 Ying Deng
Ying Deng Coen Govers
Coen Govers Ellen ter Beest1,2
Ellen ter Beest1,2 Aalt-Jan van Dijk
Aalt-Jan van Dijk Kasper Hettinga
Kasper Hettinga Harry J. Wichers
Harry J. Wichers
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Nutr. , 13 January 2021
Sec. Nutritional Immunology
Volume 7 - 2020 | https://doi.org/10.3389/fnut.2020.612397
Allergen recognition and processing by antigen presenting cells is essential for the sensitization step of food allergy. Macrophages and dendritic cells are both phagocytic antigen presenting cells and play important roles in innate immune responses and signaling between the innate and adaptive immune system. To obtain a model system with a homogeneous genetic background, we derived macrophages and dendritic cells from THP-1 monocytes. The difference between macrophages and dendritic cells was clearly shown by differences in their transcription response (microarray) and protein expression levels. Their resemblance to primary cells was analyzed by comparison to properties as described in literature. The uptake of β-lactoglobulin after wet-heating (60°C in solution) by THP-1 derived macrophages was earlier reported to be significantly increased. To analyse the subsequent immune response, we incubated THP-1 derived macrophages and dendritic cells with native and differently processed β-lactoglobulin and determined the transcription and cytokine expression levels of the cells. A stronger transcriptional response was found in macrophages than in dendritic cells, while severely structurally modified β-lactoglobulin induced a more limited transcriptional response, especially when compared to native and limitedly modified β-lactoglobulin. These results show that processing is relevant for the transcriptional response toward β-lactoglobulin of innate immune cells.
A food allergy is an immune mediated reaction against an otherwise harmless protein, also known as allergen (1, 2). Cow's milk allergy is one of the most common food allergies in the world which affects about 2.5% of all children (3). Thermal processing of milk is an essential step in the dairy industry to ensure microbiological safety and prolong shelf life (4). However, heat treatments like pasteurization, sterilization and spray drying will affect the structural characteristics of the proteins present in cow's milk, including the whey protein β-lactoglobulin (BLG) which is one of the major allergens present in milk (5–7). Structural modifications like unfolding, aggregation and glycation with lactose could occur during heat treatment, either reducing or enhancing the allergenic potential, i.e., IgE-binding of BLG (8, 9).
After passing through the epithelial cells of the intestine, BLG may be taken up by antigen-presenting cells (APC) of the innate immune system. Of these, macrophages and dendritic cells are considered to be professional APCs (10). They have the ability to capture an allergen into specialized endosomes/lysosomes, and then process it into linear peptides which are expressed on major histocompatibility complex (MHC) molecules on the outer surface, to which a naïve T cell can bind. Although both derived from the same precursor-monocytes, macrophages and dendritic cells exert different functions and possess different functional properties. Macrophages focus on scavenging allergens, manipulating inflammatory responses and recruiting T cells and B cells in situ. Dendritic cells are specialized in their ability to preserve the ingested allergens, driving T cell responses following their migration to the secondary lymphoid tissues (11).
To study the immunological response of macrophages and dendritic cells upon exposure to heat-treated BLG, macrophages and dendritic cells derived from THP-1 monocytes were used. This approach would circumvent possible effects of different genetic backgrounds on the cellular responses (12) providing a smaller variability in the read-out, and thereby a clearer picture of the effect of processing of proteins on the cellular response. THP-1 monocytes stimulated by phorbol-12-myristate-13-acetate (PMA) are regarded as macrophage-like cells, showing similarities with primary macrophages in morphology, gene transcription and expression (13–16). There are only few reports describing the differentiation of THP-1 monocytes into dendritic-like cells. THP-1 monocytes, stimulated with PMA together with IL-4, were shown to express typical cellular surface markers and exhibit properties similar to primary immature dendritic cells (17).
By using microarray analysis, the similarities and differences between THP-1 derived macrophages and dendritic cells were analyzed based on their transcriptome. Together with morphology, protein expression levels, and antigen processing capability analyses, their resemblance to primary cells was estimated in comparison with the reported properties of primary cells from literature (18). Differences in uptake capability of THP-1 derived macrophages for differently heat-treated BLG was described previously (19). To obtain an impression of the impact of exposure of THP-1 derived macrophages or dendritic cells to such differently heat-treated BLG on gene expression related to functional properties, the transcription levels of immune related genes and the production of a number of cytokines by THP-1 derived APCs, was analyzed in this study.
All chemicals were purchased from Sigma Aldrich (St Louis, Missouri, USA) unless otherwise stated.
Cells from the human monocytic leukemia cell line THP-1 (ATCC, Manassas, Virginia, USA) were differentiated into cells representative of resting macrophages (M0) as described by Chanput et al. (20). Cell suspensions with a concentration of 1 × 106 cells/mL were stimulated using 100 ng/mL phorbol-12-myristate-13-acetate (PMA) in Corning® 24 Well TC-Treated Microplates (Life Sciences, Tewksbury, Massachusetts, USA) with 500 μL volume per well, followed by 2 times washing with the same volume of medium after 48 h and another 48 h of resting. Similarly, using a cell concentration of 0.25 × 106 cells/mL, immature dendritic cells (iDC) were generated after incubating with 20 ng/mL of IL-4 (R&D Systems, Minneapolis, Minnesota, USA) and 20 ng/mL of PMA for 4 days as described by Katayama et al. (17) with the same cell suspension volume and in an identical plate as used for M0. Lipopolysaccharide (LPS) was used to differentiate M0 and iDC, generated with the previous protocol, into LPS-M and mature dendritic cells (mDC) at a concentration of 500 ng/mL and 1 μg/mL, respectively, for 48 h.
The morphology of cells in the 24 well-plate was observed by an automated digital inverted microscope Evos FL Auto 2 (Thermo Fisher, Waltham, Massachusetts, USA) using a 40 × objective lens. Scale bars were added to the picture using ImageJ software.
β-Lactoglobulin (BLG) was isolated from raw cow's milk as described in the protocol of De Jongh et al. (21). Lactose was removed by dialysis after which the purity of protein was higher than 94%, confirmed by size exclusion chromatography. In the absence or presence of glucose (molarity 80 times that of BLG), 13 mg/mL BLG was wet-heated (W), high-temperature dry-heated (H) or low-temperature dry-heated (L), centrifuged to remove insoluble material, dialyzed to remove free glucose and the protein concentration measured, as described previously (19). In the annotation of the samples, the first capital (L, H or W) referred to the type of processing, and the indication-glu in the sample name indicated the presence of glucose during the processing step. THP-1 derived M0 and iDC were incubated in Corning® 24 Well TC-Treated Microplates (Life Sciences, Tewksbury, Massachusetts, USA) with 100 μg/mL BLG, L-glu-BLG, H-glu-BLG, W-glu-BLG or the culture medium as a control for 6 h.
THP-1 derived M0 and iDC were exposed to BLG samples or medium as control in Corning® 24 Well TC-Treated Microplates (Life Sciences, Tewksbury, Massachusetts, USA) for 6 h. RNA of the cultured cells was extracted as described by Tang et al. (22), using 200 μL TRIzol (Invitrogen, Bleiswijk, The Netherlands) per well. Each cell experiment was performed in triplicate with cells from different passage numbers. RNA quality was verified using the RNA 6000 Nano assay on an Agilent 2100 Bioanalyzer (Agilent Technologies, Amstelveen, The Netherlands). Quality approved RNA was labeled and hybridized to the GeneChip® Human Gene 2.1 ST Array in a GeneTitan® platform (Affymetrix, Santa Clara, California, USA) according to standard protocols. The arrays were scanned by the high-resolution scanner of GeneChip Scanner (Affymetrix, Santa Clara, California, USA) and the signal was converted to cell intensity file (.CEL files) using GeneChip Operating Software (GCOS). The raw CEL data was normalized using the Bioconductor packages of MADMAX pipeline (23) and analyzed after data filtering. Final gene transcription levels were quantified by fold change of expression (FC). The FC of each individual gene of M0 or iDC was calculated based on the gene transcription level relative to the level of monocytes, while FC of each individual gene of M0 or iDC incubated with BLG samples was compared with the values for M0 or iDC without incubation, respectively. Genes with an FC for which the absolute value was > 2 and an intensity-based moderated t-statistics (IBMT) raw q < 0.05 were considered to be significantly differentially transcribed. The Venn diagrams were generated using Venny 2.1 (https://bioinfogp.cnb.csic.es/tools/venny/).
Pathway analysis was carried out using the program Ingenuity Pathway Analysis (IPA, Qiagen, Hilden, Germany). For M0 and iDC without BLG incubation and M0 incubated with native and low-temperature dry-heated BLG, only the significantly differentially transcribed genes were uploaded in the software. The IPA core analysis was done based on setting: “Human” in the “Species” category, and “THP-1,” ”Dendritic cells” or “Macrophages” in the category of “Tissue & Cell lines.” Pathways suggested by the software which yielded p < 0.001 were considered to be significantly associated with the gene transcription levels of this sample.
THP-1 derived M0 and iDC were cultivated in Corning® 96 Well TC-Treated Microplates (Life Sciences, Tewksbury, Massachusetts, USA) with a volume of 100 μL per well using the same protocol in the previous section on THP-1 cell Culture and Differentiation. The cells were harvested as described previously (19), first washed twice with PBS without Ca2+ and Mg2+ (Gibco, Thermo Fisher, Waltham, Massachusetts, USA) and incubated with 100 μL of 0.25% Trypsin-EDTA (Gibco, Thermo Fisher, Waltham, Massachusetts, USA) for 5 min, and subsequently centrifuged for 5 min at 450 × g. Cell surface marker staining was performed by first suspending the cell pellets after centrifugation with 100 μL PBS containing 10 μL FcR blocking reagent (Miltenyi Biotec, Auburn, CA) and incubating the cell suspensions at 4°C for 10 min. Then 90 μL PBS containing 1 μL of each detection antibody was added to the cells and incubated for 30 min at 4°C (gently mixed in between once). The following fluorescence-labeled mouse anti-human monoclonal antibodies (mAbs) were used: panel 1 [FITC-conjugated anti-CD209 (clone DCN46), PE-conjugated anti-HLA-DR (clone G46-6), PE-Cy™7-conjugated anti-CD83 (clone HB15e) and APC-conjugated anti-CD11c (clone B-ly6)]; panel 2 [FITC-conjugated anti-CD14 (clone M5E2), PE-conjugated anti-CD40 (clone 5C3), PE-Cy™7-conjugated anti-CD80 (clone L307.4) and APC-conjugated anti-HLA-ABC (clone G46-2.6)]; and panel 3 [FITC-conjugated anti-CD63 (clone H5C6) and APC-conjugated anti-CD81 (clone JS-81)] all from BD Biosciences (Franklin Lakes, New Jersey, USA). After staining, 1 mL PBS was added to the cells followed by centrifugation for 5 min at 450 × g. The cell pellet was resuspended in 150 μL PBS with Ca2+ and Mg2+ and the mean fluorescence intensity (MFI) of each fluorescence signal was analyzed for four-color immunofluorescence with an Accuri C6 flow cytometry set to compensate for overflow (BD Biosciences, Franklin Lakes, New Jersey, USA) of at least 5,000 cells for each sample using BD Accuri C6 software. The MFI of unstained cells was subtracted from each value after correcting the overflow of each fluorescence.
Supernatant of THP-1 derived M0 and iDC was collected following 16 h stimulation with medium or 100 μg/mL sample. The cytokine secretion measurement was done with the ELISA Deluxe Set Human IL-8, IL10, IL-6, TNF-α, CCL20, and IL-1β (Biolegend, San Diego, California, United States) following the manufacturer's protocol. The absorbance was measured with Infinite® 200 PRO NanoQuant (Tecan, Männedorf, Switzerland).
DQ-OVA is a self-quenched conjugate of ovalbumin which is strongly labeled with BODIPY dyes. It will exhibit bright green fluorescence upon proteolytic degradation into single, dye-labeled peptides. THP-1 derived M0 or iDC cells in Corning® 96 Well TC-Treated Microplates (Life Sciences, Tewksbury, Massachusetts, USA) with a volume of 100 μL per well using the same protocol as in the previous section on THP-1 cell Culture and Differentiation were incubated with medium containing 10 ug/mL DQ-OVA (Life Sciences, Tewksbury, Massachusetts, USA) for 1 h. After that, cells were washed twice with PBS without Ca2+ and Mg2+ (Gibco, Thermo Fisher, Waltham, Massachusetts, USA) and incubated with 100 μL of 0.25 % Trypsin-EDTA (Gibco, Thermo Fisher, Waltham, Massachusetts, USA) for 5 min. The cell suspension in Trypsin-EDTA was centrifuged for 5 min at 450 × g, after which the cell pellets were resuspended in 150 μL PBS with Ca2+ and Mg2+. The mean fluorescence intensity (MFI) of the FITC signal was recorded by flow cytometry of at least 5,000 cells for each sample using BD Accuri C6 software.
The statistical analysis of microarray data was performed as described in the previous section on Microarray Analysis. The statistical analysis of other data was done using Prism 6 software or R studio (package “agricolae”), considering p < 0.05 to be significant.
To build genetically homogenous immune cell models, we used the THP-1 monocytic cell line to obtain macrophage- and dendritic-like cells. After 2 days of culturing in medium supplemented with 100 ng/mL of PMA followed by 2 days of rest or 4 days with 20 ng/mL of PMA and 20 ng/mL of IL-4, THP-1 monocytes differentiated into macrophage-like cells in the resting stage (M0) or immature dendritic-like cells (iDC), respectively. We first compared the morphology of M0 and iDC to their precursor monocytes. Morphology of both induced cell types demonstrated an increase in size, while M0 maintained their globosity, and iDC showed an elongated shape (Figure 1). In addition, both induced cell types were adherent in contrast to the monocytes.

Figure 1. The morphologies of THP-1 monocytes and their derived macrophage-like cells at resting stage (M0) and immature dendritic-like cells (iDC) can be distinguished from each other. THP-1 monocytes were non-treated or differentiated into M0 or iDC in a 24 well-plate and analyzed under the microscope. Scale bar is 20 μm. Representative pictures of repeated differentiation experiments were shown.
In order to comprehensively analyse and compare M0 and iDC, mRNA was extracted and gene transcription was analyzed. The gene transcription of M0 and iDC was compared qualitatively and quantitatively to that of monocytes to obtain an indication of up- or down-regulation of specific macrophage and dendric cell related genes. In total, 5,307 genes were found to be significantly differentially transcribed in at least one of the cell types out of more than 25,000 tested genes. These genes revealed a moderate correlation (Figure 2A; r = 0.645, p < 0.001) between M0 and iDC in a scatter plot. Moreover, half of the significantly differentially transcribed genes were shared by both cell types, but these cell types also showed many uniquely transcribed genes (Figure 2B). These results were further analyzed based on the top 25 genes with the highest FC for M0 (Supplementary Table 1) and for iDC (Supplementary Table 2). In general, the fold changes in transcription levels of iDC were much higher than for M0. Supplementary Table 1 shows that the transcription levels of CYBB, IL7R, CD180, WIPF3, KCNMA1, MMP2, TLR7, ZC3H12D and CHODL were higher in M0 and these genes are related to antigen uptake, innate immunity and cell signaling (24–27). Supplementary Table 1 also shows increased transcription levels of ENDRA, S1PR1, CHRM3, C3AR1 in M0 which are related to G proteins and coupled receptors (28–31). Supplementary Table 2 contains four SERPINB related genes with increased transcription in iDC, which might be due to the IL-4 used for cell stimulation (32, 33). Furthermore, IPA indicated AQP9, CCL22, INHBE, CD274, MMP7, and VCAM1 (Supplementary Table 2) to be related to immunological responses (34–39), EHF, SCIMP, NR4A3, and ALDH1A2 to be related to dendritic cells (40–43), and GJB2 and VCAM1 to be related to cell adhesion (38, 44).
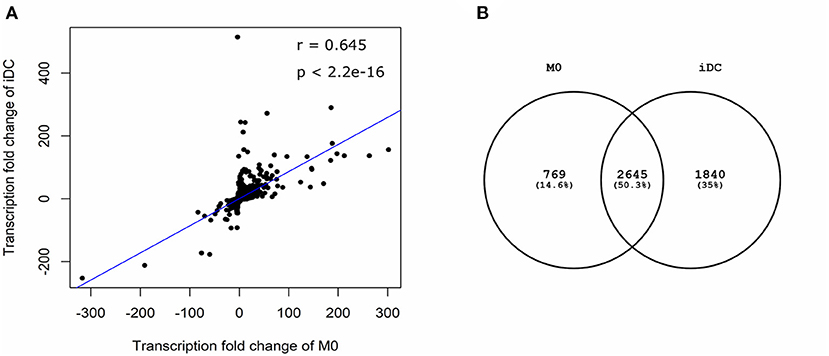
Figure 2. Differential gene transcription in THP-1 derived macrophage- (M0) and dendritic-like cells (iDC) showed similarities and differences. THP-1 monocytes were non-treated or differentiated into M0 and iDC and the transcriptome was analyzed using microarray. Microarray analysis provided data on the transcriptome which was expressed as average fold change (FC) between differentiated cells vs. monocytes of 3 independent experimental replicates. Genes with an absolute value of FC > 2 and q value < 0.05 in M0 or iDC when compared to non-treated monocytes were considered to be significant. A scatter plot (A) showed the correlation between significantly differentially expressed genes in M0 and iDC. The Pearson correlation coefficient (r) and two-tailed p-value were analyzed using R studio. The Venn diagram (B) showed the numbers of shared or exclusive significantly differentially expressed genes in M0 and iDC.
The microarray data were analyzed in a more comprehensive manner with Ingenuity Pathway Analysis (IPA) based on the significantly differentially transcribed genes for M0 and iDC. Pathway analysis is based on gene transcription of proteins that are empirically shown to interact in cellular pathways, providing evidence that a specific pathway might be activated or inhibited. Pathway analysis yielded significantly associated canonical pathways (p < 0.001) which are listed in Figure 3. Associated with both cell types were, amongst others, the pathways “Inhibition of Matrix Metalloproteases” and “Atherosclerosis Signaling and Aryl Hydrocarbon Receptor Signaling,” which are related to cell differentiation or proliferation and the pathways “IL-8 Signaling” and “Granulocyte Adhesion and Diapedesis,” which are related to inflammation and immune response. There were 9 pathways significantly associated with M0, among which “Macropinocytosis Signaling” and “Virus Entry via Endocytic Pathways” (both uptake related), “Paxillin Signaling” (cell adhesion and morphology related), “Acute Phase Response Signaling,” “IL-6 Signaling” and “Toll-like Receptor Signaling” (both proinflammatory and cytokine related). Eleven pathways were significantly associated with iDC, among which “Leukocyte Extravasation Signaling,” “IL-10 Signaling,” and “Agranulocyte Adhesion and Diapedesis” are all related to immune responses. Next to pathway analysis, IPA also predicted activation or inhibition of upstream regulators. This is done by comparing the microarray data to known gene transcription profiles linked to individual upstream regulators, which provides insights into potential activation or inhibition. As shown in Supplementary Table 3, upstream regulators ECSIT (intermediate in TLR Pathway) (45), CD40LG, TNF-α, TLR4, CD40, IL17A, IL6, and TNFRSF1A were found to be activated in both M0 and iDC.
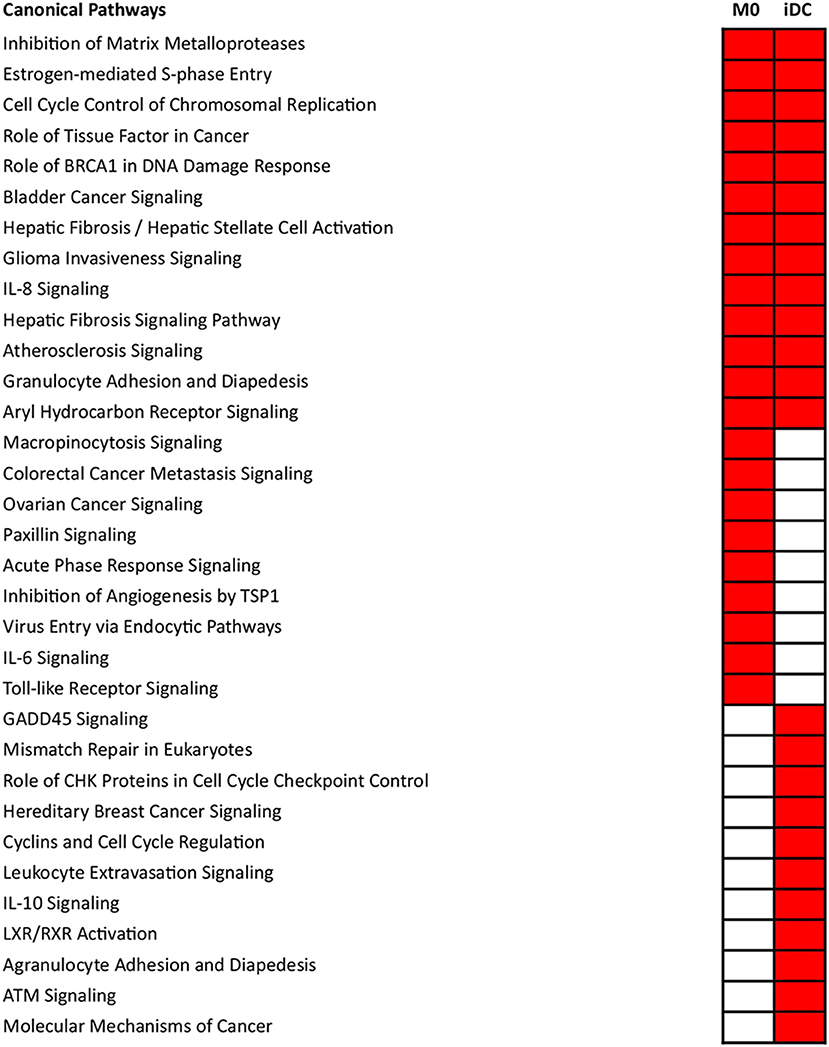
Figure 3. Significantly associated canonical pathways in THP-1 derived macrophage- (M0) and dendritic-like cells (iDC) differed from each other. THP-1 monocytes were non-treated or differentiated into M0 or iDC. Microarray analysis provided data on the transcriptome which was expressed as average fold change between differentiated cells vs. monocytes of 3 independent experimental replicates. Then, the transcriptomes were imported into IPA for canonical pathway analysis. Canonical pathways with a p < 0.001 were selected, labeled in red and listed separately according to the categories: present in both M0 and iDC, only M0 and only iDC. In each category, the pathways were listed from top to bottom following the p-value difference between M0 and iDC from large to small.
To further characterize the two different cell types generated from THP-1 monocytes, we measured their cell surface marker expression and secretion of cytokines. In addition, to compare cells in an inflammatory state, we challenged them with LPS, resulting in LPS-stimulated macrophages (LPS-M) and mature dendritic cells (mDC). As shown in Figure 4, expression of circulating monocyte-derived macrophage differentiation markers CD14 and HLA-ABC (46, 47) were significantly higher in M0 compared to monocytes or iDC, while the expression of circulating monocyte-derived dendritic cell marker and T cell activator CD209 (48) was significantly higher in iDC compared to monocytes or M0. Expression levels of primary APC surface markers CD11c and CD81 (49, 50) were similar in M0 and iDC and both significantly higher than in monocytes. Furthermore, CD63 which was reportedly expressed on circulating monocyte-derived immature dendric cells (51), showed significantly higher expression in iDC but not in M0 compared to monocytes. After LPS stimulation, expression of CD83 and CD40, which are related to dendritic cell maturation and interaction with T and B cells (52), were significantly higher in mDC than iDC or LPS-M. CD209 was significantly decreased in mDC compared to iDC. On the other hand, cell surface expression levels of CD14 and CD80 (activated circulating monocyte-derived macrophage marker) (53) were significantly higher on LPS-M than on both M0 and mDC.
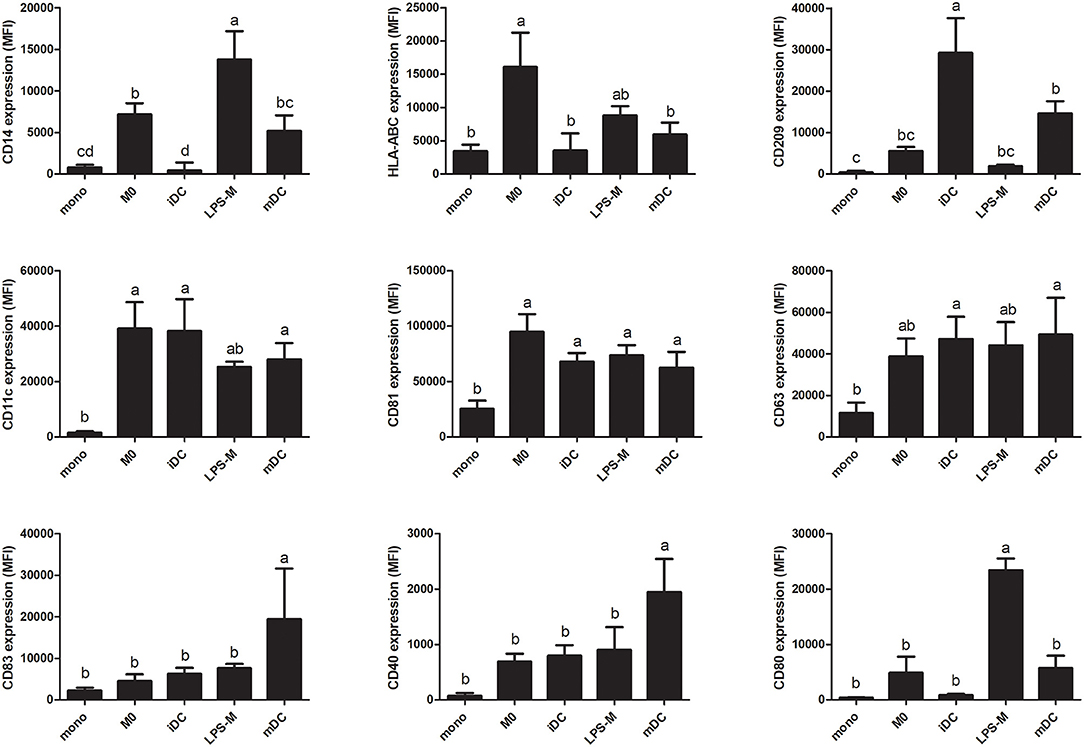
Figure 4. THP-1-derived macrophage- (M0) and dendritic-like cells (iDC) and their LPS-stimulated forms LPS-M and mDC showed different surface marker profiles compared to monocytes. THP-1 monocytes were non-treated or differentiated into M0 and iDC and/or stimulated with LPS yielding LPS-M and mDC, respectively. Expression of surface markers was analyzed through staining with fluorescent-labeled antibodies and MFI was measured by flow cytometry. The results represent the mean MFI ± SD of 5 independent experiments. The statistical differences were calculated with Fishers Least Significant Difference (LSD) test and bars with different letters represent significant differences (p < 0.05).
The secretion of cytokines also showed significant differences between M0 and iDC, and their corresponding cell types after LPS stimulation (Figure 5). Of note, none of the tested cytokines (i.e. IL-6, IL-10, IL-8, TNF-α, and IL-12) reached threshold detection levels in the monocyte culture (data not shown). We also did not observe a significant increase in IL-6 or IL-10 secretion upon differentiation to M0 or iDC. However, the secretion levels of IL-8 and TNF-α (proinflammatory cytokines) in iDC were significantly higher than in M0 (avg. 22.0 vs. 0.2 and 0.3 vs. 0 ng/mL, respectively). LPS stimulation led to higher cytokine production for both stimulated cell types compared to their corresponding non-stimulated cells for all tested cytokines. The significantly higher secretion levels of IL-8 and TNFα in iDC compared to M0 were maintained in their LPS-stimulated cells with a general higher production level (avg. 75.8 vs. 57.1 and 3.9 vs. 2.2 ng/mL). On the contrary, the production of IL-6 and IL-10 in LPS-M became significantly higher than in mDC (avg. 1.7 vs. 0.4 and 3.9 vs. 0.5 ng/mL). IL-12 production was not detected in any of the cell types or differentiation stages.
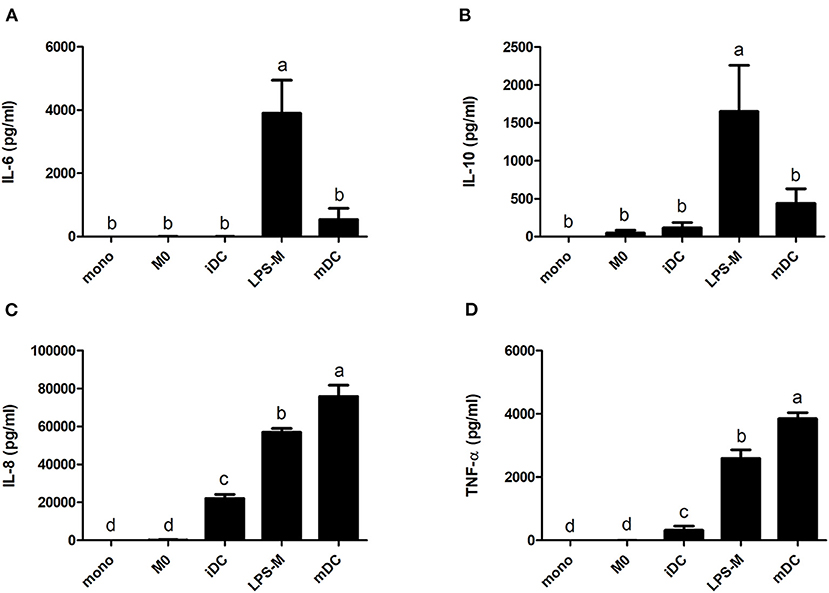
Figure 5. LPS-mediated activation resulted in significantly different increases of cytokine production for THP-1-derived macrophage- (M0) and dendritic-like cells (iDC). THP-1 monocytes were non-treated or differentiated into M0 and iDC and stimulated with LPS. The supernatant of the cell cultures was collected, and the cytokines levels of IL-6 (A), IL-10 (B), IL-8 (C), and TNF-α (D) were determined using ELISA. The results represent mean values ± SD of 2–4 independent experimental measurements. The statistical differences were calculated with Fishers Least Significant Difference (LSD) test and bars with different letters represent significant differences (p < 0.05).
A key hallmark of APCs is the internalization and processing of antigens for subsequent presentation to adaptive immune cells. Using DQ-OVA, we tested the antigen uptake and processing capacity of M0 and iDC. Comparing the numbers of cells that were able to process DQ-OVA, there were no significant differences between the phenotypes (Supplementary Figure 1). Comparing the MFI-values, both M0 and iDC showed significantly increased antigen processing capacity compared to monocytes, but no significant difference was observed between M0 and iDC (Figure 6).
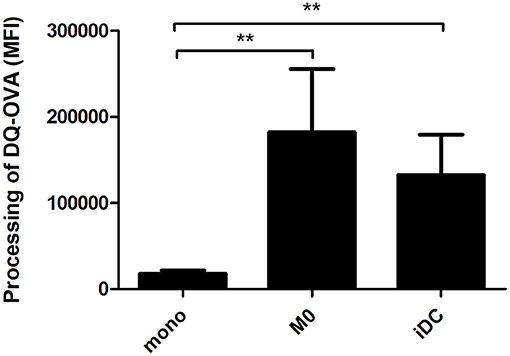
Figure 6. THP-1-derived macrophage- (M0) and dendritic-like cells (iDC) revealed a significantly increased capacity to process antigens compared to monocytes. Proteolytic cleavage of fluorophores from DQ-OVA (initially auto-quenching due to heavily labeling) result in fluorescent signals. THP-1 monocytes were non-treated or differentiated into M0 and iDC and incubated with DQ-OVA for 2 h, after which the florescence signal was measured by flow cytometry. The results represent the average MFI ± SD of 5 independent experimental measurements. The statistical differences between any two data set were calculated with two-tailed unpaired T-test, **p < 0.01.
Upon establishing the differences between M0 and iDC, and their similarities with primary monocyte derived macrophages and dendritic cells, the THP-1 derived M0 and iDC model was employed to study the impact of processed BLG samples. M0 and iDC were exposed to native, wet-heated, high- and low-temperature dry-heated BLG in the presence of glucose (BLG, W-glu-BLG, H-glu-BLG, and L-glu-BLG, respectively), after which gene transcription levels were analyzed using microarray and compared to non-treated M0 and iDC.
In general, there were more upregulated than downregulated genes in either cell type treated by either sample type. In M0 treated with BLG and L-glu-BLG, the highest total number of significantly differentially transcribed genes was found (247 and 173, respectively; Figure 7A). Figure 7C showed that most of the significantly differentially transcribed genes in M0 incubated with L-glu-BLG were also significantly differently transcribed in M0 incubated with BLG. In M0 treated with H-glu-BLG or W-glu-BLG and iDC treated with any sample, the number of significantly differentially transcribed genes was below 20 (Figures 7A,B), for which details can be found in Supplementary Tables 4, 5.
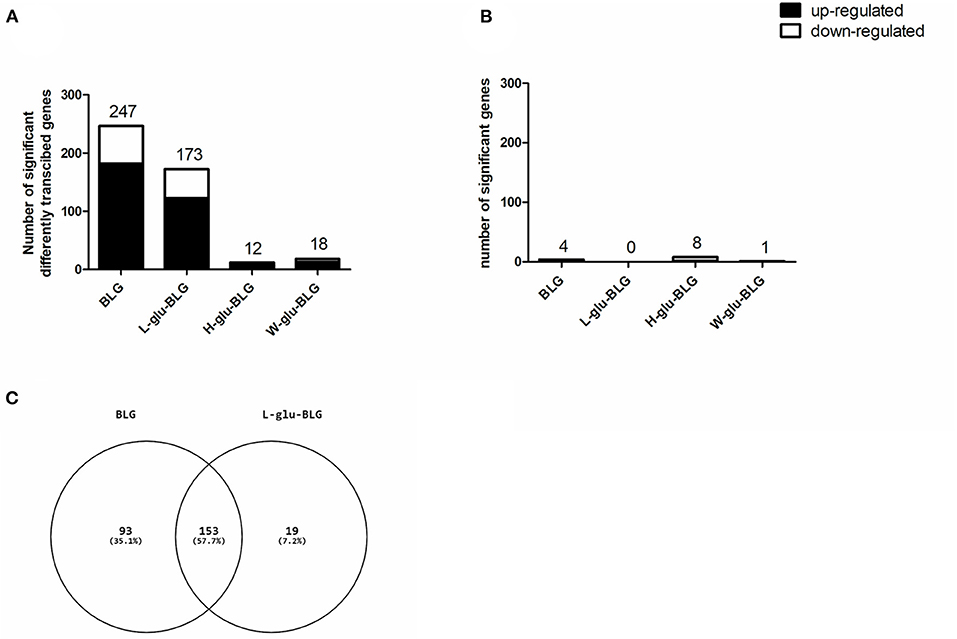
Figure 7. After exposure to BLG samples, THP-1 derived dendritic-like cells (iDC) hardly showed a response in gene transcription profiles if compared to medium control, while the response of macrophage-like cells (M0) mainly occurred in response to native and L-glu-BLG. THP-1 monocytes were differentiated into M0 and iDCs and incubated with medium (non-treated) or different BLG samples (native, L-glu-BLG, H-glu-BLG, or W-glu-BLG) for 6 h. Microarray analysis provided data on the transcriptome which was expressed as average fold change between treated vs. non-treated cells of 3 independent experimental replicates. The bar chart indicates the number of significantly (with fold change absolute value > 2 and q value < 0.05) up or down regulated genes for M0 (A) and iDC (B) treated with native or processed BLG. In a Venn diagram the numbers of significantly transcribed genes which are shared or exclusive for M0 incubated with BLG or L-glu-BLG were shown (C).
A more comprehensive analysis of gene transcription profiles of M0 treated with native or low-temperature dry-heated BLG was performed with IPA. Analysis of significantly associated canonical pathways revealed that both BLG and L-glu-BLG induced immune related pathways in M0 (Figure 8). Inflammatory-related pathways were significantly associated with M0 treated with BLG and L-glu-BLG, including “Th1 Pathway,” “NF-κB Signaling,” “TNFR1 Signaling,” “TWEAK Signaling,” “Activation of IRF by Cytosolic Pattern Recognition Receptors,” “TNFR1 Signaling,” “Toll-like Receptor Signaling,” “CD40 Signaling,” “HMGB1 Signaling,” and “TREM1 Signaling.” Additionally, pathways such as the “IL-23 Signaling Pathway” (Th17 maintenance and expansion), “Communication between Innate and Adaptive Immune Cells,” “Agranulocyte Adhesion and Diapedesis,” “Granulocyte Adhesion and Diapedesis” and “IL-15 Production” (regulatory of natural killer cells and T cells), which are all related to adaptive immunity signaling, were also significantly associated with M0 treated with BLG and L-glu-BLG. Exposure of M0 to BLG, but not L-glu-BLG, showed a significant association with the “Th17 Activation Pathway” and “IL-12 Signaling and Production in Macrophages.” On the contrary, several B cell related and Th2 pathways were only significantly associated with M0 incubated with L-glu-BLG.
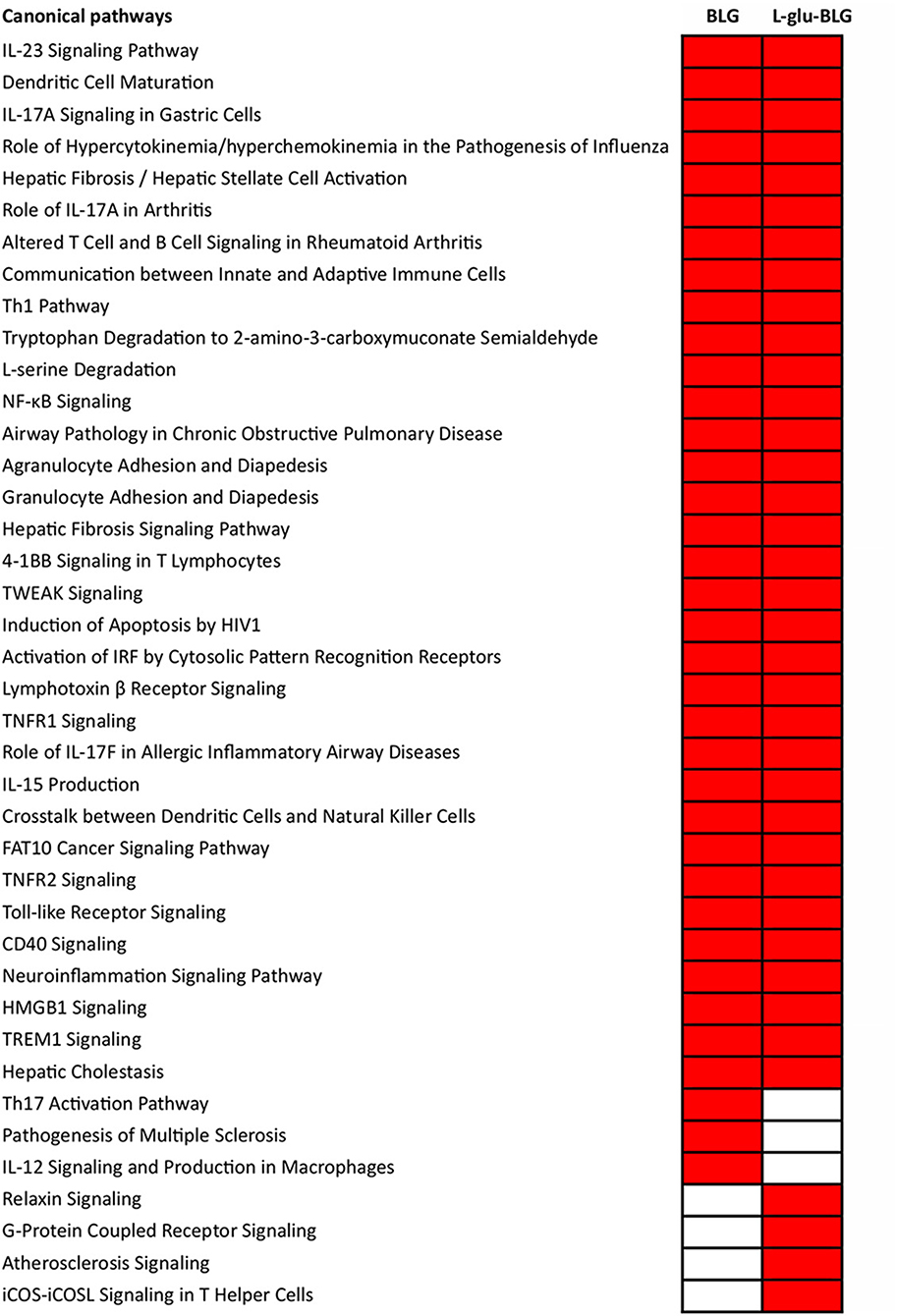
Figure 8. Significantly associated canonical pathways in THP-1 derived macrophage-like cells (M0) incubated with BLG and L-glu-BLG were mostly related to immune responses. THP-1 monocytes were differentiated into M0 cells and were incubated with medium (non-treated), native or L-glu-BLG for 6 h. Microarray analysis provided data on the transcriptome which was expressed as average fold change between treated vs. non-treated cells of 3 independent experimental replicates. Then, the transcriptomes were imported into IPA for canonical pathway analysis. Canonical pathways with a p < 0.001 were selected, labeled in red and listed separately according to the categories: present in both native and L-glu-BLG treated M0, only native treated M0 and only L-glu-BLG treated M0. In each category, the pathways were listed from top to bottom following the p-value difference between native and L-glu-BLG treated M0 from large to small.
As shown in Supplementary Table 6, in M0 treated with native BLG, innate immune related regulators ECSIT, IFNG (IFNγ), TLR4, IL27, TLR2, TNF (TNF-α), and IL-1β were significantly activated while LY6E, which has immunosuppressive effects (54), was inhibited. There was no clear difference in the type of activated or inhibited upstream regulators in M0 incubated with L-glu-BLG compared to BLG, but the activation levels (based on z-scores) were weaker in the former condition.
Similar as for establishing the M0 and iDC cell models, cytokine and chemokine analysis was performed to further characterize M0 responses to native and L-glu-BLG. From the gene transcription data, cytokine genes with a significantly differential transcription level in M0 following incubation with any BLG sample are presented in Table 1 (there were no significantly differentially transcribed cytokine gene for any of the iDC stimulations). In fact, significantly differentially transcribed cytokine or chemokine genes were found only in M0 incubated with BLG or L-glu-BLG with transcription levels being slightly higher in the former. Significant increases in transcription levels when compared to medium stimulated M0 were found for IL23A, IL32, CCL1, CCL4, CCL5, CCL19, CCL22, CXCL3, CXCL8, CXCL10, and CXCL11 following M0 stimulation with native BLG and/or L-glu-BLG. The transcriptional data was verified by protein detection for IL-8 (CXCL8), CCL20 and IL-1β. M0 incubated with native or heat-treated BLG samples showed a significantly higher secretion of all cytokines after treatment with native BLG and L-glu-BLG compared to other BLG samples (55). In iDC, BLG and L-glu-BLG similarly induced a significantly increased secretion of IL-8 and IL-1β, but not CCL20 (Figure 9).
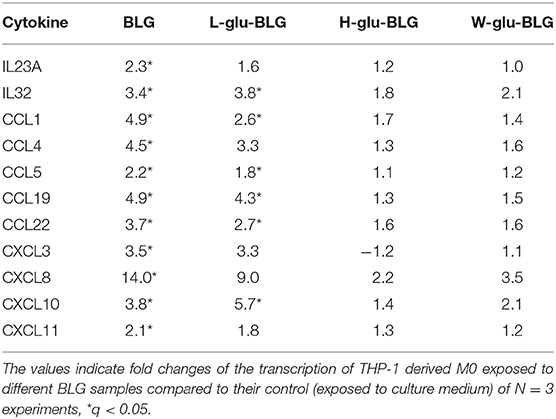
Table 1. Transcription fold change of cytokine genes in M0 incubated with BLG samples.
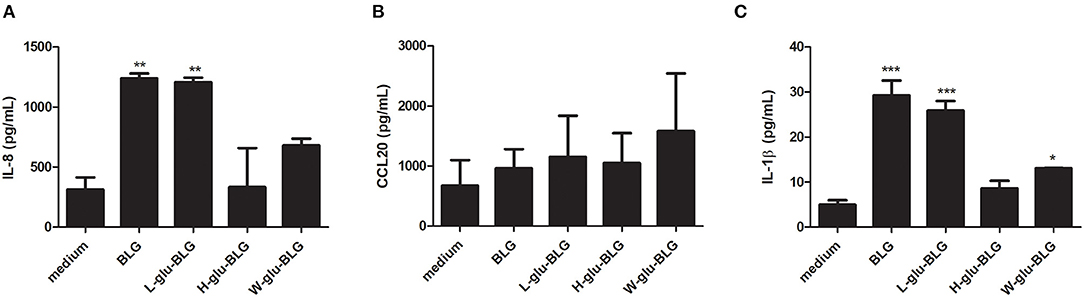
Figure 9. Cytokine response of THP-1 derived dendritic-like cells (iDC) upon exposure to native or heat-treated BLG varies. BLG was untreated or heated (W, H, or L) in the presence of glucose and incubated with iDC. The cytokine secretion of IL-8 (A), CCL20 (B), and IL-1β (C) in the supernatant was analyzed with ELISA. The results represent mean values ± SD of 3 independent experimental measurements. The statistical differences were calculated with Dunnett's Test compared to medium control: *p < 0.05; **p < 0.01; ***p < 0.001.
The differentiation of THP-1 monocytes into macrophage-like cells (M0) by PMA is a common method, whereas studies about the differentiation into dendritic-like cells are relatively limited (56). Based on the protocol of Katayama et al. (17), we generated THP-1 derived immature dendritic-like cells (iDC) using PMA and IL-4. The morphology, transcription profile, expression and secretion levels of surface markers and cytokines and chemokines, and antigen processing capability of THP-1 derived M0 and iDC were determined, and these were compared to their progenitor monocytes. Although PMA was used to derive both cell types, the lower concentration of PMA and the addition of IL-4 induced dendritic-like cells as a clearly different cell type from macrophages and monocytes. The difference of M0 and iDC from their progenitor THP-1 monocytes is clear from their morphology (Figure 1), as well as the vast number of significantly transcribed genes compared to monocytes (Figure 2). There were in total 5,254 genes significantly differentially transcribed in M0 and iDC compared to monocytes, of which about half were shared by both cell types. From the canonical pathway analysis in Figure 3, both M0 and iDC showed significant association with cell differentiation and immune response related pathways, indicating their differentiated cell identities and essential role in immune responses. Their antigen processing capabilities were illustrated by the enhanced proteolytic cleavage of DQ-OVA when compared to monocytes, as shown in Figure 6.
The difference between iDC and M0 was also obvious from their morphology, with a typical circular shape and the presence of dendrites for iDC (Figure 1). When looking at the significantly differentially transcribed genes compared to monocytes, the correlation between the fold change of M0 and iDC is moderate (Figure 2A), but with many uniquely differentially transcribed genes for each cell type, suggesting clear phenotypic differences. The fact that iDC had a higher number of significantly differentially transcribed genes than M0 might be caused by stronger activation of upstream regulators (TNF-α, TLR4, TREM1, and TSLP) and secretion of IL-8 and TNF-α compared to M0 (Supplementary Table 3 and Figure 5). TNF-α and TLR4 play essential roles in activating T cells (57, 58). This coincided with the significantly associated pathways Leukocyte Extravasation Signaling and IL-10 Signaling (Figure 3). The C-type lectin-like receptor CD209 (DC-SIGN), which was significantly stronger expressed in iDC than in THP-1 monocytes and M0, is involved in dendritic cell-T cell interaction (59). These observations indicate the additional T cell signaling function of iDC next to their role in phagocytosis and other immune regulatory functions as APCs (60). On the other hand, M0 showed exclusive and significant associations with uptake and proinflammatory related pathways (Figure 3), which is in line with the more prominent role of M0 in inducing phagocytosis and inflammation compared to iDC. Expression of CD14 was significantly higher in M0, which is reported to be a pattern recognition receptor for LPS and related to the innate immune response of macrophages (61). LPS-stimulated M0 secreted significantly higher amounts of IL-6 and IL-10 (Figure 5), which is in line with other studies (62, 63). The transcription levels of the surface markers (Supplementary Table 7) corroborated the outcomes of the surface marker expression for both M0 and iDC. Overall, our THP-1 derived M0 and iDC could be distinguished from each other and showed clearly differing properties. Moreover, they successfully represented the primary macrophages and dendritic cells by showing antigen uptake, processing and inflammatory response by M0, and the function of T cell stimulation and signaling between the innate and adaptive immune system by iDC.
Although iDC had more significantly differentially transcribed genes and higher transcription levels compared to monocytes than M0, its response to BLG samples was much weaker than M0 in both transcription and cytokine secretion levels (Figures 7, 9). BLG was reported to have limited immunomodulatory effects when incubated with murine DC (64), whereas its immune effect on macrophages has not been clearly studied yet. As shown in Table 1 and Figure 9, incubation of M0 with native or L-glu-BLG induced increased gene transcription and protein secretion of cytokines when compared to W-glu-BLG or H-glu-BLG. The latter two samples were also reported to show the strongest difference from native BLG in terms of structure and uptake by M0 as described in our previous study (19). Thus, the lower cytokine secretion levels of the M0 treated by W-glu-BLG or H-glu-BLG compared to native BLG may be related to their severe structural change after heat processing.
The inflammatory pathways and activation of innate immunity related upstream regulators in M0 that were significantly associated with incubation with BLG and L-glu-BLG (Figure 8 and Supplementary Table 6) appear to indicate a pro-inflammatory effect of BLG. A considerable number of adaptive immunity and T cell signaling pathways were activated, which is not surprising as other studies also demonstrated that macrophages have the ability to regulate adaptive immune responses and even directly contact T cells (65, 66). The pro-inflammatory effect of BLG and L-glu-BLG could possibly be linked with the significant association of T helper 1 (Th1) Pathways (67). In addition to the Th1 response, the response of M0 to incubation with the BLG sample also seemed to impact Th17 cells, as suggested by the significant association with IL-23, Th17, and several IL-17 related pathways. There was no clear difference in the activation of upstream regulators by M0 incubated with BLG or L-glu-BLG, and they also shared most of the significantly associated pathways. In a study, where human primary monocyte-derived dendritic cells were used and the cytokine profile of CD4+ T cells, co-cultured with these dendritic cells, was measured, exposure to native ovalbumin was also associated with Th1 cytokine induction (IFN-γ) (68). Endocytic uptake is the crucial and fundamental step in inducing further responses by APCs (69). As indicated by our results, M0 incubated with BLG samples showed stronger transcriptional responses than iDC. Wet-heated BLG, which has undergone the severest physicochemical modifications and also showed the highest uptake among all tested samples (19), did not cause the strongest transcriptional response by innate immune cells. There is thus no clear relationship between the uptake efficiency in M0 and the transcription-inducing effects of BLG. Although the underlying mechanisms are unclear, the data show that less modified BLG leads to a more extensive transcriptional response, indicating that extensive heating destructs immunogenic structures on BLG. Glycation, as occurring at low-temperature dry-heated conditions, might change the immunogenicity of BLG toward a Th2 differentiation enhancing direction which is related to chronic inflammatory disease and allergy (70).
In conclusion, dendritic-like cells were generated successfully from THP-1 monocytes, which clearly differed from both their progenitor and M0. In addition, a high level of uptake of processed BLG by APCs was shown to not be linked to a high level of gene transcription and cytokine production. It should be noted that this work is based on in vitro experiments, the result of which still need to be further verified in vivo, as functional responses of immune cells, rather than uptake or gene transcription alone, are required to identify the immunogenicity of proteins before and after processing-induced changes.
The datasets generated for this study can be found in the article/Supplementary Material.
YD acquired and analyzed all data. CG, EB, and A-JD contributed to the data analysis. YD, CG, KH, and HW participated in the design and interpretation of the reported experiments and results. YD drafted and edited the manuscript. CG, KH, and HW revised and edited the manuscript. All authors read and approved the final manuscript.
This work was financial supported by the China Scholarship Council (No. 201507720020), the Dutch Ministry of Economic Affairs [TKI-AF Grant No. 15269 (Sustainable Future Proteins)], and the Dutch Ministry of Agriculture, Nature and Food Quality (Grant No. KB-37-001-007).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors thank Renata Ariens and Nicole de Wit from Wageningen Food & Biobased Research for assistance in analyzing the microarray data.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnut.2020.612397/full#supplementary-material
1. Mills ENC, Tatham AS, Caballero B. ALLERGENS. In: Trugo L, Finglas PM, editor, Encyclopedia of Food Sciences and Nutrition. 2nd Edn. Waltham, MA: Academic Press (2003). p. 143–50.
2. Rahaman T, Vasiljevic T, Ramchandran L. Effect of processing on conformational changes of food proteins related to allergenicity. Trends Food Sci Technol. (2016) 49:24–34. doi: 10.1016/j.tifs.2016.01.001
3. Tsabouri S, Douros K, Priftis KN. Cow's milk allergenicity. Endocr Metab Immune Disord Drug Targets. (2014) 14:16–26. doi: 10.2174/1871530314666140121144224
4. Morr CV. Functionality of heated milk proteins in dairy and related foods. J Dairy Sci. (1985) 68:2773–81. doi: 10.3168/jds.S0022-0302(85)81165-6
5. Wal J-M. Cow's milk allergens. Allergy. (1998) 53:1013–22. doi: 10.1111/j.1398-9995.1998.tb03811.x
6. Rytkönen J, Karttunen TJ, Karttunen R, Valkonen KH, Jenmalm MC, Alatossava T, et al. Effect of heat denaturation on beta-lactoglobulin-induced gastrointestinal sensitization in rats: Denatured βLG induces a more intensive local immunologic response than native βLG. Pediatri Allergy Immunol. (2002) 13:269–77. doi: 10.1034/j.1399-3038.2002.01028.x
7. Villa C, Costa J, Oliveira MBPP, Mafra I. Bovine milk allergens: a comprehensive review. Compr Rev Food Sci Food Saf. (2018) 17:137–64. doi: 10.1111/1541-4337.12318
8. Bu GH, Luo YK, Zheng Z, Zheng H. Effect of heat treatment on the antigenicity of bovine -lactalbumin and -lactoglobulin in whey protein isolate. Food Agric Immunol. (2009) 20:195–206. doi: 10.1080/09540100903026116
9. Zenker HE, Ewaz A, Deng Y, Savelkoul HFJ, Van Neerven RJJ, De Jong NW, et al. Differential effects of dry vs. wet heating of beta-lactoglobulin on formation of sRAGE binding ligands and sIgE epitope recognition. Nutrients. (2019) 11:1432. doi: 10.3390/nu11061432
10. Schuijs MJ, Hammad H, Lambrecht BN. Professional and ‘Amateur' antigen-presenting cells in type 2 immunity. Trends Immunol. (2019) 40:22–34. doi: 10.1016/j.it.2018.11.001
11. Savina A, Amigorena S. Phagocytosis and antigen presentation in dendritic cells. Immunol Rev. (2007) 219:143–56. doi: 10.1111/j.1600-065X.2007.00552.x
12. Chanput W, Mes JJ, Wichers HJ. THP-1 cell line: an in vitro cell model for immune modulation approach. Int Immunopharmacol. (2014) 23:37–45. doi: 10.1016/j.intimp.2014.08.002
13. Auwerx J. The human leukemia cell line, THP-1: a multifacetted model for the study of monocyte-macrophage differentiation. Experientia. (1991) 47:22–31. doi: 10.1007/BF02041244
14. Kohro T, Tanaka T, Murakami T, Wada Y, Aburatani H, Hamakubo T, et al. A comparison of differences in the gene expression profiles of phorbol 12-myristate 13-acetate differentiated THP-1 cells and human monocyte-derived macrophage. J Atheroscler Thromb. (2004) 11:88–97. doi: 10.5551/jat.11.88
15. Daigneault M, Preston JA, Marriott HM, Whyte MKB, Dockrell DH. The Identification of markers of macrophage differentiation in PMA-stimulated THP-1 cells and monocyte-derived macrophages. PLoS ONE. (2010) 5:e8668. doi: 10.1371/journal.pone.0008668
16. Riddy DM, Goy E, Delerive P, Summers RJ, Sexton PM, Langmead CJ. Comparative genotypic and phenotypic analysis of human peripheral blood monocytes and surrogate monocyte-like cell lines commonly used in metabolic disease research. PLoS ONE. (2018) 13:e0197177. doi: 10.1371/journal.pone.0197177
17. Katayama S, Kukita T, Ishikawa E, Nakashima S, Masuda S, Kanda T, et al. Apple polyphenols suppress antigen presentation of ovalbumin by THP-1-derived dendritic cells. Food Chem. (2013) 138:757–61. doi: 10.1016/j.foodchem.2012.10.076
18. Ferenbach D, Hughes J. Macrophages and dendritic cells: what is the difference? Kidney Int. (2008) 74:5–7. doi: 10.1038/ki.2008.189
19. Deng Y, Govers C, Bastiaan-Net S, van Der Hulst N, Hettinga K, Wichers HJ. Hydrophobicity and aggregation, but not glycation, are key determinants for uptake of thermally processed beta-lactoglobulin by THP-1 macrophages. Food Res Int. (2019) 120:102–13. doi: 10.1016/j.foodres.2019.01.038
20. Chanput W, Mes JJ, Savelkoul HF, Wichers HJ. Characterization of polarized THP-1 macrophages and polarizing ability of LPS and food compounds. Food Funct. (2013) 4:266–76. doi: 10.1039/C2FO30156C
21. De Jongh HHJ, Groneveld T, De Groot J. Mild isolation procedure discloses new protein structural properties of beta-lactoglobulin. J Dairy Sci. (2001) 84:562–71. doi: 10.3168/jds.S0022-0302(01)74508-0
22. Tang YF, Govers C, Wichers HJ, Mes JJ. Macrophages treated with non-digestible polysaccharides reveal a transcriptionally unique phenotype. J Funct Foods. (2017) 36:280–9. doi: 10.1016/j.jff.2017.07.003
23. Lin K, Kools H, De Groot PJ, Gavai AK, Basnet RK, Cheng F, et al. MADMAX - Management and analysis database for multiple ~omics experiments. J Integr Bioinform. (2011) 8:160. doi: 10.1515/jib-2011-160
24. Vanheel A, Daniels R, Plaisance S, Baeten K, Hendriks JJA, Leprince P, et al. Identification of protein networks involved in the disease course of experimental autoimmune encephalomyelitis, an animal model of multiple sclerosis. PLoS ONE. (2012) 7:e35544. doi: 10.1371/journal.pone.0035544
25. Lundstrom W, Highfill S, Walsh STR, Beq S, Morse E, Kockum I, et al. Soluble IL7R alpha potentiates IL-7 bioactivity and promotes autoimmunity. Proc Natl Acad Sci USA. (2013) 110:E1761–70. doi: 10.1073/pnas.1222303110
26. Frazao JB, Thain A, Zhu Z, Luengo M, Condino-Neto A, Newburger PE. Regulation of CYBB gene expression in human phagocytes by a distant upstream NF-kappaB binding site. J Cell Biochem. (2015) 116:2008–17. doi: 10.1002/jcb.25155
27. Yang YH, Wang CL, Cheng PP, Zhang XB, Li XH, Hu Y, et al. CD180 Ligation inhibits TLR7-and TLR9-mediated activation of macrophages and dendritic cells through the Lyn-SHP-1/2 axis in murine lupus. Front Immunol. (2018) 9:2643. doi: 10.3389/fimmu.2018.02643
28. Shraga-Levine Z, Sokolovsky M. Functional coupling of G proteins to endothelin receptors is ligand and receptor subtype specific. Cell Mol Neurobiol. (2000) 20:305–17. doi: 10.1023/A:1007010125316
29. Cero C, Vostrikov VV, Verardi R, Severini C, Gopinath T, Braun PD, et al. The TLQP-21 peptide activates the G-protein-coupled receptor C3aR1 via a folding-upon-binding mechanism. Structure. (2014) 22:1744–53. doi: 10.1016/j.str.2014.10.001
30. Wu V, Yeerna H, Nohata N, Chiou J, Harismendy O, Raimondi F, et al. Illuminating the Onco-GPCRome: novel G protein-coupled receptor-driven oncocrine networks and targets for cancer immunotherapy. J Biol Chem. (2019) 294:11062–86. doi: 10.1074/jbc.REV119.005601
31. Patrone M, Cammarota E, Berno V, Tornaghi P, Mazza D, Degano M. Combinatorial allosteric modulation of agonist response in a self-interacting G-protein coupled receptor. Commun Biol. (2020) 3:27. doi: 10.1038/s42003-020-0752-4
32. Izuhara K, Yamaguchi Y, Ohta S, Nunomura S, Nanri Y, Azuma Y, et al. Squamous cell carcinoma antigen 2 (SCCA2, SERPINB4): an emerging biomarker for skin inflammatory diseases. Int J Mol Sci. (2018) 19:1102. doi: 10.3390/ijms19041102
33. Okawa T, Yamaguchi Y, Kou K, Ono J, Azuma Y, Komitsu N, et al. Serum levels of squamous cell carcinoma antigens 1 and 2 reflect disease severity and clinical type of atopic dermatitis in adult patients. Allergol Int. (2018) 67:124–30. doi: 10.1016/j.alit.2017.06.016
34. Buhler LA, Samara R, Guzman E, Wilson CL, Krizanac-Bengez L, Janigro D, et al. Matrix metalloproteinase-7 facilitates immune access to the CNS in experimental autoimmune encephalomyelitis. BMC Neurosci. (2009) 10:17. doi: 10.1186/1471-2202-10-17
35. Sideras P, Apostolou E, Stavropoulos A, Sountoulidis A, Gavriil A, Apostolidou A, et al. Activin, neutrophils, and inflammation: just coincidence? Semin Immunopathol. (2013) 35:481–99. doi: 10.1007/s00281-013-0365-9
36. Richter JR, Sutton JM, Belizaire RM, Friend LA, Schuster RM, Johannigman TA, et al. Macrophage-derived chemokine (CCL22) is a novel mediator of lung inflammation following hemorrhage and resuscitation. Shock. (2014) 42:525–31. doi: 10.1097/SHK.0000000000000253
37. De Santis S, Serino G, Fiorentino MR, Galleggiante V, Gena P, Verna G, et al. Aquaporin 9 contributes to the maturation process and inflammatory cytokine secretion of murine dendritic cells. Front Immunol. (2018) 9:2355. doi: 10.3389/fimmu.2018.02355
38. Kong DH, Kim YK, Kim MR, Jang JH, Lee S. Emerging roles of vascular cell adhesion molecule-1 (VCAM-1) in immunological disorders and cancer. Int J Mol Sci. (2018) 19:1057. doi: 10.3390/ijms19041057
39. Ahtiainen M, Wirta EV, Kuopio T, Seppala T, Rantala J, Mecklin JP, et al. Combined prognostic value of CD274 (PD-L1)/PDCDI (PD-1) expression and immune cell infiltration in colorectal cancer as per mismatch repair status. Mod Pathol. (2019) 32:866–83. doi: 10.1038/s41379-019-0219-7
40. Appel S, Bringmann A, Grunebach F, Weck MM, Bauer J, Brossart P. Epithelial-specific transcription factor ESE-3 is involved in the development of monocyte-derived DCs. Blood. (2006) 107:3265–70. doi: 10.1182/blood-2005-06-2480
41. Zhu B, Buttrick T, Bassil R, Zhu C, Olah M, Wu C, et al. IL-4 and retinoic acid synergistically induce regulatory dendritic cells expressing Aldh1a2. J Immunol. (2013) 191:3139–51. doi: 10.4049/jimmunol.1300329
42. Kralova J, Fabisik M, Pokorna J, Skopcova T, Malissen B, Brdicka T. The transmembrane adaptor protein SCIMP facilitates sustained dectin-1 signaling in dendritic cells. J Biol Chem. (2016) 291:16530–40. doi: 10.1074/jbc.M116.717157
43. Boulet S, Daudelin JF, Odagiu L, Pelletier AN, Yun TJ, Lesage S, et al. The orphan nuclear receptor NR4A3 controls the differentiation of monocyte-derived dendritic cells following microbial stimulation. Proc Natl Acad Sci USA. (2019) 116:15150–9. doi: 10.1073/pnas.1821296116
44. Batissoco AC, Salazar-Silva R, Oiticica J, Bento RF, Mingroni-Netto RC, Haddad LA. A cell junctional protein network associated with connexin-26. Int J Mol Sci. (2018) 19:2535. doi: 10.3390/ijms19092535
45. Moustakas A, Heldin CH. Ecsit-ement on the crossroads of Toll and BMP signal transduction. Genes Dev. (2003) 17:2855–9. doi: 10.1101/gad.1161403
46. Laupeze B, Fardel O, Onno M, Bertho N, Drenou B, Fauchet R, et al. Differential expression of major histocompatibility complex class Ia, Ib and II molecules on monocytes and monocyte-derived dendritic and macrophagic cells. Hum Immunol. (1999) 60:591–7. doi: 10.1016/S0198-8859(99)00025-7
47. Zamani F, Zare Shahneh F, Aghebati-Maleki L, Baradaran B. Induction of CD14 expression and differentiation to monocytes or mature macrophages in promyelocytic cell lines: new approach. Adv Pharm Bull. (2013) 3:329–32. doi: 10.5681/apb.2013.053
48. Cheong C, Matos I, Choi JH, Dandamudi DB, Shrestha E, Longhi MP, et al. Microbial stimulation fully differentiates monocytes to DC-SIGN/CD209(+) dendritic cells for immune T cell areas. Cell. (2010) 143:416–29. doi: 10.1016/j.cell.2010.09.039
49. Hume DA. Macrophages as APC and the dendritic cell myth. J Immunol. (2008) 181:5829–35. doi: 10.4049/jimmunol.181.9.5829
50. Peng WM, Yu CF, Kolanus W, Mazzocca A, Bieber T, Kraft S, et al. Tetraspanins CD9 and CD81 are molecular partners of trimeric FcvarepsilonRI on human antigen-presenting cells. Allergy. (2011) 66:605–11. doi: 10.1111/j.1398-9995.2010.02524.x
51. Mantegazza AR, Barrio MM, Moutel S, Bover L, Weck M, Brossart P, et al. CD63 tetraspanin slows down cell migration and translocates to the endosomal-lysosomal-MIICs route after extracellular stimuli in human immature dendritic cells. Blood. (2004) 104:1183–90. doi: 10.1182/blood-2004-01-0104
52. Banchereau J, Briere F, Caux C, Davoust J, Lebecque S, Liu YJ, et al. Immunobiology of dendritic cells. Annu. Rev. Immunol. (2000) 18:767–811. doi: 10.1146/annurev.immunol.18.1.767
53. Raggi F, Pelassa S, Pierobon D, Penco F, Gattorno M, Novelli F, et al. Regulation of human macrophage M1-M2 polarization balance by hypoxia and the triggering receptor expressed on myeloid cells-1. Front Immunol. (2017) 8:1097. doi: 10.3389/fimmu.2017.01097
54. Xu X, Qiu C, Zhu LY, Huang J, Li LS, Fu WH, et al. IFN-stimulated gene LY6E in monocytes regulates the CD14/TLR4 pathway but inadequately restrains the hyperactivation of monocytes during chronic HIV-1 infection. J Immunol. (2014) 193:4125–36. doi: 10.4049/jimmunol.1401249
55. Deng Y, Govers C, Teodorowicz M, Liobyte I, De Simone I, Hettinga K, et al. Hydrophobicity drives receptor-mediated uptake of heat-processed proteins by THP-1 macrophages and dendritic cells, but not cytokine responses. PLoS ONE. (2020) 15:e0236212. doi: 10.1371/journal.pone.0236212
56. Starr T, Bauler TJ, Malik-Kale P, Steele-Mortimer O. The phorbol 12-myristate-13-acetate differentiation protocol is critical to the interaction of THP-1 macrophages with Salmonella Typhimurium. PLoS ONE. (2018) 13:e0193601. doi: 10.1371/journal.pone.0193601
57. Bauman SK, Huffnagle GB, Murphy JW. Effects of tumor necrosis factor alpha on dendritic cell accumulation in lymph nodes draining the immunization site and the impact on the anticryptococcal cell-mediated immune response. Infect Immun. (2003) 71:68–74. doi: 10.1128/IAI.71.1.68-74.2003
58. Fang HL, Ang B, Xu XY, Huang XH, Wu YF, Sun YP, et al. TLR4 is essential for dendritic cell activation and anti-tumor T-cell response enhancement by DAMPs released from chemically stressed cancer cells. Cell Mol Immunol. (2014) 11:150–9. doi: 10.1038/cmi.2013.59
59. Geijtenbeek TBH, Torensma R, Van Vliet SJ, Van Duijnhoven GCF, Adema GJ, Van Kooyk Y, et al. Identification of DC-SIGN, a novel dendritic cell-specific ICAM-3 receptor that supports primary immune responses. Cell. (2000) 100:575–85. doi: 10.1016/S0092-8674(00)80693-5
60. Guilliams M, Ginhoux F, Jakubzick C, Naik SH, Onai N, Schraml BU, et al. Dendritic cells, monocytes and macrophages: a unified nomenclature based on ontogeny. Nat Rev Immunol. (2014) 14:571–8. doi: 10.1038/nri3712
61. Wright SD, Ramos RA, Tobias PS, Ulevitch RJ, Mathison JC. Cd14, a receptor for complexes of lipopolysaccharide (Lps) and Lps binding-protein. Science. (1990) 249:1431–3. doi: 10.1126/science.1698311
62. Iyer SS, Ghaffari AA, Cheng G. Lipopolysaccharide-mediated IL-10 transcriptional regulation requires sequential induction of type I IFNs and IL-27 in macrophages. J Immunol. (2010) 185:6599–607. doi: 10.4049/jimmunol.1002041
63. Rego D, Kumar A, Nilchi L, Wright K, Huang S, Kozlowski M. IL-6 production is positively regulated by two distinct src homology domain 2-containing tyrosine phosphatase-1 (SHP-1)-dependent CCAAT/enhancer-binding protein beta and NF-kappa B pathways and an SHP-1-independent NF-kappa B pathway in lipopolysaccharide-stimulated bone marrow-derived macrophages. J Immunol. (2011) 186:5443–56. doi: 10.4049/jimmunol.1003551
64. Brix S, Bovetto L, Fritsche R, Barkholt V, Frokiaer H. Immunostimulatory potential of beta-lactoglobulin preparations: Effects caused by endotoxin contamination. J Allergy Clin Immunol. (2003) 112:1216–22. doi: 10.1016/j.jaci.2003.08.047
65. Hilhorst M, Shirai T, Berry G, Goronzy JJ, Weyand CM. T cell-macrophage interactions and granuloma formation in vasculitis. Front Immunol. (2014) 5:432. doi: 10.3389/fimmu.2014.00432
66. Roberts CA, Dickinson AK, Taams LS. The interplay between monocytes/macrophages and CD4(+) T cell subsets in rheumatoid arthritis. Front Immunol. (2015) 6:571. doi: 10.3389/fimmu.2015.00571
67. Berger A. Science commentary: Th1 and Th2 responses: what are they? Br Med J. (2000) 321:424. doi: 10.1136/bmj.321.7258.424
68. Hilmenyuk T, Bellinghausen I, Heydenreich B, Ilchmann A, Toda M, Grabbe S, et al. Effects of glycation of the model food allergen ovalbumin on antigen uptake and presentation by human dendritic cells. Immunology. (2010) 129:437–45. doi: 10.1111/j.1365-2567.2009.03199.x
69. Frei PC, Benacerr B, Thorbeck GJ. Phagocytosis of antigen a crucial step in induction of primary response. Proc Natl Acad Sci USA. (1965) 53:20. doi: 10.1073/pnas.53.1.20
Keywords: β-lactoglobulin, THP-1, macrophage, dendritic cell, heat-treatment, microarray, cytokine, ingenuity pathway analysis
Citation: Deng Y, Govers C, Beest Et, van Dijk A-J, Hettinga K and Wichers HJ (2021) A THP-1 Cell Line-Based Exploration of Immune Responses Toward Heat-Treated BLG. Front. Nutr. 7:612397. doi: 10.3389/fnut.2020.612397
Received: 30 September 2020; Accepted: 14 December 2020;
Published: 13 January 2021.
Edited by:
Mauro Serafini, University of Teramo, ItalyReviewed by:
Wayne Robert Thomas, University of Western Australia, AustraliaCopyright © 2021 Deng, Govers, Beest, van Dijk, Hettinga and Wichers. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Harry J. Wichers, aGFycnkud2ljaGVyc0B3dXIubmw=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.