
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Pharmacol. , 26 July 2024
Sec. Inflammation Pharmacology
Volume 15 - 2024 | https://doi.org/10.3389/fphar.2024.1447677
This article is part of the Research Topic Epigenetics of inflammatory reactions and pharmacological modulation View all 5 articles
 Zhenhua Yin1,2
Zhenhua Yin1,2 Juanjuan Zhang1
Juanjuan Zhang1 Jingjing Qin1Lin Guo2
Jingjing Qin1Lin Guo2 Qingfeng Guo1
Qingfeng Guo1 Wenyi Kang2,3*
Wenyi Kang2,3* Changyang Ma2,3*
Changyang Ma2,3* Lin Chen1*
Lin Chen1*Edible fungus polysaccharides have garnered significant attention from scholars due to their safety and potential anti-inflammatory activity. However, comprehensive summaries of their anti-inflammatory properties are still rare. This paper provides a detailed overview of the anti-inflammatory effects and mechanisms of these polysaccharides, as well as their impact on inflammation-related diseases. Additionally, the relationship between their structure and anti-inflammatory activity is discussed. It is believed that this review will greatly enhance the understanding of the application of edible fungus polysaccharides in anti-inflammatory treatments, thereby significantly promoting the development and utilization of edible fungi.
Inflammation, as the initial response of the immune system, is a physiological reaction of the body to injury, infection, and stress (Medzhitov, 2008). Generally, inflammation is a natural protective response that plays a central role in the host defense system by secreting nitric oxide (NO) and pro-inflammatory cytokines. However, uncontrolled long-term or chronic inflammation can be detrimental, leading to tissue damage and numerous diseases, including fever, asthma, rheumatoid arthritis, chronic inflammatory bowel diseases, obesity, diabetes, and cancer (Varela et al., 2018; Barbu et al., 2022; Solier et al., 2023). Currently, the treatment of inflammation primarily involves chemical drugs, including non-steroidal and steroidal anti-inflammatory drugs, which are associated with numerous side effects, such as allergies, osteoporosis, hepatotoxicity, and immunosuppression (Wang and Zeng, 2019; Yu et al., 2019).
Given the critical importance of prebiotics in altering the human gut microbiota and improving host health, edible fungi are gaining attention as one of the healthiest low-calorie foods to promote overall wellbeing (Panda et al., 2024). While not widely used as food sources due to their unique and subtle taste, edible fungi are recognized for their potential in preventing or treating inflammation, cancer, diabetes, and other diseases (Chen H. Y. et al., 2023; Shamim et al., 2023). In fact, they have become very popular health foods because of their rich nutritional contents and low calories. To date, a variety of bioactive components have been extracted from edible fungi, including dietary fibers, polysaccharides, sterols, alkaloids, and terpenoids, which exhibit anti-inflammatory, hypoglycemic, immune-enhancing, and other beneficial activities (Du et al., 2018; Chopra et al., 2021; Yin et al., 2021; Mustafa et al., 2022).
As secondary metabolites, polysaccharides are among the most attractive bioactive components extracted from edible fungi. Maity et al. (2021) and Sun et al. (2022) have reviewed the structure, biological activity, and structure-activity relationship of these polysaccharides, highlighting their immunomodulatory, antibacterial, antioxidant, anti-inflammatory, and anti-tumor activities. As bioactive macromolecules, polysaccharides cannot directly enter cells. However, they can recognize pattern recognition receptors, such as β-glucan receptors and toll-like receptors (TLRs), and activate macrophages, which in turn affect the classical MAPK and NF-κ B signaling pathways, regulate the secretion of related factors, and exert anti-inflammatory effects. Additionally, due to the lack of enzymes capable of decomposing polysaccharides, they are generally considered difficult to digest and absorb in the gastrointestinal tract. Their activity primarily manifests through fermentation reactions by intestinal microorganisms.
The anti-inflammatory activities of edible fungi polysaccharides have been attracting increasing attention. To date, their potential anti-inflammatory properties have rarely been reviewed. In this work, we provide a comprehensive review of the existing anti-inflammatory activities of edible fungi polysaccharides and analyze their structure-activity relationships to elucidate the potential of edible fungi in the prevention and treatment of inflammation. We believe this review will enhance the understanding of the anti-inflammatory activities of edible fungi polysaccharides and provide valuable guidance for the development and application of new anti-inflammatory drugs. The biological activities of various polysaccharides isolated from edible fungi, such as Pleurotus ostreatus, and medicinal edible fungi, such as Ganoderma lucidum, are summarized and listed in Figure 1; Supplementary Table S1.

Figure 1. Schematic diagram of anti-inflammatory activity of polysaccharides from edible fungi.
Inflammatory cells produce various inflammatory mediators, such as NO, interleukin (IL)-1, IL-6, monocyte chemoattractant protein (MCP)-1 and tumor necrosis factor-α (TNF-α). TNF-α and IL-6 are known to play crucial roles in inflammation, apoptosis, angiogenesis, cell adhesion and transformation (Andaluz et al., 2016). NO is involved in the inflammatory response to tissue injury (Huang et al., 2011). Edible fungi polysaccharides exhibit anti-inflammatory effects by regulating cytokine secretion in inflammatory cells, including those polysaccharides from Armillaria mellea, Cordyceps cicadae, Poria cocos, G. lucidum, and Auricularia auricula-judae. For instance, a xylosyl 1,3-galactofucan (AMPS-III, 500 μg/mL) isolated from A. mellea significantly suppressed the release of TNF-α and MCP-1 in RAW264.7 macrophages and EAhy926 inducted lipopolysaccharide (LPS) and TNF-α (Chang et al., 2018). Water-soluble indigestible polysaccharides (NDPs, 40, 80, 160 μg/mL) from C. cicadae inhibited the secretion of NO, IL-1β and TNF-α in LPS-stimulated RAW264.7 macrophages (Yang C. H. et al., 2019). An alkali-soluble and carboxymethyl polysaccharide CMP33 from P. cocos (31.25–1,000 μg/mL) and a water-soluble β-1,3-D-glucan with β-1,6-D-glucosyl branches polysaccharide GLP-2 (25–100 μg/mL) from G. lucidum inhibited LPS-stimulated overproduction of NO, IL-6, TNF-α and IL-1β in RAW264.7 cells (Liu et al., 2019; Jia et al., 2022). A glucuronoxylogalactoglucomannan ME-2 (0.05–1.0 mg/mL) isolated from A. auricula-judae demonstrated anti-inflammation effects by decreasing the mRNA levels of IL-1β, INF-γ and TNF-α in a dose-dependent manner in LPS-stimulated THP-1 cells (Liang et al., 2023). Additionally, GLP1(1.25 g/L) from G. lucidum strain inhibited the expression of IL-1α in LPS-induced HaCaT inflammation model (Zhang J. C. et al., 2022). These examples illustrate that polysaccharides from edible fungi exhibit anti-inflammatory activity by regulating the secretion of inflammatory factors.
Studies have shown that the secretion of inflammatory cytokines was regulated by mitogen-activated protein kinase (MAPK), nuclear factor kappa B (NF-κB) and Janus kinase-signal transducer and activator of transcription (JAK-STAT) signal pathways. As a key transcription factor, NF-κB signal pathway is associated with pro-inflammatory cytokines and related enzymes, regulating inflammation, immune response, cell division and apoptosis, and playing a crucial role in host defense (Hu et al., 2015; Oh et al., 2019). The MAPK pathway, composed of ERΚ, p38 and JNΚ proteins, is another important signaling pathway that participates in the regulation of inflammatory process through pro-inflammatory mediators and cytokines (Xu J. et al., 2021). JAK-STAT signaling pathway also plays a significant role in regulating the inflammatory response (Schindler et al., 2007). The anti-inflammatory effects of edible fungi polysaccharides are closely related to these signaling pathway. For instance, a proteoglycan LEPS1 (100, 200, 400 μg/mL) from Lentinus edodes inhibited the secretion of factors (NO, IL-1β, IL-6 and TNF-α) by acting on p38 MAPK (p38MAPK) and JAK-STAT1 signaling pathways in LPS-induced RAW264.7 cells (Zhang et al., 2023). WPEP and NPEP from P. eryngii significantly inhibited LPS-induced inflammation in RAW264.7 macrophages by regulating the production of NO, prostaglandin E2 (PGE-2), IL-1β, TNF-α and IL-6, which was related to MAPK and NF-κB signaling pathways (Ma et al., 2020). A exopolysaccharide EPS produced by the medicinal fungus Cordyceps sinensis Cs-HK1 significantly inhibited the secretion of NO, TNF-α and IL-1β in LPS-induced THP-1 and RAW264.7 cells, likely related to the inhibition of NF-kB signaling pathway. EPS also effectively inhibited the expression of TNF-α, IL-1β, IL-10 and inducible nitric oxide synthase (iNOS) in LPS-induced acute intestinal injury in mice, alleviating intestinal injury (Li L. Q. et al., 2020).
Immunocyte-involved inflammation is thought to regulate the damage associated with various diseases. Oxidative stress, initiated by oxidants such as LPS and reactive oxygen species (ROS), is closely related to chronic inflammation. Additionally, NF-κB signaling pathway and NOD-like receptor thermal protein domain associated protein 3 (NLRP3) inflammasome activation are key mechanisms that regulate the expression of inflammatory cytokines (Du et al., 2019; Hung et al., 2019). A polysaccharide fraction from Craterellus cornucopioides (CCPP-1) inhibited LPS-induced accumulation of ROS and NO, reduced the production of TNF-α, IL-1β and IL-18, and the expression of iNOS. The mechanism was related to its inhibition of the NF-κB signaling pathway and the activation of NLRP3 inflammasome (Xu J. J. et al., 2021).
Polysacchadrides FVP and FFVP from Fermented Flammulina velutipes inhibited the secretion and expression of IL-1β, IL-6, IL-18 and TNF-β in colon tissue, and significantly decreased the expression of NLRP3, ASC, Caspase-1 and IL-1β protein in LPS- induced mice model, indicating the anti-inflammatory activity was related to inbibition of the NLRP3 signaling pathway (Ma et al., 2022). Polysaccharide from P. citrinopileatus (PCPS) inhibited the secretion of pro-inflammatory cytokines and chemokines by macrophages activated by LPS/INF-γ, and promoted the expression of anti-inflammatory cytokine IL-10. The anti-inflammatory effect was related to Dectin-1 and TLR2 receptors (Minato et al., 2019). There is growing evidence that certain miRNAs play key regulatory roles in macrophage activation and inflammation. miR-155 is closely related to the activation of NF-κB in macrophages, playing an important role in atherosclerosis by inhibiting B cells and promoting the activation of NF-κB in macrophages (Elton et al., 2013; Mann et al., 2017). Tremella fuciformis polysaccharides (TFPS) inhibited the inflammatory response of LPS-induced macrophages by inhibiting the expression of miR155 and the activation of NF-κB (Ruan et al., 2018). Figure 2 summarized the MAPK, NF-κB and JAK-STAT signaling pathways involved in the anti-inflammatory activity of edible fungi polysaccharides.
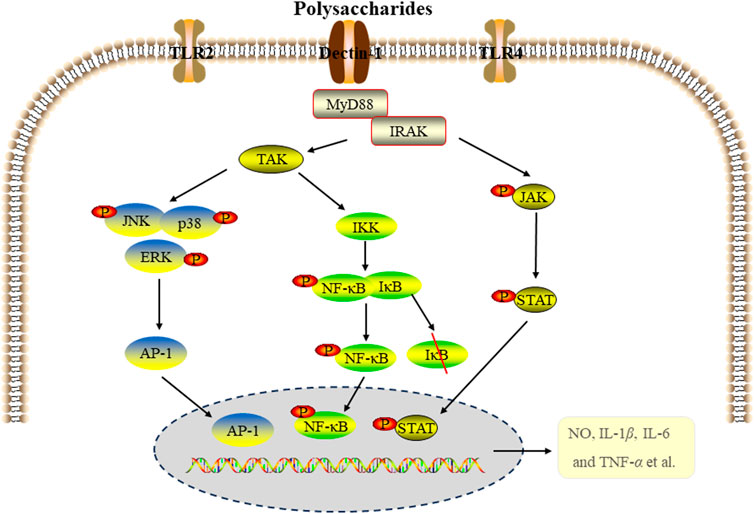
Figure 2. The signal pathways of anti-inflammatory activity of edible fungi polysaccharides.
Inflammatory bowel disease (IBD) is a chronic recurrent inflammatory disease, including crohn disease (CD) and ulcerative colitis (UC). Whlie the pathogenesis of IBD remains unclear, possible pathological mechanisms include immune response disorders, abnormal production of cellular inflammatory factors, impaired intestinal epithelial barrier function and disturbances of intestinal flora (Bisgaard et al., 2022). Edible fungi polysaccharides have been found to relieve intestinal inflammation by regulating the intestinal mucosal barrier. Both intestinal epithelial barrier function and inflammation play crucial roles in the occurrence and development of IBD. The barrier function of intestinal epithelium depends on the integrity of intestinal epithelial cells. Excessive apoptosis of these cells can compromise the barrier, allowing bacteria from the lumen to penetrate the intestinal wall and even bloodstream, potentially leading to septicemia and an inflammatory cascade (Chang, 2020). The intestinal mucosal barrier consists mainly of mechanical, biological, immune and chemical barriers, with the first three barriers being key components of intestinal mucosal immunity. The mechanical barrier is essential for maintaining the integrity of the intestinal mucosal barrier. The intestinal immune barrier involves complex interactions between immune cells and cytokines. The human gastrointestinal tract is colonized by rich and diverse microbial communities that form gut biological barrier and significantly influence the host’s physiology and health (Martínez et al., 2015; Gong et al., 2016). The effect of polysaccharides from edible fungi on inflammatory bowel disease was illustrated in Figure 3.
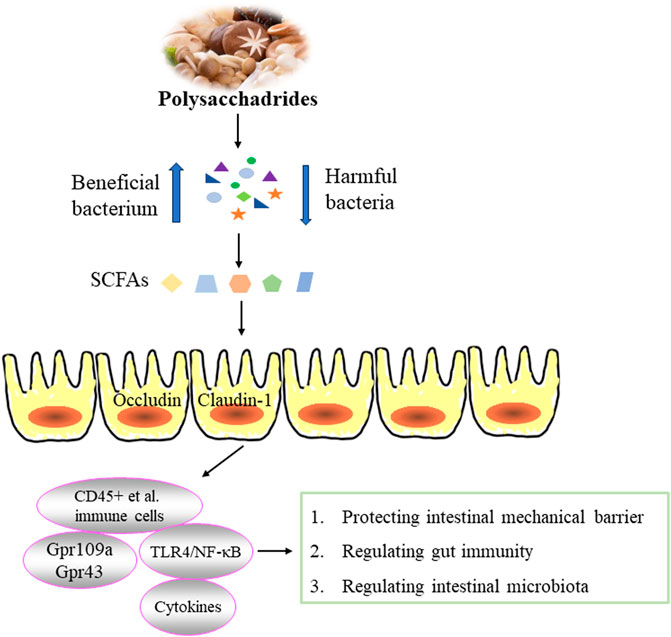
Figure 3. Effect of edible fungi polysaccharides on inflammatory bowel disease.
Intestinal epithelial cells (IECs) serve as physical and external barriers. Under specific conditions, IECs produce signal molecules such as cytokines to prevent pathogenic microorganisms from entering the intestinal tract. Research has demonstrated that LPS activate Toll-like receptors on IECs, thereby triggering the NF-κB transcription factor pathway, and leading to excessive production of TNF-α, IL-6 and IL-8, which can damage IECs (Omonijo et al., 2019; Kayama et al., 2020). Edible fungi polysaccharides exhibit protective effects on intestinal barrier. Polysaccharide from G. lucidum spores have been shown to mitigate paclitaxel-induced intestinal barrier injury by reversing microtubule polymerization and inhibiting apoptosis (Li D. et al., 2020). Furthermore, a water-soluble polysaccharide (GLSP) from G. lucidum demonstrated potent anti-inflammatory activity by inhibiting the excessive production of NO, IL-6 and IL-1β in IEC-6 cells induced by LPS, suggesting the potential of GLSP in protecting the gut barrier (Wen et al., 2022).
The host immune system also plays a crucial role in the development of IBD. Previous studies have shown that EP-1 effectively alleviates symptoms in acetic acid-induced UC rats by reducing IL-1 and IL-6 levels, increasing superoxide dismutase (SOD) levels and decreasing malondialdehyde (MDA) levels. It also lowers complement 3 (C3) and IgM levels, demonstrating anti-inflammatory, antioxidant and immune-enhancing activities (Shao et al., 2019). WPEP has been found to significantly reverse symptoms of colitis induced by dextran sulfate sodium (DSS) in mice. It reduces the concentration of pro-inflammatory cytokines and the expression of pro-inflammatory proteins, increases colon length, improves histology. These effects are associated with reduced accumulation of CD45+ immune cells, CD45+F4/80 + macrophages and CD45+ Gr1+ neutrophils (Ma et al., 2021). Studies have proved that Foxp3+T cells can inhibit inflammation and the production of IgA. T. fuciformis polysaccharides (TPs) have shown a protective effect against DSS-induced colitis in mice by regulating immune system. This effect involves reducing the Foxp3+T cells and IgA-coated bacteria, decreasing pro-inflammatory cytokines, and enhancing anti-inflammatory cytokines (Xu Y. et al., 2021).
Due to the absence of carbohydrate-active enzymes, most polysaccharides cannot be directly digested or absorbed by the body. Instead, their primary activity occurs through the fermentation by intestinal microorganisms (Kong et al., 2016; Ma et al., 2017). Abnormal changes in intestinal microflora can induce inflammation and exacerbate various inflammatory diseases (Schippa and Conte, 2014; Zuo and Ng, 2018). Edible fungi polysaccharides have been shown to regulate intestinal health by modulating intestinal microorganisms. For example, Scorias spongiosa polysaccharides SSPs have demonstrated a capacity to decrease the levels of IL-1β, IL-6 and TNF-α, while increasing IL-10 level to enhance anti-inflammation abilitiy. They also alter microbial community and composition by increasing the abundances of Firmicutes, Campilobacterota, Desulfobacterota, Proteobacteria, Actinobacteria, and Fusobacteria, Bacteroidetes, and Verrucomicrobia, while decreasing Verrucomicrobiota, Bacteroidota, Patescibacteria, and Synergistota in C57BL/6J mice (Xu Y. et al., 2022). Lentinan have been observed to inhibit the expression of cytokines (TGF-β, TNF-α, IL-1β, IL-6 and IL-8), attenuate IκBα degradation in LPS-induced inflammatory response in juvenile taimen intestine. Additionally, Lentinan increase the relative abundance of beneficial bacteria such as Lactobacillaceae, Lachnospiraceae and Ruminococcaceae while reducing harmful bacteria like Enterobacteriaceae, Fusobacteriaceae and Flavobacteriaceae. These effects suggest that the anti-inflammatory properties may correlate with NF-κB signaling pathway and improvement of intestinal microflora (Ren et al., 2019). The inhibitory effect of WPEP on DSS-induced colitis in mice is closely associated with intestinal microflora imbalance. It partially reverses this imbalance by decreasing the abundance of Ackermanella myxophilus and Clostridium cocleatum, while increasing Bifidobacterium pseudocolon, Lactobacillus, Lactobacillus saliva and Bromotococcus abundance (Ma et al., 2021). Similarly, TPS has shown significant ability to increase intestinal community diversity and restore the relative abundance of Lactobacillus, Odoribacter, Helicobacter, Ruminococcaceae, and Marinifilaceae in DSS-induced colitis in mice (Xu Y. et al., 2021).
Polysaccharides serve as an energy source for intestinal microorganisms, promote their proliferating and the production of beneficial compounds. The metabolization of polysaccharides by intestinal flora generates short-chain fatty acids (SCFAs), which possess immunomodulatory and anti-inflammatory activities critical for maintaining intestinal homeostasis, regulating immune function, and mitigating intestinal inflammation (Venegas et al., 2019). SCFAs help maintain a low pH environment that inhibits pathogen growth and stimulates the growth of butyric acid-producing bacteria, thereby reinforcing intestinal immune barriers (Gonçalves and Martel, 2016). For example, G. lucidum polysaccharide (GLP) have shown potential in alleviating DSS-induced colitis in mice by increasing SCFAs-producing bacteria, reducing pathogens in the small intestine and cecum, and enhancing SCFAs production (acetic acid, propionic acid and butyric acid). GLP also regulated the expression of genes involved in six inflammation-related KEGG pathways, thereby enhancing immunity, reducing inflammatory response and potentially lowering the risk of colon cancer (Xie et al., 2019). Similarly, the starch-free β-type glycosidic polysaccharide FVP from F. velutipes exerted protective effects in DSS-induced UC mice. FVP regulated the relative mRNA expression of cytokines (TNF-α, IFN-γ, IL-1β, IL-6, MCP-1 and MIP-1α) and tight junction proteins (caludin-1, occluding and zonulae occludens-1), modified intestinal microflora, and increased levels of acetic acid, propionic acid and n-butyric acid (Zhao et al., 2020). Administration of FVP (50 mg/kg and 100 mg/kg) reduced inflammatory response in DSS-induced colitis, significantly inhibited myeloperoxidase (MPO) activity, decreased levels of DAO and NO, and effectively restored the metabolic balance of intestinal microorganisms, especially promoting butyric acid production. These actions contribute to down-regulating the Toll-like receptor 4 (TLR4)/NF-κB inflammatory signaling pathway, thus improving colitis symptoms (Zhang et al., 2020).
SCFAs exert their immunomodulatory and anti-inflammatory effects by inhibiting histone deacetylases and activating G protein-coupled receptors (Gpr) such as Gpr43 and Gpr109a on the surface of intestinal epithelial cells and immune cells (Kim, 2018). These actions are crucial for maintaining intestinal homeostasis and controlling inflammation. For example, EP-1 has been shown to effectively alleviate acetic acid-induced UC in rats by regulating intestinal microflora. EP-1 also increased the relative proportions of acetic acid and butyric acid in feces and inhibited the expression of Gpr41 and Gpr43 (Shao et al., 2019). Similarly, the heteropolysaccharide FVP from F. velutipes demonstrated anti-inflammatory properties by inhibiting intestinal inflammation, regulating intestinal permeability, and reducing intestinal injury. FVP also prevented the downregulation of tight junction genes (Occludin and Claudin-1) and Gpr43 and Gpr109a induced by Cd. Moreover, PVP altered intestinal flora composition, and enhanced SCFAs production. These mechanisms collectively contributed to FVP’s ability to mitigate CdCl2-induced intestinal toxicity and damage by modulating SCFA-mediated intestinal inflammation and energy metabolism related to intestinal microbiota (Hao et al., 2023).
A caspase-independent form programmed cell death known as necroptosis has been identified as playing a significant role in in the pathogenesis of IBD (Rosenbaum et al., 2010). Polysaccharides extracted from L. edodes have shown dose-dependent inhibition of DSS-induced colitis in mice and have been observed to suppress necrotic cell death in Caco-2 cells. Notably, these polysaccharides exerted pronounced inhibitory effect on the necroptosis signaling cascade involving receptor-interacting protein kinase receptor-interacting protein kinase 1/receptor-interacting protein kinase 3/mixed kinase-region-like proteins (RIPK1-RIPK3-MLKL), resulting in decreased levels of phosphorylated MLKL in colitis mice. This inhibition of necroptotic cell death in the colon might contribute to the anti-inflammatory effects of L. edodes polysaccharides (Alagbaos and Mizuno, 2021). Furthermore, L. edodes polysaccharides have been shown to mitigate weight loss and reduce the expression of pro-inflammatory cytokines such as TNF-α, IL-6, IL-1β and IFN-γ in DSS-induced colitis in mice, suggesting their potential therapeutic efficacy in colitis treatment. Additionlly, evidence indicated that necroptosis might be linked to the expression of pro-inflammatory cytokines, further underscoring the anti-inflammatory mechanisms (Alagbaoso and Mizuno, 2022).
The Liver, a crucial organ in the human body, is susceptible to damage from toxic substances and drugs (Asrani et al., 2019). In recent years, chemical-induced liver injury has become increasingly significant in term of its impact on human health (Meng et al., 2018b). Studies have consistently shown that acute liver injury is often involved excessive oxidative stress and inflammatory responses (Dai et al., 2021; Yan et al., 2023). Tetrachloromethane (CCl4) is well-known environmental toxin widely used to induce experimental liver damage due to its ability to induce oxidative stress and inflammation (Liu et al., 2017; Wang et al., 2022). Edible fungi polysaccharides have demonstrated protective effects against CCl4-induced liver injury. For example, Morchella importuna polysaccharides (Mw 35.54 kDa) and F. velutipes polysaccharides (FVPs) have shown to mitigate CCl4-induced liver injury by enhancing antioxidant defenses and reducing inflammatory (Xu et al., 2021; Xu Y. et al., 2022; Xu et al., 2022 Y. Y.). P. ostreatus polysaccharide POP has exhibited hepatoprotective effects against CCl4-induced acute lung injury (ALI), attributed to its antioxidant properties that regulate metabolic pathway disorders and mitigate liver mitochondrial apoptosis (Zhu et al., 2019). Additionally, phosphorylated POP (PPOP) has shown stronger protective effects compared to POP, possibly through scavenging free radicals, preventing lipid peroxidation, and enhancing the endogenous antioxidant defense system (Duan et al., 2020). Abnormal activation of the NLRP3 inflammasome, leading to the release of pro-inflammatory cytokines, played a significant role in various types of ALI (Shao et al., 2020; Yu et al., 2023). G. lucidum polysaccharides (GLPS) have also demonstrated anti-inflammatory and hepatoprotective effects against CCl4-induced liver injury by inhibiting the inhibiting of NLRP3 activation, reducing inflammation, and inhibiting lipid peroxidation induced by free radicals (Chen et al., 2019).
Certainly, various factors such as alcohol, DSS and LPS can cause liver injury, with oxidative stress and inflammation being pathological mechanisms. For example, a heteropolysaccharide (EPS) from P. geesteranus has been shown to protect against alcohol-induced liver injury. The protective effect is likely achieved through enhancing antioxidant defenses and reducing anti-inflammatoion (Song et al., 2018). Similarly, a hepatoprotective polysaccharide PSP-1b1 (80 and 160 mg/kg/day) from Coriolus versicolor demonstrated protective effects against alcohol-induced liver injury by mitigating oxidative stress and modulating immune responses (Wang et al., 2019). M. esculenta polysaccharides have also bene studied for their protective effects against DSS-induced liver injury, primarily attributed to their ability to reduce oxidative stress, inhibit inflammation and enhance the activity of liver antioxidant enzymes (Chen S. T. et al., 2023). Additionally, GFP has shown efficacy in reducing liver injury induced by LPS/D-galactosamine (D-GalN) in mice. Its protective mechanism involved antioxidant defense and anti-inflammatory effects, likely mediated through inhibition of the miR-122/nuclear factor erythroid 2-related factor 2 (Nrf2)/anti-oxidative response element (ARE) pathway (Meng et al., 2021).
Dysregulation of gut microbes can compromise the integrity of intestinal barrier, allowing translocated bacteria and intestinal by products to enter the liver through the portal vein. This process increases oxidative stress and inflammation in the liver, posing a threat to live health and function. Increasing evidence suggests that gut microbes play a crucial role in protective effects of natural products on liver health (Meng et al., 2018a). For example, FVPs has been shown to alter the composition of intestinal microflora. It regulated bacterial pathways involved in fatty acid biosynthesis, tryptophan metabolism and exogenous metabolism via cytochrome P450, thereby protecting the liver from the toxic effects of CCl4 (Xu Y. Y. et al., 2022). Similarly, Coprinus comatus polysaccharide (CCP) has demonstrated the ability to modify the structure of intestinal flora. It inhibited the proliferation of Clostridium perfringens, Enterobacteriaceae and Enterococcus, while promoting the growth of Lactobacillus and Bifidobacterium in the gut of ALD mice. This modulation contributed to its anti-alcoholic liver injury effects (Yu et al., 2024). The relevant hepatoprotective effect was shown in Figure 4.
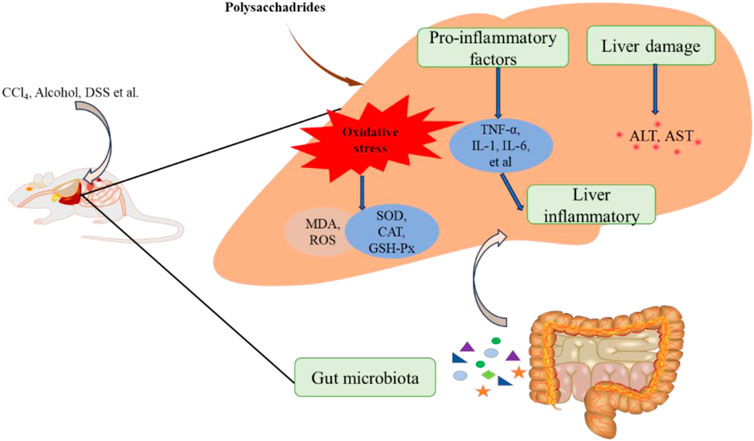
Figure 4. The hepatoprotective effect of edible fungi polysaccharides.
Since the outbreak of COVID-19 in 2019, there have been more than 500 million clinically confirmed cases worldwide. The outbreak of COVID-19 has led to significant increase in the number of acute pneumonia cases (Song P. P. et al., 2020). Acute pneumonia is a respiratory disease characterized by diffuse inflammatory lung injury, which can be caused by lung tissue contusion, bacterial, or virus infections. Activation of immune cells and excessive release of pro-inflammatory mediators are fundamental aspects of its pathogenesis (Bakowitz et al., 2012; He et al., 2021). LPS, as an endotoxin, is used to induce acute lung injury, triggering robust inflammation and immune response. Polysaccharides of edible fungi have shown protective effects against LPS-induced lung injury. For instance, G. lucidum polysaccharides (GLP, 25, 50 and 100 mg/kg/d) inhibited inflammatory cells infiltration, reduced the release of granulocyte macrophage-colony stimulating factor (GM-CSF) and IL-6, and decreased gene expression of IL-1β, IL-6, TNF-α and serum amyloid A3 (Saa3). Additionally, GLP inhibited neuropilin-1 (Nrp1) activation, upregulated B-cell lymphoma-2/Bcl-2-associated X protein (Bcl2/Bax) and Lc3 levels, and downregulated C-Caspase3/Caspase3 and p62 expression. These actions suggested that GLP protected against pneumonia by blocking inflammatory cells infiltration, suppressing cytokines secretion, inhibiting Nrp1 activation, regulating alveolar cell apoptosis, and modulating autophagy (Zhang X. L. et al., 2022). Residue polysaccharides (RPS) from L. edodes residue powder and its hydrolysates, acidic-RPS (ARPS) and enzymatic-RPS (ERPS), also showed lung protective effects in LPS-induced lung injured in mice. ERPS showed superior effects by reducing lung wet-to-dry weight ratio, inflammatory factors (TNF-α, IL-6, and IL-1β), complement C3 (C3) and high-sensitivity C-reactive protein (hs-CRP), while mitigating oxidative stress (Ren et al., 2018). L. edodes polysaccharides (PLE), primarily composed of Glc, Gal, GlcA, and Man, protected lung tissue by reducing hs-CRP and C3 levels, inhibiting gamma-glutamyltransferase (GGT) activity, decreasing TNF-α, IL-1β and IL-6 levels, and enhancing antioxidant enzymes SOD and catalase (CAT) activities (Zhang Y. W. et al., 2022).
Inhalation of fine particulate matter (PM2.5) can induce systemic inflammation, thereby increasing the risk of lung injury (Yan et al., 2017). Alveolar macrophages, upon encountering PM2.5 particles in the lungs, become activated and release cytokines and chemokines that recruit inflammatory cells to the lung, leading to inflammation (Bekki et al., 2016). NF-κB is a key transcription factor dimer that regulates the expression of pro-inflammatory cytokines such as TNF-α and IL-1β, and it is central to the pathogenesis of PM2.5-induced lung disease (Li et al., 2018). Sulfated polysaccharide from M. esculenta (SFMP-1) has shown protective effects against cell death, apoptosis, production of TNF-α and IL-1β in rat alveolar macrophages (NR8383 cells) induced by PM2.5, Its mechanism was involved inhibition of NF-κB activation (Li et al., 2019). PM2.5 exposure also induces antioxidant damage. Trametes orientalis polysaccharide (TOP-2) attenuated PM2.5-induced lung injury in mice through its antioxidant and anti-inflammatory effects, partly mediated by activating Nrf2/Heme oxygenase-1 (HO-1) pathway and inhibiting NLRP3 inflammasome (Zheng et al., 2019). The relevant protective effect was shown in Figure 5.
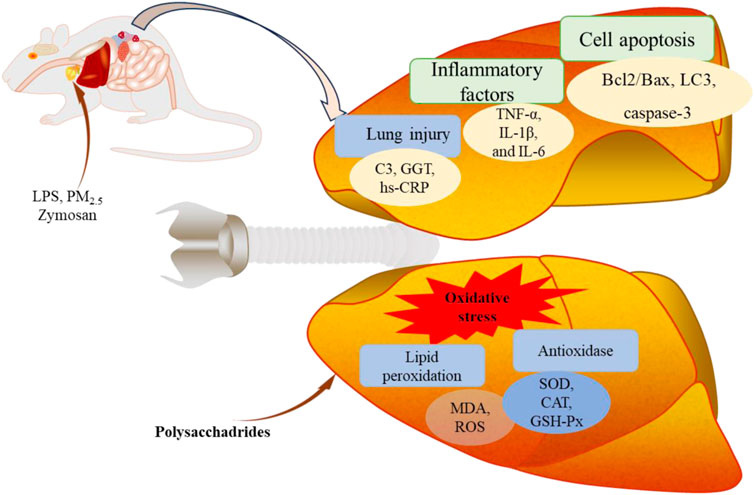
Figure 5. The protective effect of edible fungi polysaccharides on lung injury.
Obesity is characterized by chronic low-grade inflammation, and the relationship between chronic inflammation and obesity has been extensively studied. Two polysaccharides (CPA-1 and CPB-2) isolated from C. cicadae have been found to possess protective effects on HFD/HF-induced insulin resistance, metabolic abnormalities, hepatic oxidative stress (MDA, GSH-Px, SOD and CAT) and inflammatory response (TNF-α、IL-1β and IL-6) (Zhang et al., 2021). In mice fed a high-fat diet, water-soluble glucan from Grifola frondosa (GFPA) has demonstrated the ability to mitigate liver steatosis and inflammation, along with promoting significant weight loss. These beneficial effects were attributed its inhibition of chronic inflammation through the TLR4/NF-kB signaling pathway (Jiang et al., 2022).
Intestinal microflora plays a crucial role in the onset and progression of obesity by influencing host energy metabolism, substrate metabolism, and inflammation. Dictyophora indusiata polysaccharides have been shown to exert anti-obesogenic and anti-inflammatory effects by modulating the intestinal microbiome and inflammatory pathways in mice HFD-fed diet (Kanwal et al., 2020). Polysaccharide extracted from the sporoderm-broken spores of G. lucidum (BSGLP) has demonstrated significant reduction in fat accumulation, liver steatosis, inflammation, and hyperlipidemia in high-fat diet-fed mice. Its effects were believed to involve regulation of the intestinal microflora, enhancement of intestinal barrier function, promotion of SCFAs production, activation of GPR43 and inhibition of TLR4/myeloid differentiation factor 88 (MYD88)/NF-κB signaling pathway (Sang et al., 2021). Polysaccharide from Agrocybe cylindracea (ACP) has been found to ameliorate obesity in high-fat diet-induced obese mice by significantly reducing the levels of obesity-related TNF-α and IL-6. This effect partly results from decreasing the abundance of Desulfovibrio and increasing the abundance of Parabacteroides, along with related changes in solaventivone levels (Zhu et al., 2022).
Edible fungi polysaccharides have demonstrated potential in inhibiting periodontal inflammation, asthma, and providing renal protection. For instance, crude polysaccharides (CGLPs) from G. lucidum sourced from Changbai Mountain were found to regulate the expression of IL-1β, TNF-α and IL-10 in a concentration-dependent manner, effectively inhibiting alveolar bone loss caused by periodontitis (Chen Z. et al., 2023). In the context of allergic asthma, C. militaris polysaccharide CMP has been shown by Song et al. to mitigate oxidative stress and inflammation in mice with allergic asthma. The effect was achieved through the activating of NRF2/HO-1 signaling pathway and inhibiting of NF-κB signaling pathway. Importantly, these mechanisms were closely linked to maintaining the stability of intestinal microflora, highlighting the multifaceted protective roles of polysaccharides from edible fungi (Song et al., 2023).
ASMCP extracted from spent mushroom compost of L. edodes has been shown to decrease the levels of TNF-α, IL-6 and IL-1β, demonstrating potential anti-inflammatory activity on LPS-induced renal injury in mice (Song et al., 2020b). C. cicadae polysaccharide CCP has been reported to alleviate renal injury and tubulointerstitial fibrosis in rats with high-fat diet and STZ-induced diabetic nephropathy. This effect was achieved by CCP inhibiting the TLR4/NF-κB and TGF-β1/Smad signaling pathways, thereby reducing inflammatory reactions and modulating intestinal microflora (Yang et al., 2020). Additionally, crude polysaccharides from Floccularia luteovirens (FLPs) have shown effectiveness in improving renal tissue injury induced by high glucose. They targeted and regulated phosphorylated glycogen synthase kinase3β (GSK-3β), and inhibited the accumulation of inflammatory factors, highlighting their potential in renal protection (Wang et al., 2023).
The biological activities of polysaccharides are decisively influenced by their structures and physicochemical properties. Key factors include molecular weight, monosaccharide composition, type of linkage, degree of branching, conformation and solubility, all of which contribute to their anti-inflammatory activities. These structural attributes determine how polysaccharides interact with biological systems, influencing their efficacy in modulating immune responses and inflammatory processes. Understanding these structural features are crucial for elucidating and optimizing the therapeutic potential of polysaccharides in various health applications.
Research indicates that the specific glucans’ recognition and binding by immune cells can lead to subsequent immunomodulatory and anti-inflammatory activities (Camilli et al., 2018; Nakashima et al., 2018). Factors such as molecular weight and connection mode of polysaccharides are crucial in determining their receptor binding properties and anti-inflammatory activities. For example, the high molecular weight fraction of G. frondosa (GF70-F1, 1,260 kDa) has been shown to inhibit TNF-α and IL-6 production, along with NF-κB activation in LPS-induced RAW264.7 cells. This activity was likely mediated through interaction with TLR2 receptors rather than Dectin-1 or CR3 receptors due to its (1→6)-branched (1→4)-β-D-glucan structure (Su et al., 2020). In another study, polysaccharides (wHEP-1, wHEP-2 and wHEP-3) isolated from the mycelium of Hericium erinaceus demonstrated varied anti-inflammatory activities, with the high molecular weight wHEP-1 exhibiting the most potent effects in LPS-induced Caco-2 cells and a rat model (Wang et al., 2021). Similarly, β-Glucan H6PC20 (2,390 kDa) and α-heteropolysaccharide HPB-3 (15 kDa) isolated from H. erinaceus showed protective effects against alcoholic gastric ulcer in rats, with HPB-3 focusing specifically on anti-inflammation actions (Chen et al., 2020). Furthermore, Ma et al. (2020) have highlighted that low molecular weight FFVP (15,702 Da) exhibited was higher anti-inflammatory ability than that of high molecular weight FVP (15,961 Da). These findings underscore the importance of polysaccharide structural characteristics in influencing their biological activities, particularly in modulating inflammation through various receptor interactions.
The conformations of polysaccharides play significant roles in their anti-inflammatory activities. Yang et al., confirmed this by studying water-soluble indigestible polysaccharide NDPs (24.4 kDa) from C. cicadae. They found that NDPs laking a triple helix conformation strongly inhibited the production of NO, IL-1β and TNF-α by LPS-stimulated RAW264.7 macrophages compared to the crude polysaccharide CP (3.1 kDa, 21.5 kDa, 678.2 kDa) that possessed a triple helix conformation (Yang C. H. et al., 2019). This suggests that the absence of a triple helix conformation in NDPs enhances their anti-inflammatory effectiveness, highlighting the importance of polysaccharide conformation in influencing their biological activities.
The monosaccharide composition of polysaccharides indeed plays a significant role in their anti-inflammatory activities. For example, in the case of polysaccharides derived from the residue of L. edodes (RPS, ARPS, and ERPS), they have shown notable pulmonary protective effects. Among these, ERPS demonstrated superior efficacy, with Rha presumed to be essential for its lung protective activity (Ren et al., 2018). This highlights the importance of specific monosaccharide compositions in influencing the biological properties and therapeutic potential of polysaccharides.
Chemical modification and the addition of new chemical groups can indeed enhance the activities of polysaccharides or impart them with new therapeutic properties. For example, in studies on polysaccharides from M. angusticeps Peck, phosphorylation (PMEP) and three acetylated polysaccharides Ac-PMEP1-3 were employed to modify their structures. Among these, Ac-PMEP3, with a highly branched structure, demonstrated stronger anti-inflammatory effects by inhibiting the excessive production of NO and TNF-α in LPS-induced RAW264.7 cells (Yang Y. X. et al., 2019). Similarly, acetylated polysaccharide AcPPS from P. ostreatus exhibited lung protective effects in zymosan-induced acute lung injury mice, with the mechanism linked to the NF-κB signaling pathway (Song et al., 2020c). Phosphorylated polysaccharides PMPS from P. djamor mycelia showed antioxidant, anti-inflammatory, and anti-fibrotic effects in adenine-induced chronic renal failure (CRF) mice, highlighting phosphorylation as an effective modification method (Li et al., 2021). Additionally, sulfated polysaccharide from L. edodes (SPLE), characterized by sulfation of its β-glucan structure, demonstrated anti-inflammatory effects in zymosan-induced multiple organ dysfunction syndrome in mice (Sun et al., 2021). These examples underscore how chemical modifications can enhance or alter the biological activities of polysaccharides, expanding their therapeutic potential in various disease contexts.
Polysaccharides derived from edible fungi have garnered significant scholarly interest due to their safety profile and promising biological activities. This paper comprehensively reviews their anti-inflammatory activities, mechanism of action, and the effects on related inflammatory diseases. Special emphasis is placed on exploring the relationship between polysaccharide structure and anti-inflammatory activity. By synthesizing current knowledge, this review aims to deepen our understanding of how edible fungi polysaccharides can be applied in inflammation-related contexts. Ultimately, this research is expected to catalyze advancements in the development and utilization of edible fungi for therapeutic purposes.
Edible fungi polysaccharides exert their anti-inflammatory effects by inhibiting the release of relevant factors through interactions with membrane receptors and suppression of specific signaling pathways. Additionally, they can mitigate inflammation by enhancing intestinal mechanical and intestinal immune barriers, as well as modulating intestinal microorganisms. These polysaccharides have shown potential in addressing inflammation-related conditions such as liver injury, obesity, asthma, glomerulonephritis, and periodontitis. However, most research on their anti-inflammatory activities has been conducted in cell cultures or animal models, which may not fully reflect their effects in humans. Therefore, further clinical studies are needed to validate their therapeutic potential in human applications. The precise mechanisms underlying the anti-inflammatory actions of edible fungi polysaccharides remain incompletely understood. It is anticipated that future research will uncover additional signaling pathways and molecular targets relevant to inflammation. Structure-activity relationship analyses indicate that the structural characteristics of polysaccharides significantly influence their anti-inflammatory activities. Additionally, anti-inflammatory activities of polysaccharides also depend on the degradation, absorption and utilization processes. The complex structure of polysaccharides allows them to evade the action of human digestive enzymes, thereby forming a specific digestive pattern. Therefore, understanding the metabolic process of polysaccharides is of great significance for exploring the benefits and scientific applications of polysaccharides on host health. However, the complexity of polysaccharide structures presents challenges in fully elucidating these relationships. Therefore, there is a critical need for continued investigation into the structure-function dynamics of polysaccharides, with a focus on structural modifications to optimize their therapeutic efficacy. This approach holds promise for maximizing the beneficial effects of edible fungi polysaccharides in combating inflammation and advancing their clinical applications.
ZY: Funding acquisition, Writing–original draft, Writing–review and editing. JZ: Supervision, Writing–original draft. JQ: Writing–original draft. LG: Writing–review and editing. QG: Investigation, Writing–review and editing. WK: Conceptualization, Funding acquisition, Supervision, Writing–review and editing. CM: Funding acquisition, Supervision, Writing–review and editing. LC: Conceptualization, Supervision, Writing–review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This research was supported by Major Public Welfare Projects in Henan Province (201300110200), Key Research and Development Project of Henan Province (221111110600), Key Project in Science and Technology Agency of Henan Province (232102110140), National Natural Science Foundation of China (32202055), National Nand Innovation and Intelligence introduction Base of Precision Nutrition and Health Food Research of Henan Province (CXJD2021006), National College Student Innovation and Entrepreneurship Project (202311834009).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2024.1447677/full#supplementary-material
Alagbaos, C. A., and Mizuno, M. (2021). Polysaccharides from shiitake culinary-medicinal mushroom Lentinus edodes (Agaricomycetes) suppress pMLKL-mediated necroptotic cell death and colitis in mice. Int. J. Med. Mushrooms 23, 13–26. doi:10.1615/IntJMedMushrooms.2021038850
Alagbaoso, C. A., and Mizuno, M. (2022). Lentinula edodes polysaccharides suppressed pro-inflammatory cytokines expression and colitis in mice. Arq. Gastroenterol. 59, 288–295. doi:10.1590/S0004-2803.202202000-51
Andaluz, A., Yeste, M., Rodríguez-Gil, J., Rigau, T., García, F., and del Álamo, M. M. R. (2016). Pro-inflammatory cytokines: useful markers for the diagnosis of canine mammary tumours. Vet. J. 210, 92–94. doi:10.1016/j.tvjl.2016.01.016
Asrani, S. K., Devarbhavi, H., Eaton, J., and Kamath, P. S. (2019). Burden of liver diseases in the world. J. Hepatol. 70, 151–171. doi:10.1016/j.jhep.2018.09.014
Bakowitz, M., Bruns, B., and McCunn, M. (2012). Acute lung injury and the acute respiratory distress syndrome in the injured patient. Scand. J. Trauma. Resusc. Emerg. Med. 20, 54–63. doi:10.1186/1757-7241-20-54
Barbu, E., Popescu, M. R., Popescu, A. C., and Balanescu, S. M. (2022). Inflammation as a precursor of atherothrombosis, diabetes and early vascular aging. Int. J. Mol. Sci. 23, 963. doi:10.3390/ijms23020963
Bekki, K., Ito, T., Yoshida, Y., He, C. Y., Arashidani, K., He, M., et al. (2016). PM2.5 collected in China causes inflammatory and oxidative stress responses in macrophages through the multiple pathways. Environ. Toxicol. Pharmacol. 45, 362–369. doi:10.1016/j.etap.2016.06.022
Bisgaard, T. H., Allin, K. H., Keefer, L., Ananthakrishnan, A. N., and Jess, T. (2022). Depression and anxiety in inflammatory bowel disease: epidemiology, mechanisms and treatment. Nat. Rev. Gastroenterol. Hepatol. 19, 717–726. doi:10.1038/s41575-022-00634-6
Camilli, G., Tabouret, G., and Quintin, J. (2018). The complexity of fungal β-glucan in health and disease: effects on the mononuclear phagocyte system. Front. Immunol. 9, 673. doi:10.3389/fimmu.2018.00673
Chang, C. C., Cheng, J. J., Lee, I. J., and Lu, M. K. (2018). Purification, structural elucidation, and anti-inflammatory activity of xylosyl galactofucan from Armillaria mellea. Int. J. Biol. Macromol. 114, 584–591. doi:10.1016/j.ijbiomac.2018.02.033
Chang, J. T. (2020). Pathophysiology of inflammatory bowel diseases. N. Engl. J. Med. 383, 2652–2664. doi:10.1056/NEJMra2002697
Chen, H. Y., Lei, J. Y., Li, S. L., Guo, L. Q., Lin, J. F., Wu, G. H., et al. (2023). Progress in biological activities and biosynthesis of edible fungi terpenoids. Crit. Rev. Food Sci. Nutr. 63, 7288–7310. doi:10.1080/10408398.2022.2045559
Chen, S. T., Wang, M., Veeraperuma, S., Teng, B., Li, R., Qian, Z. M., et al. (2023). Antioxidative and protective effect of Morchella esculenta against dextran sulfate sodium-induced alterations in liver. Foods 12, 1115. doi:10.3390/foods12051115
Chen, W. C., Wu, D., Jin, Y. L., Li, Q. Z., Liu, Y. F., Qiao, X. X., et al. (2020). Pre-protective effect of polysaccharides purified from Hericium erinaceus against ethanol-induced gastric mucosal injury in rats. Int. J. Biol. Macromol. 159, 948–956. doi:10.1016/j.ijbiomac.2020.05.163
Chen, Y. S., Chen, Q. Z., Wang, Z. J., Hua, C., et al. (2019). Anti-inflammatory and hepatoprotective effects of Ganoderma lucidum polysaccharides against carbon tetrachloride-induced liver injury in kunming mice. Pharmacology 103, 143–150. doi:10.1159/000493896
Chen, Z., Qin, W. G., Lin, H. B., Lin, Y., Tian, Y., Zhao, X. T., et al. (2023c). Inhibitory effect of polysaccharides extracted from Changbai mountain Ganoderma lucidum on periodontal inflammation. Heliyon 9, e13205. doi:10.1016/j.heliyon.2023.e13205
Chopra, H., Mishra, A. K., Baig, A. A., Mohanta, T. K., Yugal Kishore Mohanta, Y. K., and Kwang-Hyun Baek, K. H. (2021). Narrative review: bioactive potential of various mushrooms as the treasure of versatile therapeutic natural product. J. Fungi 7, 728. doi:10.3390/jof7090728
Dai, C. S., Li, H., Wang, Y., Tang, S. S., Velkov, T., and Shen, J. Z. (2021). Inhibition of oxidative stress and ALOX12 and NF-κB pathways contribute to the protective effect of baicalein on carbon tetrachloride-induced acute liver injury. Antioxidants 10, 976. doi:10.3390/antiox10060976
Du, B., Zhu, F. M., and Xu, B. J. (2018). An insight into the anti-inflammatory properties of edible and medicinal mushrooms. J. Funct. Foods 47, 334–342. doi:10.1016/j.jff.2018.06.003
Du, L., Li, J. K., Zhang, X. T., Wang, L. F., Zhang, W. M., Yang, M., et al. (2019). Pomegranate peel polyphenols inhibits inflammation in LPS-induced RAW264.7 macrophages via the suppression of TLR4/NF-κB pathway activation. Food Nutr. Res. 63, 1–12. doi:10.29219/fnr.v63.3392
Duan, Z., Zhang, Y., Zhu, C. P., Wu, Y., Du, B. Q., Ji, H. J., et al. (2020). Structural characterization of phosphorylated Pleurotus ostreatus polysaccharide and its hepatoprotective effect on carbon tetrachloride-induced liver injury in mice. Int. J. Biol. Macromol. 162, 533–547. doi:10.1016/j.ijbiomac.2020.06.107
Elton, T. S., Selemon, H., Elton, S. M., and Parinandi, N. L. (2013). Regulation of the MIR155 host gene in physiological and pathological processes. Gene 2532, 1–12. doi:10.1016/j.gene.2012.12.009
Gonçalves, P., and Martel, F. (2016). Regulation of colonic epithelial butyrate transport: focus on colorectal cancer. Porto. Biomed. J. 1, 83–91. doi:10.1016/j.pbj.2016.04.004
Gong, Y., Li, H., and Li, Y. (2016). Effects of Bacillus subtilis on epithelial tight junctions of mice with inflammatory bowel disease. J. Interferon Cytokine Res. 36, 75–85. doi:10.1089/jir.2015.0030
Hao, R. L., Zhou, X., Zhao, X. Y., Lv, X. Q., Zhu, X. Y., Gao, N. N., et al. (2023). Flammulina velutipes polysaccharide counteracts cadmium-induced gut injury in mice via modulating gut inflammation, gut microbiota and intestinal barrier. Sci. Total Environ. 877, 162910. doi:10.1016/j.scitotenv.2023.162910
He, Y. Q., Zhou, C. C., Yu, L. Y., Wang, L., Deng, J. L., Tao, Y. L., et al. (2021). Natural product derived phytochemicals in managing acute lung injury by multiple mechanisms. Pharmacol. Res. 163, 105224. doi:10.1016/j.phrs.2020.105224
Hu, W., Zhou, P. H., Rao, T., Zhang, X. B., Wang, W., and Zhang, L. J. (2015). Adrenomedullin attenuates interleukin-1β-induced inflammation and apoptosis in rat Leydig cells via inhibition of NFκB signaling pathway. Exp. Cell Res. 339, 220–230. doi:10.1016/j.yexcr.2015.10.024
Huang, M., Mei, X. D., and Zhang, S. (2011). Mechanism of nitric oxide production in macrophages treated with medicinal mushroom extracts (review). Int. J. Med. Mushrooms. 13, 1–6. doi:10.1615/intjmedmushr.v13.i1.10
Hung, Y. L., Wang, S. C., Suzuki, K., Fang, S. H., Chen, C. S., Cheng, W. C., et al. (2019). Bavachin attenuates LPS-induced inflammatory response and inhibits the activation of NLRP3 inflammasome in macrophages. Phytomedicine 59, 152785. doi:10.1016/j.phymed.2018.12.008
Jia, X. W., Ma, B. J., Xue, F., Xing, Y. Q., Wu, P., Li, F. C., et al. (2022). Structure characterization and anti-Inflammatory activity of polysaccharides from Lingzhi or Reishi medicinal mushroom Ganoderma lucidum (Agaricomycetes) by microwave-assisted freeze-thaw extraction. Int. J. Med. Mushrooms. 24, 49–61. doi:10.1615/IntJMedMushrooms.2022045268
Jiang, X., Hao, J., Zhu, Y. F., Liu, Z. J., Li, L. Z., Zhou, Y. L., et al. (2022). The anti-obesity effects of a water-soluble glucan from Grifola frondosa via the modulation of chronic inflammation. Front. Immunol. 13, 962341. doi:10.3389/fimmu.2022.962341
Kanwal, S., Aliya, S., and Xin, Y. (2020). Anti-obesity effect of Dictyophora indusiata mushroom polysaccharide (DIP) in high fat diet-induced obesity via regulating inflammatory cascades and intestinal microbiome. Front. Endocrinol. 11, 558874. doi:10.3389/fendo.2020.558874
Kayama, H., Okumura, R., and Takeda, K. (2020). Interaction between the microbiota, epithelia, and immune cells in the intestine. Annu. Rev. Immunol. 38, 23–48. doi:10.1146/annurev-immunol-070119-115104
Kim, C. H. (2018). Immune regulation by microbiome metabolites. Immunology 154, 220–229. doi:10.1111/imm.12930
Kong, Q., Dong, S., Gao, J., and Jiang, C. (2016). In vitro fermentation of sulfated polysaccharides from E. prolifera and L. japonica by human fecal microbiota. Int. J. Biol. Macromol. 91, 867–871. doi:10.1016/j.ijbiomac.2016.06.036
Li, B., Guo, L., Ku, T. T., Chen, M. J., Li, G. K., and Sang, N. (2018). PM2.5 exposure stimulates COX-2-mediated excitatory synaptic transmission via ROS-NF-κB pathway. Chemosphere 190, 124–134. doi:10.1016/j.chemosphere.2017.09.098
Li, D., Gao, L., Li, M. X., Luo, Y. Y., Xie, Y. Z., Luo, T., et al. (2020a). Polysaccharide from spore of Ganoderma lucidum ameliorates paclitaxel-induced intestinal barrier injury: apoptosis inhibition by reversing microtubule polymerization. Biomed. Pharmacother. 130, 110539. doi:10.1016/j.biopha.2020.110539
Li, H. P., Feng, Y. B., Sun, W. X., Kong, Y., and Jia, L. (2021). Antioxidation, anti-inflammation and anti-fibrosis effect of phosphorylated polysaccharides from Pleurotus djamor mycelia on adenine-induced chronic renal failure mice. Int. J. Biol. Macromol. 170, 652–663. doi:10.1016/j.ijbiomac.2020.12.159
Li, L. Q., Song, A. X., Yin, J. Y., Siu, K. C., Wong, W. T., and Wu, J. Y. (2020b). Anti-inflammation activity of exopolysaccharides produced by a medicinal fungus Cordyceps sinensis Cs-HK1 in cell and animal models. Int. J. Biol. Macromol. 149, 1042–1050. doi:10.1016/j.ijbiomac.2020.02.022
Li, W., Cai, Z. N., Mehmood, S., Liang, L. L., Liu, Y., Zhang, H. Y., et al. (2019). Anti-inflammatory effects of Morchella esculenta polysaccharide and its derivatives in fine particulate matter-treated NR8383 cells. Int. J. Biol. Macromol. 129, 904–915. doi:10.1016/j.ijbiomac.2019.02.088
Liang, J., Rao, Z. H., Jiang, S. L., Wang, S., Kuang, H. X., and Xia, Y. G. (2023). Structure of an unprecedent glucuronoxylogalactoglucomannan from fruit bodies of Auricularia auricula-judae (black woody ear). Carbohydr. Polym. 315, 120968. doi:10.1016/j.carbpol.2023.120968
Liu, J. M., Fu, Y. Y., Zhang, H., Wang, J. D., Zhu, J., Wang, Y. Q., et al. (2017). The hepatoprotective effect of the probiotic Clostridium butyricum against carbon tetrachloride-induced acute liver damage in mice. Food Funct. 8, 4042–4052. doi:10.1039/c7fo00355b
Liu, X. F., Wang, X. Q., Xu, X. F., and Zhang, X. W. (2019). Purification, antitumor and anti-inflammation activities of an alkali-soluble and carboxymethyl polysaccharide CMP33 from Poria cocos. Int. J. Biol. Macromol. 127, 39–47. doi:10.1016/j.ijbiomac.2019.01.029
Ma, G. M., Kimatu, B. M., Zhao, L. Y., Yang, W. J., Pei, F., and Hu, Q. H. (2017). In vivo fermentation of a Pleurotus eryngii polysaccharide and its effects on fecal microbiota composition and immune response. Food Funct. 8, 1810–1821. doi:10.1039/c7fo00341b
Ma, G. X., Hu, Q. H., Han, Y. H., Du, H. J., Yang, W. J., Pan, C., et al. (2021). Inhibitory effects of β-type glycosidic polysaccharide from Pleurotus eryngii on dextran sodium sulfate-induced colitis in mice. Food Funct. 12, 3831–3841. doi:10.1039/d0fo02905j
Ma, G. X., Kimatu, B. M., Yang, W. J., Pei, F., Zhao, L. Y., Du, H. J., et al. (2020). Preparation of newly identified polysaccharide from Pleurotus eryngii and its anti-inflammation activities potential. J. Food Sci. 85, 2822–2831. doi:10.1111/1750-3841.15375
Ma, S., Xu, J. X., Lai, T., Xu, W. N., Zhang, J., Zhang, H. C., et al. (2022). Inhibitory effect of fermented Flammulina velutipes polysaccharides on mice intestinal inflammation. Front. Nutr. 9, 934073. doi:10.3389/fnut.2022.934073
Maity, P., Sen, I. K., Chakraborty, I., Mondal, S., Bar, H., Bhanja, S. K., et al. (2021). Biologically active polysaccharide from edible mushrooms: a review. Int. J. Biol. Macromol. 172, 408–417. doi:10.1016/j.ijbiomac.2021.01.081
Mann, M., Mehta, A., Zhao, J. L., Lee, K., Marinov, G. K., Flores, Y. G., et al. (2017). An NF-κB-microRNA regulatory network tunes macrophage inflammatory responses. Nat. Commun. 8, 851. doi:10.1038/s41467-017-00972-z
Martínez, I., Stegen, J. C., Maldonado-Gómez, M. X., Eren, A. M., Siba, P. M., Greenhill, A. R., et al. (2015). The gut microbiota of rural papua new guineans: composition, diversity patterns, and ecological processes. Cell Rep. 28, 527–538. doi:10.1016/j.celrep.2015.03.049
Medzhitov, R. (2008). Origin and physiological roles of inflammation. Nature 454, 428–435. doi:10.1038/nature07201
Meng, M., Zhang, R., Han, R., Kong, Y., Wang, R. H., and Hou, L. H. (2021). The polysaccharides from the Grifola frondosa fruiting body prevent lipopolysaccharide/d-galactosamine-induced acute liver injury via the miR-122-Nrf2/ARE pathways. Food Funct. 12, 1973–1982. doi:10.1039/d0fo03327h
Meng, X., Li, S., Li, Y., Gan, R. Y., and Li, H. B. (2018a). Gut microbiota’s relationship with liver disease and role in hepatoprotection by dietary natural products and probiotics. Nutrients 10, 1457. doi:10.3390/nu10101457
Meng, X., Li, Y., Li, S., Gan, R. Y., and Li, H. B. (2018b). Natural products for prevention and treatment of chemical-induced liver injuries. Compr. Rev. Food Sci. Food Saf. 17, 472–495. doi:10.1111/1541-4337.12335
Minato, K. I., Laan, L. C., Die, I. v., and Mizuno, M. (2019). Pleurotus citrinopileatus polysaccharide stimulates anti-inflammatory properties during monocyte-to-macrophage differentiation. Int. J. Biol. Macromol. 122, 705–712. doi:10.1016/j.ijbiomac.2018.10.157
Mustafa, F., Chopra, H., Baig, A. A., Avula, S. K., Kumari, S., Mohanta, T. K., et al. (2022). Edible mushrooms as novel myco-therapeutics: effects on lipid level, obesity and BMI. J. Fungi 8, 211. doi:10.3390/jof8020211
Nakashima, A., Yamada, K., Iwata, O., Sugimoto, R., Atsuji, K., Ogawa, T., et al. (2018). β-Glucan in foods and its physiological functions. J. Nutr. Sci. Vitaminol. 64, 8–17. doi:10.3177/jnsv.64.8
Oh, H., Park, S. H., Kang, M. K., Kim, Y. H., Lee, E. J., Kim, D. Y., et al. (2019). Asaronic acid attenuates macrophage activation toward M1 phenotype through inhibition of NF-κB pathway and JAKSTAT signaling in glucose-loaded murine macrophages. J. Agric. Food Chem. 67, 10069–10078. doi:10.1021/acs.jafc.9b03926
Omonijo, F. A., Hui, L. S., Zhang, Q., Lahaye, H., Bodin, L., Yang, J. C., et al. (2019). Thymol improves barrier function and attenuates inflammatory responses in porcine intestinal epithelial cells during lipopolysaccharide (LPS)-induced inflammation. J. Agr. Food Chem. 67, 615–624. doi:10.1021/acs.jafc.8b05480
Panda, J., Mishra, A. K., Nath, P. C., Mahanta, S., Sharma, M., Nayak, P. K., et al. (2024). Wild edible mushrooms to achieve sustainable development goals: novel sources for food security, health, and well-being. Food Biosci. 60, 104277. doi:10.1016/j.fbio.2024.104277
Ren, G. M., Xu, L. M., Lu, T. Y., Zhang, Y. Q., Wang, Y. Y., and Yin, J. S. (2019). Protective effects of lentinan on lipopolysaccharide induced inflammatory response in intestine of juvenile taimen (Hucho taimen, Pallas). Int. J. Biol. Macromol. 121, 317–325. doi:10.1016/j.ijbiomac.2018.09.121
Ren, Z. Z., Li, J., Song, X. L., Zhang, J. J., Wang, W. S., Wang, X. X., et al. (2018). The regulation of inflammation and oxidative status against lung injury of residue polysaccharides by Lentinula edodes. Int. J. Biol. Macromol. 106, 185–192. doi:10.1016/j.ijbiomac.2017.08.008
Rosenbaum, D. M., Degterev, A., David, J., Rosenbaum, P. S., Roth, S., Grotta, J. C., et al. (2010). Necroptosis, a novel form of caspase-independent cell death, contributes to neuronal damage in a retinal ischemia-reperfusion injury model. J. Neurosci. Res. 88, 1569–1576. doi:10.1002/jnr.22314
Ruan, Y., Li, H., Pu, L. M., Shen, T., and Jin, Z. N. (2018). Tremella fuciformis polysaccharides attenuate oxidative stress and inflammation in macrophages through miR-155. Anal. Cell Pathol. (Amst). 2018, 5762371. doi:10.1155/2018/5762371
Sang, T. T., Guo, C. J., Guo, D. D., Wu, J. J., Wang, Y. J., Wang, Y., et al. (2021). Suppression of obesity and inflammation by polysaccharide from sporoderm-broken spore of Ganoderma lucidum via gut microbiota regulation. Carbohydr. Polym. 256, 117594. doi:10.1016/j.carbpol.2020.117594
Schindler, C., Levy, D. E., and Decker, T. (2007). JAK-STAT signaling: from interferons to cytokines. J. Biol. Chem. 282, 20059–20063. doi:10.1074/jbc.R700016200
Schippa, S., and Conte, M. P. (2014). Dysbiotic events in gut microbiota: impact on human health. Nutrients 6, 5786–5805. doi:10.3390/nu6125786
Sha, M., Gao, Y. R., Deng, C., Wan, Y. M., Zhuang, Y., Hu, X. C., et al. (2020). Therapeutic effects of AdipoRon on liver inflammation and fibrosis induced by CCl4 in mice. Int. Immunopharmacol. 79, 106157. doi:10.1016/j.intimp.2019.106157
Shamim, M. Z., Mishra, A. K., Kausar, T., Mahanta, S., Sarma, B., Kumar, V., et al. (2023). Exploring edible mushrooms for diabetes: unveiling their role in prevention and treatment. Molecules 28, 2837. doi:10.3390/molecules28062837
Shao, S., Wang, D. D., Zheng, W., Li, X. Y., Zhang, H., Zhao, D. Q., et al. (2019). A unique polysaccharide from Hericium erinaceus mycelium ameliorates acetic acid-induced ulcerative colitis rats by modulating the composition of the gut microbiota, short chain fatty acids levels and GPR41/43 respectors. Int. Immunopharmacol. 71, 411–422. doi:10.1016/j.intimp.2019.02.038
Solier, S., Müller, S., Cañeque, T., Versini, A., Mansart, A., Sindikubwabo, F., et al. (2023). A druggable copper-signalling pathway that drives inflammation. Nature 617, 386–394. doi:10.1038/s41586-023-06017-4
Song, L. Y., Yang, J. T., Kong, W. H., Liu, Y., Liu, S. Y., and Su, L. (2023). Cordyceps militaris polysaccharide alleviates ovalbumin-induced allergic asthma through the Nrf2/HO-1 and NF-κB signaling pathways and regulates the gut microbiota. Int. J. Biol. Macromol. 238, 124333. doi:10.1016/j.ijbiomac.2023.124333
Song, P. P., Li, W., Xie, J. Q., Hou, Y. L., and You, C. G. (2020a). Cytokine storm induced by SARS-CoV-2. Clin. Chim. Acta. 509, 280–287. doi:10.1016/j.cca.2020.06.017
Song, X. L., Liu, Z. H., Zhang, J. J., Yang, Q. H., Ren, Z. Z., Zhang, C., et al. (2018). Anti-inflammatory and hepatoprotective effects of exopolysaccharides isolated from Pleurotus geesteranus on alcohol-induced liver injury. Sci. Rep. 8, 10493. doi:10.1038/s41598-018-28785-0
Song, X. L., Ren, Z. Z., Wang, X. X., Jia, L., and Zhang, C. (2020b). Antioxidant, anti-inflammatory and renoprotective effects of acidic-hydrolytic polysaccharides by spent mushroom compost (Lentinula edodes) on LPS-induced kidney injury. Int. J. Biol. Macromol. 151, 1267–1276. doi:10.1016/j.ijbiomac.2019.10.173
Song, X. L., Zhang, J. J., Li, J., and Jia, L. (2020c). Acetylated polysaccharides from Pleurotus geesteranus alleviate lung injury via regulating NF-κB signal pathway. Int. J. Mol. Sci. 21, 2810. doi:10.3390/ijms21082810
Su, C. H., Lu, M. K., Lu, T. J., Lai, M. N., and Ng, L. T. (2020). A (1→6)-Branched (1→4)-β-d-Glucan from Grifola frondosa inhibits lipopolysaccharide-induced cytokine production in RAW264.7 macrophages by binding to TLR2 rather than dectin-1 or CR3 receptors. J. Nat. Prod. 83, 231–242. doi:10.1021/acs.jnatprod.9b00584
Sun, W. X., Feng, Y. B., Zhang, M. Y., Song, X. L., and Jia, L. (2021). Protective effects of sulfated polysaccharides from Lentinula edodes on the lung and liver of MODS mice. Food Funct. 12, 6389–6402. doi:10.1039/d1fo00399b
Sun, Y. J., He, H. Q., Wang, Q., Yang, X. Y., Jiang, S. J., and Wang, D. B. (2022). A review of development and utilization for edible fungal polysaccharides: extraction, chemical characteristics, and bioactivities. Polym. (Basel) 14, 4454. doi:10.3390/polym14204454
Varela, M. L., Mogildea, M., Moreno, I., and Lopes, A. (2018). Acute inflammation and metabolism. Inflammation 41, 1115–1127. doi:10.1007/s10753-018-0739-1
Venegas, D. P., Fuente, M. K. D. L., Landskron, G., González, M. J., Quera, R., Dijkstra, G., et al. (2019). Short chain fatty acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 10, 277. doi:10.3389/fimmu.2019.00277
Wang, B. L., Cui, S. M., Mao, B. Y., Zhang, Q. X., Tian, F. W., Zhao, J. X., et al. (2022). Supervision cyanidin alleviated CCl4-induced acute liver injury by regulating the Nrf2 and NF-κB signaling pathways. Antioxidants (Basel) 11, 2383. doi:10.3390/antiox11122383
Wang, D. D., Xu, D. D., Zhao, D. Q., and Wang, M. X. (2021). Screening and comparison of anti-intestinal inflammatory activities of three polysaccharides from the mycelium of lion’s mane culinary-medicinal mushroom, Hericium erinaceus (Agaricomycetes). Int. J. Med. Mushrooms. 23, 63–71. doi:10.1615/IntJMedMushrooms.2021039951
Wang, H., Yang, Y., Wang, S. M., Li, C. T., Chen, C. B., Wan, X. L., et al. (2023). Polysaccharides of Floccularia luteovirens alleviate oxidative damage and inflammatory parameters of diabetic nephropathy in db/db Mice. Front. Biosci. (Landmark Ed) 28, 82. doi:10.31083/j.fbl2804082
Wang, K. L., Lu, Z. M., Mao, X. J. Z., Chen, L., Gong, J. S., Ren, Y. L., et al. (2019). Structural characterization and anti-alcoholic liver injury activity of a polysaccharide from Coriolus versicolor mycelia. Int. J. Biol. Macromol. 137, 1102–1111. doi:10.1016/j.ijbiomac.2019.06.242
Wang, Y. H., and Zeng, K. W. (2019). Natural products as a crucial source of anti-inflammatory drugs: recent trends and advancements. Tradit. Med. Res. 4, 257–268. doi:10.53388/tmr20190831133
Wen, L. R., Sheng, Z. L., Wang, J. P., Jiang, Y. M., and Yang, B. (2022). Structure of water-soluble polysaccharides in spore of Ganoderma lucidum and their anti-inflammatory activity. Food Chem. 373, 131374. doi:10.1016/j.foodchem.2021.131374
Xie, J. L., Liu, Y. H. X., Chen, B. H., Zhang, G. W., Ou, S. Y., Luo, J. M., et al. (2019). Ganoderma lucidum polysaccharide improves rat DSS-induced colitis by altering cecal microbiota and gene expression of colonic epithelial cells. Food Nutr. Res. 63, 1559. doi:10.29219/fnr.v63.1559
Xu, J., Xiao, C. M., Xu, H. S., Yang, S. X., Chen, Z. M., Wang, H. Z., et al. (2021b). Anti-inflammatory effects of Ganoderma lucidum sterols via attenuation of the p38 MAPK and NF-κB pathways in LPS-induced RAW 264.7 macrophages. Food Chem. Toxicol. 150, 112073. doi:10.1016/j.fct.2021.112073
Xu, J. J., Gong, L. L., Li, Y. Y., Zhou, Z. B., Yang, W. W., Wan, C. X., et al. (2021a). Anti-inflammatory effect of a polysaccharide fraction from Craterellus cornucopioides in LPS-stimulated macrophages. J. Food Biochem. 45, e13842. doi:10.1111/jfbc.13842
Xu, Y., Xie, L., Zhang, Z., Zhang, W., Tang, J., He, X. L., et al. (2021d). Tremella fuciformis polysaccharides inhibited colonic inflammation in dextran sulfate sodium-treated mice via Foxp3+ T cells, gut microbiota, and bacterial metabolites. Front. Immunol. 12, 648162. doi:10.3389/fimmu.2021.648162
Xu, Y., Zhang, Z., Feng, H., Tang, J., Peng, W., Chen, Y., et al. (2022a). Scorias spongiosa polysaccharides promote the antioxidant and anti-Inflammatory capacity and its effect on intestinal microbiota in mice. Front. Microbiol. 13, 865396. doi:10.3389/fmicb.2022.865396
Xu, Y. Y., Xie, L. Y., Tang, J., He, X. L., Zhang, Z. Y., Chen, Y., et al. (2021c). Morchella importuna polysaccharides alleviate carbon tetrachloride-induced hepatic oxidative injury in mice. Front. Physiol. 12, 669331. doi:10.3389/fphys.2021.669331
Xu, Y. Y., Zhang, Z. Y., Wang, B., He, X. L., Tang, J., Peng, W. H., et al. (2022b). Flammulina velutipes polysaccharides modulate gut microbiota and alleviate carbon tetrachloride-induced hepatic oxidative injury in mice. Front. Microbiol. 13, 847653. doi:10.3389/fmicb.2022.847653
Yan, J. K., Wang, C., Chen, T. T., Li, L. Q., Liu, X. Z., and Li, L. (2023). Structural characteristics and ameliorative effect of a polysaccharide from Corbicula fluminea industrial distillate against acute liver injury induced by CCl4 in mice. Int. J. Biol. Macromol. 227, 391–404. doi:10.1016/j.ijbiomac.2022.12.138
Yan, J. Y., Lai, C. H., Lung, S. C. C., Chen, C. J., Wang, W. C., Huang, P. I., et al. (2017). Industrial PM2.5 cause pulmonary adverse effect through RhoA/ROCK pathway. Sci. Total Environ. 599–600, 1658–1666. doi:10.1016/j.scitotenv.2017.05.107
Yang, C. H., Su, C. H., Liu, S. C., and Ng, L. T. (2019). Isolation, anti-Inflammatory activity and physico-chemical properties of bioactive polysaccharides from fruiting bodies of cultivated Cordyceps cicadae (Ascomycetes). Int. J. Med. Mushrooms 21, 995–1006. doi:10.1615/IntJMedMushrooms.2019031922
Yang, J. L., Dong, H. B., Wang, Y., Jiang, Y., Zhang, W. N., Lu, Y. M., et al. (2020). Cordyceps cicadae polysaccharides ameliorated renal interstitial fibrosis in diabetic nephropathy rats by repressing inflammation and modulating gut microbiota dysbiosis. Int. J. Biol. Macromol. 163, 442–456. doi:10.1016/j.ijbiomac.2020.06.153
Yang, Y. X., Chen, J. L., Lei, L., Li, F. H., Tang, Y., Yuan, Y., et al. (2019). Acetylation of polysaccharide from Morchella angusticeps peck enhances its immune activation and anti-inflammatory activities in macrophage RAW264.7 cells. Food Chem. Toxicol. 125, 38–45. doi:10.1016/j.fct.2018.12.036
Yin, Z. H., Liang, Z. H., Li, C. Q., Wang, J. M., Ma, C. Y., and Kang, W. Y. (2021). Immunomodulatory effects of polysaccharides from edible fungus: a review. Food Sci. Hum. Wellness 10, 393–400. doi:10.1016/j.fshw.2021.04.001
Yu, C. Q., Chen, P., Miao, L. Y., and Di, G. H. (2023). The role of the NLRP3 inflammasome and programmed cell death in acute liver injury. Int. J. Mol. Sci. 24, 3067. doi:10.3390/ijms24043067
Yu, H. L., Mei, S. H., Zhao, L., Zhao, M., Wang, Y. J., Zhu, H. M., et al. (2015). RGD-peptides modifying dexamethasone: to enhance the anti-inflammatory efficacy and limit the risk of osteoporosis. Med. Chem. Comm. 6, 1345–1351. doi:10.1039/c5md00215j
Yu, J. Y., Sun, J. G., Sun, M., Li, W. D., Qi, D. M., Zhang, Y. Q., et al. (2024). Protective mechanism of Coprinus comatus polysaccharide on acute alcoholic liver injury in mice, the metabolomics and gut microbiota investigation. Food Sci. Hum. Well. 13, 401–413. doi:10.26599/FSHW.2022.9250034
Zhang, J. C., Shi, X. Q., Cheng, W. J., Wang, Q., Wang, C. T., Li, M., et al. (2022a). An quan. comparison of the anti-Inflammatory and antioxidant activities of mycelial polysaccharides from different strains of Lingzhi or Reishi medicinal mushroom, Ganoderma lucidum (Agaricomycetes). Int. J. Med. Mushrooms. 24, 77–90. doi:10.1615/IntJMedMushrooms.2022044274
Zhang, R. J., Yuan, S. J., Ye, J. F., Wang, X. D., Zhang, X. D., Shen, J., et al. (2020). Polysaccharide from Flammuliana velutipes improves colitis via regulation of colonic microbial dysbiosis and inflammatory responses. Int. J. Biol. Macromol. 149, 1252–1261. doi:10.1016/j.ijbiomac.2020.02.044
Zhang, X., Li, J. P., Yang, B., Leng, Q. N., Li, J., Wang, X. T., et al. (2021). Alleviation of liver dysfunction, oxidative stress, and inflammation underlines the protective effects of polysaccharides from Cordyceps cicadae on high sugar/high fat diet-induced metabolic syndrome in rats. Chem. Biodivers. 18, e2100065. doi:10.1002/cbdv.202100065
Zhang, X. L., Wu, D. S., Tian, Y., Chen, X. D., Lan, J., Wei, F., et al. (2022b). Ganoderma lucidum polysaccharides ameliorate lipopolysaccharide-induced acute pneumonia via inhibiting NRP1-mediated inflammation. Pharm. Biol. 60, 2201–2209. doi:10.1080/13880209.2022.2142615
Zhang, Y. W., Cui, Y. F., Feng, Y. B., Jiao, F. P., and Jia, L. (2022c). Lentinus edodes polysaccharides alleviate acute lung injury by inhibiting oxidative stress and inflammation. Molecules 27, 7328. doi:10.3390/molecules27217328
Zhang, Z., Wu, D., Li, W., Chen, W. C., Liu, Y. F., Zhang, J. S., et al. (2023). Structural elucidation and anti-inflammatory activity of a proteoglycan from spent substrate of Lentinula edodes. Int. J. Biol. Macromol. 224, 1509–1523. doi:10.1016/j.ijbiomac.2022.10.239
Zhao, R. Q., Ji, Y., Chen, X., Su, A. X., Ma, G. X., Chen, G. T., et al. (2020). Effects of a β-type glycosidic polysaccharide from Flammulina velutipes on anti-inflammation and gut microbiota modulation in colitis mice. Food Funct. 11, 4259–4274. doi:10.1039/c9fo03017d
Zheng, Y., Fan, J., Chen, H. W., and Liu, E. Q. (2019). Trametes orientalis polysaccharide alleviates pm2.5-induced lung injury in mice through its antioxidant and anti-inflammatory activities. Food Funct. 10, 8005–8015. doi:10.1039/c9fo01777a
Zhu, B., Li, Y. Z., Hu, T., and Zhang, Y. (2019). The hepatoprotective effect of polysaccharides from Pleurotus ostreatus check for on carbon tetrachloride-induced acute liver injury rats. Int. J. Biol. Macromol. 131, 1–9. doi:10.1016/j.ijbiomac.2019.03.043
Zhu, Z. J., Huang, R., Huang, A., Wang, J., Liu, W., Wu, S. J., et al. (2022). Polysaccharide from Agrocybe cylindracea prevents diet-induced obesity through inhibiting inflammation mediated by gut microbiota and associated metabolites. Int. J. Biol. Macromol. 209, 1430–1438. doi:10.1016/j.ijbiomac.2022.04.107
Keywords: edible fungi, polysaccharides, anti-inflammatory effect, structureactivity relationship, health-promotion
Citation: Yin Z, Zhang J, Qin J, Guo L, Guo Q, Kang W, Ma C and Chen L (2024) Anti-inflammatory properties of polysaccharides from edible fungi on health-promotion: a review. Front. Pharmacol. 15:1447677. doi: 10.3389/fphar.2024.1447677
Received: 12 June 2024; Accepted: 12 July 2024;
Published: 26 July 2024.
Edited by:
Cinzia Parolini, University of Milan, ItalyReviewed by:
Sukkum Chang, Tulane University, United StatesCopyright © 2024 Yin, Zhang, Qin, Guo, Guo, Kang, Ma and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Wenyi Kang, a2FuZ3dlbnlAaG90bWFpbC5jb20=; Changyang Ma, bWFjYXlhMTAyNEBzaW5hLmNvbQ==; Lin Chen, bGNoZW5jaGluYUAxNjMuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.