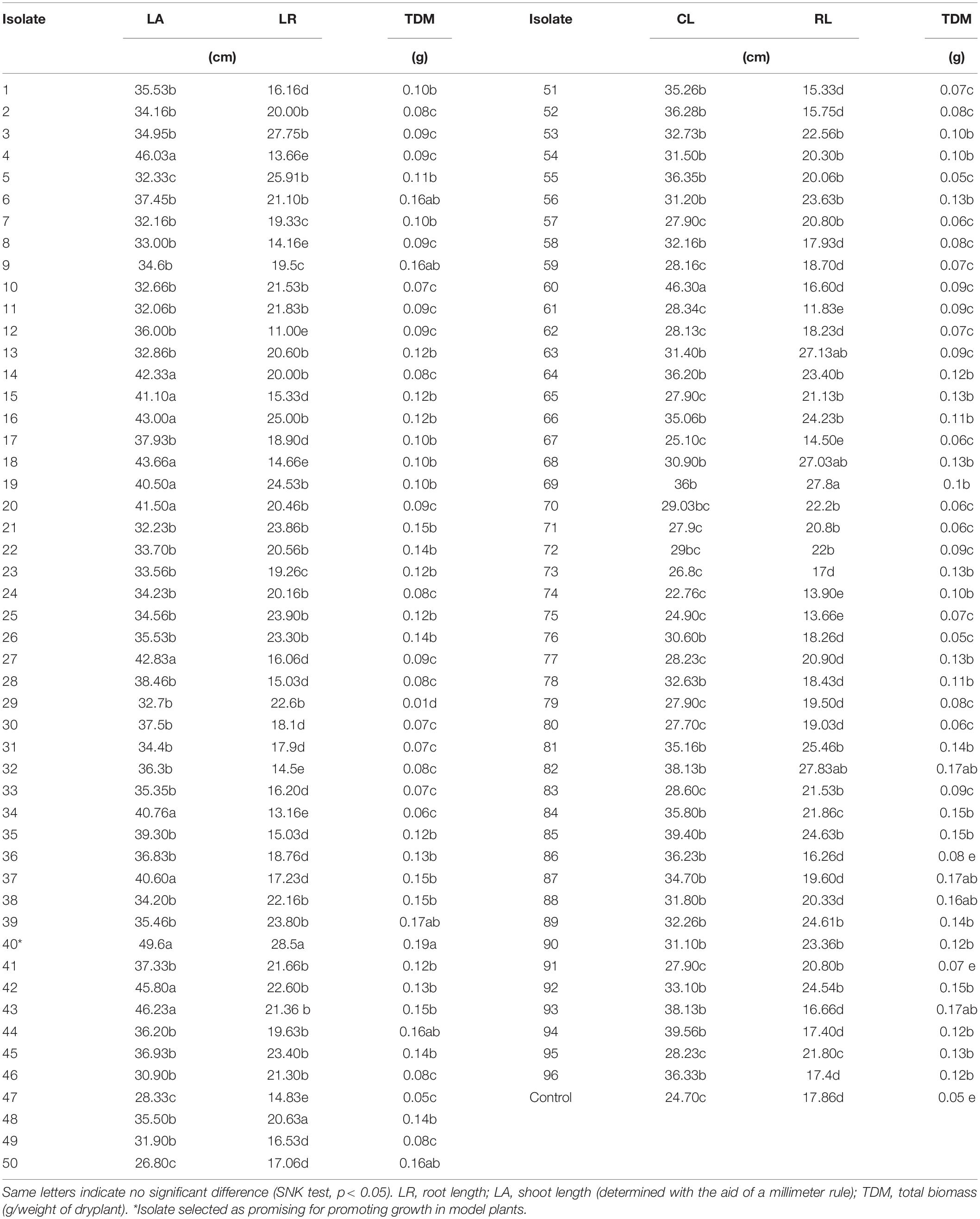
Table 1. Selection of growth-promoting rhizobacteria in rice, isolated from the rhizosphere of commercially grown green dwarf coconut trees from Santa Isabel, PA, Brazil.
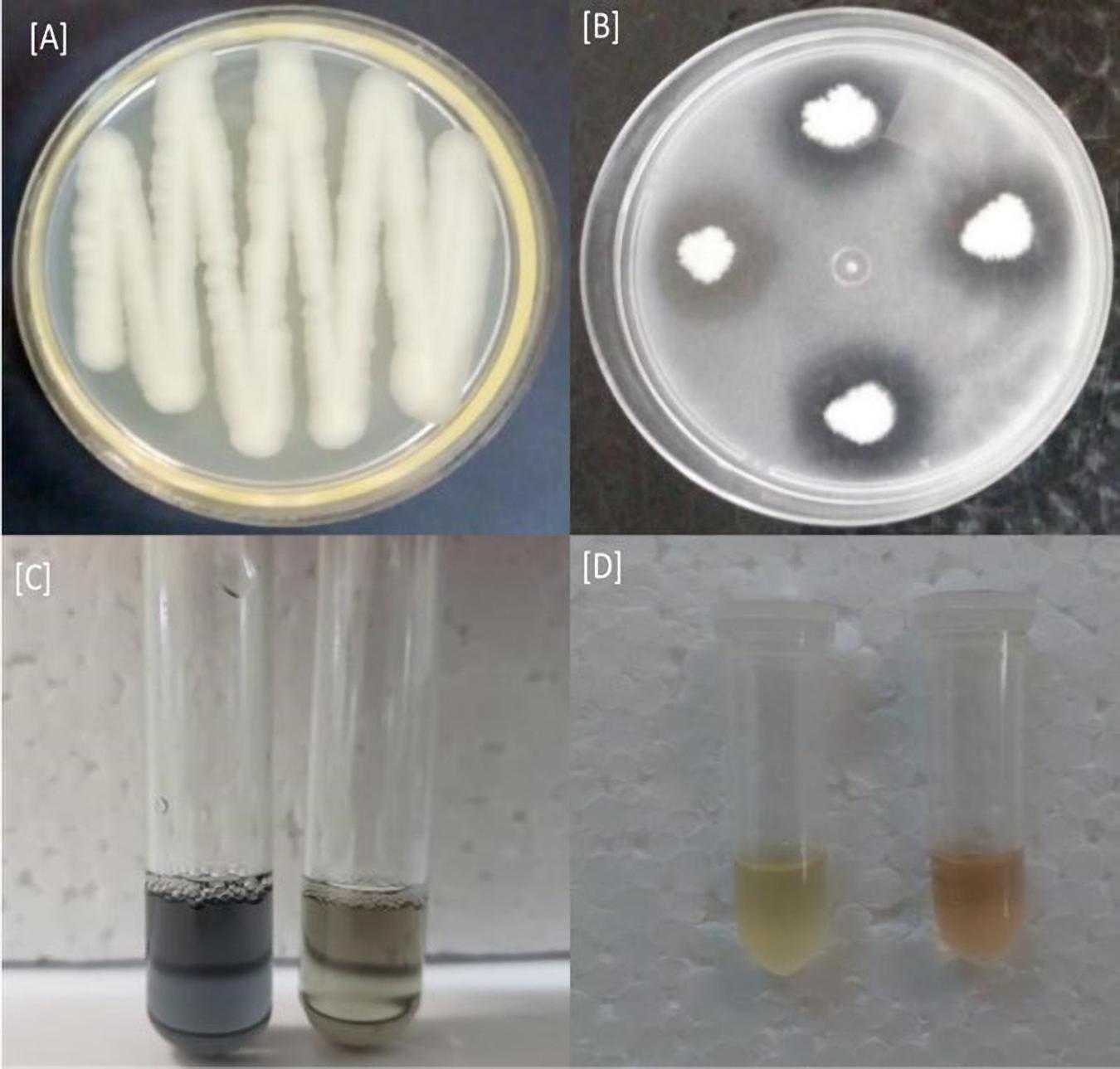
Figure 1. R40 isolate in vitro biochemical tests. (A) R40 isolate colony in Petri plate, (B) phosphate solubilization detection, (C) siderophore production, and (D) indole acetic acid (IAA) production.
Growth-Promotion Coconut Seedlings
The R40 isolate sequence was compared in GenBank using the BLASTn tool. The isolate showed 100% identity with the genus Bacillus (ATCC14579T). Based on the construction of the phylogenetic tree from 29 accesses, it was possible to identify the isolate as B. cereus. The sequence was deposited in GenBank as B. cereus (UFRABC40) with accession number MN393059 (Supplementary Table 1 and Supplementary Figure 1).
The inoculation of strain B. cereus promoted the growth of coconut seedlings even in the presence of lower levels of chemical fertilizers (Figure 2). The application of B. cereus significantly increased shoot dry weight (47%), root dry weight (122%), total dry weight (35%), height (26%), and diameter (30%) compared with the control treatment (Figure 3).

Figure 2. Green dwarf coconut seedlings from Brazil. (A) Control coconut seedlings with 100% chemical fertilization and (B) coconut seedlings inoculated with Bacillus cereus and 50% chemical fertilization.
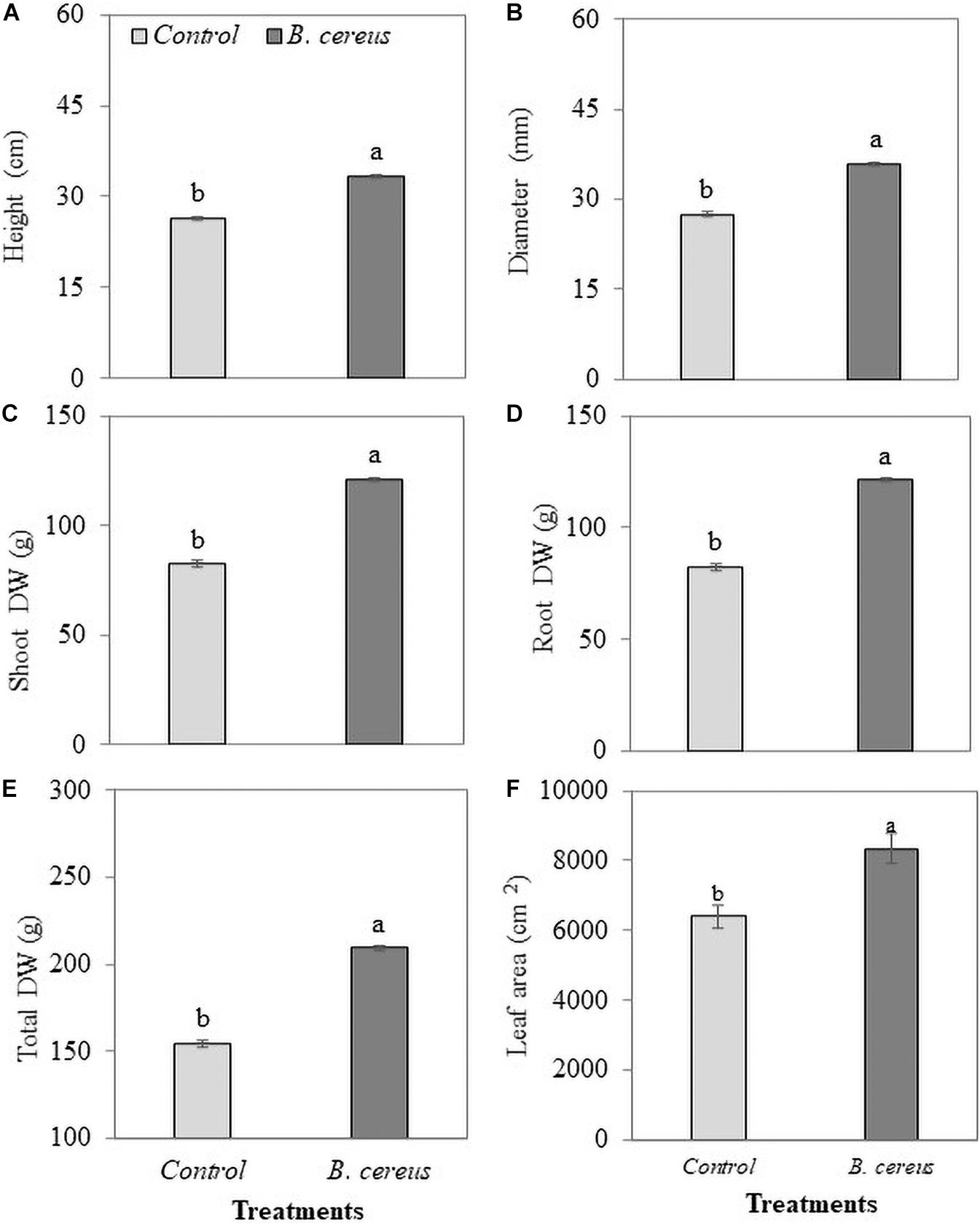
Figure 3. Biometrics of green dwarf coconut seedlings from Brazil (uninoculated and inoculated with B. cereus). (A) Height, (B) stem diameter, (C) shoot dry matter, (D) root dry matter, (E) total dry matter (E), and (F) leaf area in plants with 100% chemical fertilization (control) and plants inoculated with Bacillus cereus with 50% chemical fertilization. The same letters indicate no significant difference (t-test, p < 0.05).
Gas exchange was also influenced by inoculation with B. cereus from the third month of age in Brazilian green dwarf coconut tree seedlings. The nursery trial results indicated a maximum increase of 48% in A, 35% in gs, 33% in E, and 57% in A/Ci in plants inoculated with B. cereus compared with the uninoculated control (Figure 4). B. cereus inoculation led to an increase of 190% in IAA, 31% in GA3, and 17% in GA4 in coconut seedlings compared with the uninoculated control (Figure 5).
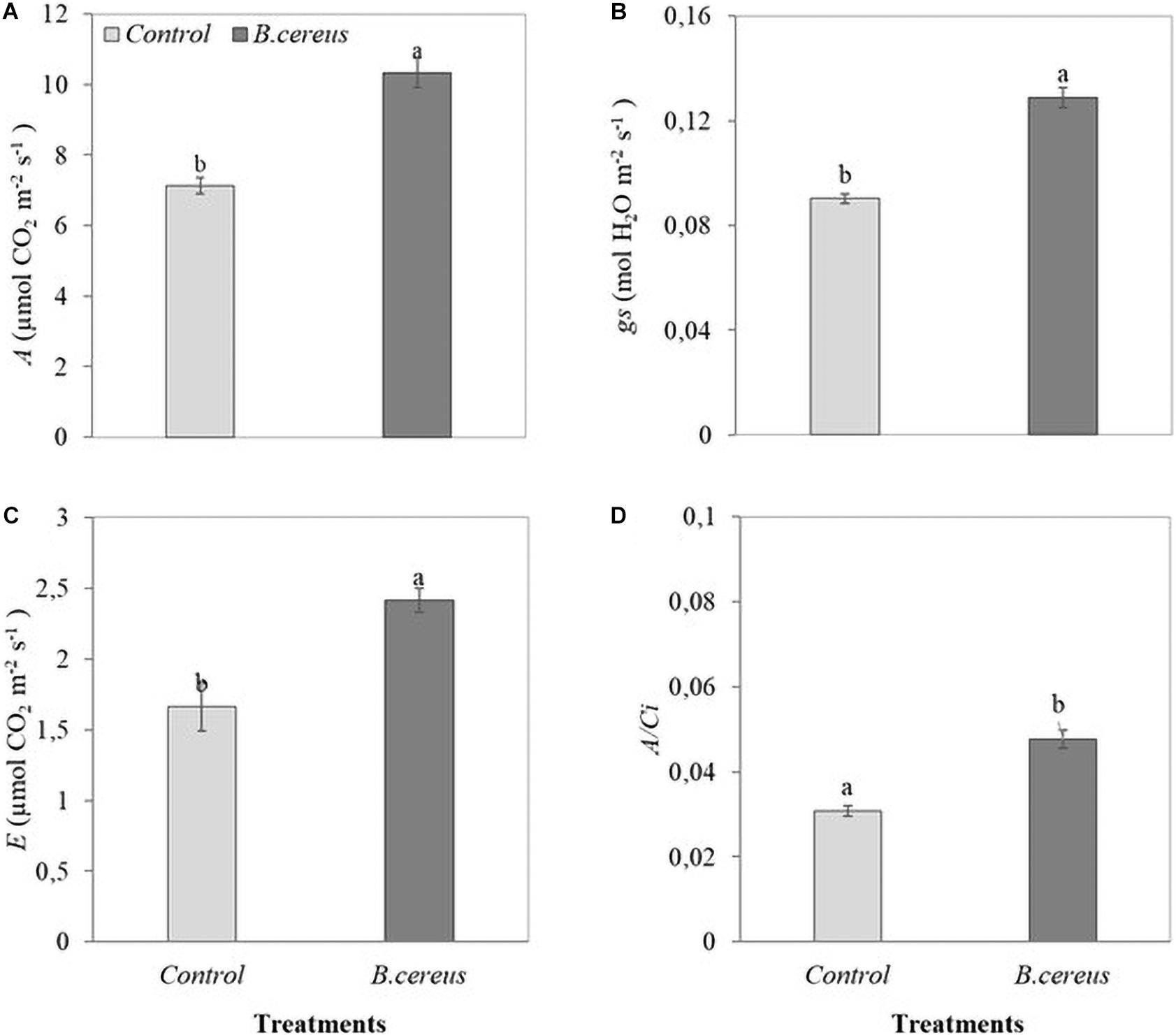
Figure 4. Leaf gas exchange in green dwarf coconut seedlings. (A) The net assimilation of CO2 (A), (B) stomatal conductance to water vapor (gs), (C) transpiration (E), and instantaneous carboxylation efficiency (A/Ci) in plants with 100% chemical fertilization (control) and plants inoculated with Bacillus cereus with 50% chemical fertilization. The same letters indicate no significant difference (t-test, p < 0.05).
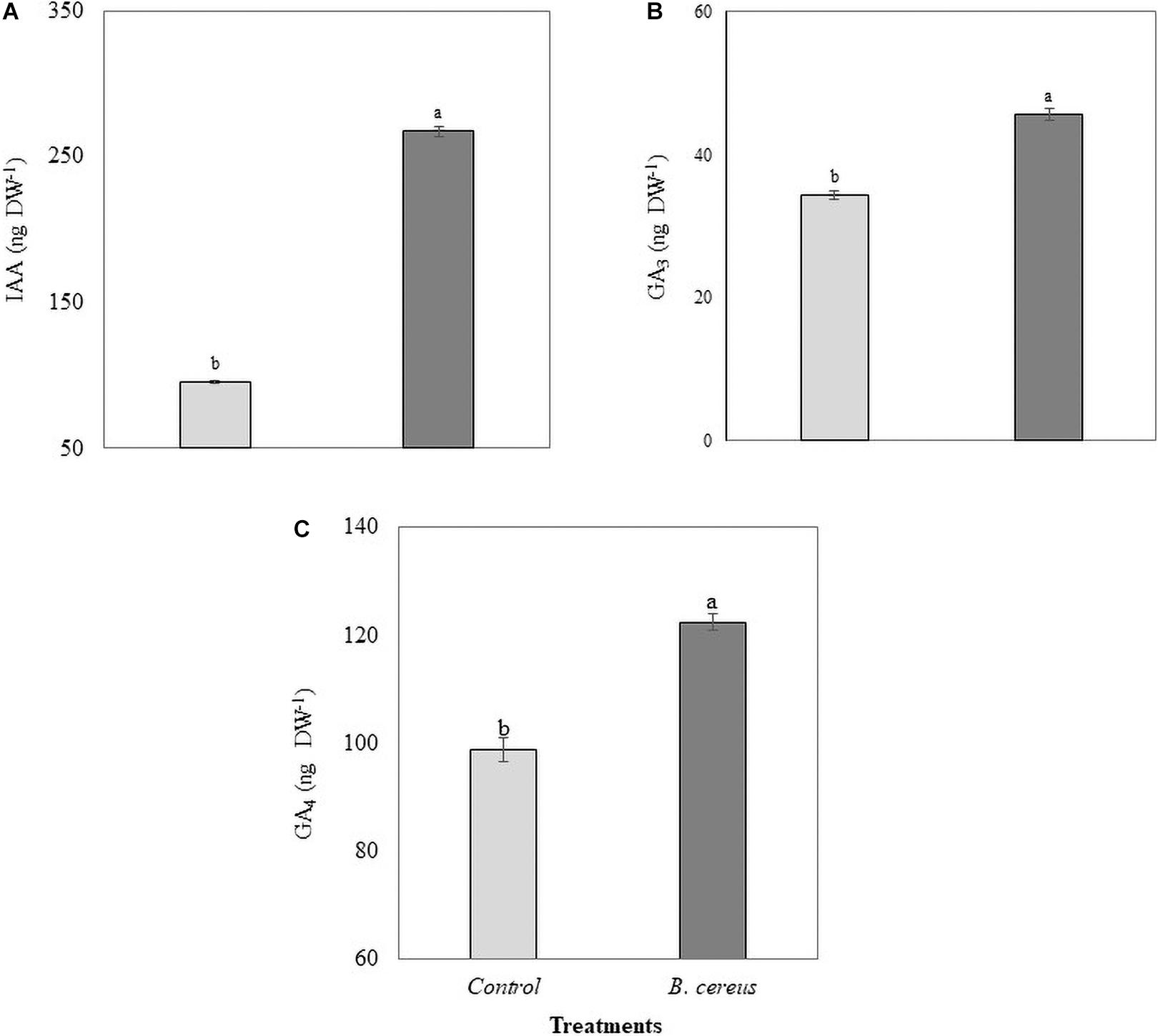
Figure 5. Phytohormone quantification in coconut seedlings. (A) Indoleacetic acid (IAA), (B) gibberellic acid GA3, and (C) gibberellic acid GA4 in plants with 100% chemical fertilization (control, T1) and plants inoculated with Bacillus cereus with 50% chemical fertilization (T2). The same letters indicate no significant difference (t-test, p < 0.05).
The B. cereus application significantly increased macronutrients and micronutrients in coconut plants by 95%, 44%, 82%, 103%, 46%, and 84% for N, P, K, Ca, Fe, and B, respectively (Table 2), compared with the control. Treatment with B. cereus shows greater efficiency in the use of nutrients (Table 3).

Table 2. Nutritional content in the shoots of green dwarf coconut seedlings from Brazil.
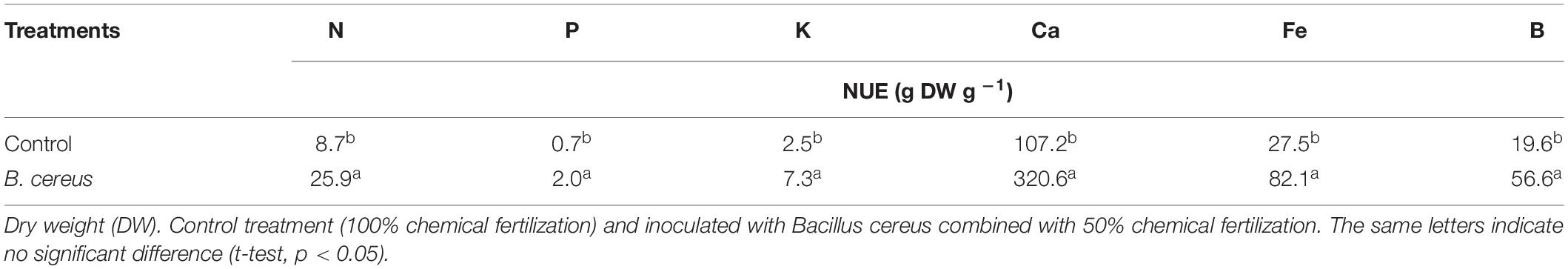
Table 3. Nutrient use efficiency (NUE) of green dwarf coconut seedlings from Brazil.
Discussion
The rhizobacteria B. cereus promoted the growth of green dwarf coconut seedlings. Bacterial inoculation induced changes in the metabolism of coconut tree seedlings, by stimulating hormonal modulation, and photosynthetic performance and efficient use of nutrients, which ultimately resulted in a greater growth of coconut tree seedlings. These results may be due to the increase in IAA concentrations derived from the enhanced production by B. cereus strain (as shown in Figure 1) or due to the regulation of its biosynthesis in the plants (Figure 5). Indole acetic acid is responsible for modulating the differentiation and elongation of lateral roots, as well as increasing the number of root hairs, therefore promoting greater nutrient absorption (Costa et al., 2015; Cassán et al., 2020). Combined with the positive effects of IAA, it is possible that bioinoculation has reduced ethylene levels in the roots by the activity of the enzyme 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase. This enzyme regulates ethylene synthesis by cleaving its acid precursor, ACC (Belimov et al., 2004; Siddikee et al., 2011), thus decreasing the negative effects of ethylene on growth and allowing the development of a better root system in inoculated plants (Glick, 2012). Bacillus cereus inoculation resulted in the more efficient use of macro- and micronutrients. A significant positive effect on the levels of the seven nutrients evaluated was observed (Tables 2, 3). Recommendations for the use of CFs are based on soil analysis and the mechanisms of macro- and micronutrient loss, such as volatilization, leaching, and adsorption, resulting in low absorption by the plant (Biswas et al., 2000; Ahmad et al., 2019). These root regions have a high influx of available ions, resulting in greater water and nutrient absorption. Root system changes induced by microorganisms improve NUE. This phenomenon has been reported in the interaction between banana and the bacterial strains Pseudomonas fluorescens Ps006 and Bacillus amyloliquefaciens Bs006 (Gamez et al., 2018) resulting in greater plant biomass. Another mechanism involved in the NUE of biostimulated coconut plants is the mineralization rate of the coconut fiber substrate by bioinoculant enzymatic activity, resulting in increased nutrient availability.
For N, B. cereus may make N available from the organic N contained in the coconut fiber via ammonium and nitrite oxidation. This was described by Di Benedetto et al. (2016), who found that Pseudomonas and Bacillus strains were able to oxidize ammonia to NO2– ions (nitrosification) and then to NO3– ions (nitrification). In a study with Triticum aestivum, the inoculation of Bacillus megaterium SNji (BmeSNji) and Azospirillum brasilense 65B (Abr65B) provided greater availability of nutrients to plants, such as N, from decomposition of organic matter resulting in greater accumulation of biomass in plants (Nguyen et al., 2019).
Coconut plants inoculated with B. cereus also had enhanced K uptake. This might be explained by its ability to produce organic acids that act in the mineralization of K present in the coconut fiber substrate, making K+ ions available for plant absorption. According to Sheng and He (2006), the Bacillus edaphicus NBT strain and its mutants can chelate metals and mobilize K from K-containing minerals using organic acids such as citric, oxalic, tartaric, and succinic.
Solubilization of P by rhizobacteria requires the production of phosphatases and phytases that mineralize the organic material by esters and H3PO4 anhydride hydrolysis (Tabatabai, 1994; Nannipieri et al., 2011). In maize, Pseudomonas plecoglossicida (PSB5) inoculation increased P production by 18% and its total absorption by 46%. This was due to the increased activity of enzymes such as dehydrogenases and phytases (Kaur and Reddy, 2013). In this study, B. cereus was able to solubilize P in vitro, and plants inoculated with this strain showed higher levels of P than control ones. As observed in vitro, B. cereus can produce siderophores, which are low molecular weight iron-chelating compounds with a great affinity and selectivity for binding and forming a Fe complex (III), reducing Fe3+ to Fe2+ (Hider and Kong, 2010; Fukushima et al., 2013). In mustard, the capacity of Bacillus sp. PZ-1 to produce siderophores resulted in higher Fe levels available for the plant (Yu et al., 2017).
Boron is a micronutrient essential for plant growth and development, and coconut plants have high B requirements (Moura et al., 2013). Brazilian Amazonian soils are generally deficient in B, and appropriate CF use is critical to avoid B deficiency. However, the application of excess exogenous B can easily be lethal to plants. Boron is not described as an essential nutrient for PGPR growth; however, in Arthrobacter nicotinovorans strain C, phenylboronic acid (PBA) catabolism was demonstrated, releasing B as orthoboric acid [B(OH)3] (Negrete-Raymond et al., 2003). Rhizobacterium-mediated B availability to plants occurs through the production of organic acids in the rhizosphere region, resulting in medium acidification and pH decrease, the latter being the main limiting factor in B availability (Deubel et al., 2000; Turan et al., 2006). Concomitant B uptake by the plant occurs through the mass flow from transpiration (Alpaslan and Gunes, 2001). Therefore, the increased transpiration rate provided by PGPR inoculation can influence B absorption (Mayak et al., 2004; Dodd and Pérez-Alfocea, 2012). In the present study, plants inoculated with B. cereus had almost twice the accumulated B in the shoots and twice the transpiration rate compared with the control. According to our analysis, this may be due to the enzymatic activity of B. cereus on the coconut fiber substrate that contained 70.35 ppm of B. The positive relationship between P and B absorption found in plants inoculated with Bacillus sp. has been recorded in canola; Bacillus improved B and P availability by 37% and 30%, respectively, in native soil (Samreen et al., 2019). These results are like ours, obtained with B. cereus coconut seedlings (Table 2). The increasing use of CFs especially with NPK and B is rapidly making them polluting agents. When not absorbed by the plants, they are leached and deposited in watercourses, or immobilized and accumulated in the soil. Implementing microbial technology in coconut seedlings tree production systems improves the physiological performance and NUE of plants, reducing the need for CFs.
In this study, the increases in the root system induced by B. cereus provided coconut seedlings with a greater possibility of absorption and translocation of nutrients, contributing to a greater growth in the aerial part (Amir et al., 2005). The increase in the aerial part growth and leaf expansion promoted by B. cereus can be attributed to the greater synthesis of gibberellins, according to the results obtained in this study (Figure 3). Increased active gibberellin concentration in the leaf tissue was stimulated by the activity of Azospirillum spp. because this microorganism promoted a significant synthesis and consequent increase in the concentration of this phytohormone (Lucangeli and Bottini, 1997; Piccoli et al., 1997; Cassán et al., 2001). Similar results were observed in alder (Alnus glutinosa) plants, where the use of Bacillus sp. enhanced the production of several isomers of gibberellins (GA1, GA3, GA4, and GA20) that were responsible for leaf area expansion and increased leaf emission rate (Gutiérrez-Mañero et al., 2001; Chauhan et al., 2015). Larger leaf areas resulted in a higher biomass accumulation in B. cereus inoculated plants.
This leaf modulation promoted by the microorganisms led to increased light capture and, consequently, increased CO2 assimilation. The increased CO2 input is due to a larger stomatal opening, verified in the present study, allowing greater CO2 diffusion, and reducing stomatal resistance (Flexas et al., 2012; Zhang et al., 2017). B. cereus regulates most of the Rubisco carboxylation, the electron transport rate, and increases the supply of ATP and NADPH molecules for photosynthesis (Shi et al., 2010), increasing carbon fixation and, consequently, increasing carbohydrate production as observed in the improved growth parameters of B. cereus inoculated plants.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/genbank/, MN393059.
Author Contributions
AC was present in all study assays and was responsible for fieldwork, statistical analysis of data, interpretation of results, and writing the manuscript. SC was present in all study assays and was responsible for fieldwork. DS, AM, EA, and PL were responsible for statistical analysis, analysis of economic indicators, and interpretation of results and assisted in writing and revising the manuscript. GS was responsible for project management, guiding the students through all stages of the process and also responsible for the verification and monitoring of field data and writing the manuscript. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank reviewers for their valuable comments and suggestions. Discussions with Gledson Luiz Salgado de Castro (Universidade Federal Rural da Amazônia, Brazil) were highly valuable in the development of this work. We thank the Federal University of Amazonia and the Plant Protection Laboratory for providing the facilities. We also thank the Amazon Foundation for the Support of Studies and Research of the State of Pará (FAPESPA) and Sococo S/A for their partnership and support, in particular the team from the phytopathology department.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2021.649487/full#supplementary-material
Supplementary Figure 1 | Phylogenetic trees analysis of R40 (UFRABC40). Major Bacillus cereus clades and groups are indicated comparing the selected strain with the reference strains.
Supplementary Table 1 | Identification of access and phylogenetic trees obtained by comparing the selected strain (R40) with the reference strains.
Footnotes
- ^ https://www.ncbi.nlm.nih.gov
- ^ https://www.phylo.org/portal2/home.action
- ^ http://tree.bio.ed.ac.uk/software/figtre
References
Ahmad, M., Adil, Z., Hussain, A., Mumtaz, M. Z., Nafees, M., Ahmad, I., et al. (2019). Potential of phosphate solubilizing Bacillus strains for improving growth and nutrient uptake in mungbean and maize crops. Pak. J. Agric. Sci. 56, 283–289. doi: 10.21162/PAKJAS/19.7285
Alpaslan, M., and Gunes, A. (2001). Interactive effects of boron and salinity stress on the growth, membrane permeability and mineral composition of tomato and cucumber plants. Plant Soil 236, 123–128. doi: 10.1023/A:1011931831273
Amir, H. G., Shamsuddin, Z. H., Halimi, M. S., Marziah, M., and Ramlan, M. F. (2005). Enhancement in nutrient accumulation and growth of oil palm seedlings caused by PGPR under field nursery conditions. Commun. Soil Sci. Plant Anal. 36, 2059–2066. doi: 10.1080/00103620500194270
Angulo, J., Martínez-Salgado, M. M., Ortega-Blu, R., and Fincheira, P. (2020). Combined effects of chemical fertilization and microbial inoculant on nutrient use efficiency and soil quality indicators. Sci. Agropecu. 11, 375–380. doi: 10.17268/sci.agropecu.2020.03.09
Belimov, A. A., Dodd, I. C., Hontzeas, N., Theobald, J. C., Safronova, V. I., and Davies, W. J. (2004). Rhizosphere bacteria containing 1-aminocyclopropane-1-carboxylate deaminase increase yield of plants grown in drying soil via both local and systemic hormone signalling. New Phytol. 181, 413–423. doi: 10.1111/j.1469-8137.2008.02657.x
Bhardwaj, D., Ansari, M. W., Sahoo, R. K., and Tuteja, N. (2014). Biofertilizers function as key player in sustainable agriculture by improving soil fertility plant tolerance and crop productivity. Microb. Cell Fact. 13:66. doi: 10.1186/1475-2859-13-66
Biswas, J. C., Ladha, J. K., and Dazzo, F. B. (2000). Rhizobia inoculation improves nutrient uptake and growth of lowland rice. Soil Sci. Soc. Am. J. 64, 1644–1650. doi: 10.2136/sssaj2000.6451644x
Blanco-Canqui, H., and Schlegel, A. J. (2013). Implications of inorganic fertilizer application of irrigated corn on soil properties: lessons learned after 50 years. J. Environ. Qual. 42, 861–871. doi: 10.2134/jeq2012.0451
Carmo, C. D. S., de Araujo, W. S., Bernardi, A. D. C., and Saldanha, M. F. C. (2000). Métodos de Análise de Tecidos Vegetais Utilizados na Embrapa Solos. Rio de Janeiro: Embrapa Solos-Circular Técnica (INFOTECA-E).
Cassán, F., Bottini, R., Schneider, G., and Piccoli, P. (2001). Azospirillum brasilense and Azospirillum lipoferum hydrolyze conjugates of GA20 and metabolize the resultant aglycones to GA1 in seedlings of rice dwarf mutants. Plant Physiol. 125, 2053–2058. doi: 10.1104/pp.125.4.2053
Cassán, F., Coniglio, A., López, G., Molina, R., Nievas, S., Carlan, C. L., et al. (2020). Everything you must know about Azospirillum and its impact on agriculture ans beyond. Biol. Fertil. Soils 56, 461–479. doi: 10.1007/s00374-020-01463-y
Chauhan, H., Bagyaraj, D. J., Selvakumar, G., and Sundaram, S. P. (2015). Novel plant growth promoting rhizobacteria—prospects and potential. Appl. Soil Ecol. 95, 38–53. doi: 10.1016/j.apsoil.2015.05.011
Conijn, J. G., Bindraban, P. S., Schröder, J. J., and Jongschaap, R. (2018). Can our food system meet food demand within planetary boundaries? Agric. Ecosyst. Environ. 251, 244–256. doi: 10.1016/j.agee.2017.06.001
Costa, R. R. G. F., Quirino, G. S. F., Naves, D. C. F., Santos, C. B., and Rocha, A. F. S. (2015). Efficiency of inoculant with Azospirillum brasilense on the growth and yield of second-harvest maize. Pesqui. Agropecu. Trop. 45, 304–311. doi: 10.1590/1983-40632015v4534593
de Castro, G. L. S., Rêgo, M. C. F., Silvestre, W. V. D., Batista, T. F. V., and da Silva, G. B. (2020). Açaí palm seedling growth promotion by rhizobacteria inoculation. Braz. J. Microbiol. 51, 205–216. doi: 10.1007/s42770-019-00159-2
Deubel, A., Gransee, A., and Merbach, W. (2000). Transformation of organic rhizodeposits by rhizoplane bacteria and its influence on the availability of tertiary calcium phosphate. J. Plant Nutr. Soil Sci. 163, 387–392. doi: 10.1002/1522-2624(200008)163
Di Benedetto, N. A., Campaniello, D., and Bevilacqua, A. (2016). “Characterization of autochthonous plant growth promoting bacteria in relation to durum wheat nitrogen use efficiency,” in Proceedings of the Plant Biology Europe Congress EPSO/FESPB, (Prague Czech Republic), 26–30. doi: 10.3934/microbiol.2017.3.413
Dodd, I. C., and Pérez-Alfocea, F. (2012). Microbial amelioration of crop salinity stress. J. Exp. Bot. 63, 3415–3428. doi: 10.1093/jxb/ers033
Fageria, N. K., Baligar, V. C., and Li, Y. C. (2008). The role of nutrient efficient plants in improving crop yields in the twenty first century. J. Plant Nutr. 31, 1121–1157.
FAO (2018). Food and Agriculture Data. Available online at: http://www.fao.org/faostat/es/#rankings/countries_by_commodity_exports (accessed September 16, 2019).
Filippi, M. C. C., Da Silva, G. B., Silva-Lobo, V. L., Côrtes, M. V. C., Moraes, A. J. G., and Prabhu, A. S. (2011). Leaf blast (Magnaporthe oryzae) suppression and growth promotion by rhizobacteria on aerobic rice in Brazil. Biol. Control 58, 160–166. doi: 10.1016/j.biocontrol.2011.04.016
Flexas, J., Flexas, J., Barbour, M. M., Brendel, O., Cabrera, H. M., Carriquí, M., et al. (2012). Mesophyll diffusion conductance to CO2: an unappreciated central player in photosynthesis. Plant Sci. 19, 70–84. doi: 10.1016/j.plantsci.2012.05.009
Fukushima, T., Allred, B. E., Sia, A. K., Nichiporuk, R., Andersen, U. N., and Raymond, K. N. (2013). Gram positive siderophore-shuttle with iron-exchange from Fe-siderophore to apo-siderophore by Bacillus cereus YxeB. Proc. Natl. Acad. Sci. U.S.A. 110, 13821–13826. doi: 10.1073/pnas.1304235110
Gamez, R., Cardinale, M., Montes, M., Ramirez, S., Schnell, S., and Rodriguez, F. (2018). Screening, plant growth promotion and root colonization pattern of two rhizobacteria (Pseudomonas fluorescens Ps006 and Bacillus amyloliquefaciens Bs006) on banana cv. Williams (Musa acuminata Colla). Microbiol. Res. 220, 12–20. doi: 10.1016/j.micres.2018.11.006
Gange, A. C., and Gadhave, K. R. (2018). Plant growth-promoting rhizobacteria promote plant size inequality. Sci. Rep. 8:13828. doi: 10.1038/s41598-01832111-z
George, P., Gupta, A., Gopal, M., Thomas, L., and Thomas, G. V. (2018). Systematic screening strategies for identifying elite plant growth promoting rhizobacteria for coconut (Cocos nucifera L.). Int. J. Curr. Microbiol. App. Sci. 7, 1051–1074. doi: 10.20546/ijcmas.2018.705.130
Glick, B. R. (2012). Plant growth-promoting bacteria: mechanisms and applications. Scientifica 2012:963401. doi: 10.6064/2012/963401
Glick, B. R., Patten, C. L., Holguin, G., and Penrose, D. M. (1999). Biochemical and Genetic Mechanisms Used by Plant Growth Promoting Bacteria. London: Imperial College Press.
Good, A. G., and Beatty, P. H. (2011). Fertilizing nature: a tragedy of excess in the commons. PLoS Biol. 9:e1001124. doi: 10.1371/journal.pbio.1001124
Gordon, S. A., and Weber, R. P. (1951). Colorimetric estimation of indoleacetic acid. Plant Physiol. 26, 192–195. doi: 10.1104/pp.26.1.192
Gutiérrez-Mañero, F. J., Ramos-Solano, B., Probanza, A. N., Mehouachi, J. R., Tadeo, F., and Talon, M. (2001). The plant-growth-promoting rhizobacteria Bacillus pumilus and Bacillus licheniformis produce high amounts of physiologically active gibberellins. Physiol. Plant. 111, 206–211. doi: 10.1034/j.1399-3054.2001.1110211.x
Gyaneshwa, P., Kumar, G. N., Parekh, L., and Poole, P. S. (2002). Role of soil microorganisms in improving P nutrition of plants. Plant Soil 245, 83–93. doi: 10.1023/A:1020663916259
Hider, R. C., and Kong, X. (2010). Chemistry and biology of siderophores. Nat. Prod. Rep. 27, 637–657. doi: 10.1039/b906679a
IBGE (2019). Indicadores. Available online at: https://www.ibge.gov.br/estatisticas-novoportal/economicas/agricultura-e pecuaria/92 01-levantamento-sistematico-da-producao-agricola.html?&t=resultado (accessed July 10, 2019).
Kado, C. I., and Heskett, M. G. (1970). Selective media for isolation of Agrobacterium, Corynebacterium, Erwinia, Pseudomonas and Xanthomonas. Phytopathology 60, 969–979.
Kaur, G., and Reddy, M. S. (2013). Phosphate solubilizing rhizobacteria from an organic farm and their influence on the growth and yield of maize (Zea mays L.). J. Gen. Appl. Microbiol. 59, 295–303. doi: 10.2323/jgam.59.295
Lamari, L. (2002). Assess 2.0: Image Analysis Software For Plant Disease Quantification. Minnesota, MN: St. Paul:APS Press.
Lane, D. J. B., Pace, G. J., Olsen, D. A., Stahl, M. L., Sogin, N., and Pace, R. (1985). Rapid determination of 16S ribosomal RNA sequences for phylogenetic analyses. Proc. Natl. Acad. Sci. U.S.A. 82, 6955–6959. doi: 10.1073/pnas.82.20.6955
Lima, J. V., Tinôco, R. S., Olivares, F. L., de Moraes, A. J. G., Chia, G. S., and da Silva, G. B. (2020). Hormonal imbalance triggered by rhizobacteria enhance nutrient use efficiency and biomass in oil palm. Sci. Hortic. 264:109161. doi: 10.1016/j.scienta.2019.109161
Lins, P., and Viégas, I. (2008). Adubação do Coqueiro no Pará. Belém: Embrapa Amazônia Oriental-Documentos (INFOTECA-E).
Lucangeli, C., and Bottini, R. (1997). Effects of Azospirillum spp on endogenous gibberellin content and growth of maize (Zea mays L.) treated with uniconazole. Symbiosis 23, 63–72.
Lucy, M., Reed, E., and Glick, B. R. (2004). Applications of free living plant growth promoting rhizobacteria. Antonie van Leeuwenhoek 86, 1–25. doi: 10.1023/B:ANTO.0000024903.10757.6e
Lwin, K. M., Myint, M. M., Tar, T., and Aung, W. Z. M. (2012). Isolation of plant hormone (indole-3-acetic acid-IAA) producing rhizobacteria and study on their effects on maize seedling. Eng. J. 16, 137–144. doi: 10.4186/ej.2012.16.5.137
Malavolta, E., Vitti, G. C., and Oliveira, S. A. D. (1997). Avaliação do Estado Nutricional Das Plantas: Princípios e Aplicações. Piracicaba: Potafos.
Mariano, R. D. L. R., and Silveira, E. D. (2005). Manual de Práticas em Fitobacteriologia. Recife: Universidade Federal Rural de Pernambuco.
Mayak, S., Tirosh, T., and Glick, B. R. (2004). Plant growth-promoting bacteria confer resistance in tomato plants to salt stress. Plant Physiol. Biochem. 42, 565–572. doi: 10.1016/j.plaphy.2004.05.009
Mogollon, J. M., Beusen, A. H. W., Van Grinsven, H. J. M., Westhoek, H., and Bouwman, A. F. (2018). Future agricultural phosphorus demand according to the shared socioeconomic pathways. Glob. Environ. Chang. 50, 149–163. doi: 10.1016/j.gloenvcha.2018.03.007
Moura, J. Z., Prado, R. M., Benvindo, R. N., and Chaves, A. L. (2013). Applying boron to coconut palm plants: effects on the soil, on the plant nutritional status and on productivity boron to coconut palm trees. J. Soil Sci. Plant Nutr. 13, 79–85. doi: 10.4067/S0718-95162013005000008
Moustaine, M., Elkahkahi, R., Benbouazza, A., Benkirane, R., and Achbani, E. H. (2017). Effect of plant growth promoting rhizobacterial (PGPR) inoculation on growth in tomato (Solanum lycopersicum L.) and characterization for direct PGP abilities in Morocco. Int. J. Environ. Agric. Biotech. 2, 590–595. doi: 10.22161/ijeab/2.2.5
Munné-Bosch, S., Oñate, M., Oliveira, P. G., and Garcia, Q. S. (2011). Changes in phytohormones and oxidative stress markers in buried seeds of Vellozia alata. Flora Morphol. Distrib. Funct. Ecol. Plants 206, 704–711. doi: 10.1016/j.flora.2010.11.012
Naher, U. A., Panhwar, Q. A., Othman, R., Shamshuddin, J., Ismail, M. R., and Zhou, E. (2018). Proteomic study on growth promotion of PGPR inoculated aerobic rice (Oryza sativa L.) cultivar MR219-9. Pak. J. Bot. 50, 1843–1852.
Nannipieri, P., Giagnoni, L., Landi, L., and Renella, G. (2011). “Role of phosphatase enzymes in soil,” in Phosphorus in Action: Biological Processes in Soil Phosphorus Cycling, Soil Biology, Vol. 26, eds E. Bunemann, A. Oberson, and E. Frossard (Heidelberg: Springer), 215–243.
Nautiyal, C. S. (1999). An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 170, 265–270. doi: 10.1111/j.1574-6968.1999.tb13383.x
Negrete-Raymond, A. C., Weder, B., and Wackett, L. P. (2003). Catabolism of arylboronic acids by Arthrobacter nicotinovorans strain PBA. Appl. Environ. Microbiol. 69, 4263–4267. doi: 10.1128/AEM.69.7.4263-4267.2003
Nguyen, M. L., Spaepen, S., du Jardin, P., and Delaplace, P. (2019). Biostimulant effects of rhizobacteria on wheat growth and nutrient uptake depend on nitrogen application and plant development. Arch. Agron. Soil Sci. 65, 58–73. doi: 10.1080/03650340.2018.1485074
Nylander, J. A. A. (2004). MrModeltest v2. Program Distributed by the Author. Uppsala University: Evolutionary Biology Centre, 575.
Pahari, A., and Mishra, B. B. (2017). Characterization of Siderophore producing rhizobacteria and its effect on growth performance of different vegetables. Int. J. Curr. Microbiol. Appl. Sci. 6, 1398–1405. doi: 10.20546/ijcmas.2017.605.152
Paungfoo-Lonhienne, C., Redding, M., Pratt, C., and Wang, W. (2019). Plant growth promoting rhizobacteria increase the efficiency of fertilisers while reducing nitrogen loss. J. Environ. Manage. 233, 337–341. doi: 10.1016/j.jenvman.2018.12.052
Piccoli, P., Lucangeli, D., Schneider, G., and Bottini, R. (1997). Hydrolysis of [1717-2H2] gibberellin A20-glucoside and [1717-2H2] gibberellin A20-glucosyl ester by Azospirillum lipoferum cultured in a nitrogen-free biotin-based chemically-defined medium. Plant Growth Regul. 23, 179–182. doi: 10.1073/pnas.89.17.8045
R Core Team (2017). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rabelo, T. K., Melo, M. D. V., Loiola, C. M.,, and Aragão, W. M. (2006). Germinação de Sementes de Cultivares de Coqueiros. Comunicado Técnico. Aracaju: Embrapa Tabuleiros Costeiros.
Ronquist, F., Teslenko, M., van de Mark, P., Ayres, D. L., Darling, A., Höhna, S., et al. (2012). MrBayes 3.2: efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542.
Samaniego-Gámez, B. Y., Garruña, R., Tun-Suárez, J. M., Kantun-Can, J., Reyes Ramírez, A., and Cervantes-Díaz, L. (2016). Bacillus spp. inoculation improves photosystem II efficiency and enhances photosynthesis in pepper plants. Chil. J. Agric. Res. 76, 409–416. doi: 10.4067/S071858392016000400003
Samreen, T., Zahir, Z. A., Naveed, M., and Asghar, M. (2019). Boron tolerant phosphorus solubilizing Bacillus spp. MN-54 improved canola growth in alkaline calcareous soils. Int. J. Agric. Biol. 21, 538–546. doi: 10.17957/IJAB/15.0926
Savci, S. (2012). Investigation of effect of chemical fertilizers on environment. Apcbee Procedia 1, 287–292. doi: 10.1016/j.apcbee.2012.03.047
Schwyn, B., and Neilands, J. B. (1987). Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 160, 47–56. doi: 10.1016/0003-2697(87)90612-9
Sheng, X. F., and He, L. Y. (2006). Solubilization of potassium-bearing minerals by a wild-type strain of Bacillus edaphicus and its mutants and increased potassium uptake by wheat. Can. J. Microbiol. 52, 66–72. doi: 10.1139/w05-117
Shi, Y., Lou, K., and Li, C. (2010). Growth and photosynthetic efficiency promotion of sugar beet (Beta vulgaris L.) by endophytic bacteria. Photosynth. Res. 105, 5–13. doi: 10.1007/s11120-010-9547-7
Siddikee, M. A., Glick, B. R., Chauhan, P. S., and Yim, W. S. T. (2011). Enhancement of growth and salt tolerance of red pepper seedlings (Capsicum annuum L.) by regulating stress ethylene synthesis with halotolerant bacteria containing 1-aminocyclopropane-1carboxylic acid deaminase activity. Plant Physiol. Biochem. 49, 427–434. doi: 10.1016/j.plaphy.2011.01.015
Sindcoco (2017). Boletim Conjuntural - Importações de Coco Ralado e de Água de Coco. Novembro de 2017; Novembro de 2018. Available online at: http://www.sindcoco.com.br (accessed Dec 11, 2018).
Staden, R., Beal, K. F., and Bonfield, J. K. (1998). The staden package. Methods Mol. Biol. 132, 115–130.
Tabatabai, M. A. (1994). “Soil enzymes,” in Methods of Soil Analysis:Part 2. Microbial and Biochemical Properties, SSSA Book Series 5, ed. R. W. Weaver (Madison, WI: Soil Science Society of America).
Turan, M., Ataoglu, N., and Sahin, F. (2006). Evaluation of the capacity of phosphate solubilizing bacteria and fungi on different forms of phosphorus in liquid culture. J. Sustain. Agric. 28, 99–108. doi: 10.1300/J064v28n03_08
Vinodhini, C. C., and Deshmukh, K. V. (2017). An economic analysis of coconut farming in Karur district of Tamil Nadu India. Int. J. Curr. Microbiol. App. Sci. 6, 1566–1573. doi: 10.20546/ijcmas.2017.612.176
Wang, Z. H., and Li, S. X. (2019). Chapter three–nitrate N loss by leaching and surface runoff in agricultural land: a global issue. Adv. Agron. 156, 159–217.
Yu, S., Teng, C., Bai, X., Liang, J., Song, T., Dong, L., et al. (2017). Optimization of siderophore production by Bacillus sp. PZ-1 and its potential enhancement of phytoextration of Pb from soil. J. Microbiol. Biotechnol. 27, 1500–1512. doi: 10.4014/jmb.1705.05021
Keywords: Bacillus cereus, coconut palm, sustainable agriculture, growth promotion, PGPR
Citation: Cardoso AF, Alves EC, da Costa SDA, de Moraes AJG, da Silva Júnior DD, Lins PMP and da Silva GB (2021) Bacillus cereus Improves Performance of Brazilian Green Dwarf Coconut Palms Seedlings With Reduced Chemical Fertilization. Front. Plant Sci. 12:649487. doi: 10.3389/fpls.2021.649487
Received: 04 January 2021; Accepted: 26 April 2021;
Published: 15 October 2021.
Edited by:
Sabine Dagmar Zimmermann, Délégation Languedoc Roussillon (CNRS), FranceReviewed by:
Sofia I. A. Pereira, Universidade Católica Portuguesa, PortugalChristel Baum, University of Rostock, Germany
Brenda Román-Ponce, University of Salamanca, Spain
Copyright © 2021 Cardoso, Alves, da Costa, de Moraes, da Silva Júnior, Lins and da Silva. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gisele Barata da Silva, Z2lzZWxlYmFyYXRhc2lsdmFAZ21haWwuY29t
†These authors have contributed equally to this work