 Veronica D'Antonio
Veronica D'Antonio Mauro Serafini
Mauro Serafini Natalia Battista
Natalia Battista
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Nutr., 26 February 2021
Sec. Nutritional Epidemiology
Volume 8 - 2021 | https://doi.org/10.3389/fnut.2021.642551
This article is part of the Research TopicEdible Insects: From Farm to ForkView all 6 articles
Edible insects are proposed as a nutritious and environmentally sustainable alternative source to animal proteins, due to their numerous advantages in terms of reduced ecological impact and high nutritional value. However, the novelty for edible insects relies on the content of bioactive ingredients potentially able to induce a functional effect in the body. The present review summarizes the main findings on the antioxidant properties of edible insects available in the literature. A total of 30 studies involving animals, cell cultures, or in vitro experimental studies evaluating the antioxidant effect of edible insects are presented in this work. When the antioxidant activity was investigated, using a wide variety of in vitro tests and in cellular models, positive results were shown. Dietary supplementation with edible insects was also able to counteract dietary oxidative stress in animal models, restoring the balance of antioxidant enzymes and reducing the formation of oxidation damage markers. On the basis of the reviewed evidences, edible insects might represent a source of novel redox ingredients at low ecological impact able to modulate oxidative stress. However, due to the fact that majority of these evidences have been obtained in vitro and in cellular and animal models, dietary intervention trials are needed to assess the efficacy of edible insect consumption to modulate redox status in humans.
Entomophagy, the practice of eating insects and invertebrates, has been part of human history for centuries, playing a significant role in cultural and religious practices. Recently, a new global interest in edible insects and invertebrates arises from the impellent necessity of preserving agriculture resources to feed the 9 billion world's population predicted for 2050 and to obtain a drastic reduction of the ecological impact of food production, accounting for between 20 and 30% of the total environmental impact (1). In terms of ecological impact, edible insects are characterized by a negligible greenhouse gas (GHG) emission as well as water and ecological footprints, meeting the population's need for a more sustainable protein supply. In this view, Onnincx et al. (2) have described lower NH3 emission level, higher average daily gain, and a comparable or lower CO2 (g/kg mass gain) production of insects, which result in a higher feed conversion efficiency, with respect to conventional livestock. Moreover, although the fossil energy needed to mealworms rearing is comparable to or higher than conventional food sources such as milk or different meats, these insects produce reduced GHG—one of the main factors inducing climate changes—and the space required for their rearing is much lower than conventional livestock (3).
Edible insects are characterized by a high nutritional value, and they are good potential sources of proteins, amino acids, minerals, and lipids (4). The protein content of the various species of insects is generally very high (50–70% on dry basis), while lipids represent the second largest fraction of the nutritional composition, ranging from 10 to 50% on dry basis and depending on life stage (higher in the larval stage) and species. Interestingly, it has been reported that they have a higher value of the essential fatty acids linoleic (18:2 n-6) and α-linolenic (18:3 n-3) when compared to other conventional sources (4). However, the innovative feature of edible insects relies on the content of bioactive ingredients and on their ability to induce a functional effect in the body and potentially able to provide a protective effect against diseases, entomotherapy, as previously suggested (5). In the last years, scientific evidences on the functional properties of edible insects have been provided in different experimental models, with majority of the studies focused on the understanding of the antioxidant role of edible insects and their extracts (6, 7). In order to understand if edible insects might play a role in the dietary modulation of oxidative stress, in this mini-review, we aim to summarize the available evidences regarding the antioxidant role of edible insects and invertebrates in in vitro, ex vivo, and in vivo models.
A search for literature investigating the antioxidant effect of edible insects was carried out by PubMed database (National Library of Medicine, Bethesda, MD) using the following keywords: “edible insect,” “oxidative,” and “antioxidant.” Eligible studies for this mini-review have included randomized controlled trials in humans, experimental animals, or cell cultures or in vitro experimental studies published in English, leading to the selection of 30 studies. No studies on human subjects were available.
Table 1 (6–24) shows the studies investigating the in vitro antioxidant activity of edible insects and invertebrate fractions. Several methods were taken into account: the most used was the 1,1-diphenyl-2-picrylhydrazyl (DPPH), performed in 14 papers (7–15, 18–20, 23, 24), while 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) and ferric reducing antioxidant power (FRAP) were utilized in 11 (6, 7, 10, 11, 13, 14, 16, 17, 19, 20, 22) and 8 studies (6, 10–12, 14, 15, 19, 21), respectively. Antioxidant activity was investigated also as scavenging activity against different radicals, metal ion chelating activity, reducing power, or, only for the paper of Sun et al. (25), with β-carotene and linolenic acid bleaching tests. Nineteen studies were carried out, involving 30 species of insects; of these, the most studied was Tenebrio molitor, cited in nine papers (6–10, 13–16), followed by Acheta domesticus (6, 8, 9) and Gryllodes sigillatus (7, 19, 20), tested in three papers, and Bombyx mori (6, 11), Hermetia illucens (21, 22), and Lethocerus indicus (6, 24) in two papers. Water-soluble fractions were tested in 12 papers (6, 9–15, 17, 18, 20, 24), protein hydrolysates were evaluated in 6 research articles (7, 8, 16, 19, 21, 22), while lipo-soluble fractions were reported in 2 papers only (6, 23). Interestingly, all tested fractions showed a significant antioxidant activity with the only exception of the cricket Gryllus sigillatus protein hydrolysates that did not show any positive results using the FRAP method, while an antioxidant capacity was recorded using ABTS, DPPH, and metal ion chelating activity methods (19). The study conducted by Di Mattia et al. (6) was the only one providing a comparison between the antioxidant activity of water- and lipo-soluble fraction edible insects and foods like fresh orange juice and olive oil. Results showed that water-soluble extracts of grasshoppers, silkworm, and crickets display the highest values of antioxidant capacity, expressed as Trolox Equivalent Antioxidant Capacity (TEAC), 5-fold higher than fresh orange juice. Furthermore, water-soluble extracts of grasshoppers, African caterpillars, and crickets had a reducing power (FRAP) double than that of fresh orange juice. As regards the lipo-soluble fraction, silkworm, evening cicada, and African caterpillars showed a TEAC twice than that of olive oil. Differently from other studies, Dutta et al. (18) showed that aqueous extract of Vespa affinis was able to increase the activity of the endogenous antioxidant enzymes catalase (CAT) and glutathione S-transferase (GST) (18).
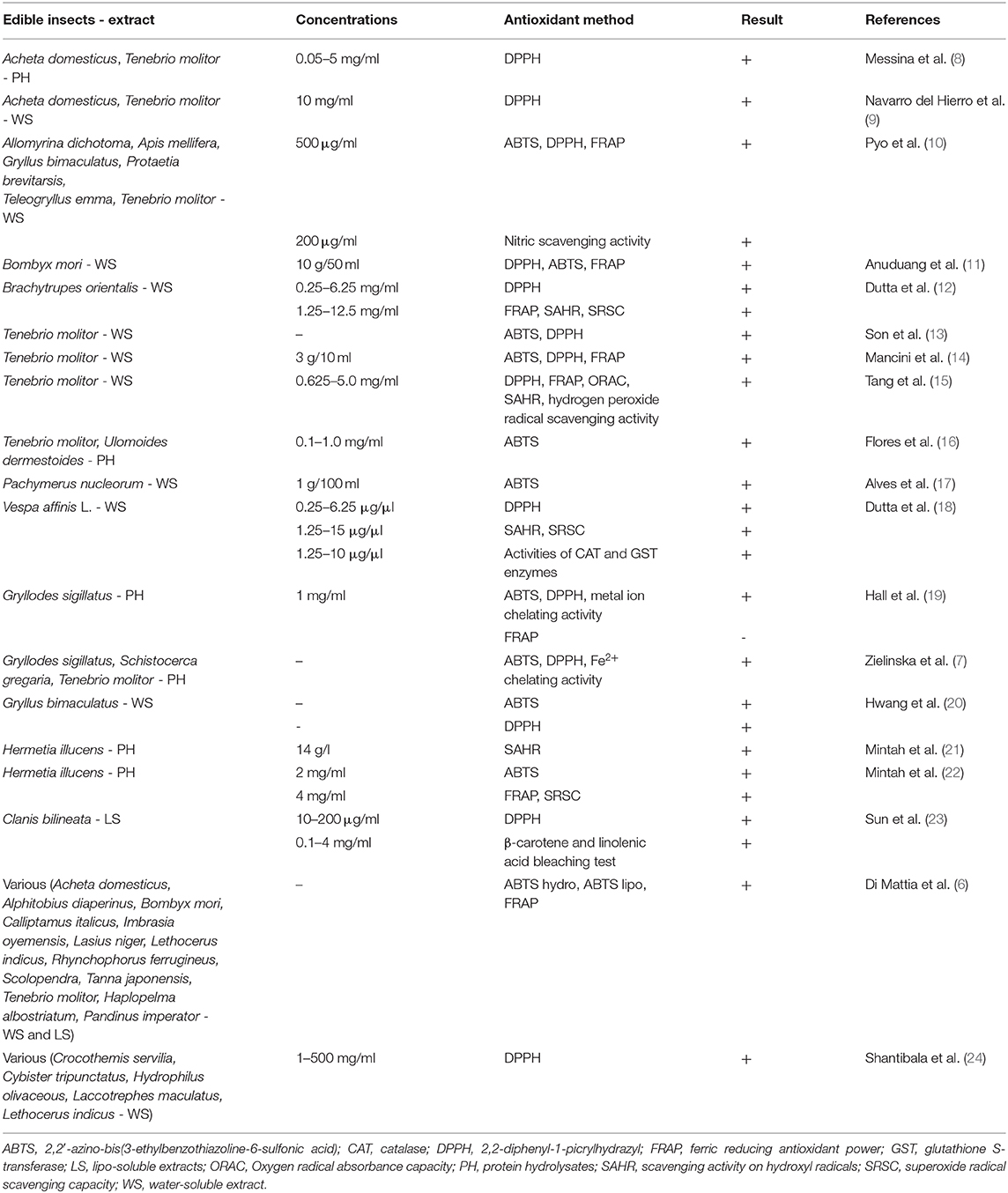
Table 1. In vitro antioxidant activity of edible insects and invertebrates.
Table 2 summarizes the results obtained on the antioxidant activity of different insects and invertebrates; eight were tested in cellular models (12, 13, 18, 20, 26–29) and as many in animal models (20, 27, 30–35). For what concerns cellular models, the effects of the water-soluble extract of dung beetles of Onitis sp., mole crickets of Gryllotalpa sp., grasshopper of Caelifera sp. (28), Oryctes boas, and Zonocerus variegatus (29) on the oxidative status of human peripheral blood lymphocytes were evaluated, showing that, at lower concentrations of all of their WEs, they display antioxidant activities, but at higher concentrations, their effects switched to prooxidant. Interestingly, intermediate concentrations did not affect the antioxidant efficiency. According to the results obtained in a cell-free system (18), the aqueous extract of V. affinis was able to increase the activity of both GST and CAT and to reduce reactive oxygen species (ROS) also in THP-1 human monocytes and human plasma. Oxidative stress induced by high glucose treatment caused a significant decrease of both Nrf2 and GST protein expression in C2C12, a murine myotubes cell line, and that the supplementation with the hydro-alcoholic extract of Brachytrupes orientalis re-established the normal levels of both proteins and prevented the high glucose-induced oxidative impairment in terms of ROS and malondialdehyde (MDA) levels (12). Three different studies have underlined the capacity of aqueous extracts of Gryllus bimaculatus (20), the methanolic extract of defatted powder and usaponifiable lipids, obtained by T. molitor (13), and B. mori protein hydrolysates to reduce nitric oxide (NO) production in lipopolysaccharide-induced RAW 264.7, a murine macrophage cell line. NO production was reduced also in D-HMVECs, i.e., diabetic type 2 microvascular endothelial cells, by glycosaminoglycan from G. bimaculatus (27). In agreement with the paper of Yoon and coworkers (26), protein hydrolysates of T. molitor and G. bimaculatus did not exert any effect on NO release.
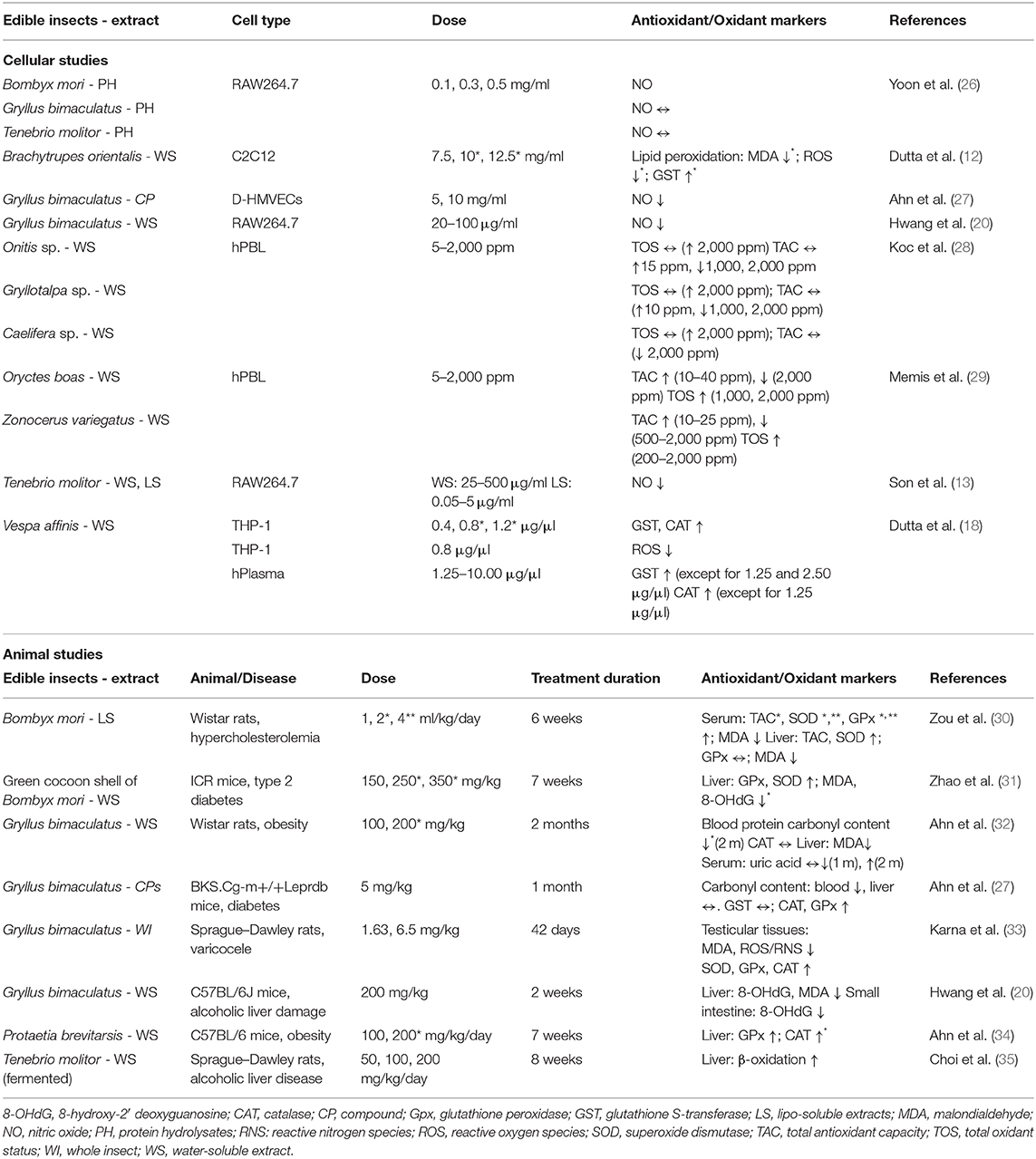
Table 2. Antioxidant activity of edible insects and invertebrates in cellular and animal models.
A total of eight intervention studies on animal models characterized by hypercholesterolemia, diabetes, obesity, and alcoholic liver damage have been published. In more detail, Zou et al. (30) reported that in Wistar rats with hypercholesterolemia, B. mori pupae oil supplementation was able to restore superoxide dismutase (SOD) levels, increasing total antioxidant capacity (TAC) levels, reducing MDA in liver and serum, and restoring the activity of glutathione peroxidase (GPx) in rats' liver, stressed by a high-cholesterol diet. Moreover, the supplementation with ethanol extract of the sericin layer from the green cocoon shell of B. mori increased liver GPx and SOD in obese mice with type 2 diabetes, and the treatment also reduced the liver content of MDA and 8-hydroxy-2′-deoxyguanosine (8-OHdG), as markers of lipid and DNA oxidative damage (31). Ethanolic extract of G. bimaculatus, added to a high-fat diet, did not affect serum CAT in obese rats (32). However, the prolonged treatment with ethanolic extract of G. bimaculatus repaired the protein and lipid oxidative damage caused by high-fat diet in both liver and blood, where serum uric acid—the final oxidation product of purine metabolism—and carbonyl—a marker of protein oxidation—concentrations were reduced (32). In a study carried out in 2019 by Hwang et al. (20), they reported the positive action of the aqueous extract of G. bimaculatus in restoring the normal physiological levels of 8-OHdG levels and MDA content in liver and small intestine of C57BL/6J mice liver damage caused by acute alcohol exposure. Moreover, treatment with G. bimaculatus significantly restored the increased levels of MDA, ROS, and reactive nitrogen species (RNS) and the reduced levels of SOD, GPx, and CAT in testicular tissue of Sprague–Dawley rats affected by varicocele (33). Glycosaminoglycan extracted from G. bimaculatus and administered for 1 month to BKS.Cg-m+/+Leprdb diabetic mice reduced blood carbonyl content, but not that of liver. It also did not affect GST, but improved CAT and GPx levels (27). The ethanolic extracts of Protaetia brevitaris larvae, administered with a high-fat diet for 7 weeks, increased GPx and CAT in liver of obese C57BL/6J mice (34). Finally, the treatment fermented defatted T. molitor powder of Sprague–Dawley rats fed with a chronic alcohol diet dose-dependently increased hepatic β-oxidation (35).
In this review, we showed that different species of edible insects display an antioxidant activity in in vitro models and are able to modulate induced oxidative stress in cellular and animal models. All the insects tested in vitro and in different cellular models, except one, display radical scavenging or metal ion chelation properties, as well as modulation of antioxidant enzymes. Results in animal models have clearly shown, in all the studies, that the increased content of markers of oxidative damage markers, induced by the dietary stress, was reversed following the treatment with edible insects, restoring the impaired activity of antioxidant enzymes, and by reducing oxidation products. Redox status was evaluated through the ability of the insect extracts to reduce ROS (three studies) or to increase total antioxidant status (two studies) in cellular models. Moreover, specific oxidation markers were chosen: the urinary excretion of 8-OHdG, a predictive risk factor for cancer, atherosclerosis, and diabetes (36), and MDA that plays a critical role in atherosclerosis by its capacity to drive inflammatory processes (37). These oxidation products were studied respectively in four in vivo interventions, one in cells for MDA and in two in vivo interventions for 8-OhdG, also in those cases with positive results. One intervention (32) has also evaluated the content of serum uric acid—the final oxidation product of purine metabolism—and two interventions (27, 32) have evaluated that of carbonyl—a marker of protein oxidation. In this context, we should recall that protein carbonyl levels are elevated in several pathological conditions, including neurodegenerative diseases, obesity, or diabetes (38); on the other hand, serum uric acid levels can be a marker of renal and cardiovascular risk, in particular as a consequence of diabetes (39). The antioxidant enzymes that were taken into account in these studies were CAT, which was studied in six different interventions (four in vivo, one in a cellular model, and one in a cell-free system); GPx, which was investigated in five in vivo studies; SOD, which was analyzed in three in vivo studies; and GSTs, which were evaluated in one in vivo intervention, two interventions in cellular models, and one in vitro. Moreover, Nrf-2—a transcription factor that acts as a master regulator of the antioxidant response system and whose activity declines with age as well as with degenerative disorders (38)—was reported to increase only in one study carried out in a myotube cell line. Conflicting results arose from human studies that evaluated the relationship between diseases or aging and antioxidant enzymes: indeed, an increase in antioxidant enzymes can also be related to a high response to oxidation (38). However, in all the examined studies, the oxidative stress induced a reduction of these enzyme levels that was prevented by insect supplementation. Based on these evidences, it can be stated that, in the applied conditions, edible insects exert mostly a positive effect on the modulation of antioxidant enzymes.
On the basis of the positive results of the three studies focused on NO production in macrophages, radical involved in the modulation of inflammation and immune regulation (40), it might be speculated that edible insect extracts might also have a potential anti-inflammatory activity due to their ability to reduce the release of NO.
As regards the insects, 35 different species were investigated; the ones that arouse major interest in researchers were T. molitor and those belonging to the Gryllydae family, respectively studied in 13 and 10 different researches. However, it is interesting to note that the experimental studies carried out in vivo are mainly focused on G. bimaculatus, P. brevitaris, and B. mori, while curiously, all the studies performed using T. molitor are in vitro, with only one exception (35).
On the basis of the reviewed evidences, edible insects might represent a source of novel redox ingredients at low ecological impact able to modulate oxidative stress. However, due to the fact that majority of these evidences have been obtained in vitro and in animal models, dietary intervention trials are needed to confirm the antioxidant efficacy of edible insects in humans.
MS conceived the review topics. VD'A wrote the initial draft. MS and NB revised and supervised the entire work. All authors approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
8-OHdG, 8-hydroxy-2′-deoxyguanosine; ABTS, 2, 2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid); CAT, catalase; DPPH, 1, 1-diphenyl-2-picrylhydrazyl; CP, compound; FRAP, ferric reducing antioxidant power; GHG, greenhouse gas emissions; GPx, glutathione peroxidase; GST, glutathione S-transferase; LS, lipo-soluble extract; MDA, malondialdehyde; NO, nitric oxide; Nrf2, Nuclear factor erythroid 2-related factor; ORAC, oxygen radical absorbance capacity; PH, protein hydrolysates; RNS, reactive nitrogen species; ROS, reactive oxygen species; SAHR, scavenging activity on hydroxyl radicals; SOD, superoxide dismutase; SRSC, superoxide radical scavenging capacity; TAC, total antioxidant capacity; TEAC, Trolox equivalent antioxidant capacity; TOS, total oxidant status; WI, whole insect; WS, water-soluble extract.
1. Tukker A, Jansen B. Environmental impacts of products: a detailed review of studies. J Ind Ecol. (2006) 10:159–82. doi: 10.1162/jiec.2006.10.3.159
2. Oonincx DGAB, van Itterbeeck J, Heetkamp MJW, van den Brand H, van Loon JJA, van Huis A. An exploration on greenhouse gas and ammonia production by insect species suitable for animal or human consumption. PLoS ONE. (2010) 5:1–7. doi: 10.1371/journal.pone.0014445
3. Oonincx DGAB, de Boer IJM. Environmental impact of the production of mealworms as a protein source for humans - a life cycle assessment. PLoS ONE. (2012) 7:e0051145. doi: 10.1371/journal.pone.0051145
4. Jantzen da Silva Lucas A, Menegon de Oliveira L, da Rocha M, Prentice C. Edible insects: an alternative of nutritional, functional and bioactive compounds. Food Chem. (2020) 311:126022. doi: 10.1016/j.foodchem.2019.126022
5. Costa-Neto EM. Entomotherapy, or the medicinal use of insects. J Ethnobiol. (2005) 25:93–114. doi: 10.2993/0278-0771(2005)25[93:EOTMUO]2.0.CO;2
6. Di Mattia C, Battista N, Sacchetti G, Serafini M. Antioxidant activities in vitro of water and liposoluble extracts obtained by different species of edible insects and invertebrates. Front Nutr. (2019) 6:106. doi: 10.3389/fnut.2019.00106
7. Zielińska E, Baraniak B, Karaś M. Antioxidant and anti-inflammatory activities of hydrolysates and peptide fractions obtained by enzymatic hydrolysis of selected heat-treated edible insects. Nutrients. (2017) 9:1–14. doi: 10.3390/nu9090970
8. Messina CM, Gaglio R, Morghese M, Tolone M, Arena R, Moschetti G, et al. Microbiological profile and bioactive properties of insect powders used in food and feed formulations. Foods. (2019) 8:1–16. doi: 10.3390/foods8090400
9. Navarro del Hierro J, Gutiérrez-Docio A, Otero P, Reglero G, Martin D. Characterization, antioxidant activity, and inhibitory effect on pancreatic lipase of extracts from the edible insects Acheta domesticus and Tenebrio molitor. Food Chem. (2020) 309:125742. doi: 10.1016/j.foodchem.2019.125742
10. Pyo SJ, Kang DG, Jung C, Sohn HY. Anti-thrombotic, anti-oxidant and haemolysis activities of six edible insect species. Foods. (2020) 9:1–13. doi: 10.3390/foods9040401
11. Anuduang A, Loo YY, Jomduang S, Lim SJ, Mustapha WAW. Effect of thermal processing on physico-chemical and antioxidant properties in mulberry silkworm (Bombyx mori L.) Powder. Foods. (2020) 9:1–11. doi: 10.3390/foods9070871
12. Dutta P, Dey T, Dihingia A, Manna P, Kalita J. Antioxidant and glucose metabolizing potential of edible insect, Brachytrupes orientalis via modulating Nrf2/AMPK/GLUT4 signaling pathway. Biomed Pharmacother. (2017) 95:556–63. doi: 10.1016/j.biopha.2017.08.094
13. Son YJ, Choi SY, Hwang IK, Nho CW, Kim SH. Could defatted mealworm (Tenebrio molitor) and mealworm Oil be used as food ingredients? Foods. (2020) 9:1–13. doi: 10.3390/foods9010040
14. Mancini S, Fratini F, Turchi B, Mattioli S, Dal Bosco A, Tuccinardi T, et al. Former foodstuff products in tenebrio molitor rearing: effects on growth, chemical composition, microbiological load, and antioxidant status. Animals. (2019) 9:484. doi: 10.3390/ani9080484
15. Tang Y, Debnath T, Choi EJ, Kim YW, Ryu JP, Jang S, et al. Changes in the amino acid profiles and free radical scavenging activities of Tenebrio molitor larvae following enzymatic hydrolysis. PLoS ONE. (2018) 13:e0196218. doi: 10.1371/journal.pone.0196218
16. Flores DR, Casados LE, Velasco SF, Ramírez AC, Velázquez G. Comparative study of composition, antioxidant and antimicrobial activity of two adult edible insects from Tenebrionidae family. BMC Chem. (2020) 14:55. doi: 10.1186/s13065-020-00707-0
17. Alves AV, Argandoña EJS, Linzmeier AM, Cardoso CAL, Macedo MLR. Chemical composition and food potential of Pachymerus nucleorum larvae parasitizing acrocomia aculeata kernels. PLoS ONE. (2016) 11:e0152125. doi: 10.1371/journal.pone.0152125
18. Dutta P, Dey T, Manna P, Kalita J. Antioxidant potential of Vespa affinis L., a traditional edible insect species of North East India. PLoS ONE. (2016) 11:e0156107. doi: 10.1371/journal.pone.0156107
19. Hall F, Johnson PE, Liceaga A. Effect of enzymatic hydrolysis on bioactive properties and allergenicity of cricket (Gryllodes sigillatus) protein. Food Chem. (2018) 262:39–47. doi: 10.1016/j.foodchem.2018.04.058
20. Hwang BB, Chang MH, Lee JH, Heo W, Kim JK, Pan JH, et al. The edible insect gryllus bimaculatus protects against gut-derived inflammatory responses and liver damage in mice after acute alcohol exposure. Nutrients. (2019) 11:40857. doi: 10.3390/nu11040857
21. Mintah BK, He R, Dabbour M, Agyekum AA, Xing Z, Golly MK, Ma H. Sonochemical action and reaction of edible insect protein: influence on enzymolysis reaction-kinetics, free-Gibbs, structure, and antioxidant capacity. J Food Biochem. (2019) 43:1–11. doi: 10.1111/jfbc.12982
22. Mintah BK, He R, Dabbour M, Xiang J, Agyekum AA, Ma H. Techno-functional attribute and antioxidative capacity of edible insect protein preparations and hydrolysates thereof: effect of multiple mode sonochemical action. Ultrason Sonochem. (2019) 58:104676. doi: 10.1016/j.ultsonch.2019.104676
23. Sun M, Xu X, Zhang Q, Rui X, Wu J, Dong M. Ultrasonic-assisted aqueous extraction and physicochemical characterization of oil from clanis bilineata. J Oleo Sci. (2018) 67:151–65. doi: 10.5650/jos.ess17108
24. Shantibala T, Lokeshwari RK, Debaraj H. Nutritional and antinutritional composition of the five species of aquatic edible insects consumed in Manipur, India. J Insect Sci. (2014) 14:1–10. doi: 10.1673/031.014.14
25. Longvah T, Manghtya K, Qadri SSYH. Eri silkworm: a source of edible oil with a high content of α-linolenic acid and of significant nutritional value. J Sci Food Agric. (2012) 92:1988–93. doi: 10.1002/jsfa.5572
26. Yoon S, Wong NAK, Chae M, Auh JH. Comparative characterization of protein hydrolysates from three edible insects: mealworm larvae, adult crickets, and silkworm pupae. Foods. (2019) 8:563. doi: 10.3390/foods8110563
27. Ahn MY, Kim BJ, Kim HJ, Jin JM, Yoon HJ, Hwang JS, et al. Anti-diabetic activity of field cricket glycosaminoglycan by ameliorating oxidative stress. BMC Complement Med Ther. (2020) 20:1–10. doi: 10.1186/s12906-020-03027-x
28. Koc K, Incekara U, Turkez H. Biomonitoring of the genotoxic effects and oxidative potentials of commercial edible dung beetles (Onitis sp.), grasshopper (Caelifera sp.) and mole crickets (Gryllotalpa sp.) in vitro. Toxicol Ind Health. (2014) 30:683–9. doi: 10.1177/0748233712457451
29. Memiş E, Türkez H, Incekara Ü, Banjo AD, Fasunwon BT, Togar B. In vitro biomonitoring of the genotoxic and oxidative potentials of two commonly eaten insects in southwestern Nigeria. Toxicol Ind Health. (2013) 29:52–9. doi: 10.1177/0748233712446721
30. Zou Y, Hu T, Shi Y, Liao S, Liu J, Mu L, et al. Silkworm pupae oil exerts hypercholesterolemic and antioxidant effects in high-cholesterol diet-fed rats. J Sci Food Agric. (2017) 97:2050–6. doi: 10.1002/jsfa.8009
31. Zhao J-G, Wang H-Y, Wei Z-G, Zhang Y-Q. Therapeutic effects of ethanolic extract from the green cocoon shell of silkworm Bombyx mori on type 2 diabetic mice and its hypoglycaemic mechanism. Toxicol Res (Camb). (2019) 8:407–20. doi: 10.1039/C8TX00294K
32. Ahn MY, Kim MJ, Kwon RH, Hwang JS, Park KK. Gene expression profiling and inhibition of adipose tissue accumulation of G. bimaculatus extract in rats on high fat diet. Lipids Health Dis. (2015) 14:1–15. doi: 10.1186/s12944-015-0113-3
33. Karna KK, Choi NY, Kim CY, Kim HK, Shin YS, Park JK. Gui-a-gra attenuates testicular dysfunction in varicocele-induced rats via oxidative stress, er stress and mitochondrial apoptosis pathway. Int J Mol Sci. (2020) 21:1–16. doi: 10.3390/ijms21239231
34. Ahn EM, Myung NY, Jung HA, Kim SJ. The ameliorative effect of Protaetia brevitarsis Larvae in HFD-induced obese mice. Food Sci Biotechnol. (2019) 28:1177–86. doi: 10.1007/s10068-018-00553-w
35. Choi RY, Ji M, Lee MK, Paik MJ. Metabolomics study of serum from a chronic alcohol-fed rat model following administration of defatted tenebrio molitor larva fermentation extract. Metabolites. (2020) 10:1–16. doi: 10.3390/metabo10110436
36. Wu LL, Chiou CC, Chang PY, Wu JT. Urinary 8-OHdG: a marker of oxidative stress to DNA and a risk factor for cancer, atherosclerosis and diabetics. Clin Chim Acta. (2004) 339:1–9. doi: 10.1016/j.cccn.2003.09.010
37. Papac-Milicevic N, Busch CJL, Binder CJ. Malondialdehyde Epitopes as Targets of Immunity and the Implications for Atherosclerosis. 1st ed. Amsterdam: Elsevier Inc. (2016). doi: 10.1016/bs.ai.2016.02.001
38. Marrocco I, Altieri F, Peluso I. Measurement and clinical significance of biomarkers of oxidative stress in humans. Oxid Med Cell Longev. (2017) 2017:e6501046. doi: 10.1155/2017/6501046
39. Lytvyn Y, Perkins BA, Cherney DZI. Uric acid as a biomarker and a therapeutic target in diabetes. Can J Diabetes. (2015) 39:239–46. doi: 10.1016/j.jcjd.2014.10.013
Keywords: entomophagy, antioxidants, edible insects, novel foods, functional foods, oxidative stress, sustainable nutrition
Citation: D'Antonio V, Serafini M and Battista N (2021) Dietary Modulation of Oxidative Stress From Edible Insects: A Mini-Review. Front. Nutr. 8:642551. doi: 10.3389/fnut.2021.642551
Received: 16 December 2020; Accepted: 25 January 2021;
Published: 26 February 2021.
Edited by:
Alessandra Bordoni, University of Bologna, ItalyReviewed by:
Lenka Kourimska, Czech University of Life Sciences Prague, CzechiaCopyright © 2021 D'Antonio, Serafini and Battista. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mauro Serafini, bXNlcmFmaW5pQHVuaXRlLml0
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.