 Chuanyou Xia1†
Chuanyou Xia1† Thale Kristin Olsen2†
Thale Kristin Olsen2† A. Ali Zirakzadeh3Radwa Almamoun4Louise K. Sjöholm4Jenny Dahlström1Jan Sjöberg1Hans-Erik Claesson1
A. Ali Zirakzadeh3Radwa Almamoun4Louise K. Sjöholm4Jenny Dahlström1Jan Sjöberg1Hans-Erik Claesson1 John Inge Johnsen2Ola Winqvist3
John Inge Johnsen2Ola Winqvist3 Dawei Xu1
Dawei Xu1 Tomas J. Ekström4Magnus Björkholm1*‡
Tomas J. Ekström4Magnus Björkholm1*‡ Klas Strååt1*‡
Klas Strååt1*‡- 1Department of Medicine, Division of Hematology, BioClinicum and Centre for Molecular Medicine, Karolinska University Hospital Solna and Karolinska Institutet, Stockholm, Sweden
- 2Childhood Cancer Research Unit, Department of Women’s and Children’s Health, Karolinska Institutet, Stockholm, Sweden
- 3Unit of Translational Immunology, Department of Medicine, Karolinska Institutet, Stockholm, Sweden
- 4Department of Clinical Neuroscience, Karolinska Institutet, Center for Molecular Medicine, Stockholm, Sweden
We studied DNA methylation profiles in four different cell populations from a unique constellation of monozygotic triplets in whom two had developed Hodgkin Lymphoma (HL). We detected shared differences in DNA methylation signatures when comparing the two HL-affected triplets with the non-affected triplet. The differences were observed in naïve B-cells and marginal zone-like B-cells. DNA methylation differences were also detected when comparing each of the HL-affected triplets against each other. Even though we cannot determine whether treatment and/or disease triggered the observed differences, we believe our data are important on behalf of forthcoming studies, and that it might provide important clues for a better understanding of HL pathogenesis.
Introduction
We previously reported Hodgkin lymphoma (HL) development in two monozygotic triplets where all three were homozygotic through a constitutional deletion in the first intron of the megakaryoblastic leukemia 1 gene (MKL1) (1). Two of the HL affected triplets were diagnosed with HL at the age of 40 and 63, respectively. Clinical details of the triplets are presented in reference (1) and in supplementary material. The significance of the MKL1 gene deletion in the same set of triplets has recently been studied further by Record et al (2). In this paper, we showed that lymphoblastoid cell lines derived from the HL-free triplet had higher MKL1 mRNA and protein levels when compared to the HL affected triplets, while cells derived from the HL treated triplets resembled the healthy controls used in the study. Additionally, we could show that cells from the HL-free sibling had increased cell proliferation, a larger number of hyperploid cells, and further, that cells derived from this triplet were able to form large tumors in vivo. This indicates that the HL-free sibling has pre-malignant cells emerging while the HL-affected triplets do not, possibly due to previous treatment. Together, these data suggest that dysregulated MKL1 activity may participate in B cell transformation and HL pathogenesis (2). After our initial report of the HL triplet cases (1), we have not seen any other publications addressing the role of MKL1 deletions in HL pathogenesis.
It is well recognized that first-degree relatives of HL-patients carry an increased risk of developing HL (3) and that the risk increases in monozygotic twins (4). HL tumors have unique features since the tumor tissue only consists of a minute number of the actual cancer cells which are called Hodgkin/Reed-Sternberg (HRS) cells. These cancerous cells are believed to originate from a germinal or post germinal center B-cell. The rest of the tumor microenvironment mainly harbors inflammatory cells of the immune system, such as lymphocytes, eosinophils, neutrophils, histocytes, plasma cells, and dendritic cells, reviewed in (5, 6).
DNA methylation is thought to play an important role in carcinogenesis. Hypermethylation in CpG islands of promoters is assumed to facilitate the silencing of gene transcription. Indeed, hypermethylation of B-cell specific genes such as CD79B, BOB1, SYK, PU.1, CD19, and CD20 is implicated in HL (7).
Previous studies carried out on these triplets used peripheral blood mononuclear cells (PBMCs) collected from whole blood with no specific selection of cells (1), or isolated CD19+ B-cells transformed into lymphoblastoid cells (2).
Methods
PBMCs were isolated using density centrifugation with Ficoll-Paque (GE Healthcare). The number of cells was counted before they were stored in AIM V medium (Gibco) in +4°C overnight. CD34+ cells were sorted out by positive selection using the CD34+ isolation kit (Miltenyi). After CD34+ cell separation, B-cells were sorted from the PBMCs using the CD20+ positive selection isolation kit (Miltenyi), naïve B-cells, marginal zone-like B cells, and switched memory B-cells were sorted using FACS Aria (BD). Genomic DNA (gDNA) was isolated from CD34+ cells and naïve, marginal zone-like and switched memory B-cells using the DNeasy Blood & Tissue Kit (Qiagen) in accordance with the manufacturer’s instructions.
Subsequently, genome-wide DNA methylation was analyzed using the Illumina EPIC array with 850.000 CpG site-specific probes. Raw.idat files were analyzed using the minfi R package (version 1.28.4) (8). Beta and M values were extracted and preprocessed using the preprocessFunnorm command. Loci with known SNPs and/or detection p values > 0.01 were filtered from subsequent analyses. For each of the four cell types, pairwise comparisons between individuals (A vs. B, A vs. C, B vs. C) were made using the DMRforPairs R package (version 1.18.0) (9). Results were then cross-checked to identify all regions that were differentially methylated between the non-affected triplet sample C and each of the HL-affected samples A and B. In the analysis, default settings were used as described in the DMRforPairs vignette. Microarray data are available at GEO under accession number GSE142202.
The study was approved by the local ethics committee, Dnr: 2008/1764-31, and informed consent was given before samples were collected. Methods and bioinformatic analysis in more detail can be found in the supplementary material.
Results
Unsupervised hierarchical clustering demonstrated that the four cell types studied were distinct from each other in terms of methylation levels (Figure 1A). This cell type distinction was also reflected in the principal component analysis (PCA) presented in (Figure 1C). When comparing the methylation profile of each HL-triplet against the non-HL-triplet, shared differences in DNA methylation were found in naïve B-cells and marginal zone-like B-cells (Figure 1B). In contrast, we found no shared changes in DNA methylation in switched memory B-cells or CD34+ cells. A list of genes located in close proximity to these differentially methylated CpG loci is presented in Table 1.
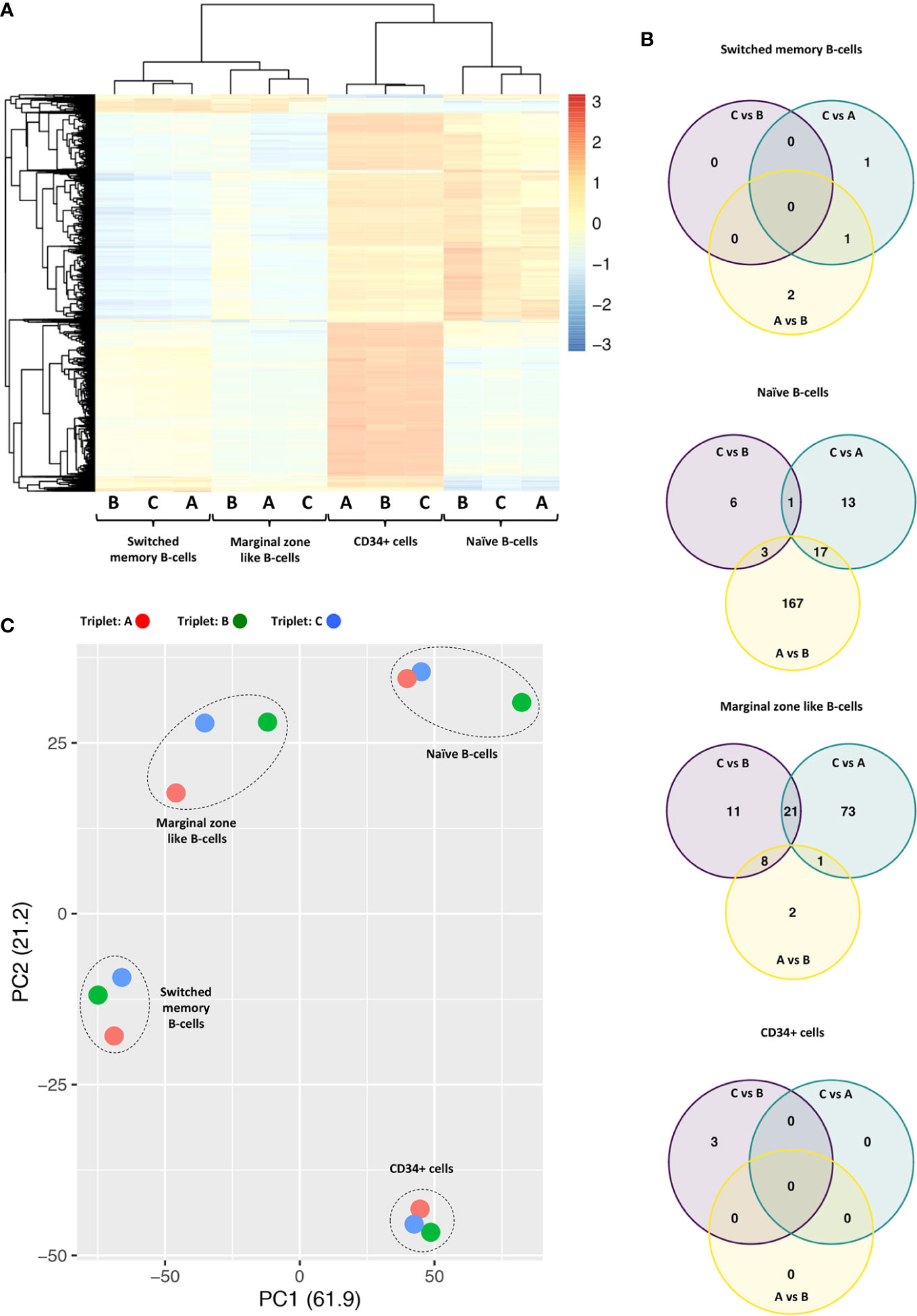
Figure 1 (A) Unsupervised hierarchical clustering of methylation levels from the 10,000 most variable probes across all samples derived from HL-affected triplets A, B, and non-affected triplet C Probe variability is quantified by standard deviation. M values are scaled by row (z score); red color indicates M values above mean; blue color indicates M values below mean. Samples from the four cell types primarily cluster by cell type. (B) Venn diagrams showing overlaps of differentially methylated regions when each triplet is compared to the others in pairwise comparisons. Of particular interest is the overlap between the “C vs. B” and the “C vs. A” sets (n =1 for naïve B cells; n = 21 for marginal zone-like B cells). (C) Principal component analysis (PCA) of all samples. PC1 and PC2 explain 61.9% and 21.2% of variability, respectively. Each triplet is labeled by color and each cell type is indicated in the plot.
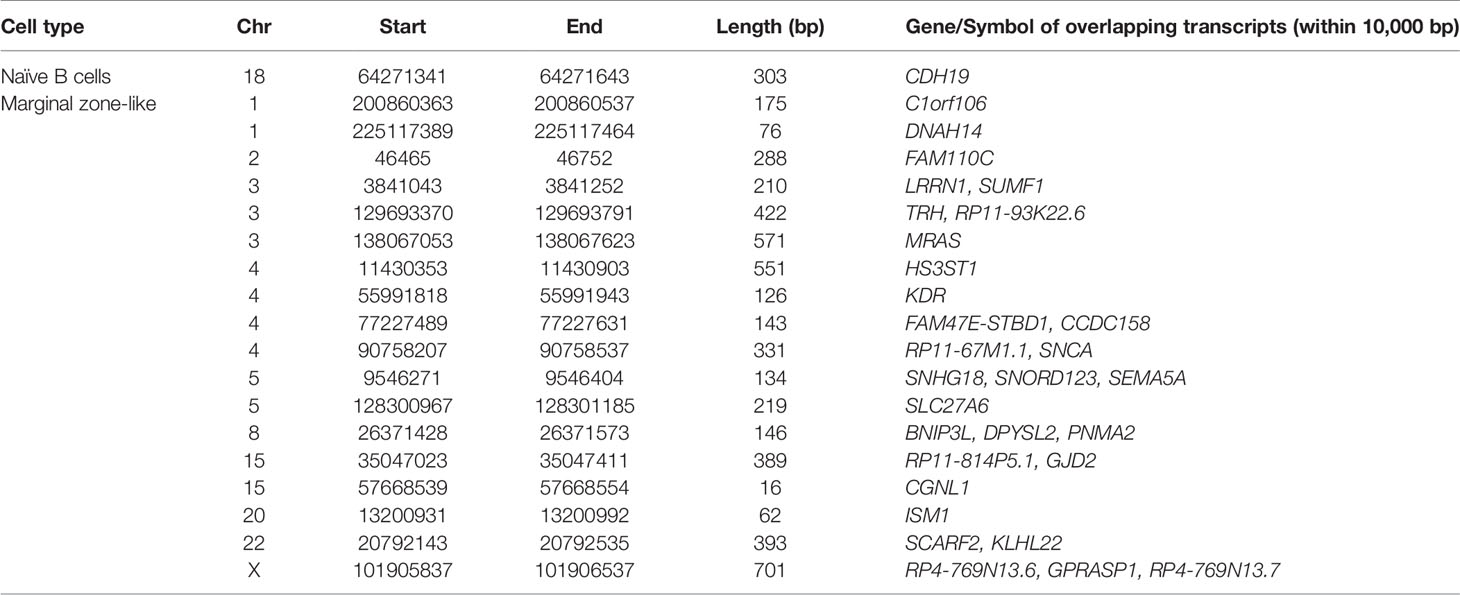
Table 1 List of genes with shared differences between the two HL-affected triplets as compared to the non-affected sibling.
In the naïve B-cell subpopulation, we found one region on chromosome 18 that differed in DNA methylation. Here, samples from both HL-affected triplets displayed hypermethylation when compared with the non-affected triplet. In marginal zone-like B-cells, we found 21 genomic regions on 10 different chromosomes which were all hypermethylated in both of the HL-affected triplets as compared to the non-affected triplet. In CD34+ and switched memory B cells, we found no common regions that differed in methylation signature (Figure 1B). The identified probe regions and corresponding genes in the studied cell populations are listed in detail in Table 1 and Supplementary Material.
Discussion
Several genes related to HL, cancer or hematopoiesis are located in or near the differentially methylated regions in marginal zone-like B-cells. For example, ISM1 has been detected in lymphocytes, bone marrow, and embryonic blood islands and found to contribute to the hematopoiesis in zebrafish (10). KLHL22 is likely involved in tumorigenesis through the activation of the amino-acid-dependent mTOR pathway (11). SNHG18 was identified to play as a tumor suppressor gene in hepatocellular carcinoma (12). Recent studies have shown that exogenous expression of SNCA through the transcription factor ΔNp63α could induce the migration of breast cancer cells (13). Down-regulation of SNCA is also associated with ovarian cancer drug resistance (14). MRAS is known to play a key role in tumor growth (15), and it contributes to the activation of several important signalling cascades such as the MAPK and ERK pathways. KDR is known to regulate angiogenesis, vascular development, vascular permeability, and embryonic hematopoiesis. In a cohort study (16), including 17 HL-prone families, a predisposing mutation (p.A 1065T) in the KDR gene was identified and suggested to enhance tumor proliferation.
We could also detect differences in DNA methylation when comparing the HL-affected triplets against each other. Similarly, we observed shared differences between the HL-affected triplet and the non-affected triplet (Figure 1B). Such differences might occur due to inter-individual variation, environmental factors, or stochastic events. Nevertheless, we believe that the results obtained here are of potential value in forthcoming studies related to DNA methylation and pathogenesis of HL.
The identified DNA hypermethylation could possibly govern a lower expression of affected genes. Recent studies suggest that inherited leukemia-associated somatic mutations might have a low impact on clonal hematopoiesis (CH) (17, 18), suggesting that DNA methylation has a larger impact on CH than anticipated. Thus, it would be of interest to further investigate DNA methylation in more detail in patients with hematological diseases. Studies of DNA methylation in patients with various hematological disorders will probably increase our understanding of their pathogenesis. Our and others’ results emphasize the importance of understanding the role of genome-wide DNA methylation in malignancies.
We are well aware that the study of this unique triplet constellation has its limitations regarding statistical strength. Even though the two HL patients A and B ended terminated their chemotherapy treatment 33 and 8 years, respectively, before samples were collected, we cannot fully rule out that chemotherapy has contributed to the observed differences in DNA methylation. This would be important to know since it would demonstrate that established methylation alterations in the adult can be sustained over very long periods.
The two HL triplets described in this study were 40 and 63 years of age at the time of diagnosis, and both cases were EBV positive (1). The familiar risk of HL in siblings as well as monozygotic twins is primarily associated with early-onset disease (3, 4). Although heritability of EBV-positive HL in older patients is not clearly established (19), we cannot rule out heritable factors contributing to disease in these cases.
There are studies indicating that chemotherapy has an effect on DNA-methylation. For example, a study on short-term (4 months) effects of chemotherapeutic treatment showed that leukocyte DNA-methylation was altered by chemotherapy in breast cancer patients (20). In this study, CpG islands in or near VMP1, CORO1B, SDK1, and SUMF2 were found to be significantly altered after chemotherapy. None of these genes were differentially methylated in our study. However, the SUMF2 paralog SUMF1 was hypermethylated in the marginal zone-like cells of HL affected triplets (Table 1). SUMF2 is known to inhibit the sulphatase-enhancing activity of SUMF1 (21). It is possible that chemotherapy might influence expression of SUMF2/SUMF1 and thus the regulation of sulphatase activity in cells. It will be of great interest to see if similar findings may be observed by others in twins or triplets discordant for the diagnosis of HL.
In summary, we report a unique constellation of monozygotic triplets where DNA methylation levels in marginal zone-like B-cells and naïve B-cells were observed to be different in two HL-affected triplets as compared to the non-affected triplet. These differences in DNA methylation levels may contribute to HL-pathogenesis.
Data Availability Statement
The original data presented in this study is publicly available at the Gene Expression Omnibus (GEO), accession number GSE142202.
Ethics Statement
The studies involving human participants were reviewed and approved by Etikprövningsnämnden Stockholm. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
CX interpreted data and wrote the manuscript. TO performed and interpreted bioinformatic analysis and took part in writing the manuscript. AZ and OW performed FACS sorting to isolate the different cell types analyzed in the study. RA and LS prepared DNA samples for analysis by Illumina EPIC. JS, H-EC, JJ, DX, and TE took part in interpreting data and reviewing the manuscript. MB, DX, and TE took part in the design of the study, interpreted data, and reviewed the manuscript. KS took part in the design of the study, interpreted obtained results as well as writing the manuscript and preparing figures and tables. All authors contributed to the article and approved the submitted version.
Funding
This study was supported by grants from Swedish Cancer Society, Cancer Society in Stockholm, Stockholm County Council and the Swedish Research Council. PhD student CX was partially supported by a PhD scholarship from Chinese Scholarship Council. The authors declare that this study received funding from Takeda Pharma AB. The funder was not involved in the study design, collection, analysis, interpretation of data, the writing of this article or the decision to submit it for publication.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Methylation profiling was performed by the SNP&SEQ Technology Platform in Uppsala (www.genotyping.se). The facility is part of the National Genomics Infrastructure (NGI) Sweden and Science for Life Laboratory. The SNP&SEQ Platform is also supported by the Swedish Research Council and the Knut and Alice Wallenberg Foundation.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fonc.2020.598872/full#supplementary-material
References
1. Björkholm M, Sjöberg J, Nygell UA, Porwit A, Björck E. Development of Hodgkin lymphoma in homozygotic triplets with constitutional deletion in MKL1. Blood (2013) 121:4807. doi: 10.1182/blood-2013-02-469031
2. Record J, Sendel A, Kritikou JS, Kuznetsov NV, Brauner H, He M, et al. An intronic deletion in megakaryoblastic leukemia 1 is associated with hyperproliferation of B cells in triplets with Hodgkin lymphoma. Haematologica (2020) 105:1339–50. doi: 10.3324/haematol.2019.216317
3. Goldin LR, Björkholm M, Kristinsson SY, Turesson I, Landgren O. Highly increased familial risks for specific lymphoma subtypes. Br J Haematol (2009) 146:91–4. doi: 10.1111/j.1365-2141.2009.07721.x
4. Mack TM, Cozen W, Shibata DK, Weiss LM, Nathwani BN, Hernandez AM, et al. Concordance for Hodgkin’s disease in identical twins suggesting genetic susceptibility to the young-adult form of the disease. N Engl J Med (1995) 332:413–8. doi: 10.1056/nejm199502163320701
5. Mathas S, Hartmann S, Küppers R. Hodgkin lymphoma: Pathology and biology. Semin Hematol (2016) 53:139–47. doi: 10.1053/j.seminhematol.2016.05.007
6. Carbone A, Gloghini A. Epstein Barr Virus-Associated Hodgkin Lymphoma. Cancers (Basel) (2018) 10(6):163–9. doi: 10.3390/cancers10060163
7. Ushmorov A, Leithäuser F, Sakk O, Weinhaüsel A, Popov SW, Möller P, et al. Epigenetic processes play a major role in B-cell-specific gene silencing in classical Hodgkin lymphoma. Blood (2006) 107:2493–500. doi: 10.1182/blood-2005-09-3765
8. Aryee MJ, Jaffe AE, Corrada-Bravo H, Ladd-Acosta C, Feinberg AP, Hansen KD, et al. Minfi: a flexible and comprehensive Bioconductor package for the analysis of Infinium DNA methylation microarrays. Bioinformatics (2014) 30:1363–9. doi: 10.1093/bioinformatics/btu049
9. Rijlaarsdam MA, van der Zwan YG, Dorssers LC, Looijenga LH. DMRforPairs: identifying differentially methylated regions between unique samples using array based methylation profiles. BMC Bioinf (2014) 15:141. doi: 10.1186/1471-2105-15-141
10. Berrun A, Harris E, Stachura DL. Isthmin 1 (ism1) is required for normal hematopoiesis in developing zebrafish. PloS One (2018) 13:e0196872. doi: 10.1371/journal.pone.0196872
11. Chen J, Ou Y, Yang Y, Li W, Xu Y, Xie Y, et al. KLHL22 activates amino-acid-dependent mTORC1 signalling to promote tumorigenesis and ageing. Nature (2018) 557:585–89. doi: 10.1038/s41586-018-0128-9
12. Liu XF, Thin KZ, Ming XL, Shuo L, Ping L, Man Z, et al. Small Nucleolar RNA Host Gene 18 Acts as a Tumor Suppressor and a Diagnostic Indicator in Hepatocellular Carcinoma. Technol Cancer Res Treat (2018) 17:1533033818794494. doi: 10.1177/1533033818794494
13. Dang TT, Westcott JM, Maine EA, Kanchwala M, Xing C, Pearson GW. ΔNp63α induces the expression of FAT2 and Slug to promote tumor invasion. Oncotarget (2016) 7:28592–611. doi: 10.18632/oncotarget.8696
14. Zou J, Yin F, Wang Q, Zhang W, Li L. Analysis of microarray-identified genes and microRNAs associated with drug resistance in ovarian cancer. Int J Clin Exp Pathol (2015) 8(6):6847–58.
15. Young LC, Hartig N, Muñoz-Alegre M, Oses-Prieto JA, Durdu S, Bender S, et al. An MRAS, SHOC2, and SCRIB complex coordinates ERK pathway activation with polarity and tumorigenic growth. Mol Cell (2013) 52:679–92. doi: 10.1016/j.molcel.2013.10.004
16. Rotunno M, McMaster ML, Boland J, Bass S, Zhang X, Burdett L, et al. Whole exome sequencing in families at high risk for Hodgkin lymphoma: identification of a predisposing mutation in the KDR gene. Haematologica (2016) 101:853–60. doi: 10.3324/haematol.2015.135475
17. Hansen JW, Pedersen DA, Larsen LA, Husby S, Clemmensen SB, Hjelmborg J, et al. Clonal hematopoiesis in elderly twins: concordance, discordance, and mortality. Blood (2020) 135:261–68. doi: 10.1182/blood.2019001793
18. Fabre MA, McKerrell T, Zwiebel M, Vijayabaskar MS, Park N, Wells PM, et al. Concordance for clonal hematopoiesis is limited in elderly twins. Blood (2020) 135:269–73. doi: 10.1182/blood.2019001807
19. Lin AY, Kingma DW, Lennette ET, Fears TR, Whitehouse JM, Ambinder RF, et al. Epstein-Barr virus and familial Hodgkin’s disease. Blood (1996) 88:3160–5. doi: 10.1182/blood.V88.8.3160.bloodjournal8883160
20. Yao S, Hu Q, Kerns S, Yan L, Onitilo AA, Misleh J, et al. Impact of chemotherapy for breast cancer on leukocyte DNA methylation landscape and cognitive function: a prospective study. Clin Epigenetics (2019) 11:45. doi: 10.1186/s13148-019-0641-1
Keywords: Hodgkin lymphoma, monozygotic triplets, DNA methylation, marginal zone-like B-cells, CD34+ cells, naïve B cells, switched memory B-cells
Citation: Xia C, Olsen TK, Zirakzadeh AA, Almamoun R, Sjöholm LK, Dahlström J, Sjöberg J, Claesson H-E, Johnsen JI, Winqvist O, Xu D, Ekström TJ, Björkholm M and Strååt K (2020) Hodgkin Lymphoma Monozygotic Triplets Reveal Divergences in DNA Methylation Signatures. Front. Oncol. 10:598872. doi: 10.3389/fonc.2020.598872
Received: 25 August 2020; Accepted: 09 November 2020;
Published: 09 December 2020.
Edited by:
Liren Qian, Sixth Medical Center of PLA General Hospital, ChinaReviewed by:
Thomas Mack, University of Southern California, United StatesGiancarlo Castellano, Institut de Recerca Biomèdica August Pi i Sunyer (IDIBAPS), Spain
Yi Miao, Nanjing Medical University, China
Copyright © 2020 Xia, Olsen, Zirakzadeh, Almamoun, Sjöholm, Dahlström, Sjöberg, Claesson, Johnsen, Winqvist, Xu, Ekström, Björkholm and Strååt. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Klas Strååt, a2xhcy5zdHJhYXRAa2kuc2U=; Magnus Björkholm, bWFnbnVzLmJqb3JraG9sbUBzbGwuc2U=
†These authors share first authorship
‡These authors share senior authorship