
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Nutr., 10 June 2022
Sec. Nutrition and Food Science Technology
Volume 9 - 2022 | https://doi.org/10.3389/fnut.2022.875270
This article is part of the Research TopicNatural Bioactives Used as Additives in Food ApplicationsView all 7 articles
 Uchechukwu Edna Obianwuna1
Uchechukwu Edna Obianwuna1 Vivian U. Oleforuh-Okoleh2
Vivian U. Oleforuh-Okoleh2 Jing Wang1
Jing Wang1 Hai-Jun Zhang1
Hai-Jun Zhang1 Guang-Hai Qi1
Guang-Hai Qi1 Kai Qiu1*
Kai Qiu1* Shu-Geng Wu1*
Shu-Geng Wu1*Albumen quality is recognized as one of the major yardsticks in measuring egg quality. The elasticity of thick albumen, a strong bond in the ovomucin-lysozyme complex, and excellent biological properties are indicators of high-quality albumen. The albumen quality prior to egg storage contribute to enhance egg’s shelf life and economic value. Evidence suggests that albumen quality can deteriorate due to changes in albumen structure, such as the degradation of β-ovomucin subunit and O-glyosidic bonds, the collapse of the ovomucin-lysozyme complex, and a decrease in albumen protein-protein interaction. Using organic minerals, natural plants and animal products with antioxidant and antimicrobial properties, high biological value, no residue effect and toxicity risk could improve albumen quality. These natural products (e.g., tea polyphenols, marigold extract, magnolol, essential oils, Upro (small peptide), yeast cell wall, Bacillus species, a purified amino acid from animal blood, and pumpkin seed meal) are bio-fortified into eggs, thus enhancing the biological and technological function of the albumen. Multiple strategies to meeting laying hens’ metabolic requirements and improvement in albumen quality are described in this review, including the use of amino acids, vitamins, minerals, essential oils, prebiotics, probiotics, organic trace elements, and phytogenic as feed additives. From this analysis, natural products can improve animal health and consequently albumen quality. Future research should focus on effects of these natural products in extending shelf life of the albumen during storage and at different storage conditions. Research in that direction may provide insight into albumen quality and its biological value in fresh and stored eggs.
Globally, meeting the demands of an ever-increasing population and per catput protein requirement is quite a challenge. Chicken eggs are among the most commonly consumed animal protein worldwide due to their low cost, nutritional contents, and biological functions (1), thus significantly contributing to human nutrition. Rapid growth in poultry production has reflected in direct increase in egg production. Critical concern in egg production is the issue of maintenance of egg quality, particularly the albumen quality, since most of the functional properties of the egg is related to the albumen.
The albumen contains proteins (ovomucin, ovalbumin, ovotransferrin, and lysozyme), peptides and amino acids which are natural antioxidant compounds (2). The ovomucin is considered as one of the core albumen proteins that plays a key role in maintaining albumen structure by stabilizing protein-protein bond interaction, viscous-gel nature, and height of the thick albumen, culminating in increased Haugh unit value (3, 4). The Haugh unit (HU) value is often used to measure albumen quality, high HU value depicts thick albumen content and gelly nature with strong viscosity which reflects better albumen quality while low HU unit value is vice versa (5). The improved albumen structure and proteins, confers on the albumen its biological functions and excellent technological properties such as foaming, gelling, emulsifying and water holding capacity. Albumen of improved quality could be utilized as raw material for food processing and health industry; consumption of food-derived antioxidants such as egg white peptides and egg white powder as functional foods exert beneficial effects on human health conditions by scavenging reactive oxygen species and free-radical mediated chain reactions (6). On the other hand, poor albumen quality is probably due to reduced ovomucin-lysozyme complex interaction, disaggregation of ovomucin subunits (α- and β-) and reduction of highly glycosylated β- subunit in thick albumen gel (3, 7, 8). All these changes eventually collapses gelatinous structure of thick albumen into a clear liquid (albumen thinning) as albumen consistency is lost (9) as well as the rheological, nutritional and technological properties which consumers rely on. Therefore, exploring the potentials of egg albumen as functional food and ingredient is reliant on the quality of albumen (technological and biological properties) obtained from the egg.
Albumen quality are influenced by various factors; animal feed (10), birds age (11) and storage environment (12). The role of diets in animal health and consequently performance cannot be overemphasized; cottonseed meal reduced albumen proteins and viscosity, and impaired ovary health (13), vanadium, an inorganic mineral, caused cell apoptosis in the magnum and reduced the immune capacity function of laying hens (10) while dietary fluoride reduced albumen quality and immune function of the animals (14). Therefore, adoption of nutritional modulations as a strategy for improving albumen quality and animal health is advocated for.
However, the use of non-natural feed source is often associated with challenges such as antinutritional factors, accurate inclusion levels, negative effect on animal health (15), and consumer concerns about residue, toxicity, pollution, and low biological value limit their optimal use. In recent times, the use of natural products from plants and animal origin as feed additives in diets of laying hens have gained attention. The use of natural-source feed such as natural products with antioxidant and antimicrobial properties derived from plants and animals are advantageous because they pose no residue effect, pollution, or toxicity risk. Natural products including antimicrobial peptides (16), purified amino acids from animal blood (17), small peptide synthesized from corn (18), prebiotics (19), probiotics (20), essential oil (21) and organic trace elements in the form of chelates or amino acid complexes (22) have been reported to influence albumen quality. The improvement in albumen quality with natural products are associated with their high bioavailability, antioxidant function and capacity to maintain gut microflora, which is important for animal health and bio-fortification of animal products. The improved albumen quality is important to both consumers and producers. As a result, this review provides an in-depth understanding of the impact of various natural diets of plant and animal origin and their inclusion levels on egg albumen quality and overall health of laying hens.
Egg albumen is a homogeneous, non-crystalline substance (liquid) formed around the egg yolk (Figure 1). The albumen structurally is composed of different layers, each with its peculiar function, namely: the chalaziferous white or inner thick layer resting around the yolk which stabilizes the yolk movement such that it maintains its center position; the inner thin layer lying very close to the chalazae which serves as a protecting capsule enveloping the yolk (this layer has more fluid and contains most of the albumen protein); the outer thick white layer, lying adjacent to the inner thin white layer, provides further fluid and texture to the albumen fluids; and the outer thin white layer which holds further protein-based nutrients and compounds that play essential role in the growth and development of embryo in the fertilized egg (23, 24). The principal function of the albumen is as a natural defense system for the yolk in table eggs and embryo in fertile eggs.
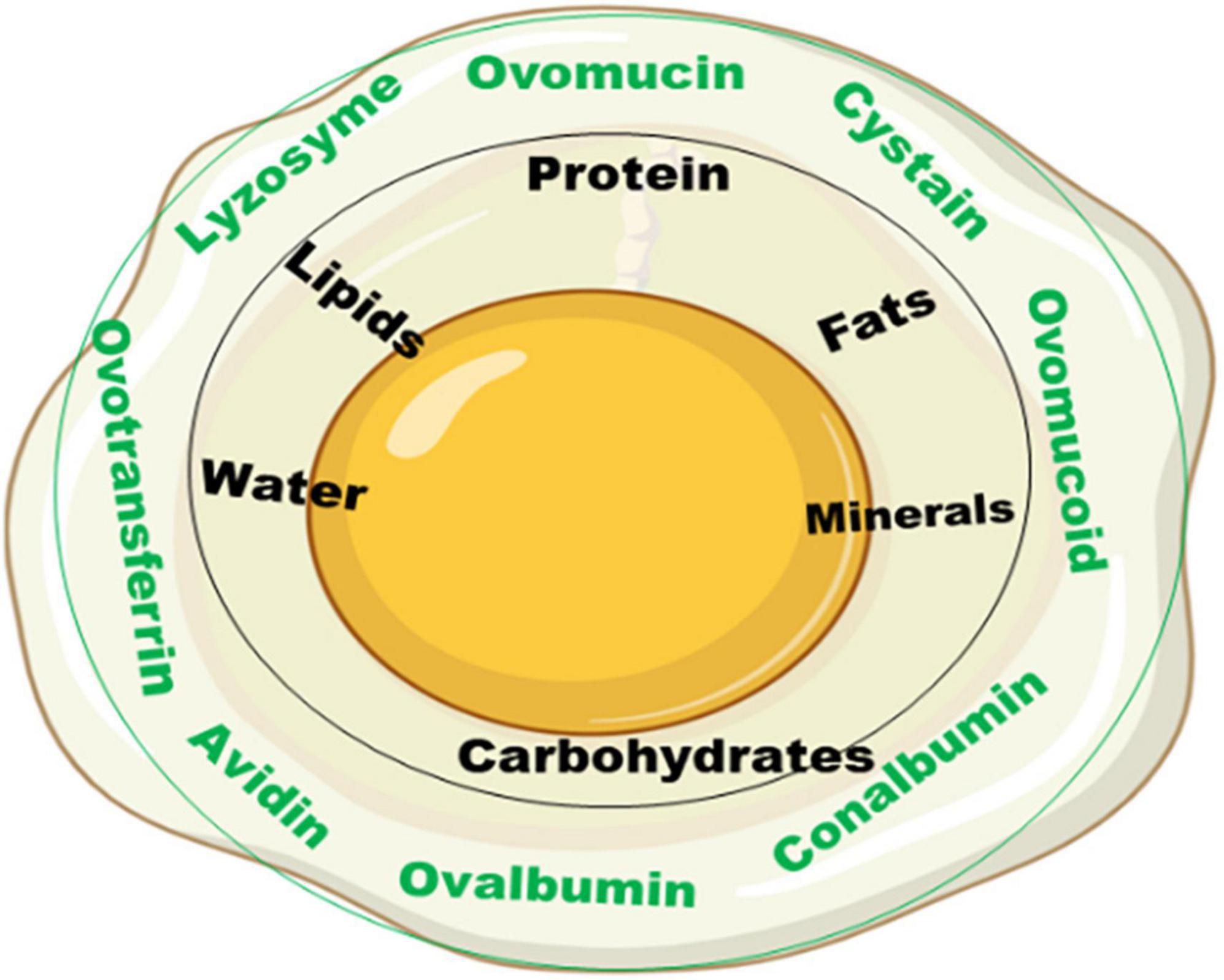
Figure 1. Albumen structure and its composition.
Chemically, the albumen consists primarily of water and proteins, containing about 88% water, and 10–11% proteins. Other nutrients such as trace minerals, vitamins, lipids and carbohydrates are also found in the albumen. The albumen proteins are, however, of much more biological relevance than other nutrients. The major albumen proteins include ovalbumin, ovomucoid, conalbumin, ovomucin and lysozyme (Figure 2). The ovoglobulins, ovomaroglobulin, avidin, ovoinhibitor, cystatin, ovoglycoprotein, and ovoflavoprotein are considered minor albumen proteins (23). However, each of the albumen proteins has specific physical and chemical characteristics which confer critical biological functions on the albumen, as shown in Figure 3.
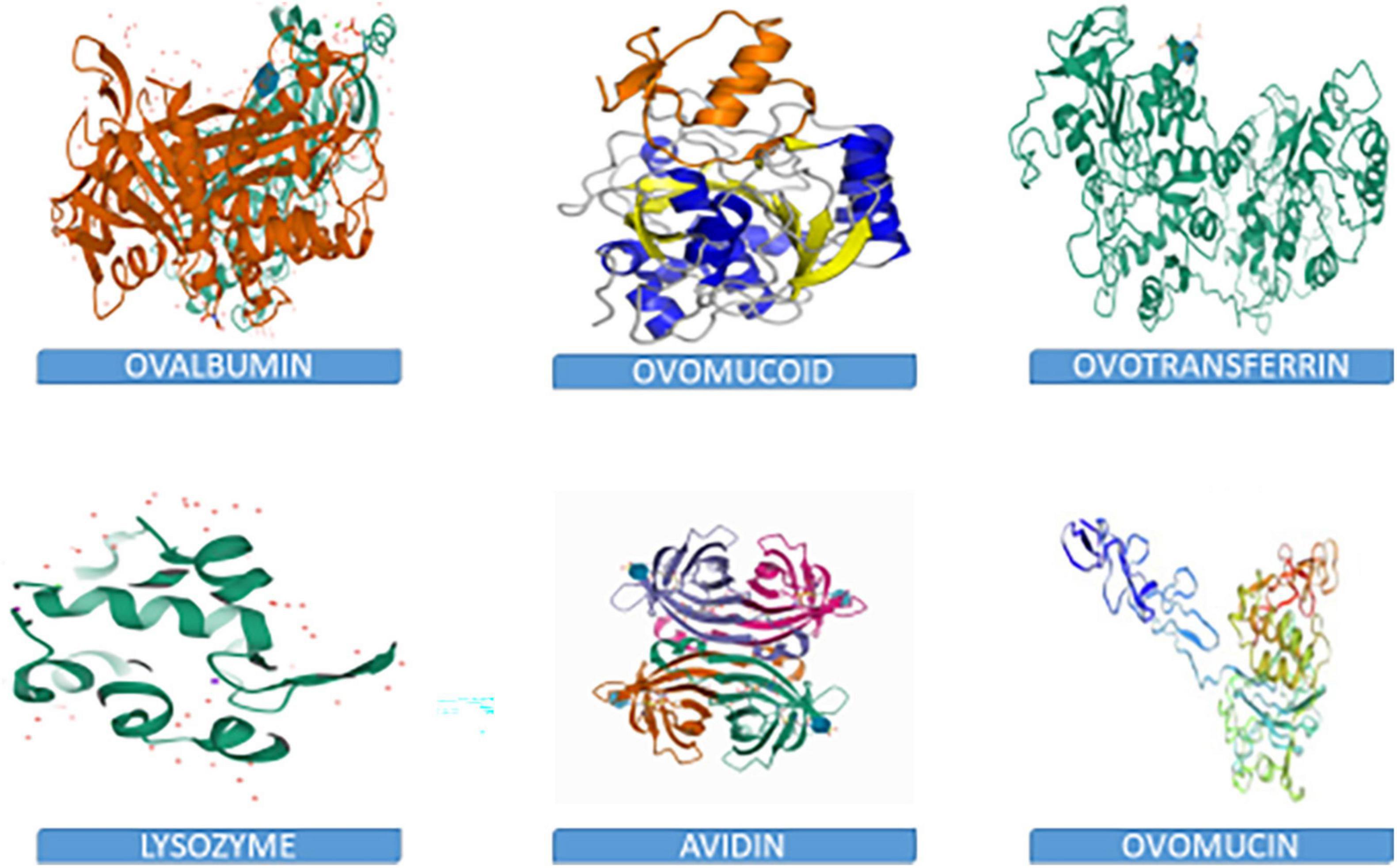
Figure 2. 3D structures of albumen proteins.
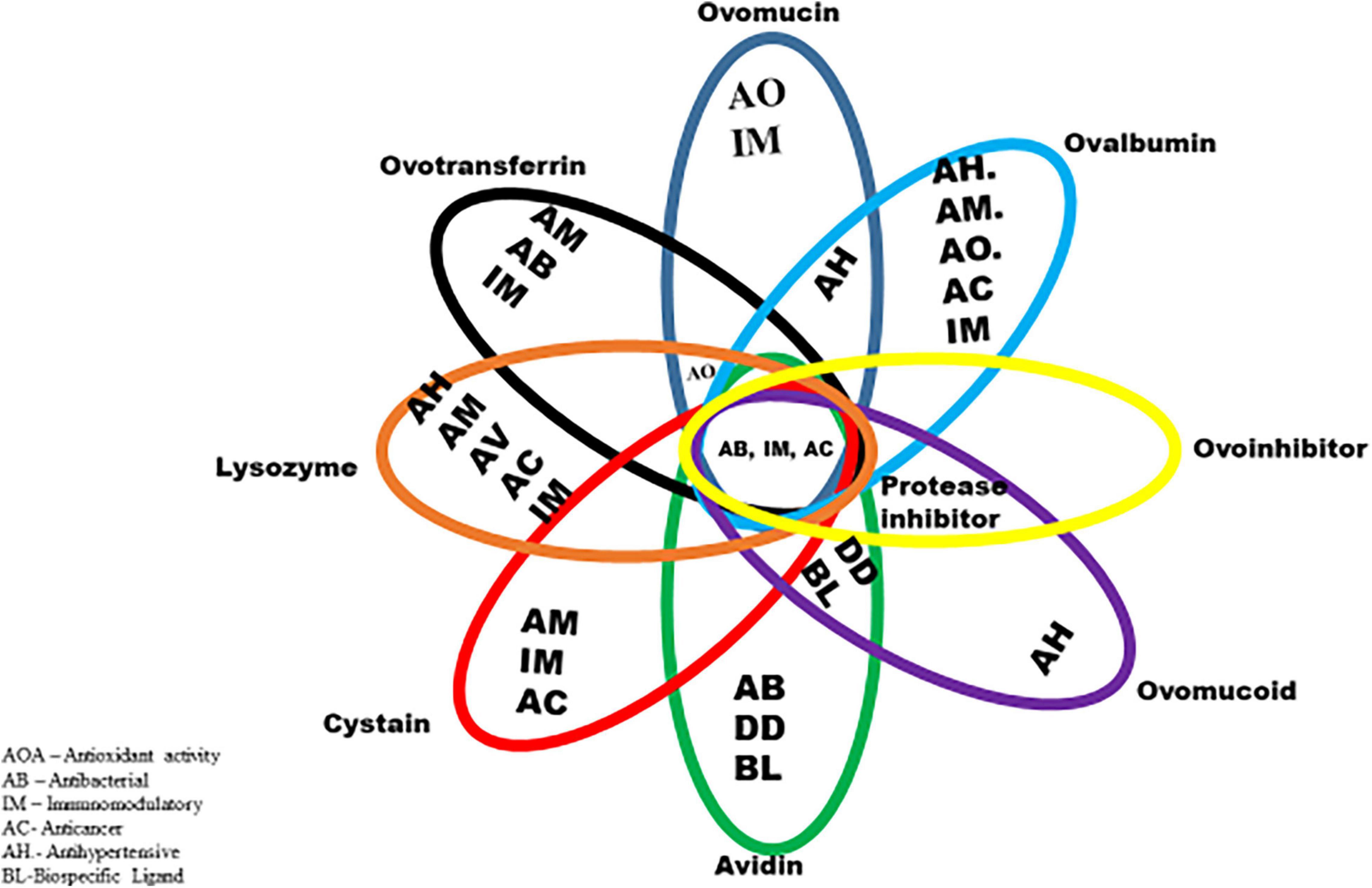
Figure 3. Biological functions of albumen proteins.
The foaming (whippability), gelling (coagulation) and emulsifying properties are conferred on the albumen by (ovalbumin, lysozyme, and ovomucin); antimicrobial properties (lysozyme and ovotransferrin), which allows its exploitation as a functional food material in many food, pharmaceuticals and other material industries (24). The mechanisms through which these proteins exert biological functions, may include bacterial cell wall degradation and binding to vitamins and metals (25). Of all the different types of protein found in the egg albumen, the ovomucin content has been demonstrated by various studies to, play a significant role in determining the albumen nature and hence its quality (3, 8, 26, 27).
Ovomucin is a glycoprotein accounting for nearly 3.5% of total egg white protein. It is made up of α- and β-subunits with molecular weights of 5.5 and 8.3, and isoelectric points (pI) of 4 and 5.5, respectively (3). α-ovomucin is homogenous and has lower carbohydrate content (15%), whereas β-ovomucin is heterogeneous and rich in carbohydrate (50%) (26). The carbohydrate chain of ovomucin consists of sialic acid (2.5–8%), hexose (15–18.6%), and hexosamine (7–12%) (6). The amino acid content of the two ovomucin subunits differs: α- and β-subunits of the ovomucin, respectively, contain acidic (glutamic acid and aspartic acid) and hydroxyl amino acids (threonine and serine) (28). Albumen’s gel-like properties are due to the complex formed by the two subunits of ovomucin, resulting in insoluble thick albumen (29). Ovomucin is strongly heat-stable because of its random coil nature, and, as such, it does not exhibit thermal transition temperature. Albumen liquefaction occurs when the α- and β-subunits, making up the thick albumen, are depolarized. This result in a lower proportion of insoluble ovomucin with a high carbohydrate content (26). There is a strong relationship between ovomucin and albumen quality because the β-ovomucin is fundamental to albumen quality determination.
It has been reported that ovomucin polymers and the ovomucin-lysozyme complex interaction are fundamental to albumen height, viscosity, elasticity, and the gelatinous nature (30). The ovomucin content and height of thick albumen are interrelated reflecting in the HU value, because of the relationship between HU value and ovomucin abundance (9). In addition, the thick albumen of eggs with high HU values had strongly glycosylated β-ovomucin compared to eggs with low HU values (27). A thick gelatinous nature, strong viscoelasticity, rich glycosylated β-ovomucin, and high HU values are albumen quality indicators largely driven by ovomucin content and activity. Improved thick albumen quality due to ovomucin content is important for producers, consumers, and breeders.
The ovomucin-albumen quality relationship is influenced by various factors, including; magnum morphology, the rate of albumen secretion, diets, storage time, and temperature. The β-ovomucin secreted by the magnum maintains magnum health (4), viscoelasticity of albumen (9), and increases albumen durability due to complex formation with proteins and polysaccharides (31). Hence, the quality of β-ovomucin secreted by the magnum indicates albumen quality. The incomplete mucosal columnar epithelial cell layer of the magnum and low magnum index reduces rate of ovomucin deposition in the albumen, thereby decreasing albumen secretion (32). Antinutritional factors impair protein digestion and secretion, thus reducing albumen secretion and ovomucin deposition in the albumen (33). Diets such as amino acids (28, 34) and green tea (4) enhanced β-ovomucin content in thick albumen while cotton seed meal (CSM) decreased ovomucin content (13). Increased storage time decreased ovomucin content in the albumen of stored eggs (35). The mechanism of how other albumen proteins contribute to albumen quality requires further investigation. Freshness of eggs is often assessed based on HU value and there exists a relationship between ovomucin abundance and HU value, thus HU value is a measure of albumen quality.
The HU measures egg quality and stability based on the height of thick albumen and the egg’s weight (36). Also, the HU value is determined by the stability of the protein chains in the albumen. The viscous-gelly nature of the thick albumen reflected in the HU value is influenced by the ovomucin content (3). A strong correlation between the two indices (i.e., thick albumen and HU) was reported by Wang et al. (9). In addition, (8) found that birds of genetic lines with high HU scores had higher ovomucin content and isoleucine levels than those with low HU scores. The USDA used HU to classify egg quality as; AA-grade at HU > 72; A-grade at HU = 60–72; B-grade at HU < 60, C-grade < 31 (37), and high HU values indicate high albumen quality. The effects of various diets on HU values are presented in Supplementary Table 1. From the analysis: green tea, organic trace elements, phytobiotics and natural oil significantly improved HU values compared to other diets.
Egg white proteins are excellent sources of bioactive peptides; thus, albumen of high quality is critical to the food and health sectors. Over the past decades, most bioactive peptides were derived from milk and soybean proteins, but peptides from egg white proteins have been produced recently. These food protein-derived bioactive peptides, are health enhancers and have quite less safety risk. Food bioactive peptides which exert physiological effects on humans, are inactive within the sequence of their parent protein and can be activated by enzymatic hydrolysis during food processing (38). Under different conditions, these active egg white peptides have been hydrolyzed with pepsin, trypsin, and α-chymotrypsin enzymes. The hydrolysis of egg white enhances the activities (antioxidant, antimicrobial, anticarcinogenic, mineral-binding, and antihypertensive) of albumen proteins and their peptides (39), confirming the multi-functionality of these egg-derived peptides. Egg white derived-bioactive peptides could be considered as natural antioxidants owing to its capacity to control lipid peroxidation and supply essential amino acids, thus have promising potential over synthetic antioxidants in the food-processing industry. Also, egg white bioactive peptides can be consumed as functional foods to replace synthetic antioxidants without dosage or safety concerns.
The reports of Manso et al. (40), Rupa et al. (41), Sun et al. (42), Garcés-Rimón et al. (43) demonstrated that functional peptides from egg white and various egg white proteins have beneficial effects on human health and the food industry; hence, they could be used as drugs and functional foods. Hydrolysis of albumen proteins yields peptides with strong antioxidant capacity. Alcalase peptide with the sequence (Tyr-Ala-Glu-Glu-Arg-Tyr-Pro-Ile-Leu) has a high antioxidant capacity and free radical scavenging ability (21, 43). In addition, peptides with the sequence (Try-Leu-Gly-Ala and Lys) and (Gly-Gly-Leu-Glu-Pro-Ile-Phe-Glu) exhibited significant antioxidant effects in 2,2-diphenyl-1-picrylhydrazyl radical scavenging capacity (DPPH) and lipid peroxidation inhibition assays (44). Moreover, (45) identified another peptide showing higher antioxidant capacity in oxygen radical absorbance capacity fluorescein (ORAC-FL) assay. The high-antioxidant nature of these egg white peptides harnesses their potentials to be consumed or used in food processing as natural food antioxidants.
The angiotensin-converting enzyme “ACE” suppressing activity of peptides promotes their use for mitigating high blood pressure. Ovalbumin peptides exert a high inhibitory effect on ACE activity (46). Chiang et al. (47) discovered that a bioactive peptide produced by hydrolyzing egg white protein with thermolysin inhibited ACE activity in hypertensive rats. Similarly, egg white peptides ameliorated oxidative stress and hyperlipidemia in hypertensive rats (40). Also, Garcés-Rimón et al. (43) demonstrated that bioactive peptides of egg white exerted strong antioxidant effects leading to prevention of abnormalities in experimental models that are obese and hypertensive. On the other hand, enzymatic hydrolysate of lysozyme produced LPH2, showing a strong ACE inhibitory effect, high antioxidant activity in ORAC-Fl assay, and resistance to gram-negative and gram-positive bacteria (48). These egg white derived bioactive peptides as natural products would provide a safe alternative to hypertension therapy over synthetic agents that may be associated with side effects.
Egg white proteins and its peptide derivatives have immunomodulatory properties, which allows them to be used to improve immune responses for cancer and other diseases. Egg white peptides can stimulate macrophage activity and block the tumor necrosis factor (TNF)-mediated NF-κB pathway (42), while tumor growth suppressions are linked to ovomucin glycopeptides (49). In addition, cystatin induces TNF and interleukin (IL)-10 synthesis while avidin enhances antitumor effect via improved upregulation of TNF-α and thus acts as an anticancer agent and drug carrier (50). Also, feeding ovalbumin peptides to hypertensive rats reduced blood pressure and had an immunomodulatory effect by increasing the production of anti-inflammatory cytokines (IL-12, 227 IL-17, and IL-10) (40), while inhibiting the production of IL-4 (41). Ovomucin and ovomucin-derived peptides induce macrophage synthesis in vitro, making them immune modulators (25). Ovotransferrin peptides have been shown to exert anticancer effect by improving the activity of superoxide dismutase and resistance of human breast and colon cells to cancer (51). Also, ovotransferrin peptides can form complexes easily with iron, and the bound iron can be in a free form at a pH of < 4.5 (52). Hence, ovotransferrin peptides can be used as human drugs in the form of iron supplements (53). This means they are useful in human medicine. It has been reported that feeding ovalbumin peptides to hypertensive rats reduced blood pressure and had an immunomodulatory effect by increasing the production of anti-inflammatory cytokines (40). Also, peptides of lysozyme stimulate immunoglobulins production and increase pro-inflammatory cytokine production (54). Also, ovomucoid could be used as a drug carrier since it possesses a bispecific ligand. Ovomucoid derived peptides demonstrated immunomodulating activity against T-cells; hence, they have promising potentials to be employed for pharmaceutical use in humans. All these highlight the usefulness of albumen bioactive peptides for human drugs with anticancer, antitumor and anti-hypertensive effects.
Egg white hydrolysates also protect the body against neurological and cardiovascular dysfunction induced by metals as well as preserve sperm viability by preventing membrane impairment, testis and epididymis histopathological damage (55). In rats administered 4.6 μg of Hgcl2 (mercury chloride) intramuscularly, the resultant effect was cardiovascular dysfunction due to high level of oxidative stress. Whereas, in rats exposed to both Hgcl2 and egg white hydrolysate, the egg white hydrolysate mitigated cardiovascular damage by neutralizing the activities of free radicals (56). Hence, egg white hydrolysate could be used as ingredients for functional foods that could act as a substitute for synthetic drugs to treat cardiovascular damage induced by mercury.
Egg albumen has potential in the food industry. Albumen bioactive peptides act in capacity of maintaining microbial safety of albumen as raw materials for industries and in processed foods.; ovomucoid inhibits the trypsin protease, lysozyme exerts bacteriolytic activity on food pathogens, ovomacroglobulin suppresses viral hemagglutination and ovoinhibitor inhibits bacterial and fungal serine proteases (57, 58). The bacteriolytic and antimicrobial effect of ovotransferrin on pathogenic bacteria, including Pseudomonas spp., E. coli, and Streptococcus mutans has been reported (59). Ovotransferrin peptide, OTAP-92, destroy Gram-negative bacteria by invading the bacterial membrane using autolysis, thereby damaging the cytoplasmic membrane (60). According to (61), ovotransferrin regulates food-poisoning activities of E. coli O157:H7 and Listeria monocytogenes. The antimicrobial capacity of lysozyme is strong compared to other albumen proteins because of its potential to regulate the proliferation of bacteria and fungi that cause food spoilage (57). Listeria monocytogenes and Clostridium botulinum which cause problems in the food industry due to its formation of toxins in plants and animals, have been successfully regulated by lysozyme activity (62). Albumen bioactive peptides could be used as natural antimicrobials to ensure food safety and reduce food spoilage.
Fresh juices, beer and wine characterized with haziness challenge consumer acceptance. To meet consumer demands in terms of clarity and sensory quality, clarifying agents are incorporated into wine during the brewing process. Albumen is used as a clarifying agent in wines to remove excess tannins and decrease wine’s astringency (55). Albumen proteins are used as raw materials in industries; egg white proteins are used as antioxidants in processed food products like cheese and sausages. Lysozyme is used as an ingredient for dentistry care products, including mouthwash, toothpaste, and chewing gums (25), because it exerts an antimicrobial effect on the oral mucosa and an inhibitory effect on periodontitis-causing bacteria. Ovomucin confers excellent texture in food products due to its protein-resistant characteristics. Hydrogels of egg white can be developed and used as bioactive material for tissue engineering applications. Owing to the health benefits of egg white bioactive peptides, they are used for health-promoting foods and drugs. It then becomes imperative to produce eggs with high albumen quality, in order to harness its usefulness as beneficial raw materials for food processing and health industry.
Nutrition plays a key role in relation of laying hen’s health and product quality. Laying hens are subjected to various physiological and environmental stressors, affecting their health and productivity. In aged laying hens, lipid metabolism and fat accumulation changes are common and the fatty liver hemorrhagic syndrome is a common cause of mortality (63). Thus, regulating lipid metabolism is critical for animal health and welfare, and maintaining egg production and product quality. The lower part of the hen oviduct is open to the cloaca and subject to colonization by various microorganisms via the vagina; this part of the oviduct is more susceptible to infection by microbes (64), resulting in a decrease in lymphocytes and an increase in macrophages during lay. This may negatively affect laying performance and consequently egg quality. The chicken’s gastrointestinal tract is crucial to animal health; the gut microbial population and intestine morphology facilitate nutrient utilization and absorption, enhancing performance and egg quality. In late laying hens, a decline in egg production and egg quality, could be due to impaired nutrient utilization, protein metabolism, alteration of gut beneficial microbial population and low synthesis of reproductive hormones (65). Therefore, age or health status could exert detrimental influence on intestinal health and functioning capacity, leading to decline in laying performance and egg quality. In same vein, serum parameters including antioxidant enzymes; glutathione peroxidase (GSH-Px), glutathione S-transferase (GST), catalase (CAT), superoxide dismutase (SOD), total antioxidant capacity (TAOC), and oxidative biomarker: malondialdehyde (MDA) are related to oxidative stress, while immunoglobulins (IgA, IgM, and IgG) are related to immunity. The antioxidant capacity and immune functions are often used as indicators for assessing physiological status of laying hens. Nutritional strategies could be used to improve animal health via increased activity of antioxidant enzymes, immunoglobulin synthesis, maintenance of gut integrity which may translate into improvement in laying performance and egg quality.
According to (66), yeast can modulate the immune response and maintain a balance between innate and protective immunity in chickens by lowering 1L-1 and 1L-2. Reduced 1L-1 expression indicates a fewer number of macrophages, implying that the hens’ health is suitable for producing high-quality eggs. Recent reports showed that diet supplementation such as phytogenic feed additive (67) and natural astaxanthin (68) improved intestine morphology, small organic peptide (18) and organic selenium (69) enhanced intestinal health while alfalfa meal (70) enhanced beneficial gut microbial population. Diets such as tea polyphenols at 600 mg/kg (71) and 400 mg/kg (72), and phytogenic extracts (73) have been found to enhance immune function capacity, while magnolol enhanced the antioxidant capacity of the ovary (74). In addition, prebiotics (75, 76) improved serum antioxidant capacity, and probiotics exerted positive effect on beneficial gut microbes (77) and nutrient utilization (78). Organic trace element such as glycerol monolaurate enhanced egg quality via improved gut function and antioxidant capacity in late-laying hens with low health status (65). These natural products that exert positive effect on animal health and egg quality may present a mechanism of action based; on the alteration of intestinal microbiota, increased enzyme secretion, improved immune response, antioxidant activity, and morpho-histological maintenance of the GIT. Nevertheless, Miao et al. (14) reported that 800 mg/kg dietary fluoride caused damage to the liver, ovary and kidney with decreased albumen quality as resultant effect. Similarly, higher caffeine concentration in green tea decreased albumen quality and did not improve serum biochemical indices (71). These adverse effects suggest that the threshold for the inclusion level have been exceeded and animals could not tolerate this range. This is often a problem associated with synthetic feed additives.
There are several attempts to improve laying hens’ antioxidant capacity, to reduce stress and maintain high egg production. These efforts are anticipated to decrease the production of toxins likely to be transferred into the eggs and ensure safe product quality. Animal health and egg quality are inter-related because animal health influences egg quality. Thus, animal health indicators such as immune response, serum antioxidant capacity, hematology, and intestinal morphology and function may be improved with nutritional strategies, which finally culminates to enhanced egg production and quality.
One of the factors that negatively affect albumen quality is oxidative stress. Oxidative stress is a physiological condition in the system where the outputs of reactive oxygen species (ROS) tends to alter the vigor of antioxidant systems at either the cellular or system level (10, 79). Oxidative stress in laying hens could be due to type of diet (10, 80) and age (81–83). Oxidative stress culminate in surge of biochemical reactions including: lipid and protein peroxidation, oxidative alteration of amino acid residues, and destabilization of protein complex and function due to increased ROS (84). All these may alter the normal physiological process in the body of the animal with consequent negative effect on egg production and quality.
Increased level of oxidative products like thiobarbituric acid reactive substances and Malondialdehyde (79), methane dicarboxylic aldehyde, ketones and carbonyl protein complexes (85) both in the blood plasma and egg are linked with oxidative stress. The increased level of oxidative products often causes a reduction in activities of antioxidant enzymes (SOD, CAT, GSH-Px, and T-SOD) which are fundamental to protection of cellular structures from damaging effect of ROS (2). Adverse effects of oxidative products including; damaging of the oviduct and consequent reduction in protein synthesis (10), impacts negatively on the taste and flavor of albumen, reduce the nutritive and antioxidant capacity of egg white (86), hence poor albumen quality. Also, oxidative stress often destabilizes homeostasis of cecal microflora and destroy the immune capacity of the intestinal mucosa (77), this invariably impairs the health of the animal, nutrient utilization and consequently egg quality. Further, diet-induced oxidative stress may degrade albumen proteins and invariably albumen quality. One study reported that vanadium-based diet caused a reduction in albumen quality by decreasing the abundance of an egg white protein; lysozyme (Q6LEL2) which possesses antimicrobial properties while increasing protease inhibitors that restrain activities of some proteins (87). It is crucial to enhance endogenous antioxidant enzymes in order to mitigate the adverse effects of oxidative stress on animal and its product as well.
Endogenous antioxidant enzymes SOD and CAT that make up the antioxidant cellular enzymatic system, function as the first line of defense and are important indicators of the oxidation status of animal tissues (84). SOD catalyzes the dismutation of a superoxide anion (O2–) into hydrogen peroxide (H202) and an oxygen molecule (O2) to reduce the damage caused by the former. SOD also works with CAT, which converts H2O2 to H2O (88). This implies that enhanced antioxidant enzymes in the animal scavenge for ROS and protect the animals’ cellular structure and consequently product quality. In this line, the antioxidant capacity of egg white is enhanced, when the content and activities of antioxidant enzymes and antioxidant indicators, such as T-AOC and oxygen radical absorbance capacity “ORAC,” are increased as MDA levels are decreased (78, 86). Therefore, the use of natural antioxidants would enhance animal health and albumen synthesis and quality via: prevention of oxidative tissue damage, inhibition of oxidation and modification of nutrients.
Introduction of green/safe feed additives of natural products such as prebiotics, probiotics, phytobiotics, organic trace elements and minerals, and vitamins in poultry diets could improve albumen quality. In addition, albumen quality often depends on protein secretion, which more or less is dependent on the dietary protein and amino acid intake. There is also the need to supplement in the diet of laying hens, natural antioxidants which are safer, less toxic, and have high bioavailability compared to synthetic antioxidants. These may provide eggs with albumen of high HU values, strong oxidative stability of albumen proteins, excellent rheological and technological properties, thus, better and acceptable egg quality. Adoption of nutritional strategies to enhance albumen structure with strong stability of albumen proteins, has therefore been promoted as essential means of improving albumen quality prior to lay while also improving animal health. The effects of various diets on albumen quality and animal health are presented in Supplementary Table 1.
Probiotics and prebiotics as natural feed additives that are non-toxic and safe for consumers, are gaining popularity as antibiotic-free alternatives in poultry nutrition. Probiotics could be used in diet of laying hens as single strains or combination of various strains (89, 90). Most commonly microorganisms used as probiotics in poultry production include colonizing species of Streptococcus, Lactobacillus, Clostridium, Bacillus, and Enterococcus.
Some studies have illustrated the additive effect of C. butyricum in improving albumen crude protein content (90) and HU value (91). In addition, probiotics such as En. faecium and L. fermentum (92), B. subtilis (20), B. licheniformis and B. subtilis (77), B. velezensis (78) and B. amyloliquefaciens (93) enhanced albumen quality and HU value. Similarly, albumen quality (i.e., albumen height) was improved in aged laying hens (20). Albumen nitrogen (94) and crude protein content (90) was increased in laying hens fed Saccharomyces fermented product and C. butyricum, respectively. In a likely manner like other probiotics, effective microorganisms (EM), a natural product consisting of various microbes (photosynthetic bacteria, actinomycetes, yeast, Lactobacillus) (95), enhanced albumen height and HU values (96). The improvement in albumen quality due to dietary probiotics may be associated with influence of probiotics on animal health and physiological status. Probiotics have been used in poultry nutrition to enhance activities of digestive enzymes, produce volatile bacteriostatic substances which favors breakdown of nutrient in feed, maintain gut microecological environment and improve intestinal villi morphometrics (97–99). All these facilitate nutrients absorption and utilization by the animals for albumen synthesis. For example, probiotics have been reported to enhance protein metabolism with consequent improvement in albumen quality (100). In addition, probiotics have been found to improve to enhance redox balance in animals evidenced by enhanced activities of antioxidant enzymes and secretion of immunoglobulins (93, 98). Probiotics may present improvement of antioxidant capacity, gut integrity, and immunity function as mechanisms that boost process of egg formation and albumen synthesis in laying hens. However, there are other evidences that probiotics had no significant effect on albumen quality (98, 101–104). These variations among studies may be due to probiotics constituents, dosage level and physiological status of the birds.
Prebiotics which are non-digestible oligosaccharides have been reported to exert various effects on albumen quality of laying hens. There are evidences that prebiotics such as dietary marine-derived polysaccharide (105), mannan oligosaccharides (106), yeast cell wall supplement (107) and sugar beet syrup (19), supplemented in the diets of laying hens improved albumen quality. The significant improvement in egg quality due to dietary prebiotics could be linked with beneficial effects of prebiotics on intestinal villi structures, activities of intestinal enzymes, gut environment by increasing beneficial microbes and suppression of pathogens (108, 109). These beneficial effects facilitate nutrient absorption and utilization, which are translated into improved egg production and protein synthesis. Also, prebiotics have been found to enhance ovary health, increase antioxidant and immune function (110, 111). Prebiotics influence on albumen quality may be explained by its protective effect on gut integrity, reproductive tract and oxidative stability in both serum and tissue. In contrast, Zhou et al. (112), Sozcu and Ipek (113) found no significant effect of dietary xylooligosaccharides and lignocellulose, respectively, on albumen traits. All these highlight the crucial role of diets in maintaining animal health and animal product quality, although there is less literature on influence of prebiotics on laying hens.
The use of phytobiotics as antibiotics replacement in the poultry industry has gained much attention; consequently, its employment for enhancing performance and egg quality is expected. Phytogenics could enhance the safety and stability of eggs, which is critical for the egg-food industry. Some phytogenics supplemented in laying hen diets include some plant extracts: Chinese herbal extracts – Lonicera confuse and Astragali Radix extracts (114), ginger extract (115), and the combination of probiotics and plant extracts (116), have been reported to improved albumen quality. Other studies have also reported that phytogenics in powdery form by combination of various phytogenic feed additives (73, 117, 118), alfalfa meal (70), fruit – Ligustrum lucidum (119) enhanced albumen quality. However, a higher level of mulberry leaf inclusion reduced egg weight and increased HU but had no effect on albumen height (120). Furthermore, some phytogenic compounds such as Chinese herbal mixture (121), garlic and ginger root powder (122), garlic and onion powder (123), and Ricinus communis leaf powder at graded levels (124) had no significant effect on albumen indices and HU values. Fermented brown algae by-products, on the other hand, significantly decreased the HU value of eggs (125).
The phytogenic enhanced effect could be attributed to the beneficial effects on the oviposition process, antioxidant capacity, and efficient conversion of digested feed into eggs. The bioactive ingredients in phytogenics, such as menthol in peppermint leaves (126), the chlorogenic acid (CA) a type of phenolic acid in Chinese extracts (114), anthocyanins in grape seeds (127), and gingerols in ginger (128) have shown antimicrobial, antifungal, and antioxidant effects. This can reduce β-ovomucin degradation and invariably enhance its content in the thick albumen. In addition, the bioactive ingredients of herbal plants have been shown to improve the integrity of the magnum and uterus while aiding digestion and nutrient absorption (129). These result in increased protein synthesis, albumen secretion and ultimately higher HU values. The presence of antinutritional factors, the level of inclusion, and the nature of active ingredients may all play a role as was reported in previous works.
One economic important phytogenic additive is the green tea powder (GTP). GTP contains tea polyphenols TP, a natural antioxidant of typical flavonoids (130). The major compounds in TP are epicatechin (EC), epigallocatechin (EGC), epicatechin gallate (ECG), and epigallocatechin-3-gallate (EGCG) (131). The use of GTP as a feed additive in laying hens’ diets to improve albumen quality have been widely reported (72, 132, 133). Positive effect of tea polyphenols on albumen quality have been demonstrated in literature (134). Also, tea extracts enhanced the strength of albumen gels (135). All of which significantly enhanced albumen quality and HU value. Specifically, the improvement in albumen quality may be linked with its influence on albumen proteins. Tea polyphenols have the capacity to bind to proteins and influence their structures (136). Also, GTP supplementation increased the β-ovomucin content of thick albumen (72). In contrast, increased levels of GTP supplementation decreased albumen weight and HU value, indicating that a high dosage of caffeine may impair albumen quality (71). However, 137 reported that TP supplementation did not affect albumen indices and HU values.
The varying effects of dietary polyphenols on egg quality could be related to various factors, such as different TP components (EC, EGC, EGCG, and caffeine), TP inclusion level (71), the difference in the polyphenol compounds isolated from different plants (138), and duration of feeding. The study by Zhang et al. (5) observed significant effect of dietary EGCG on albumen quality after 8 weeks of feeding. The study of Wang et al. (80) reported that EGCG could be more effective when fed to laying birds under oxidative stress or aging. Thus, age of laying hens, tea components, physiological status and duration of feeding could account for the variations in different studies. The effect of green TP on albumen quality may be explained by some underlying proteomic mechanisms.
Proteomics reports have highlighted some mechanisms underlying the improved albumen quality of eggs from birds fed diets supplemented with green tea polyphenols. EGCG increases the level of ovalbumin (OVA)-related Y protein and decrease OVA-related X protein (32, 80). The free sulfhydryl and increased hydrophobicity properties of OVA may influence the degradation of thick albumen protein and enhance albumen height and HU values (139). TP improves albumen quality by upregulating the genes associated with cell proliferation, metal-binding mediation, and immune function-related proteins (80). Further studies are needed on the upregulation of proteins that enhance albumen quality due to dietary effects. It is imperative to investigate whether these green additives’ antioxidant and antimicrobial capacity could maintain the oxidative balance in stored eggs, thereby extending the shelf life of the eggs and ensuring the safety of consumer eggs.
The inclusion of organic and inorganic forms of trace elements (zinc, iron, and selenium), minerals (clay-chelates), organic acids, and vitamins in the diets of laying hens have been reported to influence albumen quality differently. Trace elements and inorganic minerals like oxides and sulfates sources (14, 140) traditionally supplemented in feeds of laying hens are presently replaced with organic microelements, especially mineral amino acid chelates and complexes due to their roles in egg quality (140). Challenges such as excretion into the environment, low bioavailability, high oxidation and destruction of nutrients (141) limits the use of the inorganic forms which also produce free radicals that could adversely affect animal health (14). Preference for the organic mineral form is promoted due to ease of absorption, high biological value, environmental friendliness, non-toxicity and safety (140). The study by Yu et al. (22) reported no significant difference between organic and inorganic zinc on egg quality. Organic forms of trace elements include selenium yeast (142), zinc-methionine (140) and iron-glycine chelate (143), which are more metabolized than inorganic forms. This suggests higher bioavailability of the organic form compared to inorganic form, such bio-fortification with microelements improves eggs’ nutritive value and biological function.
Selenium is a trace element necessary for animal and human health. Selenium have been reported to enhance reproductive performance, antioxidant and immunomodulatory function in laying hens (144, 145). In same vein, biofortification of eggs with selenium, evidenced by high content of selenium in albumen have been explored (145–149). Other evidences showed that (Se-enriched insect protein) (150) and selenium enriched yeast (148, 151), enhanced albumen height and HU. Although selenium improved albumen indices (152), no significant difference in egg quality traits was found between the Se-diet and the control group in other studies (146, 148, 153). The improvement in albumen quality may be linked with; more absorbable form of organic selenium which facilitates ease of utilization in laying hens, positive effects of selenium on oviduct health and activities of antioxidant enzymes, which may boost protein metabolism and utilization. The variations could be selenium source and dosage level. There is limited information of influence of organic selenium on albumen quality of fresh eggs.
Organic forms of zinc have been demonstrated to influence albumen quality. Qi et al. (154) reported that zinc-methionine enhanced albumen quality via improved T-AOC, decreased MDA level, promoted methionine synthesis, threonine and glutathione metabolism and protein metabolism. Inclusion of zinc-methionine (Zn-Met) complexes (140, 155) and zinc-methionine hydroxyl analog chelate (156) enhanced albumen HU values. (157) reported that Zn-Met improved both albumen percentage and HU. Also, Zn-Met mitigated the adverse effect of increasing age on HU values due to watery albumen (155). Besides, improving albumen quality, nutritive contents of eggs are improved via biofortification of eggs with increased zinc content (154, 158). The improvement in albumen quality due to dietary zinc may be accrued to the antioxidant nature of zinc and its role in metabolic process.
The liver is the primary site of zinc bioaccumulation in chickens, and it also plays a role in energy metabolism. Zinc is not an antioxidant, but it activates antioxidant enzymes and competes with redox-active transition metals such as copper and iron for binding sites. Zinc is crucial to the structure and function of Cu/Zn-SOD, accounting for about 90% of total SOD, which exerts protective effects on tissues from oxidative damage (159). In another study (160), found that dietary zinc increased metallothionein production, which inhibited lipid peroxidation, reduced MDA production, and increased Cu/Zn –SOD levels. Zinc plays an important role in protein synthesis and other biochemical reactions. (161) reported that dietary zinc regulates intestinal amino acid and protein metabolism in animals. Organic zinc has been reported to enhance total protein and globulin, serum urea nitrogen, indicating enhanced protein catabolism. Further, zinc plays a positive role in the magnum during deposition of albumen and in the isthmus during the formation of shell membrane (162). It could be deduced that the beneficial effects of zinc on intestinal integrity, protein metabolism and catabolism, antioxidant system and oviduct health may account for its positive influence on albumen quality.
Iron is one of the most critical elements for poultry and participates in various metabolic processes, including transport and storage of oxygen, protein metabolism, antioxidant and immune activity (163). Inorganic iron, FeSO4, is mostly used in poultry but has low absorption, pollution, and low biological value. The use of organic iron had better results due to its high bioavailability. For example, supplementation of an amino acid complexed iron (Fe-Gly) at inclusion level of 80 mg/kg in diet of laying hens, enhanced albumen quality compared to inorganic Fe (143). The result was premised on the organic iron’s higher absorption potential and easy transportation within the body, which enhances its metabolism.
In same line, organic minerals such as dietary carbo-amino-phospho-chelates in comparison to the sulphate form (inorganic) enhanced HU values (90.2 vs. 84.95) and other albumen indices in laying hens (164). Calcium in laying pigeons promoted albumen transparency (165). Dietary montmorillonite (Mineral clay) improved albumen height and HU value (166). These mineral clays can bind to pathogens and toxins in the GIT, create favorable gut microecological environment that can lead to improved performance, health status and egg quality. Glycerol monolaurate (GML), a monoglyceride of medium chain fatty acids, enhanced flavor amino acids content in the albumen (65), thereby improving the nutritive value of egg. In contrast, calcium montmorillonite (167) and amino acid-complexed manganese (168) had no significant effect on HU values and albumen quality. The variations may be due to nature of organic element or mineral used and inclusion levels in the diets.
Organic acids such as benzoic acid enhanced albumen height and HU values (169), when supplemented in diet of laying hens. In one study, supplementation of inorganic dietary fluoride at 800 and 1200 mg/kg drastically reduced albumen height and HU values, probably because of the adverse effect on the ovary and liver, leading to oxidative stress and impaired protein synthesis (14). Various studies have shown that some products extracted from plants and animals, used as feed additives influence albumen quality (74, 170, 171). Natural products including; octacosanol (171), resveratrol (170), magnolol (74), and quercetin (172) enhanced albumen quality whereas natural astaxanthin (68, 173) and antimicrobial peptide; cecropin (16) did not affect albumen quality. Age of laying hens and inclusion levels may account for inconsistencies in results among various studies. In addition, vitamins influence albumen quality; vitamin A enhanced albumen quality during heat stress via improved reproductive tract development (15), vitamin E (152) and 25-hydroxyvitamin D3 (174, 175), enhanced albumen HU whereas others found non-significant differences between treated groups (25-hydroxyvitamin D3) and the control (176). The improvement due to dietary vitamins could be that vitamins ensures utilization of proteins and energy by animals for health enhancement and reproduction (177).
The efficiency of protein utilization in diets depends on amino acid content, composition, and digestibility, hence, the deposition of albumen and yolk relies on nutrient supply (amino acids and fatty acids). Adequate essential amino acids (AA) has been traced to enhance internal and external egg quality (73), thus it is critical to supplement layers diets with AA, which cannot be synthesized in the body of laying hens. Such supplementation is critical in maintaining the structural integrity and function of the gut, decrease intestinal dysfunction and mitigate oxidative stress (178), which could lead to improved nutrient absorption and albumen synthesis. Also, egg proteins are made of AA, and the concentration of total, essential and flavor AA plays an important role in egg nutritive value and flavor. Therefore, the albumen AA profile could influence its nutritive value. Protein synthesis in the magnum may be influenced by amino acid concentration in the blood (33). Change in the level of AA supplementation in diets may alter protein synthesis resulting in a positive or negative effect on albumen quality. Liu et al. (179) reported that low methionine levels decreased the albumen ratio. In sum, amino acid plays a great role in the body’s metabolic functions, translating to an improved animal product.
Amino acid supplementation in the diets of laying hens has varying effects on albumen quality. For instance, dietary L-arginine (73), dietary L-carnitine (180), dietary threonine in the diet of old hens (181), and 0.05% purified amino acid extracted from animal blood (17) improved albumen height and HU value. Similarly, lysine supplementation enhanced albumen percentage (182). Also, dietary threonine supplemented in a dose-dependent manner increased albumen weight and percentage (183). Improved albumen indices with AA supplementation may be due to increased nutrient concentration and protein levels in the diet (184), which enhance protein synthesis (185). This, in turn, promotes the metabolic rate in the magnum, increases the activity of the shell gland, and produces eggshells with enhanced integrity, reducing the loss of carbon dioxide and improving albumen quality. In another study, threonine improved albumen percentage in a dose-dependent manner via upregulation of genes related to AA transport and protein deposition (183). In another study, (34), suggested that supplementing diets that enhance upregulation of AA transporters would enhance albumen synthesis and quality. On the other hand, dietary threonine (186), total sulfur amino acid (178), D-lysine (187), amino acid-complexed manganese (168) and L-citrulline, a non-protein AA (188) did not affect albumen quality. However, decreasing albumen indices with dietary AA supplementation may be due to decreased albumen synthesis (33) and the feeding regime of the AA (179). The varying results may be related to the AA inclusion level, denaturation by high temperature, and source. It has been reported that the requirements for certain essential AA for laying hens increase with an increase in the dietary level of protein (189).
Protein synthesis, which facilitates egg formation, may depend on protein utilization. The study by Kowalska et al. (190), Kuzniacka et al. (191) demonstrated that eggs of hens fed diets from different protein sources with high crude protein levels had increased albumen height, thick albumen content and HU values comparable to sources with lower crude protein level. Increased dietary crude protein (CP) level improved albumen weight, egg weight and HU value (192). Lowering CP levels from 16.49 to 14.05% (193) and from 16 to 13% (194) caused a lower HU value of 89.50 vs. 86.74 and 83.95 vs. 76.82, respectively, and invariably poor albumen quality. However, (195) reported no dietary protein levels influence on HU values. Decreased albumen indices in birds fed different dietary protein sources at different levels may be due to low dietary protein intake. (33) reported that low dietary protein intake reduces egg mass, albumen weight, and albumen solids because of the reduction in albumen secretion. Another report by (28, 196) suggests that reduction in albumen height and HU values was due to low dietary protein intake and not gossypol content of the feed. Furthermore, low albumen quality may be linked to low levels of ovomucin, a major component of albumen. The albumen ovomucin content was reduced in birds given various protein sources and levels (28). It is speculated that supplementing diets low in CP with AA could enhance albumen quality. For instance isoleucine in a low CP diet improved albumen quality (197) while threonine improved egg quality in birds fed diets with low CP levels via; enhanced secretion of digestive enzymes and increase in population of beneficial gut microflora (198). Therefore, diets from protein sources that can positively influence albumen quality via enhanced magnum development, increased protein synthesis, and albumen secretion should be adopted.
Reducing feed cost is the utmost priority of poultry farmers. Soybean meal (SBM) which is the conventional source of plant protein in most layers diet is not only expensive but has highly competitive value as food by man and other Agro-allied industries. The best strategy to decrease feed cost and competition for SBM as protein source is to formulate diets based on local alternative feedstuffs which are readily available.
Replacing SBM with 100 g/kg cotton seed meal (CSM) in layers diet enhanced albumen indices (34, 196). However, camelina or flaxseed (199), increased levels of sunflower meal (200), low gossypol cottonseed meal (LCSM), or double-zero rapeseed meal (DRM) used as the sole replacement for SBM in a layer diet (34, 35) decreased albumen height and HU values. Also, CSM at 100 g/kg reduced albumen weight, Haugh unit and albumen height (13). The reduction in albumen quality may be due to decreased tubular gland cells and epithelial growth in the oviduct magnum, slowing down protein synthesis (13). This is because synthesis of albumen proteins often occurs in the oviduct (201). However, some of the alternative feedstuffs used as a replacement for SBM such as CSM (202, 203), untreated field peas (204), wheat bran supplementation (205), full-fat flaxseed and sunflower seeds (206) had no significant effect on albumen indices and HU value. Evaluating albumen quality based on HU and albumen height may not provide enough evidence of dietary effects; investigation of albumen proteins could provide further insight into dietary effects. The study by He et al. (13) used a proteomic technique to show that albumen proteins (ovalbumin, ovotransferrin, ovomucin, lysozyme, ovoinhibitor, and clusterin) decreased in birds fed CSM at 100 g/kg. The decreased content of lysozyme and ovomucin altered the stability of the ovomucin-lysozyme complex thereby reducing albumen viscosity. Given that the albumen proteins are fundamental to albumen’s functional and technological properties, more knowledge about the protein content is needed to reach conclusions about albumen quality.
The reduction or non- significant effect on albumen indices may be due to free gossypol (FG) content of the unconventional feedstuffs. For instance, Wang et al. (34) reported that 195 g/kg low gossypol cottonseed meal impaired magnum development and function; >70 mg/kg of gossypol caused inflammatory response and damaged the mucosal immune system (203). This indicates that the gossypol level in laying hens’ diets should not exceed the above thresholds. Other evidences reported no adverse effect of FG on albumen quality and HU value (203, 207). Notably, such adverse effects could be due to the capacity of FG to inhibit several enzymes activities in the digestive tract, causing dyspepsia and growth retardation in laying birds (208). The condition also suppresses gastric secretion leading to distension of the abdomen and negative growth performance, causing clinical poisoning and liver damage (209). Alternative feedstuffs may also be low in amino acids that are crucial to protein metabolism and albumen synthesis. There are evidences that feedstuffs deficient in certain digestible AAs (arginine, phenylalanine, histidine, and leucine) (33), and indispensable amino acids (lysine, threonine, and sulfur AA) (210). Also some of alternative feedstuffs may contain antinutritional factors (i.e., phytic acid and tannin, trypsin inhibitor, vicine, and convicine) (211). Taken together, utilization of alternative feedstuffs for protein sources are limited by presence of antinutritional factors, low amino acid content and inclusion levels, which could impair nutrient absorption in the intestine, albumen secretion dimensions in the magnum and deposition of ovomucin in the albumen in vivo. Utilization of alternative feedstuffs may be enhanced by addition of enzymes.
Recently, the effects of these feedstuffs supplemented with exogenous enzymes have been reported (212, 213). The hypothesis developed was that the enzymes would mask the feedstuff’s antinutrients. Dietary supplementation of < 25% faba beans with enzyme (212), xylanase at 900 U/kg supplemented to corn-soybean-meal-wheat based diets (213), and 1% of non-starch polysaccharide multi-enzyme (214) enhanced albumen quality. The improvement may be due to capacity of the enzymes to unbound nutrients and make it available for utilization by animals. In contrast, exogenous xylanase at 12000 BXU/kg supplemented to non-soluble polysaccharide diets (215), xylanase at 200 mg/kg supplemented to brasetto hybrid rye diet (216), and multi-carbohydrase at 200 mg/kg supplemented to wheat-based diet (217) did not affect albumen quality. The variations may be due to feedstuff content, inclusion levels of the enzymes and type of enzyme used. Fermentation process using microorganisms may be a better strategy to degrade antinutritional factors and increase the protein content of alternative feedstuffs, thus increasing its utilization. Fermented feeds improved albumen height and HU in laying hens, probably due to favorable microbial environment which may enhance protein synthesis (218). The use of insect meal may act as a replacement of SBM as source of protein. The study of demonstrated that Black soldier larvae meal enhanced albumen weight and HU value (219). The use of readily available non-conventional feedstuffs of high biological value is most often advocated to reduce feed costs.
Inclusion of natural essential oils of plant origin in birds’ diets has been reported to influence egg quality due to their antioxidant property (220). The peppermint oil (126), peppermint oil alone or combined with thyme oil (221), fish oil (222), and essential oils (223–225), were found to improve albumen indices and HU value. The antioxidant capacity of the oils conferred by vitamin E on the plants could explain the cumulative effect of oil from diverse sources on albumen quality (226). The antioxidant compounds in the natural oil can be transferred to the egg conferring on them higher antioxidant capacity (21), that can sustain albumen quality. Natural oil from varying sources contain phenolic compounds that acts as hydrogen donors, retarding hydrogen peroxides from generating free radicals that may alter albumen synthesis. For example, oil from grape seeds improved albumen height by reducing level of oxidative products and enhancing β-ovomucin content (127). Antibacterial action of phenolic compounds such as thymol and carvacrol, may increase population of beneficial gut microbes while suppressing pathogenic bacteria (221). This would help physiological mechanisms that boost protein synthesis, by reducing competition for nutrients between the host and its microflora. In same vein, essential oils have been found to improve gut villi morphology and reproductive tract, increase secretion of digestive enzymes, leading to enhanced digestion of protein and fat and absorption in the intestine (129, 223, 227), with consequent improvement in albumen quality. It could be deduced that the improvement in albumen quality due to dietary oil, could probably be its positive effect on uterine health, oviposition process, increased pancreatic secretions, improved gut villi structures, enhanced nutrient digestion and utilization. However, camelina and flaxseed oil (199) and, soybean and linseed oil (228) reduced albumen weight but did not affect the HU values and yolk to albumen ratio. In addition, wheat bran supplemented with oil (205), combined encapsulated essential oil and organic acid (224), glycerol oil (229), essential oils (220, 227) and Lavandula angustifolia and/or Mentha spicata essential oils (194) did not exert any effect on egg weight, HU, and albumen height. The reduction in albumen quality due to natural oil supplementation may be due to anti-nutritional factors in the seeds used for oil extraction. For example, trypsin inhibitors can impair protein digestibility and albumen secretion (230). Also, high concentrations of some bioactive compounds in oils such as thymol or carvacrol may negatively influence intestinal functions and alter composition of beneficial gut microbes (231), thus harmful to the birds and invariably cause a decline in egg quality. The varying effects of oil supplementation could be due to oil sources, duration of feeding, level of supplementation and presence of antinutritional factors. Therefore, exploring oil as natural antioxidants in maintaining albumen quality of fresh and stored eggs, would be a promising research direction for improving animal product quality.
In summary, improving the albumen quality of laying hens fed natural feed ingredients depends on the underlying mechanism of upregulation and downregulation of oxidative products by antioxidant enzymes (Figure 4). Natural products that improve animal health would result in better albumen quality without transferring toxins to the eggs. Table 1 lists the bioactive compounds in selected natural plants and products. The bioactive compounds of these products mostly from plants, have antioxidant, antimicrobial, anti-inflammatory properties which accounts for the positive effect of natural products on albumen quality.

Figure 4. Mechanism of action of diets on albumen quality.
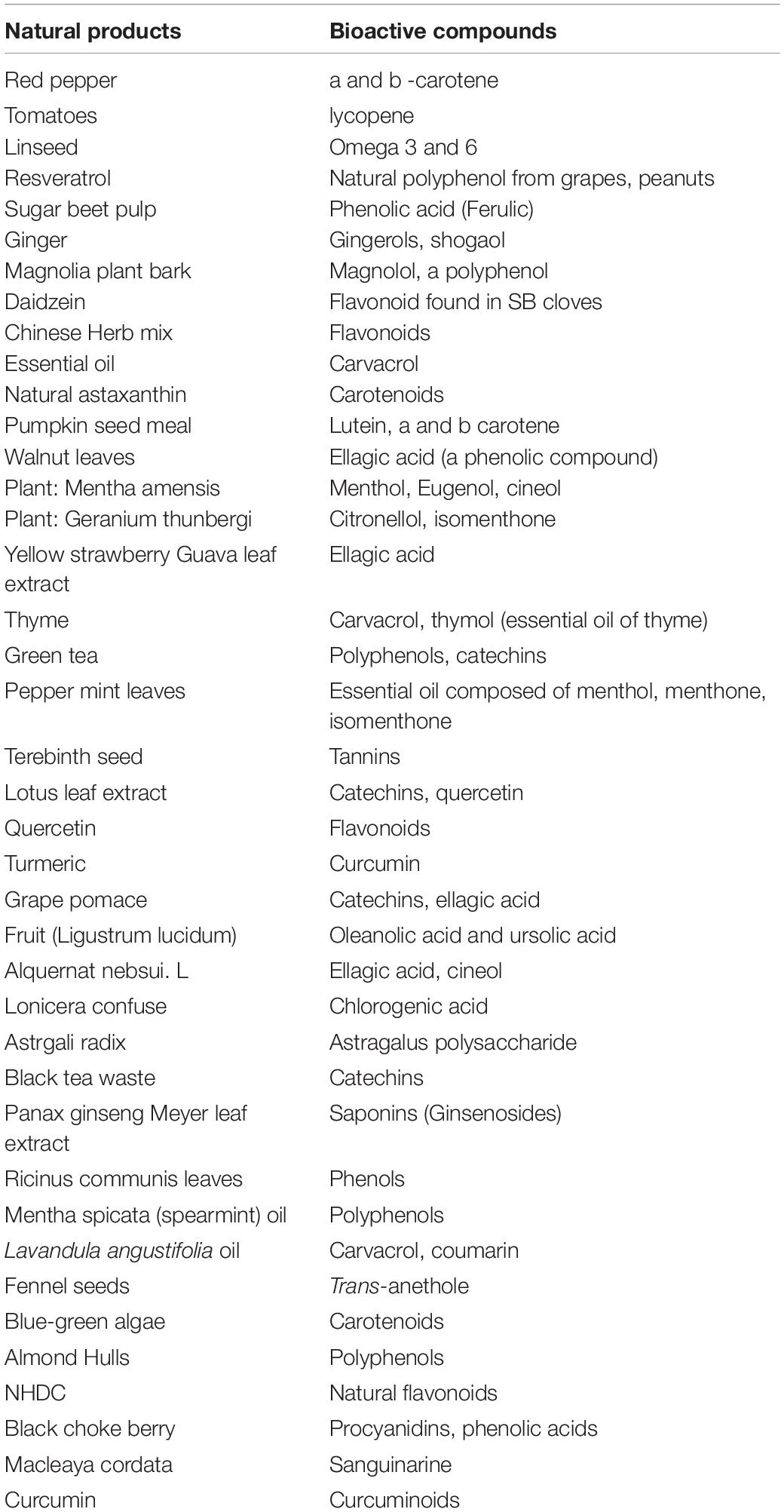
Table 1. Plant natural products and their bioactive compounds.
Albumen quality is of paramount importance to consumers, producers, breeders, and the food industry because of its biological and functional properties. In previous decades, research has focused on the performance and health of chickens, with less attention to maintaining albumen quality through nutritional modules. Various diets have been demonstrated to influence albumen quality, providing novel nutritional strategies to preserve the albumen’s integrity and the ensuing by-products. We recommend conducting similar dietary studies on albumen quality during storage to determine which diets can preserve albumen quality and reduce egg spoilage rate over time. Specifically, proteomic studies should be used to investigate the effect of diets on metabolic pathways, microbiota populations, and the proteins involved in maintaining albumen quality of fresh and stored eggs.
KQ and S-GW: conceptualization. KQ and UO: resources data, writing, and editing. UO and VO-O: writing—original draft. KQ, JW, H-JZ, and S-GW: supervision. S-GW and G-HQ: funding. All authors contributed to the reviewed and approved final version of the manuscript.
This study was supported by the National Natural Science Foundation of China (32072774) and the Agricultural Science and Technology Innovation Program (ASTIP) of the Chinese Academy of Agricultural Sciences (CAAS).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnut.2022.875270/full#supplementary-material
1. Réhault-Godbert S, Guyot N, Nys Y. The golden egg: nutritional value, bioactivities, and emerging benefits for human health. Nutrients. (2019) 11:684. doi: 10.3390/nu11030684
2. Nimalaratne C, Wu J. Hen egg as an antioxidant food commodity: a review. Nutrients. (2015) 7:8274–93. doi: 10.3390/nu7105394
3. Omana DA, Wang J, Wu J. Ovomucin–a glycoprotein with promising potential. Trends Food Sci Technol. (2010) 21:455–63. doi: 10.1016/j.tifs.2010.07.001
4. Wang XC, Wang XH, Wang J, Wang H, Zhang HJ, Wu SG, et al. Dietary tea polyphenol supplementation improved egg production performance, albumen quality, and magnum morphology of Hy-Line Brown hens during the late laying period. J Anim Sci. (2018) 96:225–35. doi: 10.1093/jas/skx007
5. Zhang J, Zhang M, Liang W, Geng Z, Chen X. Green tea powder supplementation increased viscosity and decreased lysozyme activity of egg white during storage of eggs from Huainan partridge chicken. Ital J Anim Sci. (2020) 19:586–92.
6. Abeyrathne E, Lee H, Ahn DU. Egg white proteins and their potential use in food processing or as nutraceutical and pharmaceutical agents—a review. Poult Sci. (2013) 92:3292–9. doi: 10.3382/ps.2013-03391
7. Kato A, Ogato S, Matsudomi N, Kobayashi K. Comparative study of aggregated and disaggregated ovomucin during egg white thinning. J Agric Food Chem. (1981) 29:821–3. doi: 10.1021/jf00106a035
8. Toussant MJ, Latshaw JD. Ovomucin content and composition in chicken eggs with different interior quality. J Sci Food Agric. (1999) 79:1666–70.
9. Wang Y, Wang Z, Shan Y. Assessment of the relationship between ovomucin and albumen quality of shell eggs during storage. Poult Sci. (2019) 98:473–9. doi: 10.3382/ps/pey349
10. Yuan ZH, Zhang KY, Ding XM, Luo YH, Bai SP, Zeng QF, et al. Effect of tea polyphenols on production performance, egg quality, and hepatic antioxidant status of laying hens in vanadium-containing diets. Poult Sci. (2016) 95:1709–17. doi: 10.3382/ps/pew097
11. Perić L, Đukić Stojčić M, Bjedov S. The effect of storage and age of hens on the quality of table eggs. Adv Res Life Sci. (2017) 1:64–7.
12. Marzec A, Damaziak K, Kowalska H, Riedel J, Michalczuk M, Koczywąs E, et al. Effect of hens age and storage time on functional and physiochemical properties of eggs. J Appl Poul Res. (2019) 28:290–300.
13. He T, Zhang H, Wang J, Wu S, Yue H, Qi G. Proteomic comparison by iTRAQ combined with mass spectrometry of egg white proteins in laying hens (Gallus gallus) fed with soybean meal and cottonseed meal. PLoS One. (2017) 12:e0182886. doi: 10.1371/journal.pone.0182886
14. Miao LP, Li LL, Zhu MK, Dong XY, Elwan HAM, Zou XT. Excess dietary fluoride affects laying performance, egg quality, tissue retention, serum biochemical indices, and reproductive hormones of laying hens. Poult Sci. (2019) 98:6873–9. doi: 10.3382/ps/pez443
15. Abd El-Hack ME, Alagawany M, Mahrose KM, Arif M, Saeed M, Arain MA, et al. Productive performance, egg quality, hematological parameters and serum chemistry of laying hens fed diets supplemented with certain fat-soluble vitamins, individually or combined, during summer season. J Anim Nutr. (2019) 5:49–55. doi: 10.1016/j.aninu.2018.04.008
16. Chen X, Zhan Y, Ma W, Zhu Y, Wang Z. Effects of antimicrobial peptides on egg production, egg quality and caecal microbiota of hens during the late laying period. Anim Sci J. (2020) 91:e13387. doi: 10.1111/asj.13387
17. Joshi N, Wandita T, Yang S, Park H, Hwang S. Effects of supplementing laying hens with purified amino acid prepared from animal blood. Trop Anim Sci J. (2019) 42:46–52.
18. Chang X, Qiu K, Wang J, Zhang H, You S, Mi S, et al. The evaluation of UPro as a new nutrient on high-quality egg production from the perspective of egg properties, intestinal histomorphology, and oviduct function of laying hens. Front Nutr. (2021) 8:706067. doi: 10.3389/fnut.2021.706067
19. Hussein AS, Ayoub MA, Elhwetiy AY, Ghurair JA, Sulaiman M, Habib HM. Effect of dietary inclusion of sugar syrup on production performance, egg quality and blood biochemical parameters in laying hens. Anim Nutr. (2018) 4:59–64. doi: 10.1016/j.aninu.2017.11.001
20. Neijat M, Shirley RB, Barton J, Thiery P, Welsher A, Kiarie E. Effect of dietary supplementation of Bacillus subtilis DSM29784 on hen performance, egg quality indices, and apparent retention of dietary components in laying hens from 19 to 48 weeks of age. Poult Sci. (2019) 98:5622–35. doi: 10.3382/ps/pez324
21. Migliorini MJ, Boiago MM, Stefani LM, Zampar A, Roza LF, Barreta M, et al. Oregano essential oil in the diet of laying hens in winter reduces lipid peroxidation in yolks and increases shelf life in eggs. J Therm Biol. (2019) 85:102409. doi: 10.1016/j.jtherbio.2019.102409
22. Yu Q, Liu H, Yang K, Tang X, Chen S, Ajuwon KM, et al. Effect of the level and source of supplementary dietary zinc on egg production, quality, and zinc content and on serum antioxidant parameters and zinc concentration in laying hens. Poult Sci. (2020) 99:6233–8. doi: 10.1016/j.psj.2020.06.029
23. Alleoni ACC. Albumen protein and functional properties of gelation and foaming. Sci Agric. (2006) 63:291–8.
25. Kovacs-Nolan J, Phillips M, Mine Y. Advances in the value of eggs and egg components for human health. J Agric Food Chem. (2005) 53:8421–31. doi: 10.1021/jf050964f
26. Offengenden M, Fentabil MA, Wu J. N-glycosylation of ovomucin from hen egg white. Glycoconj J. (2011) 28:113–23. doi: 10.1007/s10719-011-9328-3
27. Shan Y, Tang D, Wang R, Tu A, Yi Y, Wang X, et al. Rheological and structural properties of ovomucin from chicken eggs with different interior quality. Food Hydrocoll. (2020) 100:105393.
28. Wang X, Zhang H, Wu S, Yue H, Wang J, Li J, et al. Dietary protein sources affect internal quality of raw and cooked shell eggs under refrigerated conditions. Asian Australas J Anim Sci. (2015) 28:1641. doi: 10.5713/ajas.15.0181
29. Hiidenhovi J. Isolation and Characterization of Ovomucin-a Bioactive Agent of Egg White. Doctoral thesis. Turku: Food Chemistry and Food Development, Department of Biochemistry (2015).
30. Aobai T, Xue Z, Yuanyuan S, Xin L. Potential role of ovomucin and its peptides in modulation of intestinal health: a review. Int J Biol Macromol. (2020) 162:385–93. doi: 10.1016/j.ijbiomac.2020.06.148
31. Bravo L. Polyphenols: chemistry, dietary sources, metabolism, and nutritional significance. Nutr Rev. (1998) 56:317–33. doi: 10.1111/j.1753-4887.1998.tb01670.x
32. Wang X, Wu S, Cui Y, Qi G, Wang J, Zhang H. Effects of dietary tea polyphenols on performance, egg quality and antioxidant ability of laying hens. Chin J Anim Nutr. (2017) 29:193–201.
33. Novak C, Yakout H, Scheideler S. The effect of dietary protein level and total sulfur amino acid: lysine ratio on egg production parameters and egg yield in Hy-Line W-98 hens. Poult Sci. (2006) 85:2195–206. doi: 10.1093/ps/85.12.2195
34. Wang XC, Zhang HJ, Wang H, Yue HY, Wang J, Wu SG, et al. Effect of different protein ingredients on performance, egg quality, organ health, and jejunum morphology of laying hens. Poult Sci. (2017) 96:1316–24. doi: 10.3382/ps/pew396
35. Wang X, Wu S, Zhang H, Yue H, Qi G, Li J. Effect of dietary protein sources and storage temperatures on egg internal quality of stored shell eggs. Anim Nutr. (2015) 1:299–304. doi: 10.1016/j.aninu.2015.12.003
37. USDA.United States Standards, Grades, and Weight Classes for Shell Eggs, Ams 56. Washington, DC: USDA (2000).
38. Erdmann K, Cheung BW, Schröder H. The possible roles of food-derived bioactive peptides in reducing the risk of cardiovascular disease. J Nutr Biochem. (2008) 19:643–54. doi: 10.1016/j.jnutbio.2007.11.010
39. Abeyrathne E, Huang X, Ahn DU. Antioxidant, angiotensin-converting enzyme inhibitory activity and other functional properties of egg white proteins and their derived peptides–a review. Poult Sci. (2018) 97:1462–8. doi: 10.3382/ps/pex399
40. Manso MA, Miguel M, Even J, Hernández R, Aleixandre A, López-Fandiño R. Effect of the long-term intake of an egg white hydrolysate on the oxidative status and blood lipid profile of spontaneously hypertensive rats. Food Chem. (2008) 109:361–7. doi: 10.1016/j.foodchem.2007.12.049
41. Rupa P, Schnarr L, Mine Y. Effect of heat denaturation of egg white proteins ovalbumin and ovomucoid on CD4+ T cell cytokine production and human mast cell histamine production. J Funct Foods. (2015) 18:28–34.
42. Sun X, Ganzle M, Field CJ, Wu J. Effect of proteolysis on the sialic acid content and bifidogenic activity of ovomucin hydrolysates. Food Chem. (2016) 212:78–86. doi: 10.1016/j.foodchem.2016.05.153
43. Garcés-Rimón M, González C, Hernanz R, Herradón E, Martín A, Palacios R, et al. Egg white hydrolysates improve vascular damage in obese Zucker rats by its antioxidant properties. J Food Biochem. (2019) 43:e13062. doi: 10.1111/jfbc.13062
44. Chen C, Chi Y-J, Zhao M-Y, Lv L. Purification and identification of antioxidant peptides from egg white protein hydrolysate. Amino Acids. (2012) 43:457–66. doi: 10.1007/s00726-011-1102-0
45. Davalos A, Miguel M, Bartolome B, Lopez-Fandino R. Antioxidant activity of peptides derived from egg white proteins by enzymatic hydrolysis. J Food Prot. (2004) 67:1939–44. doi: 10.4315/0362-028x-67.9.1939
46. Fujita H, Yamagami T, Ohshima K. Effects of an ACE-inhibitory agent, katsuobushi oligopeptide, in the spontaneously hypertensive rat and in borderline and mildly hypertensive subjects. Nutr Res. (2001) 21: 1149–58.
47. Chiang W, Lee M, Guo W, Tsai T. Protein hydrolysate batch production with angiotensin I-converting enzyme inhibitory activity from egg whites. J Food Drug Anal. (2006) 14:385.
48. Rao S, Sun J, Liu Y, Zeng H, Su Y, Yang Y. ACE inhibitory peptides and antioxidant peptides derived from in vitro digestion hydrolysate of hen egg white lysozyme. Food Chem. (2012) 135:1245–52. doi: 10.1016/j.foodchem.2012.05.059
49. Oguro T, Watanabe K, Tani H, Ohishi H, Ebina T. Morphological observations on antitumor activities of 70 kDa fragment in α-subunit from pronase-treated ovomucin in a double grafted tumor system. Food Sci Technol Res. (2000) 6:179–85.
50. Vidovic D, Graddis T, Chen F, Slagle P, Diegel M, Stepan L, et al. Antitumor vaccination with HER-2-derived recombinant antigens. Int J Cancer. (2002) 102:660–4. doi: 10.1002/ijc.10785
51. Ibrahim HR, Kiyono T. Novel anticancer activity of the autocleaved ovotransferrin against human colon and breast cancer cells. J Agric Food Chem. (2009) 57:11383–90. doi: 10.1021/jf902638e
52. Ko K, Ahn DU. An economic and simple purification procedure for the large-scale production of ovotransferrin from egg white. Poult Sci. (2008) 87:1441–50. doi: 10.3382/ps.2007-00434
53. Galla R, Grisenti P, Farghali M, Saccuman L, Ferraboschi P, Uberti F. Ovotransferrin supplementation improves the iron absorption: an in vitro gastro-intestinal model. Biomedicines. (2021) 9:1543. doi: 10.3390/biomedicines9111543
54. Pellegrini A, Thomas U, Wild P, Schraner E, Von Fellenberg R. Effect of lysozyme or modified lysozyme fragments on DNA and RNA synthesis and membrane permeability of Escherichia coli. Microbiol Res. (2000) 155:69–77.
55. Karamanidou A, Kallithraka S, Hatzidimitriou E. Fining of red wines: effects on their analytical and sensory parameters. J Int Sci. (2011) 45:47–60.
56. Rizzetti DA, Martín Á, Corrales P, Fernandez F, Simões MR, Peçanha FM, et al. Egg white-derived peptides prevent cardiovascular disorders induced by mercury in rats: role of angiotensin-converting enzyme (ACE) and NADPH oxidase. Toxicol Lett. (2017) 281:158–74. doi: 10.1016/j.toxlet.2017.10.001
57. Vilcacundo R, Méndez P, Reyes W, Romero H, Pinto A, Carrillo W. Antibacterial activity of hen egg white lysozyme denatured by thermal and chemical treatments. Sci Pharm. (2018) 86:48. doi: 10.3390/scipharm86040048
59. Valenti P, Antonini G, Von Hunolstein C, Visca P, Orsi N, Antonini E. Studies of the antimicrobial activity of ovotransferrin. Int J Tissue React. (1983) 5:97–105.
60. Ibrahim HR, Sugimoto Y, Aoki T. Ovotransferrin antimicrobial peptide (OTAP-92) kills bacteria through a membrane damage mechanism. Biochim Biophys Acta (BBA) Gen Subj. (2000) 1523:196–205. doi: 10.1016/s0304-4165(00)00122-7
61. Ko K, Mendonca AF, Ahn DU. Influence of zinc, sodium bicarbonate, and citric acid on the antibacterial activity of ovotransferrin against Escherichia coli O157: H7 and Listeria monocytogenes in model systems and ham. Poult Sci. (2008) 87:2660–70. doi: 10.3382/ps.2007-00503
62. Cegielska-Radziejewska R, Lesnierowski G, Kijowski J. Antibacterial activity of hen egg white lysozyme modified by thermochemical technique. Eur Food Res Technol. (2009) 228:841–5.
63. Wang Z, Wang D, Jiang K, Guo Y, Li Z, Jiang R, et al. A comprehensive proteome and acetyl-proteome atlas reveals molecular mechanisms adapting to the physiological changes from pre-laying to peak-laying stage in liver of hens (Gallus gallus). Front Vet Sci. (2021) 8:700669. doi: 10.3389/fvets.2021.700669
64. Assersohn K, Brekke P, Hemmings N. Physiological factors influencing female fertility in birds. R Soc Open Sci. (2021) 8:202274. doi: 10.1098/rsos.202274
65. Liu T, Li C, Li Y, Feng F. Glycerol Monolaurate enhances reproductive performance, egg quality and albumen amino acids composition in aged hens with gut microbiota alternation. Agriculture. (2020) 10:250.
66. Khochamit N, Duangjinda M, Siripornadulsil S, Wongtangtintharn S, Siripornadulsil W. Effects of dried yeast, a byproduct of the brewery industry, on the egg production and quality and the immune response of laying hens. Ital J Anim Sci. (2021) 20:1135–46.
67. Sharma MK, Dinh T, Adhikari PA. Production performance, egg quality, and small intestine histomorphology of the laying hens supplemented with phytogenic feed additive. J Appl Poult Res. (2020) 29:362–71.
68. Heng N, Gao S, Guo Y, Chen Y, Wang L, Sheng X, et al. Effects of supplementing natural astaxanthin from Haematococcus pluvialis to laying hens on egg quality during storage at 4° C and 25° C. Poult Sci. (2020) 99:6877–83. doi: 10.1016/j.psj.2020.09.010
69. Muhammad AI, Mohamed DAA, Chwen LT, Akit H, Samsudin AA. Effect of sodium selenite, selenium yeast, and bacterial enriched protein on chicken egg yolk color, antioxidant profiles, and oxidative stability. Foods. (2021) 10:871. doi: 10.3390/foods10040871
70. Zheng M, Mao P, Tian X, Guo Q, Meng L. Effects of dietary supplementation of alfalfa meal on growth performance, carcass characteristics, meat and egg quality, and intestinal microbiota in Beijing-you chicken. Poult Sci. (2019) 98:2250–9. doi: 10.3382/ps/pey550
71. Zhu YF, Wang JP, Ding XM, Bai SP, Qi SRN, Zeng QF, et al. Effect of different tea polyphenol products on egg production performance, egg quality and antioxidative status of laying hens. Anim Feed Sci Technol. (2020) 267:114544.
72. Ding X, Du J, Zhang K, Bai S, Zeng Q, Peng H, et al. Tandem mass tag-based quantitative proteomics analysis and gelling properties in egg albumen of laying hens feeding tea polyphenols. Poult Sci. (2020) 99:430–40. doi: 10.3382/ps/pez523
73. Fascina VB, Pasquali GAM, Berto DA, Silva AdL, Garcia EA, Pezzato AC, et al. Effects of arginine and phytogenic additive supplementation on performance and health of brown-egg layers. Rev Bras Zootecnia. (2017) 46:502–14.
74. Chen F, Zhang H, Du E, Jin F, Zheng C, Fan Q, et al. Effects of magnolol on egg production, egg quality, antioxidant capacity, and intestinal health of laying hens in the late phase of the laying cycle. Poult Sci. (2021) 100:835–43. doi: 10.1016/j.psj.2020.10.047
75. Selim S, Hussein E. Production performance, egg quality, blood biochemical constituents, egg yolk lipid profile and lipid peroxidation of laying hens fed sugar beet pulp. Food Chem. (2020) 310:125864. doi: 10.1016/j.foodchem.2019.125864
76. Bozkurt M, Bintas E, Kirkan S, Aksit H, Kucukyilmaz K, Erbas G, et al. Comparative evaluation of dietary supplementation with mannan oligosaccharide and oregano essential oil in forced molted and fully fed laying hens between 82 and 106 weeks of age. Poult Sci. (2016) 95:2576–91. doi: 10.3382/ps/pew140
77. Yang J, Zhan K, Zhang M. Effects of the use of a combination of two Bacillus species on performance, egg quality, small intestinal mucosal morphology, and cecal microbiota profile in aging laying hens. Probiotics Antimicrob Proteins. (2020) 12:204–13.
78. Ye M, Wei C, Khalid A, Hu Q, Yang R, Dai B, et al. Effect of Bacillus velezensis to substitute in-feed antibiotics on the production, blood biochemistry and egg quality indices of laying hens. BMC Vet Res. (2020) 16:400. doi: 10.1186/s12917-020-02570-6
79. Zhu Y-W, Lu L, Li W-X, Zhang L-Y, Ji C, Lin X, et al. Effect of dietary manganese on antioxidant status and expression levels of heat-shock proteins and factors in tissues of laying broiler breeders under normal and high environmental temperatures. Br J Nutr. (2015) 114:1965–74. doi: 10.1017/S0007114515003803
80. Wang J, Bai X, Ding X, Bai S, Zeng Q, Mao X, et al. Quantitative proteomic analysis reveals the role of tea polyphenol EGCG in egg whites in response to vanadium stress. Nutrition. (2017) 39–40:20–9. doi: 10.1016/j.nut.2017.02.007
81. Wan Y, Ma R, Qi R, Li Y, Liu W, Li J, et al. Dietary fresh lemon improves the albumen quality, immune status and lipid metabolism of Jingfen laying hens during the late laying period. Ital J Anim Sci. (2021) 20:834–41.
82. Afzal M, Safer A, Menon M. Green tea polyphenols and their potential role in health and disease. Inflammopharmacology. (2015) 23:151–61. doi: 10.1007/s10787-015-0236-1
83. Liu X, Lin X, Mi Y, Li J, Zhang C. Grape seed proanthocyanidin extract prevents ovarian aging by inhibiting oxidative stress in the hens. Oxid Med Cell Longev. (2018) 2018:9390810. doi: 10.1155/2018/9390810
84. Surai PF. Antioxidant systems in poultry biology: superoxide dismutase. J Anim Res Nutr. (2016) 1:8.
85. Galli F. Amino acid and protein modification by oxygen and nitrogen species. Amino Acids. (2012) 42:1–4. doi: 10.1007/s00726-010-0670-8
86. Ariana M, Samie A, Edriss MA, Jahanian R. Effects of powder and extract form of green tea and marigold, and α-tocopheryl acetate on performance, egg quality and egg yolk cholesterol levels of laying hens in late phase of production. J Med Plants Res. (2011) 5:2710–6.
87. Bai X, Wang J, Ding X, Bai S, Zeng Q, Xuan Y, et al. Proteomic alteration of albumen by dietary vanadium in commercial egg-type layers. Poult Sci. (2020) 99:1705–16. doi: 10.1016/j.psj.2019.10.056
88. Ighodaro O, Akinloye O. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): their fundamental role in the entire antioxidant defence grid. Alex J Med. (2018) 54:287–93.
89. Zhang Z, Kim I. Effects of probiotic supplementation in different energy and nutrient density diets on performance, egg quality, excreta microflora, excreta noxious gas emission, and serum cholesterol concentrations in laying hens. J Anim Sci. (2013) 91:4781–7. doi: 10.2527/jas.2013-6484
90. Xiang Q, Wang C, Zhang H, Lai W, Wei H, Peng J. Effects of different probiotics on laying performance, egg quality, oxidative status, and gut health in laying hens. Animals (Basel). (2019) 9:1110. doi: 10.3390/ani9121110
91. Upadhaya SD, Rudeaux F, Kim IH. Efficacy of dietary Bacillus subtilis and Bacillus licheniformis supplementation continuously in pullet and lay period on egg production, excreta microflora, and egg quality of Hyline-Brown birds. Poult Sci. (2019) 98:4722–8. doi: 10.3382/ps/pez184
92. Arpášová H, Kaèániová M, Pistová V, Gálik B, Fik M, Hleba L, et al. Effect of probiotics and humic acid on egg production and quality parameters of laying hens eggs. Sci Pap Anim Sci Biotechnol. (2016) 49:1–9.
93. Zhou Y, Li S, Pang Q, Miao Z. Bacillus amyloliquefaciens BLCC1-0238 can effectively improve laying performance and egg quality via enhancing immunity and regulating reproductive hormones of laying hens. Probiotics Antimicrob Proteins. (2020) 12:246–52.
94. Martinez JS, Blount R, Park J, McIntyre D, Pavlidis H, Carey J. Effects of feeding original XPC™ to laying hens on egg production, component yield and composition. J Appl Poul Res. (2018) 27:603–8.
95. Towett G. What Are Effective Microorganisms. The Channon, NSW: The Premaculture Research Institute (2016).
96. Atsbeha AT, Hailu TG. The impact of effective microorganisms (EM) on egg quality and laying performance of chickens. Int J Food Sci. (2021) 2021:8895717. doi: 10.1155/2021/8895717
97. Jazi V, Ashayerizadeh A, Toghyani M, Shabani A, Tellez G. Fermented soybean meal exhibits probiotic properties when included in Japanese quail diet in replacement of soybean meal. Poult Sci. (2018) 97:2113–22. doi: 10.3382/ps/pey071
98. Zhan H, Dong X, Li L, Zheng Y, Gong Y, Zou X. Effects of dietary supplementation with Clostridium butyricum on laying performance, egg quality, serum parameters, and cecal microflora of laying hens in the late phase of production. Poult Sci. (2019) 98:896–903.
99. Wang W-w, Wang J, Zhang H-j, Wu S-g, Qi G-h. Effects of Clostridium butyricum on production performance and intestinal absorption function of laying hens in the late phase of production. Anim Feed Sci Technol. (2020) 264:114476.
100. Lei K, Li Y, Yu D, Rajput I, Li W. Influence of dietary inclusion of Bacillus licheniformis on laying performance, egg quality, antioxidant enzyme activities, and intestinal barrier function of laying hens. Poult Sci. (2013) 92:2389–95. doi: 10.3382/ps.2012-02686
101. Guo JR, Dong XF, Liu S, Tong JM. Effects of long-term Bacillus subtilis CGMCC 1.921 supplementation on performance, egg quality, and fecal and cecal microbiota of laying hens. Poult Sci. (2017) 96:1280–9. doi: 10.3382/ps/pew389
102. Liu X, Peng C, Qu X, Guo S, Chen JF, He C, et al. Effects of Bacillus subtilis C-3102 on production, hatching performance, egg quality, serum antioxidant capacity and immune response of laying breeders. J Anim Physiol Anim Nutr (Berl). (2019) 103:182–90.
103. Deng Y, Xiong X, Liu X, He C, Guo S, Tang S, et al. Palygorskite combined probiotics improve the laying performance, hatching performance, egg quality, plasma antioxidative status, and immune response of broiler breeders. Ital J Anim Sci. (2021) 20:1292–301.
104. Fathi M, Al-Homidan I, Al-Dokhail A, Ebeid T, Abou-Emera O, Alsagan A. Effects of dietary probiotic (Bacillus subtilis) supplementation on productive performance, immune response and egg quality characteristics in laying hens under high ambient temperature. Ital J Anim Sci. (2018) 17:804–14.
105. Guo Y, Zhao ZH, Pan ZY, An LL, Balasubramanian B, Liu WC. New insights into the role of dietary marine-derived polysaccharides on productive performance, egg quality, antioxidant capacity, and jejunal morphology in late-phase laying hens. Poult Sci. (2020) 99:2100–7. doi: 10.1016/j.psj.2019.12.032
106. Jahanian R, Ashnagar M. Effect of dietary supplementation of mannan-oligosaccharides on performance, blood metabolites, ileal nutrient digestibility, and gut microflora in Escherichia coli-challenged laying hens. Poult Sci. (2015) 94:2165–72. doi: 10.3382/ps/pev180
107. Koiyama N, Utimi N, Santos B, Bonato M, Barbalho R, Gameiro A, et al. Effect of yeast cell wall supplementation in laying hen feed on economic viability, egg production, and egg quality. J Appl Poult Res. (2018) 27:116–23.
108. Xu Z, Hu C, Xia M, Zhan X, Wang M. Effects of dietary fructooligosaccharide on digestive enzyme activities, intestinal microflora and morphology of male broilers. Poult Sci. (2003) 82:1030–6. doi: 10.1093/ps/82.6.1030
109. Kim S, Jang MJ, Kim SY, Yang Y, Pavlidis HO, Ricke SC. Potential for prebiotics as feed additives to limit foodborne Campylobacter establishment in the poultry gastrointestinal tract. Front Microbiol. (2019) 10:91.
110. Donalson L, Kim W, Chalova V, Herrera P, McReynolds J, Gotcheva V, et al. In vitro fermentation response of laying hen cecal bacteria to combinations of fructooligosaccharide prebiotics with alfalfa or a layer ration. Poult Sci. (2008) 87:1263–75. doi: 10.3382/ps.2007-00179
111. Ding X, Li D, Bai S, Wang J, Zeng Q, Su Z, et al. Effect of dietary xylooligosaccharides on intestinal characteristics, gut microbiota, cecal short-chain fatty acids, and plasma immune parameters of laying hens. Poult Sci. (2018) 97:874–81. doi: 10.3382/ps/pex372
112. Zhou JM, Zhang HJ, Wu SG, Qiu K, Fu Y, Qi GH, et al. Supplemental xylooligosaccharide modulates intestinal mucosal barrier and cecal microbiota in laying hens fed oxidized fish oil. Front Microbiol. (2021) 12:635333. doi: 10.3389/fmicb.2021.635333
113. Sozcu A, Ipek A. The effects of lignocellulose supplementation on laying performance, egg quality parameters, aerobic bacterial load of eggshell, serum biochemical parameters, and jejunal histomorphological traits of laying hens. Poult Sci. (2020) 99:3179–87. doi: 10.1016/j.psj.2020.01.024
114. Xie T, Bai SP, Zhang KY, Ding XM, Wang JP, Zeng QF, et al. Effects of Lonicera confusa and Astragali Radix extracts supplementation on egg production performance, egg quality, sensory evaluation, and antioxidative parameters of laying hens during the late laying period. Poult Sci. (2019) 98:4838–47.
115. Wen C, Gu Y, Tao Z, Cheng Z, Wang T, Zhou Y. Effects of ginger extract on laying performance, egg quality, and antioxidant status of laying hens. Animals (Basel). (2019) 9:857. doi: 10.3390/ani9110857
116. Song D, Wang Y, Lu Z, Wang W, Miao H, Zhou H, et al. Effects of dietary supplementation of microencapsulated Enterococcus fecalis and the extract of Camellia oleifera seed on laying performance, egg quality, serum biochemical parameters, and cecal microflora diversity in laying hens. Poult Sci. (2019) 98:2880–7.
117. Abdel-Wareth AA. Effect of dietary supplementation of thymol, synbiotic and their combination on performance, egg quality and serum metabolic profile of Hy-Line Brown hens. Br Poult Sci. (2016) 57:114–22. doi: 10.1080/00071668.2015.1123219
118. Abou-Elkhair R, Selim S, Hussein E. Effect of supplementing layer hen diet with phytogenic feed additives on laying performance, egg quality, egg lipid peroxidation and blood biochemical constituents. Anim Nutr. (2018) 4:394–400. doi: 10.1016/j.aninu.2018.05.009
119. Li XL, He WL, Yang ML, Yan YM, Xue YH, Zhao ST. Effect of dietary supplementation of Ligustrum lucidum on performance, egg quality and blood biochemical parameters of Hy-Line Brown hens during the late laying period. Animal. (2017) 11:1899–904. doi: 10.1017/S1751731117000532
120. Al-kirshi R, Alimon AR, Zulkifli I, Sazili A, Wan Zahari M, Ivan M. Utilization of mulberry leaf meal (Morus alba) as protein supplement in diets for laying hens. Ital J Anim Sci. (2016) 9:e51.
121. Li XL, He WL, Wang ZB, Xu TS. Effects of Chinese herbal mixture on performance, egg quality and blood biochemical parameters of laying hens. J Anim Physiol Anim Nutr (Berl). (2016) 100:1041–9. doi: 10.1111/jpn.12473
122. Gurbuz Y, Salih Y. Influence of sumac (Rhus Coriaria L.) and ginger (Zingiber officinale) on egg yolk fatty acid, cholesterol and blood parameters in laying hens. J Anim Physiol Anim Nutr (Berl). (2017) 101:1316–23. doi: 10.1111/jpn.12652
123. Omer HAA, Ahmed SM, Abdel-Magid SS, El-Mallah GMH, Bakr AA, Abdel Fattah MM. Nutritional impact of inclusion of garlic (Allium sativum) and/or onion (Allium cepa L.) powder in laying hens’ diets on their performance, egg quality, and some blood constituents. Bull Natl. Res Centre. (2019) 43:23.
124. Su B-W, Lin W-C, Lin L-J, Huang C-M, Chuang W-Y, Wu D-J, et al. Laying diet supplementation with Ricinus communis L. leaves and evaluation of productive performance and potential modulation of antioxidative status. J Poult Sci. (2020) 57:259–69. doi: 10.2141/jpsa.0190077
125. Choi Y, Lee EC, Na Y, Lee SR. Effects of dietary supplementation with fermented and non-fermented brown algae by-products on laying performance, egg quality, and blood profile in laying hens. Asian Australas J Anim Sci. (2018) 31:1654–9. doi: 10.5713/ajas.17.0921
126. Abdel-Wareth A, Lohakare J. Productive performance, egg quality, nutrients digestibility, and physiological response of bovans brown hens fed various dietary inclusion levels of peppermint oil. Anim Feed Sci Technol. (2020) 267:114554.
127. Kaya A, Yıldırım BA, Kaya H, Gül M, Çelebi Ç. The effects of diets supplemented with crushed and extracted grape seed on performance, egg quality parameters, yolk peroxidation and serum traits in laying hens. Europ Poult Sci. (2014) 78:1–10.
128. Santana-Gálvez J, Cisneros-Zevallos L, Jacobo-Velázquez DA. Chlorogenic acid: recent advances on its dual role as a food additive and a nutraceutical against metabolic syndrome. Molecules. (2017) 22:358. doi: 10.3390/molecules22030358
129. Radwan Nadia L, Hassan R, Qota E, Fayek H. Effect of natural antioxidant on oxidative stability of eggs and productive and reproductive performance of laying hens. Int J Poult Sci. (2008) 7:134–50.
130. Wang J, Yuan Z, Zhang K, Ding X, Bai S, Zeng Q, et al. Epigallocatechin-3-gallate protected vanadium-induced eggshell depigmentation via P38MAPK-Nrf2/HO-1 signaling pathway in laying hens. Poult Sci. (2018) 97:3109–18. doi: 10.3382/ps/pey165
132. Wang J, Jia R, Celi P, Ding X, Bai S, Zeng Q, et al. Green tea polyphenol epigallocatechin-3-gallate improves the antioxidant capacity of eggs. Food Funct. (2020) 11:534–43. doi: 10.1039/c9fo02157d
133. Chen X, Li T, He K, Geng Z, Wan X. Dietary green tea powder supplementation enriched egg nutrients and physicochemical property in an indigenous chicken breed. Poult Sci. (2021) 100:388–95. doi: 10.1016/j.psj.2020.10.001
134. Xia B, Liu Y, Sun D, Liu J, Zhu Y, Lu L. Effects of green tea powder supplementation on egg production and egg quality in laying hens. J Appl Anim Res. (2018) 46:927–31.
135. Hatanaka Y, Yamauchi A, Kobayashi O, Muro T. Electron microscopic analysis of the effects of tea extract on strength improvement of egg white gels. Food Sci Technol Res. (2009) 15:5–10.
136. Kanakis C, Hasni I, Bourassa P, Tarantilis P, Polissiou M, Tajmir-Riahi H-A. Milk β-lactoglobulin complexes with tea polyphenols. Food Chem. (2011) 127:1046–55.
137. Zhang H, Tsao R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr Opin Food Sci. (2016) 8:33–42.
138. Surai P. Polyphenol compounds in the chicken/animal diet: from the past to the future. J Anim Physiol Anim Nutr (Berl). (2014) 98:19–31. doi: 10.1111/jpn.12070
139. Smith D, Galazka VB, Wellner N, Sumner IG. High pressure unfolding of ovalbumin. Int J Food Sci Technol. (2000) 35:361–70.
140. Li LL, Gong YJ, Zhan HQ, Zheng YX, Zou XT. Effects of dietary Zn-methionine supplementation on the laying performance, egg quality, antioxidant capacity, and serum parameters of laying hens. Poult Sci. (2019) 98:923–31. doi: 10.3382/ps/pey440
141. Shengfeng C, Li Y, Meirong C, Luyong C, Keqiang C. Effects of feeding high-iron and high-iodine diet to hens on the iron and iodine content in egg and egg quality. J Shanghai Agric Coll. (1999) 17:248–54.
142. Pan C, Zhao Y, Liao SF, Chen F, Qin S, Wu X, et al. Effect of selenium-enriched probiotics on laying performance, egg quality, egg selenium content, and egg glutathione peroxidase activity. J Agric Food Chem. (2011) 59:11424–31. doi: 10.1021/jf202014k
143. Xie C, Elwan HAM, Elnesr SS, Dong XY, Zou XT. Effect of iron glycine chelate supplementation on egg quality and egg iron enrichment in laying hens. Poult Sci. (2019) 98:7101–9. doi: 10.3382/ps/pez421
144. Meng T, Liu Y-l, Xie C-y, Zhang B, Huang Y-q, Zhang Y-w, et al. Effects of different selenium sources on laying performance, egg selenium concentration, and antioxidant capacity in laying hens. Biol Trace Elem Res. (2019) 189:548–55. doi: 10.1007/s12011-018-1490-z
145. Qiu K, Zheng J-J, Obianwuna UE, Wang J, Zhang H-J, Qi G-H, et al. Effects of dietary selenium sources on physiological status of laying hens and production of selenium-enriched eggs. Front Nutr. (2021) 8:726770. doi: 10.3389/fnut.2021.726770
146. Han XJ, Qin P, Li WX, Ma QG, Ji C, Zhang JY, et al. Effect of sodium selenite and selenium yeast on performance, egg quality, antioxidant capacity, and selenium deposition of laying hens. Poult Sci. (2017) 96:3973–80. doi: 10.3382/ps/pex216
147. Asadi F, Shariatmadari F, Karimi-Torshizi M, Mohiti-Asli M, Ghanaatparast-Rashti M. Comparison of different selenium sources and vitamin E in laying hen diet and their influences on egg selenium and cholesterol content, quality and oxidative stability. Iran J Appl Anim Sci. (2017) 7:83–9.
148. Lu J, Qu L, Ma M, Li Y, Wang X, Yang Z, et al. Efficacy evaluation of selenium-enriched yeast in laying hens: effects on performance, egg quality, organ development, and selenium deposition. Poult Sci. (2020) 99:6267–77. doi: 10.1016/j.psj.2020.07.041
149. Chantiratikul A, Chinrasri O, Chantiratikul P. Effect of selenium from selenium-enriched kale sprout versus other selenium sources on productivity and selenium concentrations in egg and tissue of laying hens. Biol Trace Elem Res. (2018) 182:105–10. doi: 10.1007/s12011-017-1069-0
150. Qiu K, Ma Y, Obianwuna UE, Wang J, Zhang H, Qi G, et al. Application of selenium conjugated to animal protein in laying hens’ diet for the production of selenium-enriched eggs. Foods. (2021) 10:1224. doi: 10.3390/foods10061224
151. Zhang X, Tian L, Zhai S, Lin Z, Yang H, Chen J, et al. Effects of selenium-enriched yeast on performance, egg quality, antioxidant balance, and egg selenium content in laying ducks. Front Vet Sci. (2020) 7:591. doi: 10.3389/fvets.2020.00591
152. Nemati Z, Ahmadian H, Besharati M, Lesson S, Alirezalu K, Dominguez R, et al. Assessment of dietary selenium and vitamin E on laying performance and quality parameters of fresh and stored eggs in Japanese quails. Foods. (2020) 9:1324. doi: 10.3390/foods9091324
153. Liu H, Yu Q, Fang C, Chen S, Tang X, Ajuwon KM, et al. Effect of selenium source and level on performance, egg quality, egg selenium content, and serum biochemical parameters in laying hens. Foods. (2020) 9:68. doi: 10.3390/foods9010068
154. Qi X, Ma S, Liu X, Wang Y, Liu Y, Gao Y, et al. Effects of the methionine hydroxyl analogue chelate zinc on antioxidant capacity and liver metabolism using 1H-NMR-based metabolomics in aged laying hens. Animals (Basel). (2019) 9:898. doi: 10.3390/ani9110898
155. Behjatian Esfahani M, Moravej H, Ghaffarzadeh M, Nehzati Paghaleh GA. Comparison the Zn-threonine, Zn-methionine, and Zn oxide on performance, egg quality, Zn bioavailability, and zn content in egg and excreta of laying hens. Biol Trace Elem Res. (2021) 199:292–304. doi: 10.1007/s12011-020-02141-8
156. Sun Q, Guo Y, Li J, Zhang T, Wen J. Effects of methionine hydroxy analog chelated Cu/Mn/Zn on laying performance, egg quality, enzyme activity and mineral retention of laying hens. J Poult Sci. (2010) 49:20.
157. Abd El-Hack ME, Alagawany M, Amer SA, Arif M, Wahdan KMM, El-Kholy MS. Effect of dietary supplementation of organic zinc on laying performance, egg quality and some biochemical parameters of laying hens. J Anim Physiol Anim Nutr (Berl). (2018) 102:e542–9. doi: 10.1111/jpn.12793
158. Bahakaim A, Abdel Magied H, Osman S, Omar A, Abdelmalak N, Ramadan N. Effect of using different levels and sources of zinc in layer’s diets on egg zinc enrichment. Egypt Poult Sci. (2014) 34:39–56.
159. Noor R, Mittal S, Iqbal J. Superoxide dismutase–applications and relevance to human diseases. Med Sci Monit. (2002) 8:RA210–5.
160. Min Y, Liu F, Qi X, Ji S, Cui L, Wang Z, et al. Effects of organic zinc on tibia quality, mineral deposit, and metallothionein expression level of aged hens. Poult Sci. (2019) 98:366–72. doi: 10.3382/ps/pey386
161. Wang X, Ou D, Yin J, Wu G, Wang J. Proteomic analysis reveals altered expression of proteins related to glutathione metabolism and apoptosis in the small intestine of zinc oxide-supplemented piglets. Amino Acids. (2009) 37:209–18. doi: 10.1007/s00726-009-0242-y
162. Amem MH, Al-Daraji HJ. Zinc improves egg quality in Cobb500 broiler breeder females. Int J Poult Sci. (2011) 10:471–6.
163. Abbasi M, Zaghari M, Ganjkhanlo M, Khalaji S. Is dietary iron requirement of broiler breeder hens at the late stage of production cycle influenced by phytase supplementation? Appl. Anim. Res. (2015) 43:166–76.
164. Pereira GCC, Costa FGP, Silva JHVd, Pascoal LAF, Lima CABd, Bittencourt LC, et al. Different trace mineral sources and recommendations in the performance and quality of eggs from Dekalb White layers. Rev Bras Zootecnia. (2018) 47:e20170235.
165. Chang L, Zhang R, Fu S, Mu C, Tang Q, Bu Z. Effects of different dietary calcium levels on the performance, egg quality, and albumen transparency of laying pigeons. Animals (Basel). (2019) 9:110. doi: 10.3390/ani9030110
166. Chen J, Kuang Y, Qu X, Guo S, Kang K, He C. The effects and combinational effects of Bacillus subtilis and montmorillonite supplementation on performance, egg quality, oxidation status, and immune response in laying hens. Livest Sci. (2019) 227:114–9.
167. Qu XY, Chen JF, He CQ, Chi F, Johnston SL. Effects of modified montmorillonite adsorbent on performance, egg quality, serum biochemistry, oxidation status, and immune response of laying hens in late production. Livest Sci. (2018) 210:15–20.
168. Cui Y-m, Zhang H-j, Zhou J-m, Wu S-g, Zhang C, Qi G-h, et al. Effects of long-term supplementation with amino acid-complexed manganese on performance, egg quality, blood biochemistry and organ histopathology in laying hens. Anim Feed Sci Technol. (2019) 254:114203.
169. Gong H, Yang Z, Celi P, Yan L, Ding X, Bai S, et al. Effect of benzoic acid on production performance, egg quality, intestinal morphology, and cecal microbial community of laying hens. Poult Sci. (2021) 100:196–205. doi: 10.1016/j.psj.2020.09.065
170. Feng Z, Gong J, Zhao G, Lin X, Liu Y, Ma K. Effects of dietary supplementation of resveratrol on performance, egg quality, yolk cholesterol and antioxidant enzyme activity of laying hens. Br Poult Sci. (2017) 58:544–9. doi: 10.1080/00071668.2017.1349295
171. Long L, Wu SG, Yuan F, Zhang HJ, Wang J, Qi GH. Effects of dietary octacosanol supplementation on laying performance, egg quality, serum hormone levels, and expression of genes related to the reproductive axis in laying hens. Poult Sci. (2017) 96:894–903. doi: 10.3382/ps/pew316
172. Ýskender H, Yenice G, Dokumacioglu E, Kaynar O, Hayirli A, Kaya A. Comparison of the effects of dietary supplementation of flavonoids on laying hen performance, egg quality and egg nutrient profile. Br Poult Sci. (2017) 58:550–6. doi: 10.1080/00071668.2017.1349297
173. Dansou DM, Wang H, Nugroho RD, He W, Zhao Q, Zhang J. Assessment of response to moderate and high dose supplementation of astaxanthin in laying hens. Animals. (2021) 11:1138. doi: 10.3390/ani11041138
174. Chen C, Turner B, Applegate TJ, Litta G, Kim WK. Role of long-term supplementation of 25-hydroxyvitamin D3 on egg production and egg quality of laying hen. Poult Sci. (2020) 99:6899–906.
175. Silva FA. Effects of Dietary 25-Hydroxycholecalciferol on Growth, Production Performance, Eggshell Quality and Bone Traits of Brown Egg Layers Housed Under Commercial Conditions. Master’s thesis. Edmonton, AB: University of Alberta (2017).
176. Adhikari R, White D, House J, Kim W. Effects of additional dosage of vitamin D3, vitamin D2, and 25-hydroxyvitamin D3 on calcium and phosphorus utilization, egg quality and bone mineralization in laying hens. Poult Sci. (2020) 99:364–73.
178. Castro F, Kim Y, Xu H, Kim W. The effect of total sulfur amino acid levels on growth performance and bone metabolism in pullets under heat stress. Poult Sci. (2020) 99:5783–91. doi: 10.1016/j.psj.2020.06.081
179. Liu Y, Wan D, Zhou X, Ruan Z, Zhang T, Wu X, et al. Effects of dynamic feeding low-and high-methionine diets on the variation of glucose and lipid metabolism-related genes in the liver of laying hens. Poult Sci. (2019) 98:2231–40. doi: 10.3382/ps/pey589
180. Ghods-Alavi B, Samie H, Jahanian R. Effects of supplementary dietary L-carnitine on performance and egg quality of laying hens fed diets different in fat level. Ital J Anim Sci. (2017) 9:e3.
181. Azzam MM, Dong XY, Dai L, Zou XT. Effect of excess dietary L-valine on laying hen performance, egg quality, serum free amino acids, immune function and antioxidant enzyme activity. Br Poult Sci. (2015) 56:72–8. doi: 10.1080/00071668.2014.989487
182. Alagawany M, Mahrose KM. Influence of different levels of certain essential amino acids on the performance, egg quality criteria and economics of Lohmann brown laying hens. Asian J Poult Sci. (2014) 8:82–96.
183. Jiang S, El-Senousey HK, Fan Q, Lin X, Gou Z, Li L, et al. Effects of dietary threonine supplementation on productivity and expression of genes related to protein deposition and amino acid transportation in breeder hens of yellow-feathered chicken and their offspring. Poult Sci. (2019) 98:6826–36. doi: 10.3382/ps/pez420
184. Wu G, Bryant M, Gunawardana P, Roland D Sr. Effect of nutrient density on performance, egg components, egg solids, egg quality, and profits in eight commercial leghorn strains during phase one. Poult Sci. (2007) 86:691–7. doi: 10.1093/ps/86.4.691
185. Sun F, Cai Y, Liu G, Zhang S, Chang W, Yan H. The research progress of arginine poultry nutrition. Feed Res. (2010) 6:24–5.
186. Azzam M, Dong X, Zou X. Effect of dietary threonine on laying performance and intestinal immunity of laying hens fed low-crude-protein diets during the peak production period. J Anim Physiol Anim Nutr (Berl). (2017) 101:e55–66. doi: 10.1111/jpn.12559
187. Scappaticcio R, García J, Fondevila G, de Juan A, Cámara L, Mateos G. Influence of the energy and digestible lysine contents of the diet on performance and egg quality traits of brown-egg laying hens from 19 to 59 weeks of age. Poult Sci. (2021) 100:101211. doi: 10.1016/j.psj.2021.101211
188. Uyanga VA, Jiao H, Zhao J, Wang X, Lin H. Dietary L-citrulline supplementation modulates nitric oxide synthesis and anti-oxidant status of laying hens during summer season. J Anim Sci Biotechnol. (2020) 11:103. doi: 10.1186/s40104-020-00507-5
189. Adeyemo GO, Abioye SA, Aderemi FA. The effect of varied dietary crude protein levels with balanced amino acids on performance and egg quality characteristics of layers at first laying phase. Food Nutr Sci. (2012) 03:526–9.
190. Kowalska E, Kucharska-Gaca J, Kuzniacka J, Lewko L, Gornowicz E, Biesek J, et al. Egg quality depending on the diet with different sources of protein and age of the hens. Sci Rep. (2021) 11:2638. doi: 10.1038/s41598-021-82313-1
191. Kuzniacka J, Biesek J, Banaszak M, Grabowicz M, Adamski M. The quality of eggs from rosa 1 hens fed diets containing seeds of legume plants (Lupinus luteus L., Lupinus angustifolius, and Pisum sativum) in two laying phases. Animals (Basel). (2020) 10:1942. doi: 10.3390/ani10111942
192. Shim MY, Song E, Billard L, Aggrey SE, Pesti GM, Sodsee P. Effects of balanced dietary protein levels on egg production and egg quality parameters of individual commercial layers. Poult Sci. (2013) 92:2687–96. doi: 10.3382/ps.2012-02569
193. Zhou J-m, Qiu K, Wang J, Zhang H-j, Qi G-h, Wu S-g. Effect of dietary serine supplementation on performance, egg quality, serum indices, and ileal mucosal immunity in laying hens fed a low crude protein diet. Poultry Sci. (2021) 100:101465. doi: 10.1016/j.psj.2021.101465
194. Torki M, Mohebbifar A, Mohammadi H. Effects of supplementing hen diet with Lavandula angustifolia and/or Mentha spicata essential oils on production performance, egg quality and blood variables of laying hens. Vet Med Sci. (2021) 7:184–93. doi: 10.1002/vms3.343
195. Alagawany M, Abd El-Hack ME, Ashour EA, Salah AS, Hussein E-SOS, Alowaimer AA, et al. Raw faba bean (Vicia faba) as an alternative protein source in laying hen diets. J Appl Poult Res. (2019) 28:808–17.
196. He T, Zhang HJ, Wang J, Wu SG, Yue HY, Qi GH. Application of low-gossypol cottonseed meal in laying hens’ diet. Poult Sci. (2015) 94:2456–63. doi: 10.3382/ps/pev247
197. Parenteau IA, Stevenson M, Kiarie EG. Egg production and quality responses to increasing isoleucine supplementation in Shaver white hens fed a low crude protein corn-soybean meal diet fortified with synthetic amino acids between 20 and 46 weeks of age. Poult Sci. (2020) 99:1444–53. doi: 10.1016/j.psj.2019.10.064
198. Dong X, Azzam M, Zou X. Effects of dietary threonine supplementation on intestinal barrier function and gut microbiota of laying hens. Poult Sci. (2017) 96:3654–63. doi: 10.3382/ps/pex185
199. Cherian G, Quezada N. Egg quality, fatty acid composition and immunoglobulin Y content in eggs from laying hens fed full fat camelina or flax seed. J Anim Sci Biotechnol. (2016) 7:15. doi: 10.1186/s40104-016-0075-y
200. Kocer B, Bozkurt M, Ege G, Tuzun AE. Effects of sunflower meal supplementation in the diet on productive performance, egg quality and gastrointestinal tract traits of laying hens. Br Poult Sci. (2021) 62:101–9. doi: 10.1080/00071668.2020.1814202
202. Mu Y, Zhu LY, Yang A, Gao X, Zhang N, Sun L, et al. The effects of dietary cottonseed meal and oil supplementation on laying performance and egg quality of laying hens. Food Sci Nutr. (2019) 7:2436–47. doi: 10.1002/fsn3.1112
203. Wang L, Li A, Shi J, Liu K, Cheng J, Song D, et al. Effects of different levels of cottonseed meal on laying performance, egg quality, intestinal immunity and hepatic histopathology in laying hens. Food Agric Immunol. (2020) 31:803–12.
204. Ciurescu G, Panã CO. Effect of dietary untreated field pea (Pisum sativum L.) as substitute for soybean meal and enzymes supplementation on egg production and quality of laying hens. Rom Biotechnol Lett. (2017) 22: 12204–13.
205. Wanzenböck E, Schreiner M, Zitz U, Bleich B, Figl S, Kneifel W, et al. A combination of wheat bran and vegetable oils as feedstuff in laying hens’ diet: impact on egg quality parameters. Agric Sci. (2018) 9:676–91.
206. Aguillón-Páez YJ, Romero LA, Diaz G. Effect of full-fat sunflower or flaxseed seeds dietary inclusion on performance, egg yolk fatty acid profile and egg quality in laying hens. Anim Nutr. (2020) 6:179–84. doi: 10.1016/j.aninu.2019.12.005
207. Jiao H, Hou N, Jiang M, Song Z, Lin H. Effects of dietary free gossypol levels on liver gossypol accumulation, performance and egg quality of laying hens. Chin J Anim Nutr. (2015) 27:3207–17.
208. Świątkiewicz S, Arczewska-Włosek A, Krawczyk J, Szczurek W, Puchała M, Józefiak D. Effect of selected feed additives on egg performance and eggshell quality in laying hens fed a diet with standard or decreased calcium content. Ann Anim Sci. (2018) 18:167–83.
209. Mena H, Santos J, Huber J, Tarazon M, Calhoun M. The effects of varying gossypol intake from whole cottonseed and cottonseed meal on lactation and blood parameters in lactating dairy cows. J Dairy Sci. (2004) 87:2506–18. doi: 10.3168/jds.S0022-0302(04)73375-5
210. Fru-Nji F, Niess E, Pfeffer E. Effect of graded replacement of soybean meal by faba beans (Vicia faba L.) or field peas (Pisum sativum L.) in rations for laying hens on egg production and quality. J Poult Sci. (2007) 44:34–41.
211. Laudadio V, Ceci E, Tufarelli V. Productive traits and meat fatty acid profile of broiler chickens fed diets containing micronized fava beans (Vicia faba L. var. minor) as the main protein source. J Appl Poult Res. (2011) 20:12–20.
212. Abd El-Hack ME, Alagawany M, Laudadio V, Demauro R, Tufarelli V. Dietary inclusion of raw faba bean instead of soybean meal and enzyme supplementation in laying hens: effect on performance and egg quality. Saudi J Biol Sci. (2017) 24:276–85. doi: 10.1016/j.sjbs.2015.05.009
213. Lei X, Lee K, Kim I. Performance, egg quality, nutrient digestibility, and excreta microbiota shedding in laying hens fed corn-soybean-meal-wheat-based diets supplemented with xylanase. Poult Sci. (2018) 97:2071–7. doi: 10.3382/ps/pey041
214. Sun HY, Kim IH. Effects of multi-enzyme on production performance, egg quality, nutrient digestibility, and excreta noxious gas emission of early phase Hy-line brown hens. Poult Sci. (2019) 98:4889–95. doi: 10.3382/ps/pez237
215. Nguyen H, Wu S-B, Bedford M, Nguyen X, Morgan N. Dietary soluble non-starch polysaccharide level and xylanase influence the gastrointestinal environment and nutrient utilisation in laying hens. Br Poult Sci. (2021) 6:1–11. doi: 10.1080/00071668.2021.2003754
216. Bederska-Łojewska D, Arczewska-Włosek A, Świątkiewicz S, Orczewska-Dudek S, Schwarz T, Puchała M, et al. The effect of different dietary levels of hybrid rye and xylanase addition on the performance and egg quality in laying hens. Br Poult Sci. (2019) 60:423–30. doi: 10.1080/00071668.2019.1605149
217. Abbasi Arabshahi H, Ghasemi HA, Hajkhodadadi I, Khaltabadi Farahani AH. Effects of multicarbohydrase and butyrate glycerides on productive performance, nutrient digestibility, gut morphology, and ileal microbiota in late-phase laying hens fed corn-or wheat-based diets. Poult Sci. (2021) 100:101066. doi: 10.1016/j.psj.2021.101066
218. Guo L, Lv J, Liu Y, Ma H, Chen B, Hao K, et al. Effects of different fermented feeds on production performance, cecal microorganisms, and intestinal immunity of laying hens. Animals. (2021) 11:2799. doi: 10.3390/ani11102799
219. Irawan AC, Astuti DA, Wibawan IWT, Hermana W. Impact of the feeding with the black soldier fly (Hermetia illucens) on egg physical quality, egg chemical quality and lipid metabolism of laying hens. J Phys Conf Ser. (2019) 1351:012081.
220. Ding X, Yu Y, Su Z, Zhang K. Effects of essential oils on performance, egg quality, nutrient digestibility and yolk fatty acid profile in laying hens. Anim Nutr. (2017) 3:127–31. doi: 10.1016/j.aninu.2017.03.005
221. Akbari M, Torki M, Kaviani K. Single and combined effects of peppermint and thyme essential oils on productive performance, egg quality traits, and blood parameters of laying hens reared under cold stress condition (6.8±3 °C). Int J Biometeorol. (2016) 60:447–54. doi: 10.1007/s00484-015-1042-6
222. Dong X, Liu S, Tong J. Comparative effect of dietary soybean oil, fish oil, and coconut oil on performance, egg quality and some blood parameters in laying hens. Poult Sci. (2018) 97:2460–72. doi: 10.3382/ps/pey094
223. Marume U, Mokagane JM, Shole CO, Hugo A. Citrullus lanatus essential oils inclusion in diets elicit nutraceutical effects on egg production, egg quality, and physiological characteristics in layer hens. Poult Sci. (2020) 99:3038–46. doi: 10.1016/j.psj.2020.01.029
224. Wang H, Liang S, Li X, Yang X, Long F, Yang X. Effects of encapsulated essential oils and organic acids on laying performance, egg quality, intestinal morphology, barrier function, and microflora count of hens during the early laying period. Poult Sci. (2019) 98:6751–60. doi: 10.3382/ps/pez391
225. Ramirez SY, Peñuela-Sierra LM, Ospina MA. Effects of oregano (Lippia origanoides) essential oil supplementation on the performance, egg quality, and intestinal morphometry of Isa Brown laying hens. Vet World. (2021) 14:595. doi: 10.14202/vetworld.2021.595-602
226. Irías-Mata A, Stuetz W, Sus N, Hammann S, Gralla K, Cordero-Solano A, et al. Tocopherols, tocomonoenols, and tocotrienols in oils of costa rican palm fruits: a comparison between six varieties and chemical versus mechanical extraction. J Agric Food Chem. (2017) 65:7476–82. doi: 10.1021/acs.jafc.7b02230
227. Feng J, Lu M, Wang J, Zhang H, Qiu K, Qi G, et al. Dietary oregano essential oil supplementation improves intestinal functions and alters gut microbiota in late-phase laying hens. J Anim Sci Biotechnol. (2021) 12:1–15. doi: 10.1186/s40104-021-00600-3
228. Batkowska J, Drabik K, Brodacki A, Czech A, Adamczuk A. Fatty acids profile, cholesterol level and quality of table eggs from hens fed with the addition of linseed and soybean oil. Food Chem. (2021) 334:127612. doi: 10.1016/j.foodchem.2020.127612
229. Cufadar Y, Gocmen R, Kanbur G. The effect of replacing soya bean oil with glycerol in diets on performance, egg quality and egg fatty acid composition in laying hens. Animal. (2016) 10:19–24. doi: 10.1017/S1751731115001950
230. Valkonen E, Venäläinen E, Tupasela T, Hiidenhovi J, Valaja J. Effect of Camelina sativa meal on fatty acid composition of egg yolk and sensory quality of eggs. Prceedings of the 12th European Poultry Conference. Verona: (2006).
Keywords: albumen quality, natural feed additives, laying hens, safe product, high-quality eggs
Citation: Obianwuna UE, Oleforuh-Okoleh VU, Wang J, Zhang H-J, Qi G-H, Qiu K and Wu S-G (2022) Natural Products of Plants and Animal Origin Improve Albumen Quality of Chicken Eggs. Front. Nutr. 9:875270. doi: 10.3389/fnut.2022.875270
Received: 14 February 2022; Accepted: 23 May 2022;
Published: 10 June 2022.
Edited by:
Celestino Santos-Buelga, University of Salamanca, SpainReviewed by:
Kai Zhan, Anhui Academy of Agricultural Sciences (CAAS), ChinaCopyright © 2022 Obianwuna, Oleforuh-Okoleh, Wang, Zhang, Qi, Qiu and Wu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kai Qiu, cWl1a2FpQGNhYXMuY24=; Shu-Geng Wu, d3VzaHVnZW5nQGNhYXMuY24=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.