
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Nutr., 03 January 2022
Sec. Eating Behavior
Volume 8 - 2021 | https://doi.org/10.3389/fnut.2021.760914
 Coleen Roger1,2*
Coleen Roger1,2* Adèle Lasbleiz2,3
Adèle Lasbleiz2,3 Maxime Guye1,2
Maxime Guye1,2 Anne Dutour3
Anne Dutour3 Bénédicte Gaborit3
Bénédicte Gaborit3 Jean-Philippe Ranjeva1,2
Jean-Philippe Ranjeva1,2Hypothalamus (HT), this small structure often perceived through the prism of neuroimaging as morphologically and functionally homogeneous, plays a key role in the primitive act of feeding. The current paper aims at reviewing the contribution of magnetic resonance imaging (MRI) in the study of the role of the HT in food intake regulation. It focuses on the different MRI techniques that have been used to describe structurally and functionally the Human HT. The latest advances in HT parcellation as well as perspectives in this field are presented. The value of MRI in the study of eating disorders such as anorexia nervosa (AN) and obesity are also highlighted.
Energy expenditure appears as a continuous process, while energy refilling through food intake is by nature discontinuous. In order to keep the body fat mass stable, a mechanism guaranteeing an efficient balance between energy expenditure and energy intake is therefore needed (1). Modern ways of life in developed countries tend to disturb such energetic balance. Indeed, this phenomenon is leading to drastic increases in the prevalence of eating disorders such as obesity, often referred as “globesity” (2) or AN, which holds the highest mortality rate of any mental illness (3). Although it is true that eating behavior is influenced by our lifestyle, it is the result of the complex association of genetic, metabolic and neurological alterations that have not been completely elucidated to date.
One key structure in the control of the energetic balance is the hypothalamus (HT), a small and complex deep brain structure. HT is composed of four sub-regions: the pre-optic, anterior, tuberal and mammillary regions. These regions project to the autonomous system, and the primary and associative systems. They each host multiple nuclei which bear different physiological-anatomical characteristics. Along the rostral-caudal axis, HT is composed by, (i) at the anterior level: the preoptic nuclei (lateral and medial), the anterior hypothalamic nuclei, the paraventricular nuclei (PVN), the anteroventral periventricular nuclei, the supraoptic nuclei and the suprachiasmatic nuclei; (ii) at the medial level: the dorsomedial nuclei, the ventromedial nuclei, and the arcuate nuclei; (iii) laterally: the lateral nucleus and (iv) and at the posterior level: the posterior nucleus and the mammillary bodies (Figure 1). Each of these nuclei has specific roles in homeostatic regulation of energy, food intake, thirst but also temperature, sexual dimorphism, sleep and circadian rhythm (6).
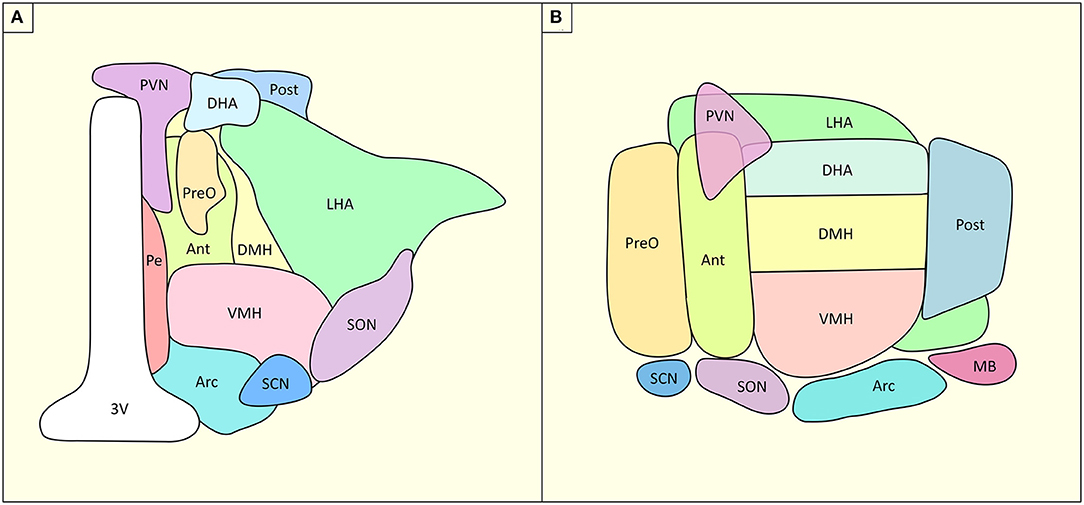
Figure 1. Structural organization of HT and its nuclei. (A) Coronal slice of the hypothalamus, (B) Sagittal slice of the hypothalamus. Ant, anterior hypothalamus; Arc, arcuate nucleus, DHA, dorsal hypothalamic area; DMH, dorsomedial hypothalamus; LHA, lateral hypothalamic area; MB, mammillary body; Pe, periventricular nucleus; Post, posterior hypothalamic nucleus; PreO, preoptic nucleus; PVN, paraventricular nucleus; SCN, suprachiasmatic nucleus; SON, supraoptic nucleus; VMH, ventromedial hypothalamus; 3V, third ventricle. Based on (4, 5).
Most of the knowledge regarding the functional role of HT nuclei in food intake processes have been derived from animal studies (7–9). Figure 2 is an attempt to summarize the current knowledge on such networks. Pioneer works in rats showed that bilateral lesions of the ventromedial HT (VMH) induced hyperphagia while bilateral lesions of the lateral hypothalamic area (LHA) provoked hypophagia (11). These early lesion approaches led to the definition of VMH as the “center of satiety”, responsible for the food intake inhibition, and LHA as the “center of hunger”, whose role is to stimulate food intake (11). The PVN sends information to the endocrine system while the LHA carries input to the cortex and limbic systems (12).
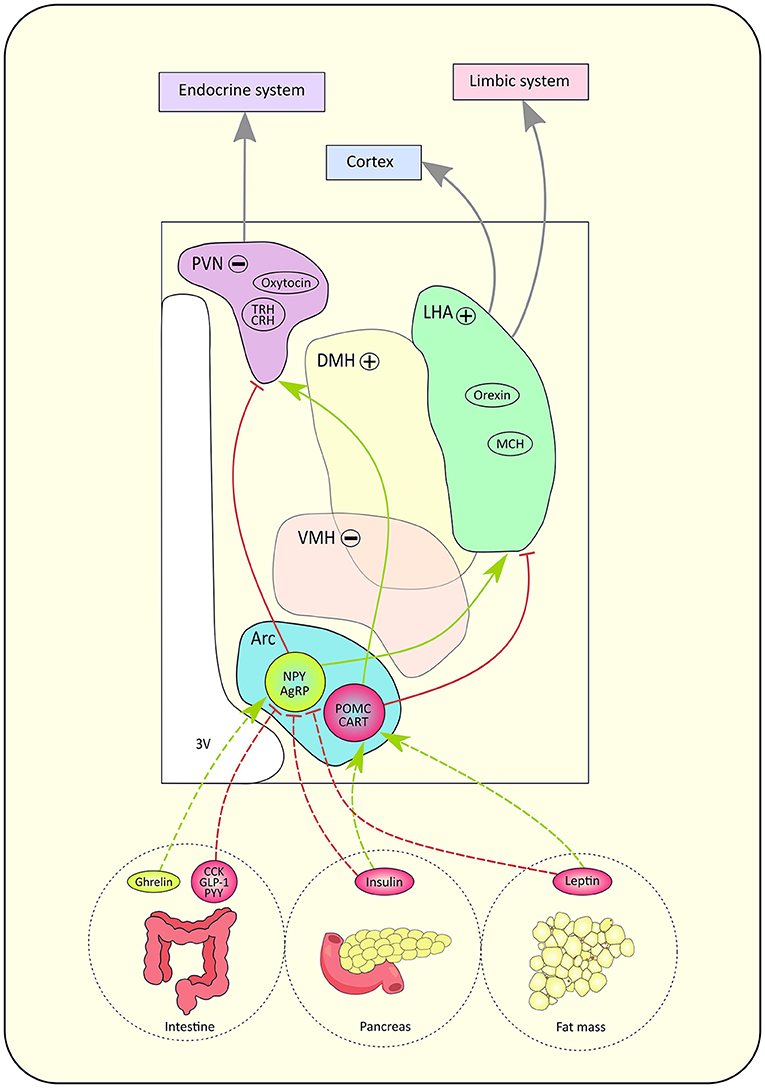
Figure 2. Schematic pathway of the lipostat and its peptidergic circuitry. According to hunger state, hormones are released by the intestine, pancreas and fat mass. Those signals directly influence neuronal activity in the hypothalamus through their action on antagonistic neurons located in the arcuate nucleus (Arc): orexinergic neurons expressing neuropeptide Y (NPY)/ agouti-related peptide (AgRP) and anorexinergic neurons expressing pro-opio-melanocortine (POMC)/ cocaine- and amphetamine- regulated transcript (CART). Gut hormones such as cholecystokinin (CCK), peptide YY (PYY), glucagon-like peptide 1 (GLP-1) inhibit NPY/AgRP neurons while ghrelin stimulates NPY/AgRP neurons. The pancreas secretes insulin while leptin is secreted by fat mass cells. Both hormones trigger the inhibition of NPY/AgRP neurons but the stimulation of POMC/CART neurons. Red arrows represent inhibitory connections, green arrows represent stimulatory connections, gray connections represent indirect pathways. The paraventricular nucleus (PVN) hosts neurons expressing oxytocin, thyrotropin-releasing hormone (TRH) and corticotropin-releasing hormone (CRH), while the lateral hypothalamic area (LHA) contains neurons expressing orexin and melanin-concentrating hormone (MCH). The '+' sign means the nucleus has an orexinergic effect on food intake when activated, whereas the '−' sign indicates that the nucleus has an anorexinergic effect on food intake when activated. Indeed, when activated, the dorsomedial hypothalamus (DMH) and lateral hypothalamic area (LHA) have an orexinergic action while the ventromedial hypothalamus (VMH) and paraventricular nucleus (PVN) have an anorexinergic action on food intake. Finally, the PVN sends information to the endocrine system while the LHA carries input to the cortex and limbic systems (24). 3V, third ventricle.
Advances in deciphering such mechanisms were also possible through the characterization of a network called the ‘hypothalamic lipostat', which refers to the neuronal pathways involved in the integration of information related to food intake (13). HT ensures energy balance in the long term by integrating factors circulating by humoral or vagal pathways such as satiety signals and adipokines [see (10) for review]. More recently, the “gut-brain axis” was also associated to the regulation of food intake. Indeed, serotonin-secreting cells found in the gut communicate with the brain through the vagus nerve (14, 15). Visceral signals from the gut activate neurons in the nucleus of the solitary tract (NTS), which stimulates regions known to mediate feeding behavior, such as the lateral parabrachial nucleus (PBN) localized in the pons (16). The gut-brain axis is also involved in food reward (17) and sugar preference (18).
Five hypothalamic nuclei have been associated to food intake regulation: the lateral, ventromedial, dorsomedial, PVN and arcuate nuclei (19). The arcuate nuclei hosts first order neurons, which impose antagonistic effects on food intake. On one hand, neurons expressing the neuropeptide Y (NPY) and neuropeptide agouti-gene related peptide (AgRP) project to second order neurons localized in the PVN to stimulate appetite (12). On the other hand, neurons expressing proopiomelanocortin (POMC) and the neuropeptide cocain and amphetamine related transcript (CART) project to second order neurons contained in the LHA in order to inhibit food intake (12), although POMC neurons' role seems more contrasted than previously described [see (20) for review]. Depending on energetic state, different types of signals activate or inhibit these neurons. Indeed, when lipid reserves are high, plasmatic leptin concentration increases (21). Leptin is an anorexigenic hormone, which acts on food intake in the long term. It reduces appetite by increasing the activity of POMC/CART neurons and inhibiting NPY/ AgRP neurons (22). The regulation of food intake is ensured in the short and medium term by insulin as well. After a meal, insulin secretion rises and exercises an anorexigenic effect (22). Intestinal peptides also modulate appetite. For example, cholecystokinin (CCK), PYY 3–36 and glucagon-like peptide-1 (GLP-1) stimulate satiety whereas ghrelin promotes food intake (22). New hypothalamic regions have recently been shown to be involved in food intake mechanisms. For example, the tuberal lateral nucleus seems to promote an orexigenic effect on food intake by inhibiting the PVN through somatostatin (SST) interneuron signaling (23).
In addition, food intake is subjected to a non-homeostatic control influenced by the palatability of food as well as the environment and emotional state (24). Second order hypothalamic neurons constitute intermediate relays to some regions of the limbic system such as the amygdala, the hippocampus, the insula, the striatum and the orbitofrontal cortex (OFC). These structures, already known to be involved in emotions, memory or addictions, seem to have an important role in modulating eating behavior (22). Furthermore, the limbic system undergoes an inhibitory control from the prefrontal cortex which promotes meal termination (25).
HT dysfunction has long been implicated in eating disorders such as obesity (26) and anorexia nervosa (AN). With 650 million people affected worldwide in 2016, obesity is defined by a body mass index (BMI) over 30, resulting from an abnormal fat accumulation, which presents a risk to health (WHO). Rat studies showed that obesity induced by high fat diet led to a chronic low-grade HT inflammation with insulin and leptin resistance which promoted weight gain (27, 28). Besides, obese individuals showed T2 hyperintensity in the mediobasal HT, reflecting gliosis (29). More generally, HT appears to be an open window for some saturated fatty acids (SFAs) such as palmitate passing through the partially incomplete blood brain barrier in the arcuate nuclei and the median eminence (30–32). Furthermore, it was shown that the presence of SFAs in the HT induces neuronal responses leading to stimulated inflammatory cytokine expression, reactive oxygen species (ROS) production, endoplasmic reticulum stress and microglia recruitment [see (33) for review]. These findings collectively support the hypothalamic inflammatory hypothesis. Malfunction of the reward system has also been associated to a decreased availability of dopamine D2 receptor (D2R) proportionally to the BMI of patients (34).
Concurrently, AN affects approximately 4% of females and 0.3% males during their lifetime (35). This pathology is characterized by an important self-inflicted restriction of food intake leading to a BMI below 17.5, intense fear of gaining weight, as well as altered body perception influencing self-esteem and causing denial of the severity of current thinness (Diagnostic and Statistical Manual of Mental Disorders 5th Ed. (DSM-5) criteria http://www.dsm5.org/meetus/pages/eatingdisorders.aspx). In comparison to obesity, the cellular mechanisms and physiological substrates of AN have not been as thoroughly characterized (36). However, the administration of D2/3 receptors (D2/3R) antagonists to a rodent model of AN, the “activity-based anorexia” (ABA) model, was found to reduce weight loss and hypophagia, and increase survival (37). Furthermore, D2/3R density or affinity seem to increase in AN patients (38). As it is the case for obesity, this result support the existence of a dysfunction of the dopaminergic system in AN. Therefore D2/3R appear to be interesting therapeutic targets for AN. Furthermore, serotonin levels have been associated to almost all the behavioral changes observed in AN patients, such as extreme dieting weight loss, hyperactivity, depression/anxiety, self-control, and behavioral impulsivity (39). However, one of the most interesting hypothesis on the pathophysiology of AN is that chronic food restriction alters the ghrelin signaling pathway and leads to the development of abnormal behaviors such as addiction to food starvation (40).
Overall, translation of the cellular mechanisms observed in animal models to Human is not easy and straightforward. In that context, neuroimaging has appeared as a game changer to explore food intake and the consequences of its deregulation from an anatomical, functional or even metabolic point of view (41, 42). However, in order to define the precise function of each hypothalamic nucleus in human, it is necessary to clarify their location and their respective roles within the different functional brain networks. Given the small size of the HT, its location and the large number of hypothalamic nuclei, fine characterization of HT is still ongoing. This paper gives an overview of the different neuroimaging studies which have attempted to characterize the HT, as well as the different ways to progress in the field. We will also highlight the undeniable value of MRI in the study of eating disorders such as AN and obesity.
HT is generally poorly defined or individualized in the numerous subcortical brain atlases proposed in the literature (43–48). Despite the fact that brain MRI stands out as the most suitable tool in the study of the central nervous system, the poor contrasts between hypothalamic nuclei has led to consider the HT as a single homogeneous structure, which prevents the precise morphological characterization of each nuclei. Moreover, hypothalamic MRI is subject to artifacts as a consequence of the vicinity of the HT with sinuses and bones (41). Nevertheless, standardized guidelines have been proposed for manual HT delineation as a single structure (49). Recently, a few MRI studies aimed at characterizing hypothalamic sub-regions based on anatomical references. The first neuroimaging atlas of the HT was based on 3D-MRI acquired at 1.5T in twenty healthy subjects (50). Thus, by combining anatomical, histological and magnetic resonance images, the human HT was parcellated into PVN, VMH, arcuate nuclei, and the posterior hypothalamic area (50).
One year later, combination of in vivo 1.5T MRI and ex vivo 7T MRI data allowed Makris and co-workers to individualize five hypothalamic sub-regions: the lower anterior HT, the upper anterior HT, the lower tuberical HT, the upper tuberical HT and the posterior HT (51). This approach was sufficiently robust to perform quantitative morphometry of HT sub-parts, showing an overall larger HT in men relative to women after correction for brain size. Indeed, significant differences in sizes were reported bilaterally in the five HT subregions, especially in the tuberal region which includes the LHA, infundibular, PVN, VMH, and supraoptic nucleus (51).
A more detailed parcellation was proposed by Lemaire and collaborators, based on a 1.5T MRI database (52). Using precise anatomical landmarks, especially the white matter (WM) bundles neighboring the HT, most internal hypothalamic structures were located: posterior nucleus, dorsomedial nucleus, infundibular nucleus, lateral nucleus, mammillary body, PVN, preoptic nuclei, suprachiasmatic nucleus, supraoptic nucleus, tuberomamillaris nucleus and VMH (52).
More recently, the first digital human brain atlas to gather neuroimaging, high-resolution histology, and chemo-architecture was created (53). This atlas has the particularity to be based on 7T MRI and 3T diffusion-weighted imaging (DWI) scans, as well as 1,356 large-format cellular resolution (1 μm/pixel) Nissl and immunohistochemistry anatomical plates from a complete adult female brain. Using this strategy enabled the description of every hypothalamic nuclei, including the most challenging structures to describe such as the suprachiasmatic nucleus.
However, a step forward was reached recently by Neudorfer and colleagues (4) who proposed a template based on a large set of 3T data composed by 990 3D-MPRAGE T1-weighted MRI (isotropic voxel size 1 mm3) of healthy controls from the Human Connectome Project database (https://www.humanconnectome.org/study/hcp-young-adult/document/1200-subjects-data-release). Using state-of-the-art multiscale template-building methods including non-linear registration procedures [ANTS, http://stnava.github.io/ANTs (54)], the authors proposed the first complete high-resolution in-vivo anatomical atlas of the human HT with a spatial resolution up to 0.25 × 0.25 × 0.25 mm voxel size. Based on this high resolved template, three experts were able to delineate the thirteen hypothalamic nuclei and individualized them. Projection to individual subjects using non-linear registration showed differences in volumes of hypothalamic nuclei which are dependent on brain hemisphere and subject gender in healthy subjects. These improvements in delineation and quantification were highly dependent on the artificially increased spatial resolution brought by the use of a large number of exams, which limited partial volume effects and allowed for a better contrast between HT nuclei.
In this context, spatially more resolved raw data, brought by in vivo ultra-high field (UHF) 7T MRI, should play a major role in the finer characterization of HT nuclei at the individual level. Indeed, it should open new opportunities to better understand individual structural organization of tiny HT sub-structures. Thus, unbiased MP2RAGE acquisitions performed at 7T (55) should provide T1 weighted images and unbiased quantitative T1 maps at high spatial resolution (typically 0.6 x 0.6 x 0.6 mm) and allow for a better description of HT sub-structures. As an example, projection of the Neudorfer hypothalamic atlas to a home-made 7T MP2RAGE Uni-Den template on a single subject is presented in Figure 3.
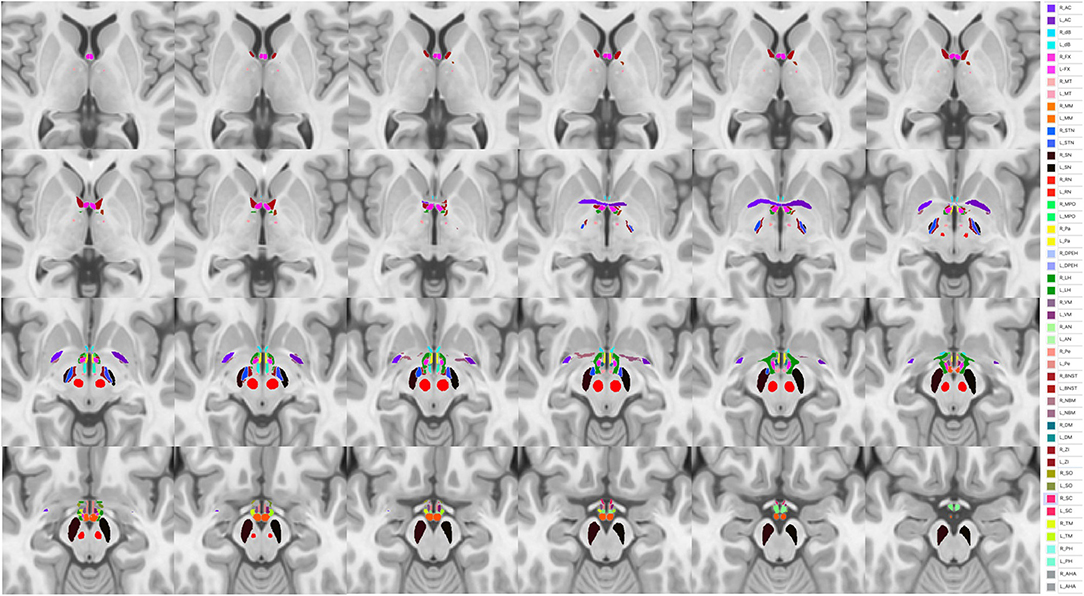
Figure 3. Projection of the Neudorfer hypothalamic atlas into a homemade 7T MP2RAGE Uni-Den template. Ac, anterior commissure; AHA, anterior hypothalamic area; AN, arcuate nucleus; BNST, bed nucleus of stria terminalis; dB, diagonal band of Broca; DM, dorsomedial hypothalamic nucleus; DPEH, dorsal periventricular hypothalamus; FX, fornix; LH, lateral hypothalamus; MM, mammillary bodies; MPO, medial preoptic nucleus of the hypothalamus; MT, mammillothalamic tract; NBM, nucleus basalis of Meynert; Pa, paraventricular nucleus; Pe, periventricular nucleus; PH, posterior hypothalamus; RN, red nucleus; SC, suprachiasmatic nucleus; SN, substantia nigra; SO, supraoptic nucleus; STN, subthalamic nucleus, TM, tuberomammillary nucleus; VM, ventromedial nucleus; ZI, zona incerta.
Figure 4 compares two different parcellation processes of the human HT using MRI.
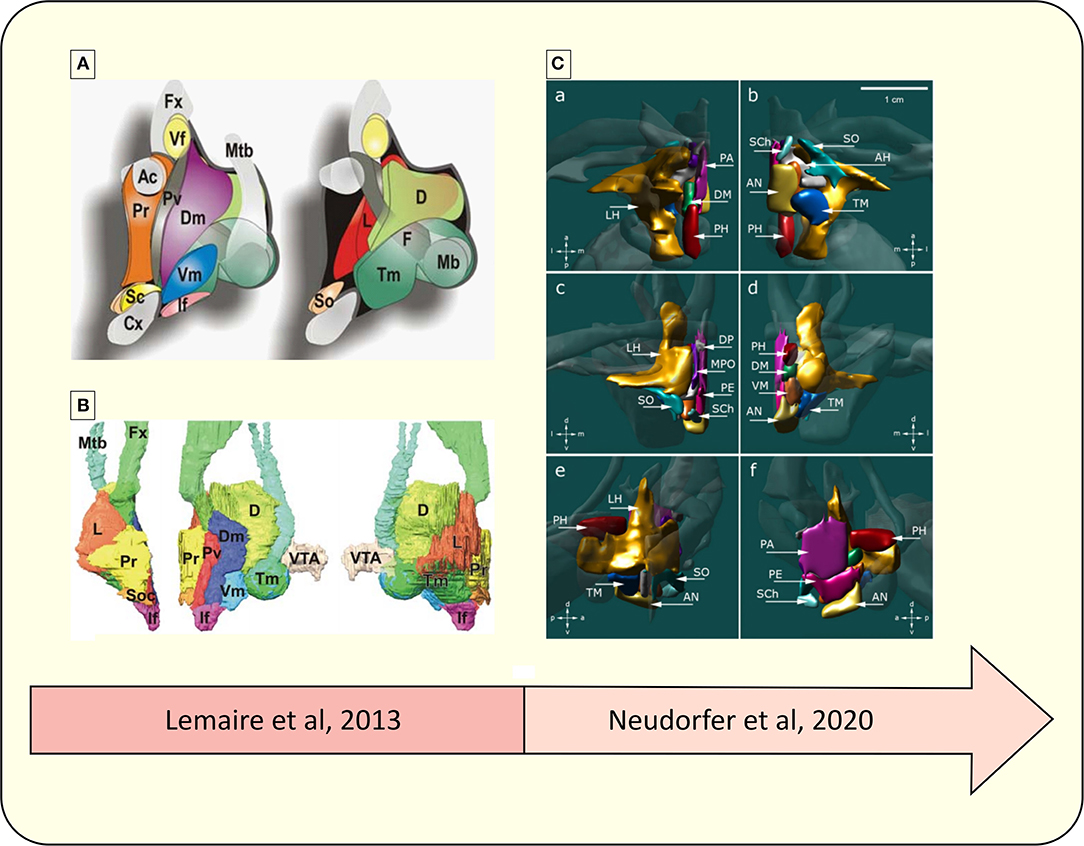
Figure 4. Representative diagram of the advances in the parcellation of the hypothalamus. (A) Schematic drawing of hypothalamic nuclei, lateral view from the midline. Left, nuclei directly located under the wall of the third ventricle; right, more deeply located nuclei up to the lateral region (52). (B) 3D overview of hypothalamic nuclei, constructed from a 3D high-field MRI data set. Left, frontal view; middle, lateral view; right, medial view. VTA, ventral tegmental area; Ac, anterior commissure; D, dorsal or posterior nucleus: Dm, dorsomedial nucleus; Fx, fornix; F, fornix nuclei; If, infundibular (arcuate) nucleus; L, lateral nucleus; Mb, mammillary body; Mtb, Fmillo-thalamic bundle; Cx, optic chiasma; Pv, paraventricular nucleus; Pr, preoptic nuclei; Sc, suprachiasmatic (ovoid) nucleus; Vf, ventricular foramen; So, supraoptic (tangential) nucleus; Tm, tuberomamillaris (mamilloinfundibularis) nucleus; Vm, ventromedial (tuber principal) nucleus (52). (C) 3D reconstruction of hypothalamic nuclei and their neuroanatomical relationships. (a) top view, (b) bottom view, (c) frontal view, (d) occipital view, (e) sagittal view depicting the outer surface of hypothalamic nuclei, (f) sagittal view depicting the inner surface of hypothalamic nuclei. AH, anterior hypothalamic area; AN, arcuate nucleus; DP, dorsal periventricular nucleus; DM, dorsomedial hypothalamic nucleus; LH, lateral hypothalamus; MPO, medial preoptic nucleus; PA, paraventricular nucleus; PE, periventricular nucleus; PH, posterior hypothalamus; SCh, suprachiasmatic nucleus; SO, supraoptic nucleus; TM, tuberomammillary nucleus; VM, ventromedial nucleus (4).
In line with these technological improvements regarding acquisition and post-processing methods, the finer structural characterization of HT nuclei using UHF MRI should play a major role in the depiction of structural abnormalities in the different HT nuclei of patients with eatingdisorders.
Very few structural hypothalamic abnormalities have been observed in obesity, while whole brain morphometry showed subtle gray matter (GM) atrophy in some regions of the frontal lobe implicated in behavioral control such as the post-central gyrus, the frontal operculum as well as the putamen which is involved in the regulation of taste and reward (56–58). BMI has been positively associated with GM volume in the right middle occipital gyrus involved in visual processing and systematically activated in food-cue task (57). Such high GM volumes of occipital regions of obese individuals could be associated to the selective attention bias toward appetitive food cue (59) and could constitute a predictor of future weight gain (57). In contrast, total body fat appeared negatively correlated with volumes of subcortical GM structures, with greater associations in men compared to women (58).
On the other hand, a lot of structural hypothalamic abnormalities were found in AN. HT GM volume has been found to be correlated with BMI in AN patients (60). Moreover, while a higher decrease in GM volume was observed in the cerebellum of AN patients with longer disease duration (> 9 years), lower hypothalamic GM was found significant only in AN patients with shorter disease duration (<3 years) (60).
Furthermore, a meta-analysis of voxel-based morphometry in AN revealed regional GM decreases in the left HT and other areas implicated in somatosensory perception and appetite, such as the left inferior parietal lobe, right lentiform nucleus and right caudate (61). Nevertheless, and surely due to the lack of spatial resolution, the largest variations were observed outside the HT, with enlargement of ventricles, cortical sulci (62), interhemispheric fissure and generalized volume reduction (63) associated with cortical thinning (64). GM atrophy has been specifically observed in somatosensory and reward regions which could explain abnormal reward responses to food stimuli and distortion of body perception (61). However, the nutritional status in AN affects cortical folding (65), and a longitudinal study on adolescent and young adults with acute and recovered AN patients reported that weight restoration seems to have a rapid reversible effect on cortical thinning (66).
The appetitive and self-control brain networks underlying food intake regulation can be represented by three interconnected sub-networks. First, the “lipostat” regulating energy balance signals including the HT, the ventral tegmental area (VTA) and the Substantia Nigra (SN) participating in the dopaminergic system. Second, the “limbic system” receiving sensory inputs and programming action to fulfill energy. This sub-network is composed by the amygdalar and hippocampal formations (memory, emotion, learning), the insula (ingestive cortex), the OFC and the ventromedial prefrontal cortex (VMPFC) (eating value), the accumbens nuclei and the striatum (motivation and value to action). Finally, the “control system” is composed by the dorsolateral prefrontal cortex (DLPFC) and the anterior cingulate cortex (ACC) which participates in self-regulation (67). Through the participation of its numerous nuclei and their WM connections, the HT appears as a central node for each of these different sub-networks.
Diffusion-weighted MRI has helped to better characterize in vivo this organization. Kamali and coworkers showed that the stria terminalis connects the amygdala to the anterior HT (68). The stria terminalis projects to the mammillary body region as well as multiple septal and hypothalamic nuclei, which could not be identified due to the lack of spatial resolution (68). Atlases of human neuroanatomy reported that the ventral amygdalo-fugal fibers originating from the basolateral and central nucleus of the amygdala, project to the nucleus accumbens, basal forebrain, medial dorsal nucleus of the thalamus (69). The ventral amygdalo-fugal fibers also reach the septal nuclei, LHA, lateral preoptic, anterior and tuberal nuclei of the HT (70, 71).
The fornix originates from the hippocampal formation and links the hippocampus to major structures involved in the homeostatic (HT), hedonic valence (amygdala) and reward control (OFC, nucleus accumbens) of food intake (12, 68, 72). The fornix fibers were also shown to terminate in the region of mammillary bodies and septal nuclei (68). The pathway connecting the hippocampus to the HT might be involved in the motivated and conditioned pattern of food intake (73).
Recently, Kamali's team was the first to reconstruct the dorsal thalamo-hypothalamic tract, revealing in the meantime the existence of a direct connection from the dorsal thalamus to the ventral hypothalamic nuclei (74). The authors hypothesized that since the thalamus constitutes a hub for somatosensory information, the dorsal thalamic nuclei could carry sensory information from the thalamus to the limbic system, in particular to the HT. These findings are in line with studies done in rats suggesting the association of the thalamus and the HT with eating behavior, food motivation/ reward and the circadian rhythms (75–77).
A high-spatial and angular resolution diffusion weighted tractography technique revealed the detailed pathway of the parieto-occipito-hypothalamic tract. The parieto-occipito-hypothalamic tract seems to allow for visuosensory information to be carried from the parietal and occipital cortices to the limbic system, especially the HT and the bed nucleus of the stria terminalis. This finding stresses the point that many limbic functions, such as hunger, fear or circadian rhythm rely on visuo-sensory information (78).
The cerebellum might be involved in the locomotor and foraging aspects of eating behavior (79). Lesional animal studies showed that ablation of the cerebellar nuclei induced degeneration in the HT, reflecting the existence of a cerebello-hypothalamic connection (80, 81). In human, high-resolution diffusion-weighted tractography at 3T revealed that the cerebellar-ponto-hypothalamic tract is responsible for the reciprocal communication between the cerebellum and the limbic system, especially the HT and the septum (82). More recently, a direct cerebello-hypothalamic tract was described in Human. Indeed, DTI results reported fibers projecting from the cerebellar nuclei to the contralateral anterior HT and ipsilateral posterior HT (83). The cerebellar-hypothalamic connection seems to allow for the convergence of non-limbic information to the limbic system through the hypothalamic and septal nuclei. Numerous animal studies support the modulatory action of the cerebellum on food intake through its action on the HT (84). Therefore, these findings suggest that the cerebellar-ponto-hypothalamic tract might carry the modulatory action of the cerebellum on hypothalamic feeding centers.
Finally, structural connectivity derived from DTI tractography was used to parcel the HT by studying the orientations and projections of the different efferent and afferent WM fibers connecting the HT to other brain areas. Lemaire and co-workers characterized parts of the macroscopic internal structure of the HT and its extrinsic connectivity with deep brain and cortical regions (85). Thus, preoptic, anteroventral and lateral hypothalamic regions were shown to predominantly project to the right frontal hemisphere, largely involved in behavioral control (85). Figure 5 summarizes the structural extra-hypothalamic networks of food intake discussed above. Using color coded directional fractional anisotropy (FA) maps of the HT, three different subregions of the HT were defined: an anteromedial region with dorsoventral diffusion direction, a posteromedial region with rostro-caudal direction, and a lateral region with mediolateral direction (86). The anteromedial region was assigned to the PVN, anterior, and dorsomedial hypothalamic nucleus and part of the LHA, the lateral region to the VMH and supraoptic nucleus and finally the posteromedial region to the suprachiasmatic, infundibular, VMH, posterior hypothalamic, medial and lateral mammillary nucleus.
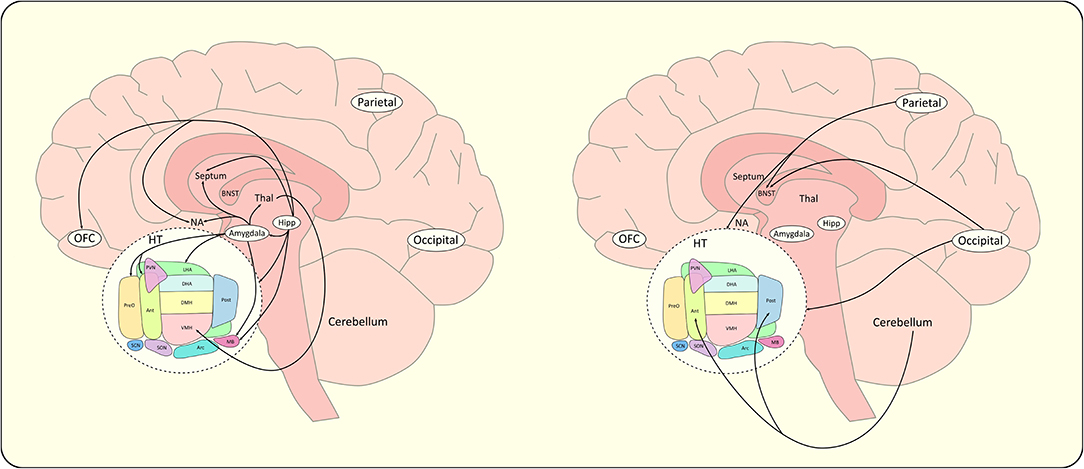
Figure 5. Schematic representations of extra-hypothalamic networks of food intake. For clarity concern, extra-hypothalamic connections reported in the literature have been divided in an appetitive network (left) and a control network (right). BNST, bed nucleus of the stria terminalis; Hipp, hippocampus; HT, hypothalamus; NA, nucleus accumbens; Parietal, parietal cortex; Occipital, occipital cortex; OFC, orbitofrontal cortex; Thal, thalamus. The schematic sagittal slice of the hypothalamus (HT) includes; Ant, anterior hypothalamus; Arc, arcuate nucleus; DHA, dorsal hypothalamic area; DMH, dorsomedial hypothalamus; LHA, lateral hypothalamic area; MB, mammillary body; Pe, periventricular nucleus; Post, posterior hypothalamic nucleus; PreO, preoptic nucleus; PVN, paraventricular nucleus; SCN, suprachiasmatic nucleus; SON, supraoptic nucleus; VMH, ventromedial hypothalamus.
Obesity has been associated with abnormal WM volume and microstructure as well as microstructural brain diffusion changes in HT and other structures involved in satiety and hunger (87). Indeed, DWI studies revealed that the apparent diffusion coefficients (ADCs) of HT, hippocampal gyrus, amygdala, insula, cerebellum and midbrain were significantly increased in obese patients. More specifically, hypothalamic longitudinal diffusivity λ1, a DTI metric related to axonal integrity, has been found to be negatively correlated with fat mass in obese subjects (88). Furthermore, altered hypothalamic microstructure, characterized by a higher mean diffusivity (MD) within the HT, has been associated with a higher BMI (89). Indeed, alteration in the hypothalamic microstructure visible on DTI has been associated with the presence of inflammatory factors, such as C-reactive protein (CRP) or lipopolysaccharide-binding protein (LBP) (88). However, there is no consensus regarding the sign of the correlations between BMI and WM volumes. Most studies have established an increase of WM volume in the frontal, temporal, parietal and occipital lobes, fusiform and parahippocampal gyri, brainstem and cerebellum in obese relative to lean participants (90, 91). On the contrary, a decrease in WM volume has been reported in overweight and obese older adults within the frontal lobe, anterior cingulum and corona radiata (92). The percentage of total body fat has been positively associated with global FA in men and women, and negatively associated with global MD in women. These results suggest that obese women globally present higher integrity but lower magnitude of WM microstructure, showing differential gender effects of obesity on WM microstructure (58). Generally, the association between markers of obesity and diffusion parameters seems stronger in women than in men (93). However, regionally, obese individuals demonstrate a negative correlation between BMI and FA in the corpus callosum and fornix (94). Furthermore, BMI was found to be negatively associated with the microstructural integrity of numerous WM bundles such as callosal and limbic tracts (95). Interestingly, obese subjects show partial recovery of WM volumes after only 6 weeks of dieting (90).
Alteration of WM microstructure has also been observed in AN patients (96). DTI studies reported low FA and/or high MD of the fornix, amygdala and the prefrontal region in AN (97). Low FA was also found in the lateral part of the cerebellum of AN patients (84, 98). This result is in line with animal studies stating that cerebellar nuclei would modulate lateral hypothalamic response to satiety and hunger through interaction with gastric vagal nerves (99).
Using a probabilistic tracking algorithm, the same team who implemented the first 3D-MRI atlas of the HT (50) found a significant reduction in the number of fibers in the arcuate nucleus of AN patients and leanness controls compared to normal-weight controls but higher connectivity of LHA in AN compared to leanness and normal-weight controls (42). In the same study, high basal levels of HT glutamatergic markers (Glutamate-Glutamine/Cr ratios) derived from magnetic resonance spectroscopy (MRS) dropped abnormally following food intake in AN compared to controls (42). Therefore, lower BMI seems to be associated to a specific altered structural connectivity of the arcuate nucleus and LHA as well as a different HT metabolic response to food intake (42).
Moreover, AN seems to be characterized by a disrupted fronto-accumbal structural connectivity since a longitudinal neuroimaging study revealed that the abnormal increase in structural connectivity within the fronto-accumbal network found in AN patients persisted even after weight recovery (100).
However, the full structural connectome of HT nuclei is far from being entirely described, both at the intra-HT and extra-HT connectivity levels. DTI techniques cannot provide crucial information such as the afferent or efferent nature of fiber tracts. Only significant improvements in DWI acquisitions and models including high angle resolution diffusion imaging (HARDI), orientation diffusion functions, multi-shell acquisitions and/or UHF protocols increasing spatial resolution (101), and correlation with post-mortem data should allow for the full characterization of these complex hypothalamic networks. In that sense, improvement of post-processing pipelines validated in large cohorts (102) will help to correct distortion artifacts (eddy currents, susceptibility effects, motions…) in UHF DWI and will allow for high quality data to be accurately overlaid with HT nucleus structural atlases.
Appetite networks have also been explored through the prism of functional connectivity (FC) by using resting-state functional MRI (rs-fMRI). rs-fMRI consists in recording temporal correlations of low frequencies blood oxygen level dependent (BOLD) fluctuations between various brain areas during a “resting” period (103). This technique is commonly used to identify functional networks when no task is performed (104).
Such paradigm was used to explore the effect of fasting and satiation on the FC patterns in healthy subjects (105). Interestingly, variations of blood glucose levels between scans accounted for numerous altered functional connectivities. Indeed, FC between the left HT and inferior frontal gyrus strengthened during fasting whereas FC increased between the right HT and the superior parietal cortex during satiation (105).
In healthy subjects, a depleted energy state seems to favor connectivity of the HT with the inferior frontal gyrus, which is implicated in suppressing the desire for food and preventing temptation to eat (106, 107). On the contrary, satiation is associated with a stronger connectivity between the HT and the superior parietal cortex, which is hypothesized to be involved in decreasing the eating drive by being less distracted by thoughts of food or eating (105). Thus, the HT connectivity seems to be lateralized and to depend on the energy state.
Hunger may increase effective connectivity (EC) from posterior insula to anterior insula while EC from the anterior insula to posterior insula may be decreased (108). Therefore, individuals' energy states affect the strength and directionality of connections between sub-regions of the insula. However, this same study failed to provide evidence of an effect of hunger on the connection between the HT and the insula.
Seed-based resting-state connectivity maps derived from a 3T MRI database of 49 healthy subjects were generated from the LHA (109) considered as the hunger center, and from the medial HT (including the VMH) considered as the satiety center (11, 110). The LHA was shown to be strongly connected to the dorsal striatum (caudate, putamen), anterior cingulum, thalamus, and frontal operculum, regions involved in goal-directed behavior (111), and to the lateral OFC implicated in coding stimulus reward value (112). The LHA was also shown to receive preferentially inputs from the lateral OFC (113). In contrast, the medial HT showed stronger connections with the ventral striatum (nucleus accumbens), the medial OFC and the occipital gyrus, regions involved in reward and motivation (114) and with the medial OFC (113).
Improvement of statistical power through a better temporal resolution and higher spatial resolution available with UHF MRI scanners (115) should be a game changer in the study of functional reorganization of HT networks in healthy subjects and eating disorders patients.
Functional connectivity is altered in obese patients when compared to healthy subjects. On one hand, high BMI subjects show a decreased global brain connectivity (GBC) in the lateral and prefrontal cortices, the insula and the limbic system (116), and a reduced FC between HT and regions involved in cognitive control such as the superior parietal lobule (117). On the other hand, obese patients show increased global brain connectivity in the visual, parietal, and premotor cortices (116). Functional connectivity of the LHA with the ventral striatum, anterior insula and OFC were found to be enhanced in obese compared to healthy individuals (118). In addition, high BMI subjects show increased FC between HT and regions involved in motivation processes (insula, thalamus, globus pallidus, cerebellum) (117), and increased FC between medial HT and regions involved in reward processing such as OFC and nucleus accumbens (109). Therefore, BMI is positively associated with FC between HT and brain regions involved in motivated feeding, reward processes and motricity, while FC between HT and areas involved in cognitive control of food intake is negatively correlated to BMI (117).
Modulation of hunger states showed that obese patients exhibit higher connectivity between the HT and the medial prefrontal cortex (PFC) and the dorsal striatum when fasted compared to lean subjects (119). Furthermore, in contrast to lean subjects, FC between HT and the PFC did not decrease when fed, which could explain the increased craving for food obese individuals experience during a fast (119). Additionally, lower FC was found between the HT and brainstem in response to glucose administration in obese compared to normal-weight control (120). These results stress the presence of FC alterations between the HT and regions involved in the inhibition of the eating drive, thus limiting the control of the balance between energy intake and expenditure (119). However, weight loss surgery allowed reversibility of FC abnormalities within HT networks, with restored lower FC between the HT and the OFC and somatosensory cortices (121).
Even though numerous resting-state fMRI studies were conducted in AN, most of them did not look at HT connectivity. Indeed, they rather focused on altered local- and large-scale- connectivity of “conventional” resting state networks (RSN) such as the default mode network (DMN), the visual network and auditory networks, as well as the reward and cognitive control systems. The DMN is a network associated with stimulus-independent thoughts and self-reflection (122). Within this network, increased FC was observed between the dorsal ACC and regions such as the precuneus and the retro-splenial cortex, which was positively correlated with body shape questionnaire scores (123). Such FC increase could represent a supplementary effort of the cognitive control system to restrain appetite (124). Data driven approach using independent component analysis (ICA) showed increased FC between the angular gyrus and the frontoparietal network in AN, which was also correlated with stronger cognitive control scores (125). In AN patients compared to healthy subjects, EC was increased from the bilateral orbitofrontal gyrus to the right IFG and from the bilateral insula to the left IFG while EC was decreased from the right inferior frontal gyrus to the mid-cingulum (109). These findings suggest that connectivity within the control network is reduced whereas connectivity between salience-related systems is increased in AN.
The ventral attentional circuit was also shown to be abnormally highly connected in AN, especially the inferior frontal gyrus (126). Since the ventral attention circuit allows for the implementation of appropriate behavioral responses, especially when an unexpected or salient stimulus occurs, its alteration could be responsible for lower interoceptive sensations awareness and struggles in adapting one's behavior to environmental needs (127). Visuospatial and auditory processing involved in self-body image have also appeared to be disturbed in AN patients, with reduced FC within the lateral visual cortices and the auditory network (104), and between the sensorimotor and visual circuits (128).
Furthermore, hyperconnectivity in AN of the dorsal caudate with prefrontal regions involved in motor and cognitive functions (129), have been interpreted as altered habit learning deficiencies responsible for the maintenance of AN symptoms (130).
Within the reward system, AN patients showed increased FC between the left nucleus accumbens and the left medial OFC compared to control subjects (100). Moreover, the connectivity within the ventral fronto-striatal circuit also seems to be dysfunctional in AN. Furthermore, hypoconnectivity between nucleus accumbens and superior frontal gyrus, found in AN, correlated with greater cognitive eating disorder symptoms (Eating Disorder Examination Global scores). Collectively, these findings suggest that FC is altered in AN in key brain regions involved in reward processing, homeostatic communication and habit.
Half of the AN individuals who received medical care need to be re-hospitalized within a year of discharge (131). Given the persistence and high risk of relapse of this long-term disease, the study of the resting state functional connectivity (rsFC) in weight-recovered AN individuals is particularly important.
Complete restoration of rsFC in weight-recovered AN individuals for at least 12 months has been observed in the PFC, sensorimotor, precuneal, insular, left parietal and left temporal cortices (132). In contrast, altered FC persists after 6 months recovery in the frontoparietal network with reduced FC with the DLPFC and increased FC with the angular gyrus and between the precuneus and DLPFC (133). Overall, rsFC alterations in AN tend to normalize after weight restoration, with the noticeable exception of the cognitive control network, which could in part explain the high risk of relapse of this disease.
However, more resting-state studies are needed since findings on the same networks are inconsistent (134). Moreover, undernutrition was found to alter reward and habit learning related networks (135) as well as to influence rsFC results (108). Therefore, rsFC AN studies might observe the consequence of starvation and not the neuronal correlates of AN (136). On the other hand, it can be hypothesized that a malfunction of the reward system could be the trigger of AN in genetically predisposed individuals exposed to certain environmental conditions such as emotional shock or diet [see (137) for review]. Furthermore, most studies did not take into account the different subtypes of AN (restrictive and binge eating/purging), although they were not found to influence results (100).
Different methods of functional imaging have been used to study appetite networks. Tracking variations of BOLD signal (fMRI), regional cerebral blood flow (rCBF) (PET or fMRI) or ADCs (diffusion weighted MRI) have been conducted (i) during different hunger states, (ii) before, during and after glucose (or other nutriment) ingestion, or (iii) during different food cue image display paradigms, including (or not) different satiety states.
Hunger was shown to enhance BOLD signal and rCBF within the HT, the ACC, the insula, the parahippocampal gyrus, and the hippocampus, core regions of the limbic system (138). Concurrently, during satiation, increase in rCBF was observed in the VMPFC, the DLPFC, and the inferior parietal lobe, which are all involved in the control of food intake (138).
Glucose ingestion in healthy subjects induce a dose-dependent decrease in the BOLD signal within the PVN and the VMH, which have an anorexigenic role (139). Besides, oral glucose intake triggers two peaks of response in healthy subjects: one just after the ingestion, followed by another peak approximately 10 min later (140). This delayed response was found to be associated with a negative response in the medial HT, and the fasting plasma insulin concentrations (140) reflecting the important role of HT in the modulation of insulin secretion to regulate glucose levels (141).
The type of ingestion may also play a role since both sweet taste and energy content are necessary to trigger a hypothalamic response (142). However, the limited spatial resolution of these techniques may prevent the ability to detect such changes in HT during glucose ingestion due to the different roles and activity patterns (stimulatory or inhibitory) of close-by hypothalamic nuclei (143).
However, more recently, Osada and colleagues described the glucose metabolism of individual major HT nuclei by using areal parcellation based on areal profiles of resting state functional connectivity (144). They observed a decreased activity in the VMH but an increased activity in the LHA between 10 and 40 min after glucose ingestion. Furthermore, the decrease in activity found in the arcuate nucleus was followed by a rise in blood insulin during the first 10 min after glucose ingestion (144).
Sun and colleagues demonstrated that the amygdala response to the administration of a milkshake predicted weight in satiated but not in hungry individuals (145). Looking at EC, sated subjects showed an unidirectional gustatory input from basolateral amygdala to the HT, whereas during hunger, HT drove bilateral connectivity with the amygdala (145).
Another popular neuroimaging method commonly used to investigate appetite networks is food-cue reactivity paradigms, which consist in presenting images of food under various forms and energy-content as well as non-related food images. Comparison of BOLD signals between conditions helped to decipher appetite processes. Indeed, high-calorie food were shown to trigger a significant bilateral activation of the HT (146, 147), the medial and DLPFC, the medial dorsal thalamus, the cingular cortex, and the cerebellum compared to the non-food condition (146). The cerebellum was also shown to be activated when viewing high fat relative to low fat food images (146). Furthermore, numerous animal studies revealed that the cerebellum modulates the activity of the LHA through its connections with gastric vagal nerves involved in hunger and satiety signaling (99).
Similarly to results obtained during the glucose ingestion, presentation of rewarding food tastes and images during satiation decreased BOLD signal in the reward system, including the HT, vmPFC, nucleus accumbens, OFC, and insula but increased BOLD response in the DLPFC, which is part of the control network (89). Similar manipulation of food types through hunger states showed that presentation of high-energy food during fasting might activate the HT and ventral striatum to a greater extent compared to low-energy food (148). fMRI is also used in the study of emotional connotation of food, which could be a factor in unhealthy food choices. One of this studies found an association between lower food healthiness evaluation of food in a food-cue task, and higher activation in the amygdala (149). Therefore, the ability to differentiate healthy from unhealthy foods is affected by emotional state.
Nevertheless, reproducibility across these studies appears to be highly dependent on various external factors, limiting the fine characterization of the role of HT within food intake networks. One meta-analysis (148) revealed that no more than 41% of experiments contributed to the activated voxel clusters for the contrast between food and non-food pictures, which could be explained by biases of subject selection according to age (150), BMI scores (151), sex, hunger states (152), emotional state (153), dietary restriction (154) and subject genotype (155). Minimization of potential biases relative to menstrual cycle, which could trigger cravings, has been proposed through the recommendation of performing MRI scans during the follicular phase (156). Indeed, mean diffusivity and metabolism of HT appear to be modulated by artificial menstrual cycle, leading to a decrease in ADCs and increase in Choline/NAA ratios after oral contraceptive use compared to pill free period in young healthy women (157).
Regarding study designs, food paradigms differ between studies in the nature and nutritional properties of the food presented, in task instruction and even in sample sizes, which are generally small, and differences in the acquisition and analyses methods (148). In regard to these irregularities, which hinder the understanding of the role of the HT in the regulation of food intake, standardized databases of food and non-food images have been implemented (158, 159). Moreover, the duration of the fasting period before scan sessions should be harmonized to create a more homogenous hunger state in subjects across studies.
Recent studies have tried to minimize the bias related to individuals' food preferences. Some studies take into account the participant's preferred food in their script (160), while others ask them to rate the appetence of the presented food (161). One study even had volunteers rate their hunger, satiety, thirst, fullness and emptiness, before and after the scanning session (162). However, taking into account those ratings did not refine the model fit in this food-cue reactivity study.
During satiation, obese individuals showed deactivation of the precuneus and the superior parietal cortex, with lower rCBF in the HT, cingulate, nucleus accumbens, in the limbic and paralimbic regions (parahippocampal gyrus, insular cortex, amygdala), caudate nucleus, frontal and temporal regions relative to controls (25, 163). In contrast, sated obese women exhibited significantly higher rCBF in the ventral PFC and frontal operculum (25, 163). These findings are in agreement with the hypothesis that the PFC is implicated in the termination of a meal by its inhibiting action on limbic and paralimbic areas (25).
Concurrently, glucose ingestion in obese patients relative to controls induced a delayed and attenuated dose-dependent decrease in BOLD signal in the PVN and VMH (139). In addition, the decrease of rCBF in HT during a meal associated with normal satiety processes in healthy subjects has appeared to be attenuated in obese patients (163).
When administered a liquid meal, obese and former obese who succeeded in decreasing their BMI from at least 35 to a BMI of 25 kg/m2 and in keeping their weight stable for no less than 3 months before the start of the study, exhibited an increased rCBF in the middle insular cortex while no change in activity was registered in lean individuals (164). Additionally, when given a satiating amount of the same liquid, obese presented a different neural response in the posterior hippocampus, posterior cingulate cortex and amygdala. Indeed, the posterior hippocampus rCBF decreased in a similar manner in obese and former obese while it increased in lean individuals (164).
Persisting differential activities in regions implicated in gustation and rewarding/ hedonic aspect of food (insular cortex) or enteroception and learning/memory (hippocampus) could partly explain the high risk for relapse of former obese. Interestingly, a study on smokers and non-smokers revealed that the hypothalamic response to oral intake of milkshake in non-smokers was lower, which is associated with long-term weight change in this population (165).
Food-cue task fMRI studies have reported elevated responsivity of somatosensory (Rolandic operculum), gustatory (insula, frontal operculum) and reward brain areas (caudate, putamen, amygdala, OFC) in obese subjects compared to lean individuals when presented with palatable food cues (166–168). Abnormal activity patterns in obese patients have been found in the HT, PFC, parietal and temporal cortex, cingulate cortex, nucleus acumens, amygdala, midbrain, insula, and OFC (25, 164).
During high-calorie food cue, obesity has been associated with hyperactivation of the right putamen, left caudate body, left anterior insula, left hippocampus, and the left parietal lobe relative to control, while low-calorie food cue elicited exacerbate responses in the left superior frontal, right middle and inferior frontal gyrus, middle occipital gyrus, and left superior temporal gyrus in obese compared to healthy controls (166). Thus, BMI was positively associated with higher response to high-calorie in brain regions involved in the processing of taste information (anterior insula, lateral OFC), motivation (OFC), emotions and memory (posterior cingulate cortex), salience-guided orienting (claustrum) and reward anticipation (dorsal striatum) (166, 169). BMI is also positively associated with selective attention toward appetitive food-cue and higher responsivity in reward regions such as the ventrolateral prefrontal cortex (VLPFC), lateral OFC, temporal operculum and anterior insula (59, 170). However, when asked to inhibit responses to high-calorie food images, obese individuals exhibited less activation in frontal inhibitory regions, such as the superior and middle frontal gyrus, VLPFC and VMPFC, and OFC, relative to lean individuals, which is consistent with behavioral evidence of impulsivity often found in overweight individuals (170). These findings suggest that a higher weight is preferably linked to hypo-activation of inhibition control regions and hyper-activation of reward regions in response to palatable food. Failure of inhibition control regions to inhibit reward regions when presented with appetitive food could increase the susceptibility to overeat (171). Moreover, obese-like activity in the middle insular cortex and hippocampus persists in former obese (164). These results suggest that higher activation in reward and somatosensory regions in response to food-cue constitute a risk for future weight gain.
During reduced weight maintenance, obese individuals show high FC between the HT and visual (occipital fusiform and temporal fusiform areas, superior lateral occipital cortex and cuneus), memory (hippocampus) and attention areas (dorsal ACC, left middle and inferior frontal gyri) when viewing food compared to non-food images (172). Moreover, the inferior frontal gyrus, implicated in inhibitory control (173), has been found to be more activated by food images in people able to maintain their weight loss than in obese or lean subjects (174) and its activation in satiation is lower in obese than in normal weight participants (175). These results suggest that during weight loss, obese patients have a higher sensitivity to food cues. Furthermore, the inhibitory action of the inferior frontal gyrus seems to be necessary to maintain weight loss (174).
Overall, obesity seems to be associated with a concurrent blunted hypothalamic reactivity to glucose ingestion and an exacerbated hedonic reactivity, which do not coincide with homeostatic state and lead to overconsumption of high fat food.
Considering the ability of AN patients to fight against the natural instinct to feed oneself, the hypothesis of a dysregulation in hypothalamic glucose-sensitivity was investigated. AN and controls exhibited the same decrease in neuronal activity when administered with a glucose solution (176). Therefore, the hypothalamic reactivity to glucose does not seem to be altered in AN. However, contrary to normal-weight controls and obese subjects, AN patients did not display an increase of FC between the HT and brains regions implicated in reward processes (amygdala, nucleus accumbens) during infusion of water when compared to glucose infusion, with the exception of the posterior insula (120). As opposed to control, AN did not show deactivation in the mesocorticolimbic reward circuit including structures such as the caudate nucleus, putamen, insular cortex, medial OFC, and inferior operculum after glucose administration (120).
Furthermore, AN exhibited a higher FC between the HT and the left ventral striatum in response to glucose administration relative to normal-weight controls and obese subjects (120). The abnormal hyper-activation of the ventral striatum is a neural signature of patients suffering from AN and is hypothesized to be involved in the onset and maintenance of this illness (177). Interestingly, a study comparing the response to pictures of underweight women demonstrated a higher activation in the left ventral striatum in AN patients compared to lean control (178).
Moreover, patients with AN do not show satiety state–dependent connectivity between the HT and the meso-corticolimbic reward circuit that is commonly found in normal-weight controls. This observation could explain the reduced hedonic coding aspect of food associated with AN (120). AN individuals show hypoactivation of the food motivation network in response to high-calorie foods (179). Indeed, both fasted women with weight-restored (maintenance of 90%−110% ideal body weight for at least 6 months) and active AN exhibited hypoactivity in the HT, amygdala and anterior insula in response to high-calorie foods vs. objects when compared to control. Hypoactivation in the anterior insula persisted in women with active disease after they were served a meal (18% calories from protein, 23% from fat and 59% from carbohydrates).
In contrast, weight-restored AN women show similar activation to controls after a meal (179). Compared with controls, women with active disease show hypoactivation in the HT, amygdala, hippocampus, OFC and anterior insula when fasted and in the amygdala and insula after they ingested a meal (179). Interestingly, activations in the HT, amygdala and anterior insula are significantly associated with appetite (hunger) and appetence ratings (how appealing is the food) in controls and weight-restored AN women but not in those with active disease (179). The authors provided evidence of a dysfunction of the food motivation network, including the HT, in women with active AN, which tends to persist even after weight-restoration. AN has been shown to be associated with lower food-cue processing activity in brain structures implicated in reward and salience (HT, striatum, hippocampus, amygdala, cerebellum, insula) but greater activity in regions involved with cognitive control (dorsolateral PFC, mPFC, OFC, ACC) (180, 181). To sum up, AN has been associated with a reduced hypothalamic reactivity and connectivity with the reward system, which could be responsible for the repressed motivational responses to food cue, in relation with an alteration in food reward processing.
Despite the important progress in the neuroimaging of HT and the better characterization of its functional activation pattern across different hunger states or eating disorders, its role within the structural and functional food-intake networks remains to be elucidated. UHF MRI combined with the use of the highly resolved atlases (4) should allow for a better understanding of the role of each hypothalamic nuclei in a normal and pathological context. Indeed, advanced magnetic resonance (MR) techniques provide new significant anatomical, functional metabolic and connectomic insights of the HT, through the improvement of quantitative MRI, spectral/spatial resolution of MRS, spatial resolution of anatomic MRI, diffusion and functional MRI. Furthermore, the current paper briefly mentioned the possible contribution of metabolic MRI in the study of obesity. In addition, MRI is an undeniable interesting tool in the study of the effectiveness of treatments for eating disorders in humans since this technic is non-invasive and partially replace for the lack of AN animal models. Finally, combining the study of the HT at the structural, functional and metabolic levels could bring new perspectives to better understand and better treat patients with eating disorders, as it is already done in the study of pathologies such as Alzheimer's (182) or epilepsy (183).
All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
CR had received a PhD grant from the Neuroschool, a support from the French government under the Programme Investissements d'Avenir, Initiative d'Excellence d'Aix-Marseille Université via A*Midex (AMX-19-IET-004) and ANR (ANR-17-EURE-0029) funding.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Morton GJ, Meek TH, Schwartz MW. Neurobiology of food intake in health and disease. Nat Rev Neurosci. (2014) 15:367–78. doi: 10.1038/nrn3745
2. World Health Organisation. Controlling The Global Obesity Epidemic. (2001). Available Online at: https://www.who.int/activities/controlling-the-global-obesity-epidemic.
3. Arcelus J, Mitchell AJ, Wales J, Nielsen S. Mortality rates in patients with anorexia nervosa and other eating disorders. A meta-analysis of 36 studies. Arch Gen Psychiatry. (2011) 68:724–31. doi: 10.1001/archgenpsychiatry.2011.74
4. Neudorfer C, Germann J, Elias GJB, Gramer R, Boutet A, Lozano AM. A high-resolution in vivo magnetic resonance imaging atlas of the human hypothalamic region. Sci Data. (2020) 7:305. doi: 10.1038/s41597-020-00644-6
5. Atlas of the Human Hypothalamus - 1st Edition. [WWW Document] (2021). Available online at: https://www.elsevier.com/books/atlas-of-the-human-hypothalamus/dudas/978-0-12-822051-1
6. Saper CB, Lowell BB. The hypothalamus. Curr Biol. (2014) 24:R1111–1116. doi: 10.1016/j.cub.2014.10.023
7. Krieg WJS. The hypothalamus of the albino rat. J Comp Neurol. (1932) 55:19–89. doi: 10.1002/cne.900550104
8. Anand BK, Brobeck JR. Localization of a “feeding center” in the hypothalamus of the rat. Proc Soc Exp Biol Med. (1951) 77:323–4. doi: 10.3181/00379727-77-18766
9. Smith PM, Ferguson AV. Neurophysiology of hunger and satiety. Dev Disabil Res Rev. (2008) 14:96–104. doi: 10.1002/ddrr.13
10. De Silva A, Salem V, Matthews PM, Dhillo WS. The Use of Functional MRI to Study Appetite Control in the CNS. Exp Diabetes Res. (2012) 2012:1–13. doi: 10.1155/2012/764017
11. Hetherington AW, Ranson SW. Hypothalamic lesions and adiposity in the rat. Anat Rec. (1940) 78:149–72. doi: 10.1002/ar.1090780203
12. Berthoud H-R. Multiple neural systems controlling food intake and body weight. Neurosci Biobehav Rev. (2002) 26:393–428. doi: 10.1016/S0149-7634(02)00014-3
13. Jais A, Brüning JC. Hypothalamic inflammation in obesity and metabolic disease. J Clin Invest. (2017) 127:24–32. doi: 10.1172/JCI88878
14. Mawe GM, Hoffman JM. Serotonin signalling in the gut–functions, dysfunctions and therapeutic targets. Nat Rev Gastroenterol Hepatol. (2013) 10:473–86. doi: 10.1038/nrgastro.2013.105
15. O'Mahony SM, Clarke G, Borre YE, Dinan TG, Cryan JF. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav Brain Res. (2015) 277:32–48. doi: 10.1016/j.bbr.2014.07.027
16. Roman CW, Derkach VA, Palmiter RD. Genetically and functionally defined NTS to PBN brain circuits mediating anorexia. Nat Commun. (2016) 7:11905. doi: 10.1038/ncomms11905
17. de Araujo IE, Schatzker M, Small DM. Rethinking Food Reward. Annu Rev Psychol. (2020) 71:139–64. doi: 10.1146/annurev-psych-122216-011643
18. Tan H-E, Sisti AC, Jin H, Vignovich M, Villavicencio M, Tsang KS, et al. The gut-brain axis mediates sugar preference. Nature. (2020) 580:511–6. doi: 10.1038/s41586-020-2199-7
19. Koutcherov Y, Mai JK, Paxinos G. Hypothalamus of the human fetus. J Chem Neuroanat. (2003) 26:253–70. doi: 10.1016/j.jchemneu.2003.07.002
20. Quarta C, Claret M, Zeltser LM, Williams KW, Yeo GSH, Tschöp MH, et al. POMC neuronal heterogeneity in energy balance and beyond: an integrated view. Nat Metab. (2021) 3:299–308. doi: 10.1038/s42255-021-00345-3
21. Considine RV, Sinha MK, Heiman ML, Kriauciunas A, Stephens TW, Nyce MR, et al. Serum immunoreactive-leptin concentrations in normal-weight and obese humans. N Engl J Med. (1996) 334:292–5. doi: 10.1056/NEJM199602013340503
22. Berthoud H-R, Morrison C. The Brain, Appetite, and Obesity (SSRN Scholarly Paper No. ID 1141913). Rochester, NY: Social Science Research Network. (2008).
23. Luo SX, Huang J, Li Q, Mohammad H, Lee C-Y, Krishna K, et al. Regulation of feeding by somatostatin neurons in the tuberal nucleus. Science. (2018) 361:76–81. doi: 10.1126/science.aar4983
24. Berthoud H-R. The neurobiology of food intake in an obesogenic environment. Proc Nutr Soc. (2012) 71:478–87. doi: 10.1017/S0029665112000602
25. Gautier JF, Del Parigi A, Chen K, Salbe AD, Bandy D, Pratley RE, et al. Effect of satiation on brain activity in obese and lean women. Obes Res. (2001) 9:676–84. doi: 10.1038/oby.2001.92
26. Thaler JP, Schwartz MW. Minireview: Inflammation and Obesity Pathogenesis: The Hypothalamus Heats Up. Endocrinology. (2010) 151:4109–15. doi: 10.1210/en.2010-0336
27. De Souza CT, Araujo EP, Bordin S, Ashimine R, Zollner RL, Boschero AC, et al. Consumption of a fat-rich diet activates a proinflammatory response and induces insulin resistance in the hypothalamus. Endocrinology. (2005) 146:4192–9. doi: 10.1210/en.2004-1520
28. Ávalos Y, Kerr B, Maliqueo M, Dorfman M. Cell and molecular mechanisms behind diet-induced hypothalamic inflammation and obesity. J Neuroendocrinol. (2018) 30:e12598. doi: 10.1111/jne.12598
29. Thaler JP, Yi C-X, Schur EA, Guyenet SJ, Hwang BH, Dietrich MO, et al. Obesity is associated with hypothalamic injury in rodents and humans. J Clin Invest. (2012) 122:153–62. doi: 10.1172/JCI59660
30. Posey KA, Clegg DJ, Printz RL, Byun J, Morton GJ, Vivekanandan-Giri A, et al. Hypothalamic proinflammatory lipid accumulation, inflammation, and insulin resistance in rats fed a high-fat diet. Am J Physiol Endocrinol Metab. (2009) 296:E1003–12. doi: 10.1152/ajpendo.90377.2008
31. Morselli E, Fuente-Martin E, Finan B, Kim M, Frank A, Garcia-Caceres C, et al. Hypothalamic PGC-1α Protects Against High-Fat Diet Exposure by Regulating ERα. Cell Rep. (2014) 9:633–45. doi: 10.1016/j.celrep.2014.09.025
32. Valdearcos M, Robblee MM, Benjamin DI, Nomura DK, Xu AW, Koliwad SK. Microglia Dictate the Impact of Saturated Fat Consumption on Hypothalamic Inflammation and Neuronal Function. Cell Rep. (2014) 9:2124–38. doi: 10.1016/j.celrep.2014.11.018
33. Cai D, Khor S. “Hypothalamic Microinflammation” Paradigm in Aging and Metabolic Diseases. Cell Metab. (2019) 30:19–35. doi: 10.1016/j.cmet.2019.05.021
34. Wang GJ, Volkow ND, Logan J, Pappas NR, Wong CT, Zhu W, et al. Brain dopamine and obesity. Lancet. (2001) 357:354–7. doi: 10.1016/S0140-6736(00)03643-6
35. van Eeden AE, van Hoeken D, Hoek HW. Incidence, prevalence and mortality of anorexia nervosa and bulimia nervosa. Curr Opin Psychiatry. (2021) 34:515–24. doi: 10.1097/YCO.0000000000000739
36. Rask-Andersen M, Olszewski PK, Levine AS, Schiöth HB. Molecular mechanisms underlying anorexia nervosa: focus on human gene association studies and systems controlling food intake. Brain Res Rev. (2010) 62:147–64. doi: 10.1016/j.brainresrev.2009.10.007
37. Klenotich SJ, Ho EV, McMurray MS, Server CH, Dulawa SC. Dopamine D2/3 receptor antagonism reduces activity-based anorexia. Transl Psychiatry. (2015) 5:e613. doi: 10.1038/tp.2015.109
38. Frank GK, Bailer UF, Henry SE, Drevets W, Meltzer CC, Price JC, et al. Increased dopamine D2/D3 receptor binding after recovery from anorexia nervosa measured by positron emission tomography and [11c]raclopride. Biol Psychiatry. (2005) 58:908–12. doi: 10.1016/j.biopsych.2005.05.003
39. Haleem DJ. Serotonin neurotransmission in anorexia nervosa. Behav Pharmacol. (2012) 23:478–95. doi: 10.1097/FBP.0b013e328357440d
40. Méquinion M, Langlet F, Zgheib S, Dickson S, Dehouck B, Chauveau C, et al. Ghrelin: Central and Peripheral Implications in Anorexia Nervosa. Front Endocrinol (Lausanne). (2013) 4:15. doi: 10.3389/fendo.2013.00015
41. Salem V, Dhillo WS. Imaging in endocrinology: The use of functional MRI to study the endocrinology of appetite. Eur J Endocrinol. (2015) 173:R59–68. doi: 10.1530/EJE-14-0716
42. Florent V, Baroncini M, Jissendi-Tchofo P, Lopes R, Vanhoutte M, Rasika S, et al. Hypothalamic Structural and Functional Imbalances in Anorexia Nervosa. Neuroendocrinology. (2020) 110:552–62. doi: 10.1159/000503147
43. Brodmann K, Gary LJ. Brodmann's Localisation in the Cerebral Cortex: The Principles of Comparative Localisation in the Cerebral Cortex Based on Cytoarchitectonics. 1st ed New York, NY: Springer (2006). p. 286.
44. Desikan RS, Ségonne F, Fischl B, Quinn BT, Dickerson BC, Blacker D, et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. Neuroimage. (2006) 31:968–80. doi: 10.1016/j.neuroimage.2006.01.021
45. Lancaster JL, Tordesillas-Gutiérrez D, Martinez M, Salinas F, Evans A, Zilles K, et al. Bias between MNI and Talairach coordinates analyzed using the ICBM-152 brain template. Hum Brain Mapp. (2007) 28:1194–205. doi: 10.1002/hbm.20345
46. Mai JK, Majtanik M, Paxinos G. Atlas of the Human Brain. 4th ed. San Diego, SD: Academic Press (2016). p. 350.
47. Pauli WM, Nili AN, Tyszka JM. A high-resolution probabilistic in vivo atlas of human subcortical brain nuclei. Scientific Data. (2018) 5:180063. doi: 10.1038/sdata.2018.63
48. Tian Y, Margulies DS, Breakspear M, Zalesky A. Topographic organization of the human subcortex unveiled with functional connectivity gradients. Nat Neurosci. (2020) 23:1421–32. doi: 10.1038/s41593-020-00711-6
49. Nieuwenhuys R, Voogd J, van Huijzen C. (1988). The Human Central Nervous System: A Synopsis and Atlas, 3rd edn. Berlin Heidelberg: Springer-Verlag. doi: 10.1007/978-3-662-10343-2
50. Baroncini M, Jissendi P, Balland E, Besson P, Pruvo J-P, Francke J-P, et al. MRI atlas of the human hypothalamus. Neuroimage. (2012) 59:168–80. doi: 10.1016/j.neuroimage.2011.07.013
51. Makris N, Swaab DF, van der Kouwe A, Abbs B, Boriel D, Handa RJ, et al. Volumetric parcellation methodology of the human hypothalamus in neuroimaging: normative data and sex differences. Neuroimage. (2013) 69:1–10. doi: 10.1016/j.neuroimage.2012.12.008
52. Lemaire J-J, Nezzar H, Sakka L, Boirie Y, Fontaine D, Coste A, et al. Maps of the adult human hypothalamus. Surg Neurol Int. (2013) 4:S156–63. doi: 10.4103/2152-7806.110667
53. Ding S-L, Royall JJ, Sunkin SM, Ng L, Facer BAC, Lesnar P, et al. Comprehensive cellular-resolution atlas of the adult human brain. J Comp Neurol. (2016) 524:3127–481. doi: 10.1002/cne.24097
54. Avants BB, Epstein CL, Grossman M, Gee JC. Symmetric diffeomorphic image registration with cross-correlation: evaluating automated labeling of elderly and neurodegenerative brain. Med Image Anal. (2008) 12:26–41. doi: 10.1016/j.media.2007.06.004
55. Marques JP, Kober T, Krueger G, van der Zwaag W, Van de Moortele P-F, Gruetter R. MP2RAGE, a self bias-field corrected sequence for improved segmentation and T1-mapping at high field. Neuroimage. (2010) 49:1271–81. doi: 10.1016/j.neuroimage.2009.10.002
56. Pannacciulli N, Del Parigi A, Chen K, Le DSNT, Reiman EM, Tataranni PA. Brain abnormalities in human obesity: a voxel-based morphometric study. Neuroimage. (2006) 31:1419–25. doi: 10.1016/j.neuroimage.2006.01.047
57. Yokum S, Ng J, Stice E. Relation of Regional Grey and White Matter Volumes to Current BMI and Future Increases in BMI: A Prospective MRI Study. Int J Obes (Lond). (2012) 36:656–64. doi: 10.1038/ijo.2011.175
58. Dekkers IA, Jansen PR, Lamb HJ. Obesity, brain volume, and white matter microstructure at MRI: A cross-sectional UK biobank study. Radiology. (2019) 291:763–71. doi: 10.1148/radiol.2019194010
59. Yokum S, Ng J, Stice E. Attentional bias to food images associated with elevated weight and future weight gain: an fMRI study. Obesity (Silver Spring). (2011) 19:1775–83. doi: 10.1038/oby.2011.168
60. Boghi A, Sterpone S, Sales S, D'Agata F, Bradac GB, Zullo G, et al. In vivo evidence of global and focal brain alterations in anorexia nervosa. Psychiatry Res. (2011) 192:154–9. doi: 10.1016/j.pscychresns.2010.12.008
61. Titova OE, Hjorth OC, Schiöth HB, Brooks SJ. Anorexia nervosa is linked to reduced brain structure in reward and somatosensory regions: a meta-analysis of VBM studies. BMC Psychiatry. (2013) 13:110. doi: 10.1186/1471-244X-13-110
62. Artmann H, Grau H, Adelmann M, Schleiffer R. Reversible and non-reversible enlargement of cerebrospinal fluid spaces in anorexia nervosa. Neuroradiology. (1985) 27:304–12. doi: 10.1007/BF00339562
63. Miles AE, Voineskos AN, French L, Kaplan AS. Subcortical volume and cortical surface architecture in women with acute and remitted anorexia nervosa: An exploratory neuroimaging study. J Psychiatr Res. (2018) 102:179–85. doi: 10.1016/j.jpsychires.2018.04.010
64. King JA, Geisler D, Ritschel F, Boehm I, Seidel M, Roschinski B, et al. Global cortical thinning in acute anorexia nervosa normalizes following long-term weight restoration. Biol Psychiatry. (2015) 77:624–32. doi: 10.1016/j.biopsych.2014.09.005
65. Bernardoni F, King JA, Geisler D, Birkenstock J, Tam FI, Weidner K, et al. Nutritional Status Affects Cortical Folding: Lessons Learned From Anorexia Nervosa. Biol Psychiatry. (2018) 84:692–701. doi: 10.1016/j.biopsych.2018.05.008
66. Bernardoni F, King JA, Geisler D, Stein E, Jaite C, Nätsch D, et al. Weight restoration therapy rapidly reverses cortical thinning in anorexia nervosa: A longitudinal study. Neuroimage. (2016) 130:214–22. doi: 10.1016/j.neuroimage.2016.02.003
67. Harris RBS. (2017). Appetite and Food Intake: Central Control, 2nd edn. CRC Press/Taylor & Francis, Boca Raton (FL). doi: 10.1201/9781315120171
68. Kamali A, Yousem DM, Lin DD, Sair HI, Jasti SP, Keser Z, et al. Mapping the trajectory of the stria terminalis of the human limbic system using high spatial resolution diffusion tensor tractography. Neurosci Lett. (2015) 608:45–50. doi: 10.1016/j.neulet.2015.09.035
69. Noback CR, Ruggiero DA, Demarest RJ, Strominger NL. (2007). The Human Nervous System: Structure and Function, 6th ed. 2005 edition. edn. Totowa, N.J.: Humana Press.
70. Nolte J, Sundsten JW. The Human Brain: An Introduction to Its Functional Anatomy. 5th ed. St. Louis, MO: Mosby (2002). p. 650.
71. Kamali A, Sair HI, Blitz AM, Riascos RF, Mirbagheri S, Keser Z, et al. Revealing the ventral amygdalofugal pathway of the human limbic system using high spatial resolution diffusion tensor tractography. Brain Struct Funct. (2016) 221:3561–9. doi: 10.1007/s00429-015-1119-3
72. Metzler-Baddeley C, Baddeley RJ, Jones DK, Aggleton JP, O'Sullivan MJ. Individual Differences in Fornix Microstructure and Body Mass Index. PLoS ONE. (2013) 8:e59849. doi: 10.1371/journal.pone.0059849
73. Tracy AL, Jarrard LE, Davidson TL. The hippocampus and motivation revisited: appetite and activity. Behav Brain Res. (2001) 127:13–23. doi: 10.1016/S0166-4328(01)00364-3
74. Kamali A, Karbasian N, Ghazi Sherbaf F, Wilken LA, Aein A, Sair HI, et al. Uncovering the dorsal thalamo-hypothalamic tract of the human limbic system. Neuroscience. (2020) 432:55–62. doi: 10.1016/j.neuroscience.2020.02.021
75. Moga MM, Weis RP, Moore RY. Efferent projections of the paraventricular thalamic nucleus in the rat. J Comp Neurol. (1995) 359:221–38. doi: 10.1002/cne.903590204
76. Kelley AE, Baldo BA, Pratt WE. A proposed hypothalamic-thalamic-striatal axis for the integration of energy balance, arousal, and food reward. J Comp Neurol. (2005) 493:72–85. doi: 10.1002/cne.20769
77. Poulin A-M, Timofeeva E. The dynamics of neuronal activation during food anticipation and feeding in the brain of food-entrained rats. Brain Res. (2008) 1227:128–41. doi: 10.1016/j.brainres.2008.06.039
78. Kamali A, Ghazi Sherbaf F, Rahmani F, Khayat-Khoei M, Aein A, Gandhi A, et al. A direct visuosensory cortical connectivity of the human limbic system. Dissecting the trajectory of the parieto-occipito-hypothalamic tract in the human brain using diffusion weighted tractography. Neurosci Lett. (2020) 728:134955. doi: 10.1016/j.neulet.2020.134955
79. Risold PY, Thompson RH, Swanson LW. The structural organization of connections between hypothalamus and cerebral cortex. Brain Res Brain Res Rev. (1997) 24:197–254. doi: 10.1016/S0165-0173(97)00007-6
80. Jacobs VL. The Cerebellofugal System in the Tarsius (Tarsiidae Carbonarius) and the Marmoset (Oedipomidas Oedipus). (Ph.D.thesis) University of Kansas, Kansas, United States (1965).
81. Martin GF, Dom R, Katz S, King JS. The organization of projection neurons in the opossum red nucleus. Brain Res. (1974) 78:17–34. doi: 10.1016/0006-8993(74)90350-3
82. Kamali A, Karbasian N, Rabiei P, Cano A, Riascos RF, Tandon N, et al. Revealing the cerebello-ponto-hypothalamic pathway in the human brain. Neurosci Lett. (2018) 677:1–5. doi: 10.1016/j.neulet.2018.04.024
83. Çavdar S, Esen Aydin A, Algin O, Aydogmuş E. Fiber dissection and 3-tesla diffusion tensor tractography of the superior cerebellar peduncle in the human brain: emphasize on the cerebello-hypthalamic fibers. Brain Struct Funct. (2020) 225:121–8. doi: 10.1007/s00429-019-01985-8
84. Zhu J-N, Wang J-J. The cerebellum in feeding control: possible function and mechanism. Cell Mol Neurobiol. (2008) 28:469–78. doi: 10.1007/s10571-007-9236-z
85. Lemaire J-J, Frew AJ, McArthur D, Gorgulho AA, Alger JR, Salomon N, et al. White matter connectivity of human hypothalamus. Brain Res. (2011) 1371:43–64. doi: 10.1016/j.brainres.2010.11.072
86. Schönknecht P, Anwander A, Petzold F, Schindler S, Knösche TR, Möller HE, et al. Diffusion imaging-based subdivision of the human hypothalamus: a magnetic resonance study with clinical implications. Eur Arch Psychiatry Clin Neurosci. (2013) 263:497–508. doi: 10.1007/s00406-012-0389-5
87. Van den Eynde F, Treasure J. Neuroimaging in Eating Disorders and Obesity: Implications for Research. Child Adolesc Psychiatr Clin N Am. (2009) 18:95–115. doi: 10.1016/j.chc.2008.07.016
88. Puig J, Blasco G, Daunis-I-Estadella J, Molina X, Xifra G, Ricart W, et al. (2015). Hypothalamic damage is associated with inflammatory markers and worse cognitive performance in obese subjects. J Clin Endocrinol Metab 100, E276–281. doi: 10.1210/jc.2014-2682
89. Thomas K, Beyer F, Lewe G, Zhang R, Schindler S, Schönknecht P, et al. Higher body mass index is linked to altered hypothalamic microstructure. Sci Rep. (2019) 9:17373. doi: 10.1038/s41598-019-53578-4
90. Haltia LT, Viljanen A, Parkkola R, Kemppainen N, Rinne JO, Nuutila P, et al. Brain white matter expansion in human obesity and the recovering effect of dieting. J Clin Endocrinol Metab. (2007) 92:3278–84. doi: 10.1210/jc.2006-2495
91. Walther K, Birdsill AC, Glisky EL, Ryan L. Structural brain differences and cognitive functioning related to body mass index in older females. Hum Brain Mapp. (2010) 31:1052–64. doi: 10.1002/hbm.20916
92. Raji CA, Ho AJ, Parikshak NN, Becker JT, Lopez OL, Kuller LH, et al. Brain structure and obesity. Hum Brain Mapp. (2009) 31:353–64. doi: 10.1002/hbm.20870
93. Mueller K, Anwander A, Möller HE, Horstmann A, Lepsien J, Busse F, et al. Sex-dependent influences of obesity on cerebral white matter investigated by diffusion-tensor imaging. PLoS ONE. (2011) 6. doi: 10.1371/journal.pone.0018544
94. Stanek KM, Grieve SM, Brickman AM, Korgaonkar MS, Paul RH, Cohen RA, et al. Obesity is associated with reduced white matter integrity in otherwise healthy adults. Obesity (Silver Spring). (2011) 19:500–4. doi: 10.1038/oby.2010.312
95. Xu J, Li Y, Lin H, Sinha R, Potenza MN. Body mass index correlates negatively with white matter integrity in the fornix and corpus callosum: a diffusion tensor imaging study. Hum Brain Mapp. (2013) 34:1044–52. doi: 10.1002/hbm.21491
96. Miles AE, Kaplan AS, French L, Voineskos AN. White matter microstructure in women with acute and remitted anorexia nervosa: an exploratory neuroimaging study. Brain Imaging Behav. (2020) 14:2429–37. doi: 10.1007/s11682-019-00193-6
97. Martin Monzon B, Hay P, Foroughi N, Touyz S. White matter alterations in anorexia nervosa: A systematic review of diffusion tensor imaging studies. World J Psychiatry. (2016) 6:177–86. doi: 10.5498/wjp.v6.i1.177
98. Nagahara Y, Nakamae T, Nishizawa S, Mizuhara Y, Moritoki Y, Wada Y, et al. A tract-based spatial statistics study in anorexia nervosa: abnormality in the fornix and the cerebellum. Prog Neuropsychopharmacol Biol Psychiatry. (2014) 51:72–7. doi: 10.1016/j.pnpbp.2014.01.009
99. Chen EY, Zeffiro TA. Hunger and BMI modulate neural responses to sweet stimuli: fMRI meta-analysis. Int J Obes. (2020) 44:1636–52. doi: 10.1038/s41366-020-0608-5
100. Cha J, Ide JS, Bowman FD, Simpson HB, Posner J, Steinglass JE. Abnormal reward circuitry in anorexia nervosa: A longitudinal, multimodal MRI study. Hum Brain Mapp. (2016) 37:3835–46. doi: 10.1002/hbm.23279
101. Fritz FJ, Sengupta S, Harms RL, Tse DH, Poser BA, Roebroeck A. Ultra-high resolution and multi-shell diffusion MRI of intact ex vivo human brains using kT-dSTEAM at 9.4T. NeuroImage. (2019) 202:116087. doi: 10.1016/j.neuroimage.2019.116087
102. Glasser MF, Sotiropoulos SN, Wilson JA, Coalson TS, Fischl B, Andersson JL, et al. The minimal preprocessing pipelines for the Human Connectome Project. Neuroimage. (2013) 80:105–24. doi: 10.1016/j.neuroimage.2013.04.127
103. Biswal B, Yetkin FZ, Haughton VM, Hyde JS. Functional connectivity in the motor cortex of resting human brain using echo-planar MRI. Magn Reson Med. (1995) 34:537–41. doi: 10.1002/mrm.1910340409
104. Scaife JC, Godier LR, Filippini N, Harmer CJ, Park RJ. Reduced resting-state functional connectivity in current and recovered restrictive anorexia nervosa. Front Psychiatry. (2017) 8:30. doi: 10.3389/fpsyt.2017.00030
105. Wright H, Li X, Fallon NB, Crookall R, Giesbrecht T, Thomas A, et al. Differential effects of hunger and satiety on insular cortex and hypothalamic functional connectivity. Eur J Neurosci. (2016) 43:1181–9. doi: 10.1111/ejn.13182
106. Hollmann M, Hellrung L, Pleger B, Schlögl H, Kabisch S, Stumvoll M, et al. Neural correlates of the volitional regulation of the desire for food. Int J Obes (Lond). (2012) 36:648–55. doi: 10.1038/ijo.2011.125
107. Lopez RB, Hofmann W, Wagner DD, Kelley WM, Heatherton TF. Neural predictors of giving in to temptation in daily life. Psychol Sci. (2014) 25:1337–44. doi: 10.1177/0956797614531492
108. Al-Zubaidi A, Iglesias S, Stephan KE, Buades-Rotger M, Heldmann M, Nolde JM, et al. Effects of hunger, satiety and oral glucose on effective connectivity between hypothalamus and insular cortex. Neuroimage. (2020) 217:116931. doi: 10.1016/j.neuroimage.2020.116931
109. Kullmann S, Heni M, Linder K, Zipfel S, Häring H-U, Veit R, et al. Resting-state functional connectivity of the human hypothalamus. Hum Brain Mapp. (2014) 35:6088–96. doi: 10.1002/hbm.22607
110. Mayer J, Barrnett RJ. Obesity following unilateral hypothalamic lesions in rats. Science. (1955) 121:599–600. doi: 10.1126/science.121.3147.599
111. Dosenbach NUF, Visscher KM, Palmer ED, Miezin FM, Wenger KK, Kang HC, et al. A core system for the implementation of task sets. Neuron. (2006) 50:799–812. doi: 10.1016/j.neuron.2006.04.031
112. O'Doherty JP. Reward representations and reward-related learning in the human brain: insights from neuroimaging. Curr Opin Neurobiol. (2004) 14:769–76. doi: 10.1016/j.conb.2004.10.016
113. Hirose S, Osada T, Ogawa A, Tanaka M, Wada H, Yoshizawa Y, et al. Lateral-Medial Dissociation in Orbitofrontal Cortex-Hypothalamus Connectivity. Front Hum Neurosci. (2016) 10:244. doi: 10.3389/fnhum.2016.00244
114. Tomasi D, Volkow ND. Striatocortical pathway dysfunction in addiction and obesity: differences and similarities. Crit Rev Biochem Mol Biol. (2013) 48:1–19. doi: 10.3109/10409238.2012.735642
115. Gulban OF, De Martino F, Vu AT, Yacoub E, Ugurbil K, Lenglet C. Cortical fibers orientation mapping using in-vivo whole brain 7 T diffusion MRI. Neuroimage. (2018) 178:104–18. doi: 10.1016/j.neuroimage.2018.05.010
116. Geha P, Cecchi G, Todd Constable R, Abdallah C, Small DM. Reorganization of brain connectivity in obesity: Obesity and Altered Brain Connectivity. Hum Brain Mapp. (2017) 38:1403–20. doi: 10.1002/hbm.23462
117. Le TM, Liao D-L, Ide J, Zhang S, Zhornitsky S, Wang W, et al. The interrelationship of body mass index with gray matter volume and resting-state functional connectivity of the hypothalamus. Int J Obes. (2020) 44:1097–107. doi: 10.1038/s41366-019-0496-8
118. Martín-Pérez C, Contreras-Rodríguez O, Vilar-López R, Verdejo-García A. Hypothalamic networks in adolescents with excess weight: stress-related connectivity and associations with emotional eating. J Am Acad Child Adolesc Psychiatry. (2019) 58:211–220.e5. doi: 10.1016/j.jaac.2018.06.039
119. Lips MA, Wijngaarden MA, van der Grond J, van Buchem MA, de Groot GH, Rombouts SARB, et al. Resting-state functional connectivity of brain regions involved in cognitive control, motivation, and reward is enhanced in obese females. Am J Clin Nutr. (2014) 100:524–31. doi: 10.3945/ajcn.113.080671
120. Simon JJ, Stopyra MA, Mönning E, Sailer S, Lavandier N, Kihm LP, et al. Neuroimaging of hypothalamic mechanisms related to glucose metabolism in anorexia nervosa and obesity. J Clin Invest. (2020) 130:4094–103. doi: 10.1172/JCI136782
121. van de Sande-Lee S, Pereira FRS, Cintra DE, Fernandes PT, Cardoso AR, Garlipp CR, et al. Partial reversibility of hypothalamic dysfunction and changes in brain activity after body mass reduction in obese subjects. Diabetes. (2011) 60:1699–704. doi: 10.2337/db10-1614
122. Cowdrey FA, Filippini N, Park RJ, Smith SM, McCabe C. Increased resting state functional connectivity in the default mode network in recovered anorexia nervosa. Hum Brain Mapp. (2014) 35:483–91. doi: 10.1002/hbm.22202
123. Lee S, Ran Kim K, Ku J, Lee J-H, Namkoong K, Jung Y-C. Resting-state synchrony between anterior cingulate cortex and precuneus relates to body shape concern in anorexia nervosa and bulimia nervosa. Psychiatry Res. (2014) 221:43–8. doi: 10.1016/j.pscychresns.2013.11.004
124. Kim KR, Ku J, Lee J-H, Lee H, Jung Y-C. Functional and effective connectivity of anterior insula in anorexia nervosa and bulimia nervosa. Neurosci Lett. (2012) 521:152–7. doi: 10.1016/j.neulet.2012.05.075
125. Boehm I, Geisler D, King JA, Ritschel F, Seidel M, Deza Araujo Y, et al. Increased resting state functional connectivity in the fronto-parietal and default mode network in anorexia nervosa. Front Behav Neurosci. (2014) 8. doi: 10.3389/fnbeh.2014.00346
126. Collantoni E, Michelon S, Tenconi E, Degortes D, Titton F, Manara R, et al. Functional connectivity correlates of response inhibition impairment in anorexia nervosa. Psychiatry Res Neuroimaging. (2016) 247:9–16. doi: 10.1016/j.pscychresns.2015.11.008
127. Corbetta M, Shulman GL. Control of goal-directed and stimulus-driven attention in the brain. Nature Reviews Neuroscience. (2002) 3:201–15. doi: 10.1038/nrn755
128. Phillipou A, Abel LA, Castle DJ, Hughes ME, Nibbs RG, Gurvich C, et al. Resting state functional connectivity in anorexia nervosa. Psychiatry Res Neuroimaging. (2016) 251:45–52. doi: 10.1016/j.pscychresns.2016.04.008
129. Di Martino A, Scheres A, Margulies DS, Kelly AMC, Uddin LQ, Shehzad Z, et al. Functional connectivity of human striatum: a resting state FMRI study. Cereb Cortex. (2008) 18:2735–47. doi: 10.1093/cercor/bhn041
130. Steinglass JE, Walsh BT. Neurobiological model of the persistence of anorexia nervosa. J Eat Disord. (2016) 4:19. doi: 10.1186/s40337-016-0106-2
131. Pike KM. Long-term course of anorexia nervosa: response, relapse, remission, and recovery. Clin Psychol Rev. (1998) 18:447–75. doi: 10.1016/S0272-7358(98)00014-2
132. Lotter LD, von Polier G, Offermann J, Buettgen K, Stanetzky L, Eickhoff SB, et al. Recovery-associated resting-state activity and connectivity alterations in anorexia nervosa. Biol Psychiatry Cogn Neurosci Neuroimaging. (2021) 6:1023–33. doi: 10.1016/j.bpsc.2021.03.006
133. Boehm I, Geisler D, Tam F, King JA, Ritschel F, Seidel M, et al. Partially restored resting-state functional connectivity in women recovered from anorexia nervosa. J Psychiatry Neurosci. (2016) 41:377–85. doi: 10.1503/jpn.150259
134. Gaudio S, Wiemerslage L, Brooks SJ, Schiöth HB. A systematic review of resting-state functional-MRI studies in anorexia nervosa: Evidence for functional connectivity impairment in cognitive control and visuospatial and body-signal integration. Neurosci Biobehav Rev. (2016) 71:578–89. doi: 10.1016/j.neubiorev.2016.09.032
135. Zink CF, Weinberger DR. Cracking the moody brain: The rewards of self starvation. Nat Med. (2010) 16:1382–3. doi: 10.1038/nm1210-1382
136. Haynos AF, Hall LMJ, Lavender JM, Peterson CB, Crow SJ, Klimes-Dougan B, et al. Resting state functional connectivity of networks associated with reward and habit in anorexia nervosa. Hum Brain Mapp. (2019) 40:652–62. doi: 10.1002/hbm.24402
137. Keating C, Tilbrook AJ, Rossell SL, Enticott PG, Fitzgerald PB. Reward processing in anorexia nervosa. Neuropsychologia. (2012) 50:567–75. doi: 10.1016/j.neuropsychologia.2012.01.036
138. Tataranni PA, Gautier J-F, Chen K, Uecker A, Bandy D, Salbe AD, et al. Neuroanatomical correlates of hunger and satiation in humans using positron emission tomography. Proc Natl Acad Sci U S A. (1999) 96:4569–74. doi: 10.1073/pnas.96.8.4569
139. Matsuda M, Liu Y, Mahankali S, Pu Y, Mahankali A, Wang J, et al. Altered hypothalamic function in response to glucose ingestion in obese humans. Diabetes. (1999) 48:1801–6. doi: 10.2337/diabetes.48.9.1801
140. Liu Y, Gao JH, Liu HL, Fox PT. The temporal response of the brain after eating revealed by functional MRI. Nature. (2000) 405:1058–62. doi: 10.1038/35016590
141. Benzo CA. Minireview. The hypothalamus and blood glucose regulation. Life Sci. (1983) 32:2509–15. doi: 10.1016/0024-3205(83)90231-X
142. Smeets PAM, de Graaf C, Stafleu A, van Osch MJP, van der Grond J. Functional MRI of human hypothalamic responses following glucose ingestion. Neuroimage. (2005) 24:363–8. doi: 10.1016/j.neuroimage.2004.07.073
143. Purnell JQ, Klopfenstein BA, Stevens AA, Havel PJ, Adams SH, Dunn TN, et al. Brain functional magnetic resonance imaging response to glucose and fructose infusions in humans. Diabetes Obes Metab. (2011) 13:229–34. doi: 10.1111/j.1463-1326.2010.01340.x
144. Osada T, Suzuki R, Ogawa A, Tanaka M, Hori M, Aoki S, et al. Functional subdivisions of the hypothalamus using areal parcellation and their signal changes related to glucose metabolism. Neuroimage. (2017) 162:1–12. doi: 10.1016/j.neuroimage.2017.08.056
145. Sun X, Kroemer NB, Veldhuizen MG, Babbs AE, de Araujo IE, Gitelman DR, et al. Basolateral amygdala response to food cues in the absence of hunger is associated with weight gain susceptibility. J Neurosci. (2015) 35:7964–76. doi: 10.1523/JNEUROSCI.3884-14.2015
146. Killgore WDS, Young AD, Femia LA, Bogorodzki P, Rogowska J, Yurgelun-Todd DA. Cortical and limbic activation during viewing of high- versus low-calorie foods. Neuroimage. (2003) 19:1381–94. doi: 10.1016/S1053-8119(03)00191-5
147. Schur EA, Kleinhans NM, Goldberg J, Buchwald D, Schwartz MW, Maravilla K. Activation in brain energy regulation and reward centers by food cues varies with choice of visual stimulus. Int J Obes (Lond). (2009) 33:653–61. doi: 10.1038/ijo.2009.56
148. van der Laan LN, de Ridder DTD, Viergever MA, Smeets PAM. The first taste is always with the eyes: A meta-analysis on the neural correlates of processing visual food cues. Neuroimage. (2011) 55:296–303. doi: 10.1016/j.neuroimage.2010.11.055
149. Herwig U, Dhum M, Hittmeyer A, Opialla S, Scherpiet S, Keller C, et al. Neural Signaling of Food Healthiness Associated with Emotion Processing. Front Aging Neurosci. (2016) 8:16. doi: 10.3389/fnagi.2016.00016
150. Killgore WDS, Yurgelun-Todd DA. Developmental changes in the functional brain responses of adolescents to images of high and low-calorie foods. Dev Psychobiol. (2005) 47:377–97. doi: 10.1002/dev.20099
151. Killgore WDS, Yurgelun-Todd DA. Body mass predicts orbitofrontal activity during visual presentations of high-calorie foods. Neuroreport. (2005) 16:859–63. doi: 10.1097/00001756-200505310-00016
152. Uher R, Treasure J, Heining M, Brammer MJ, Campbell IC. Cerebral processing of food-related stimuli: Effects of fasting and gender. Behav Brain Res. (2006) 169:111–9. doi: 10.1016/j.bbr.2005.12.008
153. Killgore WDS, Yurgelun-Todd DA. Affect modulates appetite-related brain activity to images of food. Int J Eat Disord. (2006) 39:357–63. doi: 10.1002/eat.20240
154. Coletta M, Platek S, Mohamed FB, van Steenburgh JJ, Green D, Lowe MR. Brain activation in restrained and unrestrained eaters: an fMRI study. J Abnorm Psychol. (2009) 118:598–609. doi: 10.1037/a0016201
155. Kaurijoki S, Kuikka JT, Niskanen E, Carlson S, Pietiläinen KH, Pesonen U, et al. Association of serotonin transporter promoter regulatory region polymorphism and cerebral activity to visual presentation of food. Clin Physiol Funct Imaging. (2008) 28:270–6. doi: 10.1111/j.1475-097X.2008.00804.x
156. Buffenstein R, Poppitt SD, McDevitt RM, Prentice AM. Food intake and the menstrual cycle: A retrospective analysis, with implications for appetite research. Physiol Behav. (1995) 58:1067–77. doi: 10.1016/0031-9384(95)02003-9
157. Baroncini M, Jissendi P, Catteau-Jonard S, Dewailly D, Pruvo J-P, Francke J-P, et al. Sex steroid hormones-related structural plasticity in the human hypothalamus. Neuroimage. (2010) 50:428–33. doi: 10.1016/j.neuroimage.2009.11.074
158. Blechert J, Meule A, Busch NA, Ohla K. Food-pics: an image database for experimental research on eating and appetite. Front Psychol. (2014) 5. doi: 10.3389/fpsyg.2014.00617
159. Charbonnier L, van Meer F, van der Laan LN, Viergever MA, Smeets PAM. Standardized food images: A photographing protocol and image database. Appetite. (2016) 96:166–73. doi: 10.1016/j.appet.2015.08.041
160. Jastreboff AM, Sinha R, Lacadie C, Small DM, Sherwin RS, Potenza MN. Neural Correlates of Stress- and Food Cue–Induced Food Craving in Obesity. Diabetes Care. (2013) 36:394–402. doi: 10.2337/dc12-1112
161. Mehta S, Melhorn SJ, Smeraglio A, Tyagi V, Grabowski T, Schwartz MW, et al. Regional brain response to visual food cues is a marker of satiety that predicts food choice. Am J Clin Nutr. (2012) 96:989–99. doi: 10.3945/ajcn.112.042341
162. Chin S-H, Kahathuduwa CN, Stearns MB, Davis T, Binks M. Is hunger important to model in fMRI visual food-cue reactivity paradigms in adults with obesity and how should this be done? Appetite. (2018) 120:388–97. doi: 10.1016/j.appet.2017.09.012
163. Gautier JF, Chen K, Salbe AD, Bandy D, Pratley RE, Heiman M, et al. Differential brain responses to satiation in obese and lean men. Diabetes. (2000) 49:838–46. doi: 10.2337/diabetes.49.5.838
164. DelParigi A, Chen K, Salbe AD, Hill JO, Wing RR, Reiman EM, et al. Persistence of abnormal neural responses to a meal in postobese individuals. Int J Obes Relat Metab Disord. (2004) 28:370–7. doi: 10.1038/sj.ijo.0802558
165. Geha PY, Aschenbrenner K, Felsted J, O'Malley SS, Small DM. Altered hypothalamic response to food in smokers. Am J Clin Nutr. (2013) 97:15–22. doi: 10.3945/ajcn.112.043307
166. Rothemund Y, Preuschhof C, Bohner G, Bauknecht H-C, Klingebiel R, Flor H, et al. Differential activation of the dorsal striatum by high-calorie visual food stimuli in obese individuals. Neuroimage. (2007) 37:410–21. doi: 10.1016/j.neuroimage.2007.05.008
167. Stice E, Spoor S, Bohon C, Veldhuizen MG, Small DM. Relation of reward from food intake and anticipated food intake to obesity: a functional magnetic resonance imaging study. J Abnorm Psychol. (2008) 117:924–35. doi: 10.1037/a0013600
168. Stoeckel LE, Weller RE, Cook EW, Twieg DB, Knowlton RC, Cox JE. Widespread reward-system activation in obese women in response to pictures of high-calorie foods. Neuroimage. (2008) 41:636–47. doi: 10.1016/j.neuroimage.2008.02.031
169. Smith JB, Watson GDR, Liang Z, Liu Y, Zhang N, Alloway KD. A Role for the Claustrum in Salience Processing? Front Neuroanat. (2019) 13. doi: 10.3389/fnana.2019.00064
170. Batterink L, Yokum S, Stice E. Body mass correlates inversely with inhibitory control in response to food among adolescent girls: an fMRI study. Neuroimage. (2010) 52:1696–703. doi: 10.1016/j.neuroimage.2010.05.059
171. Wang G-J, Volkow ND, Telang F, Jayne M, Ma Y, Pradhan K, et al. Evidence of gender differences in the ability to inhibit brain activation elicited by food stimulation. Proc Natl Acad Sci U S A. (2009) 106:1249–54. doi: 10.1073/pnas.0807423106
172. Hinkle W, Cordell M, Leibel R, Rosenbaum M, Hirsch J. Effects of reduced weight maintenance and leptin repletion on functional connectivity of the hypothalamus in obese humans. PLoS ONE. (2013) 8:e59114. doi: 10.1371/journal.pone.0059114
173. Swick D, Ashley V, Turken AU. Left inferior frontal gyrus is critical for response inhibition. BMC Neurosci. (2008) 9:102. doi: 10.1186/1471-2202-9-102
174. Sweet LH, Hassenstab JJ, McCaffery JM, Raynor HA, Bond DS, Demos KE, et al. Brain response to food stimulation in obese, normal weight, and successful weight loss maintainers. Obesity (Silver Spring). (2012) 20:2220–5. doi: 10.1038/oby.2012.125
175. Le DSNT, Chen K, Pannacciulli N, Gluck M, Reiman EM, Krakoff J. Reanalysis of the obesity-related attenuation in the left dorsolateral prefrontal cortex response to a satiating meal using gyral regions-of-interest. J Am Coll Nutr. (2009) 28:667–73. doi: 10.1080/07315724.2009.10719799
176. van Opstal AM, Westerink AM, Teeuwisse WM, van der Geest MAM, van Furth EF, van der Grond J. Hypothalamic BOLD response to glucose intake and hypothalamic volume are similar in anorexia nervosa and healthy control subjects. Front Neurosci. (2015) 9:159. doi: 10.3389/fnins.2015.00159
177. Bergh C, Södersten P. Anorexia nervosa, self–starvation and the reward of stress. Nat Med. (1996) 2:21–2. doi: 10.1038/nm0196-21
178. Fladung A-K, Schulze UME, Schöll F, Bauer K, Grön G. Role of the ventral striatum in developing anorexia nervosa. Transl Psychiatry. (2013) 3:e315. doi: 10.1038/tp.2013.88
179. Holsen LM, Lawson EA, Blum J, Ko E, Makris N, Fazeli PK, et al. Food motivation circuitry hypoactivation related to hedonic and nonhedonic aspects of hunger and satiety in women with active anorexia nervosa and weight-restored women with anorexia nervosa. J Psychiatry Neurosci. (2012) 37:322–32. doi: 10.1503/jpn.110156
180. Brooks SJ, Rask-Andersen M, Benedict C, Schiöth HB. A debate on current eating disorder diagnoses in light of neurobiological findings: is it time for a spectrum model? BMC Psychiatry. (2012) 12:76. doi: 10.1186/1471-244X-12-76
181. Sanders N, Smeets PAM, van Elburg AA, Danner UN, van Meer F, Hoek HW, et al. Altered food-cue processing in chronically ill and recovered women with anorexia nervosa. Front Behav Neurosci. (2015) 9:46. doi: 10.3389/fnbeh.2015.00046
182. Schouten TM, Koini M, de Vos F, Seiler S, van der Grond J, Lechner A, et al. Combining anatomical, diffusion, and resting state functional magnetic resonance imaging for individual classification of mild and moderate Alzheimer's disease. NeuroImage: Clinical. (2016) 11:46–51. doi: 10.1016/j.nicl.2016.01.002
183. Chang Y-HA, Marshall A, Bahrami N, Mathur K, Javadi SS, Reyes A, et al. Differential sensitivity of structural, diffusion, and resting-state functional MRI for detecting brain alterations and verbal memory impairment in temporal lobe epilepsy. Epilepsia. (2019) 60:935–47. doi: 10.1111/epi.14736
Keywords: MRI, networks, anorexia, obesity, resting state, diffusion, hypothalamus
Citation: Roger C, Lasbleiz A, Guye M, Dutour A, Gaborit B and Ranjeva J-P (2022) The Role of the Human Hypothalamus in Food Intake Networks: An MRI Perspective. Front. Nutr. 8:760914. doi: 10.3389/fnut.2021.760914
Received: 19 August 2021; Accepted: 07 December 2021;
Published: 03 January 2022.
Edited by:
John David Herrington, Children's Hospital of Philadelphia, United StatesReviewed by:
Vincent Prevot, Institut National de la Santé et de la Recherche Médicale (INSERM), FranceCopyright © 2022 Roger, Lasbleiz, Guye, Dutour, Gaborit and Ranjeva. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Coleen Roger, Y29sZWVuLnJvZ2VyQGV0dS51bml2LWFtdS5mcg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.