
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Nutr., 12 June 2023
Sec. Food Chemistry
Volume 10 - 2023 | https://doi.org/10.3389/fnut.2023.1186161
This article is part of the Research TopicPlant’s Bioactivity in Modern Health and Diet: Benefits, Limitations and Trendy ApplicationView all 5 articles
 Wang Cheng-yuan
Wang Cheng-yuan Dai Jian-gang*
Dai Jian-gang*Hyperuricemia is another common metabolic disease, which is considered to be closely related to the development of many chronic diseases, in addition to the “three highs.” Currently, although drugs show positive therapeutic effects, they have been shown to produce side effects that can damage the body. There is growing evidence that medicinal and edible plants and their bioactive components have a significant effect on hyperuricemia. In this paper, we review common medicinal and edible plants with uric acid-lowering effects and summarize the uric acid-lowering mechanisms of different bioactive components. Specifically, the bioactive components are divided into five categories: flavonoids, phenolic acids, alkaloids, polysaccharides, and saponins. These active substances exhibit positive uric acid-lowering effects by inhibiting uric acid production, promoting uric acid excretion, and improving inflammation. Overall, this review examines the potential role of medicinal and edible plants and their bioactive components as a means of combating hyperuricemia, with the hope of providing some reference value for the treatment of hyperuricemia.
Hyperuricemia (HUA) is a metabolic disease caused by abnormal purine metabolism in the body. Clinically, HUA is a higher-than-normal saturated concentration of urate in the blood in the fasting state. The standard for men is >7 mg/dL, for women >6 mg/dL, and for adolescents and children >5.5 mg/dL (1). Uric acid (UA) is synthesized by the body’s organs through the conversion of purines produced by their own nucleic acid metabolism and/or food purines (2). Excess uric acid can be deposited in the kidneys and synovial fluid to form monosodium urate crystals, causing inflammatory reactions and gout (3). With the change in people’s diet, the incidence of Hpegloticasepeople’s dietUA is increasing year by year in the world, especially in China and the United States (4, 5). Studies have found that high uric acid not only causes gout, but also causes other metabolic diseases, including cardiovascular disease, hypertension, and intestinal flora disorders (1, 6, 7).
It was found that UA can be oxidized by urate oxidase to 5-hydroxyurate, which in turn is converted to allantoin (8). However, humans and primates have evolved without urate oxidase and are therefore unable to break down uric acid into allantoin. Therefore, most UA is excreted only by the kidneys, with a very small proportion excreted by the intestine, and the excretion of UA often relies on the synergistic action of multiple uric acid transporters. For example, uric acid transporter 1 (URAT1), organic anion transporter 4 (OAT4), and glucose transporter 9 (GLUT9) mainly regulate UA reabsorption, while adenosine triphosphate-binding transporter protein G superfamily member 2 (ABCG2), organic anion transporter 1 (OAT1) and 3 (OAT3) are responsible for regulating renal UA excretion (9–11). It has been demonstrated that xanthine oxidase (XOD) catalyzes the oxidation of hypoxanthine and xanthine to uric acid and is a key enzyme in the control of UA production (12). However, the production and metabolism of UA are complex physiological processes. In addition to XOD, it also involves phosphoribosyl pyrophosphate synthetase (PRPS), hypoxanthine-guanine phosphoribosyl transferase (HGPRT), phosphoribosyl pyrophosphate amino-transferase (PRPPAT) (13). Figure 1 shows the factors affecting hyperuricemia. Therefore, reducing uric acid production by adjusting the activity of uric acid metabolism enzymes, increasing uric acid excretion by promoting the expression of UA transporters, and controlling the intake of foods containing high purine can achieve the goal of reducing uric acid. However, the current treatment for HUA is more through drug therapy, which can be divided into synthesis inhibitors and excretion promoters according to the formation mechanism of UA. The former includes Allopurinol and Febuxostat, the latter includes Propofol and Benzbromarone; oxidative breakdown of uric acid can also be achieved by ingestion of pegloticase and rasburicase. Although the above-mentioned drugs can positively affect the lowering of uric acid, they can also have some harmful effects on the body. It has been found that allopurinol and benzbromarone cause skin sensitization and nephrotoxicity, respectively, in clinical treatment (14, 15).
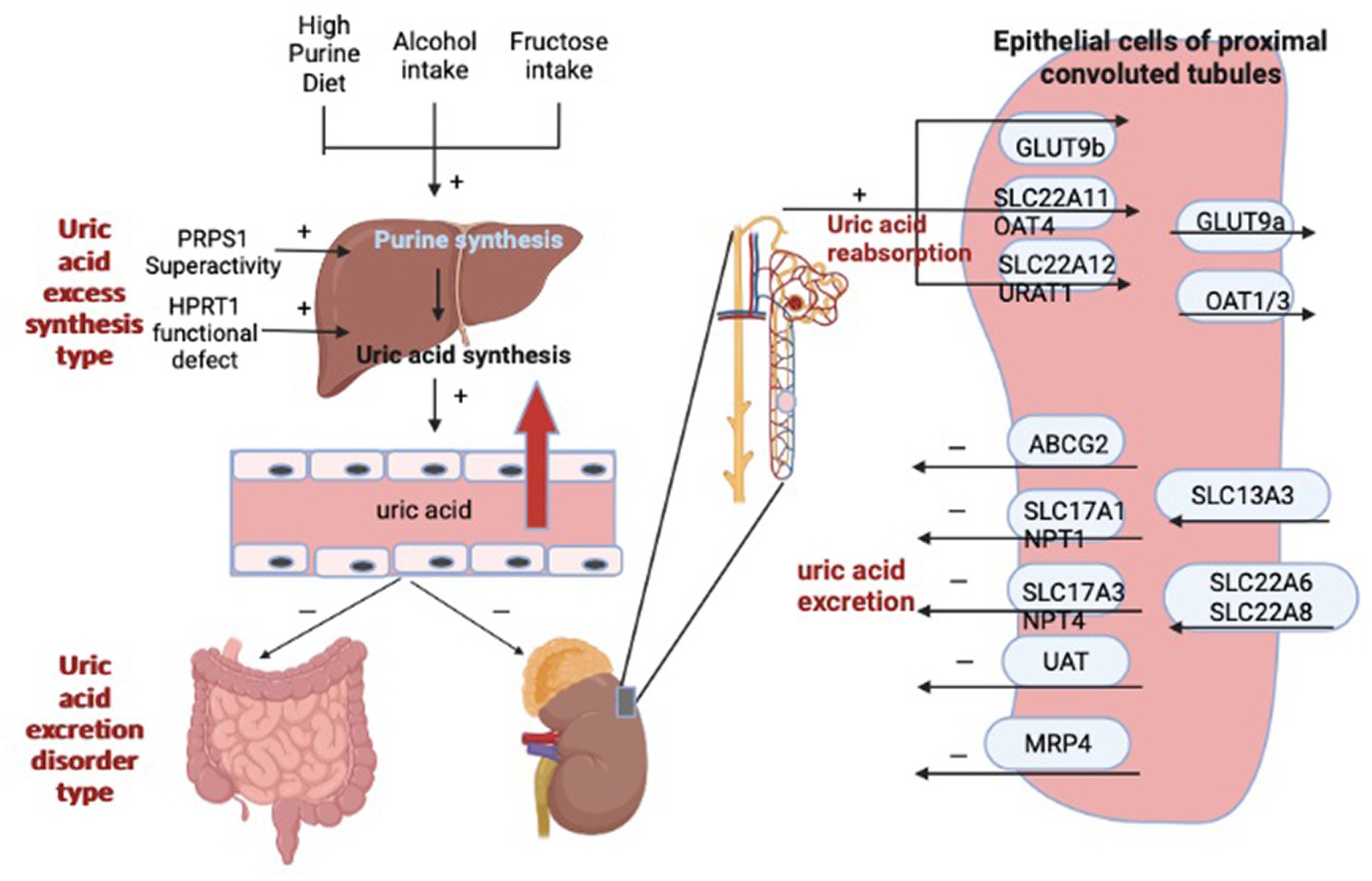
Figure 1. The factors influencing hyperuricemia.
Therefore, it is necessary and of great importance to find safer and more effective natural drugs to replace HUA therapeutic drugs.
Chinese medicine believes that food and medicine originated at the same time, hence the term medicine and food harmony. Drugs and food are relative: drugs are also food, while food is also drugs; food has fewer side effects, while drugs have more. Medicine and food harmony are scientifically combined for both food and medicine; they meet nutritional needs as well as the function of preventing and treating diseases (16). Interestingly, as research has progressed, many medicinal and edible plants have been linked to a reduction in the incidence of various diseases, such as diabetes, hypertension, and vascular disease (17–19). Therefore, there is an increasingly growing interest in the research and development of medicinal and food plants (20–22). In recent years, functional foods developed from medicinal and edible plants for the treatment of HUA have received greater attention. For example, it is found that chicory extract can significantly reduce serum uric acid levels as well as increase intestinal uric acid excretion in rats with high glucose-induced HUA (p < 0.05), which in turn was found to enhance the mRNA expression of ABCG2 and promote uric acid excretion (23). Nelumbinis folium leaf extract showed positive inhibition of XOD activity, thus showing potential for alternative drug treatment of HUA (24). This review highlights medicinal and edible plants with UA-lowering activity and their bioactive compounds for potential UA-lowering mechanisms. We expect that this paper will contribute to the understanding of the potential applications of medicinal and edible plants in the prevention and treatment of HUA.
Chinese quince (Chaenomeles sinensis) is often used in Chinese medicine to treat HUA and gout. Modern pharmacological studies have also shown that it contains many esters, acids, alcohols, aldehydes, ketones, heterocycles, and alkanes (25). Zhang et al. (26) mixed dried Chinese quinces with 30% ethanol and extracted them at 50°C for 1.5 h, followed by centrifugation at 5,000 rmp for 15 min, after which the crude extract was obtained by rotary evaporation of the solvent. Finally, the ethyl acetate fraction of Chaenomeles sinensis fruit extract (CSF-E) was obtained by AB-8 macroporous resin. They used the obtained CSF-E to treat HUA mice induced by potassium oxyzinate (PO) and showed that CSF-E significantly reduced XOD activity in the serum and liver of mice (p < 0.05). It also reduced serum UA, creatinine (CR), and blood urea nitrogen (BUN) levels, and further analysis revealed downregulation of URAT1 as well as upregulation of OAT1 protein expression in the kidney, which increased UA excretion.
Lemon (Citrus limon) is one of the best-known and most used species in the citrus genus Citrus, which contains high levels of citric acid, proven to have anti-inflammatory, anti-bacterial, anti-cancer, and anti-parasitic activities (27). A recent study found that 30 mL of freshly squeezed pure lemon juice administered daily for six weeks significantly reduced serum uric acid levels (p < 0.01), and the same results were found in PO- induced HUA mice (28). Lin et al. (29) found that the water-soluble extract of lemon significantly reduced the serum uric acid level in PO-induced HUA mice (p < 0.05) and hardly damaged the kidney. They further found that it was mainly the potassium citrate contained in lemons that exerted the uric acid-lowering effect. This study suggests that potassium citrate has the potential to be developed as a drug for the treatment of HUA.
In Chinese traditional medicine, Chicory (Cichorium intybus L.) is considered a diuretic and cholagogue, and its effects are often related to the bioactive substances it contains, such as polyphenols, inulin, oligofructose, and sesquiterpene lactones (30). As previously mentioned, chicory extract enhances uric acid excretion by down-regulating ABCG2 mRNA and up-regulating OAT3 mRNA expression in the kidney (23). Similarly, previous work has found that chicory powder has a positive effect on serum metabolites of UA, lipids, glucose, and abdominal fat deposition in a quail model induced by a purine-rich diet. Jin et al. (31) induced HUA rats by gavage of yeast (15 g/kg/d) and adenine (80 mg/kg/d). They found that chicory extracts enhanced 24 h UA excretion and CR clearance in rats, demonstrating that chicory extract significantly reduced serum UA and CR levels in rats (p < 0.05). A decrease in GLUT9 protein expression was also found in rat kidneys and human renal tubular epithelial cells (HKC cells) in vitro, demonstrating that chicory extract exerted a blood UA-lowering effect by affecting GLUT9 protein expression. Meng Bian’s team used network pharmacology to predict the possible components and corresponding mechanisms of the anti-HUA effect of chicory (32). They selected quail with the same UA metabolic pathway as humans to establish an HUA model to evaluate the potential anti-HUA mechanism of chicory extract. The results suggest that luteolin and β-sterols in chicory may be the key active components of chicory in lowering UA, and the mechanism involved in the regulation of lipolysis in adipocytes. After treatment with chicory extract, the serum UA levels of quail gradually decreased, and serum total cholesterol (TC), triglyceride (TG), and low-density lipoprotein-cholesterol (LDL-C) levels were significantly reduced (p < 0.05).
Tea (Camellia sinensis) originated in China, with coffee and cocoa being known as the world’s three major beverages. With the popularity of tea, more and more research is being done on tea. The various components and functions of tea are also gradually being explored, with the most researched being tea polyphenols and antioxidant properties (33–35). It is the presence of antioxidants in tea that can act as inhibitors of many enzymes, including XOD. It has been found that green tea extract can reduce serum UA levels and modulate UA gene transporters to increase renal uric acid clearance in HUA mice by inhibiting XOD enzyme activity (36). Additional studies have found that ester catechins in tea extracts exert stronger XOD inhibitory activity than non-ester catechins (37, 38). Li et al. (39) jointly verified through in vivo and in vitro experiments that epigallocatechin gallate (EGCG) showed significant anti-HUA effects by inhibiting XOD activity, reducing serum uric acid levels, enhancing renal OAT1 expression, and decreasing GLUT9 expression in rats. Other studies found that theaflavin was able to reduce serum BUN and CR values by inhibiting adenosine deaminase (ADA) and XOD activities, down-regulating gene and protein expression of GLUT9 and URAT1, and up-regulating gene expression of ABCG2, OCTN1, and OAT2, to improve hyperuricemic symptoms and renal damage in HUA mice (40). Putranty et al. (41) studied the effect of green tea extract on the kidney of HUA rats, and they found that 600 mg/kg of green tea extract reduced malondialdehyde (MDA) and CR levels in the kidney while repairing kidney damage. To investigate the effect of each tea on HUA, Sang et al. (42) used each of these six basic teas (Green Tea, White Tea, Yellow Tea, Oolong Tea, Black Tea, and Dark Tea) in HUA rats and interestingly, the results showed that all except green tea showed a lowering effect of serum UA, while yellow tea significantly improved HUA by modulating inflammatory response, autophagy, and apoptosis (p < 0.05). In the introductory section, we mentioned that HUA causes disturbances in the gut microbiota. Wu et al. (43) applied aqueous extracts of six teas (TWE) to HUA mice to study the changes in their UA, and found that a significant decrease in xanthine dehydrogenase (XDH) expression in the liver, enhanced ABCG2, OAT1, and OAT3 expression in the kidney, and reduced URAT1 expression in the kidney occurred after TWE intervention. Enhanced expression of ABCG2 in the gut and improved gut microbiota were also found to occur. There are many studies on tea to lower UA, but we can conclude that the gallic acid, polyphenols and theaflavins contained in tea mainly play the role of inhibiting XOD enzyme activity and lowering UA. The mechanisms involved are simply the down-regulation of the expression of genes involved in uric acid reabsorption, such as GLUT9 and URAT1, and the up-regulation of the expression of genes involved in uric acid transport or excretion, such as ABCG2, OAT1, and OAT3, the inhibition of XOD enzyme activity, and also the anti-HUA effect through the modulation of the inflammatory response or gut microbes (44–46).
Flos Sophorae (Sophora japonica Linn), the flower of the Chinese scholar tree, has remarkable biological activity and is used in several Asian countries to treat diseases such as bleeding hemorrhoids, hematuria, vomiting of blood, and inflammatory conditions (47). The main bioactive components in Flos Sophorae include kaempferol, quercetin, rutin, and isorhamnetin (48). Wang et al. (49) screened the XOD inhibitors of the main components of the aqueous extract of Flos Sophorae using HPLC-MS/MS and found that isorhamnetin significantly reduced serum uric acid levels and inhibited XOD activity, and also reduced serum CR and BUN levels and protected the kidneys. In addition, molecular docking showed that isorhamnetin binds well to XOD and is a promising XOD inhibitor. In a recent study, Li et al. (50) analyzed the active components in Flos Sophorae and also investigated the type and mechanism of XOD inhibition. They found that the IC50 of quercetin, kaempferol, isorhamnetin, rutin, and gibberellin in Flos Sophorae against XOD were 0.03, 0.11, 0.07, 5.62, 11.48, and 22.13 mg/mL, respectively. Only quercetin, isorhamnetin, and kaempferol were mixed competitive inhibitors and significantly inhibited the fluorescence intensity of XOD (p < 0.05).
Lotus leaf (Folium nelumbinis) is the dried leaf of Nelumbo nucifera Gaertn. It is used as a herbal medicine to relieve summer heat and spleen deficiency and diarrhea in traditional Chinese medical knowledge (51). Modern research has also found that lotus leaves can lower blood pressure, blood lipids, and blood sugar, and can be used as a new weight loss strategy (52, 53). Previously, we mentioned that the total alkaloids of lotus leaves could significantly inhibit XOD activity with an IC50 of 3.313 μg/mL. Roemerine were analyzed by UHPLC-Q-TOF-MS and 3D molecular docking to identify them as potential active substances (24). A recent study found that both crude extract and the total alkaloid fraction of lotus leaves showed positive XOD inhibitory effects. They inhibit UA production by reducing the mRNA and protein expression of hepatic XOD, and further analysis revealed that the mechanism is to down-regulate renal GLUT9 and URAT1 expression to inhibit uric acid reabsorption and up-regulate renal OAT1 to promote uric acid excretion (54).
Chrysanthemum flowers (Dendranthema morifolium) are not only used as ornamental flowers in urban landscapes but also as a traditional medicine or food for people in some Asian countries (55–57). After a long-term research, more than 190 chemical substances have been isolated and identified from chrysanthemum, mainly flavonoids, terpenoids, and phenolic acids, which exert antioxidant, anti-inflammatory, anti-tumor, as well as antibacterial and uric acid-lowering effects (58–60). In a 2017 Japanese study, researchers found that intake of chrysanthemum extract led to a significant reduction in serum UA after week four in young Japanese men with high serum uric acid (p < 0.01) (61). Peng et al. (62) compared the inhibition rates of 11 chrysanthemum extracts on XOD and found that Chrysanthemum morifolium Ramat “Boju” extract (CBE) showed excellent inhibition. Their analysis revealed that the flavonoid glycosides (luteolin, apigenin, diosgenin, and acacetin, accounting for up to 79.8% of the total) played the main inhibitory role. In addition, administration of CBE to HUA rats revealed a significant decrease in serum UA levels and XOD activity (p < 0.05), upregulation of renal ABCG2 expression as well as decreased expression of URAT1 and GLUT9, and reduced serum CR and inflammatory factor levels, demonstrating a protective effect on the kidney. As mentioned earlier, the mechanism of UA lowering may involve adipocyte lipolysis regulation (32). Similarly, in this article by Peng (62), the analysis of serum metabolites using UPLC-ESI-QTQF/MS revealed that CBE prevents the pathological process of HUA by regulating 16 biomarkers related to the metabolism of tryptophan, sphingolipids, glycerophospholipids, and arachidonic acid.
Lou et al. (63, 64) found that macroporous resin extract of Dendrobium officinale Leaves (DLE) could inhibit HUA by inhibiting XOD and ADA activity in the liver and kidney, reducing uric acid production, regulating URAT1, ABCG2, and GLUT9 expression, and inhibiting NF-κB inflammatory pathway. Alpinia officinarum extract and Pueraria lobata extract also showed positive inhibition of XOD activity, attenuation of oxidative stress, downregulation of URAT1 and GLUT9 protein expression, and reduction of serum urea and creatinine levels (65, 66). Liang et al. (67) used HPLC-DAD-MS/MS to analyze nine compounds from Poria cocos, namely 5-O-caffeoylmangiferine, neo-caffeine, paclitaxel, neo-isocaffeine, isocaffeine, and trans-resveratrol, which were able to inhibit XOD activity. These substances significantly reduced serum tumor necrosis factor (TNF)-α, interleukin-1β (IL-1β), interleukin-6 (IL-6), interleukin-12(IL-12), UA, and BUN (p < 0.01), and protected chondrocytes from erosive damage. Chen et al. (68) found a flavonoid extract in saffron (Crocus sativus). After treatment of HUA rats with this extract, on the one hand, serum UA, CR, and BUN levels were significantly reduced in HUA rats (p < 0.05), serum and liver XOD activities were inhibited, and serum TG and LDL-C levels were significantly reduced (p < 0.05). On the other hand, it ameliorated intestinal microbial dysbiosis and disorders of lipid and amino acid metabolites in HUA rats. Agrocybe aegerita has a unique taste and function and is used worldwide as food or medicine. Yong et al. (69) extracted Agrocybe aegerita with ethanol and water, respectively, and found that they both had an inhibitory effect on hepatic XOD activity and an elevated effect on renal OAT1 expression. 2-Formyl-3,5-dihydroxybenzyl acetate, 2,4-dihydroxy-6-methylbenzaldehyde, 2-(6-hydroxy-1H-indol-3-yl) acetamide and 6-hydroxy-1H-indole-3-carboxaldehyde (HHC) were identified as potential active compounds by molecular docking. It is hypothesized that the potential mechanism is that the active compounds compete with the substrate xanthine for the XOD active site.
From the above summary of 14 common medicinal and food plant resources used in traditional medicine for the treatment of HUA in some Asian countries, it is easy to find that some plant extracts are, they can be used to treat HUA and prevent kidney inflammation by inhibiting XOD activity and lowering serum UA, CR, and BUN levels. At a deeper level, by down-regulating the expression of proteins that promote uric acid reabsorption, such as GLUT9 and URAT1, and up-regulating the expression of proteins that promote uric acid excretion, such as ABCG2, OAT1, OAT2, and OAT3, the anti-HUA is mechanistically achieved by regulating signaling pathways such as lipid metabolism and inflammatory response. And most of these extracts are flavonoids and phenolic substances, such as luteolin, catechins, and isorhamnetin, which play a major role in lowering uric acid and inhibiting XOD activity. Therefore, we next further summarized these bioactive substances with uric acid-lowering effects and their uric acid-lowering mechanisms.
In 1806, morphine with analgesic effects was successfully isolated from plants and is also considered the first active ingredient isolated from plants in the world (70). Over the next few hundred years, scientists have continuously isolated active ingredients from plants and studied their pharmacological effects and mechanisms, leading to the continuous development of modern drug innovation. It has been confirmed that plant-active ingredients have a wide range of biological activities. Table 1 shows some biologically active ingredients and their effects. In our previous summary of the various types of medicinal and edible plants, we briefly mentioned that some of these active substances can exert anti-HUA effects by inhibiting XOD activity and lowering serum uric acid levels (87). Therefore, in the next section, we will focus on summarizing the anti-hyperuricemia mechanism of various bioactive ingredients.
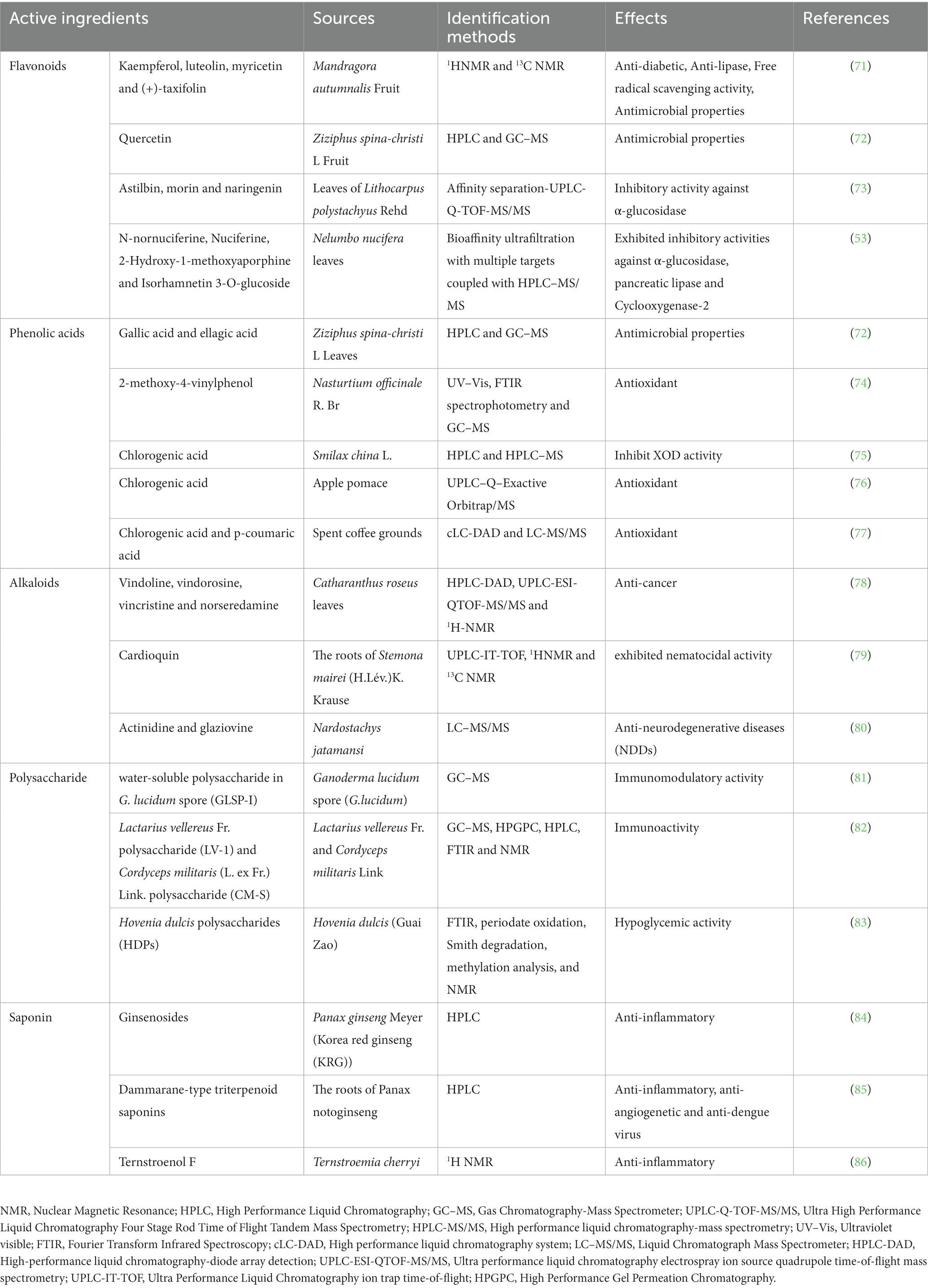
Table 1. Effects of active ingredients from different sources and its identification methods.
Flavonoids generally refer to a series of compounds formed by the interconnection of two benzene rings with phenolic hydroxyl groups through central three carbon atoms. It is among the most studied natural compounds and has received a lot of attention for its uric acid-lowering effects (88, 89). In a previous study, researchers selected 15 flavonoids to study their uric acid-lowering effects in HUA mice. It was found that continuous oral administration of 50 and 100 mg/kg of quercetin, mulberry pigment, kaempferol, apigenin, and gerberinol for 3 days significantly reduced hepatic uric acid levels and significantly inhibited hepatic xanthine oxidase activity in mice (90) (p < 0.01).
Yuan et al. (91) extracted and identified flavonoid components from corn beard and found that the flavonoid components eluted from 60% ethanol solution had the best uric acid-lowering effect. It could reduce serum uric acid level by 26.69% and XOD activity by 11.29%. It was also hypothesized that apigenin-6-C-glucoside-7-O-glucoside, kaempferol-3-globoside, lignan-7-glucoside, lignan-3′,7-di-O-glucoside and naringenin among them would affect the XOD activity.
Quercetin is one of the most abundant flavonoids in the daily diet and has been studied as an anti-cardiovascular, anti-cancer, anti-viral, and antioxidant drug (92–96). In recent years, studies have found that quercetin has the potential to treat HUA, and as a result, more and more research has been conducted on quercetin for the treatment of HUA (97). Zhang et al. (98) investigated the dual-substrate kinetics and inhibition mechanism of quercetin on XOD by combining kinetic analysis, molecular docking techniques, and multispectral approaches. The IC50 of quercetin against XOD was found to be 2.74 ± 0.04 mol/L. It also reversibly inhibited the production of uric acid and O2- in a mixed manner, and the binding process with XOD was mainly dominated by van der Waals forces and hydrogen bonds. In a study at the University of Leeds, Sarka et al. (99) found that quercetin and its metabolites were able to inhibit the activity of xanthine oxidoreductase (XOR, one of the key rate-limiting enzymes of purine metabolism) exerting a lowering effect on uric acid in vivo. Also in a study by Yu et al. (45), quercetin was found to show relatively strong URAT1 inhibitory activity with an IC50 of 12.6 μM. Zhu et al. (100) found that quercetin derivatives (quercetin-3-O-β-D-galactopyranoside, quercetin-3-O-γ-D-glucopyranoside, and quercetin-3-O-α-L-rhamnoside) inhibited XOD activity and reduced the expression of intracellular reactive oxygen species (ROS), IL-1β, IL-1β, and NOD-like receptor superfamily containing 3 (NLRP3) inflammatory vesicles, showing significant therapeutic effects on HUA and renal inflammation.
Isorhamnetin is a dimethylated flavonol that is found in sea buckthorn and ginkgo leaves (101). In a previous study by Wang (49), isorhamnetin has been shown to significantly reduce serum uric acid levels and inhibit hepatic XOD activity in mice. Also in a study by Adachi et al. (102), they found that oral administration of isorhamnetin reduced uric acid levels in blood and liver in a dose-dependent manner and decreased XOD activity. Luteolin is a natural flavonoid found in a variety of plants and has a wide range of pharmacological effects, such as anti-inflammatory, anti-tumor, anti-viral, and uric acid-lowering (103–105). Back in 2013, the team of Yan (106) found that luteolin reversibly competitively inhibited XOD with an inhibition constant (Ki) value of (2.38 ± 0.05) × 10–6 mol/L. They found that luteolin interacts with amino acid residues of the first order within the active site pocket of XOD by fluorescence and circular dichroism. In a 2017 clinical trial in Japan, researchers found that a continuous intake of 10 mg of luteolin for four weeks was able to reduce serum uric acid levels (61). Lin et al. (107) also investigated the therapeutic effects of luteolin and luteolin-4’-O-glucoside in HUA mice. The results revealed that both showed positive anti-HUA effects by decreasing URAT1 levels and inhibiting XOD activity. The levels of IL-1β and TNF-α were also reduced to improve inflammation. Rajesh et al. (108) extracted luteolin from millet and found a binding energy of −9.7 kcal/mol with XOR by molecular docking, while the binding energy of allopurinol with XOR was −8.0 kcal/mol, indicating the potential of luteolin as an XOR inhibitor. It has been found that luteolin inhibited XOD activity and mRNA expression of XDH while inducing increased expression levels of OAT1 and OAT3 proteins as well as decreased GLUT9 and URAT1 expression. In addition, in a study by Zhu et al. (109), it was found that luteolin reversed the decrease in OAT1 levels and inhibited XOD activity as well as TLR4/MyD88/NLRP3 signaling pathway in the liver.
Overall, flavonoids reduce uric acid production by inhibiting XOD and XOR enzyme activities and regulating the expression of GLUT9, UART1, OAT1, and other related proteins. It also promotes uric acid excretion to reduce HUA and reduces the expression of inflammatory vesicles such as IL-1β, IL-1β and NLRP3 inflammatory vesicles and TLR4/MyD88/NLRP3 signaling pathway to protect the kidney or joints from inflammatory damage. The established hypouricemic mechanisms of flavonoids in bioactive substances are summarized in Table 2.
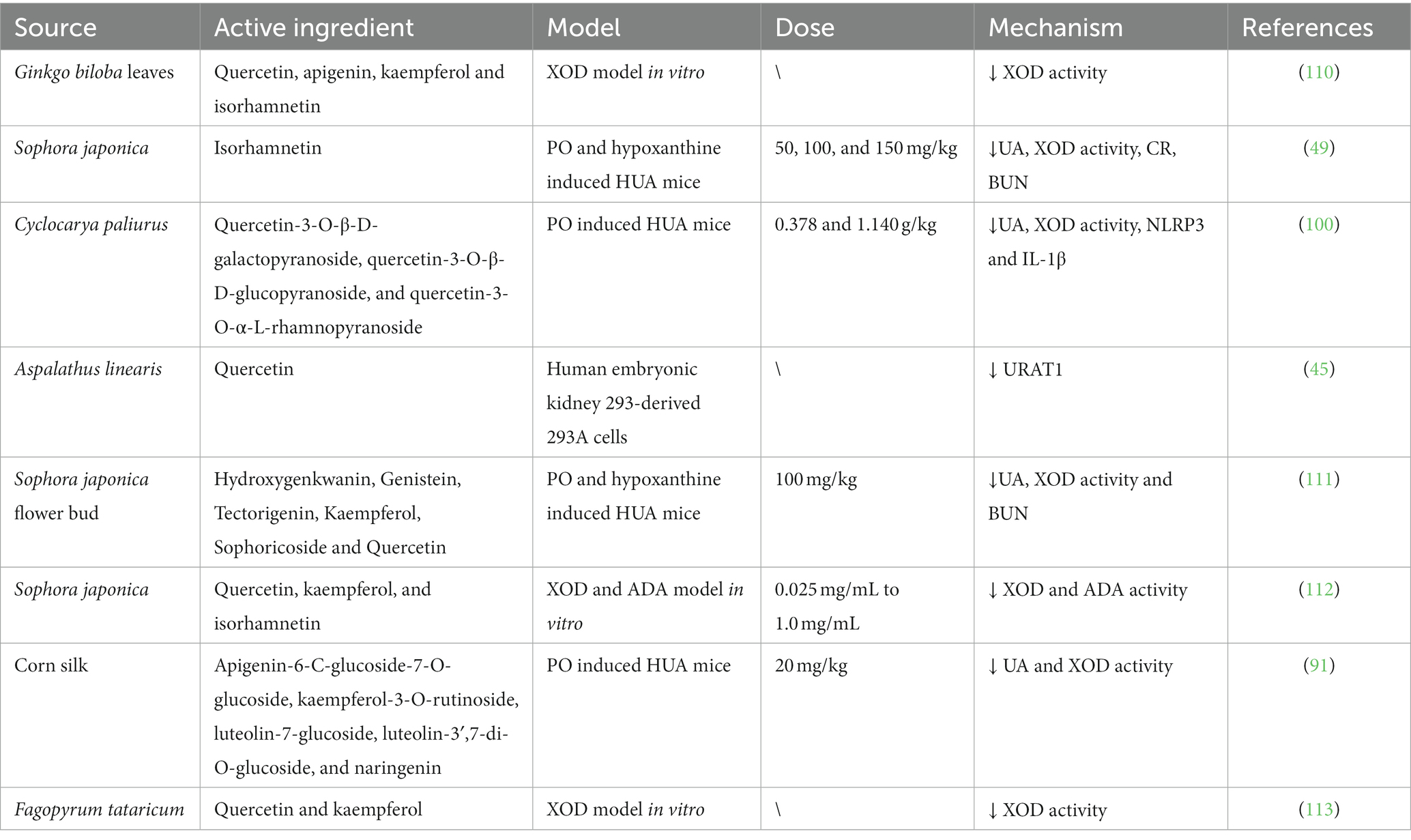
Table 2. Anti-hyperuricemic effect of flavonoids from different sources.
Phenolic acids are a class of organic acids containing phenolic rings and are one of the main types of polyphenols (114). Phenolic acid compounds refer to a class of compounds with several hydroxyl groups on the same benzene ring, and there are two main types of corresponding carbon frameworks. One type is C6-C1 type, which is benzoic acid type, such as p-hydroxybenzoic acid, gallic acid, protocatechu, and the other type is C6-C3 type, which is cinnamic acid type, such as caffeic acid, ferulic acid, p-coumaric acid, and mustard acid (115). It has been shown that Phenolic acids have numerous health benefits such as antioxidant properties, anti-inflammatory, antibacterial, and neuroprotective effects (115–117). In recent years, phenolic acid compounds have been found to have XOD inhibitory activity and have been suggested to have potential effects against HUA. Lin et al. (118) identified 10 phenolic acid compounds from Coix lacryma bran-free polyphenol extract by UPLC-QTOF-MS/MS. For the first time, mustard acid was found to inhibit XOD in a mixed non-competitive manner. In a recent study, researchers compared free polyphenols in Lactobacillus acidophilus NCUF202.2 fermented artemisia selengensis turcz extracts (FASTE) and artemisia selengensis turcz (AST) and the therapeutic effects on HUA rats. The results revealed that coffee quinic acid in FASTE significantly increased the content of free polyphenols (p < 0.05), enhanced the inhibitory effect on XOD, and better reduced serum ADA activity. And it increases the expression of uric acid secretory protein OAT1 and decreases the expression of reabsorption proteins URAT1 and GLUT9 in model rats, thus reducing serum uric acid levels in model rats (119).
Chlorogenic acid is an ester produced by caffeic acid and quinic acid (1-hydroxyhexahydro gallic acid), a phenylpropanoid compound produced by the plant body during aerobic respiration via the mangiferous acid pathway (120, 121). Zilma et al. (122) found that the aqueous extract of Tabebuia roseoalba leaves had an inhibitory effect on hepatic XOD and reduced serum uric acid levels. Further identification by HPLC analysis revealed that caffeic and chlorogenic acids may be responsible for the activity exhibited by the extract. In a study by Zhou et al. (123), chlorogenic acid was found to decrease UA, BUN, CR levels and inhibited XOD activity in hypoxanthine and oxokalate induced HUA mice. Chlorogenic acid also down-regulated the mRNA expression of UA secreted proteins. In addition, chlorogenic acid significantly reduced the mRNA expression of IL-1β, TNF-α, NLRP3 and Caspase-1 (p < 0.05), inhibited TLR4/MyD88/NF-κB signaling pathway in the kidney and attenuated renal inflammation in HUA mice.
Caffeic acid is also one of the products of the phytocannabinoid pathway. Extensive studies have confirmed the biological activities of caffeic acid such as anti-microbial, anti-cancer, antioxidant and anti-inflammatory (124–127). In a study by Wan et al. (128), PO-induced gavage of caffeic acid (100 mg/kg) for 7 days in HUA rats significantly reduced serum uric acid and XOD activity (p < 0.01). Caffeic acid achieves its uric acid-lowering effect mainly by down-regulating the transcript levels of URAT1 and GLUT9 mRNAs and up-regulating the transcription of OAT1, UAT and ABCG2 mRNAs. In vitro studies showed that caffeic acid was competitively inhibited with XOD, with an IC50 value of 53.45 μM. Foreign researchers studied ethanolic extract of Lobelia and found that its ethanolic extract and its components (chlorogenic and caffeic acids, etc.) had inhibitory activity on hepatic XOD and were able to lessen the size of paw edema caused by urate crystals (129).
Gallic acid is a polyphenolic compound widely found in fruits, plants and nuts, with a variety of biological activities including anti-inflammatory, anti-mutagenic, antioxidant and anti-cancer (130–132). Wu et al. (37) established a HUA model using normal human hepatocytes (HL-7702). They found that gallic acid significantly inhibited uric acid production at the cellular level (p < 0.05), and the inhibition was stronger than that of tea polyphenols and theaflavins. Chinese researchers identified the active compound gallic acid (GA) by HPLC from ethyl acetate extracts of Sonneratia apetala leaves and branches and investigated its mechanism of regulating uric acid metabolism in HUA mice (38). It was found that GA inhibited ADA and XOD activities and was able to down-regulate the expression of URAT1 and GLUT9 proteins, up-regulate the expression of ABCG2 protein as well as inhibit MDA, IL-6, IL-1β, TNF-α and enhance the activity of SOD and GSH-Px in the kidney. These suggest that GA reduces the levels of uric acid and prevents renal injury by inhibiting oxidative stress and inflammation and regulating the expression of uric acid transporters in HUA mice.
Phenolic acids had inhibitory activity against ADA and XOD, increased the expression of uric acid secretory proteins (OAT1, UAT, and ABCG2) and inhibited uric acid reabsorption proteins (URAT1 and GLUT9). It also inhibited the activity of oxidative stress and related inflammatory bodies (IL-1β, TNF-α, NLRP3, and Caspase-1), and exerted the effect of lowering uric acid level to prevent HUA and protect the kidney from inflammatory damage. The established hypouricemic mechanisms of phenolic acids in bioactive substances are summarized in Table 3.
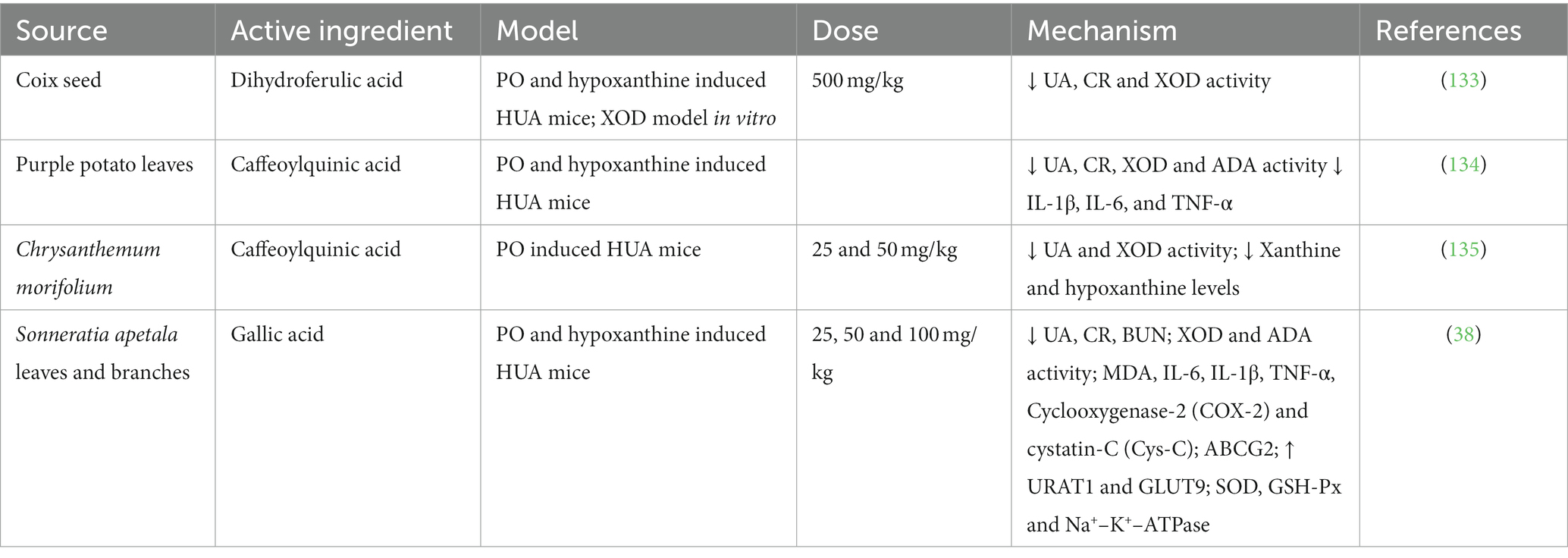
Table 3. Anti-hyperuricemic effect of phenolic acids from different sources.
Alkaloids are a category of nitrogen-containing alkaline organic compounds found in nature with significant biological activities, such as anti-cancer and anti-bacterial (136–138). It was found that the total lotus leaves alkaloids showed positive inhibition of XOD enzyme activity with an IC50 value of 3.313 μg/mL, and UHPLC-Q-TOF-MS and 3D docking analysis revealed lotus alkaloids as potential active ingredients (24). Not only that, in PO-induced HUA mice, lotusine reduced serum uric acid levels by a possible mechanism of reversing the expression of related uric acid transporters (URAT1, GLUT9, ABCG2) as well as inhibiting TLR4/MyD88/NF-κB signaling pathway and NLRP3 inflammatory vesicles activation in the kidney, exerting anti-HUA and anti-inflammatory effects (139).
Berberine (BBR), also known as flavopiridol, is a quaternary ammonium alkaloid isolated from the Chinese medicine Huanglian, which has a wide range of antibacterial activity (140). It was found that berberine significantly reduced uric acid levels in the blood and feces of HUA rats (p < 0.05), while blood XOD levels decreased, ABCG2 levels increased and Galectin-9 levels decreased (141). Li et al. (142) found 82 targets of berberine acting on HUA as well as several inflammation-related signaling pathways by network pharmacology prediction. In vivo experiments revealed that berberine significantly reduced UA, BUN and CRE levels in HUA mice (p < 0.05), decreased IL-1β and IL-18 levels, and down-regulated the expression of URAT1, NLRP3, apoptosis-associated spot-like protein (ASC), Caspase1 and IL-1β, showing anti-HUA and prevention of renal effects on kidney injury. Berberine also reduced hepatic XOD activity at the protein and mRNA levels, down-regulated GLUT9 expression, up-regulated OAT1/3 and ABCG2 expression, and inhibited the JAK2/STAT3 signaling pathway to exert anti-HUA effects (143, 144). Zhong et al. (145) used PO and hypoxanthine to induce HUA mouse models for 7 consecutive days, followed by gavage of oxyberberrubine (oxyberberrubine is a metabolite of berberine in the liver, 5, 10 and 20 mg/kg), while febuxostat (Fex, 5 mg/kg) was used as a positive control. After OBR treated, the levels of serum uric acid was reduced and inhibition of XOD and ADA activities were enhanced, along with decreased expression of XOD, URAT1, GLUT9, NLRP3, ASC and Caspase-1 at the mRNA and protein levels; it also significantly reduced serum creatinine and blood urea nitrogen and inflammatory mediators (TNF-α, IL-1β, IL-6 and IL-18) levels (p < 0.05).
Alkaloids reduce XOD and ADA activity by inhibiting XOD expression in the liver. It also downregulates the expression of GLUT9 and URAT1, upregulates the expression of ABCG2 and OAT1/3, reduces serum uric acid levels, prevents HUA as well as inhibits the expression of inflammatory vesicles and inflammatory signaling pathways, and protects the kidney from inflammatory damage. The established hypouricemic mechanisms of alkaloids in bioactive substances are summarized in Table 4.
Table 4. Anti-hyperuricemic effect of alkaloids from different sources.
Polysaccharide is formed by condensation and water loss of several monosaccharide molecules and is a class of sugar substances with complex and large molecular structure, which has various biological activities such as anti-cancer, anti-bacterial, anti-inflammatory and anti-HUA (148–151). It can be divided into homopolysaccharides and heteropolysaccharides. The former includes starch, glycogen, and cellulose, while the latter includes hyaluronic acid, etc. Lonicera japonica polysaccharide (LJP-1) has anti-uricemia and anti-gout effects on PO and hypoxanthine induced HUA rats. The mechanism alleviated the degree of ankle joint swelling in a sodium urate crystal-induced gouty arthritis model by inhibiting XOD activity and reducing serum uric acid levels, IL-1β, IL-6, TNF-α, and COX-2-related inflammatory factor levels (152). Yu et al. (153) established a HUA model by intraperitoneal injection of PO (250 mg/kg) into eight-week-old C57BL/6 male mice for seven consecutive days and treated with a dose of Lycium barbarum polysaccharide (LBP) for seven consecutive days. The results revealed that 100 and 200 mg/kg of LBP significantly reduced serum uric acid levels and elevated urinary uric acid levels (p < 0.05). It also significantly reduced XOD activity in serum and liver (p < 0.01). It also increased the mRNA and protein expression levels of OAT1/3 as well as decreased GLUT9 levels. Ganoderma lucidum polysaccharide (GP) significantly reduced UA, BUN and alaninetransaminase (ALT) levels by inhibiting hepatic and blood ADA activity (p < 0.05) and increased UA excretion by inhibiting GLUT9 and OAT1 expression (154, 155).
In general, polysaccharides are macromolecules with low toxicity (156, 157), which can be widely used in the development of various functional foods. Polysaccharides also have a therapeutic effect on HUA by a mechanism related to the inhibition of XOD and ADA activity to reduce UA production. It is also associated with the enhancement of OAT1/3 and inhibition of GLUT9 protein expression to promote UA excretion as well as protection of the kidney and joints from inflammatory damage by reducing the levels of IL-1β, IL-6, TNF-α and COX-2-related inflammatory factors.
Saponins are composed of saponins and sugars, which can be divided into steroidal saponins and Triterpenoid saponin. It is mainly found in terrestrial higher plants and small amounts in marine organisms such as starfish and sea cucumbers (158, 159). Studies have shown that saponins have biological activities such as antioxidant and anti-inflammatory (160, 161).
The total saponin content of natural plant “Yaocha” extract is 1.0 to 1.5%, and the saponin content of Camellia sinensis is 0.4 to 0.8%. Its inhibition of XOD activity by upregulating the mRNA expression of hypoxanthine guanine phosphoribosyltransferase transferase (HPRT1) and OAT1 makes “Yaocha” a potential therapeutic agent for the treatment of HUA (162). The total saponin of Huangshan yam was extracted using 70% ethanol at 80°C, and its effects on serum uric acid level (SUA), urinary uric acid level (UUA), uric acid clearance (CUR) and creatinine clearance (CCR) as well as the expression of related uric acid transporters were investigated in HUA rats within 24h (163). The results revealed that saponin administration was dose-dependent, with increased UUA, CUR and CCR values and decreased SUA within 24 h; and decreased mRNA and protein expression of URAT1 and GLUT9 and increased OAT1/3 expression in the kidney.
In general, saponins have similar mechanisms of HUA inhibition as most plant active ingredients. Saponins reduce UA levels and increase UA excretion by inhibiting the activity of XOD, a key enzyme for uric acid synthesis. It also reduced the expression of URAT1 and GLUT9 in the kidney, promoted the expression of ABCG2 to reduce serum uric acid levels, and downregulated NLRP3 inflammatory vesicles to have a therapeutic effect on gouty arthritis (164, 165). The established hypouricemic mechanisms of saponins in bioactive substances are summarized in Table 5.

Table 5. Anti-hyperuricemic effect of saponins from different sources.
HUA has become a global public health hazard after chronic diseases such as “three highs” and diabetes, and its incidence is increasing rapidly worldwide. Traditionally, drugs for the treatment of HUA have been shown to cause a series of side effects on the human body, thus limiting the clinical treatment of HUA. Therefore, medicinal plants have been extensively studied by researchers working in medicine worldwide for their significant uric acid-lowering effects and are considered as a future therapeutic alternative to HUA. In this regard, researchers are intrigued by the large number of bioactive components contained in medicinal plants. On this basis, numerous experimental studies were conducted in various countries to understand the uric acid-lowering mechanisms of these active ingredients, leading to the discovery that active ingredients such as flavonoids, phenolic acids, and alkaloids inhibit uric acid production or promote uric acid excretion. Up to now, although a huge number of in vivo and in vitro experiments have shown that medicinal plants have significant therapeutic effects on HUA, the current research is still limited. We know that not only XOD plays a role in the production of uric acid, but also PRPS, HGPRT, and PRPPAT play indispensable roles in uric acid metabolism. However, there is currently limited research on the mechanism of action of plant extracts on uric acid metabolism enzymes, with most studies focusing on the related enzymes (ADA and/or XDH) and the expression of uric acid transporters during the process of uric acid production. Meanwhile, many of these medicinal plants have been extracted to obtain their crude extracts or to isolate certain active ingredients, but these studies have been limited to the laboratory stage only and have never been studied clinically, nor have toxicological studies been performed. In addition, the metabolic pathway of uric acid in humans is significantly different from that in rodents. The possible differences are undeniable as the enzyme uricase, which converts uric acid into allantoic acid, does not exist in humans, whereas rodents can convert uric acid into allantoic acid and excrete it through uricase.
Although there are still some pressing problems in the treatment of HUA with medicinal plants, it does not prevent it from being a potential drug for the prevention and treatment of HUA in the future. In previous studies, we found that molecular docking techniques could be used to elucidate the relationship between bioactive components and XOD and to determine the mechanism of inhibition in this way. In subsequent studies, we can use network pharmacology and molecular docking techniques to explore the binding between bioactive ingredients and HUA targets and uric acid transporters and uric acid metabolizing enzyme and use them as evidence of the effectiveness of bioactive ingredients against HUA. In conclusion, medicinal plants are available as an effective strategy for the prevention and treatment of HUA, and the active ingredients in them provide new directions for the synthesis of effective and less toxic drugs for the treatment of HUA, indicating the potential of medicinal plants for the prevention and treatment of HUA in the future.
DJ-g is responsible for writing and revising the thesis. WC-y is responsible for the revision and guidance of the thesis. All authors contributed to the article and approved the submitted version.
This study was supported by Chongqing Science and Technology Project, cstc2016zdcy-ztzx10001.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Wang, H., Zhang, H., Sun, L., and Guo, W. Roles of hyperuricemia in metabolic syndrome and cardiac-kidney-vascular system diseases. Am J Transl Res. (2018) 10:2479–63.
2. Yanai, H, Adachi, H, Hakoshima, M, and Katsuyama, H. Molecular biological and clinical understanding of the pathophysiology and treatments of hyperuricemia and its association with metabolic syndrome, cardiovascular diseases and chronic kidney disease. Int J Mol Sci. (2021) 22:221. doi: 10.3390/ijms22179221
3. Jiang, LL, Gong, X, Ji, MY, Wang, CC, Wang, JH, and Li, MH. Bioactive compounds from plant-based functional foods: a promising choice for the prevention and Management of Hyperuricemia. Foods. (2020) 9:973. doi: 10.3390/foods9080973
4. Xia, Y, Wu, Q, Wang, H, Zhang, S, Jiang, Y, Gong, T, et al. Global, regional and national burden of gout, 1990–2017: a systematic analysis of the global burden of disease study. Rheumatolog. (2019) 59:1529–38. doi: 10.1093/rheumatology/kez476
5. Chen-Xu, M, Yokose, C, Rai, SK, Pillinger, MH, and Choi, HK. Contemporary prevalence of gout and hyperuricemia in the United States and decadal trends: the National Health and nutrition examination survey, 2007–2016. Arthritis Rheumatol. (2019) 71:991–9. doi: 10.1002/art.40807
6. De Becker, B, Borghi, C, Burnier, M, and van de Borne, P. Uric acid and hypertension: a focused review and practical recommendations. J Hypertens. (2019) 37:878–83. doi: 10.1097/HJH.0000000000001980
7. Liu, X, Lv, Q, Ren, H, Gao, L, Zhao, P, Yang, X, et al. The altered gut microbiota of high-purine-induced hyperuricemia rats and its correlation with hyperuricemia. PeerJ. (2020) 8:e8664. doi: 10.7717/peerj.8664
8. Su, HY, Yang, C, Liang, D, and Liu, HF. Research advances in the mechanisms of hyperuricemia-induced renal injury. Biomed Res Int. (2020) 2020:5817348. doi: 10.1155/2020/5817348
9. Morimoto, C, Tamura, Y, Asakawa, S, Kuribayashi-Okuma, E, Nemoto, Y, Li, J, et al. ABCG2 expression and uric acid metabolism of the intestine in hyperuricemia model rat. Nucleosides Nucleotides Nucleic Acids. (2020) 39:744–59. doi: 10.1080/15257770.2019.1694684
10. Toyoda, Y, Takada, T, Miyata, H, Matsuo, H, Kassai, H, Nakao, K, et al. Identification of GLUT12/SLC2A12 as a urate transporter that regulates the blood urate level in hyperuricemia model mice. Proc Natl Acad Sci U S A. (2020) 117:18175–7. doi: 10.1073/pnas.2006958117
11. Xu, L, Shi, Y, Zhuang, S, and Liu, N. Recent advances on uric acid transporters. Oncotarget. (2017) 8:100852–62. doi: 10.18632/oncotarget.20135
12. Wang, Y, Zhang, W, Qian, T, Sun, H, Xu, Q, Hou, X, et al. Reduced renal function may explain the higher prevalence of hyperuricemia in older people. Sci Rep. (2021) 11:1302. doi: 10.1038/s41598-020-80250-z
13. Maiuolo, J, Oppedisano, F, Gratteri, S, Muscoli, C, and Mollace, V. Regulation of uric acid metabolism and excretion. Int J Cardiol. (2016) 213:8–14. doi: 10.1016/j.ijcard.2015.08.109
14. Cw, W, Rl, D, and Wh, C. Immunopathogenesis and risk factors for allopurinol severe cutaneous adverse reactions. Curr Opin Allergy Clin Immunol. (2016) 16:339–45. doi: 10.1097/ACI.0000000000000286
15. Harrold, L. New developments in gout. Curr Opin Rheumatol. (2013) 25:304–9. doi: 10.1097/BOR.0b013e32835fd5e5
16. Hou, Y, and Jiang, JG. Origin and concept of medicine food homology and its application in modern functional foods. Food Funct. (2013) 4:1727–41. doi: 10.1039/c3fo60295h
17. Gong, X, Ji, M, Xu, J, Zhang, C, and Li, M. Hypoglycemic effects of bioactive ingredients from medicine food homology and medicinal health food species used in China. Crit Rev Food Sci Nutr. (2020) 60:2303–26. doi: 10.1080/10408398.2019.1634517
18. Xia, X, and Xiao, J. Natural ingredients from medicine food homology as chemopreventive reagents against type 2 diabetes mellitus by modulating gut microbiota homoeostasis. Molecules. (2021) 26:6934 and 1420-3049. doi: 10.3390/molecules26226934
19. Song, D-X, and Jiang, J-G. Hypolipidemic components from medicine food homology species used in China: pharmacological and health effects. Arch Med Res. (2017) 48:569–81. doi: 10.1016/j.arcmed.2018.01.004
20. Yang, Z, Wang, H, Qi, G, Chen, G, Cao, C, and Wang, S. Antimicrobial effects of a compound solution of three medicine food homology plants. Food Biosci. (2022) 49:101845. doi: 10.1016/j.fbio.2022.101845
21. Guo, P, Zhang, B, Zhao, J, Wang, C, Wang, Z, Liu, A, et al. Medicine-food herbs against Alzheimer’s disease: a review of their traditional functional features, substance basis, clinical practices and mechanisms of action. Molecules. (2022) 27:901. doi: 10.3390/molecules27030901
22. Shen, D, Feng, Y, Zhang, X, Gong, L, Liu, J, Li, Y, et al. Antiosteoporosis studies of 20 medicine food homology plants containing quercetin, Rutin, and Kaempferol: TCM characteristics, in vivo and in vitro activities, potential mechanisms, and food functions. Evid Based Complement Alternat Med. (2022) 2022:5902293. doi: 10.1155/2022/5902293
23. Wang, Y, Lin, Z, Zhang, B, Nie, A, and Bian, M. Cichorium intybus L. promotes intestinal uric acid excretion by modulating ABCG2 in experimental hyperuricemia. Nutr Metab (Lond). (2017) 14:38. doi: 10.1186/s12986-017-0190-6
24. Sang, M, du, G, Hao, J, Wang, L, Liu, E, Zhang, Y, et al. Modeling and optimizing inhibitory activities of Nelumbinis folium extract on xanthine oxidase using response surface methodology. J Pharm Biomed Anal. (2017) 139:37–43. doi: 10.1016/j.jpba.2017.02.048
25. Zhou, Y, Shang, F, Zhao, W, Wang, Y, Geng, X, Meng, Y, et al. Resourcing potential of diverse functional components from Chaenomeles Sinensis immature fruits. Ekoloji. (2019) 28:153–7.
26. Zhang, R, Zhan, S, Li, S, Zhu, Z, He, J, Lorenzo, JM, et al. Anti-hyperuricemic and nephroprotective effects of extracts from Chaenomeles sinensis (Thouin) Koehne in hyperuricemic mice. Food Funct. (2018) 9:5778–90. doi: 10.1039/c8fo01480a
27. Klimek-Szczykutowicz, M, Szopa, A, and Ekiert, H. Citrus limon (lemon) phenomenon—a review of the chemistry, pharmacological properties, applications in the modern pharmaceutical, food, and cosmetics industries, and biotechnological studies. Plan Theory. (2020) 9:119. doi: 10.3390/plants9010119
28. Wang, H, Cheng, L, Lin, D, Ma, Z, and Deng, X. Lemon fruits lower the blood uric acid levels in humans and mice. Sci Hortic. (2017) 220:4–10. doi: 10.1016/j.scienta.2017.03.023
29. Chen, L, Li, M, Wu, JL, Li, JX, and Ma, ZC. Effect of lemon water soluble extract on hyperuricemia in a mouse model. Food Funct. (2019) 10:6000–8. doi: 10.1039/c9fo00509a
30. Perović, J, Tumbas Šaponjac, V, Kojić, J, Krulj, J, Moreno, DA, García-Viguera, C, et al. Chicory (Cichorium intybus L.) as a food ingredient–nutritional composition, bioactivity, safety, and health claims: a review. Food Chem. (2021) 336:127676–8146. doi: 10.1016/j.foodchem.2020.127676
31. Jin, Y-N, Lin, Z-J, Zhang, B, and Bai, Y-F. Effects of chicory on serum uric acid, renal function, and GLUT9 expression in Hyperuricaemic rats with renal injury and in vitro verification with cells. Evid Based Complement Alternat Med. (2018) 1-11:2018. doi: 10.1155/2018/1764212
32. Bian, M, Wang, J, Wang, Y, Nie, A, Zhu, C, Sun, Z, et al. Chicory ameliorates hyperuricemia via modulating gut microbiota and alleviating LPS/TLR4 axis in quail. Biomed Pharmacother. (2020) 131:110719. doi: 10.1016/j.biopha.2020.110719
33. Yan, Z, Zhong, Y, Duan, Y, Chen, Q, and Li, F. Antioxidant mechanism of tea polyphenols and its impact on health benefits. Animal. Nutrition. (2020) 6:115–23. doi: 10.1016/j.aninu.2020.01.001
34. Zhao, C-N, Tang, GY, Cao, SY, Xu, XY, Gan, RY, Liu, Q, et al. Phenolic profiles and antioxidant activities of 30 tea infusions from green, black, oolong, white, yellow and dark teas. Antioxidants. (2019) 8:215 and 2076-3921. doi: 10.3390/antiox8070215
35. Wang, X, Liu, Y, Wu, Z, Zhang, P, and Zhang, X. Tea polyphenols: a natural antioxidant regulates gut flora to protect the intestinal mucosa and prevent chronic diseases. Antioxidants. (2022) 11:2076–3921. doi: 10.3390/antiox11020253
36. Widha Nugraheni, P, Rahmawati, F, Mahdi, C, and Prasetyawan, S. Green tea extract (Camellia Sinensis L.) effects on uric acid levels on hyperuricemia rats (Rattus norvegicus). J Pure Appl Chem Res. (2017) 6:246–54. doi: 10.21776/ub.jpacr.2017.006.03.355
37. Wu, D, Chen, R, Zhang, W, Lai, X, Sun, L, Li, Q, et al. Tea and its components reduce the production of uric acid by inhibiting xanthine oxidase. Food Nutr Res. (2022) 66:8239. doi: 10.29219/fnr.v66.8239
38. Jiang, L, Wu, Y, Qu, C, Lin, Y, Yi, X, Gao, C, et al. Hypouricemic effect of gallic acid, a bioactive compound from Sonneratia apetala leaves and branches, on hyperuricemic mice. Food Funct. (2022) 13:10275–90. doi: 10.1039/D2FO02068H
39. Li, F, Liu, Y, Xie, Y, Liu, Z, and Zou, G. Epigallocatechin gallate reduces uric acid levels by regulating xanthine oxidase activity and uric acid excretion in vitro and in vivo. Ann Palliat Med. (2020) 9:331–8. doi: 10.21037/apm.2019.11.28
40. Tai, L, Liu, Z, Sun, M, Xie, Q, Cai, X, Wang, Y, et al. Anti-hyperuricemic effects of three theaflavins isolated from black tea in hyperuricemic mice. J Funct Foods. (2020) 66:103803. doi: 10.1016/j.jff.2020.103803
41. Nugraheni, PW, and Mahdi, C. Kidney evaluation in hyperuricemia rats treated with green tea leaves (Camellia sinensis L.). Ext Biol Med Nat Product Chem. (2022) 11:17–26. doi: 10.14421/biomedich.2022.111.17-26
42. Sang, S, Wang, L, Liang, T, Su, M, and Li, H. Potential role of tea drinking in preventing hyperuricaemia in rats: biochemical and molecular evidence. Chin Med. (2022) 17:108. doi: 10.1186/s13020-022-00664-x
43. Wu, D, Chen, R, Li, Q, Lai, X, Sun, L, Zhang, Z, et al. Tea (Camellia sinensis) ameliorates hyperuricemia via uric acid metabolic pathways and gut microbiota. Nutrients. (2022) 14:666. doi: 10.3390/nu14132666
44. Yuan, D, Lin, L, Peng, Y, Zhou, Y, Li, L, Xiao, W, et al. Effects of black tea and black brick tea with fungal growth on lowering uric acid levels in hyperuricemic mice. J Food Biochem. (2022) 46:e14140. doi: 10.1111/jfbc.14140
45. Toyoda, Y, Takada, T, Saito, H, Hirata, H, Ota-Kontani, A, Tsuchiya, Y, et al. Identification of inhibitory activities of dietary flavonoids against URAT1, a renal urate re-absorber: in vitro screening and fractional approach focused on rooibos leaves. Nutrients. (2022) 14:2072–6643. doi: 10.3390/nu14030575
46. Wang, Z, Zhang, Z, Lu, C, Zhou, J, Wang, Z, Han, J, et al. Effects of Sporisorium reiliana polysaccharides and Phoenix dactylifera monosaccharides on the gut microbiota and serum metabolism in mice with fructose-induced hyperuricemia. Arch Microbiol. (2022) 204:436. doi: 10.1007/s00203-022-03053-y
47. Lee, M-H, Jeong, J-H, Jeong, M-S, Chang, S-H, and Her, E. Anti-inflammatory function of the Sophora japonica extract rutin: the inhibitory effect of rutin of korean Sophora japonica on the productions of NO and TNF-alpha from mouse peritoneal macrophages. Korean journal of medicinal. Crop Sci. (2010) 18:105–12.
48. He, X, Bai, Y, Zhao, Z, Wang, X, Fang, J, Huang, L, et al. Local and traditional uses, phytochemistry, and pharmacology of Sophora japonica L.: a review. J Ethnopharmacol. (2016) 187:160–82. doi: 10.1016/j.jep.2016.04.014
49. Wang, F, Zhao, X, Su, X, Song, D, Zou, F, and Fang, L. Isorhamnetin, the xanthine oxidase inhibitor from Sophora japonica, ameliorates uric acid levels and renal function in hyperuricemic mice. Food Funct. (2021) 12:12503–12. doi: 10.1039/d1fo02719k
50. Li, J, Gong, Y, Li, J, and Fan, L. In vitro xanthine oxidase inhibitory properties of Flos Sophorae Immaturus and potential mechanisms. Food Biosci. (2022) 47:101711. doi: 10.1016/j.fbio.2022.101711
51. Zheng, H, Han, L, Shi, W, Fang, X, Hong, Y, and Cao, Y. Research advances in Lotus leaf as Chinese dietary herbal medicine. Am J Chin Med. (2022) 50:1423–45. doi: 10.1142/S0192415X22500616
52. Wan, Y, Xia, J, Xu, JF, Chen, L, Yang, Y, Wu, JJ, et al. Nuciferine, an active ingredient derived from lotus leaf, lights up the way for the potential treatment of obesity and obesity-related diseases. Pharmacol Res. (2022) 175:106002. doi: 10.1016/j.phrs.2021.106002
53. Zhang, H, Chen, G, Zhang, Y, Yang, M, Chen, J, and Guo, M. Potential hypoglycemic, hypolipidemic, and anti-inflammatory bioactive components in Nelumbo nucifera leaves explored by bioaffinity ultrafiltration with multiple targets. Food Chem. (2022) 375:131856. doi: 10.1016/j.foodchem.2021.131856
54. An, Y, Hao, J, Li, J, He, W, Wang, L, and Zhang, Y. The inhibitory effect of lotus leaf extract on hyperuricemia and its potential mechanism. Acupunc Herbal Med. (2021) 1:122–9. doi: 10.1097/hm9.0000000000000001
55. Kong, LD, Cai, Y, Huang, WW, Cheng, CHK, and Tan, RX. Inhibition of xanthine oxidase by some Chinese medicinal plants used to treat gout. J Ethnopharmacol. (2000) 73:199–207. doi: 10.1016/S0378-8741(00)00305-6
56. Lee, YS, Son, E, Kim, SH, Lee, YM, Kim, OS, and Kim, DS. Synergistic uric acid-lowering effects of the combination of Chrysanthemum indicum Linne flower and Cinnamomum cassia (L.) J. Persl bark extracts. Evid Based Complement Alternat Med. (2017) 2017:9764843. doi: 10.1155/2017/9764843
57. Lee, YS, Kim, SH, Yuk, HJ, and Kim, DS. DKB114, a mixture of Chrysanthemum Indicum Linne flower and Cinnamomum Cassia (L.) J. Presl bark extracts, improves hyperuricemia through inhibition of xanthine oxidase activity and increasing urine excretion. Nutrients. (2018) 10:1382. doi: 10.3390/nu10101381
58. Chen, L, Liu, Y, Huang, X, Zhu, Y, Li, J, Miao, Y, et al. Comparison of chemical constituents and pharmacological effects of different varieties of Chrysanthemum Flos in China. Chem Biodivers. (2021) 18:e2100206. doi: 10.1002/cbdv.202100206
59. Liang, W-L, Gong, D, and Zhang, WK. The composition of chrysanthemum extracts and their pharmacological functions. STEMedicine. (2020) 2:69. doi: 10.37175/stemedicine.v2i5.69
60. Shao, Y, Sun, Y, Li, D, and Chen, Y. Chrysanthemum indicuml.: a comprehensive review of its botany, Phytochemistry and pharmacology. Am J Chin Med. (2020) 48:871–97. doi: 10.1142/S0192415X20500421
61. Hirano, M, Takeda, S, Hitoe, S, and Shimoda, H. Luteolin-rich chrysanthemum flower extract suppresses baseline serum uric acid in Japanese subjects with mild hyperuricemia. Integrat Mol Med. (2017) 4:275. doi: 10.15761/imm.1000275
62. Peng, A, Lin, L, Zhao, M, and Sun, B. Identifying mechanisms underlying the amelioration effect of Chrysanthemum morifolium Ramat. 'Boju' extract on hyperuricemia using biochemical characterization and UPLC-ESI-QTOF/MS-based metabolomics. Food Funct. (2019) 10:8042–55. doi: 10.1039/c9fo01821b
63. Lou, XJ, Wang, YZ, Lei, SS, He, X, Lu, TT, Zhan, LH, et al. Beneficial effects of macroporous resin Extract of Dendrobium candidum leaves in rats with hyperuricemia induced by a high-purine diet. Evid Based Complement Alternat Med. (2020) 2020:3086106. doi: 10.1155/2020/3086106
64. Wang, YZ, Zhou, C, Zhu, LJ, He, XLS, Li, LZ, Zheng, X, et al. Effects of macroporous resin Extract of Dendrobium officinale leaves in rats with hyperuricemia induced by fructose and potassium Oxonate. Comb Chem High Throughput Screen. (2022) 25:1294–303. doi: 10.2174/1386207324666210528114345
65. Zhang, D, Zhao, M, Li, Y, Zhang, D, Yang, Y, and Li, L. Natural xanthine oxidase inhibitor 5-O-Caffeoylshikimic acid ameliorates kidney injury caused by hyperuricemia in mice. Molecules. (2021) 26:7307. doi: 10.3390/molecules26237307
66. Lin, L, Liu, X, and Zhao, M. Screening of xanthine oxidase inhibitor from selected edible plants and hypouricemic effect of Rhizoma Alpiniae Officinarum extract on hyperuricemic rats. J Funct Foods. (2018) 50:26–36. doi: 10.1016/j.jff.2018.09.024
67. Liang, G, Nie, Y, Chang, Y, Zeng, S, Liang, C, Zheng, X, et al. Protective effects of Rhizoma smilacis glabrae extracts on potassium oxonate- and monosodium urate-induced hyperuricemia and gout in mice. Phytomedicine. (2019) 59:152772. doi: 10.1016/j.phymed.2018.11.032
68. Chen, N, Wang, R, Li, H, Wang, W, Wang, L, Yin, X, et al. Flavonoid extract of saffron by-product alleviates hyperuricemia via inhibiting xanthine oxidase and modulating gut microbiota. Phytother Res. (2022) 36:4604–19. doi: 10.1002/ptr.7579
69. Yong, T, Chen, S, Xie, Y, Shuai, O, Li, X, Chen, D, et al. Hypouricemic effects of extracts from Agrocybe aegerita on hyperuricemia mice and virtual prediction of bioactives by molecular docking. Front Pharmacol. (2018) 9:498. doi: 10.3389/fphar.2018.00498
70. Nosek, K, Leppert, W, Puchala, L, and Lon, K. Efficacy and safety of topical morphine: a narrative review. Pharmaceutics. (2022) 14:1499. doi: 10.3390/pharmaceutics14071499
71. Al-Maharik, N, Jaradat, N, Bassalat, N, Hawash, M, and Zaid, H. Isolation, identification and pharmacological effects of Mandragora autumnalis fruit flavonoids fraction. Molecules. (2022) 27:1046. doi: 10.3390/molecules27031046
72. El-Shahir, AA, El-Wakil, DA, Abdel Latef, AAH, and Youssef, NH. Bioactive compounds and antifungal activity of leaves and fruits Methanolic extracts of Ziziphus spina-christi L. Plants (Basel). (2022) 11:746. doi: 10.3390/plants11060746
73. Fang, HL, Liu, ML, Li, SY, Song, WQ, Ouyang, H, Xiao, ZP, et al. Identification, potency evaluation, and mechanism clarification of alpha-glucosidase inhibitors from tender leaves of Lithocarpus polystachyus Rehd. Food Chem. (2022) 371:131128. doi: 10.1016/j.foodchem.2021.131128
74. Bustanussalam, HY, Rachman, F, Septiana, E, and Simanjuntak, P. Identification of Antioxidant Active Compounds from Watercress (Nasturtium officinale R.Br). Atlantis Press. (2023). doi: 10.2991/978-94-6463-112-8_35
75. Li, X, Liu, S, Jin, W, Zhang, W, and Zheng, G. Identification of the constituents of ethyl acetate fraction from Smilax china L. and determination of xanthine oxidase inhibitory properties. Int J Mol Sci. (2023) 24:5158. doi: 10.3390/ijms24065158
76. Li, W, Yang, R, Ying, D, Yu, J, Sanguansri, L, and Augustin, MA. Analysis of polyphenols in apple pomace: a comparative study of different extraction and hydrolysis procedures. Ind Crop Prod. (2020) 147:112250. doi: 10.1016/j.indcrop.2020.112250
77. Ramón-Gonçalves, M, Gómez-Mejía, E, Rosales-Conrado, N, León-González, ME, and Madrid, Y. Extraction, identification and quantification of polyphenols from spent coffee grounds by chromatographic methods and chemometric analyses. Waste Manag. (2019) 96:15–24. doi: 10.1016/j.wasman.2019.07.009
78. Al-Amin, M, Eltayeb, NM, Rahiman, SSF, Khairuddean, M, and Salhimi, SM. UPLC-ESI-QTOF-MS/MS and 1H-NMR identification of alkaloids in potent fraction of Catharanthus roseus leaves inhibits migration and invasion of MDA-MB-231 cells. Biologia. (2022) 77:3291–303. doi: 10.1007/s11756-022-01185-6
79. Shi, B-B, Kongkiatpaiboon, S, Chen, G, Schinnerl, J, and Cai, X-H. Nematocidal alkaloids from the roots of Stemona mairei (H.Lév.)K.Krause and identification of their pharmacophoric moiety. Bioorg Chem. (2023) 130:106239. doi: 10.1016/j.bioorg.2022.106239
80. Krishnan, N, Mariappanadar, V, Dhanabalan, AK, Devadasan, V, Gopinath, SCB, and Raman, P. Purification, identification and in silico models of alkaloids from Nardostachys jatamansi — bioactive compounds for neurodegenerative diseases. Biomass Conver Biorefin. (2022) 1–12. doi: 10.1007/s13399-022-03237-y
81. Sheng, Z, Wen, L, and Yang, B. Structure identification of a polysaccharide in mushroom Lingzhi spore and its immunomodulatory activity. Carbohydr Polym. (2022) 278:118939. doi: 10.1016/j.carbpol.2021.118939
82. Ding, X, Song, Z, and Hou, Y. Comparative study on the structure characterization and immune activity of Lactarius vellereus Fr. polysaccharide (LV-1) and Cordyceps militaris (L. ex Fr.) Link. polysaccharide (CM-S). J Food Measure Charact. (2022) 16:901–19. doi: 10.1007/s11694-021-01215-y
83. Yang, B, Luo, Y, Sang, Y, and Kan, J. Isolation, purification, structural characterization, and hypoglycemic activity assessment of polysaccharides from Hovenia dulcis (Guai Zao). Int J Biol Macromol. (2022) 208:1106–15. doi: 10.1016/j.ijbiomac.2022.03.211
84. Lee, J-O, Yang, Y, Tao, Y, Yi, Y-S, and Cho, JY. Korean red ginseng saponin fraction exerts anti-inflammatory effects by targeting the NF-κB and AP-1 pathways. J Ginseng Res. (2022) 46:489–95. doi: 10.1016/j.jgr.2022.02.004
85. Zheng, Y-R, Fan, CL, Chen, Y, Quan, JY, Shi, LZ, Tian, CY, et al. Anti-inflammatory, anti-angiogenetic and antiviral activities of dammarane-type triterpenoid saponins from the roots of Panax notoginseng. Food Funct. (2022) 13:3590–602. doi: 10.1039/D1FO04089H
86. Raju, R, Mathew, S, Reddell, P, and Münch, G. Ternstroenol F: a new pentacyclic triterpenoid saponin isolated from the Australian rainforest plant Ternstroemia cherryi. Nat Prod Res. (2022) 1-6:1–6. doi: 10.1080/14786419.2022.2039139
87. Song, HP, Wang, H, Liang, JX, Qian, C, Wu, SQ, Xu, WJ, et al. Integration of multiple analytical and computational tools for the discovery of high-potency enzyme inhibitors from herbal medicines. ChemMedChem. (2016) 11:2588–97. doi: 10.1002/cmdc.201600489
88. Shoaib, M, Xin, L, and Xin, Y. Oral administration of Lactobacillus acidophilus alleviates exacerbations in Pseudomonas aeruginosa and Staphylococcus aureus pulmonary infections. Pak J Pharm Sci. (2019) 32:1621–30.
89. Yen, S-C, Chen, LC, Huang, HL, Ngo, ST, Wu, YW, Lin, TE, et al. Investigation of selected flavonoid derivatives as potent FLT3 inhibitors for the potential treatment of acute myeloid leukemia. J Nat Prod. (2021) 84:1–10. doi: 10.1021/acs.jnatprod.0c00589
90. Mo, S-F, Zhou, F, Lv, YZ, Hu, QH, Zhang, DM, and Kong, LD. Hypouricemic action of selected flavonoids in mice: structure–activity relationships. Biol Pharm Bull. (2007) 30:1551–6. doi: 10.1248/bpb.30.1551
91. Yuan, L, Bao, Z, Ma, T, and Lin, S. Hypouricemia effects of corn silk flavonoids in a mouse model of potassium oxonated-induced hyperuricemia. J Food Biochem. (2021) 45:13856. doi: 10.1111/jfbc.13856
92. Patel, RV, Mistry, BM, Shinde, SK, Syed, R, Singh, V, and Shin, HS. Therapeutic potential of quercetin as a cardiovascular agent. Eur J Med Chem. (2018) 155:889–904. doi: 10.1016/j.ejmech.2018.06.053
93. Tang, S-M, Deng, XT, Zhou, J, Li, QP, Ge, XX, and Miao, L. Pharmacological basis and new insights of quercetin action in respect to its anti-cancer effects. Biomed Pharmacother. (2020) 121:109604. doi: 10.1016/j.biopha.2019.109604
94. Di Petrillo, A, Orrù, G, Fais, A, and Fantini, MC. Quercetin and its derivates as antiviral potentials: a comprehensive review. Phytother Res. (2022) 36:266–78. doi: 10.1002/ptr.7309
95. Asgharian, P, Tazekand, AP, Hosseini, K, Forouhandeh, H, Ghasemnejad, T, Ranjbar, M, et al. Potential mechanisms of quercetin in cancer prevention: focus on cellular and molecular targets. Cancer Cell Int. (2022) 22:1-20 and 1475-2867. doi: 10.1186/s12935-022-02677-w
96. Remigante, A, Spinelli, S, Basile, N, Caruso, D, Falliti, G, Dossena, S, et al. Oxidation stress as a mechanism of aging in human erythrocytes: protective effect of quercetin. Int J Mol Sci. (2022) 23:7781 and 1422-0067. doi: 10.3390/ijms23147781
97. Nutmakul, T. A review on benefits of quercetin in hyperuricemia and gouty arthritis. Saudi Pharm J. (2022) 30:918–26. doi: 10.1016/j.jsps.2022.04.013
98. Zhang, C, Wang, R, Zhang, G, and Gong, D. Mechanistic insights into the inhibition of quercetin on xanthine oxidase. Int J Biol Macromol. (2018) 112:405–12. doi: 10.1016/j.ijbiomac.2018.01.190
99. Tumova, S, Shi, Y, Carr, IM, and Williamson, G. Effects of quercetin and metabolites on uric acid biosynthesis and consequences for gene expression in the endothelium. Free Radic Biol Med. (2021) 162:191–201. doi: 10.1016/j.freeradbiomed.2020.10.017
100. Zhu, L-H, Xu, Y-Y, Zhu, L-p, Zheng, X, Jiang, C-H, Liu, J-J, et al. Protective effects of Cyclocarya paliurus on hyperuricemia and urate-induced inflammation. J Funct Foods. (2022) 94:5130. doi: 10.1016/j.jff.2022.105130
101. Gong, G, Guan, YY, Zhang, ZL, Rahman, K, Wang, SJ, Zhou, S, et al. Isorhamnetin: a review of pharmacological effects. Biomed Pharmacother. (2020) 128:110301. doi: 10.1016/j.biopha.2020.110301
102. Adachi, SI, Kondo, S, Sato, Y, Yoshizawa, F, and Yagasaki, K. Anti-hyperuricemic effect of isorhamnetin in cultured hepatocytes and model mice: structure-activity relationships of methylquercetins as inhibitors of uric acid production. Cytotechnology. (2019) 71:181–92. doi: 10.1007/s10616-018-0275-8
103. Imran, M, Rauf, A, Abu-Izneid, T, Nadeem, M, Shariati, MA, Khan, IA, et al. Luteolin, a flavonoid, as an anticancer agent: a review. Biomed Pharmacother. (2019) 112:108612. doi: 10.1016/j.biopha.2019.108612
104. Aziz, N, Kim, M-Y, and Cho, JY. Anti-inflammatory effects of luteolin: a review of in vitro, in vivo, and in silico studies. J Ethnopharmacol. (2018) 225:342–58. doi: 10.1016/j.jep.2018.05.019
105. Cook, MT. Mechanism of metastasis suppression by luteolin in breast cancer. Breast Cancer Targets Therapy. (2018) 10:89–100. doi: 10.2147/BCTT.S144202
106. Yan, J, Zhang, G, Hu, Y, and Ma, Y. Effect of luteolin on xanthine oxidase: inhibition kinetics and interaction mechanism merging with docking simulation. Food Chem. (2013) 141:3766–73. doi: 10.1016/j.foodchem.2013.06.092
107. Lin, Y, Liu, PG, Liang, WQ, Hu, YJ, Xu, P, Zhou, J, et al. Luteolin-4'-O-glucoside and its aglycone, two major flavones of Gnaphalium affine D. Don, resist hyperuricemia and acute gouty arthritis activity in animal models. Phytomedicine. (2018) 41:54–61. doi: 10.1016/j.phymed.2018.02.002
108. Pathak, RK, Gupta, A, Shukla, R, and Baunthiyal, M. Identification of new drug-like compounds from millets as xanthine oxidoreductase inhibitors for treatment of hyperuricemia: a molecular docking and simulation study. Comput Biol Chem. (2018) 76:32–41. doi: 10.1016/j.compbiolchem.2018.05.015
109. Zhu, JX, Yang, HY, Hu, WQ, Cheng, J, Liu, Y, Yi, LT, et al. Active components from Lagotis brachystachya maintain uric acid homeostasis by inhibiting renal TLR4-NLRP3 signaling in hyperuricemic mice. Inflammopharmacology. (2021) 29:1187–200. doi: 10.1007/s10787-021-00844-5
110. Wang, J, Zhang, Y, Zhou, M, Zheng, M, Cui, J, Liu, Z, et al. Rapid screening and evaluation of XOD inhibitors and O(2) (•-) scavenger from total flavonoids of Ginkgo biloba leaves by LC-MS and multimode microplate reader. Biomed Chromatogr. (2020) 34:e4852. doi: 10.1002/bmc.4852
111. Song, D, Zhao, H, Wang, L, Wang, F, Fang, L, and Zhao, X. Ethanol extract of Sophora japonica flower bud, an effective potential dietary supplement for the treatment of hyperuricemia. Food Biosci. (2023) 52:102457. doi: 10.1016/j.fbio.2023.102457
112. Li, J, Gong, Y, Li, J, and Fan, L. Improving the xanthine oxidase and adenosine deaminase inhibitory activities of Flos Sophorae Immaturus by ultrasound-assisted heating treatments. Food Biosci. (2023) 51:102245. doi: 10.1016/j.fbio.2022.102245
113. Li, J, Gong, Y, Li, J, and Fan, L. In vitro inhibitory effects of polyphenols from Tartary buckwheat on xanthine oxidase: identification, inhibitory activity, and action mechanism. Food Chem. (2022) 379:132100. doi: 10.1016/j.foodchem.2022.132100
114. Szwajgier, D, Borowiec, K, and Pustelniak, K. The neuroprotective effects of phenolic acids: molecular mechanism of action. Nutrients. (2017) 9:477. doi: 10.3390/nu9050477
115. Kumar, N, and Goel, N. Phenolic acids: natural versatile molecules with promising therapeutic applications. Biotechnol Rep. (2019) 24:e00370. doi: 10.1016/j.btre.2019.e00370
116. Chen, J, Yang, J, Ma, L, Li, J, Shahzad, N, and Kim, CK. Structure-antioxidant activity relationship of methoxy, phenolic hydroxyl, and carboxylic acid groups of phenolic acids. Sci Rep. (2020) 10:2611. doi: 10.1038/s41598-020-59451-z
117. Caruso, G, Godos, J, Privitera, A, Lanza, G, Castellano, S, Chillemi, A, et al. Phenolic acids and prevention of cognitive decline: polyphenols with a neuroprotective role in cognitive disorders and Alzheimer's disease. Nutrients. (2022) 14:819. doi: 10.3390/nu14040819
118. Mehmood, A, Zhao, L, Ishaq, M, Safdar, B, Wang, C, and Nadeem, M. Optimization of total phenolic contents, antioxidant, andin-vitroxanthine oxidase inhibitory activity of sunflower head. CyTA J Food. (2018) 16:957–64. doi: 10.1080/19476337.2018.1504121
119. Wan, Y, Wang, D, Shen, Y, Chen, Y, Qian, J, and Fu, G. Effect of Lactobacillus acidophilus fermentation on the composition of chlorogenic acids and anti-hyperuricemia activity of Artemisia selengensis Turcz. Food Funct. (2022) 13:11780–93. doi: 10.1039/d2fo01854c
120. Wianowska, D, and Gil, M. Recent advances in extraction and analysis procedures of natural chlorogenic acids. Phytochem Rev. (2019) 18:273–302. doi: 10.1007/s11101-018-9592-y
121. Wu, S, Chen, W, Lu, S, Zhang, H, and Yin, L. Metabolic engineering of Shikimic acid biosynthesis pathway for the production of Shikimic acid and its branched products in microorganisms: advances and prospects. Molecules. (2022) 27:779. doi: 10.3390/molecules27154779
122. Ferraz-Filha, ZS, Ferrari, FC, Araujo, M, Bernardes, A, and Saude-Guimaraes, DA. Effects of the aqueous Extract from Tabebuia roseoalba and phenolic acids on hyperuricemia and inflammation. Evid Based Complement Alternat Med. (2017) 2017:2712108. doi: 10.1155/2017/2712108
123. Zhou, X, Zhang, B, Zhao, X, Lin, Y, Wang, J, Wang, X, et al. Chlorogenic acid supplementation ameliorates hyperuricemia, relieves renal inflammation, and modulates intestinal homeostasis. Food Funct. (2021) 12:5637–49. doi: 10.1039/d0fo03199b
124. Khan, F, Bamunuarachchi, NI, Tabassum, N, and Kim, YM. Caffeic acid and its derivatives: antimicrobial drugs toward microbial pathogens. J Agric Food Chem. (2021) 69:2979–3004. doi: 10.1021/acs.jafc.0c07579
125. Mirzaei, S, Gholami, MH, Zabolian, A, Saleki, H, Farahani, MV, Hamzehlou, S, et al. Caffeic acid and its derivatives as potential modulators of oncogenic molecular pathways: new hope in the fight against cancer. Pharmacol Res. (2021) 171:105759. doi: 10.1016/j.phrs.2021.105759
126. Tajner-Czopek, A, Gertchen, M, Rytel, E, Kita, A, Kucharska, AZ, and Sokół-Łętowska, A. Study of antioxidant activity of some medicinal plants having high content of caffeic acid derivatives. Antioxidants. (2020) 9:412 and 2076-3921. doi: 10.3390/antiox9050412
127. Chahardoli, A, Qalekhani, F, Shokoohinia, Y, and Fattahi, A. Caffeic acid based titanium dioxide nanoparticles: blood compatibility, anti-inflammatory, and cytotoxicity. J Mol Liq. (2022) 361:119674. doi: 10.1016/j.molliq.2022.119674
128. Wan, Y, Wang, F, Zou, B, Shen, Y, Li, Y, Zhang, A, et al. Molecular mechanism underlying the ability of caffeic acid to decrease uric acid levels in hyperuricemia rats. J Funct Foods. (2019) 57:150–6. doi: 10.1016/j.jff.2019.03.038
129. de Sa, M, Muller, C, Coelho, GB, de Paula, C, Michel Araujo, M, and Saude-Guimaraes, DA. Lychnophora pinaster ethanolic extract and its chemical constituents ameliorate hyperuricemia and related inflammation. J Ethnopharmacol. (2019) 242:112040. doi: 10.1016/j.jep.2019.112040
130. Bai, J, Zhang, Y, Tang, C, Hou, Y, Ai, X, Chen, X, et al. Gallic acid: pharmacological activities and molecular mechanisms involved in inflammation-related diseases. Biomed Pharmacother. (2021) 133:110985. doi: 10.1016/j.biopha.2020.110985
131. Zhang, T, Ma, L, Wu, P, Li, W, Li, T, Gu, R, et al. Gallic acid has anticancer activity and enhances the anticancer effects of cisplatin in non-small cell lung cancer A549 cells via the JAK/STAT3 signaling pathway. Oncol Rep. (2019) 41:1779–88. doi: 10.3892/or.2019.6976
132. Nouri, A, Heibati, F, and Heidarian, E. Gallic acid exerts anti-inflammatory, anti-oxidative stress, and nephroprotective effects against paraquat-induced renal injury in male rats. Naunyn Schmiedeberg's Arch Pharmacol. (2021) 394:1–9. doi: 10.1007/s00210-020-01931-0
133. Sui, Y, Xu, D, and Sun, X. Identification of anti-hyperuricemic components from Coix seed. Food Biosci. (2023) 52:102461. doi: 10.1016/j.fbio.2023.102461
134. Sun, R, Kan, J, Cai, H, Hong, J, Jin, C, and Zhang, M. In vitro and in vivo ameliorative effects of polyphenols from purple potato leaves on renal injury and associated inflammation induced by hyperuricemia. J Food Biochem. (2022) 46:e14049. doi: 10.1111/jfbc.14049
135. Ng, TL, Loh, KE, Tan, SA, Tan, HY, Yue, CS, Wee, SP, et al. Anti-Hyperuricemic effect of ethyl acetate sub-fractions from Chrysanthemum morifolium Ramat. Dried flowers on potassium Oxonate-induced Hyperuricemic rats. Appl Sci. (2022) 12:3487. doi: 10.3390/app12073487
136. Debnath, B, Singh, WS, das, M, Goswami, S, Singh, MK, Maiti, D, et al. Role of plant alkaloids on human health: a review of biological activities. Materials Today Chem. (2018) 9:56–72. doi: 10.1016/j.mtchem.2018.05.001
137. Liu, C, Yang, S, Wang, K, Bao, X, Liu, Y, Zhou, S, et al. Alkaloids from traditional Chinese medicine against hepatocellular carcinoma. Biomed Pharmacother. (2019) 120:109543. doi: 10.1016/j.biopha.2019.109543
138. Othman, L, Sleiman, A, and Abdel-Massih, RM. Antimicrobial activity of polyphenols and alkaloids in middle eastern plants. Front Microbiol. (2019) 10:911 and 1664-1302X. doi: 10.3389/fmicb.2019.00911
139. Wang, M-X, Liu, Y-L, Yang, Y, Zhang, D-M, and Kong, L-D. Nuciferine restores potassium oxonate-induced hyperuricemia and kidney inflammation in mice. Eur J Pharmacol. (2015) 747:59–70. doi: 10.1016/j.ejphar.2014.11.035
140. Wang, L., et al. The bacteriostatic effect and mechanism of berberine on methicillin resistant Staphylococcus aureus in vitro. Research Square (2020). doi: 10.21203/rs.2.9640/v3
141. Chen, Q, Li, D, Wu, F, He, X, Zhou, Y, Sun, C, et al. Berberine regulates the metabolism of uric acid and modulates intestinal Flora in hyperuricemia rats model. Comb Chem High Throughput Screen. (2023) 26:2057–66. doi: 10.2174/1386207326666221124093228
142. Li, Q, Huang, Z, Liu, D, Zheng, J, Xie, J, Chen, J, et al. Effect of Berberine on hyperuricemia and kidney injury: a network pharmacology analysis and experimental validation in a mouse model. Drug Des Devel Ther. (2021) 15:3241–54. doi: 10.2147/DDDT.S317776
143. Lin, G, Yu, Q, Xu, L, Huang, Z, Mai, L, Jiang, L, et al. Berberrubine attenuates potassium oxonate- and hypoxanthine-induced hyperuricemia by regulating urate transporters and JAK2/STAT3 signaling pathway. Eur J Pharmacol. (2021) 912:174592. doi: 10.1016/j.ejphar.2021.174592
144. Naz, H, Naz, S, Miraj, R, Zaheer, A, Azam, N, Mughal, IS, et al. The effect of Berberine, a drug from Chinese folk medicine, on serum and urinary uric acid levels in rats with hyperuricemia. Cureus. (2021) 13:e13186. doi: 10.7759/cureus.13186
145. Zhong, L, Lin, Y, Gong, S, Wu, X, Liu, Y, Chen, J, et al. Oxyberberrubine, a novel liver microsomes-mediated secondary metabolite of berberine, alleviates hyperuricemic nephropathy in mice. Phytomedicine. (2023) 108:154521. doi: 10.1016/j.phymed.2022.154521
146. Zhao, L-Q, Zhao, Y-L, He, Y-J, Yang, X-W, and Luo, X-D. Tuberindine a, a truffle alkaloid with an unprecedented skeleton exhibiting anti-hyperuricemic bioactivity. Org Lett. (2022) 24:4333–7. doi: 10.1021/acs.orglett.2c01421
147. Wang, LM, Wang, P, Teka, T, Zhang, YC, Yang, WZ, Zhang, Y, et al. (1)H NMR and UHPLC/Q-Orbitrap-MS-based metabolomics combined with 16S rRNA gut microbiota analysis revealed the potential regulation mechanism of Nuciferine in hyperuricemia rats. J Agric Food Chem. (2020) 68:14059–70. doi: 10.1021/acs.jafc.0c04985
148. Yao, W, Qiu, H-M, Cheong, K-L, and Zhong, S. Advances in anti-cancer effects and underlying mechanisms of marine algae polysaccharides. Int J Biol Macromol. (2022) 221:472–85. doi: 10.1016/j.ijbiomac.2022.09.055
149. Yang, W, Zhao, P, Li, X, Guo, L, and Gao, W. The potential roles of natural plant polysaccharides in inflammatory bowel disease: a review. Carbohydr Polym. (2022) 277:118821. doi: 10.1016/j.carbpol.2021.118821
150. Zhou, Y, Chen, X, Chen, T, and Chen, X. A review of the antibacterial activity and mechanisms of plant polysaccharides. Trends Food Sci Technol. (2022) 123:264–80. doi: 10.1016/j.tifs.2022.03.020
151. Li, X, Gao, X, Zhang, H, Liu, Y, Sarker, MMR, Wu, Y, et al. The anti-hyperuricemic effects of green alga Enteromorpha prolifera polysaccharide via regulation of the uric acid transporters in vivo. Food Chem Toxicol. (2021) 158:112630. doi: 10.1016/j.fct.2021.112630
152. Yang, Q, Wang, Q, Deng, W, Sun, C, Wei, Q, Adu-Frimpong, M, et al. Anti-hyperuricemic and anti-gouty arthritis activities of polysaccharide purified from Lonicera japonica in model rats. Int J Biol Macromol. (2019) 123:801–9. doi: 10.1016/j.ijbiomac.2018.11.077
153. Yu, X, Zhang, L, Zhang, P, Zhi, J, Xing, R, and He, L. Lycium barbarum polysaccharides protect mice from hyperuricaemia through promoting kidney excretion of uric acid and inhibiting liver xanthine oxidase. Pharm Biol. (2020) 58:944–9. doi: 10.1080/13880209.2020.1817951
154. Lin, S, Meng, J, Li, F, Yu, H, Lin, D, Lin, S, et al. Ganoderma lucidum polysaccharide peptide alleviates hyperuricemia by regulating adenosine deaminase and urate transporters. Food Funct. (2022) 13:12619–31. doi: 10.1039/d2fo02431d
155. Huang, CH, Chen, TY, and Tsai, GJ. Hypouricemic effect of submerged culture of Ganoderma lucidum in potassium Oxonate-induced Hyperuricemic rats. Meta. (2022) 12:553. doi: 10.3390/metabo12060553
156. Jiang, Y, Shang, Z, Lv, X, du, M, Ma, L, Hou, G, et al. Structure elucidation and antitumor activity of a water soluble polysaccharide from Hemicentrotus pulcherrimus. Carbohydr Polym. (2022) 292:119718. doi: 10.1016/j.carbpol.2022.119718
157. Qiu, A, Wang, Y, Zhang, G, and Wang, H. Natural polysaccharide-based Nanodrug delivery Systems for Treatment of diabetes %M. Polymers. (2022) 14:2073–4360. doi: 10.3390/polym14153217%U
158. Zhao, YC, Xue, CH, Zhang, TT, and Wang, YM. Saponins from sea cucumber and their biological activities. J Agric Food Chem. (2018) 66:7222–37. doi: 10.1021/acs.jafc.8b01770
159. Reichert, CL, Salminen, H, and Weiss, J. Quillaja saponin characteristics and functional properties. Annual review of. Food Sci Technol. (2019) 10:43–73. doi: 10.1146/annurev-food-032818-122010
160. Figueiredo, CCM, da Costa Gomes, A, Granero, FO, Bronzel Junior, JL, Silva, LP, Ximenes, VF, et al. Antiglycation and antioxidant activities of the crude extract and saponin fraction of Tribulus terrestris before and after microcapsule release. J Integr Med. (2022) 20:153–62. doi: 10.1016/j.joim.2021.12.003
161. Xu, H-L, Chen, GH, Wu, YT, Xie, LP, Tan, ZB, Liu, B, et al. Ginsenoside Ro, an oleanolic saponin of Panax ginseng, exerts anti-inflammatory effect by direct inhibiting toll like receptor 4 signaling pathway. J Ginseng Res. (2022) 46:156–66. doi: 10.1016/j.jgr.2021.05.011
162. Xiong, XY, Liang, J, Guo, SY, Dai, MZ, Zhou, JL, Zhang, Y, et al. A natural complex product Yaocha reduces uric acid level in a live zebrafish model. J Pharmacol Toxicol Methods. (2020) 102:106681. doi: 10.1016/j.vascn.2020.106681
163. Zhu, L, Dong, Y, Na, S, Han, R, Wei, C, and Chen, G. Saponins extracted from Dioscorea collettii rhizomes regulate the expression of urate transporters in chronic hyperuricemia rats. Biomed Pharmacother. (2017) 93:88–94. doi: 10.1016/j.biopha.2017.06.022
164. Wu, X-H, Ruan, J-L, Zhang, J, Wang, S-Q, and Zhang, Y-W. Pallidifloside D, a saponin glycoside constituent from Smilax riparia, resist to hyperuricemia based on URAT1 and GLUT9 in hyperuricemic mice. J Ethnopharmacol. (2014) 157, 201-205 and 0378-8741:201–5. doi: 10.1016/j.jep.2014.09.034
165. Zhou, Q, Yu, D-h, Zhang, N, and Liu, S-M. Anti-inflammatory effect of total saponin fraction from Dioscorea nipponica Makino on gouty arthritis and its influence on NALP3 inflammasome. Chin J Integr Med. (2019) 25, 663-670 and 1993-0402:663–70. doi: 10.1007/s11655-016-2741-5
Keywords: hyperuricemia, medicinal and edible plants, xanthine oxidase, uric acid transporters, bioactive components, mechanisms of action
Citation: Cheng-yuan W and Jian-gang D (2023) Research progress on the prevention and treatment of hyperuricemia by medicinal and edible plants and its bioactive components. Front. Nutr. 10:1186161. doi: 10.3389/fnut.2023.1186161
Edited by:
Dasha Mihaylova, University of Food Technologies, BulgariaReviewed by:
Lanzhou Li, Jilin Agricultural University, ChinaCopyright © 2023 Cheng-yuan and Jian-gang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dai Jian-gang, ZGFpamlhbmdhbmcxOTk3QDE2My5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.