 Jihane Balla
Jihane Balla
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Oncol. , 10 March 2025
Sec. Molecular and Cellular Oncology
Volume 15 - 2025 | https://doi.org/10.3389/fonc.2025.1548524
This article is part of the Research Topic Innovative ADC and CAR-T Therapies for High-Risk Neuroblastoma: A New Hope View all articles
Neuroblastoma (NB) is a pediatric cancer that often manifests in a high-risk form and is characterized by frequent relapses and resistance to conventional therapies. This underscores the urgent need for more effective and targeted treatment strategies. One promising avenue has been the identification of unique or overexpressed surface antigens on neoplastic cells, which has facilitated the development of antibody conjugates and related technologies. These include antibody-drug conjugates (ADCs) and immunotoxins (ITs), which deliver cytotoxic agents directly to tumor cells, as well as antibody-fluorophore conjugates (AFCs), which bind to surface antigens with high specificity to target malignant tumors. Additionally, radioimmunotherapy (RIT) allows the precise delivery of radioactive isotopes linked to a monoclonal antibody directly to the tumor cells. ADCs, ITs, and RIT represent a novel class of anti-cancer agents offering precision therapy with reduced systemic toxicity, enabling longer and potentially more effective treatment regimens. Meanwhile, AFCs are valuable tools in diagnostics, aiding in detecting and characterizing malignant tissues. Despite advancements in antibody conjugates for NB, significant challenges persist, including optimizing payload delivery, mitigating off-target effects, and addressing tumor heterogeneity. Future research should also prioritize refining and integrating these technologies into multimodal treatment protocols to improve outcomes for pediatric NB patients.
Neuroblastoma (NB) is a highly aggressive pediatric tumor that arises from neural crest cells of the sympathetic nervous system, accounting for approximately 8-10% of juvenile cancers and 15% of pediatric fatalities (1). Around the ages of 17 and 18 months, NB is frequently identified; it rises during the first year following birth and subsequently declines with age. Nearly 90% of all cases involve children younger than five (1). While it is very low in South and East Asia, the prevalence is higher in high-income nations and among certain ethnic groups. The frequency is 11.5 per million for white children and 8.5 per million for black children in the United States. Unfortunately, the prevalence and mortality rates of NB have not yet been the subject of a systematic global assessment (1). The prevalence and mortality rates among children with NB aged 0-14 years were analyzed by the Global Burden of Disease database. A total of 5560 cases and 1977 fatalities were reported in 2021. Incidence increased by 30.26%, and mortality by 20.35% from 1990 to 2021 (1). Its clinical behavior varies from spontaneous regression to relentless progression, influenced by tumor genetics, patient age, and disease stage. High-risk NB (HR-NB), often metastatic at diagnosis, has a poor prognosis, with a 5-year survival rate of 40–50% (2). Relapsed or therapy-resistant HR-NB survival drops to ~20%, highlighting the urgent need for better treatments (2). The standard treatment for HR-NB relies on multimodal protocols, involving intensive chemotherapy, surgical resection, radiation therapy, and hematopoietic stem cell transplantation, alongside targeted therapies addressing MYCN amplification, anaplastic lymphoma kinase (ALK) mutations, and signaling pathway abnormalities (3, 4). Epigenetic modulators and therapies targeting norepinephrine and somatostatin receptors are also under investigation for their potential to improve outcomes (5, 6). Despite these advances, 50-60% HR-NB patients face recurrence with limited curative options for relapsed or resistant cases. Conventional therapies lack specificity, indiscriminately targeting dividing cells, causing severe side effects and fostering chemoresistance (CR) (7). These challenges underscore the need for novel therapeutic approaches that combine efficacy with precision to minimize collateral damage and improve patient outcomes. Advances in molecular oncology provided valuable insights into potential solutions to address these limitations. One promising avenue is the identification of tumor-specific or overexpressed surface antigens on NB cells (6). These discoveries have facilitated the development of targeted therapies and diagnostic tools such as antibody conjugates (ACs), including antibody-drug conjugates (ADCs), immunotoxins (ITs), antibody-fluorophore conjugates (AFCs) and Radioimmunotherapy (RIT) (8–12) (Figure 1). By combining these innovative approaches with traditional therapies, there is potential to enhance therapeutic efficacy, address CR, and minimize the side effects of standard treatments. These emerging strategies provide renewed hope for improving outcomes in HR-NB. This review focuses on the latest progress made regarding the use of ACs for NB, underscoring their transformative potential in enhancing precision therapy and diagnosis.
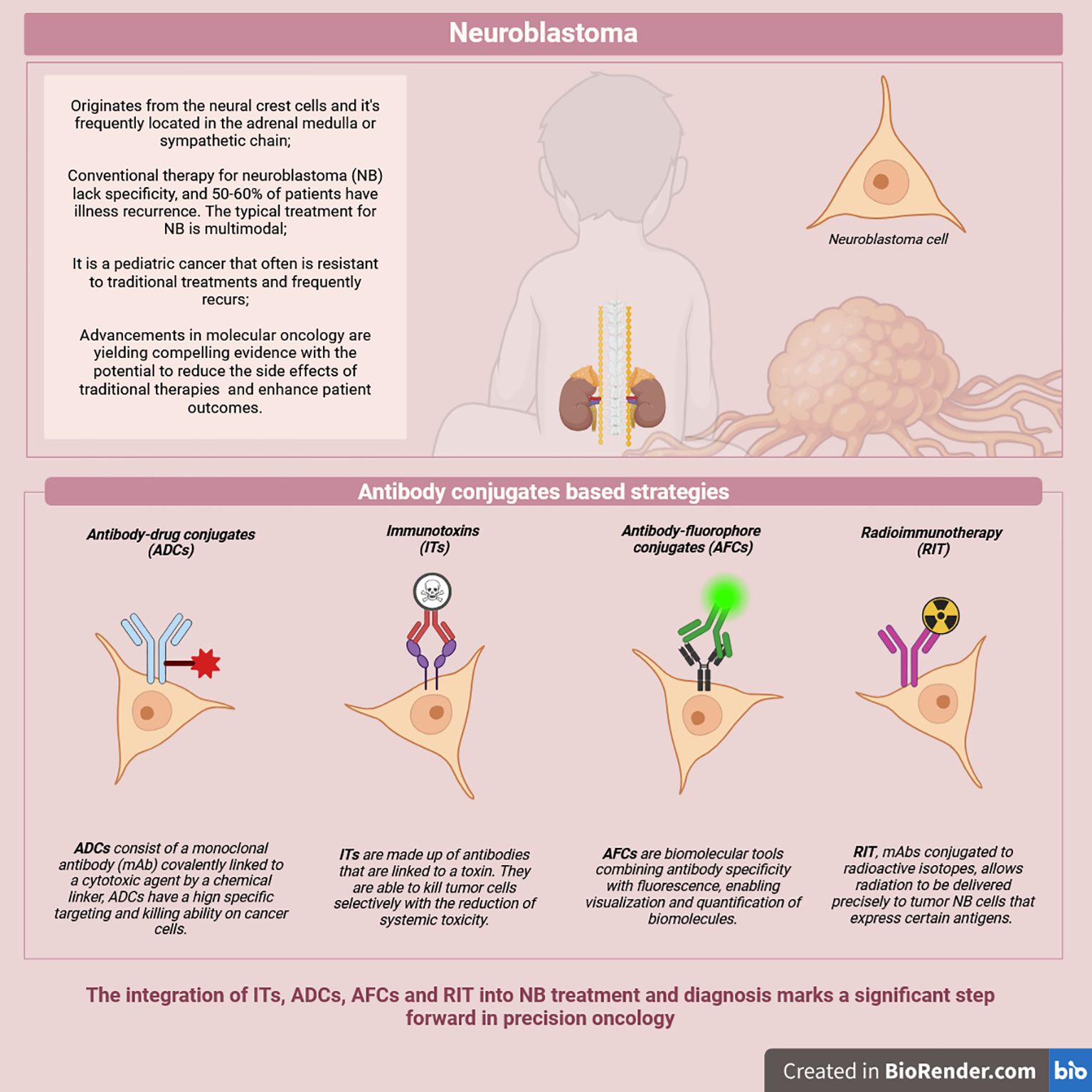
Figure 1. Overview of possible antibody-conjugates based strategies.
ACs are advanced biopharmaceutical tools designed for targeted cancer diagnosis and therapy. Conjugating antibodies to a toxin, a cytotoxic drug, a radioactive isotope, or labeling molecules such as fluorescent dyes allow specific binding to targets exclusively expressed on tumorous cells for diagnosis, monitoring, or treatment (8–12) (Figure 1).
While the use of monoclonal antibodies (mAb) revolutionized cancer therapy, they are often unable to reach the same level of efficacy displayed by classical treatments such as chemotherapy (13–15). This was successfully addressed by the development of ADCs. These drug-delivery systems are comprised of a mAb, a chemical linker, and a cytotoxic compound referred to as the payload. ADCs composition allows it to combine both the specificity of mAb and the potent cytotoxicity of the conjugated payload to meticulously achieve precise tumor eradication, avoiding both adverse effects and treatment resistance typically seen in classical cancer therapies (15, 16). ADCs gained clinical relevance with FDA approval of Gemtuzumab ozogamicin (Mylotarg®) in 2000. Since then, 14 more ADCs have been approved, underscoring their growing role in precision cancer treatment (15). Understanding NB’s complex biology is vital for precision medicine. Targeting key genetic alterations like ALK mutations and MYCN amplification with ADCs can improve outcomes. MYCN, a member of the MYC oncogene family, drives NB tumorigenesis by forming a transcriptionally active heterodimer with MAX, making it a therapeutic target (17–19). Bibbo et al. advanced Omomyc, a protein disrupting MYC-MAX interaction, by conjugating it to a HER3-targeting mAb (EV20) (20–22). This ADC, using a bifunctional linker, ensured Omomyc delivery only inside tumor cells, blocking MYC-MAX formation (22). In a pseudo-metastatic NB mouse model, EV20/Omomyc significantly reduced liver metastases, supporting its efficacy in managing NB metastasis and pediatric cancers (22) (Table 1). On the other hand, the development of an ADC targeting the ALK receptor offers significant advantages over traditional treatments for NB. ALK belongs to the insulin tyrosine kinase family and promotes mitogenic and anti-apoptotic effects by activating several downstream signaling pathways. These include phospholipase C-γ (PLCγ), janus kinase 3 (JAK3)-STAT3, extracellular signal-regulated kinase 1/2 (ERK1/2), and phosphatidylinositol 3-kinase (PI3K)-Akt. Mutations in ALK, such as F1174L and R1275Q, lead to constitutive activation of pro-survival pathways or receptor amplification, driving sustained tumor growth, cell proliferation, and migration. This associates the receptor with poor prognosis, making it a critical target for therapeutic intervention (23, 24). Research data shows that the ADC, CDX-0125-thienoindole (TEI), exhibited efficient antigen binding and internalization and demonstrated cytotoxicity in various NB cell lines (25). In vivo, CDX-0125-TEI significantly reduced tumor growth in NB mouse models, effectively targeting both wild-type (WT) and mutant ALK in patient-derived xenografts while sparing healthy cells due to its specificity (25). This dual efficacy against WT and mutant ALK broadens its applicability (Table 1). Besides focusing on key genetic aberrations in NB, precise therapy also leverages the targeting of surface proteins that are either overexpressed or uniquely expressed in cancer cells. One such target is B7-H3, a type I transmembrane glycoprotein linked to metastasis and poor outcomes in several cancers, including NB. Its ability to suppress T-cell activation leads to the successful immune evasion of the tumor while its contribution to the prompting of pathways such as the PI3K/Akt and STAT3 pathways can enhance survival and proliferation, further increasing the tumor’s resistance (26, 27). The ADC m276-SL-PBD, targeting B7-H3, extended event-free survival (EFS), reduced tumor volume in NB xenograft models, and showed low toxicity, confirming its tolerability and potency (28) (Table 1). Vobramitamab duocarmazine (vobra duo) is an anti-B7-H3 ADC showing preclinical efficacy in various solid cancers (29) and is under clinical evaluation for advanced castration-resistant metastatic prostate cancer (30). It induces apoptosis in multiple human NB cell lines (31). In co-culture experiments, B7-H3-unreactive murine NB NX-S2-luc cells showed reduced survival due to the ADC’s strong bystander activity. Using orthotopic, pseudo-metastatic, and tumor-resected NB mouse models, vobra duo delayed tumor growth and improved survival (31) (Table 1). Compared to Topotecan-Temozolomide (TOTEM), vobra duo delayed tumor relapse more effectively. Combined with TOTEM, it further slowed tumor progression and increased survival, enhancing conventional therapy (31). Repeated treatments in relapsed NB mice demonstrated superior efficacy and reduced off-target effects compared to TOTEM (31). Another ADC targeting a surface receptor, the Leucine-rich repeat-containing G-protein-coupled receptor 5 (LGR5), was assessed for its anti-NB action both in vitro and in vivo (32). LGR5 is a stem cell marker that is found to be overexpressed in various malignancies (33, 91). It enhances tumor cell survival, migration, and drug resistance in NB by potentiating the WNT signaling pathway (34). LGR5 was identified as a target for the treatment of NB thanks to its high expression in NB cell lines, as well as in primary tumor samples from NB patients (32). This has inspired the development of the anti-LGR5 ADC conjugated to pyrrolobenzodiazepine (PBD), a DNA alkylator. Assessment of the latter in vitro and in vivo using clinically relevant doses in mouse models xenografted with LGR5-positive human NB cell lines and PDX has led to the inhibition of tumor growth. This was attributed to a reduction in proliferation as well as an increase in apoptosis (32). Glypican 2 (GPC2), an MYCN-regulated oncoprotein overexpressed in NB cells, is associated with reduced overall survival and event-free survival (35, 36). In NB-PDX mouse models treated with a GPC2-targeting ADC (D3-GPC2-PBD) for 20 weeks, no tumor recurrence or weight changes were observed (37). In advanced NB models with larger tumors, D3-GPC2-PBD showed effective penetration and induced apoptosis. Similar results related to prolonged tumor regression, survival, and no recurrence were seen in PDX models with diverse NB-related mutations (MYCN, ALK, TP53) (37). The ADC’s bystander-killing effect was confirmed in co-cultures of responsive and resistant cell lines, showing cytotoxicity across all cells (37). The use of the same ADC in another study led to immunogenic cell death (ICD) both in vitro and in vivo by decreasing NB cell viability and increasing biomarker expression (38). Mice inoculated with NB cells pretreated with D3-GPC2-PBD exhibited slower tumor growth and better survival after tumor rechallenge with naïve NB cells, linked to T cell activation, a proinflammatory tumor microenvironment, enhanced immune cell infiltration, and increased phagocytosis (38). D3-GPC2-PBD also boosted CD40 expression on antigen-presenting cells (APCs). Combining the ADC with CD40 agonists or CD47 blockade further reduced tumor burden, improved survival, enhanced T cell infiltration, and induced long-term immunological memory, protecting against rechallenge (38). This highlights ADCs’ dual cytotoxic and immunoregulatory effects, as well as their potentiating effect when combined with immunotherapy.
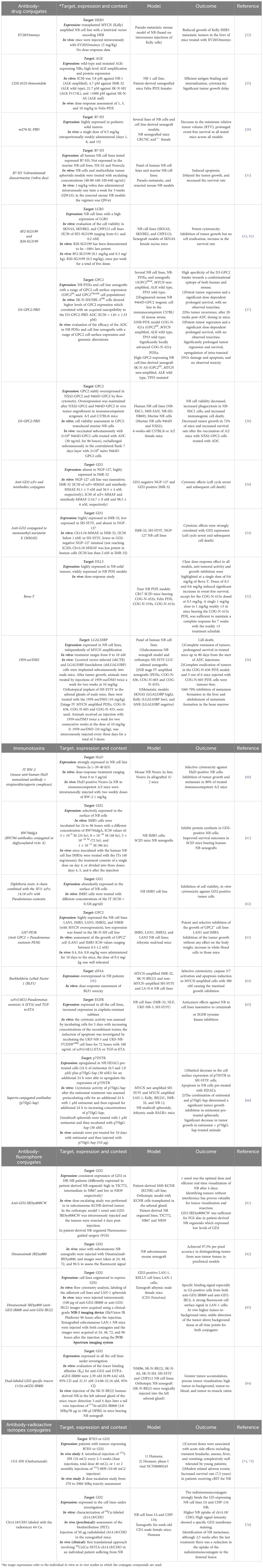
Table 1. Antibody conjugates-based strategies in the treatment and diagnosis of neuroblastoma.
The beneficial use of ADCs as a therapeutic option for NB is in part summarized in its specific drug delivery that singles out tumor cells expressing the targeted antigen, circumventing problems such as unwanted side effects and treatment resistance that are observed with conventional therapy (39). Taking into account that NB is a pediatric cancer with a median age of diagnosis of 17 months (1, 40), prolonged treatments, drug resistance, off-target effects, and any subsequent organ damage can lead to short- or long-term health and growth complications in children (41, 42), making ADCs an optimal therapeutic choice. Nevertheless, treating solid tumors with ADCs is more challenging. The high intertumoral variability and NB’s cellular heterogeneity, particularly in target expression levels, can represent an obstacle in ADC-oriented therapies (43, 44). Dense vascularization, a complex tumor microenvironment, and the large molecular weight of ADC antibodies further limits penetration and efficacy (45, 46). To address these issues, miniaturized antibodies have been developed to improve ADC diffusion and effectiveness in solid tumors, including NB (15). Ganglioside (GD2) is a sialic acid-containing glycosphingolipid that can be used as a biomarker for NB as it is highly expressed on NB cell surface, especially in HR-NB (47). Anti-GD2mAbs such as dinutuximab and naxitamab, have been extensively used as part of therapeutic protocols for the treatment of patients with HR-NB (48, 49). However, to overcome the limitations linked to the antibody’s size, two antibody fragment formats were developed: a single-chain variable fragment (scFv) from the ch14.18 antibody and a minibody combining scFv with an IgG1 heavy chain (50). These were conjugated to tubulin inhibitors, forming fragment-drug conjugates (FDCs). In GD2-positive IMR32 NB cells, FDCs showed cytotoxicity, with minibodies outperforming scFv due to superior kinetics and bivalent binding, enabling efficient payload delivery, cell cycle arrest, and cell death (50) (Table 1). Intertumoral differences in antigen expression also play a crucial role in determining the efficacy of ADCs in NB. A study exploring the neuroendocrine characteristics of NB targeted the delta-like ligand 3 (DLL3), an inhibitory Notch ligand selectively upregulated in neuroendocrine tumors where it potentiates tumor proliferation and invasion by activating the Notch and the SNAI1/Snail signaling pathways (51–53). The study demonstrated that Rova-T, an anti-DLL3 ADC previously proven to be ineffective in small cell lung cancer (SCLC), exhibited preclinical therapeutic potency in NB-PDX models. Nonetheless, this efficacy was closely dependent on DLL3 expression, emphasizing the need to restrict its application to NB cases with high DLL3 levels. This underscores the critical impact of antigen expression variability across tumors in dictating responsiveness to ADCs (52) (Table 1).
The impact of NB intertumoral heterogeneity and ADC composition on therapy has been studied extensively (44). Anti-GPC2 ADCs with different payloads showed enhanced internalization in NB cells with dense GPC2 expression. However, TP53-mutated NB cells exhibited resistance to ADCs with DNA binding and DNA topoisomerase 1 inhibiting payload. MYCN-amplified cells were more sensitive to ADCs with N-acetyl-calicheamicin g1 compared to non-MYCN-amplified cells, while cells with high ATP-binding cassette transporter B1 (ABCB1) expression, driven by MYCN, resisted tubulin inhibitor payloads. Notably, ALK-related mutations did not influence resistance to ADCs, regardless of payload type (44). The link between payload selection and NB heterogeneity was studied using monomethyl auristatin E (MMAE) and F (MMAF) as payloads (54). The efficacy of this ADC was evaluated across various NB cell lines with varying GD2 expressions, showing cytotoxicity positively correlated with GD2 levels (54). However, payload type also influenced outcomes; anti-GD2 conjugated with MMAE was more cytotoxic in SH-SY5Y cells than IMR-32 cells (54) (Table 1). These findings underscore the critical roles of NB heterogeneity, antigen density, and payload choice in ADC therapeutic efficacy. The tumor microenvironment plays a critical role in ADC efficacy as well, particularly through components like extracellular vesicles (EVs), which contribute to NB progression by transferring DNA, RNA, and proteins from tumor to healthy cells, promoting metastasis (55). Leveraging its high expression on EVs, Galectin-3 binding protein (LGALS3BP) was targeted by 1959-sss/DM3, a non-internalizing ADC shown to bind EVs effectively and yield therapeutic benefits in NB models. This ADC demonstrated efficacy in SKNAS subcutaneous NB xenografts, orthotopic SH-SY5Y-LUC adrenal xenografts, and patient-derived stage 4 NB xenografts (COG-N-636 and COG-N-603) with high LGALS3BP expression. Notably, tumor elimination was achieved in both PDX models, with COG-N-636 exhibiting a longer recurrence-free period than COG-N-603 (56) (Table 1). Success with targeted therapies depends on the application context. For heterogeneous tumors like NB, ADCs must match the tumor’s molecular profile and microenvironment for optimal efficacy. Molecular profiling aids ADC design while targeting EVs and other non-tumor-cell elements offer new ways to combat NB progression. Key design factors-antigen, mAb size, linker, and payload-are crucial for ADC efficacy and clinical potential.
ADCs and ITs are therapeutic complexes created by binding a toxic element to a mAb. However, in ADCs, the mAb is conjugated to a payload consisting of a cytotoxic drug. This includes as mentioned before: tubulin inhibitors, DNA-damaging agents, or immune-stimulatory compounds (15, 57). In contrast, the conjugates in ITs are not drugs, but rather toxins, which are poisons that are bacterial, animal, or plant-based (11, 58). One promising therapeutic target is the HuD antigen, a neuronal-specific RNA binding protein that is highly expressed in both NB and SCLC cells and was shown to promote NB tumor growth and survival through suppressing apoptotic pathways and downregulating mTORC1 activity (59). The conjugation of an anti-HuD antibody with the ribosomal toxin saporin, forming the IT-BW-2, has shown selective cytotoxicity against HuD-positive NB cells (50). Another well-established target is GD2, overexpressed in NB cells. Antibodies like BW704, conjugated to deglycosylated ricin A (BW704dgA), inhibit protein synthesis in GD2-positive NB cells. In vitro studies demonstrated the potent cytotoxic effects of BW-2 at low concentrations, while BW704dgA improved survival outcomes in SCID mice bearing human NB xenografts (60, 61). ITs based on the anti-GD2 scFv 5F11 have also demonstrated high potency. This recombinant fusion protein combines the diphtheria toxin A chain with the 5F11 scFv, allowing it to directly kill GD2-positive tumor cells without relying on Fc-mediated mechanisms like ADCC or CMC. Similarly, a recombinant fusion of 14.18 scFv with Pseudomonas exotoxin A has shown elevated in vitro cytotoxicity against GD2-positive tumor cells (62). GPC2 has also been targeted by ITs. This was done by using single-domain antibodies that bind specifically to GPC2 and conjugating them with a truncated Pseudomonas exotoxin (PE38) to create chimeric proteins capable of delivering the toxin directly to NB cells. These ITs exhibited potent cytotoxicity in GPC2-positive NB cell lines at low concentrations (IC50 values of 0.5-1.2 nM) while sparing GPC2-low-expressing cells. In mouse models, the immunotoxin LH7-PE38 significantly inhibited tumor growth at 0.4 mg/kg with minimal side effects, though higher doses showed toxicity, underscoring the importance of dose optimization (63). Another promising therapeutic agent is Burkholderia Lethal Factor 1 (BLF1), which exhibits selective cytotoxicity against MYCN-amplified NB cells. By inhibiting eukaryotic initiation translation factor 4A (eIF4A), BLF1 suppresses the translation of oncogenic proteins, inducing apoptosis in cancer cells while sparing non-transformed cells. BLF1 demonstrated a 70% reduction in viability in MYCN-amplified cells, with minimal impact on non-MYCN-amplified cells. Furthermore, BLF1 downregulated critical proteins such as MYCN and CDK4 and increased caspase 3/7 activity threefold, highlighting its potential for targeted NB therapy with reduced side effects compared to conventional treatments (64). NB cells often express the epidermal growth factor receptor (EGFR), making them susceptible to EGFR-targeted therapies. Cisplatin-resistant NB cells express higher levels of EGFR, making them particularly susceptible to EGFR-targeted toxins such as scFv(14E1)-Pseudomonas exotoxin A (ETA) and TGF-α-ETA. These toxins, especially when combined with cisplatin, significantly enhanced apoptosis and reduced cell viability compared to monotherapies. This combination approach shows promise for addressing chemoresistant NB (65). Novel strategies that aim at rendering NB more vulnerable are garnering interest. Notably, combination therapy is becoming increasingly popular. Given that the success of ACs relies on targeting molecules that are over- or uniquely expressed on tumor cells, identifying novel targets of this nature is crucial for advancing the development of therapeutical options, especially ACs. Fittingly, previous studies have shown that NB cells are particularly ‘impressionable’ when it comes to cellular target overexpression. For instance, exposure of NB cells to entinostat, a histone deacetylase inhibitor (HDACi), has led to the upregulation of the neurotrophin receptor p75NTR (66). Targeting p75NTR with saporin-conjugated antibodies (p75IgG-Sap) effectively induced apoptosis in these pretreated cells, taking advantage of the heightened receptor expression (66) (Table 1). Therefore, the priming of NB cells with agents that can induce the overexpression of specific receptors can be strategically used to increase NB susceptibility to AC-based approaches. This synergistic effect can be further explored using other compounds, especially those used in conventional cancer therapy such as chemotherapeutic agents and cytokines, to explore their potential modulatory effect on the expression of pre-existing underutilized targets or the induction of novel ones that can then be identified and precisely targeted. In summary, ITs are a promising strategy for treating NB, offering precise tumor targeting while sparing normal tissues. Advances in delivery systems, enhanced antigen expression and mitigation of side effects continue to refine their clinical utility, making them a promising avenue for improving NB treatment outcomes.
Radioimmunotherapy (RIT) involves using mAbs conjugated to radioactive isotopes, enabling targeted delivery of radiation to tumor cells expressing specific antigens. This approach aims to maximize the eradication of tumors while minimizing damage to surrounding healthy tissues. Similarly to the other ACs, identifying suitable target antigens, that are highly expressed in NB cells but show limited expression in normal tissues is a critical component of RIT. With GD2 being a notable target for ACs as previously described, the anti-GD2 mAb, such as 3F8 facilitates the direct delivery of therapeutic agents to NB cells. It is important to emphasize that these drugs may act as radiosensitizers. When appropriately timed, radiolabeled mAbs can be most effective when tumor cells are radiosensitive during the cell cycle phases. This rationale underpins the combination of chemotherapy and RIT (67–72). Radiolabeled anti-GD2 antibodies, such as 131I-3F8 and 131I-14G2a, have demonstrated efficacy in targeting NB cells and have been utilized for radioimmunodetection. Phase I and II studies have evaluated the toxicity, biological distribution, and dosage of intravenously administered 131I-3F8 in NB patients, revealing side effects such as pain and allergic reactions. In cases of leptomeningeal NB, 131I-3F8 was delivered intraventricularly, with no long-term toxicities observed (73). Metastatic NB remains one of the most challenging forms of NB to treat. Notably, survival rates have improved for patients with recurrent NB treated with a rescue regimen incorporating RIT. This includes intrathecal delivery of 131I-labeled mAbs targeting GD2 or B7-H3 following surgery and radiation to address minimal residual disease. A study reported that 14 out of 21 patients treated with this RIT-based salvage regimen were alive 7 to 74 months after relapse, with the others remaining NB-free (74) (Table 1). Another study from the same group evaluated the intraventricular injection of the anti-B7-H3 murine mAb omburtamab. The findings indicated that omburtamab can be safely used for RIT, even in patients previously exposed to high doses of chemotherapy and radiation. Remarkably, 44% of patients with recurrent NB survived for 13–17 years following RIT, with only 13% experiencing another relapse (75) (Table 1). Preclinical studies have also emphasized the potential of the ch14.18/CHO antibody labeled with the radiotracer 64Cu for highly specific targeting of NB cells. Clinical PET/MR scans confirmed its ability to detect NB metastases and enhance GD2-directed immunotherapy. Importantly, tumor binding by the radiolabeled antibody was blocked by excess unlabeled antibodies, validating its specificity. This technique enables the detection of otherwise inaccessible lesions, improving treatment planning and predicting therapeutic efficacy (76) (Table 1). Despite these advancements, the integration of RIT in complex treatment protocols remains unexplored, with no clinical or preclinical studies combining these modalities. This reflects the experimental nature of these approaches and the complexity of developing a unified treatment regimen. Nevertheless, RIT holds significant promise for the treatment of NB, offering a targeted approach with reduced systemic toxicity. Continued research and clinical evaluation are essential to fully realize the potential of this therapeutic modality in pediatric oncology.
AFCs combine antibody specificity with fluorescence for biomolecule visualization and quantification. Introduced by Albert Coons in 1941, they transformed immunology and microscopy (77). AFCs are essential in imaging, diagnostics, and cancer research, aiding in detecting breast, ovarian, colorectal, and lung cancer (78). Their precision in targeting tumor-associated antigens enables accurate imaging of malignant tissues (79). Fluorophores, from dyes to quantum dots and NIR fluorophores, improve sensitivity and tissue penetration (80). AFCs play key roles in immunohistochemistry, fluorescence-guided surgery, and flow cytometry, with real-time imaging advancements enhancing therapies, including for NB.
Advancements in imaging techniques and probes are transforming NB surgery, offering promising strategies to improve tumor detection, resection precision, and outcomes for pediatric patients. One notable innovation is the anti-GD2-IRDye800CW fluorescent probe, which specifically binds to NB cells, enhancing fluorescence-guided surgery (FGS). Preclinical studies using orthotopic tumor models demonstrated its effectiveness in identifying tumors without interference from prior immunotherapy with dinutuximab-beta, a clinically approved antibody. This combination approach shows potential for pediatric clinical trials, supported by GD2 expression across NB stages and patient-derived organoid models. Anti-GD2-IRDye800CW has proven valuable for tumor visualization and resection, with the potential to improve survival rates for children with high-risk NB (81) (Table 1). Multispectral short-wave infrared (SWIR) fluorescence imaging is another promising development for FGS. Using NB-specific NIR-I probes like Dinutuximab IRDye800, SWIR imaging achieved 97.5% per-pixel accuracy in distinguishing tumor from non-tumor tissues in preclinical models. This precision was further enhanced through machine learning methods, including principal component analysis (PCA) combined with the k-nearest neighbor (KNN) algorithm, which classified tumor tissues with 97.1% accuracy and background areas with 99.2% accuracy (82). SWIR imaging outperforms NIR-I imaging, offering higher tumor-to-background ratios (4.6 vs. 2.6), detecting small tumor volumes (0.9 mm³), and penetrating up to 3 mm into tissues. These capabilities underscore its potential to improve surgical precision by minimizing residual malignancy (83). In addition, dual-labeled GD2-specific tracers, such as 111In-αGD2-IR800, have been evaluated for intraoperative molecular imaging (IMI). In NB xenograft models, these tracers demonstrated high tumor-to-background, tumor-to-blood, and tumor-to-muscle ratios, facilitating precise tumor visualization with imaging tools like the Neoprobe and SPY-PHI NIR camera. This enabled the identification of residual disease and complete tumor excision. Compared to control tracers, 111In-αGD2-IR800 exhibited significantly increased tumor accumulation and negligible uptake in non-target organs, highlighting its sensitivity and specificity for NB detection (84) (Table 1). Organoid-based platforms are also advancing personalized FGS probe development. These platforms identified tumor-specific membrane targets, such as GD2 and L1CAM, for NB and breast cancer (BC) using patient-derived organoids (PDO) and xenograft models. By analyzing RNA sequencing datasets and verifying limited expression in healthy tissues, the study identified five potential NB probes and underscored the need for additional BC probes to address tumor heterogeneity. These findings suggest that organoid-based platforms can improve FGS accuracy by reducing false positives and ensuring robust tumor labeling (85). Collectively, these imaging innovations-from fluorescent and radiolabeled probes to SWIR imaging and organoid-based platforms-represent a transformative leap forward in NB surgery. They enhance tumor visualization, guide precise resections, and enable tailored therapeutic approaches. By addressing current challenges in detecting and excising tumors, these technologies promise to reduce residual malignancies, improve surgical precision, and ultimately enhance outcomes for children with this challenging disease.
Advancements in ACs, including ADCs, ITs, RIT, and AFCs, have significantly improved the precision in the treatment and diagnosis of NB by delivering cytotoxic agents and labeling molecules directly to tumor cells. NB’s heterogeneous nature, its diverse molecular profile, and complex microenvironments require careful consideration when selecting or designing the appropriate ACs to maximize therapeutic efficacy and diagnostic reliability. As of now, no ADCs, RIT or ITs have been FDA-approved specifically for NB management. Finding the adequate conditions of ACs’ conception and application in NB are still investigated preclinically, as previously reported above. Nonetheless, some ACs are currently in clinical trials (NCT06041516; NCT06224855; NCT00089245 (Table 1) and NCT06395103). Although it is the most studied subclass of ACs, only one clinical study investigating ADCs has been completed. The latter reported limited efficacy of the ADC Lorvotuzumab mertansine, with 25% of the treated NB patients showing stable disease progression through 6 cycles of treatment (86). Partial to complete delayed responses were only observed, in a reserved manner, in the case of Rhabdomyosarcoma and synovial sarcoma (86). On the other hand, the reliance on ADCs that have previously demonstrated efficacy in other malignancies is a good ground to start upon. For instance, the clinical testing of Zilovertamab vedotin (NCT06395103) has a strong rationale thanks to its targeting of the receptor tyrosine kinase-like orphan receptor 1 (ROR1), a surface protein that is overexpressed in NB and that plays a crucial role in its oncogenesis, with high levels of its mRNA correlating with poor prognosis, as it activates several signaling pathways that promote survival, invasion and metastasis (87, 88). Its previous efficacy in treating lymphoid cancers makes it a potentially good candidate for the treatment of other malignancies (89). Nevertheless, Zilovertamab vedotin, as well as the efficacy of other ACs, remains to be proven in the case of NB, as several challenges related to NB’s nature and ACs delivery within the tumor’s microenvironment continue to persist. Further advancements in ACs design, combined with deeper insights into NB-specific molecular targets, offer hope for improved efficacy and higher transability into clinical settings. Recent research has also highlighted the potential of cytokine-based treatments and epigenetic drugs to sensitize NB cells and render them more vulnerable to these targeted therapies. However, no approved therapeutic strategies currently combine these antibody conjugates with standard chemotherapy for aggressive NB, emphasizing the need for further research to validate these innovations in combination therapy and clinical trials to improve outcomes for HR-NB patients by addressing CR and systemic toxicity.
JB: Investigation, Supervision, Writing – original draft, Writing – review & editing. CS: Writing – original draft, Resources, Visualization. MS: Resources, Visualization, Writing – original draft. PF: Resources, Visualization, Writing – original draft. SD: Visualization, Writing – original draft, Conceptualization, Investigation, Supervision, Validation, Writing – review & editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Nong J, Su C, Li C, Wang C, Li W, Li Y, et al. Global, regional, and national epidemiology of childhood neuroblastoma (1990-2021): a statistical analysis of incidence, mortality, and DALYs. EClinicalMedicine. (2024) 79:102964. doi: 10.1016/j.eclinm.2024.102964
2. Lerone M, Ognibene M, Pezzolo A, Martucciello G, Zara F, Morini M, et al. Molecular genetics in neuroblastoma prognosis. Children (Basel). (2021) 8:456. doi: 10.3390/children8060456
3. Huang M, Weiss WA. Neuroblastoma and MYCN. Cold Spring Harb Perspect Med. (2013) 3:a014415. doi: 10.1101/cshperspect.a014415
4. Tucker ER, Poon E, Chesler L. Targeting MYCN and ALK in resistant and relapsing neuroblastoma. Cancer Drug Resist. (2019) 2:803–12. doi: 10.20517/cdr.2019.009
5. Zafar A, Wang W, Liu G, Wang X, Xian W, McKeon F, et al. Molecular targeting therapies for neuroblastoma: Progress and challenges. Med Res Rev. (2021) 41:961–1021. doi: 10.1002/med.21750
6. Matthay KK, George RE, Yu AL. Promising therapeutic targets in neuroblastoma. Clin Cancer Res. (2012) 18:2740–53. doi: 10.1158/1078-0432.CCR-11-1939
7. Zhou X, Wang X, Li N, Guo Y, Yang X, Lei Y. Therapy resistance in neuroblastoma:Mechanisms and reversal strategies. Front Pharmacol. (2023) 14:1114295. doi: 10.3389/fphar.2023.1114295
8. Dumontet C, Reichert JM, Senter PD, Lambert JM, Beck A. Antibody-drug conjugates come of age in oncology. Nat Rev Drug Discovery. (2023) 22:641–61. doi: 10.1038/s41573-023-00709-2
9. Phuna ZX, Kumar PA, Haroun E, Dutta D, Lim SH. Antibody-drug conjugates: Principles and opportunities. Life Sci. (2024) 347:122676. doi: 10.1016/j.lfs.2024.122676
10. Knutson S, Raja E, Bomgarden R, Nlend M, Chen A, Kalyanasundaram R, et al. Development and evaluation of a fluorescent antibody-drug conjugate for molecular imaging and targeted therapy of pancreatic cancer. PloS One. (2016) 11:e0157762. doi: 10.1371/journal.pone.0157762
11. Turner MA, Lwin TM, Amirfakhri S, Nishino H, Hoffman RM, Yazaki PJ, et al. The use of fluorescent anti-CEA antibodies to label, resect and treat cancers: A review. Biomolecules. (2021) 11:1819. doi: 10.3390/biom11121819
12. Dosio F, Brusa P, Cattel L. Immunotoxins and anticancer drug conjugate assemblies: the role of the linkage between components. Toxins (Basel). (2011) 3:848–83. doi: 10.3390/toxins3070848
13. Shefet-Carasso L, Benhar I. Antibody-targeted drugs and drug resistance–challenges and solutions. Drug Resist Updat. (2015) 18:36–46. doi: 10.1016/j.drup.2014.11.001
14. Carter PJ, Lazar GA. Next generation antibody drugs: pursuit of the ‘high-hanging fruit’. Nat Rev Drug Discovery. (2018) 17:197–223. doi: 10.1038/nrd.2017.227
15. Fu Z, Li S, Han S, Shi C, Zhang Y. Antibody drug conjugate: the “biological missile” for targeted cancer therapy. Sig Transduct Target Ther. (2022) 7:1–25. doi: 10.1038/s41392-022-00947-7
16. Tonon G, Rizzolio F, Visentin F, Scattolin T. Antibody drug conjugates for cancer therapy: from metallodrugs to nature-inspired payloads. Int J Mol Sci. (2024) 25:8651. doi: 10.3390/ijms25168651
17. Kohl NE, Kanda N, Schreck RR, Bruns G, Latt SA, Gilbert F, et al. Transposition and amplification of oncogene-related sequences in human neuroblastomas. Cell. (1983) 35:359–67. doi: 10.1016/0092-8674(83)90169-1
18. Müller M, Trunk K, Fleischhauer D, Büchel G. MYCN in neuroblastoma: The kings’ new clothes and drugs. EJC Paediatric Oncol. (2024) 4:100182. doi: 10.1016/j.ejcped.2024.100182
19. Wenzel A, Schwab M. The mycN/max protein complex in neuroblastoma. Short review. Eur J Cancer. (1995) 31A:516–9. doi: 10.1016/0959-8049(95)00060-V
20. Massó-Vallés D, Soucek L. Blocking myc to treat cancer: reflecting on two decades of omomyc. Cells. (2020) 9:883. doi: 10.3390/cells9040883
21. Sala G, Rapposelli IG, Ghasemi R, Piccolo E, Traini S, Capone E, et al. EV20, a novel anti- erbB-3 humanized antibody, promotes erbB-3 down-regulation and inhibits tumor growth in vivo. Transl Oncol. (2013) 6:676–84. doi: 10.1593/tlo.13475
22. Bibbò S, Capone E, Lovato G, Ponziani S, Lamolinara A, Iezzi M, et al. EV20/Omomyc: A novel dual MYC/HER3 targeting immunoconjugate. J Control Release. (2024) 374:171–80. doi: 10.1016/j.jconrel.2024.08.009
23. George RE, Sanda T, Hanna M, Fröhling S, Luther W, Zhang J, et al. Activating mutations in ALK provide a therapeutic target in neuroblastoma. Nature. (2008) 455:975–8. doi: 10.1038/nature07397
24. Carpenter EL, Mossé YP. Targeting ALK in neuroblastoma–preclinical and clinical advancements. Nat Rev Clin Oncol. (2012) 9:391–9. doi: 10.1038/nrclinonc.2012.72
25. Sano R, Krytska K, Larmour CE, Raman P, Martinez D, Ligon GF, et al. An antibody-drug conjugate directed to the ALK receptor demonstrates efficacy in preclinical models of neuroblastoma. Sci Transl Med. (2019) 11:eaau9732. doi: 10.1126/scitranslmed.aau9732
26. Bottino C, Vitale C, Dondero A, Castriconi R. B7-H3 in pediatric tumors: far beyond neuroblastoma. Cancers (Basel). (2023) 15:3279. doi: 10.3390/cancers15133279
27. Zhang H, Zhang J, Li C, Xu H, Dong R, Chen CC, et al. Survival association and cell cycle effects of B7H3 in neuroblastoma. J Korean Neurosurg Soc. (2020) 63:707–16. doi: 10.3340/jkns.2019.0255
28. Kendsersky NM, Lindsay J, Kolb EA, Smith MA, Teicher BA, Erickson SW, et al. The B7-H3-targeting antibody-drug conjugate m276-SL-PBD is potently effective against pediatric cancer preclinical solid tumor models. Clin Cancer Res. (2021) 27:2938–46. doi: 10.1158/1078-0432.CCR-20-4221
29. Scribner JA, Brown JG, Son T, Chiechi M, Li P, Sharma S, et al. Preclinical development of MGC018, a duocarmycin-based antibody-drug conjugate targeting B7-H3 for solid cancer. Mol Cancer Ther. (2020) 19:2235–44. doi: 10.1158/1535-7163.MCT-20-0116
30. Jang S, Powderly JD, Spira AI, Bakkacha O, Loo D, Bohac GC, et al. Phase 1 dose escalation study of MGC018, an anti-B7-H3 antibody-drug conjugate (ADC), in patients with advanced solid tumors. JCO. (2021) 39:2631–1. doi: 10.1200/JCO.2021.39.15_suppl.2631
31. Brignole C, Calarco E, Bensa V, Giusto E, Perri P, Ciampi E, et al. Antitumor activity of the investigational B7-H3 antibody-drug conjugate, vobramitamab duocarmazine, in preclinical models of neuroblastoma. J Immunother Cancer. (2023) 11:e007174. doi: 10.1136/jitc-2023-007174
32. Di Giannatale A, Dias-Gastellier N, Devos A, Mc Hugh K, Boubaker A, Courbon F, et al. Phase II study of temozolomide in combination with topotecan (TOTEM) in relapsed or refractory neuroblastoma: A European Innovative Therapies for Children with Cancer-SIOP-European Neuroblastoma study. Eur J Cancer. (2014) 50:170–7. doi: 10.1016/j.ejca.2013.08.012
33. Tu J, Toh Y, Aldana AM, Wen JJ, Wu L, Jacob J, et al. Antitumor activity of a pyrrolobenzodiazepine antibody-drug conjugate targeting LGR5 in preclinical models of neuroblastoma. Pharmaceutics. (2024) 16:943. doi: 10.3390/pharmaceutics16070943
34. Clark-Corrigall J, Myssina S, Michaelis M, Cinatl J, Ahmed S, Carr-Wilkinson J. Elevated expression of LGR5 and WNT signaling factors in neuroblastoma cells with acquired drug resistance. Cancer Invest. (2023) 41:173–82. doi: 10.1080/07357907.2022.2136682
35. Li N, Spetz MR, Ho M. The role of glypicans in cancer progression and therapy. J Histochem Cytochem. (2020) 68:841–62. doi: 10.1369/0022155420933709
36. Bosse KR, Raman P, Zhu Z, Lane M, Martinez D, Heitzeneder S, et al. Identification of GPC2 as an oncoprotein and candidate immunotherapeutic target in high-risk neuroblastoma. Cancer Cell. (2017) 32:295. doi: 10.1016/j.ccell.2017.08.003
37. Raman S, Buongervino SN, Lane MV, Zhelev DV, Zhu Z, Cui H, et al. A GPC2 antibody-drug conjugate is efficacious against neuroblastoma and small-cell lung cancer via binding a conformational epitope. Cell Rep Med. (2021) 2:100344. doi: 10.1016/j.xcrm.2021.100344
38. Pascual-Pasto G, McIntyre B, Shraim R, Buongervino SN, Erbe AK, Zhelev DV, et al. GPC2 antibody-drug conjugate reprograms the neuroblastoma immune milieu to enhance macrophage- driven therapies. J Immunother Cancer. (2022) 10:e004704. doi: 10.1136/jitc-2022-004704
39. Alessi I, Caroleo AM, de Palma L, Mastronuzzi A, Pro S, Colafati GS, et al. Short and long-term toxicity in pediatric cancer treatment: central nervous system damage. Cancers. (2022) 14:1540. doi: 10.3390/cancers14061540
40. Mahapatra S, Challagundla KB. Neuroblastoma. In: StatPearls. StatPearls Publishing, Treasure Island (FL (2023).
41. Roman J, Villaizán CJ, García-Foncillas J, Salvador J, Sierrasesúmaga L. Growth and growth hormone secretion in children with cancer treated with chemotherapy. J Pediatr. (1997) 131:105–12. doi: 10.1016/S0022-3476(97)70132-X
42. Hovi L, Saarinen-Pihkala UM, Vettenranta K, Lipsanen M, Tapanainen P. Growth in children with poor-risk neuroblastoma after regimens with or without total body irradiation in preparation for autologous bone marrow transplantation. Bone Marrow Transplant. (1999) 24:1131–6. doi: 10.1038/sj.bmt.1702021
43. Ngan ESW. Heterogeneity of neuroblastoma. Oncoscience. (2015) 2:837–8. doi: 10.18632/oncoscience.v2i10
44. Buongervino S, Lane MV, Garrigan E, Zhelev DV, Dimitrov DS, Bosse KR. Antibody-drug conjugate efficacy in neuroblastoma: role of payload, resistance mechanisms, target density, and antibody internalization. Mol Cancer Ther. (2021) 20:2228–39. doi: 10.1158/1535-7163.MCT-20-1034
45. Singh AP, Guo L, Verma A, Wong GGL, Thurber GM, Shah DK. Antibody coadministration as a strategy to overcome binding-site barrier for ADCs: a quantitative investigation. AAPS J. (2020) 22:28. doi: 10.1208/s12248-019-0387-x
46. Xu S. Internalization, trafficking, intracellular processing and actions of antibody-drug conjugates. Pharm Res. (2015) 32:3577–83. doi: 10.1007/s11095-015-1729-8
47. Balis FM, Busch CM, Desai AV, Hibbitts E, Naranjo A, Bagatell R, et al. The ganglioside, GD2, as a circulating tumor biomarker for neuroblastoma. Pediatr Blood Cancer. (2020) 67:e28031. doi: 10.1002/pbc.28031
48. Mora J. Dinutuximab for the treatment of pediatric patients with high-risk neuroblastoma. Expert Rev Clin Pharmacol. (2016) 9:647–53. doi: 10.1586/17512433.2016.1160775
50. Kalinovsky DV, Kholodenko IV, Kibardin AV, Doronin II, Svirshchevskaya EV, Ryazantsev DY, et al. Minibody-based and scFv-based antibody fragment-drug conjugates selectively eliminate GD2-positive tumor cells. Int J Mol Sci. (2023) 24:1239. doi: 10.3390/ijms24021239
51. Peddio A, Pietroluongo E, Lamia MR, Luciano A, Caltavituro A, Buonaiuto R, et al. DLL3 as a potential diagnostic and therapeutic target in neuroendocrine neoplasms: A narrative review. Crit Rev Oncol Hematol. (2024) 204:104524. doi: 10.1016/j.critrevonc.2024.104524
52. Krytska K, Casey CE, Pogoriler J, Martinez D, Rathi KS, Farrel A, et al. Evaluation of the DLL3-targeting antibody-drug conjugate rovalpituzumab tesirine in preclinical models of neuroblastoma. Cancer Res Commun. (2022) 2:616–23. doi: 10.1158/2767-9764.CRC-22-0137
53. Serrano AG, Rocha P, Freitas Lima C, Stewart A, Zhang B, Diao L, et al. Delta-like ligand 3 (DLL3) landscape in pulmonary and extra-pulmonary neuroendocrine neoplasms. NPJ Precis Oncol. (2024) 8:268. doi: 10.1038/s41698-024-00739-y
54. Kalinovsky DV, Kibardin AV, Kholodenko IV, Svirshchevskaya EV, Doronin II, Konovalova MV, et al. Therapeutic efficacy of antibody-drug conjugates targeting GD2-positive tumors. J Immunother Cancer. (2022) 10:e004646. doi: 10.1136/jitc-2022-004646
55. Dhamdhere MR, Spiegelman VS. Extracellular vesicles in neuroblastoma: role in progression, resistance to therapy and diagnostics. Front Immunol. (2024) 15:1385875. doi: 10.3389/fimmu.2024.1385875
56. Capone E, Lamolinara A, Pastorino F, Gentile R, Ponziani S, Di Vittorio G, et al. Targeting vesicular LGALS3BP by an antibody-drug conjugate as novel therapeutic strategy for neuroblastoma. Cancers (Basel). (2020) 12:2989. doi: 10.3390/cancers12102989
57. Lambert JM, Morris CQ. Antibody-drug conjugates (ADCs) for personalized treatment of solid tumors: A review. Adv Ther. (2017) 34:1015–35. doi: 10.1007/s12325-017-0519-6
58. Janthur WD, Cantoni N, Mamot C. Drug conjugates such as Antibody Drug Conjugates (ADCs), immunotoxins and immunoliposomes challenge daily clinical practice. Int J Mol Sci. (2012) 13:16020–45. doi: 10.3390/ijms131216020
59. Bishayee K, Habib K, Nazim U, Kang J, Szabo A, Huh SO, et al. RNA binding protein HuD promotes autophagy and tumor stress survival by suppressing mTORC1 activity and augmenting ARL6IP1 levels. J Exp Clin Cancer Res. (2022) 41:18. doi: 10.1186/s13046-021-02203-2
60. Ehrlich D, Wang B, Lu W, Dowling P, Yuan R. Intratumoral anti-HuD immunotoxin therapy for small cell lung cancer and neuroblastoma. J Hematol Oncol. (2014) 7:91. doi: 10.1186/s13045-014-0091-3
61. Manzke O, Russello O, Leenen C, Diehl V, Bohlen H, Berthold F. Immunotherapeutic strategies in neuroblastoma: antitumoral activity of deglycosylated Ricin A conjugated anti-GD2 antibodies and anti-CD3xanti-GD2 bispecific antibodies. Med Pediatr Oncol. (2001) 36:185–9. doi: 10.1002/1096-911X(20010101)36:1<185::AID-MPO1044>3.0.CO;2-J
62. Tur MK, Sasse S, Stöcker M, Djabelkhir K, Huhn M, Matthey B, et al. An anti-GD2 single chain Fv selected by phage display and fused to Pseudomonas exotoxin A develops specific cytotoxic activity against neuroblastoma derived cell lines. Int J Mol Med. (2001) 8:579–84. doi: 10.3892/ijmm.8.5.579
63. Li N, Fu H, Hewitt SM, Dimitrov DS, Ho M. Therapeutically targeting glypican-2 via single-domain antibody-based chimeric antigen receptors and immunotoxins in neuroblastoma. Proc Natl Acad Sci United States America. (2017) 114:E6623. doi: 10.1073/pnas.1706055114
64. Rust A, Shah S, Hautbergue GM, Davletov B. Burkholderia lethal factor 1, a novel anti-cancer toxin, demonstrates selective cytotoxicity in MYCN-amplified neuroblastoma cells. Toxins (Basel). (2018) 10:261. doi: 10.3390/toxins10070261
65. Michaelis M, Bliss J, Arnold SC, Hinsch N, Rothweiler F, Deubzer HE, et al. Cisplatin-resistant neuroblastoma cells express enhanced levels of epidermal growth factor receptor (EGFR) and are sensitive to treatment with EGFR-specific toxins. Clin Cancer Res. (2008) 14:6531–7. doi: 10.1158/1078-0432.CCR-08-0821
66. Dedoni S, Olianas A, Manconi B, Collu M, Tuveri B, Vincis ME, et al. Upregulation of p75NTR by histone deacetylase inhibitors sensitizes human neuroblastoma cells to targeted immunotoxin-induced apoptosis. Int J Mol Sci. (2022) 23:3849. doi: 10.3390/ijms23073849
67. Goldenberg DM. Targeted therapy of cancer with radiolabeled antibodies. J Nucl Med. (2002) 43:693–713.
68. Sharkey RM, Goldenberg DM. Perspectives on cancer therapy with radiolabeled monoclonal antibodies. J Nucl Med. (2005) 46 Suppl 1:115S–27S.
69. Gold DV, Modrak DE, Schutsky K, Cardillo TM. Combined 90Yttrium-DOTA-labeled PAM4 antibody radioimmunotherapy and gemcitabine radiosensitization for the treatment of a human pancreatic cancer xenograft. Int J Cancer. (2004) 109:618–26. doi: 10.1002/ijc.v109:4
70. Stein R, Chen S, Reed L, Richel H, Goldenberg DM. Combining radioimmunotherapy and chemotherapy for treatment of medullary thyroid carcinoma: effectiveness of dacarbazine. Cancer. (2002) 94:51–61. doi: 10.1002/cncr.10157
71. Milenic DE, Baidoo KE, Shih JH, Wong KJ, Brechbiel MW. Evaluation of platinum chemotherapy in combination with HER2-targeted α-particle radiation. Cancer Biother Radiopharm. (2013) 28:441–9. doi: 10.1089/cbr.2012.1423
72. Blumenthal RD, Leone E, Goldenberg DM. Tumor-specific dose scheduling of bimodal radioimmunotherapy and chemotherapy. Anticancer Res. (2003) 23:4613–9.
73. Modak S, Kramer K, Pandit-Taskar N. Radioimmunotherapy of Neuroblastoma. In: Baum RP, editor. Therapeutic Nuclear Medicine. Medical Radiology. Springer, Berlin, Heidelberg (2013). p. 629–38.
74. Kramer K, Kushner BH, Modak S, Pandit-Taskar N, Smith-Jones P, Zanzonico P, et al. Compartmental intrathecal radioimmunotherapy: results for treatment for metastatic CNS neuroblastoma. J Neurooncol. (2010) 97:409–18. doi: 10.1007/s11060-009-0038-7
75. Kramer K, Pandit-Taskar N, Kushner BH, Zanzonico P, Humm JL, Tomlinson U, et al. Phase 1 study of intraventricular 131I-omburtamab targeting B7H3 (CD276)-expressing CNS Malignancies. J Hematol Oncol. (2022) 15:165. doi: 10.1186/s13045-022-01383-4
76. Schmitt J, Schwenck J, Maurer A, Przybille M, Sonanini D, Reischl G, et al. Translational immunoPET imaging using a radiolabeled GD2-specific antibody in neuroblastoma. Theranostics. (2022) 12:5615–30. doi: 10.7150/thno.56736
77. Coons AH, Creech HJ, Jones RN, Berliner E. The demonstration of pneumococcal antigen in tissues by the use of fluorescent antibody. J Immunol. (1942) 45:159–70. doi: 10.4049/jimmunol.45.3.159
78. Parit S, Manchare A, Gholap AD, Mundhe P, Hatvate N, Rojekar S, et al. Antibody-Drug Conjugates: A promising breakthrough in cancer therapy. Int J Pharm. (2024) 659:124211. doi: 10.1016/j.ijpharm.2024.124211
79. Warram JM, de Boer E, Sorace AG, Chung TK, Kim H, Pleijhuis RG, et al. Antibody-based imaging strategies for cancer. Cancer Metastasis Rev. (2014) 33:809–22. doi: 10.1007/s10555-014-9505-5
80. Nagaya T, Nakamura YA, Choyke PL, Kobayashi H. Fluorescence-guided surgery. Front Oncol. (2017) 7:314. doi: 10.3389/fonc.2017.00314
81. Wellens LM, Deken MM, Sier CFM, Johnson HR, de la Jara Ortiz F, Bhairosingh SS, et al. Anti-GD2-IRDye800CW as a targeted probe for fluorescence-guided surgery in neuroblastoma. Sci Rep. (2020) 10:17667. doi: 10.1038/s41598-020-74464-4
82. Waterhouse DJ, Privitera L, Anderson J, Stoyanov D, Giuliani S. Enhancing intraoperative tumor delineation with multispectral short-wave infrared fluorescence imaging and machine learning. J BioMed Opt. (2023) 28:094804. doi: 10.1117/1.JBO.28.9.094804
83. Privitera L, Waterhouse DJ, Preziosi A, Paraboschi I, Ogunlade O, Da Pieve C, et al. Shortwave infrared imaging enables high-contrast fluorescence-guided surgery in neuroblastoma. Cancer Res. (2023) 83:2077–89. doi: 10.1158/0008-5472.CAN-22-2918
84. Rosenblum LT, Sever RE, Gilbert R, Guerrero D, Vincze SR, Menendez DM, et al. Dual-labeled anti-GD2 targeted probe for intraoperative molecular imaging of neuroblastoma. J Transl Med. (2024) 22:940. doi: 10.1186/s12967-024-05728-0
85. Jeremiasse B, van Ineveld RL, Bok V, Kleinnijenhuis M, de Blank S, Alieva M, et al. A multispectral 3D live organoid imaging platform to screen probes for fluorescence guided surgery. EMBO Mol Med. (2024) 16:1495–514. doi: 10.1038/s44321-024-00084-4
86. Geller JI, Pressey JG, Smith MA, Kudgus RA, Cajaiba M, Reid JM, et al. ADVL1522: A phase 2 study of lorvotuzumab mertansine (IMGN901) in children with relapsed or refractory wilms tumor, rhabdomyosarcoma, neuroblastoma, pleuropulmonary blastoma, Malignant peripheral nerve sheath tumor, or synovial sarcoma-A Children’s Oncology Group study. Cancer. (2020) 126:5303–10. doi: 10.1002/cncr.v126.24
87. Quezada MJ, Lopez-Bergami P. The signaling pathways activated by ROR1 in cancer. Cell Signal. (2023) 104:110588. doi: 10.1016/j.cellsig.2023.110588
88. Dave H, Butcher D, Anver M, Bollard CM. ROR1 and ROR2-novel targets for neuroblastoma. Pediatr Hematol Oncol. (2019) 36:352–64. doi: 10.1080/08880018.2019.1646365
89. Wang ML, Barrientos JC, Furman RR, Mei M, Barr PM, Choi MY, et al. Zilovertamab vedotin targeting of ROR1 as therapy for lymphoid cancers. NEJM Evid. (2022) 1:EVIDoa2100001. doi: 10.1056/EVIDoa2100001
90. Skofler C, Kleinegger F, Krassnig S, Birkl-Toeglhofer AM, Singer G, Till H, et al. Eukaryotic translation initiation factor 4AI: A potential novel target in neuroblastoma. Cells. (2021) 10:301. doi: 10.3390/cells10020301
Keywords: neuroblastoma, antibody-drug conjugates, antibody-fluorophore conjugates, immunotoxin, precision medicine, radioimmunotherapy
Citation: Balla J, Siddi C, Scherma M, Fadda P and Dedoni S (2025) Antibody conjugates in neuroblastoma: a step forward in precision medicine. Front. Oncol. 15:1548524. doi: 10.3389/fonc.2025.1548524
Received: 19 December 2024; Accepted: 07 February 2025;
Published: 10 March 2025.
Edited by:
Hugo de Jonge, University of Pavia, ItalyReviewed by:
Charalampos Theocharopoulos, National and Kapodistrian University of Athens, GreeceCopyright © 2025 Balla, Siddi, Scherma, Fadda and Dedoni. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Simona Dedoni, dedoni@unica.it
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.