
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ophthalmol. , 29 June 2023
Sec. Inflammatory Eye Diseases
Volume 3 - 2023 | https://doi.org/10.3389/fopht.2023.1183167
This article is part of the Research Topic Women in Inflammatory Eye Diseases: 2022 View all 5 articles
 Warlen Miiller Rocha Araujo1
Warlen Miiller Rocha Araujo1 Christiane Maria Ayo1†
Christiane Maria Ayo1† Mariana Previato1,2
Mariana Previato1,2 Geraldo Magela de Faria Jr.1
Geraldo Magela de Faria Jr.1 Fábio Batista Frederico1,2Rubens Camargo Siqueira1Gildásio Castello de Almeida Jr.1,2
Fábio Batista Frederico1,2Rubens Camargo Siqueira1Gildásio Castello de Almeida Jr.1,2 Vera Lúcia Pereira-Chioccola3
Vera Lúcia Pereira-Chioccola3 Luiz Carlos de Mattos1
Luiz Carlos de Mattos1 Cinara Cássia Brandão1†*
Cinara Cássia Brandão1†*Introduction: Ocular toxoplasmosis (OT) is an intraocular inflammation caused by Toxoplasma gondii infection that affects the retina and choroid, giving rise to posterior uveitis. Genetic polymorphisms in cytokine genes may exert influence in the expression of these molecules and play a significant role in inflammatory responses and susceptibility to OT. The aim of this study was to evaluate the role of polymorphisms rs16944 (–511 C > T) of the interleukin (IL) 1β gene and rs1800896 (–1082 G > A) of the IL10 gene on OT in Brazilian individuals with a serologic diagnosis of T. gondii and after conducting fundoscopic exams.
Methods: Participants with a positive serology were classified into two distinct groups according to the presence (G1; n = 110) or absence (G2; n = 104) of OT. The control group (G3) consisted of individuals without the infection (n = 108).
Results: It was observed that the C/C genotype of the IL1β gene polymorphism was a protective factor for OT (p = 0.02, OR = 0.28, 95% CI 0.08–0.78 for G1 vs. G2; p = 0.03; OR = 0.29, 95% CI 0.09–0.82 for G1 vs. G3), according to the recessive inheritance model.
Conclusions: The -511C.T polymorphisms of the IL1β gene seems to play an important role in the pathogenesis of OT in Brazilian individuals.
Ocular toxoplasmosis (OT) is an intraocular inflammation caused by Toxoplasma gondii infection that affects the retina and choroid (retinochoroiditis), giving rise to posterior uveitis. It can be of congenital origin or (in most cases) acquired after birth, and old lesions can reactivate at any time (1–4). In some cases, OT can leave serious sequelae, including complete loss of vision when the optic nerve is affected (1). Studies have shown that the prevalence and severity of OT are much higher in Brazil than in many other parts of the world, such as Europe and North America (5, 6).
In addition to the infective strain of the parasite, host genetic factors related to immune response can determine the final clinical outcome of infection (7, 8). Cytokines are crucial to the development and functioning of immune responses because they are important signaling molecules involved in cellular communication. Some of these molecules may play an important role in immune protection, but can also be pathological when unregulated (9).
Regarding toxoplasmosis, specific cytokines are important in the pathogenic process of OT (10). Genotypes and promoter sequences of genes related to the coding of cytokines have been associated with the development of human OT (11–17), as genetic polymorphisms in cytokine genes may exert influence on the expression of these molecules (14, 18) and play a significant role in inflammatory responses and susceptibility to OT.
IL-1β and IL-10 are cytokines that participate in the host immune response. T. gondii-infected human retinal pigment epithelium cells produce cytokines that activate neutrophils and lead to the production of proinflammatory cytokines such as IL-1β (19). Although inflammatory responses are critical for host defense against the parasite, excessive inflammation can result in tissue damage associated with OT. Furthermore, T. gondii can induce IL-1β production by monocytes depending on the specific strain (20). In contrast, an important role for the anti-inflammatory cytokine IL-10 in modulating ocular disease has been demonstrated, as this cytokine contributes to the balance between protective and pathological T-cell responses. (21). The IL1β gene is located on chromosome 2. The –511 polymorphism, located in the IL1β promoter region, affects the regulatory sites of IL1-β, thus influencing gene expression and circulatory levels of this cytokine. The literature shows that the T allele was associated with lower levels of mRNA expression for IL1-β (22). The IL10 gene is located on chromosome 1. The A allele of the –1082 polymorphism in the IL10 promoter region was associated with lower levels of production of IL-10 (23).
Considering that the expression of cytokines is regulated by genetic variants and evidence suggests that cytokine gene polymorphisms may contribute to the presence of ocular lesions in patients infected by T. gondii, the aim of this study was to evaluate the role of polymorphisms rs16944 (–511 C > T) of the IL1β gene and rs1800896 (–1082 G > A) of the IL10 gene in OT in Brazilian individuals.
This study was approved by the Research Ethics Committee of the Medicine School in São José do Rio Preto (CAAE 32259714.8.00005415). All individuals who agreed to participate signed informed consent forms.
This was a case–control study. Between January 2015 and December 2017, a total of 322 consecutive and unrelated individuals seen at the Retinopathy Outpatient Clinic of the Specialty Hospital, Faculty of Medicine de São José do Rio Preto (HB-FUNFARME), participated in this study. These patients were classified into three groups. Group 1 (G1) contained 110 individuals with a positive serology for T. gondii and with OT present. Group 2 (G2) contained 104 individuals with a positive serology for T. gondii and the ocular diseases other than toxoplasmosis present, such as age-related macular degeneration (24.0%), cataracts (19.2%), retinal detachment (14.4%), diabetic retinopathy (10.6%), glaucoma (6.7%), macular edema (3.8%), pterygium (3.8%), optic neuropathy (2.9%), macular atrophy (1.9%), and other ocular diseases (12.5%). Group 3 (G3) contained 108 individuals with a non-reactive serology for T. gondii (control group) who attended the clinic to check their visual acuity.
All individuals who participated in this study were residents of municipalities in the northwest region of the State of São Paulo—located in the southeast region of Brazil, between 20°49′13″S and 49°22′47″W. The individuals were monitored and evaluated for clinical symptoms, T. gondii serology, and epidemiological data. In addition, they were defined as belonging to a population of mixed ethnicity owing to the high heterogeneity of the Brazilian population (24).
The inclusion criteria for individuals with OT were a positive laboratory diagnosis for T. gondii and the presence of scars/eye lesions due to toxoplasmosis. Inclusion criteria for individuals without OT were a positive laboratory diagnosis for T. gondii and an absence of eye scars/lesions due to toxoplasmosis. The inclusion criterion for the individuals who made up the control group was a negative laboratory diagnosis for T. gondii.
Exclusion criteria were individuals with other infectious and parasitic diseases, individuals with some type of mental disability, individuals with blood dyscrasia and who used oral anticoagulants, and individuals related to patients already enrolled.
Blood samples were collected in tubes without anticoagulant to obtain serum. The presence of anti-T. gondii IgM and IgG was confirmed by enzyme-linked immunosorbent assay (ELISA) (DiaSorin S.p.A., Saluggia, Italy) using the ETI-TOXOK-M reverse plus kit for IgM and the ETI-TOXOK-G plus kit for IgG, and by enzyme-linked fluorescence assay (ELFA) (BioMérieux, Marcy-l’Étoile, France) using the Vidas® Toxo IgM (TXM) kit for screening for IgM; and the Vidas® Toxo IgG II (TXG) kit for screening for IgG. All tests were performed according to the manufacturer’s instructions.
All individuals were clinically evaluated by two experienced ophthalmologists using retinography according to the Early Treatment Diabetic Retinopathy Study Report (ETDRS) standards and criteria (25, 26). As no invasive test was performed, this was a presumptive diagnosis for OT.
Genomic DNA was extracted using a silica column QIAamp Blood Mini kit (Qiagen) from peripheral blood collected with ethylenediaminetetraacetic acid (EDTA). DNA quantity and quality were measured using a spectrophotometer (BioTek, Epoch Instruments).
The respective cytokines’ polymorphisms were selected based on data from previous studies carried out in the context of ocular toxoplasmosis or other human diseases. In addition, single nucleotide polymorphisms (SNPs) were chosen based on their predicted effect on the level of gene expression, which was obtained through STIFF tool analysis, available at the Ensembl genome browser (https://www.ensembl.org/index.html), and through previous reports of an association with any disease condition. Genotyping for the identification of the rs16944 (–511 C/T) polymorphism of the IL1β gene and rs1800896 (–1082 G/A) polymorphism of the IL10 gene was performed using the PCR-restriction fragment length polymorphism (PCR-RFLP) technique. The primer pair used for the IL1β gene was sense 5′-TGGCATTGATCTGGTTCATC-3′ and antisense 5′-GTTTAGGAATCTTCCCACTT-3′, and the primer pair used for the IL10 gene was sense 5′-CCAAGACAACACTACTAAGGCTCCTTT-3′ and antisense 5′-GCTTCTTATATGCTAGTCAGGTA-3′. The PCR reaction conditions were previously described by Piatto et al. (27). The PCR products were digested with Eco88I (Thermo Fisher Scientific, Waltham, MA, USA) for the IL1β gene and with the FaqI (Thermo Fisher Scientific) for the IL10 gene. The digested PCR products were subsequently separated by agarose gel electrophoresis and detected using the SYBR™ Green nucleic acid gel stain (Invitrogen Life Technologies, Grand Island, NY, USA).
Allele and genotypic frequencies were obtained by direct counting, and the Hardy–Weinberg equilibrium (HWE) was verified according to the method described by Guo and Thompson (28). Comparisons of these frequencies between the groups of patients were performed using the chi-squared test (χ2) with Yates’ correction or by Fisher’s exact test, using a 3 × 3 contingency table for genotypes and a 3 × 2 contingency table for alleles. The genotypic frequencies were evaluated against each other using the models of dominant, recessive, and codominant inheritance, by means of a 2 × 2 contingency table. The odds ratio (OR) was calculated to verify association estimates using the GraphPad InStat program (InStat 3.06, 2003). The mean ages were compared using a t-test. All tests produced a 95% confidence interval (CI) and differences with p-values < 0.05 were considered statistically significant.
The general characteristics of the study individuals are shown in Table 1. Of the total number of individuals analyzed in G1, 60 (54.5%) were male and 50 (45.5%) were female; in G2, 50 (48.0%) were male and 54 (52.0%) were female; and in G3, 55 (51.9%) were male and 53 (49.1%) were female. The mean age was statistically different between the groups (G1 vs. G2 vs. G3: p < 0.0001; G1 vs. G2: p < 0.0001, t = 5.38; G1 vs. G3: p = 0.002, t = 3.04; G2 vs. G3: p < 0.0001, t = 10.25); in G1 the mean age was 72.7 years (± 21.4 years), in G2 it was 56.9 years (± 17.0 years), and in G3 it was 35.2 years (± 13.6 years).
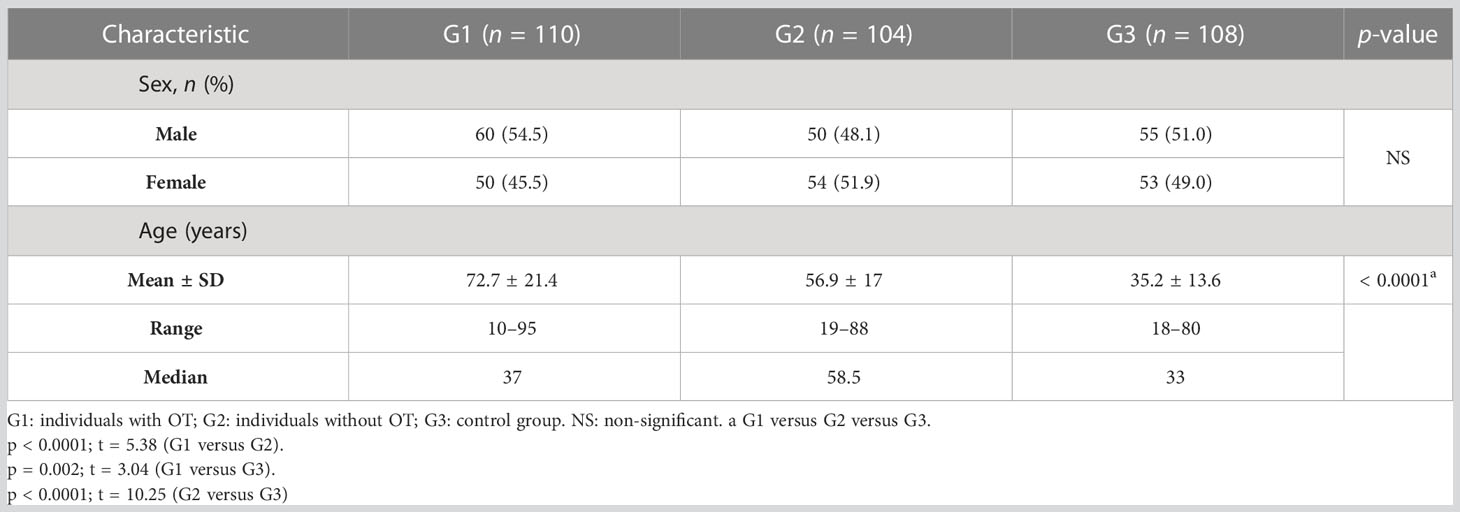
Table 1 General characteristics of individuals infected by Toxoplasma gondii with and without ocular toxoplasmosis and of individuals without the infection.
The distribution of alleles and genotypes for the –511 IL1β and –1082 IL10 polymorphisms was in the HWE in the control group (p > 0.05).
In relation to the IL1β gene polymorphism (position –511), there was a statistically significant difference between the groups in genotype frequency (p = 0.0028; χ2 = 16.13; gl = 4). According to the inheritance models, a significant difference was found using the recessive model for the C allele as a protective factor against OT, showing that it would be necessary to have two C alleles (C/C genotype) to exert protection against the occurrence of OT (p = 0.02, OR = 0.28, 95% CI 0.08–0.78 for G1 vs. G2; and p = 0.03, OR = 0.29, 95% CI 0.09–0.82 for G1 vs. G3). The alleles expressed as heterozygous (T/C genotype) were more frequent in G1 than in G2 (p = 0.056, OR = 2.05, 95% CI 1.03–4.16) and G3 (p = 0.0004, OR = 3.30, 95% CI 1.72–6.52). The presence of two T alleles (T/T genotype) was more frequent in G3 than in G1 (p = 0.02, OR = 0.39, 95% CI 0.17–0.83) and G2 (p = 0.059, OR = 0.45, 95% CI 0.20–0.96). However, both the T/C and T/T genotype were not associated with the occurrence of OT, as shown by the codominant and dominant inheritance models, respectively (Table 2).
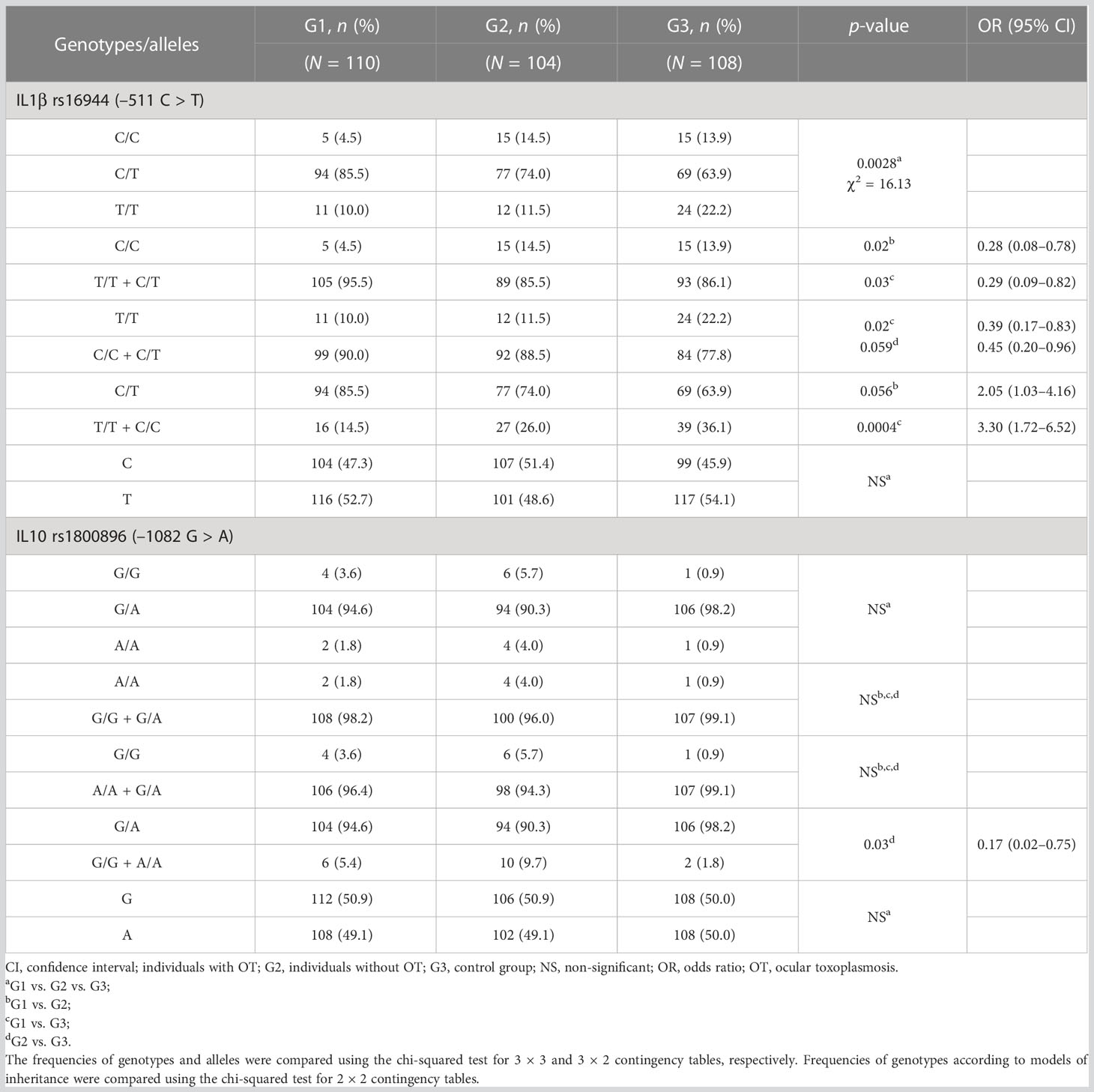
Table 2 Genotype and allele distribution for IL1β –511 and IL10 –1082 polymorphisms in individuals infected by Toxoplasma gondii with and without ocular toxoplasmosis and in individuals without the infection.
The comparison of G1 + G2 with G3 showed that the wild-type homozygote (T/T) genotype of the IL1β –511 polymorphism was more frequent in G3 (p = 0.007, OR = 0.042, 95% CI 0.22–0.78) and the heterozygous genotype was more frequent in individuals with the infection (G1 + G2) (p = 0.002, OR = 2.24, 95% CI 1.33–3.76) (Supplementary Table 1).
Regarding the –1082 polymorphism of the IL10 gene, statistically significant differences were not found for genotype and allele distribution between the groups. Considering the analyses through the inheritance models, the codominant model showed that the heterozygous G/A genotype was more frequent in G3 than in G2 (p = 0.03, OR = 0.17, 95% CI 0.02–0.75) (Table 2). This same observation was found in the comparison of G1 + G2 with G3, in which the heterozygous genotype was more frequent in patients without the infection (G3) (p = 0.04, OR = 0.23, 95% CI 0.05–1.03) (Supplementary Table 1).
Despite numerous discoveries in recent years, OT remains a poorly understood disease, despite its epidemiological relevance (29). The importance of age in the clinical course of OT has been reported, and studies have shown that the disease most often affects patients between the second and fourth decades of life (2, 30, 31). However, OT can develop at any time in life (2, 32) and older patients appear to be at greater risk of developing eye injuries (2, 3, 5).
In this study, the group of individuals with eye lesions resulting from a T. gondii infection (G1) had a higher mean age than the other groups (G2 and G3). Ferreira et al. (33) demonstrated that a higher average age is a risk factor for the development of OT (33). In addition, as most cases of OT occur due to infections acquired after birth (1), the difference observed in terms of age between the groups in this study can be attributed to longer exposure to the parasite, both for the development of OT and for the acquisition of infection. A possible explanation for the higher mean age observed in G2 than in G3 is that eye diseases, other than OT, are more prevalent in older patients (34).
CD4+ and CD8+ T cells, B cells, and macrophages have been identified as the effector cells that attack and destroy infected neurons in the retina. The expansion, migration, and activation of these cells may be associated with cytokine signaling functions (29). Individuals with OT present abnormal expression in the levels of some cytokines (i.e., elevated or reduced levels), and these changes in expression levels may be associated with polymorphisms in cytokine genes (14, 18).
The data obtained in this study showed that the homozygous C/C genotype of the IL1β gene rs16944 polymorphism was a protective factor for OT in a Brazilian population; this finding has never been previously published. According to the literature, the C allele of this mutation is related to the high expression of the gene, i.e., increased production of the cytokine (11, 15–17, 35–37). Therefore, we suggest that the homozygous presence of the C allele may generate an efficient immune response for parasite control, so that individuals with this polymorphism are protected against the occurrence of OT.
IL-1β upregulates other cytokines that are known to be potent neutrophil chemoattractants (19). Therefore, neutrophil recruitment to the T. gondii-infected human retinal pigment epithelium cells, enhanced by IL-1β, may play an important role in the pathogenesis of the infection. It is known that IL-1β can modulate pro-inflammatory T helper (Th) 1 (Th1) response by impeding IL-6-induced phosphorylation of signal transducer and activator of transcription 1 (38). A regulated Th1 response is necessary for the efficient control of the parasites and to avoid tissue damage. High levels of IL-1β inhibit the growth of the parasite in endothelial cells in the retinal region (10, 36). Machado et al. (39) showed that in newborns with a positive serology for T. gondii, the increase in IL-1β production was an important indicator of protection against or resolution of eye injury. In addition, Brunton et al. (36) demonstrated that low levels of IL-1β expression can result in OT. However, a recent study conducted by Naranjo-Galvis et al. in Colombian subjects found no association between the rs16944 polymorphism in the –511 region of the IL1β gene and OT (17).
In this study, the genotypes and alleles of the rs1800896 polymorphism (–1082 G > A) of the IL10 gene were not associated with the occurrence of OT. Our results contrast with the results reported by other studies. In a population from southeastern Brazil, evidence of association was found between the A allele (rs1800896) and the presence of OT (11). In Colombian patients the G/G genotype (rs1800896) and the IL10 gene “AG” haplotype (rs1800896, rs1800871) were associated with the presence of OT (17). The A allele at position –1082 is associated with a low level of production of IL-10 (40). The discrepancies observed between studies may be caused, in part, by differences in methodology, sample size, and patient selection. In addition, additional factors that are difficult to control in the context of toxoplasmosis, such as duration of infection, may also have an effect on eye disease and serve as a bias. Moreover, as allelic frequency of the cytokine genes can differ between populations according to regional variations (41, 42), associations involving IL1β and IL10 polymorphisms could result in different clinical and immune phenotypes in patients with OT from different populations.
Previous studies analyzed the influence of other cytokine polymorphisms on OT.
The C allele of the IL-6 −174 G > C polymorphism, which is related to a decreased production of IL-6, may be associated with the occurrence of OT (18). In another study, the recurrence of OT was associated with the IL1A −889 C > T polymorphism (T allele), which is related to an increase in IL-1α expression (13). The A/A genotype of the interferon gamma (IFN-γ) gene polymorphism (position +874 T > A), which is related to reduced IFN-γ production, was associated with the presence of OT (14). A study on the tumor necrosis factor alpha (TNF-α) gene polymorphism (−308 G > A) did not find an association with the presence or recurrence of OT (12).
The data from this study also indicated that the homozygous T/T genotype of the IL1β –511 polymorphism and the heterozygous G/A genotype at position –1082 of the IL10 gene were more frequent in individuals without the T. gondii infection. These links between infection and genotypes, which are associated with low or intermediate production of IL-1β and IL-10, respectively, provide evidence that variations in the genetic control of cytokine expression levels may influence the immune response in toxoplasmosis, as these levels influence systemic immune modulation, altering the number and responsiveness of host T cells. T-cell-mediated immune responses exercise a relevant influence on the suppression of T. gondii replication, resulting in chronic infection (4). Because of the difference in mean age, it can be presumed that control individuals were not exposed to infection by T. gondii and this could be a bias. However, G3 was useful for comparing the frequencies of the alleles and genes with the infected groups, as it was composed of healthy individuals.
The pathogenic mechanisms involved in the ocular form of toxoplasmosis can be explained, at least in part, by a cytokine-guided immune response (10, 21, 29). It is known that the production of certain cytokines is under genetic control. Therefore, polymorphisms in the promoter regions of cytokine genes could affect gene transcription, and, consequently, the expression of these molecules, in turn causing interindividual variations. Although IL-1β is one of the important cytokines involved in the pathogenesis of OT, the mechanism appears to be complex, involving many other cytokines and factors. Studies that aim to clarify the importance of cytokines in OT can offer perspectives for immunomodulatory strategies for the treatment of this disease. The main limitation of this study was that the expression levels of IL-1β and IL-10 were not evaluated.
In conclusion, the –511 C > T polymorphism of the IL1β gene seems to play an important role in the pathogenesis of OT in Brazilian individuals, whereas the –1082 polymorphism of the IL10 gene does not seem to influence the occurrence of OT in this population.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
The studies involving human participants were reviewed and approved by the Research Ethics Committee of the Medicine School in São José do Rio Preto (CAAE 32259714.8.00005415). The patients/participants provided their written informed consent to participate in this study.
CB was head of the FAMERP Toxoplasma Research Group. CB and CA contributed equally. CB and CA conceived and designed the study. FF, MP, RS, and GA selected the patients and developed the clinical diagnosis. WA and GM developed the molecular analysis. CB, CA, WA, VLP-C, and LM were responsible for the data analysis. CA, CB, and WA were responsible for writing the manuscript. All the authors approved the final version of the manuscript. All authors contributed to the article and approved the submitted version.
This study was supported by grants from the Brazilian Ministry of Education CAPES Scholarship (Coordination of Improvement of Higher Education Personnel, Brazil—financing code 001), given to WA and to CA; by the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP #2013/25650-8 to LM, #2013/06580-9 to CA, #2015/17226-7 to GA, #2018/09448-8 and 2019/05581–8 to GM, and #2018/04709–8 to VLP-C); by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq 09/2022 to LM). The opinions, assumptions, and conclusions or recommendations expressed in this material are the responsibility of the authors and do not necessarily reflect the views of FAPESP.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fopht.2023.1183167/full#supplementary-material
1. Maenz M, Schlüter D, Liesenfeld O, Schares G, Gross U, Pleyer U. Ocular toxoplasmosis past, present and new aspects of an old disease. Prog Retin. Eye Res (2014) 39:77–106. doi: 10.1016/j.preteyeres.2013.12.005
2. Aleixo ALQ, Do C, Vasconcelos C, de Oliveira R, Cavalcanti Albuquerque M, Biancardi AL, et al. Toxoplasmic retinochoroiditis: the influence of age, number of retinochoroidal lesions and genetic polymorphism for IFN-γ +874 T/A as risk factors for recurrence in a survival analysis. PloS One (2019) 14:e0211627. doi: 10.1371/journal.pone.0211627
3. Vasconcelos-Santos DV. Ocular toxoplasmosis BT - posterior uveitis: advances in imaging and treatment. In: Rao N., Schallhorn J., Rodger D. eds. Posterior Uveitis. Essentials in Ophthalmology (2019) Springer, Cham. doi: 10.1007/978-3-030-03140-4_5
4. de Barros RAM, Torrecilhas AC, Marciano MAM, Mazuz ML, Pereira-Chioccola VL, Fux B. Toxoplasmosis in human and animals around the world. diagnosis and perspectives in the one health approach. Acta Trop (2022) 231:106432. doi: 10.1016/j.actatropica.2022.106432
5. Dubey JP, Lago EG, Gennari SM, Su C, Jones JL. Toxoplasmosis in humans and animals in Brazil: high prevalence, high burden of disease, and epidemiology. Parasitology (2012) 139:1375–424. doi: 10.1017/S0031182012000765
6. Rahimi Esboei B, Kazemi B, Zarei M, Mohebali M, Keshavarz Valian H, Shojaee S, et al. Evaluation of RE and B1 genes as targets for detection of toxoplasma gondii by nested PCR in blood samples of patients with ocular toxoplasmosis. Acta Parasitol (2019) 64:384–9. doi: 10.2478/s11686-019-00056-6
7. Hernández-de-los-Ríos A, Murillo-Leon M, Mantilla-Muriel LE, Arenas AF, Vargas-Montes M, Cardona N, et al. Influence of two major toxoplasma gondii virulence factors (ROP16 and ROP18) on the immune response of peripheral blood mononuclear cells to human toxoplasmosis infection. Front Cell Infect Microbiol (2019) 9:413. doi: 10.3389/fcimb.2019.00413
8. Mantilla-Muriel LE, Hernández-de-los-Ríos A, Rincón M, De-la-Torre A, Murillo-León M, Cardona N, et al. Serotyping, host genes and cytokines response in human ocular toxoplasmosis. Microb Pathog (2020) 148:104465. doi: 10.1016/j.micpath.2020.104465
9. Netea MG, Domínguez-Andrés J, Barreiro LB, Chavakis T, Divangahi M, Fuchs E, et al. Defining trained immunity and its role in health and disease. Nat Rev Immunol (2020) 20:375–88. doi: 10.1038/s41577-020-0285-6
10. Cordeiro CA, Moreira PR, Dutra WO, Young L, Campos WR, Oréfice F. Imunologia da retinocoroidite toxoplásmica immunology of the toxoplasmic retinochoroiditis. Arq Bras Oftalmol (2010) 73:548–51. doi: 10.1590/S0004-27492010000600018
11. Cordeiro CA, Moreira PR, Andrade MS, Dutra WO, Campos WR, Oréfice F, et al. Interleukin-10 gene polymorphism (-1082G/A) is associated with toxoplasmic retinochoroiditis. Investig Ophthalmol Vis Sci (2008) 49:1979–82. doi: 10.1167/iovs.07-1393
12. Cordeiro CA, Moreira PR, Costa GC, Dutra WO, Campos WR, Oréfice F, et al. Interleukin-1 gene polymorphisms and toxoplasmic retinochoroiditis. Br J Ophthalmol (2008) 92:986–8. doi: 10.1136/bjo.2008.140590
13. Cordeiro CA, Moreira PR, Costa GC, Dutra WO, Campos WR, Oréfice F, et al. TNF-α gene polymorphism (-308G/A) and toxoplasmic retinochoroiditis. Br J Ophthalmol (2008) 92:986–8. doi: 10.1136/bjo.2008.140590
14. de Albuquerque MC, do Couto Aleixo ALQ, Benchimol EI, Leandro ACCS, das Neves LB, Vicente RT, et al. The IFN-γ+874T/A gene polymorphism is associated with retinochoroiditis toxoplasmosis susceptibility. Mem Inst Oswaldo Cruz (2009) 104:451–5. doi: 10.1590/s0074-02762009000300009
15. Turner MD, Nedjai B, Hurst T, Pennington DJ. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim Biophys Acta - Mol Cell Res (2014) 1843:2563–82. doi: 10.1016/j.bbamcr.2014.05.014
16. Vargas-Alarcón G, Cruz-López M, Valladares A, Álvarez-León E, Juárez-Cedillo T, Pérez-Méndez Ó., et al. The interleukin-1β-511 T>C (rs16944) gene polymorphism is associated with risk of developing silent myocardial ischemia in diabetic patients. Immunol Lett (2015) 168:7–12. doi: 10.1016/j.imlet.2015.08.005
17. Naranjo-Galvis CA, de-la-Torre A, Mantilla-Muriel LE, Beltrán-Angarita L, Elcoroaristizabal-Martín X, McLeod R, et al. Genetic polymorphisms in cytokine genes in colombian patients with ocular toxoplasmosis. Infect Immun (2018) 86:e00597–17. doi: 10.1128/IAI.00597-17
18. Cordeiro CA, Moreira PR, Bessa TF, Costa GC, Dutra WO, Campos WR, et al. Interleukin-6 gene polymorphism (–174 G/C) is associated with toxoplasmic retinochoroiditis. Acta Ophthalmol (2013) 91:e311–4. doi: 10.1111/aos.12046
19. Nagineni CN, Detrick B, Hooks JJ. Toxoplasma gondii infection induces gene expression and secretion of interleukin 1 (IL-1), IL-6, granulocyte-macrophage colony-stimulating factor, and intercellular adhesion molecule 1 by human retinal pigment epithelial cells. Infect Immun (2000) 68:407–10. doi: 10.1128/IAI.68.1.407-410.2000
20. Gov L, Karimzadeh A, Ueno N, Lodoen MB. Human innate immunity to toxoplasma gondii is mediated by host caspase-1 and ASC and parasite GRA15. mBio (2013) 4:e00255–13. doi: 10.1128/mBio.00255-13
21. Garweg JG, Candolfi E. Immunopathology in ocular toxoplasmosis: facts and clues. Mem Inst Oswaldo Cruz (2009) 104:211–20. doi: 10.1590/S0074-02762009000200014
22. Iglesias Molli AE, Bergonzi MF, Spalvieri MP, Linari MA, Frechtel GD, Cerrone GE. Relationship between the IL-1β serum concentration, mRNA levels and rs16944 genotype in the hyperglycemic normalization of T2D patients. Sci Rep (2020) 10:9985. doi: 10.1038/s41598-020-66751-x
23. D’Alfonso S, Rampi M, Rolando V, Giordano M, Momigliano-Richiardi P, D’Alfonso S, et al. New polymorphisms in the IL-10 promoter region. Genes Immun (2000) 1:231–3. doi: 10.1038/sj.gene.6363666
24. Parra FC, Amado RC, Lambertucci JR, Rocha J, Antunes CM, Pena SDJJ. Color and genomic ancestry in brazilians. Proc Natl Acad Sci (2003) 100:177–82. doi: 10.1073/pnas.0126614100
25. Photocoagulation for Diabetic Macular Edema. Arch. Ophthalmol. (1985) 103:1796. doi: 10.1001/archopht.1985.01050120030015
26. Previato M, Frederico FB, Henrique Antunes Murata F, Siqueira RC, Barbosa AP, Silveira-Carvalho AP, et al. A Brazilian report using serological and molecular diagnosis to monitoring acute ocular toxoplasmosis. BMC Res Notes (2015) 8:746. doi: 10.1186/s13104-015-1650-6
27. Gabriel ML, Braga FB, Cardoso MR, Lopes AC, Piatto VB, Souza AS. The association between pro- and anti-inflammatory cytokine polymorphisms and periventricular leukomalacia in newborns with hypoxic-ischemic encephalopathy. J Inflamm Res (2016) 9:59–67. doi: 10.2147/JIR.S103697
28. Guo SW, Thompson EA. Performing the exact test of hardy-Weinberg proportion for multiple alleles. Biometrics (1992) 48:361. doi: 10.2307/2532296
29. Greigert V, Bittich-Fahmi F, Pfaff AW. Pathophysiology of ocular toxoplasmosis: facts and open questions. PloS Negl Trop Dis (2020) 14:e0008905. doi: 10.1371/journal.pntd.0008905
30. Bosch-Driessen LEH, Berendschot TTJM, Ongkosuwito JV, Rothova A. Ocular toxoplasmosis: clinical features and prognosis of 154 patients. Ophthalmology (2002) 109:869–78. doi: 10.1016/S0161-6420(02)00990-9
31. Holland GN. Ocular toxoplasmosis: a global reassessment: part II: disease manifestations and management. Am J Ophthalmol (2004) 137:1–17. doi: 10.1016/j.ajo.2003.10.032
32. Furtado JM, Winthrop KL, Butler NJ, Smith JR. Ocular toxoplasmosis I: parasitology, epidemiology and public health. Clin Exp Ophthalmol (2013) 41:82–94. doi: 10.1111/j.1442-9071.2012.02821.x
33. Ferreira AIC, De Mattos CCB, Frederico FB, Meira CS, Almeida GC, Nakashima F, et al. Risk factors for ocular toxoplasmosis in Brazil. Epidemiol. Infect (2014) 142:142–8. doi: 10.1017/S0950268813000526
35. Turner DM, Williams DM, Sankaran D, Lazarus M, Sinnott PJ, Hutchinson IV. An investigation of polymorphism in the interleukin-10 gene promoter. Eur J Immunogenet. (1997) 24:1–8. doi: 10.1111/j.1365-2370.1997.tb00001.x
36. Brunton CL, Wallace GR, Graham E, Stanford MR. The effect of cytokines on the replication of t. gondii within rat retinal vascular endothelial cells. J Neuroimmunol. (2000) 102:182–8. doi: 10.1016/S0165-5728(99)00167-8
37. Chua K, Lau T, Tee Z, Tan S, Lian L. Genetic polymorphisms of the interleukin-1 beta (IL-1 beta ) -511 and +3954 single nucleotide polymorphisms (SNPs) in Malaysian systemic lupus erythematosus (SLE) patients. J Heal Sci (2009) 55:657–62. doi: 10.1248/jhs.55.657
38. Shen X, Tian Z, Holtzman MJ, Gao B. Cross-talk between interleukin 1beta (IL- 1beta) and IL-6 signalling pathways: IL-1beta selectively inhibits IL-6-activated signal tr ansducer and activator of transcription factor 1 (STAT1) by a proteasome- dependent mechanism. Biochem J (2000) 3:913–9. doi: 10.1042/BJ3520913
39. Machado AS, Carneiro ACAV, Béla SR, Andrade GMQ, Vasconcelos-Santos DV, Januário JN, et al. Biomarker analysis revealed distinct profiles of innate and adaptive immunity in infants with ocular lesions of congenital toxoplasmosis. Mediators Inflamm (2014) 2014:ID910621. doi: 10.1155/2014/910621
40. Yousefi A, Zare Bidoki A, Shafioyoun A, Sadr M, Varzaneh FN, Shabani M, et al. Association of IL-10 and TGF-beta cytokine gene polymorphisms with autoimmune hepatitis. Clin Res Hepatol Gastroenterol (2018) 43:45–50. doi: 10.1016/j.clinre.2018.07.004
41. Ness RB, Haggerty CL, Harger G, Ferrel R. Differential distribution of allelic variants in cytokine genes among African americans and white americans. Am J Epidemiol (2004) 160:1033–8. doi: 10.1093/aje/kwh325
Keywords: ocular toxoplasmosis, cytokines, interleukins, genetic polymorphisms, IL1β and IL10
Citation: Araujo WMR, Ayo CM, Previato M, de Faria GM Jr., Frederico FB, Siqueira RC, de Almeida GC Jr., Pereira-Chioccola VL, de Mattos LC and Brandão CC (2023) Role of interleukin 1β and interleukin 10 variants on ocular toxoplasmosis in Brazilian individuals. Front. Ophthalmol. 3:1183167. doi: 10.3389/fopht.2023.1183167
Received: 09 March 2023; Accepted: 02 June 2023;
Published: 29 June 2023.
Edited by:
Alejandra de-la-Torre, Rosario University, ColombiaReviewed by:
Graham Wallace, University of Birmingham, United KingdomCopyright © 2023 Araujo, Ayo, Previato, de Faria, Frederico, Siqueira, de Almeida, Pereira-Chioccola, de Mattos and Brandão. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cinara Cássia Brandão, Y2luYXJhLmJyYW5kYW9AZmFtZXJwLmJy
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.