
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Nutr., 21 June 2021
Sec. Nutrition and Metabolism
Volume 8 - 2021 | https://doi.org/10.3389/fnut.2021.651721
This article is part of the Research TopicMilks Mean More: The Role of Milk in Nutrition, Digestion and Metabolism Across the LifespanView all 26 articles
 Ayşenur Arslan1
Ayşenur Arslan1 Merve Kaplan1
Merve Kaplan1 Hatice Duman1
Hatice Duman1 Ayşe Bayraktar1,2
Ayşe Bayraktar1,2 Melih Ertürk2Bethany M. Henrick3,4
Melih Ertürk2Bethany M. Henrick3,4 Steven A. Frese4,5
Steven A. Frese4,5 Sercan Karav1*
Sercan Karav1*Colostrum is the first milk produced post-partum by mammals and is compositionally distinct from mature milk. Bovine colostrum has a long history of consumption by humans, and there have been a number of studies investigating its potential for applications in human nutrition and health. Extensive characterization of the constituent fractions has identified a wealth of potentially bioactive molecules, their potential for shaping neonatal development, and the potential for their application beyond the neonatal period. Proteins, fats, glycans, minerals, and vitamins are abundant in colostrum, and advances in dairy processing technologies have enabled the advancement of bovine colostrum from relative limitations of a fresh and unprocessed food to a variety of potential applications. In these forms, clinical studies have examined bovine colostrum as having the substantial potential to improve human health. This review discusses the macro-and micronutrient composition of colostrum as well as describing well-characterized bioactives found in bovine colostrum and their potential for human health. Current gaps in knowledge are also identified and future directions are considered in order to elevate the potential for bovine colostrum as a component of a healthy diet for a variety of relevant human populations.
Colostrum is the earliest milk produced from the mammary glands for the first few days after giving birth and is unique in its composition of essential nutrients, immune factors, and oligosaccharides that benefit the newborn (1, 2). In the case of cows, bovine colostrum is produced immediately after calving and quickly wanes to mature milk (3), which lacks the high level of beneficial nutrients found in bovine colostrum. There are several factors affecting the composition and physical properties of colostrum such as individuality, breed, parity, pre-partum nutrition, length of the dry period of cows, and time post-partum (4). Generally, colostrum has more fat, protein, peptides, non-protein nitrogen, ash, vitamins and minerals, hormones, growth factors, cytokines, nucleotides, and less lactose compared to mature milk content. The concentration of these compounds decreases rapidly in the first 3 days of lactation with the exception of lactose content (5–7).
While the consumption of human colostrum by infants has long been recognized as a source of critical bioactive proteins for infants (8), the consumption of animal colostrum is also practiced in many locations beyond the neonatal period (9, 10). In these cultures and regions, colostrum has long been consumed as a health food or for medicinal purposes, with cultural practices centered on the belief that animal colostrum was an important component of the development of healthy children and supportive of healthy or infirmed adults (9, 11, 12). While these cultural or regional beliefs are associated with this practice, the abundance of well-characterized bioactive compounds and selective prebiotic components of this food may further support this cultural knowledge from a scientific perspective.
Historically, liquid fresh colostrum was primarily consumed, but pasteurized colostrum is also commercially available as a standalone drink, though production remains small (13). In European cultures and elsewhere such as India, and Scandinavia, colostrum is also used in the production of cheeses and other traditional foods (14). More recently, dried colostrum is collected and processed as a dietary supplement, which is widely consumed for perceived health benefits (10). In the US and EU, colostrum supplements are marketed for a variety of health benefits, including boosting immunity and gastrointestinal (GI) health. While attractive in concept, there are limitations to this use of dried colostrum, which are typically in a pill or tablet form, given the limited amount of colostrum consumed relative to clinically studied consumption rates.
Still, colostrum is a complex biological fluid and contains significant components which are natural anti-microbial factors for stimulating the maturation of calf immunity (15). In addition, the development and function of the GI tract are shaped by colostrum intake (5, 6, 16–18), and it also affects the metabolic and endocrine systems as well as the nutritional state of neonatal calves (5, 6, 17). Colostrum has muscular-skeletal repair and growth potential in addition to its immune support function and many benefits to health because of its content of bioactive proteins (19). Further, some evidence suggests that the cytokines, immunoglobulins, growth factors, antimicrobial compounds, and maternal immune cells are transferred to the newborn with the feeding of colostrum to support neonatal immunity (20–22). Bovine colostrum has even been purported to treat viral and bacterial infections as a nutraceutical (23). Together, the existing evidence in support of colostrum suggests that there is potential for colostrum to have a significant role in supporting human health as well. While there are other studies which have begun to look at colostrum from other animals (24–27), this review explores the current knowledge on the bovine colostrum in the context of nutrition, its bioactive components, and its potential for human health and nutrition.
Milk composition changes dramatically over the course of lactation and bovine colostrum is compositionally and nutritionally distinct from mature milk (28). In contrast to mature milk, colostrum has a much higher protein and moderately higher fat content, with substantially less lactose (Table 1). This reflects the needs of the developing calf, where the passive transfer of immunoglobulins is critical for health (41).
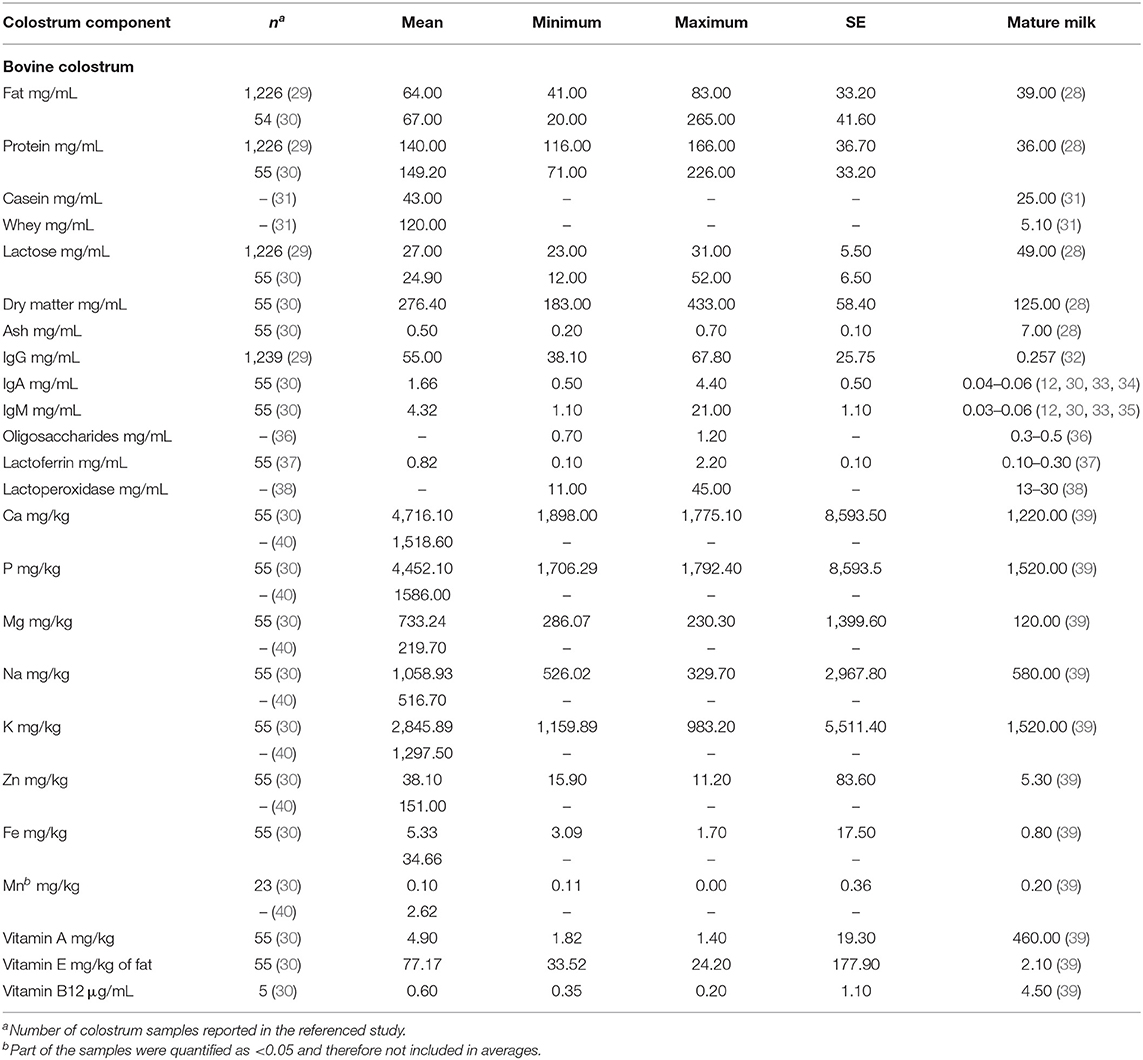
Table 1. Bovine colostrum and mature milk composition.
Further, as the volume of milk production increases over lactation, there is a concomitant decrease in the mineral content of milk (Table 1). Thus, colostrum represents a relatively high-protein and lower-carbohydrate solution that can be processed similarly to mature milk in order to reduce fat content and shape the caloric density for desired nutritional applications. Further, milk proteins are considered a “complete protein” source owing to their amino acid profile, and high protein digestibility, especially of whey proteins (42), though colostrum contains higher concentrations of immunoglobulins which are less digestible (Table 1).
While current dietary recommendations of protein intake for a healthy adult with minimal physical activity are 0.8 g per kg per day (43, 44), a growing body of evidence suggests that optimal intake may be higher [1.2–1.6 g per kg per day; (45–47)] and this intake should be balanced across meals to promote skeletal muscle protein synthesis (48). Especially in elderly populations, optimal protein intake to reduce skeletal muscle loss associated with aging is often not achieved, which is further compounded by diminished proteolytic activity associated with aging (49). Thus, colostrum may offer an attractive digestible, complete protein source that can be integrated into a calorically-appropriate diet. In addition to macronutrients, bovine colostrum includes vitamins, minerals, and a broad assortment of protein-derived bioactives which may offer additive benefits to its macronutrient profile.
The composition and quality of the bovine colostrum are highly variable due to genetic and environmental factors including individuality, breed, parity, the timing of milking, diseases, pre-partum nutrition, season, length of the dry period of cows, and time post-partum (50–52).
Bovine colostrum quality is different among individuals and between genetic backgrounds (31). For instance, the concentration of immunoglobulin G (IgG) in bovine colostrum and the volume of first milking vary among individual cattle (53, 54). The Jersey cows produce the highest (66.5 g/L) whereas Friesian-Holsteins produce the lowest (41.2 g/L) concentrations of IgG among breeds studied (55). In the case of cow parity, first-calf heifers produce a lower yield of colostrum and lower IgG concentration in colostrum than those cows in their second or greater lactation. The quality of bovine colostrum increases with parity after the second calving, and older cows generally produce the best quality colostrum (54).
Another individual factor is the disease which influences bovine colostrum quality. For instance, mastitis is an inflammation of the mammary gland of the bovine that has negative consequences including low quality of the colostrum. The volume and concentration of bovine colostrum IgG are lower in cows with infected mammary glands than cows with uninfected glands (56). The age of cows also affects the quality of colostrum. Some studies' data are in general agreement that older cows have a higher quality of colostrum than younger cows (53, 54, 57). The association between older age and good quality of colostrum is thought to be a result of increased pathogen exposure, improved immunity, and body condition score (31).
The timing of the bovine colostrum milking after parturition has significant effects on concentrations of IgG in the bovine colostrum. Early or immediate colostrum milking will significantly increase colostrum quality. Moore et al. (58) reported that colostrum collected 6, 10, and 14 h after parturition has lower IgG concentration than colostrum collected 2 h after parturition. Another study also showed that bovine colostrum quality is highest immediately after parturition of North American herds, but it decreased when milking was delayed (53, 58). Bovine colostrum quality is also affected by the calving season. Cows calving during the summer months have lower quality colostrum than those calving in the autumn months (53). The bovine colostrum fat percentage is at 24 and 48 h after birth is affected by the calving season. Animals born in autumn-winter seasons have a higher colostrum fat percentage than those in calving in spring-summer seasons. One cause may be differences in metabolism, feed, and water consumption in different seasons (59, 60).
The dry period length is an important period for cows which lasts ~6–8 weeks. This period is needed for the renewal of milk secretion tissue, preparation for lactation, and completion of fetus development (61–63). Colostrum starts to be secreted in the last 15–20 days of the dry period and its composition changes continued until parturition (62, 64). Le Cozler et al. (65) also reported that there is a positive coefficient of correlation (R2 = 0.22; P < 0.01) between IgG concentration and dry period length (65).
Colostrum contains a higher percentage of fat than mature milk (66) and the composition of these fats is also distinct. O'Callaghan et al. (67) examined the composition of colostrum and the changes observed during the transition to mature milk, reporting that colostrum is higher in palmitic, palmitoleic, and myristic acids, relative to mature milk (67). While these fat profiles are well-suited to the developing calf (68), the profiles of these fats and the higher concentration of saturated fat have been associated with long-term negative health outcomes, though there is some disagreement within the literature as to the level of support for the role of dairy fats in cardiovascular disease (69). There is evidence that these fatty acids play a role as signaling molecules and, as dietary fatty acids, contribute to the regulation of lipogenesis in the liver (70). Further, many vitamins found in milk are fat soluble (e.g., vitamin A, D) and removal of these fats also reduces the concentration of these vitamins in colostrum.
It is of relevance to consumers that the advances in dairy technology which enable efficient separation of fats from the aqueous fraction of milk (that is, the fraction which contains proteins, carbohydrates, minerals, and some vitamins) enable the reduction or removal of these fats from colostrum, ahead of downstream processing, making the potential for a low-fat or fat-free colostrum product possible. However, some have speculated that the tradeoff between dairy fats and the removal of bioactive found in the fat fraction of dairy foods may not always be a net benefit (69). To resolve this conflict in the literature, it is clear that well-controlled clinical studies investigating the relationship between the dietary fats found in colostrum and health are needed.
Bovine colostrum contains also high levels of fat-soluble and water-soluble vitamins that are critical to human health (4). Notably, vitamin A is reported to be found at high concentrations in colostrum in a variety of forms including retinol, retinal, retinoic acid, retinyl esters and as provitamin A carotenoids (71–73). Vitamin E, in the form of tocopherols and tocotrienols (~ mean 77.17 mg/kg) are found in low density lipoproteins in colostrum (4, 30). Vitamin K is also found in greater concentration in colostrum compared to mature milk in two forms, phylloquinone, and menaquinones (71). Vitamin D is found in higher concentrations in colostrum than mature milk (74). Vitamin D has important roles in immune activities and promotes the uptake of calcium and phosphorus in the small intestine (75). It has two major forms as cholecalciferol (vitamin D3) and ergocalciferol (vitamin D2) and their concentration decreases from 1.2 to 0.36 IU. g−1 during the first 5 days post-partum (76). Vitamin C and the B vitamins are also found in the water-soluble fraction of colostrum at a higher concentration compared to mature milk (77) and together provide a natural source of essential vitamins critical to human health.
Bovine colostrum and mature milk are known to be good sources of several minerals especially calcium and phosphorus (75). Recent studies revealed that the mean concentrations of several important minerals in colostrum are significantly higher than in mature bovine milk. Calcium is necessary for the maintenance of calf development and their healthy bones and teeth. Phosphorus is also crucial for the metabolic rate and physiological functions including development of skeletal tissue, energy utilization, protein synthesis, and transport of fatty acids (78). Magnesium is present in a relatively large amount, along with zinc and selenium in bovine colostrum (75).
Immunoglobulins (Igs) are complex proteins, known as antibodies, that make up a significant part of the total protein in bovine colostrum. The immunoglobulins in bovine colostrum mainly come in 3 different varieties called isotypes including IgG (IgG1 and IgG2), IgA, IgM. IgG is the dominant immunoglobulin in bovine colostrum, which makes up 85–90% of the total immunoglobulin content. IgG1 represents 80–90% of the total IgG content in bovine colostrum, followed by IgM, IgA, and IgG2 (23, 79, 80). These immunoglobulins are essential in the survival of the calves and their immune systems and they neutralize enteric pathogens such as bacteria, microbes, and viruses. Using bovine colostrum as a source of antibody preparations to support bovine and human health is an important research subject that has been studied for decades (81).
One of the key differences between mature milk and colostrum is the high concentration of IgG found in colostrum, which reaches up to 50–100 mg/mL in the first days after birth (33, 82, 83). Bovine serum IgG1 and IgG2 concentration decrease before parturition, they are transferred from the blood into the colostrum. In fact, nearly all IgG in colostrum is transferred from bovine serum into the colostrum and milk (84, 85).
The high concentration of IgG is necessary for the survival of calves, which is strongly dependent on the transfer of IgG from bovine colostrum to calves to provide passive immunity as cows cannot transfer IgG through the placenta (86). Indeed, if calves do not receive colostrum immediately after birth, they are prone to infection and will suffer from a higher risk of morbidity and mortality (31, 87, 88).
Lactoferrin is a cationic, iron-binding glycoprotein present as about 0.80 mg/mL in bovine colostrum (37). It has multiple functions including antibacterial, antifungal, antiviral, antiparasitic, antitumor and immunomodulatory (anti-inflammatory) effects (23, 35, 89, 90), and is the major protein in the milk serum of all mammals (91). Bovine colostrum-derived lactoferrin has antimicrobial activity by inhibiting the growth of disease-causing protozoa, yeasts, bacteria, and viruses, and lactoferrin may prevent the attachment of pathogens to epithelial cells and help maintain intestinal permeability and stability (83, 92, 93). Moreover, there are some studies showing that bovine colostrum-derived lactoferrin can increase the proliferation of cells involved in the bone formation such as osteoblasts, and the release of some growth factors from osteoblasts (94, 95).
Furthermore, it is known to play a role in iron uptake in the intestine and activation of phagocytes and immune responses. Receptors for lactoferrin are expressed on intestinal tissue, monocytes, macrophages, neutrophils, lymphocytes, platelets, and on some bacteria (96). Bovine lactoferrin supplements are thought to support the immune system and influence immune cell activity potentially via these antioxidant, antibacterial, and antiviral properties (97). The greatest concentration of this protein is found in colostrum, which has been determined to be four times greater than mature milk (98).
Lactoperoxidase is a major antibacterial enzyme found in bovine colostrum, it is a basic glycoprotein that catalyzes the oxidation of thiocyanate and generates intermediate compounds with antimicrobial activities (99). The concentration of lactoperoxidase is 11–45 mg/L in bovine colostrum and 13–30 mg/L in mature bovine milk (38). Its concentration in bovine colostrum is low initially, but it reaches the maximum level within 3–5 days after parturition. Lactoperoxidase catalase activity is also higher in bovine colostrum than in mature milk (100, 101).
Lactoperoxidase activity produces toxic oxidation products that inhibit bacterial metabolism by oxidation of essential sulfhydryl groups in proteins. This system is toxic to some gram-positive and negative bacteria like Pseudomonas aeruginosa, Salmonella typhimurium, Listeria monocytogenes, Streptococcus mutans, and Staphylococcus aureus (102). The lactoperoxidase system also inactivates the poliovirus, vaccinia virus, and HIV (93, 103, 104).
Bovine colostrum is a rich source of complex and highly selective oligosaccharides and glycans. The concentration of oligosaccharides in colostrum is 0.7–1.2 mg/mL and the majority of these structures are acidic oligosaccharides which are lower in mature bovine milk (36, 105). Forty distinct oligosaccharides compositions have been detected in bovine colostrum so far (106–108). The total colostrum oligosaccharides differ between cows because of their genetic variability (109). Predominant oligosaccharides in bovine colostrum are 3′ sialyllactose (3′SL), 6′ sialyllactose (6′SL), 6′ siayllactosamine (6′SLN) and disialyllactose (DSL). 3'SL is 70% of total oligosaccharide content in bovine colostrum (105, 107, 110, 111). 3′SL, 6′SL, and 6′SLN levels in colostrum were highest following parturition and decreased by 48 h post-partum, while neutral oligosaccharide level increased (105). Breed specific differences have also been identified in oligosaccharide content. Concentrations of 3′SL, 6′SL, 6′SLN and DSL were found as 867, 136, 220, and 283 μg/mL, respectively, in colostrum from Jersey cows, while these concentrations were 681, 243, 239, and 201 μg/mL, respectively, in Holstein colostrum after parturition (112). Both free oligosaccharides (bovine milk oligosaccharides, BMOs) and complex, conjugated N-glycans represent the majority of the prebiotic components of bovine colostrum (113).
While there are many distinctions between BMOs and human milk oligosaccharides (HMOs), there has been significant interest in utilizing milk and colostrum as a source of BMOs for human nutrition and health to modulate the GI microbiome (114). In contrast to HMOs, BMOs are predominantly sialylated (i.e., acidic) oligosaccharides, with a low propensity for fucosylation (106) and a lower structural diversity (106). Recent advancements in enzymatic glycosylation have provided opportunities for the structural enhancement of BMOs to alter their structure to resemble HMOs (115). Several complexities in milk processing have thus far limited the ability of BMOs to be separated from lactose found at high concentrations in milk (114), though solutions have begun to emerge (116) which complicates their utility for human nutrition and health. Further, though pilot experiments with purified BMOs in adults have not yet demonstrated generalizable changes to GI microbial populations (117), future work in infants may be more promising as recent in vitro experiments with BMOs are more promising (118, 119).
Complex and hybrid N-glycans found in bovine mature milk and colostrum may also provide a source of prebiotic glycans that can be selectively utilized in a fashion similar to HMOs/BMOs (120). Further, the conjugation of these N-glycans to milk proteins enable different strategies for their recovery. Protein separation from lactose and subsequent treatment to separate N-glycans from their protein conjugates may offer a potentially attractive avenue to purification of these glycans (121). Thus, N-glycans derived from bovine colostrum, which is exceptionally rich in N-glycosylated proteins (122), may be a potent source of bioactive glycans to serve as prebiotic substrates. Extensive characterization of complex N-glycans derived from bovine milk proteins abundant in colostrum now shows that these N-glycans are highly selective for certain bacteria in the adult GI microbiome. The bacteria able to access these glycans are further restricted, relative to larger repeated polymers of less complex oligosaccharides which are limited to select Bifidobacterium species (e.g., Bifidobacterium longum subsp. infantis). Some strains of these species have been associated with diminished enteric inflammation and improved GI barrier function in humans (123, 124). Bovine colostrum is also a potential source of anti-infective glycans and recent work provides evidence for the anti-infective activity of oligosaccharides sourced from bovine colostrum against a highly invasive strain of C. jejuni (125).
The first study investigating the role of colostrum supplementation in exercise performance was completed in 1997 and showed marked improvements on explosive muscle power and increased concentration of immunoglobulins in serum (35). This finding is relevant given intense physical activity can suppress immunity several hours after training (126). Subsequent, well-controlled studies in comparison to whey protein concentrate have demonstrated significant improvements in lean body mass and weightlifting performance (127), in athletic performance among male and female athletes (128), speed in elite cyclists with dose-dependent effects (129), and in runners for recovery (130). Duff et al. (131) indicated that bovine colostrum supplementation (60 g/d of colostrum) on male and female older adults during resistance training is beneficial for increasing leg press strength and reducing bone resorption in comparison to whey protein complex supplementation. Improvement in the upper body strength, muscle thickness, lean tissue mass, and cognitive function were noted for colostrum supplemented group as well as whey protein treated group (131).
Despite this progress, the exact mechanism behind these marked improvements is not fully elucidated. As human studies typically use whey protein with similar protein content, observed differences are unlikely to simply be a response to protein digestibility or amino acid supplementation. Given that bovine colostrum immune components are likely not providing passive immunity to the human, it is possible that bioactive compounds and/or their metabolites have a direct effect on the immune system (35, 126, 132–135). There is currently weak support for the potential for bovine colostrum supplementation to improve leukocyte function relating to adaptive immunity (126). While a 33% increase in saliva IgA was noted after supplementation of colostrum at 20 g/day for 2 weeks (134) and a 79% increase in IgA in runners fed 12 g/day for 12 weeks was reported (132), these results were not repeated in other studies (35, 135–140). Further, colostrum supplementation diminished exercise-induced intestinal permeability which was replicated in in vitro culture models of intestinal epithelial cells (141). Considering the safety profile and generally positive past research from well-controlled studies, further research is warranted to understand the underlying mechanism and explain inter-individual variations and unexplored discrepancies between the growing number of studies on colostrum supplementation in regards to body composition and athletic performance.
Non-steroidal anti-inflammatory drugs (NSAIDs) are the most common prescribed medicine and used for the symptomatic treatment of acute pain, chronic inflammatory, and degenerative joint diseases (142, 143). NSAIDs can cause gastric and intestinal damage such as peptic ulceration and injury to both the small and large intestine. Complications from NSAIDs use include increased intestinal permeability with protein and blood loss, and also stricture formation (142, 144). Approximately 2% of subjects taking NSAIDs experience adverse effects on the GI tract including bleeding, perforation, and inflammation. Acid suppressants and prostaglandin analogs are used to reduce gastric injury induced by NSAIDs, but these are not adequately effective in preventing small intestine injury. Hence, additional avenues for the mitigation of these negative side effects are needed. Some research suggests that colostrum may be an alternative, owing to the composition of growth factors like α-IGF-1, β-IGF-1, transforming growth factor (TGF), and epidermal growth factors (EGF). These growth factors are capable of stimulating the repair process of the GI tract (145) and are complementary to evidence supporting diminished GI permeability associated with exercise (141).
In a clinical examination of bovine colostrum for protection against NSAID-induced enteropathy, seven male volunteers (26–38 years old) who were taking NSAIDs or suffering from conditions likely to affect intestinal permeability (e.g., coeliac disease or previous intestinal surgery), were evaluated for the potential of bovine colostrum to alter intestinal permeability with concomitant indomethacin supplementation. In this crossover study, following an initial baseline permeability assessment, these volunteers were supplemented orally with 125 mL of bovine colostrum or a whey placebo three times daily for seven days. At the end of the trial period, intestinal permeability was reassessed and a 2-week “washout” period was performed between the crossover. Approximately a 3-fold increase in permeability was observed in the participants taking the whey placebo with indomethacin, while no significant increase in permeability was seen in the participants taking co-administration of bovine colostrum with indomethacin (146). In support of these findings, molecular characterization of the mechanism underlying these effects have been documented. Mir et al., (147) demonstrated that bovine lactoferrin can act as a carrier for NSAIDs by binding to these molecules, but with far lower affinity than the protein targets for NSAIDs which suggests that the efficacy of NSAIDs may not be affected by co-administration of a lactoferrin-containing protein source, like bovine colostrum (147). While further studies will be required to demonstrate that these compounds, when co-administered with bovine colostrum, maintain their desired efficacy, there is growing and consistent evidence supporting the potential for the use of bovine colostrum to manage the potential side effects of NSAIDs.
Bovine colostrum has led to human supplementation trials due to potential for improvement of GI health and integrity. Several conditions related to GI conditions associated with chronic or acute infections have been investigated for the potential of bovine colostrum to ameliorate symptoms associated with these conditions or infections. While the mechanisms behind these findings are difficult to disentangle given the disparate populations and disease etiologies, there are consistent themes related to the improvement of GI symptoms and reduced inflammation associated with each, though not all conditions demonstrate promising avenues for therapy.
Ulcerative colitis, an inflammatory bowel disease associated with durable inflammation and ulcers in the colon (148), was investigated as a potential target for a bovine colostrum enema treatment in a small proof-of-concept trial. The authors rationalized this approach given the high concentrations of antimicrobial peptides, immunoglobulins and growth factors found in colostrum (149). In this pilot study, fourteen patients with active mild to moderate colitis were compared. Eight patients received 100 mL (10% solution) of bovine colostrum and six patients received an albumin placebo twice per day for 4 weeks. Improvement of the symptom score including patient well-being, abdominal pain, rectal bleeding, temperature, anorexia/nausea, bowel frequency, stool consistency, abdominal tenderness and the presence of extra-intestinal manifestations was reported in seven of the eight patients in the bovine colostrum treated groups (149). While this study is small, the findings show a significant reduction in symptom scores and follow up studies in a similar population with a larger sample size may be warranted.
Necrotizing enterocolisis (NEC) is one of the most common morbidities associated with preterm birth, and among the chief causes of mortality among infants born preterm (150). Several studies have examined the impact of either human or bovine-derived colostrum on NEC outcomes and development of preterm infants. In one clinical trial of 86 low birthweight infants supplemented with bovine colostrum in a dose of 2 g, four times per day for infants between 1,000 and 1,500 g and 1.2 g, four times per day for those under 1,000 g at birth. No significant differences were observed in the occurrence of NEC, sepsis, or mortality after the administration of bovine colostrum as compared with placebo (151). In a meta-analysis examining the use of bovine and human colostrum among preterm infants, Sadeghirad et al. concluded that the cumulative findings in the literature suggest that neither human nor bovine colostrum had an effect on the incidence of severe NEC, mortality, culture-proven sepsis, feed intolerance, or length (152). The lack of effects is observed on NEC patients due to some limitations such as using commercial bovine colostrum supplement and number of patients is modest (146, 147). Given these findings, it will be difficult to rationalize continued use of bovine or human colostrum with preterm infants for improvements in these outcomes. However, the use of human colostrum in preterm infants should not be curtailed based on these outcomes as other benefits have been demonstrated for preterm infants (153).
Acute infection with enterotoxigenic Escherichia coli (ETEC) represents the most common causes of so-called Traveler's Diarrhea, associated with travel to tropic and semitropical regions throughout the world (154). As bovine colostrum plays a key role in protecting the neonatal calf from environmental pathogens via passive immunity and ETEC represents one of the primary causative agents of neonatal calf diarrhea (155), researchers have been interested in determining whether the same effects can be demonstrated in humans at risk of Traveler's Diarrhea.
Using hyperimmune bovine colostrum which is rich in immunoglobulins targeting 14 strains of ETEC, the efficacy and dose response of consuming bovine colostrum in a tablet form (400 mg of bovine colostrum protein) demonstrated a dose-dependent and significant improvement in protecting against the development of diarrhea among volunteers in a double blinded, placebo-controlled ETEC challenge study. A 400 mg serving of hyperimmune bovine colostrum protein administered with a bicarbonate buffer three times daily conferred 90.9% protection when compared to the placebo (156). Bicarbonate buffer contributes to the enhancement the protective effects of hyperimmune colostrum protein in the ETEC challenge experiments, but the difference was not statistically significant. As little as 200 mg consumed three times per day without buffer gave an estimated 58.3% protection from diarrhea symptoms, compared to the placebo group (156).
In addition to ETEC, viruses contribute to a significant proportion of both neonatal calf diarrhea and Traveler's Diarrhea (154, 155). In a double blinded, placebo-controlled study, Mitra et al., (157) reported that consuming three daily servings of 100 mL of hyperimmune bovine colostrum targeting human rotavirus for 3 days conferred a modest but significant reduction in both the duration of diarrhea and the total stool output among male infants 6–24 months of age (157). Similarly, another study reported that purified immunoglobulins from hyperimmune bovine colostrum conferred a similar effect in acute rotavirus infection, supporting these findings (158).
While neither study examined the impact of colostrum from cows which had not been immunized against the target pathogen, a clinical trial examining the differences between hyperimmune bovine colostrum and bovine colostrum among children infected with shigellosis caused by Shigella dysenteriae (S. dysenteriae) failed to find any improvements among patients relative to the concurrent antibiotic therapy. However, preclinical studies in other biomedical models (e.g., gnotobiotic pigs) have shown promise for hyperimmune bovine colostrum in preventing diarrhea caused by Clostridioides difficile (C. difficile). Together, these findings may suggest that infectious mechanisms of pathogenesis shape the ability of hyperimmune bovine colostrum to influence disease progression as S. dysenteriae invades epithelial cells (159), potentially evading hyperimmune bovine colostrum immunoglobulins, while ETEC and C. difficile utilize secreted toxins to induce epithelial damage (160, 161).
Given the biological role of colostrum for neonates (8, 162), its documented bioactive components as outlined here, and the potential for development as a functional food or food ingredient. There is a significant interest in developing colostrum as an ingredient to improve the bioactivity of foods and/or their potential health benefits. With a higher protein content and lower lactose concentration, this favorable protein/carbohydrate ratio is also nutritionally attractive and the potential for the future development of the ingredients and constituent fractions of colostrum is promising. However, overcoming processing challenges to separate bioactive fractions from colostrum remains a challenge to both study the mechanisms by which this fluid can act on humans and for practical product development. Future clinical trials should address current gaps in understanding which populations, such as those with GI disorders, may benefit most from colostrum consumption and whether whole or fractionated colostrum offers the most attractive balance of nutrition and bioactive properties.
SK organized the general content of the paper. AA was responsible for general editing and organizing the authors and also responsible for the two sections of the paper. MK contributed one section of the paper. HD was responsible for writing one section of the paper. AB was responsible for the one section of the paper. ME contributed to a section of the paper. BMH was responsible for the organizing a section. SAF contributed editing and organizing the paper. All authors contributed to the article and approved the submitted version.
BMH is an employee of Evolve Biosystems, Inc. SK has received funding from Uluova Süt Ticaret A.Ş (Uluova Milk Trading Co.), a company focused on the production of colostrum and lactoferrin. ME and AB are employees of Uluova Dairy.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Uluova Süt Ticaret A.Ş (Uluova Milk Trading Co.) is funding to achievement of this study.
1. Godden SM, Smolenski DJ, Donahue M, Oakes JM, Bey R, Wells S, et al. Heat-treated colostrum and reduced morbidity in preweaned dairy calves: results of a randomized trial and examination of mechanisms of effectiveness. J Dairy Sci. (2012) 95:4029–40. doi: 10.3168/jds.2011-5275
2. Rathe M, Müller K, Sangild PT, Husby S. Clinical applications of bovine colostrum therapy: a systematic review. Nutr Rev. (2014) 72:237–54. doi: 10.1111/nure.12089
3. Larson BL, Heary HL, Devery JE. Immunoglobulin production and transport by the mammary gland. J Dairy Sci. (1980) 63:665–71. doi: 10.3168/jds.S0022-0302(80)82988-2
4. McGrath BA, Fox PF, McSweeney PLH, Kelly AL. Composition and properties of bovine colostrum: a review. Dairy Sci Technol. (2016) 96:133–58. doi: 10.1007/s13594-015-0258-x
5. Blum JW, Hammon HM. Bovine colostrum - more than just an immunoglobulin supplier. Schweiz Arch Tierheilkd. (2000) 142:221–8.
6. Blum J, Hammon H. Colostrum effects on the gastrointestinal tract, and on nutritional, endocrine and metabolic parameters in neonatal calves. Livest Prod Sci. (2000) 66:151–9. doi: 10.1016/S0301-6226(00)00222-0
7. Uruakpa F, Ismond MA, Akobundu EN. Colostrum and its benefits: a review. Nutr Res. (2002) 22:755–67. doi: 10.1016/S0271-5317(02)00373-1
8. Gephart SM, Weller M. Colostrum as oral immune therapy to promote neonatal health. Adv Neonatal Care. (2014) 14:44–51. doi: 10.1097/ANC.0000000000000052
9. Dzik S, Miciński B, Aitzhanova I, Miciński J, Pogorzelska J, Beisenov A, et al. Properties of bovine colostrum and the possibilities of use. Polish Ann Med. (2017) 24:295–9. doi: 10.1016/j.poamed.2017.03.004
10. Silva EG, Rangel AH, Mürmam L, Bezerra MF, Oliveira JP. Bovine colostrum: benefits of its use in human food. Food Sci Technol. (2019) 39:355–62. doi: 10.1590/fst.14619
11. Godhia M, Patel N. Colostrum - its composition, benefits as a nutraceutical : a review. Curr Res Nutr Food Sci J. (2013) 1:37–47. doi: 10.12944/CRNFSJ.1.1.04
14. Rocha JM. Scientific and Medical Research Related To Bovine Colostrum Its Relationship and Use in the Treatment of Disease in Humans Selected (2016).
15. van Hooijdonk ACM, Kussendrager KD, Steijns JM. In vivo antimicrobial and antiviral activity of components in bovine milk and colostrum involved in non-specific defence. Br J Nutr. (2000) 84:127–34. doi: 10.1017/S000711450000235X
16. Hadorn U, Hammon H, Bruckmaier RM, Blum JW. Delaying colostrum intake by one day has important effects on metabolic traits and on gastrointestinal and metabolic hormones in neonatal calves. J Nutr. (1997) 127:2011–23. doi: 10.1093/jn/127.10.2011
17. Guilloteau P, Huërou-Luron Le I, Chayvialle JA, Toullec R, Zabielski R, Blum JW. Gut regulatory peptides in young cattle and sheep. J Vet Med Ser A. (1997) 44:1–23. doi: 10.1111/j.1439-0442.1997.tb01082.x
18. Bühler C, Hammon H, Rossi GL, Blum JW. Small intestinal morphology in eight-day-old calves fed colostrum for different durations or only milk replacer and treated with long-R3-insulin-like growth factor i and growth hormone. J Anim Sci. (1998) 76:758–65. doi: 10.2527/1998.763758x
19. Korhonen HJ. Production and properties of health-promoting proteins and peptides from bovine colostrum and milk. Cell Mol Biol. (2013) 59:12–24. doi: 10.1170/T943
20. Goto M, Maruyama M, Kitadate K, Kirisawa R, Obata Y, Koiwa M, et al. Detection of Interleukin-1.BETA. In sera and colostrum of dairy cattle and in sera of neonates. J Vet Med Sci. (1997) 59:437–41. doi: 10.1292/jvms.59.437
21. Reber AJ, Lockwood A, Hippen AR, Hurley DJ. Colostrum induced phenotypic and trafficking changes in maternal mononuclear cells in a peripheral blood leukocyte model for study of leukocyte transfer to the neonatal calf. Vet Immunol Immunopathol. (2006) 109:139–50. doi: 10.1016/j.vetimm.2005.08.014
22. Yamanaka H, Hagiwara K, Kirisawa R, Iwai H. Proinflammatory cytokines in bovine colostrum potentiate the mitogenic response of peripheral blood mononuclear cells from newborn calves through IL-2 and CD25 expression. Microbiol Immunol. (2003) 47:461–8. doi: 10.1111/j.1348-0421.2003.tb03371.x
23. Bagwe S, Tharappel LJP, Kaur G, Buttar HS. Bovine colostrum: an emerging nutraceutical. J Complement Integr Med. (2015) 12:175–85. doi: 10.1515/jcim-2014-0039
24. Bernabucci U, Basiricó L, Morera P. Impact of hot environment on colostrum and milk composition. Cell Mol Biol. (2013) 59:67–83. doi: 10.1170/T948
25. Hernández-Castellano LE, Almeida AM, Renaut J, Argüello A, Castro N. A proteomics study of colostrum and milk from the two major small ruminant dairy breeds from the Canary Islands: a bovine milk comparison perspective. J Dairy Res. (2016) 83:366–74. doi: 10.1017/S0022029916000273
26. Park YW, Juárez M, Ramos M, Haenlein GFW. Physico-chemical characteristics of goat and sheep milk. Small Rumin Res. (2007) 68:88–113. doi: 10.1016/j.smallrumres.2006.09.013
27. Roy D, Ye A, Moughan PJ, Singh H. Composition, structure, and digestive dynamics of milk from different species—a review. Front Nutr. (2020) 7:577759. doi: 10.3389/fnut.2020.577759
28. Foley JA, Otterby DE. Availability, storage, treatment, composition, and feeding value of surplus colostrum: a review. J Dairy Sci. (1978) 61:1033–60. doi: 10.3168/jds.S0022-0302(78)83686-8
29. Dunn A, Ashfield A, Earley B, Welsh M, Gordon A, Morrison SJ. Evaluation of factors associated with immunoglobulin G, fat, protein, and lactose concentrations in bovine colostrum and colostrum management practices in grassland-based dairy systems in Northern Ireland. J Dairy Sci. (2017) 100:2068–79. doi: 10.3168/jds.2016-11724
30. Kehoe SI, Jayarao BM, Heinrichs AJ. A survey of bovine colostrum composition and colostrum management practices on pennsylvania dairy farms. J Dairy Sci. (2007) 90:4108–16. doi: 10.3168/jds.2007-0040
31. Godden S. Colostrum management for dairy calves. Vet Clin North Am Food Anim Pract. (2008) 24:19–39. doi: 10.1016/j.cvfa.2007.10.005
32. Conesa C, Lavilla M, Sánchez L, Pérez MD, Mata L, Razquín P, et al. Determination of IgG levels in bovine bulk milk samples from different regions of Spain. Eur Food Res Technol. (2005) 220:222–5. doi: 10.1007/s00217-004-1016-0
33. Stelwagen K, Carpenter E, Haigh B, Hodgkinson A, Wheeler TT. Immune components of bovine colostrum and milk1. J Anim Sci. (2009) 87:3–9. doi: 10.2527/jas.2008-1377
34. Pakkanen R, Aalto J. Growth factors and antimicrobial factors of bovine colostrum. Int Dairy J. (1997) 7:285–97. doi: 10.1016/S0958-6946(97)00022-8
35. Mero A, Miikkulainen H, Riski J, Pakkanen R, Aalto J, Takala T. Effects of bovine colostrum supplementation on serum IGF-I, IgG, hormone, and saliva IgA during training. J Appl Physiol. (1997) 83:1144–51. doi: 10.1152/jappl.1997.83.4.1144
36. ten Bruggencate SJ, Bovee-Oudenhoven IM, Feitsma AL, van Hoffen E, Schoterman MH. Functional role and mechanisms of sialyllactose and other sialylated milk oligosaccharides. Nutr Rev. (2014) 72:377–89. doi: 10.1111/nure.12106
37. Buttar HS, Bagwe SM, Bhullar SK, Kaur G. Health benefits of bovine colostrum in children and adults. In: Watson RR, Collier RJ, Preedy VR, editors. Dairy in Human Health and Disease Across the Lifespan. Elsevier (2017). p. 3–20.
38. Hahn R, Schulz P, Schaupp C, Jungbauer A. Bovine whey fractionation based on cation-exchange chromatography. J Chromatogr A. (1998) 795:277–87. doi: 10.1016/S0021-9673(97)01030-3
39. Khan IT, Nadeem M, Imran M, Ullah R, Ajmal M, Jaspal MH. Antioxidant properties of milk and dairy products: a comprehensive review of the current knowledge. Lipids Health Dis. (2019) 18:41. doi: 10.1186/s12944-019-0969-8
40. Dande ND, Nande J. Nutritional composition of bovine colostrum: palatability evaluation of food products prepared using bovine colostrum. Int J Nutr Pharmacol Neurol Dis. (2020) 10:8–13. doi: 10.4103/ijnpnd.ijnpnd_77_19
41. Weaver DM, Tyler JW, VanMetre DC, Hostetler DE, Barrington GM. Passive transfer of colostral immunoglobulins in calves. J Vet Intern Med. (2000) 14:569–77. doi: 10.1111/j.1939-1676.2000.tb02278.x
42. Stelwagen K. Mammary gland, milk biosynthesis and secretion milk protein. In: Fuquay JW, editor. Encyclopedia of Dairy Sciences. Elsevier (2011). p. 359–66.
43. Richter M, Baerlocher K, Bauer JM, Elmadfa I, Heseker H, Leschik-Bonnet E, et al. Revised reference values for the intake of protein. Ann Nutr Metab. (2019) 74:242–50. doi: 10.1159/000499374
44. Wu G. Dietary protein intake and human health. Food Funct. (2016) 7:1251–65. doi: 10.1039/C5FO01530H
45. Dong J-Y, Zhang Z-L, Wang P-Y, Qin L-Q. Effects of high-protein diets on body weight, glycaemic control, blood lipids and blood pressure in type 2 diabetes: meta-analysis of randomised controlled trials. Br J Nutr. (2013) 110:781–9. doi: 10.1017/S0007114513002055
46. Santesso N, Akl EA, Bianchi M, Mente A, Mustafa R, Heels-Ansdell D, et al. Effects of higher- versus lower-protein diets on health outcomes: a systematic review and meta-analysis. Eur J Clin Nutr. (2012) 66:780–8. doi: 10.1038/ejcn.2012.37
47. Wycherley TP, Buckley JD, Noakes M, Clifton PM, Brinkworth GD. Comparison of the effects of weight loss from a high-protein versus standard-protein energy-restricted diet on strength and aerobic capacity in overweight and obese men. Eur J Nutr. (2013) 52:317–25. doi: 10.1007/s00394-012-0338-0
48. Norton LE, Wilson GJ. Optimal protein intake to maximize muscle protein synthesis examinations of optimal meal protein intake and frequency for athletes. Agro Food Ind Hi Tech. (2009) 20:54–57.
49. Deer RR, Volpi E. Protein intake and muscle function in older adults. Curr Opin Clin Nutr Metab Care. (2015) 18:248–53. doi: 10.1097/MCO.0000000000000162
50. Moreno-Rojas R, Amaro-Lopez MA, Zurera-Cosano G. Mkronutrients in natural cow, ewe and goat milk. Int J Food Sci Nutr. (1993) 44:37–46. doi: 10.3109/09637489309017421
51. Nowak W, Mikuła R, Zachwieja A, Paczyńska K, Pecka E, Drzazga K, et al. The impact of cow nutrition in the dry period on colostrum quality and immune status of calves. Pol J Vet Sci. (2012) 15:77–82. doi: 10.2478/v10181-011-0117-5
52. Hyrslova I, Krausova G. Goat and bovine colostrum as a basis for new probiotic functional foods and dietary supplements. J Microb Biochem Technol. (2016) 08:56–9. doi: 10.4172/1948-5948.1000262
53. Morin DE, Constable PD, Maunsell FP, McCoy GC. Factors associated with colostral specific gravity in dairy cows. J Dairy Sci. (2001) 84:937–43. doi: 10.3168/jds.S0022-0302(01)74551-1
54. Morrill KM, Conrad E, Lago A, Campbell J, Quigley J, Tyler H. Nationwide evaluation of quality and composition of colostrum on dairy farms in the United States. J Dairy Sci. (2012) 95:3997–4005. doi: 10.3168/jds.2011-5174
55. Muller LD, Ellinger DK. Colostral immunoglobulin concentrations among breeds of dairy cattle. J Dairy Sci. (1981) 64:1727–30. doi: 10.3168/jds.S0022-0302(81)82754-3
56. Maunsell FP, Morin DE, Constable PD, Hurley WL, McCoy GC, Kakoma I, et al. Effects of mastitis on the volume and composition of colostrum produced by holstein cows. J Dairy Sci. (1998) 81:1291–9. doi: 10.3168/jds.S0022-0302(98)75691-7
57. Gulliksen SM, Lie KI, Sølverød L, Østerås O. Risk factors associated with colostrum quality in norwegian dairy cows. J Dairy Sci. (2008) 91:704–12. doi: 10.3168/jds.2007-0450
58. Moore M, Tyler JW, Chigerwe M, Dawes ME, Middleton JR. Effect of delayed colostrum collection on colostral IgG concentration in dairy cows. J Am Vet Med Assoc. (2005) 226:1375–7. doi: 10.2460/javma.2005.226.1375
59. Erdem H, Okuyucu IC. Non-genetic factors affecting some colostrum quality traits in holstein cattle. Pak J Zool. (2020) 52:557–64. doi: 10.17582/journal.pjz/20190219100236
60. West JW. Effects of heat-stress on production in dairy cattle. J Dairy Sci. (2003) 86:2131–44. doi: 10.3168/jds.S0022-0302(03)73803-X
61. Annen EL, Collier RJ, McGuire MA, Vicini JL. Effects of dry period length on milk yield and mammary epithelial cells. J Dairy Sci. (2004) 87:E66–76. doi: 10.3168/jds.S0022-0302(04)70062-4
62. Collier RJ, Annen-Dawson EL, Pezeshki A. Effects of continuous lactation and short dry periods on mammary function and animal health. Animal. (2012) 6:403–14. doi: 10.1017/S1751731111002461
63. Kok A, van Hoeij RJ, Tolkamp BJ, Haskell MJ, van Knegsel ATM, de Boer IJM, et al. Behavioural adaptation to a short or no dry period with associated management in dairy cows. Appl Anim Behav Sci. (2017) 186:7–15. doi: 10.1016/j.applanim.2016.10.017
64. Barrington GM, McFadden TB, Huyler MT, Besser TE. Regulation of colostrogenesis in cattle. Livest Prod Sci. (2001) 70:95–104. doi: 10.1016/S0301-6226(01)00201-9
65. Le Cozler Y, Guatteo R, Le Dréan E, Turban H, Leboeuf F, Pecceu K, et al. IgG1 variations in the colostrum of holstein dairy cows. Animal. (2016) 10:230–7. doi: 10.1017/S1751731115001962
66. Czerniewicz M, Kielczewska K, Kruk A. Comparison of some physicochemical properties of milk from holstein-friesian and jersey cows. Polish J food Nutr Sci. (2006) 15:61.
67. O'Callaghan TF, O'Donovan M, Murphy JP, Sugrue K, Mannion D, McCarthy WP, et al. Evolution of the bovine milk fatty acid profile – from colostrum to milk five days post parturition. Int Dairy J. (2020) 104:104655. doi: 10.1016/j.idairyj.2020.104655
68. Quigley JD, Drewry JJ. Nutrient and immunity transfer from cow to calf pre- and postcalving. J Dairy Sci. (1998) 81:2779–90. doi: 10.3168/jds.S0022-0302(98)75836-9
69. German JB, Gibson RA, Krauss RM, Nestel P, Lamarche B, van Staveren WA, et al. A reappraisal of the impact of dairy foods and milk fat on cardiovascular disease risk. Eur J Nutr. (2009) 48:191–203. doi: 10.1007/s00394-009-0002-5
70. German JB, Argov-Argaman N, Boyd BJ. Milk lipids: a complex nutrient delivery system. In: Donovan SM, German JB, Lönnerdal B, Lucas A, editors. Nestle Nutrition Institute Workshop Series. (2019). p. 217–225.
71. Morrissey PA, Hill TR. Fat-soluble vitamins and vitamin c in milk and milk products. In: McSweeney P, Fox PF, editors. Advanced Dairy Chemistry. New York, NY: Springer New York (2009). p. 527–89.
72. Jensen SK, Johannsen AKB, Hermansen JE. Quantitative secretion and maximal secretion capacity of retinol, β-carotene and α-tocopherol into cows' milk. J Dairy Res. (1999) 66:511–22. doi: 10.1017/S0022029999003805
73. Debier C, Larondelle Y. Vitamins A and E: metabolism, roles and transfer to offspring. Br J Nutr. (2005) 93:153–74. doi: 10.1079/BJN20041308
74. Indyk HE, Woollard DC. The endogenous vitamin K1 content of bovine milk: temporal influence of season and lactation. Food Chem. (1995) 54:403–7. doi: 10.1016/0308-8146(95)00091-V
75. Pereira PC. Milk nutritional composition and its role in human health. Nutrition. (2014) 30:619–27. doi: 10.1016/j.nut.2013.10.011
76. Henry KM, Kon SK. A note on the vitamin D content of cow's colostrum1. Biochem J. (1937) 31:2199–201. doi: 10.1042/bj0312199
77. Marnila P, Korhonen H. Colostrum. In: Roginski H, editor. Encyclopedia of Dairy Sciences. Elsevier (2002). p. 473–8.
78. Haug A, Høstmark AT, Harstad OM. Bovine milk in human nutrition – a review. Lipids Health Dis. (2007) 6:25. doi: 10.1186/1476-511X-6-25
79. Ahmad S, Anjum FM, Huma N, Sameen A, Zahoor T. Composition and physico-chemical characteristics of buffalo milk with particular emphasis on lipids, proteins, minerals, enzymes and vitamins. J Anim Plant Sci. (2013) 23:62–74.
80. Barrington GM, Besser TE, Davis WC, Gay CC, Reeves JJ, McFadden TB. Expression of immunoglobulin G1 receptors by bovine mammary epithelial cells and mammary leukocytes. J Dairy Sci. (1997) 80:86–93. doi: 10.3168/jds.S0022-0302(97)75915-0
81. Korhonen H, Pihlanto A. Technological options for the production of health-promoting proteins and peptides derived from milk and colostrum. Curr Pharm Des. (2007) 13:829–43. doi: 10.2174/138161207780363112
82. Kramski M, Lichtfuss GF, Navis M, Isitman G, Wren L, Rawlin G, et al. Anti-HIV-1 antibody-dependent cellular cytotoxicity mediated by hyperimmune bovine colostrum IgG. Eur J Immunol. (2012) 42:2771–81. doi: 10.1002/eji.201242469
83. Korhonen H, Marnila P, Gill HS. Milk immunoglobulins and complement factors. Br J Nutr. (2000) 84:75–80. doi: 10.1017/S0007114500002282
84. Baumrucker CR, Bruckmaier RM. Colostrogenesis: IgG1 transcytosis mechanisms. J Mammary Gland Biol Neoplasia. (2014) 19:103–17. doi: 10.1007/s10911-013-9313-5
85. Sasaki M, Davis CL, Larson BL. Production and turnover of IgG1 and IgG2 immunoglobulins in the bovine around parturition. J Dairy Sci. (1976) 59:2046–55. doi: 10.3168/jds.S0022-0302(76)84486-4
86. Virtala A-M, Gröhn Y, Mechor G, Erb H. The effect of maternally derived immunoglobulin G on the risk of respiratory disease in heifers during the first 3 months of life. Prev Vet Med. (1999) 39:25–37. doi: 10.1016/S0167-5877(98)00140-8
87. Beam AL, Lombard JE, Kopral CA, Garber LP, Winter AL, Hicks JA, et al. Prevalence of failure of passive transfer of immunity in newborn heifer calves and associated management practices on US dairy operations. J Dairy Sci. (2009) 92:3973–80. doi: 10.3168/jds.2009-2225
88. McGuirk SM, Collins M. Managing the production, storage, and delivery of colostrum. Vet Clin North Am Food Anim Pract. (2004) 20:593–603. doi: 10.1016/j.cvfa.2004.06.005
89. Wakabayashi H, Oda H, Yamauchi K, Abe F. Lactoferrin for prevention of common viral infections. J Infect Chemother. (2014) 20:666–71. doi: 10.1016/j.jiac.2014.08.003
90. Chatterton DEW, Nguyen DN, Bering SB, Sangild PT. Anti-inflammatory mechanisms of bioactive milk proteins in the intestine of newborns. Int J Biochem Cell Biol. (2013) 45:1730–47. doi: 10.1016/j.biocel.2013.04.028
91. Manzoni P. Clinical benefits of lactoferrin for infants and children. J Pediatr. (2016) 173:S43–52. doi: 10.1016/j.jpeds.2016.02.075
92. Dial EJ, Hall LR, Serna H, Romero JJ, Fox JG, Lichtenberger LM. Antibiotic properties of bovine lactoferrin on helicobacter pylori. Dig Dis Sci. (1998) 43:2750–6. doi: 10.1023/A:1026675916421
93. Yamauchi K, Tomita M, Giehl TJ, Ellison RT. Antibacterial activity of lactoferrin and a pepsin-derived lactoferrin peptide fragment. Infect Immun. (1993) 61:719–28. doi: 10.1128/IAI.61.2.719-728.1993
94. Lee J, Kwon SH, Kim HM, Fahey SN, Knighton DR, Sansom A. Effect of a growth protein-colostrum fraction* on bone development in juvenile rats. Biosci Biotechnol Biochem. (2008) 72:1–6. doi: 10.1271/bbb.60695
95. Nakajima K, Kanno Y, Nakamura M, Gao X-D, Kawamura A, Itoh F, et al. Bovine milk lactoferrin induces synthesis of the angiogenic factors VEGF and FGF2 in osteoblasts via the p44/p42 MAP kinase pathway. BioMetals. (2011) 24:847–56. doi: 10.1007/s10534-011-9439-0
96. Siqueiros-Cendón T, Arévalo-Gallegos S, Iglesias-Figueroa BF, García-Montoya IA, Salazar-Martínez J, Rascón-Cruz Q. Immunomodulatory effects of lactoferrin. Acta Pharmacol Sin. (2014) 35:557–66. doi: 10.1038/aps.2013.200
97. Superti F. Lactoferrin from bovine milk: a protective companion for life. Nutrients. (2020) 12:2562. doi: 10.3390/nu12092562
98. Sánchez L, Aranda P, Pérez Md, Calvo M. Concentration of lactoferrin and transferrin throughout lactation in cow's colostrum and milk. Biol Chem Hoppe Seyler. (1988) 369:1005–8. doi: 10.1515/bchm3.1988.369.2.1005
99. Fox PF, Kelly AL. Indigenous enzymes in milk: overview and historical aspects—Part 1. Int Dairy J. (2006) 16:500–16. doi: 10.1016/j.idairyj.2005.09.013
100. Farkye NY, Bansal N. Enzymes indigenous to milk other enzymes. In: Fuquay JW, Fox PJ, McSweeney PLH, editors. Encyclopedia of Dairy Sciences. Amsterdam: Elsevier (2011). p. 327–34.
101. Shakeel-ur-Rehman, Farkye NY. Enzymes indigenous to milk lactoperoxidase. In: Roginski H, editor. Encyclopedia of Dairy Sciences. Elsevier (2002). p. 938–41.
102. Wolfson LM, SUMNER SS. Antibacterial activity of the lactoperoxidase system: a review. J Food Prot. (1993) 56:887–92. doi: 10.4315/0362-028X-56.10.887
103. Belding ME, Klebanoff SJ, Ray CG. Peroxidase-mediated virucidal systems. Science (80-). (1970) 167:195–6. doi: 10.1126/science.167.3915.195
104. Tanaka T, Xuan X, Fujisaki K, Shimazaki K. Expression and characterization of bovine milk antimicrobial proteins lactoperoxidase and lactoferrin by vaccinia virus. In: Roy PK, editors. Insight and Control of Infectious Disease in Global Scenario. IntechOpen (2012) p. 127–33.
105. Nakamura T, Kawase H, Kimura K, Watanabe Y, Ohtani M, Arai I, et al. Concentrations of sialyloligosaccharides in bovine colostrum and milk during the prepartum and early lactation. J Dairy Sci. (2003) 86:1315–20. doi: 10.3168/jds.S0022-0302(03)73715-1
106. Tao N, DePeters EJ, Freeman S, German JB, Grimm R, Lebrilla CB. Bovine milk glycome. J Dairy Sci. (2008) 91:3768–78. doi: 10.3168/jds.2008-1305
107. Tao N, DePeters EJ, German JB, Grimm R, Lebrilla CB. Variations in bovine milk oligosaccharides during early and middle lactation stages analyzed by high-performance liquid chromatography-chip/mass spectrometry. J Dairy Sci. (2009) 92:2991–3001. doi: 10.3168/jds.2008-1642
108. Barile D, Marotta M, Chu C, Mehra R, Grimm R, Lebrilla CB, et al. Neutral and acidic oligosaccharides in Holstein-Friesian colostrum during the first 3 days of lactation measured by high performance liquid chromatography on a microfluidic chip and time-of-flight mass spectrometry. J Dairy Sci. (2010) 93:3940–9. doi: 10.3168/jds.2010-3156
109. Ninonuevo MR, Park Y, Yin H, Zhang J, Ward RE, Clowers BH, et al. A strategy for annotating the human milk glycome. J Agric Food Chem. (2006) 54:7471–80. doi: 10.1021/jf0615810
110. Martín-Sosa S, Martín M-J, García-Pardo L-A, Hueso P. Sialyloligosaccharides in human and bovine milk and in infant formulas: variations with the progression of lactation. J Dairy Sci. (2003) 86:52–59. doi: 10.3168/jds.S0022-0302(03)73583-8
111. Urashima T, Kitaoka M, Asakuma S, Messer M. Milk oligosaccharides. In: McSweeney P, Fox PF, esitors. Advanced Dairy Chemistry. New York, NY: Springer New York (2009). p. 295–349.
112. McJarrow P, van Amelsfort-Schoonbeek J. Bovine sialyl oligosaccharides: seasonal variations in their concentrations in milk, and a comparison of the colostrums of jersey and friesian cows. Int Dairy J. (2004) 14:571–9. doi: 10.1016/j.idairyj.2003.11.006
113. Karav S, Bell J, Parc Le A, Liu Y, Mills DA, Block DE, et al. Characterizing the release of bioactive N-glycans from dairy products by a novel endo-β-N-acetylglucosaminidase. Biotechnol Prog. (2015) 31:1331–9. doi: 10.1002/btpr.2135
114. Zivkovic AM, Barile D. Bovine milk as a source of functional oligosaccharides for improving human health. Adv Nutr. (2011) 2:284–9. doi: 10.3945/an.111.000455
115. Weinborn V, Li Y, Shah IM, Yu H, Dallas DC, German JB, et al. Production of functional mimics of human milk oligosaccharides by enzymatic glycosylation of bovine milk oligosaccharides. Int Dairy J. (2020) 102:104583. doi: 10.1016/j.idairyj.2019.104583
116. Robinson RC, Colet E, Tian T, Poulsen NA, Barile D. An improved method for the purification of milk oligosaccharides by graphitised carbon-solid phase extraction. Int Dairy J. (2018) 80:62–68. doi: 10.1016/j.idairyj.2017.12.009
117. Westreich ST, Salcedo J, Durbin-Johnson B, Smilowitz JT, Korf I, Mills DA, et al. Fecal metatranscriptomics and glycomics suggest that bovine milk oligosaccharides are fully utilized by healthy adults. J Nutr Biochem. (2020) 79:108340. doi: 10.1016/j.jnutbio.2020.108340
118. Jakobsen LMA, Sundekilde UK, Andersen HJ, Nielsen DS, Bertram HC. Lactose and bovine milk oligosaccharides synergistically stimulate B. Longum subsp. Longum growth in a simplified model of the infant gut microbiome. J Proteome Res. (2019) 18:3086–98. doi: 10.1021/acs.jproteome.9b00211
119. Jakobsen LMA, Maldonado-Gómez MX, Sundekilde UK, Andersen HJ, Nielsen DS, Bertram HC. Metabolic effects of bovine milk oligosaccharides on selected commensals of the infant microbiome-commensalism and postbiotic effects. Metabolites. (2020) 10:167. doi: 10.3390/metabo10040167
120. Karav S, Le Parc A, Leite Nobrega de Moura Bell JM, Frese SA, Kirmiz N, Block DE, et al. Oligosaccharides released from milk glycoproteins are selective growth substrates for infant-associated bifidobacteria. Appl Environ Microbiol. (2016) 82:3622–30. doi: 10.1128/AEM.00547-16
121. Bunyatratchata A, Huang YP, Ozturk G, Cohen JL, Bhattacharya M, Mln De Moura Bell J, et al. Effects of industrial thermal treatments on the release of bovine colostrum glycoprotein n-glycans by endo-β- N-acetylglucosaminidase. J Agric Food Chem. (2020) 68:15208–15. doi: 10.1021/acs.jafc.0c05986
122. Cao X, Yang M, Yang N, Liang X, Tao D, Liu B, et al. Characterization and comparison of whey N-glycoproteomes from human and bovine colostrum and mature milk. Food Chem. (2019) 276:266–73. doi: 10.1016/j.foodchem.2018.09.174
123. Henrick BM, Chew S, Casaburi G, Brown HK, Frese SA, Zhou Y, et al. Colonization by B. Infantis EVC001 modulates enteric inflammation in exclusively breastfed infants. Pediatr Res. (2019) 86:749–57. doi: 10.1038/s41390-019-0533-2
124. Duar RM, Casaburi G, Mitchell RD, Scofield LNC, Ramirez CAO, Barile D, et al. Comparative genome analysis of bifidobacterium among commercial probiotics. Nutrients. (2020) 12:3247. doi: 10.3390/nu12113247
125. Lane JA, Mariño K, Naughton J, Kavanaugh D, Clyne M, Carrington SD, et al. Anti-infective bovine colostrum oligosaccharides: campylobacter jejuni as a case study. Int J Food Microbiol. (2012) 157:182–8. doi: 10.1016/j.ijfoodmicro.2012.04.027
126. Shing CM, Hunter DC, Stevenson LM. Bovine colostrum supplementation and exercise performance: potential mechanisms. Sport Med. (2009) 39:1033–54. doi: 10.2165/11317860-000000000-00000
127. Antonio J, Sanders MS, Van Gammeren D. The effects of bovine colostrum supplementation on body composition and exercise performance in active men and women. Nutrition. (2001) 17:243–7. doi: 10.1016/S0899-9007(00)00552-9
128. Hofman Z, Smeets R, Verlaan G, Lugt RVD, Verstappen PA. The effect of bovine colostrum supplementation on exercise performance in elite field hockey players. Int J Sport Nutr Exerc Metab. (2002) 12:461–9. doi: 10.1123/ijsnem.12.4.461
129. Coombes JS, Conacher M, Austen SK, Marshall PA. Dose effects of oral bovine colostrum on physical work capacity in cyclists. Med Sci Sport Exerc. (2002) 34:1184–8. doi: 10.1097/00005768-200207000-00020
130. Buckley JD, Abbott MJ, Brinkworth GD, Whyte PBD. Bovine colostrum supplementation during endurance running training improves recovery, but not performance. J Sci Med Sport. (2002) 5:65–79. doi: 10.1016/S1440-2440(02)80028-7
131. Duff WRD, Chilibeck PD, Rooke JJ, Kaviani M, Krentz JR, Haines DM. The effect of bovine colostrum supplementation in older adults during resistance training. Int J Sport Nutr Exerc Metab. (2014) 24:276–85. doi: 10.1123/ijsnem.2013-0182
132. Crooks CV, Wall CR, Cross ML, Rutherfurd-Markwick KJ. The effect of bovine colostrum supplementation on salivary IgA in distance runners. Int J Sport Nutr Exerc Metab. (2006) 16:47–64. doi: 10.1123/ijsnem.16.1.47
133. Główka N, Durkalec-Michalski K, Wozniewicz M. Immunological outcomes of bovine colostrum supplementation in trained and physically active people: a systematic review and meta-analysis. Nutrients. (2020) 12:1023. doi: 10.3390/nu12041023
134. Mero A, Kähkönen J, Nykänen T, Parviainen T, Jokinen I, Takala T, et al. IGF-I, IgA, and IgG responses to bovine colostrum supplementation during training. J Appl Physiol. (2002) 93:732–9. doi: 10.1152/japplphysiol.00002.2002
135. Shing CM, Peake J, Suzuki K, Okutsu M, Pereira R, Stevenson L, et al. Effects of bovine colostrum supplementation on immune variables in highly trained cyclists. J Appl Physiol. (2007) 102:1113–22. doi: 10.1152/japplphysiol.00553.2006
136. Crooks C, Cross ML, Wall C, Ali A. Effect of bovine colostrum supplementation on respiratory tract mucosal defenses in swimmers. Int J Sport Nutr Exerc Metab. (2010) 20:224–35. doi: 10.1123/ijsnem.20.3.224
137. Davison G, Diment BC. Bovine colostrum supplementation attenuates the decrease of salivary lysozyme and enhances the recovery of neutrophil function after prolonged exercise. Br J Nutr. (2010) 103:1425–32. doi: 10.1017/S0007114509993503
138. Jones AW, Cameron SJS, Thatcher R, Beecroft MS, Mur LAJ, Davison G. Effects of bovine colostrum supplementation on upper respiratory illness in active males. Brain Behav Immun. (2014) 39:194–203. doi: 10.1016/j.bbi.2013.10.032
139. Jones AW, Thatcher R, March DS, Davison G. Influence of 4 weeks of bovine colostrum supplementation on neutrophil and mucosal immune responses to prolonged cycling. Scand J Med Sci Sports. (2015) 25:788–96. doi: 10.1111/sms.12433
140. Shing CM, Peake JM, Suzuki K, Jenkins DG, Coombes JS. A pilot study: bovine colostrum supplementation and hormonal and autonomic responses to competitive cycling. J Sports Med Phys Fitness. (2013) 53:490–501.
141. Marchbank T, Davison G, Oakes JR, Ghatei MA, Patterson M, Moyer MP, et al. The nutriceutical bovine colostrum truncates the increase in gut permeability caused by heavy exercise in athletes. Am J Physiol Liver Physiol. (2011) 300:G477–84. doi: 10.1152/ajpgi.00281.2010
142. Allison MC, Howatson AG, Torrance CJ, Lee FD, Russell RI. Gastrointestinal damage associated with the use of nonsteroidal antiinflammatory drugs. N Engl J Med. (1992) 327:749–54. doi: 10.1056/NEJM199209103271101
143. Henry D, McGettigan P. Epidemiology overview of gastrointestinal and renal toxicity of NSAIDs. Int J Clin Pract Suppl. (2003) 43–49.
144. Sigthorsson G, Tibble J, Hayllar J, Menzies I, Macpherson A, Moots R, et al. Intestinal permeability and inflammation in patients on NSAIDs. Gut. (1998) 143:506–11. doi: 10.1136/gut.43.4.506
145. Playford RJ, Macdonald CE, Johnson WS. Colostrum and milk-derived peptide growth factors for the treatment of gastrointestinal disorders. Am J Clin Nutr. (2000) 72:5–14. doi: 10.1093/ajcn/72.1.5
146. Playford RJ, Macdonald CE, Calnan DP, Floyd DN, Podas T, Johnson W, et al. Co-administration of the health food supplement, bovine colostrum, reduces the acute non-steroidal anti-inflammatory drug-induced increase in intestinal permeability. Clin Sci. (2001) 100:627. doi: 10.1042/CS20010015
147. Mir R, Singh N, Vikram G, Kumar RP, Sinha M, Bhushan A, et al. The structural basis for the prevention of nonsteroidal antiinflammatory drug-induced gastrointestinal tract damage by the C-Lobe of bovine colostrum lactoferrin. Biophys J. (2009) 97:3178–86. doi: 10.1016/j.bpj.2009.09.030
148. Klotz U. The role of aminosalicylates at the beginning of the new millennium in the treatment of chronic inflammatory bowel disease. Eur J Clin Pharmacol. (2000) 56:353–62. doi: 10.1007/s002280000163
149. Khan Z, Macdonald C, Wicks AC, Holt MP, Floyd D, Ghosh S, et al. Use of the ‘nutriceutical', bovine colostrum, for the treatment of distal colitis: results from an initial study. Aliment Pharmacol Ther. (2002) 16:1917–22. doi: 10.1046/j.1365-2036.2002.01354.x
150. Niño DF, Sodhi CP, Hackam DJ. Necrotizing enterocolitis: new insights into pathogenesis and mechanisms. Nat Rev Gastroenterol Hepatol. (2016) 13:590–600. doi: 10.1038/nrgastro.2016.119
151. Balachandran B, Dutta S, Singh R, Prasad R, Kumar P. Bovine colostrum in prevention of necrotizing enterocolitis and sepsis in very low birth weight neonates: a randomized, double-blind, placebo-controlled pilot trial. J Trop Pediatr. (2017) 63:10–17. doi: 10.1093/tropej/fmw029
152. Sadeghirad B, Morgan RL, Zeraatkar D, Zea AM, Couban R, Johnston BC, et al. Human and bovine colostrum for prevention of necrotizing enterocolitis: a meta-analysis. Pediatrics. (2018) 142:e20180767. doi: 10.1542/peds.2018-0767
153. Snyder R, Herdt A, Mejias-Cepeda N, Ladino J, Crowley K, Levy P. Early provision of oropharyngeal colostrum leads to sustained breast milk feedings in preterm infants. Pediatr Neonatol. (2017) 58:534–40. doi: 10.1016/j.pedneo.2017.04.003
154. De La Cabada Bauche J, DuPont HL. New developments in traveler's diarrhea. Gastroenterol Hepatol. (2011) 7:88–95.
155. Kolenda R, Burdukiewicz M, Schierack P. A systematic review and meta-analysis of the epidemiology of pathogenic escherichia coli of calves and the role of calves as reservoirs for human pathogenic E. Coli. Front Cell Infect Microbiol. (2015) 5:23. doi: 10.3389/fcimb.2015.00023
156. Otto W, Najnigier B, Stelmasiak T, Robins-Browne RM. Randomized control trials using a tablet formulation of hyperimmune bovine colostrum to prevent diarrhea caused by enterotoxigenic escherichia coli in volunteers. Scand J Gastroenterol. (2011) 46:862–8. doi: 10.3109/00365521.2011.574726
157. Mitra A, Mahalanabis D, Ashraf H, Unicomb L, Eeckels R, Tzipori S. Hyperimmune cow colostrum reduces diarrhoea due to rotavirus: a double-blind, controlled clinical trial. Acta Pædiatrica. (1995) 84:996–1001. doi: 10.1111/j.1651-2227.1995.tb13814.x
158. Sarker SA, Casswall TH, Mahalanabis D, Alam NH, Albert MJ, Brüssow H, et al. Successful treatment of rotavirus diarrhea in children with immunoglobulin from immunized bovine colostrum. Pediatr Infect Dis J. (1998) 17:1149–54. doi: 10.1097/00006454-199812000-00010
159. Schroeder GN, Hilbi H. Molecular pathogenesis of shigella spp.: controlling host cell signaling, invasion, and death by type III secretion. Clin Microbiol Rev. (2008) 21:134–56. doi: 10.1128/CMR.00032-07
160. Mirhoseini A, Amani J, Nazarian S. Review on pathogenicity mechanism of enterotoxigenic escherichia coli and vaccines against it. Microb Pathog. (2018) 117:162–9. doi: 10.1016/j.micpath.2018.02.032
161. Kachrimanidou M, Malisiovas N. Clostridium difficile infection: a comprehensive review. Crit Rev Microbiol. (2011) 37:178–87. doi: 10.3109/1040841X.2011.556598
Keywords: bovine colostrum, human health, bioactive proteins, oligosaccharides, infants
Citation: Arslan A, Kaplan M, Duman H, Bayraktar A, Ertürk M, Henrick BM, Frese SA and Karav S (2021) Bovine Colostrum and Its Potential for Human Health and Nutrition. Front. Nutr. 8:651721. doi: 10.3389/fnut.2021.651721
Received: 10 January 2021; Accepted: 27 May 2021;
Published: 21 June 2021.
Edited by:
Nicole Clemence Roy, University of Otago, New ZealandReviewed by:
Isabelle Le Huërou-Luron, INRA Centre Bretagne-Normandie, FranceCopyright © 2021 Arslan, Kaplan, Duman, Bayraktar, Ertürk, Henrick, Frese and Karav. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sercan Karav, c2VyY2Fua2FyYXZAY29tdS5lZHUudHI=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.