
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Nutr., 21 September 2022
Sec. Nutrition and Food Science Technology
Volume 9 - 2022 | https://doi.org/10.3389/fnut.2022.966734
This article is part of the Research TopicWomen in Nutrition and Food Science TechnologyView all 9 articles
 Nelson Andrade1,2*
Nelson Andrade1,2* Juliana A. Barreto Peixoto1
Juliana A. Barreto Peixoto1 M. Beatriz P. P. Oliveira1
M. Beatriz P. P. Oliveira1 Fátima Martel2,3
Fátima Martel2,3 Rita C. Alves1*
Rita C. Alves1*Coffee is one of the most consumed products in the world, and its by-products are mainly discarded as waste. In order to solve this problem and in the context of a sustainable industrial attitude, coffee by-products have been studied concerning their chemical and nutritional features for a potential application in foodstuffs or dietary supplements. Under this perspective, coffee silverskin, the main by-product of coffee roasting, stands out as a noteworthy source of nutrients and remarkable bioactive compounds, such as chlorogenic acids, caffeine, and melanoidins, among others. Such compounds have been demonstrating beneficial health properties in the context of metabolic disorders. This mini-review compiles and discusses the potential health benefits of coffee silverskin and its main bioactive components on metabolic syndrome, highlighting the main biochemical mechanisms involved, namely their effects upon intestinal sugar uptake, glucose and lipids metabolism, oxidative stress, and gut microbiota. Even though additional research on this coffee by-product is needed, silverskin can be highlighted as an interesting source of compounds that could be used in the prevention or co-treatment of metabolic syndrome. Simultaneously, the valorization of this by-product also responds to the sustainability and circular economy needs of the coffee chain.
Coffee is one of the most produced, traded, and consumed commodities worldwide. Only in 2020/2021, around 10.13 and 10.06 million tons of coffee beans were produced and consumed, respectively, around the globe (1). Despite the multiplicity of existing and identified coffee species, Coffea arabica L. and Coffea canephora Pierre (known as arabica and robusta, respectively) are the most important in the international coffee trade, representing around 98% of the market (2). Besides the unique sensory and pleasant flavor of coffee, which alone contribute significantly to the popularity and attractiveness of this beverage, its regular consumption has been associated to several health benefits (3). The functional activity of coffee is attributed to a wide range of bioactive components such as phenolic compounds (mainly chlorogenic acids), alkaloids (caffeine and trigonelline), diterpenes (cafestol and kahweol), and other secondary metabolites (4). Improvements in mental alertness (5), reduced risk of diseases development [type 2 diabetes (6), depression (7), suicidal behavior (8), cancer (9), hepatic injuries (10, 11), neurodegenerative diseases (12), and cardiovascular disorders (13)], and positive effects on the gastrointestinal tract and gut microbiota (14) are well–documented benefits.
On the other hand, coffee processing involves different serial operations which generate distinct by-products (husks, pulp, mucilage, parchment, silverskin, and spent coffee grounds) (15). In total, coffee industries produce more than 10 million tons of solid residues per year (16), representing a serious environmental problem. Basically, all of them hold a significant nutritional and bioactive potential, therefore, their use/recycling into health promoting products is of great interest and can also bring positive socioeconomic and environmental impacts (17, 18).
Coffee silverskin, in particular, is a thin tegument that covers the green coffee beans and is detached during the roast when these expand, being the major by-product of coffee roasting companies (19). Although it is generally discarded, silverskin has also been highlighted as a natural source of dietary fiber, protein and bioactive compounds that can be further used, directly of after extraction, in the development of several functional food products (20, 21).
Metabolic syndrome (MetS) is a set of metabolic disorders (e.g., insulin resistance, hypertriglyceridemia, and abdominal obesity) that increases the risk for type 2 diabetes, cardiovascular diseases, and even cancer (22). A sedentary lifestyle, unhealthy eating habits (e.g., high sugar consumption), and increased stress, are factors that play a crucial role in this syndrome (23). Type II diabetes and obesity emerged as major causes of MetS, which is significantly increasing in modern societies (24). According to The International Diabetes Federation, one-quarter of the world's population has MetS (25). This alarming estimation highlights the urgent need for new ways of preventing and/or treating this syndrome.
Plant extracts have been attracting much attention as potential preventive or co-therapeutic agents against MetS due to their multiple targets and lower side effects (26). In this context, much attention has been given to natural bioactive compounds, namely those which can be found in coffee silverskin, as they have been presented as a weapon for the prevention and treatment of MetS (27, 28). The aim of this mini review is to explore the current evidence linking coffee silverskin and its main bioactive compounds to a lower risk of developing MetS, emphasizing the known mechanisms of action that could be involved in such protection.
Silverskin is rich in dietary fiber (~60 g/100 g, mainly insoluble one) (29) and protein (~12 g/100 g) (30), but its fat content is low (often below 3g/100 g) (29, 31). It is also a source of minerals (8 g/100 g in total), mainly potassium (5 g/100 g), magnesium (2 g/100 g), and calcium (0.5 g/100 g) (29). The principal amino acids found in silverskin protein are aspartic acid (10.2 mg/g) and glutamic acid (9.2 mg/g), however branched chain amino acids (leucine, isoleucine, and valine) are also present in significant levels (7.6, 4.7, and 5.0 mg/g) (30). Moreover, hydrosoluble vitamins B2 (0.18–0.2 μg/g), and B3 (2.5–3.1 μg/g) (32) are also present, as well as the liposoluble vitamin E, which is represented by different vitamers (being α-tocopherol the major one: 22.5 μg/g) (29). Silverskin also contains caffeine (~1 g/100 g), a widely recognized coffee stimulant that acts mainly by inhibiting adenosine receptors (33). In Figure 1, the structural similarity between caffeine and adenosine is highlighted. Other action mechanisms of caffeine are also described, such as inhibition of phosphodiesterases (cyclic AMP inactivating enzymes) or mobilization of intracellular calcium (33).
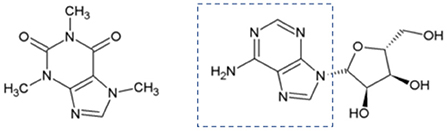
Figure 1. Chemical structures of caffeine (left) and adenosine (right).
Note: Adenosine is a potent endogenous neuromodulator, which inhibits the release of several neurotransmitters, such as glutamate, gamma-aminobutyric acid, acetylcholine and monoamines (33).
As for coffee beans, chlorogenic acids (GCA) are the major phenolics present in silverskin. CGA represent a whole set of hydroxycinnamic esters with quinic acid. It is known that this group of compounds plays crucial roles on the regulation of glucose and lipid metabolism, with several beneficial health effects described (e.g., antidiabetic, anti-inflammatory, antitumoral, etc.,) (34). The major chlorogenic acid of silverskin is 5-caffeoylquinic acid (0.1–0.2 g/100 g; Figure 2), but other CGA are also present, namely, 3-caffeoylquinic acid, 4-caffeoylquinic acid, 3-feruloylquinic acid, 5-feruloylquinic acid, and dicaffeoylquinic acids, among others (35).
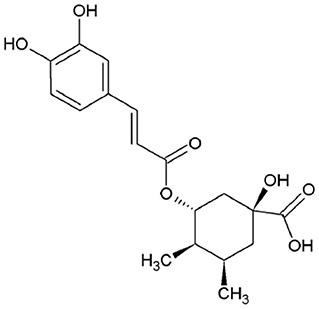
Figure 2. Chemical structure of 5-caffeoylquinic acid.
In addition to these bioactive components, melanoidins are also formed during coffee roasting. These are a complex group of polymeric structures generated during the last stage of the Maillard reaction, being responsible for the brown color, taste, and texture of foods exposed to high temperatures (36). Melanoidins result from reactions among the amino group of amino acids, proteins, or vitamins and the carbonyl group of reductive sugars or oxidized lipids. Their chemical structure comprises, thus, nitrogen-containing anionic compounds of high molecular weight (37). Phenolic compounds can also be linked to the melanoidin nucleus via covalent interactions or through non-covalent bonds, being released during intestinal digestion (36, 38). Indeed, it has been reported that some melanoidins are extensively metabolized by gut microbiota also leading to the production of short chain fatty acids and favoring the growth of beneficial bacteria (39).
The main bioactive compounds present in coffee silverskin (above detailed) can affect several pathways involved in the pathogenesis of MetS. In fact, they have been associated to beneficial effects on type II diabetes (40, 41), obesity (42), insulin resistance (43), oxidative stress (40, 44), regulation of glycemic and lipid disorders (45), and other MetS features. Those aspects and mechanisms will be presented and discussed below.
It is also important to briefly highlight the role of metabolites, although they are not the main focus of this mini review. For example, extensive research on CGA metabolism has revealed that one-third of these consumed compounds are absorbed throughout the digestive tract and further metabolized (e.g., by partial intestinal hydrolysis, phase two metabolism involving sulfation and glucuronidation…), with a wide range of variability among individuals. According to a recent study, for instance, ferulic acid undergoes extensive phase II metabolism in the liver due to the presence of hydroxyl groups (conjugation reactions—glucuronic acid and ferulic acid sulfate are the major metabolites in rat plasma and urine) (46). In turn, the unabsorbed CGA are extensively hydrolyzed by gut bacteria, resulting in phenolic acids which are then methylated, conjugated with sulfate or glycine (mostly CGA colonic metabolites: ferulic, isoferulic, and hydroxybenzoic acids), functioning as a potential prebiotic for beneficial bacteria (47, 48).
Regarding caffeine (whose biological effects are mostly dependent on its biotransformation in the body), as it is extensively metabolized in the liver by phase I (cytochrome P450) enzymes, paraxanthine, xanthines, and methyluric acids are the main metabolites found in plasma and urine (49).
Further, melanoidin's carbohydrates are typically metabolized inside the colon by gut microbiota to short-chain fatty acids (acetate, lactate, butyrate, and propionate) having a important role in gut homoeostasis, digestion, and illness prevention.
Nevertheless, because there is still some debate regarding how these metabolites are assessed (methodologically and in terms of the models utilized), more research on their bioavailability, concentrations, and activity in different tissues and organs after absorption is required.
Regarding the effect of silverskin components on glucose uptake, CGA have been found to reduce the intestinal absorption of glucose in rats, by stimulating dispersal of the Na+ electrochemical gradient, thus inhibiting Na+-dependent glucose absorption (45, 50). In fact, direct effects on glucose tolerance appear to be caused by the antagonistic effect of CGA (with/without caffeine) on glucose transport, shifting glucose absorption to more distal parts of the intestine (45). CGA (mainly 5- and 3-CQA) also affects glucose absorption through the reduction of α-amylase and α-glucosidase activities and α-amylase activity is also decreased in the presence of caffeine, resulting in reduced uptake of glucose (51) and suggesting a positive role in glycaemic control (52) (Figure 3), Further, CGA from coffee sources (coffee silverskin included) was able to significantly attenuate the postprandial release of glucose-dependent insulinotropic polypeptide (GIP), a gut hormone that potentiates insulin secretion, in the proximal part of the small intestine (53). As the amount of glucose absorbed at the intestinal barrier determines the magnitude of the GIP response, these results suggest that CGA (5-CQA) is likely to decrease the intestinal absorption of glucose (34). Additionally, melanoidin metabolites stimulate the secretion of GLP-1 by the enteroendocrine cells, causing a delay in the digestion of sugars (41).
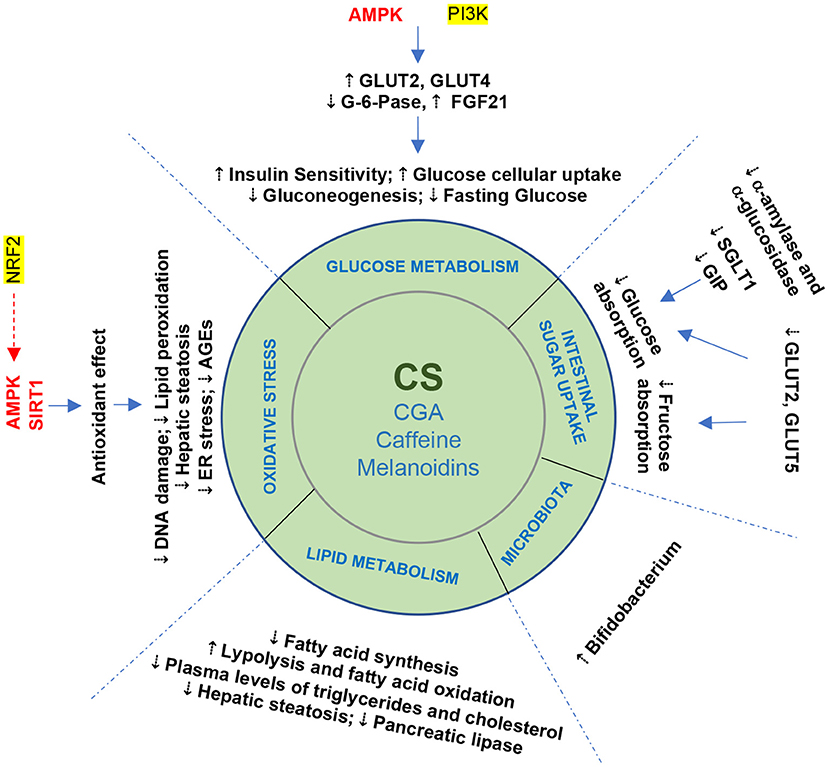
Figure 3. Beneficial effects of coffee silverskin and its bioactive compounds on metabolic syndrome: summary of the potential mechanisms involved.
In vitro, a subset of coffee CGAs reduced glucose absorption in intestinal Caco-2 cells (54). Further, among the potential bioactive-derived metabolites present in the coffee silverskin, ferulic acid has shown effects on glucose uptake. In fact, another in vitro study in adipocyte cells suggested that ferulic acid enhances glucose uptake through the PI3K-dependent pathway (55).
Moreover, a recent work suggested that coffee silverskin aqueous extract is also rich in other phenolics, such as phenolic acids (caffeic, ferulic, gallic, p-coumaric, syringic, and vanillic acids) and flavonoids (rutin, quercetin, kaempferol) (56), and these compounds might be able to interact directly with glucose and fructose intestinal transporters (Figure 3) such as GLUT2, GLUT5, and SGLT1 (57, 58). This constitutes another potential explanation for the inhibitory effect of silverskin extracts on the intestinal absorption of sugar. In reality, the evidence that coffee silverskin extracts, CGA, caffeine, and melanoidins can reduce sugar absorption in the intestinal lumen is still scarce and more studies are needed.
In what concerns to sugar consumption, fructose intake plays a prominent role, and its intestinal absorption and metabolism have been closely correlated with obesity and MetS (27). The bioactive components present in silverskin extracts (fiber, phenolics, flavonoids) were shown to inhibit the absorption of this sugar by intestinal epithelial cells in vitro (57). However, more research is still needed to better understand this effect.
The CGA family are specific competitive inhibitors of glucose-6-phosphate translocase enzyme in rat liver microsomes, which is involved in the regulation of blood glucose levels (59), thus exerting a hypoglycaemic activity (34) and modulating glucose metabolism (60).
At the intracellular level, CGA activates adenosine monophosphate-activated protein kinase (AMPK), a sensor and regulator of cellular energy balance, responsible for GLUT2 and GLUT4 translocation from intracellular to plasma membranes (Figure 3), thus increasing glucose blood removal and leading to beneficial metabolic effects, such as the inhibition of fatty acid synthesis and hepatic glucose production, contributing to the regulation of lipid and glucose metabolism (45, 59). Also, CGA presented insulin-sensitizing activity by activating glucose uptake through AMPK pathway in hepatocytes and skeletal muscle cells (60).
On the other hand, in the muscle, caffeine is able to stimulate AMPK (61) and to increase the amount of glucose transporters (GLUT4) (62), improving glucose uptake and insulin sensitivity in the body, and playing an important role in glucose homeostasis (61, 63). Moreover, in adipose cells, caffeine was able to inhibit insulin-induced Akt activation, resulting in decreased glucose transport and GLUT4 translocation (64).
As to silverskin, consumption of silverskin aqueous extract reduces fasting plasma glucose concentrations, increase the sensitivity to insulin, and slow the appearance of glucose in circulation after a glucose load (45, 65).
Likewise, a recent work suggested that aqueous extracts from coffee silverskin (125 μg/mL) stimulated phosphatidylinositol 3-kinase (PI3K) and AMPK signaling (66). The effect on insulin and PI3K pathways improved hepatic cell insulin sensitivity (in vitro), and diminished lipogenesis and gluconeogenesis (67). In this regard, ferulic acid, an important metabolite of feruloylquinic acids, has been shown to increase the expression levels of PI3-K and GLUT4, thereby increasing glucose uptake in adipocytes (55).
A recent study, using aqueous extract of coffee silverskin on hepatic cells (10 to 500 μg/mL), suggested that bioactive compounds, particularly CGA, may regulate glucose metabolism and prevent the development of NAFLD by preventing hyperglycemia and systemic insulin resistance (67).
Lastly, according to a recent randomized controlled trial, coffee melanoidins seem to be able to reduce blood glucose peak and insulin response by potentiating the effect of ghrelin and attenuating the response of orexin-A and β-endorphin (41). Therefore, because their basic chemical composition is similar, the melanoidins in the coffee silverskin extract might be expected to have a similar effect.
To summarize, the results of the numerous studies suggest that silverskin has a vast and flexible potential for impact on glucose metabolism, and that CGA and caffeine seem to be the main contributors to these effects of silverskin extract (Figure 3).
Dyslipidemia increases the risk of cardiovascular diseases and, consequently, of morbimortality (68). It is a multifactorial disorder observed in many metabolic disorders such as type II diabetes, obesity and MetS, that includes hepatic overproduction of very low-density lipoproteins (VLDL), decreased triglycerides lipolysis, impaired peripheral free fatty acid (FFA) trapping, increased FFA fluxes from adipocytes to the liver and other tissues (Figure 3), and a higher formation of small dense low-density protein (LDL). In this context, del Castillo et al. (65) have pointed out that silverskin aqueous extract (140 mg/Kg) was able to reduce total cholesterol and triglyceride plasma levels in rats and suggested that one possible mechanism of action was the inhibition of pancreatic lipase, a key enzyme for fat digestion. This liporegulatory character might have preventive and therapeutic effects in obesity and MetS contexts (65). Additionally, a work based on an animal model showed that a silverskin-based beverage had an inhibitory effect on fat accumulation, suggesting it could be a natural alternative for the prevention of obesity and MetS (69) (Figure 3).
Several studies associated the anti-obesity effect of coffee to its bioactive compounds (CGAs, caffeine and melanoidins), which are also present in coffee silverskin, and different mechanisms have been proposed by which they regulate lipid metabolism, including modulation of cell signaling, inhibition of pancreatic lipase, regulation of hepatic lipid metabolism-related enzymes, and reduction in hepatic fat accumulation in the rat model (70). In addition, a meta-analysis by Tabrizi et al. (71) suggested that the long-term consumption of caffeine sources might protect against type II diabetes through increased metabolic rate and thermogenesis, and stimulation of fat oxidation and free fatty acid release from peripheral tissues mediated by AMPK induction (Figure 3).
Using an adipocyte cell line, Rebollo-Hernanz et al. (66) demonstrated that a coffee silverskin aqueous extract (31 μg/mL) was able to inhibit cell differentiation, increase adipocyte lipid metabolism and induce lipolysis through the regulation of lipases. These effects seem to occur mainly through inactivation of ERK, JNK, and NF-κB signaling pathways (66) (Figure 3).
Furthermore, an animal model study with diabetic rats using ferulic acid (a major metabolite of feruloylquinic acids) was able to reduce the level of total cholesterol, low density cholesterol, very low-density lipoprotein, and triglycerides (72).
Recently, a meta-analysis of randomized controlled trials concluded that caffeine has been widely used as a practical approach to obesity control by increasing the release of norepinephrine and dopamine and therefore stimulating neuronal activity in different brain regions, which in turn can decrease body weight and the fat content. Furthermore, this compound can increase fat oxidation by inhibiting phosphodiesterase and suppressing the inhibitory effects of adenosine on noradrenaline release, demonstrating that caffeine intake might promote weight, body mass index and body fat reduction (71).
As aforementioned, the antioxidant capacity of coffee silverskin is related to the presence of natural constituents like CGA and caffeine, and compounds formed during coffee roasting (melanoidins) (21, 36). Regarding caffeine, some of the reported beneficial health effects are linked to its antioxidant properties (73). A work conducted by del Castillo and colleagues have suggested that caffeine (0.434 mg per dose of aqueous coffee silverskin extract) were able to protect the pancreas against oxidative stress after streptozotocin-induced diabetes in rats (74).
In this context, we should also look at the potential of metabolites of these bioactives. Thus, in a diabetes model, animals treated with a typical CGA metabolite (ferulic acid) showed improved activities of the antioxidant enzymes, SOD and catalase, and reduced glutathione in pancreatic tissue (75).
An in vitro study using hepatic cells verified that silverskin aqueous extracts (100 μg/ml) could protect cells from benzo(a)pyrene, an oxidative agent-induced DNA damage. The reduction on DNA strand breaks was suggested to be related to the presence of antioxidant agents such as CGAs and roast-associated constituents (76).
Moreover, melanoidins are known to possess antiglycative, chelating and antioxidant properties (77). Thus, these compounds together with CGA and caffeine may possess a synergic inhibitory effect on the formation of advanced glycation end products (AGEs) and related diseases (78).
Indeed, studies in animals and humans found that the ingestion of phytochemicals from coffee (and by extrapolation, its by-products) induce an adaptive cellular response characterized by upregulation and de novo synthesis of enzymes involved in cell defense and repair (79). In particular, the anti-inflammatory and antioxidant features of CGA are well–recognized (80) (Figure 3).
In this context, the nuclear factor erythroid 2-related factor 2 (Nrf2) is an important cellular target for physical binding of coffee phytochemicals (81, 82). This complex is a cellular key regulator in association with AMPK and sirtuins. Melanoidins, CGAs, and their degradation products have been reported as robust activators of Nrf2 activity (82, 83) and this effect may contribute to the possible health effects of coffee (79) and its by-products. The liver appears to be an important site of coffee bioactivity, by improving fat oxidation and thus lowering the risk of steatosis. Furthermore, these phytochemicals are linked to the preservation of functional pancreatic β cell mass via improved mitochondrial function and a reduction in endoplasmic reticulum stress (Figure 3). Indeed, long-term preservation of liver and β cell functions may account for the association between regular coffee consumption and a lower risk of type 2 diabetes (79). So, coffee silverskin extract has an interesting bioactive cluster, emphasizing the potential of this by-product as an unconventional functional constituent for the food industry (21, 36).
The colonic microflora plays a crucial role upon the fate of CGA metabolites (caffeic and ferulic acids), by converting them into dihydroferulic acid, which can then be absorbed (84). On the other hand, melanoidins appear to induce metabolic effects like those of dietary fiber from cereals linked to polyphenols (modulation of digestive enzymes, intestinal microbiota and control of oxidative processes in the gut) (41). Also, they are metabolized by the intestinal microbiota, being used as carbon and nitrogen sources by the hindgut microflora, and exerting a prebiotic activity (Figure 3) through the modulation of bacterial colon population and supporting Bifidobacteria growth (85).
CS, Coffee silverskin; CGA, Chlorogenic acids; AMPK, 5' adenosine monophosphate-activated protein kinase; FFA, Free fatty acids; GLP1, Glucagon-like peptide-1; GLUTs, Glucose transporters; SGLT1, Sodium/glucose cotransporter 1; GIP, Glucose-dependent insulinotropic polypeptide; G-6Pase, Glucose-6-phosphate translocase; PI3K, Phosphatidylinositol 3-kinases; NRF2, Nuclear factor erythroid 2-related factor 2; AGEs, Advanced glycation end products; SIRT1, Sirtuin 1.
Additionally, melanoidins contribute to the prolongation of satiety, more specifically, their metabolites stimulate the secretion of GLP-1 by the enteroendocrine cells, causing a delay in the digestion of sugars (41). Interestingly enough, a recent study showed that the preventive and therapeutic effect of CGA in relation to obesity may be related to the regulation of gut microbiota (86). Indeed, the gut microbiota is the main driver of both intra- and inter-individual variations in metabolism of dietary bioactive small molecules (47). In this regard, it is important to note that research into melanoidins metabolism by gut microbiota is valuable because these biotransformed metabolites play an important role in the protection of colorectal cells from cancer by reducing inflammation and boosting tumor cell apoptosis. Moreover, data from an animal model study showed that caffeine consumption neutralizes the shift in the obesogenic (Firmicutes/Bacteroides) ratio in gut microbiota, resulting from a western diet (87). So, the ability of main bioactive compounds from coffee silverskin to modulate gut microbiota may play an important role in the prevention/treatment of MetS.
Coffee silverskin compounds seem to contribute to a potential protective effect against MetS through complementary mechanisms of action: (a) interaction with intestinal sugar transporters, hormones, and signaling pathways acting on glucose metabolism, particularly AMPK; (b) improvement of glucose uptake and insulin sensitivity; (c) lowering total cholesterol and triglyceride plasma levels, possibly through the inhibition of pancreatic lipase; (d) reduction of inflammatory cytokine production in adipose tissue; (e) protection against oxidative DNA damage; and (f) acting as interesting players in the regulation of gut microbiota.
By this way, coffee silverskin appears as a very interesting matrix to be used directly or to develop functional ingredients to enrich foods or to produce dietary supplements with potential beneficial effects against MetS. Simultaneously, the valorization of this by-product that, up to date, is being discarded by the coffee roasting industries, would also respond to the sustainability and circular economy needed in the coffee chain.
Lastly, it is essential to note that coffee silverskin extract is not a competitor to the coffee beverage and can be consumed as a supplement or used as a functional ingredient to enrich beverages and foods. Silverskin valorization has a sustainable footprint and a wealth of bioactive compounds as key factors and can be consumed by a wide range of people, regardless of whether they are coffee drinkers or dislike the taste of coffee.
Conceptualization, Research/Investigation: NA, JP, and RA. Writing–original draft preparation: NA and JP. Writing–review and editing, Funding acquisition: BO, FM, and RA. Visualization: NA and FM. Supervision: FM and RA. Project administration: RA and BO. All authors contributed to the article and approved the submitted version.
The open access publication fee of this paper was funded by AgriFood XXI I&D&I project (NORTE-01-0145-FEDER-000041) cofinanced by European Regional Development Fund (ERDF) through the NORTE 2020 (Programa Operacional Regional do Norte 2014/2020).
This work was supported by the projects PTDC/SAU-NUT/2165/2021 (COBY4HEALTH - Can coffee by-products decrease the risk of metabolic syndrome? A comprehensive approach to reduce waste and valorize health benefits) funded by FCT; UIDB/50006/2020, funded by FCT/MCTES (Portugal); SYSTEMIC “an integrated approach to the challenge of sustainable food systems: adaptive and mitigatory strategies to address climate change and malnutrition”, Knowledge hub on Nutrition and Food Security; and UID/BIM/04293/2013 funded by FCT (to Fátima Martel).
NA is grateful to the project PTDC/SAU-NUT/2165/2021-COBY4HEALTH for a postdoctoral grant. JP and RA are grateful to Fundação para a Ciência e a Tecnologia (FCT) for their PhD grant (SFRH/BD/07329/2021) and CEECIND/01120/2017 contract, respectively.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. International Coffee Organization. Trade Statistics. Available online at: https://www.ico.org/ (2022). (accessed March, 2022).
2. Alves RC, Rodrigues F, Antónia Nunes M, Vinha AF, Oliveira MBPP. “Chapter 1 - State of the Art in Coffee Processing by-Products”. In: Galanakis CM, editor. Handbook of Coffee Processing by-Products. Academic Press (2017). p. 1–26.
3. Samoggia A, Riedel B. Consumers' perceptions of coffee health benefits and motives for coffee consumption and purchasing. Nutrients. (2019) 11:653. doi: 10.3390/nu11030653
4. de Melo Pereira GV, de Carvalho Neto DP, Magalhaes Junior AI, do Prado FG, Pagnoncelli MGB, Karp SG, et al. Chemical composition and health properties of coffee and coffee by-products. Adv Food Nutr Res. (2020) 91:65–96. doi: 10.1016/bs.afnr.2019.10.002
5. Pasman WJ, Boessen R, Donner Y, Clabbers N, Boorsma A. Effect of caffeine on attention and alertness measured in a home-setting, using web-based cognition tests. JMIR Res Protoc. (2017) 6:e169. doi: 10.2196/resprot.6727
6. Jiang X, Zhang D, Jiang W. Coffee and caffeine intake and incidence of type 2 diabetes mellitus: a meta-analysis of prospective studies. Eur J Nutr. (2014) 53:25–38. doi: 10.1007/s00394-013-0603-x
7. Navarro AM, Abasheva D, Martinez-Gonzalez MA, Ruiz-Estigarribia L, Martin-Calvo N, Sanchez-Villegas A, et al. Coffee consumption and the risk of depression in a middle-aged cohort: the sun project. Nutrients. (2018) 10:1333. doi: 10.3390/nu10091333
8. Lucas M, O'Reilly EJ, Pan A, Mirzaei F, Willett WC, Okereke OI, et al. Coffee, caffeine, and risk of completed suicide: results from three prospective cohorts of american adults. World J Biol Psychiatry. (2014) 15:377–86. doi: 10.3109/15622975.2013.795243
9. Tran KT, Coleman HG, McMenamin ÚC, Cardwell CR. Coffee consumption by type and risk of digestive cancer: a large prospective cohort study. Br J Cancer. (2019) 120:1059–66. doi: 10.1038/s41416-019-0465-y
10. Heath RD, Brahmbhatt M, Tahan AC, Ibdah JA, Tahan V. Coffee: the magical bean for liver diseases. World J Hepatol. (2017) 9:689–96. doi: 10.4254/wjh.v9.i15.689
11. Dranoff JA. Coffee consumption and prevention of cirrhosis: in support of the caffeine hypothesis. Gene Expr. (2018) 18:1–3. doi: 10.3727/105221617X15046391179559
12. Herden L, Weissert R. The impact of coffee and caffeine on multiple sclerosis compared to other neurodegenerative diseases. Front Nutr. (2018) 5:133. doi: 10.3389/fnut.2018.00133
13. Crippa A, Discacciati A, Larsson SC, Wolk A, Orsini N. Coffee consumption and mortality from all causes, cardiovascular disease, and cancer: a dose-response meta-analysis. Am J Epidemiol. (2014) 180:763–75. doi: 10.1093/aje/kwu194
14. Gonzalez S, Salazar N, Ruiz-Saavedra S, Gomez-Martin M, de Los Reyes-Gavilan CG, Gueimonde M. Long-term coffee consumption is associated with fecal microbial composition in humans. Nutrients. (2020) 12:1287. doi: 10.3390/nu12051287
15. Klingel T, Kremer JI, Gottstein V, Rajcic de. Rezende T, Schwarz S, Lachenmeier DW. A review of coffee by-products including leaf, flower, cherry, husk, silver skin, and spent grounds as novel foods within the european union. Foods. (2020) 9:665. doi: 10.3390/foods9050665
16. Echeverria MC, Nuti M. Valorisation of the residues of coffee agro-industry: perspectives and limitations. Open Waste Manag J. (2017) 10:13–22. doi: 10.2174/1876400201710010013
17. Lachenmeier DW, Schwarz S, Rieke-Zapp J, Cantergiani E, Rawel H, Martin-Cabrejas MA, et al. Coffee by-products as sustainable novel foods: report of the 2nd international electronic conference on foods-future foods and food technologies for a sustainable world. Foods. (2021) 11:3. doi: 10.3390/foods11010003
18. Schanes K, Dobernig K, Gözet B. Food waste matters - a systematic review of household food waste practices and their policy implications. J Clean Prod. (2018) 182:978–91. doi: 10.1016/j.jclepro.2018.02.030
19. Blinová L, Sirotiak M, Pastierova A, Soldán M. Review: utilization of waste from coffee production. Res Papers Fac Mat Sci Technol Slovak Univ Technol. (2017) 25:101–91. doi: 10.1515/rput-2017-0011
20. Bertolino M, Barbosa-Pereira L, Ghirardello D, Botta C, Rolle L, Guglielmetti A, et al. Coffee silverskin as nutraceutical ingredient in yogurt: its effect on the functional properties and its bioaccessibility. J Sci Food Agric. (2019) 99:4267–4275. doi: 10.1002/jsfa.9659
21. Bessada S, Alves R, Oliveira M. Coffee silverskin: a review on potential cosmetic applications. Cosmetics. (2018) 5:5. doi: 10.3390/cosmetics5010005
22. Andrade N, Marques C, Andrade S, Silva C, Rodrigues I, Guardao L, et al. Effect of chrysin on changes in intestinal environment and microbiome induced by fructose-feeding in rats. Food Funct. (2019) 10:4566–76. doi: 10.1039/C9FO01142K
23. Lee EG, Choi JH, Kim KE, Kim JH. Effects of a walking program on self-management and risk factors of metabolic syndrome in older Korean adults. J Phys Ther Sci. (2014) 26:105–9. doi: 10.1589/jpts.26.105
24. Tsai YJ, Wu MP, Hsu YW. Emerging health problems among women: inactivity, obesity, and metabolic syndrome. Gynecol Minim Invasive Ther. (2014) 3:12–4. doi: 10.1016/j.gmit.2013.07.005
25. O'Neill S, O'Driscoll L. Metabolic syndrome: a closer look at the growing epidemic and its associated pathologies. Obes Rev. (2015) 16:1–12. doi: 10.1111/obr.12229
26. Boque N, Campion J. de la Iglesia R, de la Garza AL, Milagro FI, San Roman B, et al. Screening of polyphenolic plant extracts for anti-obesity properties in wistar rats. J Sci Food Agric. (2013) 93:1226–32. doi: 10.1002/jsfa.5884
27. Andrade N, Andrade S, Silva C, Rodrigues I, Guardao L, Guimaraes JT, et al. Chronic consumption of the dietary polyphenol chrysin attenuates metabolic disease in fructose-fed rats. Eur J Nutr. (2020) 59:151–65. doi: 10.1007/s00394-019-01895-9
28. Noce A, Di Lauro M, Di Daniele F, Pietroboni Zaitseva A, Marrone G, Borboni P, et al. Natural bioactive compounds useful in clinical management of metabolic syndrome. Nutrients. (2021) 13:630. doi: 10.3390/nu13020630
29. Costa ASG, Alves RC, Vinha AF, Costa E, Costa CSG, Nunes MA, et al. Nutritional, chemical and antioxidant/pro-oxidant profiles of silverskin, a coffee roasting by-product. Food Chem. (2018) 267:28–35. doi: 10.1016/j.foodchem.2017.03.106
30. Machado S, Costa ASG, Pimentel BF, Oliveira M, Alves RC. A study on the protein fraction of coffee silverskin: protein/non-protein nitrogen and free and total amino acid profiles. Food Chem. (2020) 326:126940. doi: 10.1016/j.foodchem.2020.126940
31. Bessada SMF, Alves RC, Costa ASG, Nunes MA, Oliveira M. Coffea canephora silverskin from different geographical origins: a comparative study. Sci Total Environ. (2018) 645:1021–8. doi: 10.1016/j.scitotenv.2018.07.201
32. Nzekoue FK, Borsetta G, Navarini L, Abouelenein D, Xiao J, Sagratini G, et al. Coffee silverskin: characterization of b-vitamins, macronutrients, minerals and phytosterols. Food Chem. (2022) 372:131188. doi: 10.1016/j.foodchem.2021.131188
33. Alves RC, Casal S, Oliveira B. Benefícios do café na saúde: mito ou realidade? Química Nova. (2009) 32:2169–80. doi: 10.1590/S0100-40422009000800031
34. Tajik N, Tajik M, Mack I, Enck P. The potential effects of chlorogenic acid, the main phenolic components in coffee, on health: a comprehensive review of the literature. Eur J Nutr. (2017) 56:2215–44. doi: 10.1007/s00394-017-1379-1
35. Puga H, Alves RC, Costa AS, Vinha AF, Oliveira MBPP. Multi-frequency multimode modulated technology as a clean, fast, and sustainable process to recover antioxidants from a coffee by-product. J Clean Prod. (2017) 168:14–21. doi: 10.1016/j.jclepro.2017.08.231
36. Tores de. la Cruz S, Iriondo-DeHond A, Herrera T, Lopez-Tofino Y, Galvez-Robleno C, Prodanov M, et al. An assessment of the bioactivity of coffee silverskin melanoidins. Foods. (2019) 8:68. doi: 10.3390/foods8020068
37. Langner E, Rzeski W. Biological properties of melanoidins: a review. Int J Food Prop. (2014) 17:344–53. doi: 10.1080/10942912.2011.631253
38. Rufian-Henares JA, Morales FJ. Effect of in vitro enzymatic digestion on antioxidant activity of coffee melanoidins and fractions. J Agric Food Chem. (2007) 55:10016–21. doi: 10.1021/jf0718291
39. Perez-Burillo S, Rajakaruna S, Pastoriza S, Paliy O, Angel Rufian-Henares J. Bioactivity of food melanoidins is mediated by gut microbiota. Food Chem. (2020) 316:126309. doi: 10.1016/j.foodchem.2020.126309
40. Kaczmarczyk-Sedlak I, Folwarczna J, Sedlak L, Zych M, Wojnar W, Szuminska I, et al. Effect of caffeine on biomarkers of oxidative stress in lenses of rats with streptozotocin-induced diabetes. Arch Med Sci. (2019) 15:1073–80. doi: 10.5114/aoms.2019.85461
41. Walker JM, Mennella I, Ferracane R, Tagliamonte S, Holik A-K, Hölz K, et al. Melanoidins from coffee and bread differently influence energy intake: a randomized controlled trial of food intake and gut-brain axis response. J Funct Foods. (2020) 72:104063. doi: 10.1016/j.jff.2020.104063
42. Shaheen S, Shorbagi M, Lorenzo JM, Farag MA. Dissecting dietary melanoidins: formation mechanisms, gut interactions and functional properties. Crit Rev Food Sci Nutr. (2021) 17:1–18. doi: 10.1080/10408398.2021.1937509
43. Shi X, Xue W, Liang S, Zhao J, Zhang X. Acute caffeine ingestion reduces insulin sensitivity in healthy subjects: a systematic review and meta-analysis. Nutr J. (2016) 15:103. doi: 10.1186/s12937-016-0220-7
44. Du WY, Xiao Y, Yao JJ, Hao Z, Zhao YB. Chlorogenic acid in the oxidative stress injury triggered by shuang-huang-lian injection. Exp Ther Med. (2018) 16:2901–8. doi: 10.3892/etm.2018.6567
45. Meng S, Cao J, Feng Q, Peng J, Hu Y. Roles of chlorogenic acid on regulating glucose and lipids metabolism: a review. Evid Based Complement Alternat Med. (2013) 2013:801457. doi: 10.1155/2013/801457
46. Nankar R, Prabhakar PK, Doble M. Hybrid drug combination: combination of ferulic acid and metformin as anti-diabetic therapy. Phytomedicine. (2017) 37:10–3. doi: 10.1016/j.phymed.2017.10.015
47. Kerimi A, Kraut NU, da Encarnacao JA, Williamson G. The gut microbiome drives inter- and intra-individual differences in metabolism of bioactive small molecules. Sci Rep. (2020) 10:19590. doi: 10.1038/s41598-020-76558-5
48. Farah A, de Paula LJ. Consumption of chlorogenic acids through coffee and health implications. Beverages. (2019) 5:11. doi: 10.3390/beverages5010011
49. de Paula J, Farah A. Caffeine consumption through coffee: content in the beverage, metabolism, health benefits and risks. Beverages. (2019) 5:37. doi: 10.3390/beverages5020037
50. Nikpayam O, Najafi M, Ghaffari S, Jafarabadi MA, Sohrab G, Roshanravan N. Effects of green coffee extract on fasting blood glucose, insulin concentration and homeostatic model assessment of insulin resistance (Homa-Ir): a systematic review and meta-analysis of interventional studies. Diabetol Metab Syndr. (2019) 11:91. doi: 10.1186/s13098-019-0489-8
51. Buscemi S, Marventano S, Antoci M, Cagnetti A, Castorina G, Galvano F, et al. Coffee and metabolic impairment: an updated review of epidemiological studies. NFS J. (2016) 3:1–7. doi: 10.1016/j.nfs.2016.02.001
52. Rajan ND, Koellner S, Calabrese VT, Khan A. Inactivation of α-Amylase by caffeine: reducing the break-down of starch into sugars. Am J BioSci. (2018) 6:1. doi: 10.11648/j.bio.20180601.11
53. Tunnicliffe J, Eller L, Reimer R, Hittel D, Shearer J. Chlorogenic acid differentially affects postprandial glucose and glucose-dependent insulinotropic polypeptide response in rats. Appl Physiol Nutr Metab. (2011) 36:650–9. doi: 10.1139/h11-072
54. Williamson G. Protection against developing type 2 diabetes by coffee consumption: assessment of the role of chlorogenic acid and metabolites on glycaemic responses. Food Funct. (2020) 11:4826–33. doi: 10.1039/D0FO01168A
55. Prabhakar PK, Doble M. Interaction of cinnamic acid derivatives with commercial hypoglycemic drugs on 2-deoxyglucose uptake in 3T3-L1 adipocytes. J Agric Food Chem. (2011) 59:9835–44. doi: 10.1021/jf2015717
56. Nzekoue FK, Angeloni S, Navarini L, Angeloni C, Freschi M, Hrelia S, et al. Coffee silverskin extracts: quantification of 30 bioactive compounds by a new Hplc-Ms/Ms method and evaluation of their antioxidant and antibacterial activities. Food Res Int. (2020) 133:109128. doi: 10.1016/j.foodres.2020.109128
57. Andrade N, Araújo JR, Correia-Branco A, Carletti JV, Martel F. Effect of dietary polyphenols on fructose uptake by human intestinal epithelial (Caco-2) cells. J Funct Foods. (2017) 36:429–39. doi: 10.1016/j.jff.2017.07.032
58. Koepsell H. Glucose transporters in the small intestine in health and disease. Pflugers Arch. (2020) 472:1207–48. doi: 10.1007/s00424-020-02439-5
59. Reis CEG, Dorea JG, da Costa THM. Effects of coffee consumption on glucose metabolism: a systematic review of clinical trials. J Tradit Complement Med. (2019) 9:184–91. doi: 10.1016/j.jtcme.2018.01.001
60. Gao J, He X, Ma Y, Zhao X, Hou X, Hao E, et al. Chlorogenic acid targeting of the Akt Ph domain activates Akt/Gsk3beta/Foxo1 signaling and improves glucose metabolism. Nutrients. (2018) 10:1366. doi: 10.3390/nu10101366
61. Jensen TE, Rose AJ, Hellsten Y, Wojtaszewski JF, Richter EA. Caffeine-induced Ca(2+) release increases Ampk-dependent glucose uptake in rodent soleus muscle. Am J Physiol Endocrinol Metab. (2007) 293:E286–92. doi: 10.1152/ajpendo.00693.2006
62. Park S, Scheffler TL, Gunawan AM, Shi H, Zeng C, Hannon KM, et al. Chronic elevated calcium blocks Ampk-induced glut-4 expression in skeletal muscle. Am J Physiol Cell Physiol. (2009) 296:C106–15. doi: 10.1152/ajpcell.00114.2008
63. da Silva LA, Wouk J, Weber VMR, Eltchechem CL, de Almeida P, Martins JCL, et al. Mechanisms and biological effects of caffeine on substrate metabolism homeostasis: a systematic review. J App Pharm Sci. (2017) 7:215–21. doi: 10.7324/JAPS.2017.70632
64. Akiba T, Yaguchi K, Tsutsumi K, Nishioka T, Koyama I, Nomura M, et al. Inhibitory mechanism of caffeine on insulin-stimulated glucose uptake in adipose cells. Biochem Pharmacol. (2004) 68:1929–37. doi: 10.1016/j.bcp.2004.07.036
65. del Castillo MD, Fernandez-Gomez B, Martinez-Saez N, Iriondo-DeHond A, Martirosyan D, Mesa M. “Coffee Silverskin Extract for Aging and Chronic Diseases”. In: Martirosyan DM, editor. Functional Foods for Chronic Diseases. Scotts Valley: CreateSpace. (2016). p. 386–409.
66. Rebollo-Hernanz M, Zhang Q, Aguilera Y, Martin-Cabrejas MA, Gonzalez de. Mejia E. Phenolic compounds from coffee by-products modulate adipogenesis-related inflammation, mitochondrial dysfunction, and insulin resistance in adipocytes, via Insulin/Pi3k/Akt signaling pathways. Food Chem Toxicol. (2019) 132:110672. doi: 10.1016/j.fct.2019.110672
67. Rebollo-Hernanz M, Aguilera Y, Martín-Cabrejas MA, Gonzalez de. Mejia E. Activating effects of the bioactive compounds from coffee by-products on Fgf21 signaling modulate hepatic mitochondrial bioenergetics and energy metabolism in vitro. Front Nutr. (2022) 9:866233. doi: 10.3389/fnut.2022.866233
68. Franssen R, Monajemi H, Stroes ES, Kastelein JJ. Obesity and Dyslipidemia. Med Clin North Am. (2011) 95:893–902. doi: 10.1016/j.mcna.2011.06.003
69. Martinez-Saez N, Ullate M, Martin-Cabrejas MA, Martorell P, Genoves S, Ramon D, et al. A novel antioxidant beverage for body weight control based on coffee silverskin. Food Chem. (2014) 150:227–34. doi: 10.1016/j.foodchem.2013.10.100
70. Farias-Pereira R, Park CS, Park Y. Mechanisms of action of coffee bioactive components on lipid metabolism. Food Sci Biotechnol. (2019) 28:1287–96. doi: 10.1007/s10068-019-00662-0
71. Tabrizi R, Saneei P, Lankarani KB, Akbari M, Kolahdooz F, Esmaillzadeh A, et al. The effects of caffeine intake on weight loss: a systematic review and dos-response meta-analysis of randomized controlled trials. Crit Rev Food Sci Nutr. (2019) 59:2688–96. doi: 10.1080/10408398.2018.1507996
72. Prabhakar PK, Prasad R, Ali S, Doble M. Synergistic interaction of ferulic acid with commercial hypoglycemic drugs in streptozotocin induced diabetic rats. Phytomedicine. (2013) 20:488–94. doi: 10.1016/j.phymed.2012.12.004
73. Lee C. Antioxidant ability of caffeine and its metabolites based on the study of oxygen radical absorbing capacity and inhibition of Ldl peroxidation. Clin Chim Acta. (2000) 295:141–54. doi: 10.1016/S0009-8981(00)00201-1
74. Fernandez-Gomez B, Lezama A, Amigo-Benavent M, Ullate M, Herrero M, Martín MA, et al. Insights on the health benefits of the bioactive compounds of coffee silverskin extract. J Funct Foods. (2016) 25:197–207. doi: 10.1016/j.jff.2016.06.001
75. Roy S, Metya SK, Sannigrahi S, Rahaman N, Ahmed F. Treatment with ferulic acid to rats with streptozotocin-induced diabetes: effects on oxidative stress, pro-inflammatory cytokines, and apoptosis in the pancreatic β cell. Endocrine. (2013) 44:369–79. doi: 10.1007/s12020-012-9868-8
76. Iriondo-DeHond A, Haza AI, Avalos A. del Castillo MD, Morales P. Validation of coffee silverskin extract as a food ingredient by the analysis of cytotoxicity and genotoxicity. Food Res Int. (2017) 100:791–7. doi: 10.1016/j.foodres.2017.08.012
77. Verzelloni E, Pellacani C, Tagliazucchi D, Tagliaferri S, Calani L, Costa LG, et al. Antiglycative and neuroprotective activity of colon-derived polyphenol catabolites. Mol Nutr Food Res. (2011) 55 (Suppl. 1):S35–43. doi: 10.1002/mnfr.201000525
78. Mesías M, Navarro M, Martínez-Saez N, Ullate M, del Castillo MD, Morales FJ. Antiglycative and carbonyl trapping properties of the water soluble fraction of coffee silverskin. Food Res Int. (2014) 62:1120–6. doi: 10.1016/j.foodres.2014.05.058
79. Kolb H, Martin S, Kempf K. Coffee and lower risk of type 2 diabetes: arguments for a causal relationship. Nutrients. (2021) 13:1144. doi: 10.3390/nu13041144
80. Liang N, Kitts DD. Role of chlorogenic acids in controlling oxidative and inflammatory stress conditions. Nutrients. (2015) 8:16. doi: 10.3390/nu8010016
81. Liang N, Dupuis JH, Yada RY, Kitts DD. Chlorogenic acid isomers directly interact with keap 1-Nrf2 signaling in Caco-2 cells. Mol Cell Biochem. (2019) 457:105–18. doi: 10.1007/s11010-019-03516-9
82. Sauer T, Raithel M, Kressel J, Munch G, Pischetsrieder M. Activation of the transcription factor Nrf2 in macrophages, Caco-2 cells and intact human gut tissue by maillard reaction products and coffee. Amino Acids. (2013) 44:1427–39. doi: 10.1007/s00726-012-1222-1
83. Fratantonio D, Speciale A, Canali R, Natarelli L, Ferrari D, Saija A, et al. Low nanomolar caffeic acid attenuates high glucose-induced endothelial dysfunction in primary human umbilical-vein endothelial cells by affecting Nf-Kappab and Nrf2 Pathways. Biofactors. (2017) 43:54–62. doi: 10.1002/biof.1312
84. Naveed M, Hejazi V, Abbas M, Kamboh AA, Khan GJ, Shumzaid M, et al. Chlorogenic acid (Cga): a pharmacological review and call for further research. Biomed Pharmacother. (2018) 97:67–74. doi: 10.1016/j.biopha.2017.10.064
85. Jiménez-Zamora A, Pastoriza S, Rufián-Henares JA. Revalorization of coffee by-products. Prebiotic, antimicrobial and antioxidant properties LWT. Food Sci Technol. (2015) 61:12–8. doi: 10.1016/j.lwt.2014.11.031
86. Wang Z, Lam KL, Hu J, Ge S, Zhou A, Zheng B, et al. Chlorogenic acid alleviates obesity and modulates gut microbiota in high-fat-fed mice. Food Sci Nutr. (2019) 7:579–88. doi: 10.1002/fsn3.868
Keywords: coffee by-products, caffeine, chlorogenic acids, melanoidins, metabolic syndrome
Citation: Andrade N, Peixoto JAB, Oliveira MBPP, Martel F and Alves RC (2022) Can coffee silverskin be a useful tool to fight metabolic syndrome? Front. Nutr. 9:966734. doi: 10.3389/fnut.2022.966734
Received: 11 June 2022; Accepted: 02 September 2022;
Published: 21 September 2022.
Edited by:
Lidia Montero, University of Duisburg-Essen, GermanyReviewed by:
Iziar Amaia Ludwig, University of Navarra, SpainCopyright © 2022 Andrade, Peixoto, Oliveira, Martel and Alves. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nelson Andrade, bmVsc29uYW5kcmFkZUBvdXRsb29rLmNvbQ==; Rita C. Alves, cmNhbHZlc0BmZi51cC5wdA==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.