
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Nutr. , 25 July 2022
Sec. Nutrition and Sustainable Diets
Volume 9 - 2022 | https://doi.org/10.3389/fnut.2022.954170
 Kenya E. Fernandes1
Kenya E. Fernandes1 Elizabeth A. Frost2,3
Elizabeth A. Frost2,3 Emily J. Remnant1
Emily J. Remnant1 Kathleen R. Schell4
Kathleen R. Schell4 Nural N. Cokcetin4
Nural N. Cokcetin4 Dee A. Carter1,5*
Dee A. Carter1,5*Honey is the source of energy for the European honey bee, Apis mellifera. Beyond simple nutrition and a hedge against the seasonal, geographic, and chemical unpredictability of nectar, honey has properties that protect the hive against various stresses. Enzyme-mediated detoxification during honey ripening neutralizes potentially toxic phytochemicals, and bees that consume honey have enhanced tolerance to other ingested toxins. Catalase and antioxidant phenolics protect honey bees from oxidative damage caused by reactive oxygen species, promoting their longevity. Phytochemical components of honey and microRNAs have the potential to influence developmental pathways, with diet playing a large role in honey bee caste determination. Components of honey mediate stress response and promote cold tolerance during overwintering. Honey has a suite of antimicrobial mechanisms including osmotic pressure, low water activity, low pH, hydrogen peroxide, and plant-, honey bee-, and microbiota-derived compounds such as phytochemicals and antimicrobial peptides. Certain types of honey, particularly polyfloral honeys, have been shown to inhibit important honey bee pathogens including the bacteria responsible for American and European Foulbrood, the microsporidian Nosema ceranae, and the fungi responsible for Stonebrood. Understanding the diverse functional properties of honey has far-ranging implications for honey bee and hive health and management by beekeepers.
The European honey bee, Apis mellifera, possesses an extraordinary ability to process and store nectar in the form of honey. In the context of human health, honey is a highly valued natural product that has been used as a food and to treat various ailments since ancient times (1). Over recent decades, the specific nutritional and therapeutic effects of honey have been the focus of much research interest. Various constituents of honey have been shown to have a suite of therapeutically beneficial properties including antimicrobial, antioxidant, anti-inflammatory, and in vivo anticancer effects (2). Although there has been less research into the ecological role of honey, it has clearly evolved these functional properties to protect itself and the hive from microbial attack and spoilage, and to enhance honey bee health. In addition to producing honey, honey bees play a crucial role in agriculture, providing pollination services for a wide variety of crops globally (3).
Major challenges to global honey bee populations over recent decades including threats ranging from pesticides and pathogens to global warming and habitat transformation (4), and the imperative to reduce the use of chemical and antimicrobial agents to treat hive diseases, makes understanding the diverse functions of honey in the hive more important than ever. With our understanding of the complex actions of components of honey continually growing [reviewed by (5)], and the contribution of various colony products to honey bee health being increasingly recognized [reviewed by (6)], the fact that honey is a multi-functional food for honey bees, impacting far more than nutrition alone, has become clear [reviewed by (7)]. Here, we summarize research on the features and components of honey with a focus on its functional and ecological significance to honey bees and the hive (Table 1), including the various antimicrobial mechanisms and studies that have assayed honey against hive pathogens.
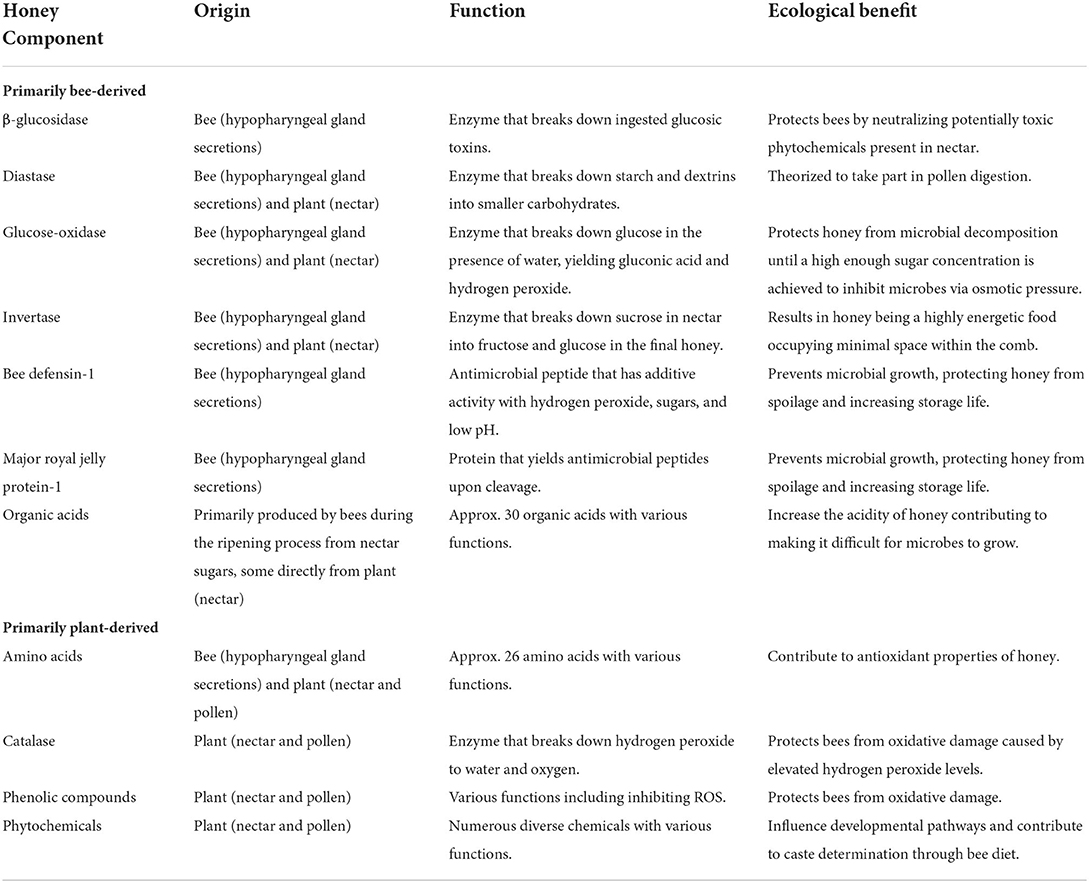
Table 1. Ubiquitous honey components and their ecological benefit to the hive.
Nectar is the main source of energy for the hive in the form of carbohydrates and is primarily composed of sugars with varying levels of moisture, mineral content, enzymes, and phytochemicals depending on the floral source (5). Once transported back to the hive by foragers, nectar is deposited into the wax cells of the honeycomb where the physicochemical transformation into honey, known as ripening, takes place (6). During this process, worker bees continue to manipulate the developing honey by secreting various enzymes from their hypopharyngeal glands into the nectar, and these can metabolize its components. The most important is invertase, which converts sucrose into fructose and glucose creating a stable, high density, and highly energetic food source that occupies a minimum amount of space within the wax cells of the hive (7). Other enzymes include protease, which breaks down proteins and polypeptides to yield smaller peptides that may influence the quality and nutritional value of honey (8), and diastase, which breaks down starch and dextrins into smaller carbohydrates and is thought to play a role in the digestion of pollen, the main source of protein for honey bees (9).
While this biochemical processing is taking place, the moisture content of the nectar is also being minimized through active and passive evaporation, making it resilient to microbial spoilage and prolonging its storage life. Active evaporation behavior by worker bees includes increasing the surface area of the nectar by sucking up, regurgitating, and holding it between their mandibles (6), and wing-fanning to increase circulation throughout the hive (10), which is done until the water content reaches ~50–60%. It is then placed into wax cells and relocated periodically, evaporating passively until a final water content of ~13–25% is reached (9, 11). The ripening process can take between 1 and 11 days to complete depending on factors such as colony size, climatic conditions, and botanical origin of the nectar (12). Once the honey is ripe, it is capped off with a thin wax layer for long-term storage to protect the hive from nectar shortages when foraging is not possible due to unfavorable seasonal and climatic conditions. In the absence of forage, honey bee colonies would collapse within a few days without an adequate store of honey (13).
Beyond providing food for the colony to maintain basic physiological functioning, the production of honey has additional nutritional benefits. The properties, both beneficial and detrimental, of foraged plant-derived compounds can vary significantly depending on the floral and geographic source. While an individual honey bee may forage from only one or a few floral sources, at the level of the colony and over the course of a season, a wide range of nectars can be collected, processed, and stored as honey, providing a more suitable and consistent diet across the colony (14). Additionally, honey bees have long been known to exhibit preferences for certain food sources over others including the specific position of one flower over another (15), and to adjust their choices amongst more and less profitable nectar sources based on the nutritional needs of the colony (16, 17). Foragers will also avoid nectar containing particular bacterial communities (18), and in laboratory studies will selectively choose among various types of honey depending on their health status (19). The capacity to store various products in the hive, including honey, resins, propolis, pollen, bee bread, and beeswax, is thought to allow honey bees to adapt their food source and behavior based on the health status of individuals or the colony in order to manage nutritional and health challenges (14).
While foraged pollen and nectar offer a diverse array of beneficial chemicals, they can also contain considerable amounts of toxic phytochemicals such as some alkaloids, coumarins, flavanols, and saponins (20). Honey bees can also be exposed to toxic pesticides though nectar, with some extremely persistent in the environment; for example neonicotinoid insecticides can be taken up by plants from environmental reservoirs in water and soil for years after their application (21). The processing of nectar into honey assists detoxification both directly and indirectly. Directly, the enzyme β-glucosidase is added by secretions from the honey bee hypopharyngeal gland and breaks down glycosidic toxins (9). Indirectly, the variety of floral resources collected by the colony allows them to be mixed and diluted thereby reducing the potential toxicity from any one nectar source (22). Ripening honey is also exposed to temperatures in the hive maintained around 35°C, which has been shown to decrease phenolic content including harmful phenolics present in some toxic nectars (23). Maintaining honey at this temperature is thought to similarly deactivate other compounds including potential toxins, reducing the concentration that must then be cleared via further enzymatic processing by the honey bee (22) but likely also reducing potentially beneficial compounds.
Honey consumption has been demonstrated to enhance honey bee tolerance to ingested natural and synthetic toxins. Specific components of honey that have been identified to play a role in the detoxification process include the flavonoid quercetin, the phenolic acid p-coumaric acid, and the plant hormone abscisic acid. RNA-sequencing analysis revealed that both low and high levels of quercetin in larval diets induced upregulation of multiple cytochrome P450 monooxygenases, one of the major enzyme subfamilies responsible for the metabolism of toxins in honey bees (24). Diet supplementation with p-coumaric acid increased metabolism of the acaricide coumaphos (25), and increased the survival of honey bees exposed to the insecticide tau-fluvalinate (26). Supplementation with abscisic acid increased the tolerance of honey bees to the pesticide carvacrol, and the acaricide oxalic acid (27). Various other phenolic acids and flavonoids present in honey including flavones, flavanones, and flavanols have also been shown to induce detoxification-related genes (25).
The accumulation of oxidative damage to lipids, proteins, and nucleic acids caused by reactive oxygen species (ROS) has been linked to aging and death in various organisms including the honey bee. Honey bee senescence accelerates with the transition from in-hive tasks to foraging behavior (28), presumed to be due to an increased metabolic rate to meet the requirements of hovering flight resulting in elevated production of several ROS including superoxide anions and hydroxyl radicals (29). Components of honey that exhibit antioxidant activity are thought to enhance the longevity of the honey bee by neutralizing ROS. The enzyme catalase, originating from pollen and nectar, metabolizes hydrogen peroxide to produce water and molecular oxygen. This protects honey bees from oxidative damage caused by high levels of hydrogen peroxide and works in combination with other antioxidant enzymes expressed in the honey bee gut such as superoxide dismutase and peroxidases (30). Phenolic compounds, peptides, vitamins, organic acids, and trace elements have also been identified as contributing to the antioxidant capacity of honey (31), which can vary significantly with floral source (32).
Honey is thought to play a role in various other ecological functions including developmental regulation, immune system enhancement, coordination of the stress response, and supporting the honey bee gut microbiome. As diet plays a large role in honey bee caste determination, phytochemical components of honey have the potential to influence developmental pathways. While queen-destined larvae are fed only royal jelly, worker-destined larvae consume a diet that includes honey along with royal jelly and beebread (33). Although present in greater quantities in pollen, components of honey including p-coumaric acid and plant microRNAs have been shown to reduce ovary development and promote differentiation into workers (33, 34). The honey bee humoral immune system involves the production and secretion of antimicrobial peptides that are active against various pathogens including bacteria, fungi, viruses, and parasites (35). Diverse phytochemicals found in honey upregulate various antimicrobial peptides including hypenoptaecin (36), and apidaecin (33). Consumption of abscisic acid, a phytohormone commonly present in honey, has been implicated in coordinating honey bee stress response including stimulating innate immune defenses such as wound healing and phagocytosis activation (11), and inducing cold tolerance during overwintering (27). The community of microbes that live in the honey bee gut, referred to as the gut microbiome, play a role in various aspects of host health including contributing to digestion and nutrient intake, and out-competing harmful microbes (37). It is likely that consumption of honey supports the health of the honey bee gut microbiome, and studies investigating honey in the context of human gut health have shown that honey oligosaccharides can promote the growth of beneficial lactobacilli and bifidobacteria (38), which are also members of the honey bee gut (39).
Honey is a supersaturated sugar solution, rendering it unfavorable for microbial growth via two mechanisms. The first is a low water content that results in high osmotic pressure, drawing water out of microorganisms and inhibiting their growth. The exact water content of honey can be influenced by various factors including the floral origin of the nectar and seasonal and climatic conditions, but normally ranges from between 13 and 25%, with around 17% being optimal as above this honey is vulnerable to fermentation by osmophilic yeasts (9). The second mechanism is low water activity resulting from the high concentration of sugar, which makes water unavailable for microorganisms to utilize for growth. The water activity of honey typically ranges from 0.5 to 0.65, which is lower than the amount needed for most bacteria (0.90), yeasts (0.80), and molds (0.70) to grow (9, 37). A water activity below 0.61 will also inhibit the growth of osmophilic yeasts (38).
Immature honey that has not yet reached a sufficient sugar concentration, and honey that has been diluted after processing, however, would still be vulnerable to microbial degradation without additional antimicrobial mechanisms. Nurse bees generally dilute honey with water before feeding it to larvae and adults due to its high viscosity (11) and ensuring protection of this diluted honey is essential. Glucose oxidase (GOX) is an enzyme that is added to nectar from the hypopharyngeal glands of honey bees during processing (39) and converts glucose to gluconolactone in diluted honey, in turn yielding gluconic acid and hydrogen peroxide (40). GOX is highly expressed in worker bees across developmental stages reaching its highest level in nectar processors and foragers (41). Gluconic acid discourages microbial overgrowth by lowering the pH of honey. Over thirty other organic acids have been identified in honey including acetic, citric, formic, lactic, and malic, which contribute to keeping the pH between 3.2 and 4.5, however gluconic acid is predominant representing 70–90% of all honey acids (9, 42, 43). The hydrogen peroxide produced by GOX activity destroys microbes via free hydroxyl radical production, which causes oxidation of microbial lipids, proteins, and nucleic acids (44). Maximum production of hydrogen peroxide is reached at 30–50% dilution, depending on the variety of honey (45, 46).
As an incredibly complex mix comprised of up to 200 different substances (47), it is no surprise that components of honey responsible for antimicrobial effects are increasingly being identified. These components originate from foraged plant sources, endogenous honey bee secretions, and associated microbiota, and are often specific to particular varieties of honey. One of the most well-known plant derived components is methylglyoxal (MGO), identified as the main antibacterial component of manuka honey produced from the nectar of certain Leptospermum species native to New Zealand and Australia (48). MGO is thought to crosslink proteins and DNA, damage cell membranes, and alter the structure of bacterial fimbriae and flagella, limiting their adherence and motility (49). Among other plant-derived components associated with antimicrobial activity are secondary metabolites such as polyphenols, flavonoids, volatile compounds, and alkaloids present in nectar (9). These diverse phytochemicals have evolved as innate plant protection systems against stresses including microbial infection and degradation (50) and thus likely perform similar ecological functions in honey. Many of these components are found at concentrations that are not sufficient to account for significant antimicrobial activity however, and are thought to work synergistically with other honey components including hydrogen peroxide (51).
Among honey bee-derived antimicrobial components are the antimicrobial peptide bee defensin-1, and major royal jelly protein-1. Bee defensin-1, also known as royalisin, is found in honey bee haemolymph as well as the hypopharyngeal gland, from where it is introduced to honey during processing (51). It is active against both gram-positive and gram-negative bacteria and is also implicated in the ability of honey to damage biofilms (52, 53). Although the mechanism of action has not yet been confirmed, bee defensin-1 is thought to act in a similar way to defensins from other species by disrupting the bacterial cell membrane, resulting in the inhibition of DNA, RNA, and protein synthesis (47). Major royal jelly protein-1 (MRJP1) is the most abundant protein found in both royal jelly and honey, and it is also introduced from the hypopharyngeal gland during nectar processing (54). Upon cleavage, MRJP1 yields the antimicrobial peptides jellein-1, jellien-2, and jellein-4, which have been shown to cause cell wall lysis and death in bacteria (55, 56). Finally, recent work suggests that honey can be a reservoir of microbe-produced antimicrobial compounds including antimicrobial peptides, surfactants, and proteolytic and cell wall-degrading enzymes that are produced by antagonistic microbial interactions in plant nectars, the honey bee gut, and the honey itself (57). The compounds produced by these microbes may account for the target-specific antimicrobial effects observed with honey that cannot be due to the non-specific broad-spectrum action caused by the principal antimicrobial mechanisms of honey. Antimicrobial peptides produced by the bacteria Lactobacillus kunkeei (58) and Bacillus subtilis (59) isolated from honey samples have shown activity against human, hive, and food spoilage pathogens including bacteria and fungi.
While the antimicrobial activity of honey against human pathogens has been extremely well-documented, there are far fewer studies that have assayed honey against ecologically relevant hive pathogens. American Foulbrood (AFB) is a highly contagious disease capable of wiping out the entire hive, and is acquired when larvae ingest spores of the bacterium Paenibacillus larvae that then germinate in the gut, causing infected larvae to die in the brood cell (60). The endospores produced by P. larvae are highly resilient and capable of surviving for many years, and infected hives and equipment often need to be burned to ensure that they are destroyed. Monofloral black locust, canola, citrus, clover, cotton, heather, honeydew, linden, and sunflower honey and polyfloral Romanian honey have been shown to inhibit the growth of P. larvae vegetative cells at varying concentrations (61–64) (Table 2). The antimicrobial peptide bee defensin-1 has also been identified as a specific component of honey with activity against P. larvae (86). European Foulbrood (EFB) is another common bacterial disease that is acquired when larvae ingest food contaminated with the bacterium Melissococcus plutonius which begins reproducing in the midgut, deriving nutrients from the larvae and causing starvation (60). Secondary invading bacteria are often present in weakened colonies suffering from EFB including Bacillus pumilus, Brevibacillus laterosporus, Enterococcus faecalis, and Paenibacillus alvei (67). Antimicrobial activity against EFB and/or its secondary invaders has been reported for monofloral almond, eucalyptus, manuka, and orange honey and polyfloral Argentinean, Australian, Brazilian, Cuban, Romanian, Spanish, and Turkish honey (63, 68–73, 87, 88) (Table 2).
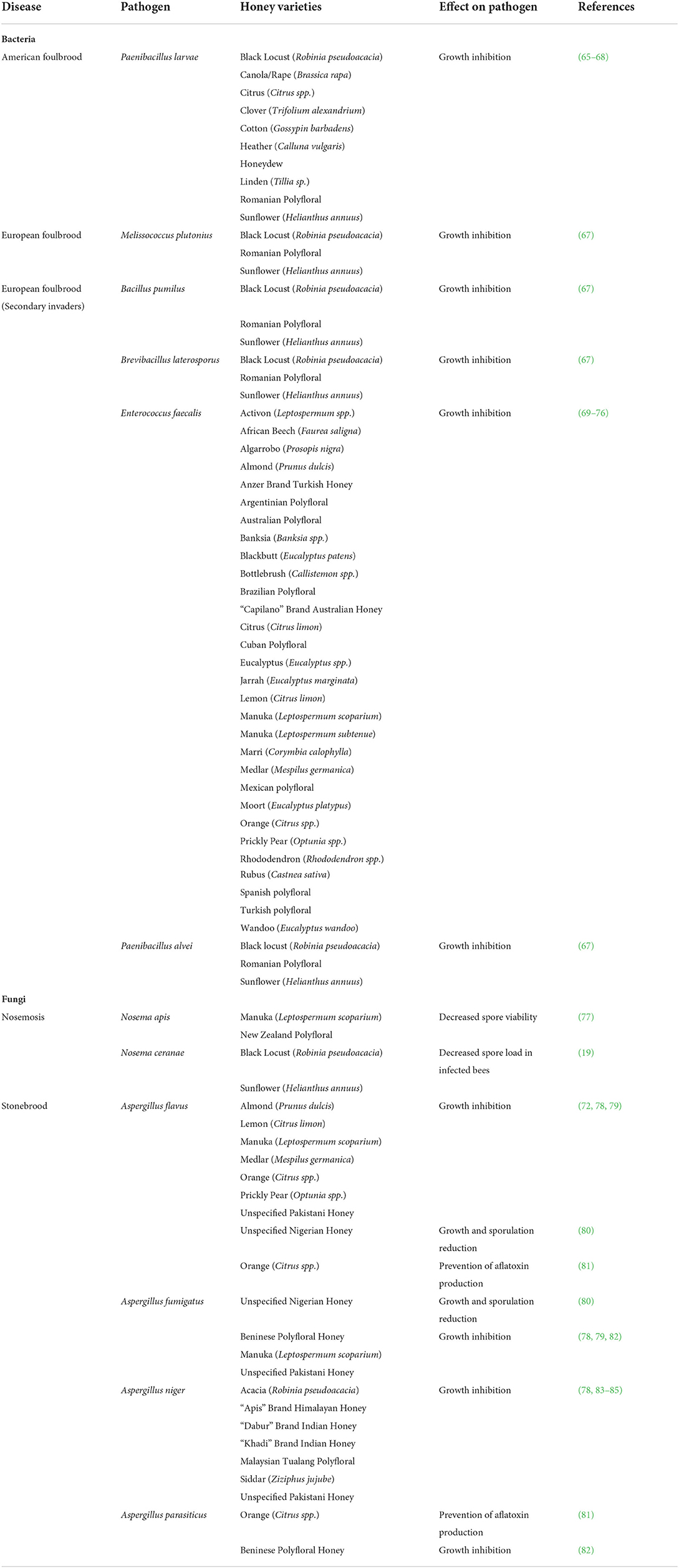
Table 2. Honey varieties with antimicrobial activity against hive pathogens.
Nosemosis is the most common disease of adult honey bees and is acquired when spores of the microsporidian fungi Nosema ceranae or Nosema apis are ingested. The spores then germinate in the midgut causing difficulty with digestion, increasing susceptibility to viral infection and weakening the honey bees, leading to reductions in lifespan, colony health, and colony performance (60, 74). A study testing the viability of N. apis spores found that it significantly decreased after only 3 days in the presence of New Zealand polyfloral honey compared to 1 month in manuka monofloral honey and 21 days in sugar syrup (75), while another study found infected honey bees ingesting sunflower honey had significantly decreased spore loads of N. ceranae compared to honey bees ingesting honeydew honey (19) (Table 2). Stonebrood is a less common fungal infection acquired when larvae ingest spores of Aspergillus species. Stonebrood can be fatal when mycotoxins produced by the fungi kill and mummify the larvae before they hatch (60). While no studies have determined whether honey is capable of killing Aspergillus spores, monofloral acacia, almond, lemon, manuka, medlar, orange, prickly pear, and siddar honey and polyfloral Beninese and Malaysian honey have been shown to inhibit the growth of various Aspergillus species including A. flavus, A. fumigatus, A. niger, and A. parasiticus (69, 76–79, 83, 89) (Table 2). Nigerian honey of unspecified botanical origin has been observed to reduce growth and sporulation in A. flavus and A. fumigatus (84), and orange honey has been observed to prevent aflatoxin production by A. flavus and A. parasiticus (85). This demonstrates the impressive ability of honey to protect the hive even from a ubiquitous environmental fungus that is capable of growth at low water activity and elevated temperatures.
Honey plays a fundamental role in maintaining colony health. Beyond simple nutrition, honey consumption has far-ranging impacts on toxin tolerance, honey bee longevity, developmental regulation, immunity, stress response, and resilience to pathogens. Beekeeping practices that substitute sugar solutions for honey may provide the hive with general nutrition, but this could have consequences on numerous aspects of honey bee and hive health. There is still a great deal we do not fully understand about how and why honey bees create and utilize honey. This includes by what mechanisms honey bees alter their foraging behaviors based on colony needs, how the microbiota of foraged plant components and the honey bee gut contribute to the functional properties of honey, whether and how various phytochemicals, endogenous secretions, and other components of honey work synergistically to exert their full effects, and how diverse foraging sources impact bee and hive health. Answering these questions could go a long way toward ensuring that honey bees and beekeepers alike prosper for decades to come.
This article was written by KF. Reviewed and edited by EF, ER, KS, NC, and DC. All authors contributed to the article and approved the submitted version.
Honey research projects undertaken by our team were supported by the NSW Bushfire Industry Recovery Package Sector Development Grant (BIP-SDG-135).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Eteraf-Oskouei T, Najafi M. Traditional and modern uses of natural honey in human diseases: a review. Iran J Basic Med Sci. (2012) 16:731–42.
2. Samarghandian S, Farkhondeh T, Samini F. Honey and health: a review of recent clinical research. Pharmacogn Res. (2017) 9:121–7. doi: 10.4103/0974-8490.204647
3. Hung KLJ, Kingston JM, Albrecht M, Holway DA, Kohn JR. The worldwide importance of honey bees as pollinators in natural habitats. Proc Royal Soc B. (2018) 285:20172140. doi: 10.1098/rspb.2017.2140
4. Potts SG, Imperatriz-Fonseca VL, Ngo HT, Biesmeijer JC, Breeze TD, Dicks LV, et al. The Assessment Report on Pollinators, Pollination and Food Production: Summary for Policymakers. Secretariat of the Intergovernmental Science-Policy Platform on Biodiversity an Ecosystem Services (2016).
5. Machado De-Melo AA, Almeida-Muradian LB de, Sancho MT, Pascual-Maté A. Composition and properties of Apis mellifera honey: a review. J Apic Res. (2018) 57:5–37. doi: 10.1080/00218839.2017.1338444
6. Erler S, Moritz RFA. Pharmacophagy and pharmacophory: mechanisms of self-medication and disease prevention in the honeybee colony (Apis mellifera). Apidologie. (2016) 47:389–411. doi: 10.1007/s13592-015-0400-z
7. Berenbaum MR, Calla B. Honey as a functional food for Apis mellifera. Annu Rev Entomol. (2020) 66:185–208. doi: 10.1146/annurev-ento-040320-074933
8. Santos-Buelga C, Gonzalez-Paramas AM. Chemical composition of honey. In: Alvarez-Suarez JM, editor. Bee Products—Chemical and Biological Properties. Cham: Springer International Publishing AG (2017). p. 43–82.
9. Wright GA, Nicolson SW, Shafir S. Nutritional physiology and ecology of honey bees. Annu Rev Entomol. (2018) 63:327–44. doi: 10.1146/annurev-ento-020117-043423
10. White JW, Doner LW. Honey composition and properties. In: Beekeeping in the United States Agriculture Handbook (1980). p. 82–91.
11. Rossano R, Larocca M, Polito T, Perna AM, Padula MC, Martelli G, et al. What are the proteolytic enzymes of honey and what they do tell us? A fingerprint analysis by 2-D zymography of unifloral honeys. PLoS ONE. (2012) 7:e49164. doi: 10.1371/journal.pone.0049164
12. Southwick EE, Moritz RFA. Social control of air ventilation in colonies of honey bees, Apis mellifera. J Insect Physiol. (1987) 33:623–6. doi: 10.1016/0022-1910(87)90130-2
13. Eyer M, Neumann P, Dietemann V. A look into the cell: honey storage in honey bees, Apis mellifera. PLoS ONE. (2016) 11:e0161059. doi: 10.1371/journal.pone.0161059
14. Somerville D. Fat Bees Skinny Bees: A Manual on Honey Bee Nutrition for Beekeepers. Rural Industries Research and Development Corporation (2005).
15. Abou-Shaara HF. The foraging behaviour of honey bees, Apis mellifera: a review. Vet Med. (2014) 59:1–10. doi: 10.17221/7240-VETMED
16. Seeley TD. Social foraging in honey bees: how nectar foragers assess their colony's nutritional status. Behav Ecol Sociobiol. (1989) 24:181–99. doi: 10.1007/BF00292101
17. Toth AL, Kantarovich S, Meisel AF, Robinson GE. Nutritional status influences socially regulated foraging ontogeny in honey bees. J Exp Biol. (2005) 208:4641–9. doi: 10.1242/jeb.01956
18. Good AP, Gauthier MPL, Vannette RL, Fukami T. Honey bees avoid nectar colonized by three bacterial species, but not by a yeast species, isolated from the bee gut. PLoS ONE. (2014) 9:e86494. doi: 10.1371/journal.pone.0086494
19. Gherman BI, Denner A, Bobiş O, Dezmirean DS, Mărghitaş LA, Schlüns H, et al. Pathogen-associated self-medication behavior in the honeybee Apis mellifera. Behav Ecol Sociobiol. (2014) 68:1777–84. doi: 10.1007/s00265-014-1786-8
20. Bernklau E, Bjostad L, Hogeboom A, Carlisle A, Arathi HS. Dietary phytochemicals, honey bee longevity and pathogen tolerance. Insects. (2019) 10:14. doi: 10.3390/insects10010014
21. Tosi S, Nieh JC, Sgolastra F, Cabbri R, Medrzycki P. Neonicotinoid pesticides and nutritional stress synergistically reduce survival in honey bees. Proc Royal Soc B. (2017) 284:20171711. doi: 10.1098/rspb.2017.1711
22. Berenbaum MR, Johnson RM. Xenobiotic detoxification pathways in honey bees. Curr Opin Insect Sci. (2015) 10:51–8. doi: 10.1016/j.cois.2015.03.005
23. Liu F, He J, Fu W. Highly controlled nest homeostasis of honey bees helps deactivate phenolics in nectar. Sci Nat. (2005) 92:297–9. doi: 10.1007/s00114-005-0629-x
24. Mao W, Schuler MA, Berenbaum MR. Disruption of quercetin metabolism by fungicide affects energy production in honey bees (Apis mellifera). Proc Nat Acad Sci USA. (2017) 114:2538–43. doi: 10.1073/pnas.1614864114
25. Mao W, Schuler MA, Berenbaum MR. Honey constituents up-regulate detoxification and immunity genes in the western honey bee Apis mellifera. Proc Natl Acad Sci USA. (2013) 110:8842–6. doi: 10.1073/pnas.1303884110
26. Mitton GA, Szawarski N, Mitton FM, Iglesias A, Eguaras MJ, Ruffinengo SR, et al. Impacts of dietary supplementation with p-coumaric acid and indole-3-acetic acid on survival and biochemical response of honey bees treated with tau-fluvalinate. Ecotoxicol Environ Saf. (2020) 189:109917. doi: 10.1016/j.ecoenv.2019.109917
27. Negri P, Ramirez L, Quintana S, Szawarski N, Maggi MD, Eguaras MJ, et al. Immune-related gene expression of Apis mellifera larvae in response to cold stress and abscisic acid (ABA) dietary supplementation. J Apic Res. (2020) 59:669–76. doi: 10.1080/00218839.2019.1708653
28. Visscher PK, Dukas R. Survivorship of foraging honey bees. Insectes Soc. (1997) 44:1–5. doi: 10.1007/s000400050017
29. Margotta JW, Roberts SP, Elekonich MM. Effects of flight activity and age on oxidative damage in the honey bee, Apis mellifera. J Exp Biol. (2018) 221:jeb183228. doi: 10.1242/jeb.183228
30. Li Z, Hou M, Qiu Y, Zhao B, Nie H, Su S. Changes in antioxidant enzymes activity and metabolomic profiles in the guts of honey bee (Apis mellifera) larvae infected with Ascosphaera apis. Insects. (2020) 11:1–12. doi: 10.3390/insects11070419
31. Gheldof N, Wang XH, Engeseth NJ. Identification and quantification of antioxidant components of honeys from various floral sources. J Agric Food Chem. (2002) 50:5870–7. doi: 10.1021/jf0256135
32. Frankel S, Robinson GE, Berenbaum MR. Antioxidant capacity and correlated characteristics of 14 unifloral honeys. J Apic Res. (1998) 37:27–31. doi: 10.1080/00218839.1998.11100951
33. Mao W, Schuler MA, Berenbaum MR. A dietary phytochemical alters caste-associated gene expression in honey bees. Sci Adv. (2015) 1:e1500795. doi: 10.1126/sciadv.1500795
34. Zhu K, Liu M, Fu Z, Zhou Z, Kong Y, Liang H, et al. Plant microRNAs in larval food regulate honeybee caste development. PLoS Genet. (2017) 13:e1006946. doi: 10.1371/journal.pgen.1006946
35. Sheehan G, Garvey A, Croke M, Kavanagh K. Innate humoral immune defences in mammals and insects: the same, with differences? Virulence. (2018) 9:1625–39. doi: 10.1080/21505594.2018.1526531
36. Palmer-Young EC, Tozkar CO, Schwarz RS, Chen Y, Irwin RE, Adler LS, et al. Nectar and pollen phytochemicals stimulate honey bee (Hymenoptera: Apidae) immunity to viralinfection. J Econ Entomol. (2017) 110:1959–72. doi: 10.1093/jee/tox193
37. Engel P, Kwong WK, McFrederick Q, Anderson KE, Barribeau SM, Chandler JA, et al. The bee microbiome: impact on bee health and model for evolution and ecology of host-microbe interactions. mBio. (2016) 7:e02164-15. doi: 10.1128/mBio.02164-15
38. Mohan A, Quek S-Y, Gutierrez-Maddox N, Gao Y, Shu Q. Effect of honey in improving the gut microbial balance. Food Qual Saf . (2017) 1:107–15. doi: 10.1093/fqs/fyx015
39. Moran NA. Genomics of the honey bee microbiome. Curr Opin Insect Sci. (2015) 10:22–8. doi: 10.1016/j.cois.2015.04.003
40. Chen C. Relationship between water activity and moisture content in floral honey. Foods. (2019) 8:30. doi: 10.3390/foods8010030
41. Grant WD, Danson MJ, Scott DJ, Halling PJ, Engberts JBFN, Ho MW, et al. Life at low water activity. Philos Trans R Soc Lond B Biol Sci. (2004) 359:1249–67. doi: 10.1098/rstb.2004.1502
42. Ohashi K, Natori S, Kubo T. Expression of amylase and glucose oxidase in the hypopharyngeal gland with an age-dependent role change of the worker honeybee (Apis mellifera L.). Eur J Biochem. (1999) 265:127–33. doi: 10.1046/j.1432-1327.1999.00696.x
43. Almasaudi S. The antibacterial activities of honey. Saudi J Biol Sci. (2021) 28:2188–96. doi: 10.1016/j.sjbs.2020.10.017
44. Bucekova M, Valachova I, Kohutova L, Prochazka E, Klaudiny J, Majtan J. Honeybee glucose oxidase - its expression in honeybee workers and comparative analyses of its content and H2O2-mediated antibacterial activity in natural honeys. Sci Nat. (2014) 101:661–70. doi: 10.1007/s00114-014-1205-z
45. Mato I, Huidobro JF, Simal-Lozano J, Sancho MT. Significance of nonaromatic organic acids in honey. J Food Prot. (2003) 66:2371–6. doi: 10.4315/0362-028X-66.12.2371
46. Ball DW. The chemical composition of honey. J Chem Educ. (2007) 84:1643–6. doi: 10.1021/ed084p1643
47. Brudzynski K, Lannigan R. Mechanism of honey bacteriostatic action against MRSA and VRE involves hydroxyl radicals generated from honey's hydrogen peroxide. Front Microbiol. (2012) 3:36. doi: 10.3389/fmicb.2012.00036
48. Bang LM, Phil M, Buntting C, Molan P. The effect of dilution on the rate of hydrogen peroxide production in honey and its implications for wound healing. J Altern Complement Med. (2003) 9:267–73. doi: 10.1089/10755530360623383
49. Molan PC. The antibacterial activity of honey. Bee World. (1992) 73:5–28. doi: 10.1080/0005772X.1992.11099109
50. Nolan VC, Harrison J, Cox JAG. Dissecting the antimicrobial composition of honey. Antibiotics. (2019) 8:251. doi: 10.3390/antibiotics8040251
51. Adams CJ, Manley-Harris M, Molan PC. The origin of methylglyoxal in New Zealand manuka (Leptospermum scoparium) honey. Carbohydr Res. (2009) 344:1050–3. doi: 10.1016/j.carres.2009.03.020
52. Rabie E, Serem JC, Oberholzer HM, Gaspar ARM, Bester MJ. How methylglyoxal kills bacteria: an ultrastructural study. Ultrastruct Pathol. (2016) 40:107–11. doi: 10.3109/01913123.2016.1154914
53. Adler LS, Fowler AE, Malfi RL, Anderson PR, Coppinger LM, Deneen PM, et al. Assessing chemical mechanisms underlying the effects of sunflower pollen on a gut pathogen in bumble bees. J Chem Ecol. (2020) 46:649–58. doi: 10.1007/s10886-020-01168-4
54. Combarros-Fuertes P, Fresno JM, Estevinho MM, Sousa-Pimenta M, Tornadijo ME, Estevinho LM. Honey: another alternative in the fight against antibiotic-resistant bacteria? Antibiotics. (2020) 9:1–21. doi: 10.3390/antibiotics9110774
55. Proaño A, Coello D, Villacrés-Granda I, Ballesteros I, Debut A, Vizuete K, et al. The osmotic action of sugar combined with hydrogen peroxide and bee-derived antibacterial peptide defensin-1 is crucial for the antibiofilm activity of eucalyptus honey. LWT. (2021) 136:110379. doi: 10.1016/j.lwt.2020.110379
56. Kwakman PHS, Zaat SAJ. Antibacterial components of honey. IUBMB Life. (2012) 64:48–55. doi: 10.1002/iub.578
57. Buttstedt A, Moritz RFA, Erler S. Origin and function of the major royal jelly proteins of the honeybee (Apis mellifera) as members of the yellow gene family. Biol. (2014) 89:255–69. doi: 10.1111/brv.12052
58. Brudzynski K, Sjaarda C, Lannigan R. MRJP1-containing glycoproteins isolated from honey, a novel antibacterial drug candidate with broad spectrum activity against multi-drug resistant clinical isolates. Front Microbiol. (2015) 6:711. doi: 10.3389/fmicb.2015.00711
59. Fontana R, Mendes MA, de Souza BM, Konno K, César LMM, Malaspina O, et al. Jelleines: a family of antimicrobial peptides from the royal jelly of honeybees (Apis mellifera). Peptides. (2004) 25:919–28. doi: 10.1016/j.peptides.2004.03.016
60. Brudzynski K. Honey as an ecological reservoir of antibacterial compounds produced by antagonistic microbial interactions in plant nectars, honey and honey bee. Antibiotics. (2021) 10:551. doi: 10.3390/antibiotics10050551
61. Endo A, Salminen S. Honeybees and beehives are rich sources for fructophilic lactic acid bacteria. Syst Appl Microbiol. (2013) 36:444–8. doi: 10.1016/j.syapm.2013.06.002
62. Lee H, Churey JJ, Worobo RW. Purification and structural characterization of bacillomycin F produced by a bacterial honey isolate active against Byssochlamys fulva H25. J Appl Microbiol. (2008) 105:663–73. doi: 10.1111/j.1365-2672.2008.03797.x
63. Goodman R. Australian Beekeeping Guide. Rural Industries Research and Development Corporation (2015).
64. Bobiş O, Dezmirean DS. Preliminary studies regarding antioxidant and antimicrobial capacity for different types of Romanian honeys. Bull UASVM Anim Sci Biotechnol. (2011) 68:91–7.
65. Bobiş O, Dezmirean DS. Preliminary studies regarding antioxidant and antimicrobial capacity for different types of Romanian honeys. Bulletin UASVM Anim Sci and Biotechno. (2011) 68:91–97. doi: 10.15835/buasvmcn-asb:68:1-2:6673
66. Bobiş O, Mǎrghitaş LA, Dezmirean DS, Gherman B, Chirilǎ F. In vitro antibacterial activity of unifloral honeys against honeybee pathogens Paenibacillus larvae and Escherichia coli. Lucrǎri Ştiinţifice-Seria Zootehnie. (2013) 60:139–144.
67. Erler S, Denner A, Bobiş O, Forsgren E, Moritz RFA. Diversity of honey stores and their impact on pathogenic bacteria of the honeybee, Apis mellifera. Ecol Evol. (2014) 4:3960–7. doi: 10.1002/ece3.1252
68. Nafea EA, Gumgumjee NM, Danial EN, Hajair AS. Physicochemical and antimicrobial properties of four Egyptian honeys with reference to American foulbrood disease. Life Sci. (2014) 11:1097–8135.
69. Isla MI, Craig A, Ordoñez R, Zampini C, Sayago J, Bedascarrasbure E, et al. Physicochemical and bioactive properties of honeys from northwestern Argentina. LWT. (2011) 44:1922–30. doi: 10.1016/j.lwt.2011.04.003
70. Chan-Rodríguez D, Ramón-Sierra J, Lope-Ayora J, Sauri-Duch E, Cuevas-Glory L, Ortiz-Vázquez E. Antibacterial properties of honey produced by Melipona beecheii and Apis mellifera against foodborn microorganisms. Food Sci Biotechnol. (2012) 21:905–9. doi: 10.1007/s10068-012-0118-x
71. Escuredo O, Silva LR, Valentão P, Seijo MC, Andrade PB. Assessing rubus honey value: pollen and phenolic compounds content and antibacterial capacity. Food Chem. (2012) 130:671–8. doi: 10.1016/j.foodchem.2011.07.107
72. Tenore GC, Ritieni A, Campiglia P, Novellino E. Nutraceutical potential of monofloral honeys produced by the Sicilian black honeybees (Apis mellifera ssp. sicula). Food Chem Toxicol. (2012) 50:1955–61. doi: 10.1016/j.fct.2012.03.067
73. da Cruz CBN, Piere FA, Carvalho-Zilse GA, Orlandi PP, Nunes-Silva CG, Leomil L. Antimicrobial activity of honeys from two stingless honeybee species and Apis mellifera (Hymenoptera: Apidae) against pathogenic microorganisms. Acta Amaz. (2014) 44:287–90. doi: 10.1590/S0044-59672014000200015
74. Gallardo-Chacón JJ, Caselles M, Izquierdo-Pulido M, Rius N. Inhibitory activity of monofloral and multifloral honeys against bacterial pathogens. J Apic Res. (2008) 47:131–6. doi: 10.1080/00218839.2008.11101439
75. Roshan N, Rippers T, Locher C, Hammer KA. Antibacterial activity and chemical characteristics of several Western Australian honeys compared to manuka honey and pasture honey. Arch Microbiol. (2017) 199:347–55. doi: 10.1007/s00203-016-1308-3
76. Morroni G, Alvarez-Suarez JM, Brenciani A, Simoni S, Fioriti S, Pugnaloni A, et al. Comparison of the antimicrobial activities of four honeys from three countries (New Zealand, Cuba, and Kenya). Front Microbiol. (2018) 9:1378. doi: 10.3389/fmicb.2018.01378
77. Malone LA, Gatehouse HS, Tregidga EL. Effects of time, temperature, and honey on Nosema apis (Microsporidia: Nosematidae), a parasite of the honeybee, Apis mellifera (Hymenoptera: Apidae). J Invertebr Pathol. (2001) 77:258–68. doi: 10.1006/jipa.2001.5028
78. Ahmad K, Khalil AT, Somayya R, Khan FN, Shan AR, Ovais M, et al. Potential antifungal activity of different honey brands from Pakistan: a quest for natural remedy. Afr J Tradit Complement Altern Med. (2017) 14:18–23. doi: 10.21010/ajtcam.v14i5.3
79. Yabes J, White B, Murray C, Sanchez C, Mende K, Beckius M, et al. In vitro activity of manuka honey and polyhexamethylene biguanide on filamentous fungi and toxicity to human cell lines. Med Mycol. (2017) 55:334–43. doi: 10.1093/mmy/myw070
80. Efem SEE, Udoh KT, Iwara CI. The antimicrobial spectrum of honey and its clinical significance. Infection. (1992) 4:227–9. doi: 10.1007/BF02033065
81. Wellford TET, Eadie T, Llewellyn GC. Evaluating the inhibitory action of honey on fungal growth, sporulation, and aflatoxin production. Z Lebensm Unters Forsch. (1978) 166:280–3. doi: 10.1007/BF01127653
82. Azonwade FE, Gbaguidi B, Paraïso A, Dah-Nouvlessounon DM, Goubalan E, Baba-Moussa F, et al. Polyphenolic profile, and antioxidant and antifungal activities of honey products in Benin. Afr J Microbiol Res. (2018) 12:9–18. doi: 10.5897/AJMR2017.8749
83. Fahim H, Dasti JI, Ali I, Ahmed S, Nadeem M. Physico-chemical analysis and antimicrobial potential of Apis dorsata, Apis mellifera and Ziziphus jujube honey samples from Pakistan. Asian Pac J Trop Biomed. (2014) 4:633–41. doi: 10.12980/APJTB.4.2014APJTB-2014-0095
84. Hamid Z, Mohamad I, Harun A, Salim R, Sulaiman SA. Antifungal effect of three local Malaysian honeys on selected pathogenic fungi of otomycosis: an in vitro evaluation. J Young Pharm. (2018) 10:414–7. doi: 10.5530/jyp.2018.10.91
85. Khan A, Jain SK. Antifungal effect of honey on fungi causing otomycosis. Bull Env Pharmacol Life Sci. (2021) 10:39–43.
86. Bobiş O, Mărghitaş LA, Dezmirean D, Gherman B, Chirilă F. In vitro antibacterial activity of unifloral honeys against honeybee pathogens Paenibacillus larvae and Escherichia coli. Lucrări Stiintifice Seria Zootehnie. (2013)60:139–44.
87. Bachanová K, Klaudiny J, Kopernický J, Šimúth J. Identification of honeybee peptide active against Paenibacillus larvae larvae through bacterial growth-inhibition assay on polyacrylamide gel. Apidologie. (2002) 33:259–69. doi: 10.1051/apido:2002015
88. Lewkowski O, Erler S. Virulence of Melissococcus plutonius and secondary invaders associated with European foulbrood disease of the honey bee. Microbiologyopen. (2019) 8:e649. doi: 10.1002/mbo3.649
Keywords: honey, honey bee, honey bee ecology, medicinal honey, hive pathogens
Citation: Fernandes KE, Frost EA, Remnant EJ, Schell KR, Cokcetin NN and Carter DA (2022) The role of honey in the ecology of the hive: Nutrition, detoxification, longevity, and protection against hive pathogens. Front. Nutr. 9:954170. doi: 10.3389/fnut.2022.954170
Received: 27 May 2022; Accepted: 04 July 2022;
Published: 25 July 2022.
Edited by:
Juraj Majtan, Institute of Molecular Biology (SAS), SlovakiaReviewed by:
May Berenbaum, University of Illinois at Urbana–Champaign, United StatesCopyright © 2022 Fernandes, Frost, Remnant, Schell, Cokcetin and Carter. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dee A. Carter, ZGVlLmNhcnRlckBzeWRuZXkuZWR1LmF1
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.