 Gökçe Polat Yemiş1
Gökçe Polat Yemiş1 Pascal Delaquis2*
Pascal Delaquis2*- 1Department of Food Engineering, Sakarya University, Serdivan, Turkey
- 2Summerland Research and Development Research Centre, Agriculture and AgriFood Canada, Summerland, BC, Canada
Bacteria from the genus Cronobacter are opportunistic foodborne pathogens capable of causing severe infections in neonates, the elderly and immunocompromised adults. The majority of neonatal infections have been linked epidemiologically to dehydrated powdered infant formulas (PIFs), the majority of which are manufactured using processes that do not ensure commercial sterility. Unfortunately, the osmotolerance, desiccation resistance, mild thermotolerance and wide-ranging minimum, optimum and maximum growth temperatures of Cronobacter spp. are conducive to survival and/or growth during the processing, reconstitution and storage of reconstituted PIFs. Consequently, considerable research has been directed at the development of alternative strategies for the control of Cronobacter spp. in PIFs, including approaches that employ antimicrobial compounds derived from natural sources. The latter include a range of phytochemicals ranging from crude extracts or essential oils derived from various plants (e.g., thyme, cinnamon, clove, marjoram, cumin, mint, fennel), to complex polyphenolic extracts (e.g., muscadine seed, pomegranate peel, olive oil, and cocoa powder extracts), purified simple phenolic compounds (e.g., carvacrol, citral, thymol, eugenol, diacetyl, vanillin, cinnamic acid, trans-cinnamaldehyde, ferulic acid), and medium chain fatty acids (monocaprylin, caprylic acid). Antimicrobials derived from microbial sources (e.g., nisin, other antibacterial peptides, organic acids, coenzyme Q0) and animal sources (e.g., chitosan, lactoferrin, antibacterial peptides from milk) have also been shown to exhibit antibacterial activity against the species. The selection of antimicrobials for the control of Cronobacter spp. requires an understanding of activity at different temperatures, knowledge about their mode of action, and careful consideration for toxicological and nutritional effects on neonates. Consequently, the purpose of the present review is to provide a comprehensive summary of currently available data pertaining to the antibacterial effects of natural antimicrobial compounds against Cronobacter spp. with a view to provide information needed to inform the selection of compounds suitable for control of the pathogen during the manufacture or preparation of PIFs by end users.
Introduction
Fatal bacterial infections in neonates caused by “yellow-pigmented coliforms” were first reported in the early 1960s (1). Early clinical isolates were classified as strains of Enterobacter cloacae until comparative analysis by DNA–DNA hybridization showed they belong to a distinct species that was initially named Enterobacter sakazakii (2). Further genomic analysis by ribotyping, amplified fragment length polymorphism and 16S rDNA sequencing eventually provided evidence to support assignment of E. sakazakii and other closely related Enterobacter species to the novel genus Cronobacter (3). The List of Prokaryotic names with Standing in Nomenclature (LPSN) presently includes seven species of Cronobacter with a validly published and correct name, including Cronobacter sakazakii, Cronobacter malonaticus, Cronobacter universalis, Cronobacter turicensis, Cronobacter muytjensii, Cronobacter dublinensis and Cronobacter condimenti (https://www.bacterio.net/genus/cronobacter). C. sakazakii is the most frequently reported clinical isolate and is considered to be the prototype species for the genus (4, 5). However, all Cronobacter spp. with the exception of C. condimenti have been recovered from clinical specimens (6). C. sakazakii and C. malonaticus are currently the major species of public health concern, followed by C. turicensis, C. universalis, C. muytjensii, and C. dublinensis (5). Cronobacter infections typically affect hosts with immature or compromised immune systems, primarily neonates and infants, and to a lesser extent the elderly or individuals from all age groups with underlying chronic disease. Consequently, Cronobacter spp. are considered opportunistic human pathogens. Infections in neonates can lead to septicemia, necrotizing enterocolitis or severe meningitis with estimated case-fatality rates of 10, 20, and 42%, respectively, and to severe neurological sequelae upon recovery (7–9). Symptoms of infection in adults include wound and urinary tract infections, gastroenteritis, appendicitis, conjunctivitis, biliary sepsis, pneumonia, septicemia, and osteomyelitis (10). Cronobacter infections were considered exceptional and sporadic occurrences until the late 1980s when several clusters were reported in neonatal care units (11). Clinical investigation of a landmark incident in a US hospital showed that neonates were infected by enteral administration of reconstituted powdered infant formula (PIF) (12). Epidemiological investigations of similar incidents in other jurisdictions have confirmed that PIFs can serve as a vehicle for the foodborne transmission of C. sakazakii to neonates (13–15). Only one suspected foodborne outbreak involving ostensibly healthy and older individuals has been reported to date. Yong et al. (16) presented evidence that food contaminated with Cronobacter spp. consumed in a senior high school canteen led to an outbreak of acute gastroenteritis that resulted in 124 suspected, 12 probable, and 20 confirmed cases. Molecular analysis of isolates recovered from clinical, food, or environmental samples revealed the presence of both C. sakazakii (four isolates from two sequence types determined by multilocus sequence typing) and C. malonaticus (two isolates from one sequence type). However, the whole genome sequences of two C. sakazakii isolates recovered from a food sample and a clinical specimen differed by only five single nucleotide polymorphisms, which was highly suggestive of an epidemiological link.
While Cronobacter infections remain uncommon, alarmingly high case-fatality rates in neonates and uncertainty about transmission outside hospital care settings have prompted considerable research to determine the origin, distribution and fate of this emerging foodborne pathogen in food chains. Despite these efforts, the primary habitats of Cronobacter spp. remain unknown. Infrequent isolation from livestock and limited survival in the animal gut are indicative of a non-zoonotic nature, although contamination of meat and milk have been reported (17–19). A review and meta-analysis of data published between 2008–14 revealed a prevalence of 5.7% in meat products and 19.0% in plant based foods or food ingredients, which is suggestive of a stronger association with plants or environments in which they are grown (20). Irrespective of their primary habitat, Cronobacter spp. have been isolated from diverse dehydrated food products (PIFs, infant cereals, dairy-based preparations, flours, pasta, candies, spices, herbs, and nuts), fresh or frozen vegetables, and both natural (soil, water, insects) and man-made (hospitals, households, food storage and processing facilities) environments (20–23).
Current understanding about the fate of Cronobacter spp. in food systems is primarily derived from research concerned with the role of PIFs in foodborne transmission. PIFs intended to serve as complete or partial substitutes for human milk at birth or after the introduction of solid food (follow-up formulas) contain mixtures of protein, fat, carbohydrates, vitamins, minerals, and other functional ingredients (e.g., essential fatty acids, nucleotides) in proportions needed to achieve nutrient contents mandated by national or international regulatory standards. Intact bovine milk powder is the most common source of protein, although specialized formulas containing hydrolyzed casein or proteins derived from plant sources such as soy bean are used for feeding of neonates with underlying pathologies or to accommodate cultural or religious practices (24). Manufacture of PIF products is accomplished by spray-drying of the mixed ingredients solubilized in water (wet processes), by mixing of heat-labile ingredients with a previously spray-dried base powder (dry processes), or by a combination of both approaches (25). Despite the application of heat at one or more stages of these processes PIF is not a sterile food; Cronobacter spp. are routinely detected in microbiological analysis of commercial products (26, 27). For example, a recent survey of 128 products in Latin American markets revealed a prevalence rate of 4.7% (28). Microbiological assessments of milk powder and PIF manufacturing environments and processes have shown that Cronobacter spp. may derive from extrinsic sources, notably dry ingredients, or intrinsically contaminated sites where specific strains may persist over long periods of time (29–33). Moreover, most strains examined to date have shown higher resistance to potentially lethal osmotic and dessication stresses than other human pathogen belonging to the family Enterobacteriaceae. Dessication resistance likely contributes to the environmental persistence of Cronobacter spp. in some niches within manufacturing plants, and to long term survival in powdered milk and PIF (34–36).
The suspected role of PIF in the transmission of infections has prompted examination of Cronobacter behavior during reconstitution in water and subsequent handling, including storage for later use. Incipient work by Nazarowec-White and Farber (37) showed that C. sakazakii could survive reconstitution with water heated to 52–60°C. Data from additional studies conducted with numerous strains over a wider range of temperatures and in different substrates suggests that most Cronobacter spp. are mildly thermotolerant, although strain-associated stress tolerance or prior heat adaptation can enhance thermal stability (38–41). In response to the risk implied by latent contamination of PIFs with infectious bacteria the WHO recommends reconstitution in water at a minimum temperature of 70°C, conditions which have been shown to reduce C. sakazakii by >5 log10 cycles (42, 43). Reconstitution at ≥70°C is often impractical, however, as high temperatures can lead to curdling or other undesirable organoleptic changes, cause depletion of heat sensitive nutrients, and introduce scald or burn hazards particularly in home settings (27, 44). Consequently, lower temperatures are endorsed in some jurisdictions despite experimental evidence of limited thermal inactivation at lower temperatures. Moreover, growth of C. sakazakii has been reported to occur in reconstituted PIF stored between 5.5 and 47°C, conditions that can occur when feeding is delayed or during storage (15, 21, 41). Additionally, recent work has shown that C. sakazakii is readily transferred from caregiver hands and utensils to reconstituted PIF, thereby highlighting the significance of contact surfaces as reservoirs of contamination (45). Adherence to and biofilm formation have been demonstrated on a wide range of materials used in the manufacture of equipment, tools and utensils used in hospital and home settings (46–48). Biofilm formation also contributes to the persistence of Cronobacter on surfaces by enhancing resistance to adverse environmental stresses, including chemicals used in cleaning and sanitation of food processing facilities and equipment (49).
The risk of contamination with Cronobacter spp. is an enduring food safety challenge for the PIF industry, public health authorities and consumers worldwide. Manufacturers have adopted risk mitigation strategies that primarily rely on rigorous microbiological analysis of raw materials, improved cleaning and sanitation of the manufacturing environment, and enhanced testing of finished products. Despite these efforts, levels of contamination detected through recent surveys clearly show that the risk persists (28). Because the modification of existing industrial processes is constrained by the heat lability of PIF ingredients, alternative non-thermal physical treatments meant to inactivate microbial contaminants without affecting ingredient stability have been investigated or are under study (reviewed in (50, 51)). To date, none have been adapted to the production of PIF on an commercial scale. The use of synthetic preservatives is likewise impractical due to regulatory restrictions and enduring concerns about the negative effects of man-made food additives on human health. Accordingly, natural antibacterial compounds (NACs) derived from plant, microbial or animal sources are under consideration as alternatives to synthetic chemical preservatives for the control of Cronobacter spp. in PIF. This approach is aligned with increasing consumer willingness to accept food additives and preservatives of natural origin over synthetic products (52). The present work is intended to provide a summary of current knowledge about NACs with antibacterial activity against Cronobacter spp., with a view to inform their application in the development of improved PIF manufacturing processes or the formulation of safer products. For example, NACs that increase the lethality of mild heat may find value in the development of alternative processes to enhance bacterial inactivation in manufacture or during reconstitution by end-users (53). Where possible, the mode of action, toxicological data, regulatory status and potential health benefits of specific NACs are provided.
NACs From Plants With Antibacterial Activity Against Cronobacter Spp.
The scope of research on plants as sources of natural antimicrobials and their applications in food preservation or safety has expanded significantly in recent years (54). Plant components and extractives thereof are attractive alternative food additives because many have a long history of use, are likely to have received regulatory consent or benefit from the increasing availability of toxicological data needed to support requests for approval (55). Recognition of PIF as a vector for foodborne transmission has led to the assessment of numerous crude extracts, essential oils recovered by distillation of whole plants or their parts, and purified phytochemicals for the control of Cronobacter spp. (Table 1). Crude extracts obtained in water or an alcoholic solvent are generally not modified after evaporation of the liquid phase and contain several bioactive constituents. For example, a tea extract examined by Li et al. (61) was reported to contain catechins, flavonoids, phenolic acids, anthocyanins, malic acid, and citric acid. The latter is typical of crude plant extracts which tend to consist of complex mixtures of compounds from different classes, notably large and diverse fractions of phenolic compounds referred to collectively as polyphenolics. Most crude extracts listed in Table 1 contain polyphenolics known to have antibacterial properties, although additive or synergistic effects due to co-extracted plant constituents, notably organic acids, likely contribute to overall antibacterial activity (57). The mode of action of crude extracts is accordingly complex, however damage to the bacterial membrane leading to loss of function and cellular integrity induced by polyphenolics contributes significantly to overall antibacterial effects (83). Two extracts (cocoa powder and polyphenolic tea extract) were shown to exert bacteriostatic effects and reduce the growth of Cronobacter spp. in reconstituted PIF. Crude plant extracts are economically attractive food additives due to their low cost of production in comparison purified phytochemicals. However, variability in antibacterial activity due to differences in composition resulting from varietal, agronomic or production factors tend to limit practical applications in foods. Concentrations of compounds with antibacterial activity may also be lower than those of co-extracted compounds that can lend undesirable physico-chemical or organoleptic properties to target food products. Similar constraints hamper food applications for essential oils, complex mixtures of volatile lipophilic terpenoids, phenylpropanoids, or short-chain aliphatic hydrocarbon derivatives (84). Results from four scientific reports listed in Table 1 show that essential oils recovered from several aromatic plant species show antibacterial activity against Cronobacter spp. Moreover, Al-Nabulsi et al. (65) found that growth of C. sakazakii in reconstituted PIF was inhibited by supplementation with cinnamon or fir essential oils and that mixtures of the two were bactericidal, reducing populations by >6 log10 after 3 h of incubation at 37°C. These results support the widely held view that the activity of essential oils stems from additive or synergistic antimicrobial effects involving multiple components, and underscores the need to determine concentrations of key active compounds needed to ensure consistent activity in foods (85).
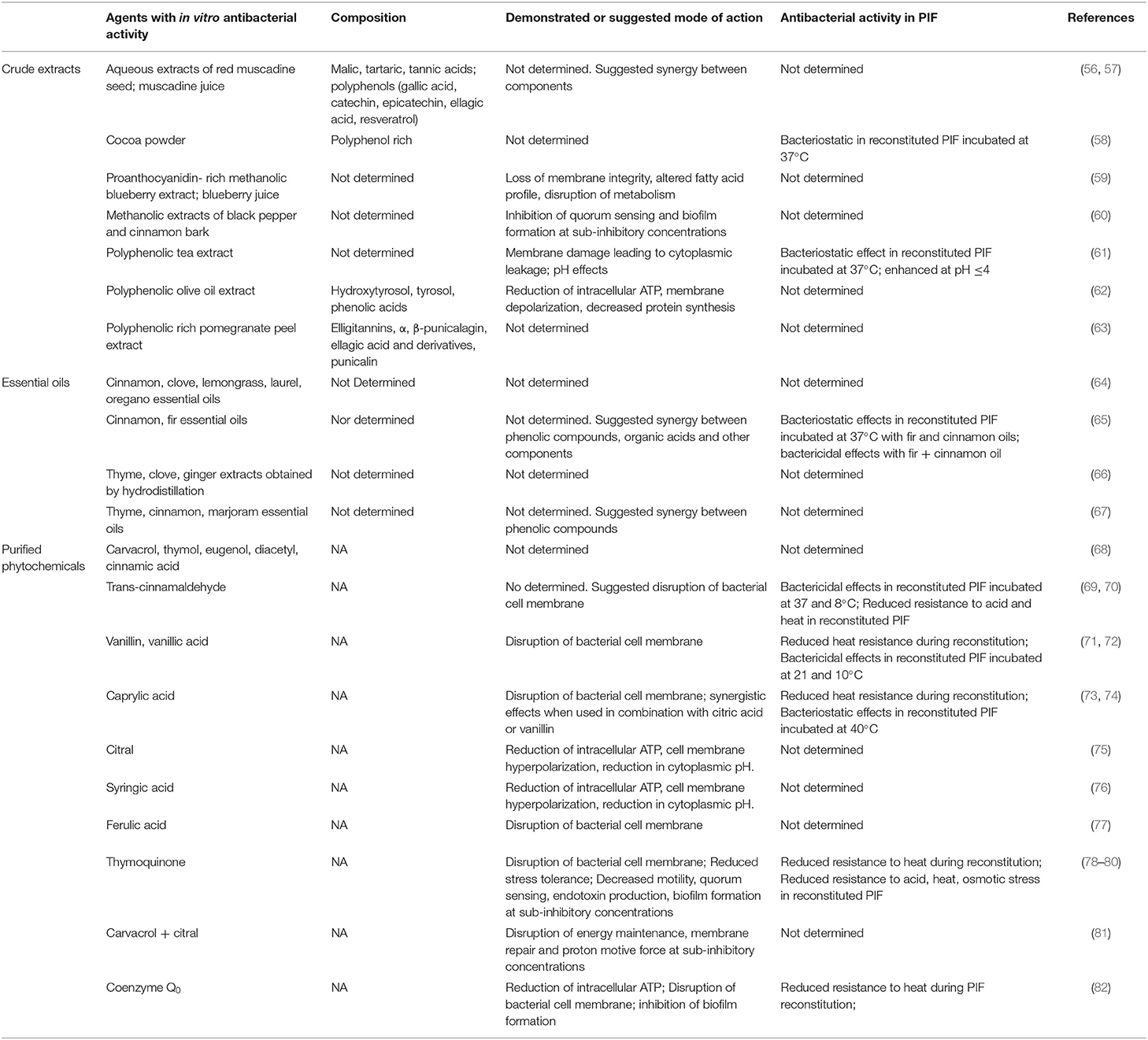
Table 1. Plant-derived crude extracts, essential oils, and purified phytochemicals with antibacterial activity against Cronobacter spp.
Extraction and purification of NACs from plants circumvents challenges occasioned by variable crude extract or essential oil composition. Purified preparations of the phytochemicals listed in Table 1 are readily available from commercial sources. Most are known to exert antimicrobial effects primarily by disruption of the bacterial cell membrane. Trans-cinnamaldehyde, vanillin, ethyl vanillin, vanillic acid, thymoquinone, and Coenzyme Q0 have been shown to reduce the thermotolerance of Cronobacter spp. during reconstitution and to provide antibacterial effects during subsequent storage. Overall, the reported nature and magnitude of measured effects vary with type of compound, level of PIF supplementation and temperature. For example, Amalaradjou et al. (69) reported that trans-cinnamaldehyde exerts time- and temperature-dependent bactericidal activity against C. sakazakii in reconstituted PIF during storage at 4, 8, 23, and 37°C. In contrast, supplementation with caprylic acid could only elicit bacteriostatic effects against the same species in reconstituted PIF stored at 40°C (74). The effects of these compounds on the thermotolerance of Cronobacter are more consistent. Amalaradjou et al. (70) showed that complete thermal inactivation of C. sakazakii could be achieved by heating reconstituted PIF supplemented with 70 μM trans-cinnamaldehyde at 60°C for 10 min, but that longer heating times were required to achieved the same effect at lower supplementation levels or temperatures. Likewise, C. sakazakii was completely inactivated by heating reconstituted PIF supplemented with 30 mmol l−1 thymoquinone at 55°C for 10 min, and longer treatment times were needed to achieved the same effect at lower concentrations or temperatures (78). Caprylic acid (73), coenzyme Q0 (82), vanillin, ethyl vanillin, and vanillic acid (72) have also been found to lower the thermotolerance of C. sakazakii during reconstitution. Moreover, the bactericidal activity of vanillin, ethyl vanillin, and vanillic acid were sustained during subsequent storage at 21 and 5°C, which illustrates that some phytochemical compounds can contribute antibacterial effects at multiple stages during the manufacture or preparation of PIF by end-users.
The volatility, susceptibility to conversion and degradation reactions, intense organoleptic character and poor solubility of many plant extracts, essentials oils or phytochemicals have long hindered wider use in food preservation. However, considerable progress has been achieved in the development of novel technologies for the delivery of food additives that are intended to overcome these limitations, notably encapsulation methods that stabilize active components and enable their release of over variable time periods, at specific temperatures or at different sites within food matrices (reviewed in (86)). To date, the merit of these technologies for the delivery of NACs for the control of Cronobacter in PIF remains unexplored. Encapsulation platforms that provide controlled release at temperatures applied during the manufacture, preparation or storage of reconstituted PIF could provide the means to ensure delivery when contaminants are most vulnerable to their effects or when the risk of proliferation is highest.
NACs From Microbial Sources With Antibacterial Activity Against Cronobacter Spp.
Several NACs from microbial sources and their antibacterial activity against Cronobacter spp. are shown in Table 2. Probiotics, live microorganisms which when administered in adequate amounts confer a health benefit on the host, are presently used in some countries for pediatric care. Commercial PIFs products containing live preparations of Lactobacillus or Bifidobacterium spp. are available in the marketplace but the effect of probiotic supplementation on the behavior of Cronobacter spp. during or after reconstitution is unknown. The use probiotic bacteria for the explicit control of Cronobacter spp. infections was investigated by Collado et al. (93) who showed that species of Lactobacillus, Bifidobacterium and Streptococcus competitively excluded, inhibited and displaced C. sakazakii in a human intestinal model system. Despite evidence of their antibacterial properties, there have been few additional attempts to exploit the use of live probiotic bacteria for the control of human pathogens in PIF, likely in response to on-going controversy about the efficacy, safety, variability, quality, labeling, and lack of standards for the use of probiotic products destined for neonatal care (94, 95). NACs derived from microbial sources continue to attract interest, however, notably whole inactivated cells or crude cell extracts that retain bioactivity, which have been termed “parabiotics” (96), or purified soluble factors (products or metabolic byproducts) secreted by live microorganisms or released after lysis, which have been described as “postbiotics” (97). Hayes et al. (87) showed that addition of a crude cell-free extract prepared from a caseinate medium fermented by the common probiotic bacterium Lactobacillus acidophilus could inhibit C. sakazakii in reconstituted PIF. Antimicrobial effects were attributed to antimicrobial peptides (caseicin A & B) derived from the degradation of bovine casein by microbial enzymes. Charchoghlyan et al. (89) found that a purified heat inactivated aqueous extract of skim milk fermented with a commercial probiotic strain of Lactobacillus acidophilus used to supplement PIF also inactivated C. sakazakii in the reconstituted product stored at 37°C. The composition of the extract was not provided by the authors of the study, who offered that acids released during fermentation were likely responsible for antibacterial effects. A similar conclusion was reached by Kim et al. (90) in reference to the mechanism responsible for inhibition of C. sakazakii by cell free supernatants of laboratory grown cultures of Lactobacillus kefiri and Lactobacillus kefiranofaciens isolated from kefir, and a commercial probiotic strain of Bifidobacterium longum. It must be noted here that current international (e.g., CODEX STAN 72-1981) or national standards for ingredients or additives permissible in PIF do not include D-lactic acid, a metabolic by-product released during fermentation by L. acidophilus, L., kefiri, and L. kefiranofaciens. Probiotic bacteria such as Bifidobacterium longum that do not produce D-lactic acid could be used to circumvent the problem. However, uncertainty concerning the composition and variability in bioactive components in crude cell-free extracts derived from any microbial species is a barrier to broader food applications. The problem can be avoided by the isolation, purification and characterization of antimicrobial compounds that accumulate in growth media. For example, the examination of cell-free supernatants from laboratory grown cultures of L. acidophilus and L. casei by Awaisheh et al. (88) revealed that both species produce heat-labile bacteriocins, proteinaceous or peptidic molecules with antibacterial activity against C. sakazakii. To date, nisin is the only antibacterial peptide to receive close scrutiny for enhancement of PIF safety. Nisin, a polycyclic peptide bacteriocin produced by Lactococcus lactis, is presently permitted as a food additive in over 50 countries, primarily for the extension of shelf-life or the prevention of quality defects in dairy products (98). The antibacterial activity of nisin is mainly due to depolarizing effects and consequent disruption of the cytoplasmic cell membrane. Gram-positive bacteria are more resistant to this effect than Gram-negative bacteria in which the cell membrane is surrounded by a lipopolysaccharide outer membrane. Consequently, means to destabilize and permeabilize the outer membrane by physical (mild heat, sonication) or chemical means (metal chelators, EDTA, disodium pyrophosphate, sodium hydrogen orthophosphate, citric acid, lactic acid) are often used in conjunction with nisin to improve antibacterial activity against Gram-negative bacteria such as Cronobacter spp. Al-Nabulsi et al. (99) found that reconstitution of PIF at 55°C did not significantly improve the antibacterial activity of nisin against 5 Cronobacter strains. In contrast, Campion et al. (92) observed strong bactericidal activity against C. sakazakii in reconstituted PIF supplemented with a commercial nisin preparation (Nisaplin) and citric acid, a food additive permitted in infant formula. This finding suggests that antibacterial strategies based on synergistic effects between nisin and additives compliant with regulatory standards for PIF merit further investigation. Corresponding efforts should be directed at the assessment of alternative sources of antimicrobial peptides, including other commercial bacteriocin products or the array of peptides derived from probiotics or bacterial starter cultures described in the scientific literature (100).

Table 2. Natural antimicrobials from microbial sources with antibacterial activity against Cronobacter spp.
NACs From Animal Sources With Antibacterial Activity Against Cronobacter Spp.
Selected NACs from animal sources and their antibacterial activity against Cronobacter spp. are shown in Table 3. The first natural antimicrobial derived from animal sources considered for the control of Cronobacter spp. in PIF was lactoperoxidase, an enzyme that occurs in milk, colostrum, tears, saliva, and other mammalian secretions (101). Lactoperoxidase catalyzes the oxidation of thiocyanate to hypothiocyanous acid and hypothiocyanate by H2O2 and generates intermediate products with antimicrobial properties. The “lactoperoxidase (LPO) system” relies on the interaction of all three components which must be present in sufficient amounts to initiate the reaction. Gurtler and Beuchat (101) showed that addition of bovine LPO to reconstituted milk-based PIF could inhibit the growth of C. sakazakii at temperatures >21°C. However, use of the LPO system in PIF is hindered by the need for addition of an exogenous source of thiocyanate, a known goitrogen (106). Another antibacterial protein also found in mammalian secretory fluids, lactoferrin, is not bound by this restriction. Lactoferrin, a small glycoprotein, inhibits bacteria indirectly by the sequestration of iron from the environment and through direct antibacterial effects resulting from disruption of the outer Gram-negative bacterial membrane, leading to alterations in cell permeability and loss of viability (107). Indirect antibacterial effects mediated by alteration of host innate immune functions have also been reported (108). Bovine milk is the most common source of lactoferrin and several manufacturers provide purified preparations for use in pharmaceutical, cosmetic and food applications, including the supplementation of PIF. Usage of lactoferrin supplemented PIF is common in some countries for the prevention of neonatal sepsis and necrotizing enterocolitis despite continued uncertainty about efficacy (109). Experimentation in vitro has shown that Cronobacter spp. are highly susceptible to the direct antibacterial effects of lactoferrin (99). However, the same authors found no evidence of antibacterial effects in reconstituted PIF during storage at 10, 21, or 37°C, a result ascribed to interactions with food components that reduced the activity of lactoferrin, notably the divalent cations Ca2+, Mg2+, and Fe3+. Harouna et al. (102) attempted to improve the activity of lactoferrin by saturation with iron cations but the saturated form of the protein had no measurable antibacterial effect against C. sakzakii. Numerous novel antimicrobial peptides with enhanced antimicrobial activity have been synthesized through chemical or enzymatic hydrolysis of lactoferrin (110). Harouna et al. (103) prepared lactoferrin hydrolysates using pepsin, chymosin and microbial rennet that exhibited enhanced antibacterial activity against C. sakazakii in a microbiological medium. None were effective in reconstituted PIF at 37°C, however, which provided further evidence that PIF ingredients interfere with the activity of proteic or peptidic antimicrobials. The majority of NACs from animal sources considered for the enhancement of PIF safety have been derived from bovine milk. Antimicrobial peptides have been detected in the milk of other food animal species including sheep and goats (111). A recent report that growth of C. sakazakii is inhibited by strong, inherent antibacterial factors in camel milk suggested they are likely derived from the protein component (105). A database assembled by Wang et al. (112) lists 1,972 known antimicrobial peptides from animal sources, in addition to 321 from plants and many from fungi, protists or other life forms. One antimicrobial peptide from a non-bovine source, bicarinalin from ants, exhibited stronger bactericidal effects against C. sakazakii than ampicillin and tetracycline (104). Clearly, animals are a rich and largely untapped source of NACs that could find value in the enhancement of PIF safety.
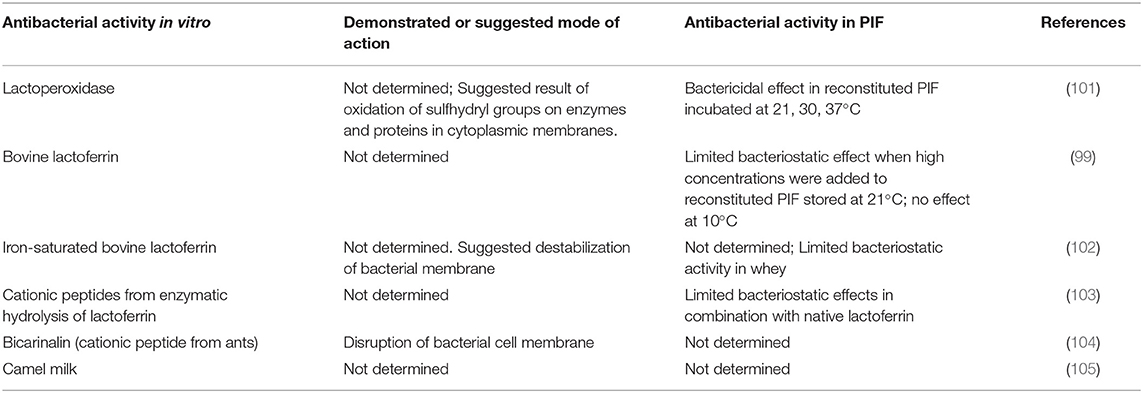
Table 3. Natural antimicrobials from animal sources with antibacterial activity against Cronobacter spp.
Nutritional, Toxicological, and Regulatory Considerations
Commercial PIF products are subject to regulations and regulatory oversight administered by national governments. All are based on the Codex Alimentarius “Standard for Infant Formula and Formulas for Special Medical Purposes Intended for Infants (CODEX STAN 72-1981),” which provides recommendations regarding the essential composition, nutritional quality and additives in formula. CODEX recommendations are adopted by national governments in various ways, but there is general consistency in the application of compositional and nutritional quality standards for PIF across the globe. In contrast, some disparities exist with respect to the use of additives due to variable legislative, legal or regulatory frameworks within different jurisdictions. In the United States, food ingredients are subject to provisions in the Code of Federal Regulations. Ingredients that are not listed in the Code can obtain Generally Recognized as Safe (GRAS) designation through a notification program which requires petitioners to provide historical and scientific evidence that a substance added to food is considered safe and suitable for exemption from the food additive tolerance requirements of the United States Food and Drug Administration. The GRAS status of selected NACs with antibacterial activity against Cronobacter spp. in PIF is shown in Table 4, along with health benefits that have been ascribed to them and pertinent toxicological data. The authors recognize that lack of knowledge about the metabolism of food additives old or new, difficulties in determining accurate levels of exposure, and susceptibility to toxicity have historically hindered safety assessments in the neonatal context (150). Likewise, the purported health benefits noted in Table 4 are largely derived from animal studies or human feeding trials conducted with children or adults, and extrapolation of results to the neonatal situation is challenging.
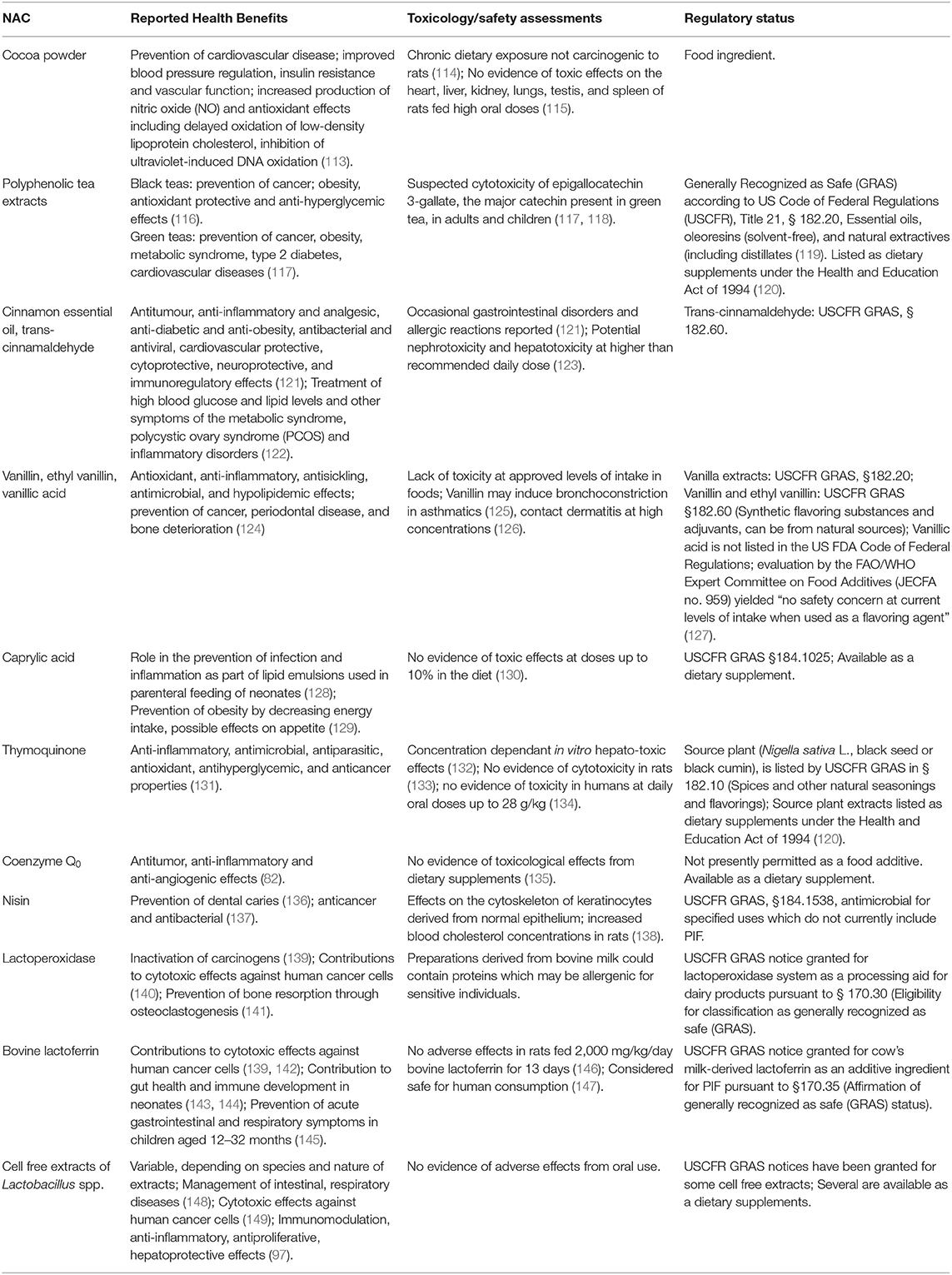
Table 4. Suggested health-promoting effects, toxicological data, safety assessments and regulatory status of selected natural antimicrobial compounds with antibacterial activity against Cronobacter spp. in PIF.
All NACs from plant sources with antibacterial activity in PIF listed in Table 4, with the exception of caprylic acid, are either purified phenolic compounds or contain high concentrations thereof. Vanillin, ethyl vanillin, and trans-cinnamaldehyde have GRAS status, and are widely consumed in food and nutritional supplements. Several vanilla-flavored PIF products containing natural and artificial flavors were once available in the marketplace but have since been withdrawn, although vanilla flavored “toddler” formula is still sold in North America. Whether the level of vanillin or ethyl vanillin supplementation used in these products was sufficient to achieve antibacterial effects against Cronobacter during PIF reconstitution or storage, as reported by Polat Yemiş et al. (72) is unknown. Supplementation of PIF with cocoa powder was also discontinued several years ago. The antibacterial activity of cocoa powder against C. sakzakii in reconstituted PIF described by Pina-Pérez et al. (58) was likely derived from polyphenols, many of which are known to interact with and damage the bacterial cell membrane (83). Polyphenols encompass several groups of compounds including phenolic acids and flavonoids, primarily flavanols present as monomeric epicatechin, catechin, and their oligomers referred to as proanthocyanidins. Research on the nutritional effects of moderate cocoa consumption suggests that the benefits likely outweigh the risks, and that beneficial effects on health are primarily derived from flavanol-mediated protection against oxidative insult by the modulation of oxygen radical generation and antioxidant enzyme and non-enzyme defenses (151, 152). There is compelling evidence that most polyphenols are largely beneficial to human health, principally for the prevention and management of chronic diseases (153). On the other hand, the pharmacological properties of some polyphenols introduce concerns about their safety in products intended for use by infants. Isoflavones (genistein, daidzein, and glycitein) derived from soya beans are known activators of estrogen receptors with demonstrable effects on reproductive and endocrine functions in animal models (154). All soy-protein based PIFs contain isoflavones, mainly genistein, but no clear consensus has emerged regarding the short or long term implications of long-term dietary exposure on the development of infants (155, 156). In contrast, consumption of the medium length straight chain fatty acid caprylic acid (octanoic acid) is considered to be comparatively free of toxicological risk (130). Caprylic acid is found naturally in the milk of mammals including humans (157) and in infant formulas as part of the medium chain triglyceride component contributed by vegetable fat, or increasingly bovine milk fat (158). Choi et al. (74) showed that low concentrations of caprylic acid in conjunction with citric acid completely inactivated C. sakazakii in PIF during reconstitution at the relatively low temperature of 45°C. A GRAS compound, it is used as an additive in a range of foods as an adjuvant or, interestingly, as a flavoring agent despite having an odor described as “slightly unpleasant and rancid-like.” Similarly, the monoterpene diketone thymoquinone, also a GRAS compound, could inactivate C. sakazakii during reconstitution (78) but it has a bitter taste and a “pencil-like” odor (159). The ubiquinone coenzyme Q0 is the only ordorless and tasteless NAC from non-microbial or animal sources identified to date with antibacterial activity against Cronobacter spp. Coenzyme Q0 extracted from the AC mushroom (Antrodia cinnamomea), a parasitic fungus that grows of the camphor tree, has a long history of use in traditional medicine but has only recently been considered for food applications. Toxicological assessment of supplements prepared from the fungus suggest they are safe for human consumption (132). However, toxicological assessments of the purified compound are lacking and coenzyme Q0 does not currently have GRAS status.
The chemistry, biology, toxicology, pharmacokinetic properties, and functionality of nisin as a food preservative have been extensively investigated. A recent reassessment of toxicological data by the European Food Safety Authority (EFSA) reaffirmed the safety of nisin as a food additive (160). Activity against Cronobacter spp. in PIF relies on synergism with citric acid (92), but the latter is a permitted additive. Hence, there appear to be few regulatory impediments to the use of nisin in foods destined for infants. As noted above, antibacterial activity of lactoperoxidase, another GRAS food additive that is used worldwide for milk preservation, is dependent on a source of thiocyanate, which Gurtler and Beuchat (101) provided exogenously in the form of sodium thiocyanate. Thiocyanates are ubiquitous in food products, however, and it is unfortunate that no attempt was made to determine if endogenous levels could have sustained the reaction. Evidence in support of this presumption was provided by Banks and Board (161) who found that lactoperoxidase catalyzed degradation of endogenous thiocyanates reduced the growth of Enterococcus, Pseudomonas spp. and Enterobacteriaceae in reconstituted PIF stored at 30°C for 48 h, which coincided with the depletion of free SCN- ions. These observations suggest that the value of lactoperoxidase for the control of Cronobacter spp. in PIF merits further investigation. There are also few regulatory obstacles to the application of lactoferrin in PIF since it is already in use for therapeutic purposes, disease prevention or health promotion in neonates (82, 143, 162), and is available in highly purified forms safe for use in infant foods (163). Cell-free extracts derived from microbial cultures present greater regulatory challenges as noted above due to the multiplicity of bioactive compounds and variable composition of extracts which add complexity to toxicological assessments. In this context, the selection of candidate microorganisms for the production of cell free-extracts among those already permitted as probiotics in PIF or that are considered GRAS on the basis of historical, safe use in food fermentations is highly advisable.
Conclusions and Future Prospects
Societal concerns and regulatory response to the risk of exposure to harmful food chemicals in early life provide strong impetus to pursue the search for alternatives. Research on NACs with antibacterial activity against Cronobacter spp. has shown that several could find value in the control of this hazardous pathogen in PIF. However, technological obstacles to practical applications persist and means to overcome them must be the focus of future research in the field. The delivery of NACs with strong antibacterial activity to food systems is often hampered by limited solubility in aqueous matrices, instability, reactions with other food components or adverse effects on sensory properties. For example, the low solubility, volatility, intense sensory characteristics, and reactivity of phenolic compounds and essential oils often hinders their incorporation in foods. Adverse effects on the sensory quality of PIF are a notable concern in light of evidence that exposure to flavors modulates neonatal feeding behavior and food acceptability and choice later in life (164, 165). Recent progress in the use of biopolymers from natural sources for the design of innovative encapsulation systems that provide means to deliver effective yet reduced doses of antibacterial agents, protect active ingredients from undesirable reactions, and provide controlled, quantitative release into food matrices will undoubtedly promote the development of delivery strategies that overcome constraints on both the choice and application of NACs (166, 167). The selection of suitable NACs must also be guided by careful consideration of the regulatory framework governing PIF composition, specifically the use of additives. In the near term, single compounds for which nutritional and toxicological data are available should take preference over preparations likely to contain multiple bioactive compounds. The latter remain eminently worthy of study, however, as possible sources of novel NACs. As a final note, it must be emphasized that the NACs described in the present work were derived from a limited number of animal, plant and bacterial species. Future efforts should be directed at the assessment of additional sources of NACs for the control of Cronobacter spp., such as mushroom species known to contain compounds with antibacterial activity against other foodborne pathogens (168).
Author Contributions
GP and PD equally contributed to a review of the scientific literature, collection of relevant references, writing, and editing of the manuscript. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Urmenyi AM, White-Franklin A. Neonatal death from pigmented coliform infection. Lancet. (1961) 1:313–15. doi: 10.1016/S0140-6736(61)91481-7
2. Farmer JJ III, Asbury MA, Hickman FW, Brenner DJ, the Enterobacteriaceae Study Group. Enterobacter sakazakii: a new species of “Enterobacteriaceae” isolated from clinical specimens. Int J Syst Bacteriol. (1980) 30:569–84. doi: 10.1099/00207713-30-3-569
3. Iversen C, Lehner A, Mullane N, Bidlasm E, Cleenwerck I, Marugg J, et al. The taxonomy of Enterobacter sakazakii: proposal of a new genus Cronobacter gen. nov. and descriptions of Cronobacter sakazakii comb. nov. Cronobacter sakazakii subsp. sakazakii, comb. nov., Cronobacter sakazakii subsp. malonaticus subsp. nov., Cronobacter turicensis sp. nov., Cronobacter muytjensii sp. nov., Cronobacter dublinensis sp. nov. and Cronobacter genomospecies 1. BMC Evol Biol. (2007) 7:64. doi: 10.1186/1471-2148-7-64
4. Feeney A, Kropp KA, O'Connor R, Sleator RD. Cronobacter sakazakii: stress survival and virulence potential in an opportunistic foodborne pathogen. Gut Microbes. (2014) 5:711–18. doi: 10.4161/19490976.2014.983774
5. Henry M, Fouladkhah A. Outbreak history, biofilm formation, and preventive measures for control of Cronobacter sakazakii in infant formula and infant care settings. Microorganisms. (2019) 7:1–10. doi: 10.3390/microorganisms7030077
6. Jang H, Gopinath GR, Eshwar A, Srikumar S, Nguyen S, Gangiredla J. The secretion of toxins and other exoproteins of Cronobacter: role in virulence, adaption, and persistence. Microorganisms. (2020) 8:229. doi: 10.3390/microorganisms8020229
7. Healy B, Cooney S, O'Brien S, Iversen C, Whyte P, Nally J, Callanan JJ, Fanning S. Cronobacter (Enterobacter sakazakii): an opportunistic foodborne pathogen. Foodborne Pathog Dis. (2017) 7:339–50. doi: 10.1089/fpd.2009.0379
8. Friedemann M. Epidemiology of invasive neonatal Cronobacter (Enterobacter sakazakii) infections. Eur J Clin Microbiol Infect Dis. (2009) 28:1297–304. doi: 10.1007/s10096-009-0779-4
9. Forsythe SJ. Updates on the Cronobacter genus. Annu Rev Food Sci Technol. (2017) 9:23–44. doi: 10.1146/annurev-food-030117-012246
10. Lai KK. Enterobacter sakazakii infections among neonates, infants, children and adults: case reports and review of the literature. Medicine. (2001) 80:113–22. doi: 10.1097/00005792-200103000-00004
11. Nazarowec-White M, Farber JM. Enterobacter sakazakii: a review. Int J Food Microbiol. (1997) 34:103–13. doi: 10.1016/S0168-1605(96)01172-5
12. Simmons BP, Gelfand MS, Haas M, Metts L, Ferguson J. Enterobacter sakazakii infections in neonates associated with intrinsic contamination of a powdered infant formula. Infect Control Hosp Epidemiol. (1989) 10:398–401. doi: 10.1086/646060
13. Clark NC, Hill BC, O'Hara CM, Steingrimsson O, Cooksey RC. Epidemiologic typing of Enterobacter sakazakii in two neonatal nosocomial outbreaks. Diagn Microbiol Infect Dis. (1990) 13:467–72. doi: 10.1016/0732-8893(90)90078-A
14. Muytjens HL, Roelofs-Willemse H, Jaspar GH. Quality of powdered substitutes for breast milk with regard to members of the family Enterobacteriaceae. J Clin Microbiol. (1998) 26:743–46. doi: 10.1128/JCM.26.4.743-746.1988
15. Iversen C, Forsythe S. Isolation of Enterobacter sakazakii and other Enterobacteriaceae from powdered infant formula milk and related products. Food Microbiol. (2004) 21:771–6. doi: 10.1016/j.fm.2004.01.009
16. Yong W, Guo B, Shi X, Cheng T, Chen M, Jiang X, et al. An investigation of an acute gastroenteritis outbreak: Cronobacter sakazakii, a potential cause of food-borne illness. Front Microbiol. (2018) 9:2549. doi: 10.3389/fmicb.2018.02549
17. Molloy C, Cagney C, Fanning S, Duffy G. Survival characteristics of Cronobacter spp. in model bovine gut and in the environment. Foodborne Pathog Dis. (2010) 7:671–5. doi: 10.1089/fpd.2009.0449
18. Zeng H, Li C, Ling N, Zhang J, Chen M, Lei T, et al. Prevalence, genetic analysis and CRISPR typing of Cronobacter spp. isolated from meat and meat products in China. Int J Food Microbiol. (2020) 321:108549. doi: 10.1016/j.ijfoodmicro.2020.108549
19. Parra-Flores J, Cerda-Leal F, Contreras A, Valenzuela-Riffo N, Rodríguez A, Aguirre J. Cronobacter sakazakii and microbiological parameters in dairy formulas associated with a food alert in Chile. Front Microbiol. (2018) 9:1708. doi: 10.3389/fmicb.2018.01708
20. Sani NA, Odeyemi OA. Occurrence and prevalence of Cronobacter spp. in plant and animal derived food sources: a systematic review and meta-analysis. SpringerPlus. (2015) 4:545. doi: 10.1186/s40064-015-1324-9
21. Beuchat LR, Kim H, Gurtler JB, Lin LC, Ryu JH, Richards GM. Cronobacter sakazakii in foods and factors affecting its survival, growth, and inactivation. Int J Food Microbiol. (2009) 136:204–13. doi: 10.1016/j.ijfoodmicro.2009.02.029
22. Kandhai MC, Reij MW, Gorris LG, Guillaume-Gentil O, van Schothorst M. Occurrence of Enterobacter sakazakii in food production environments and households. Lancet. (2004) 363:39–40. doi: 10.1016/S0140-6736(03)15169-0
23. Lou X, Liu T, Zhang W, Yu H, Wan H, Song S, et al. The occurrence and distribution characteristics of Cronobacter in diverse cereal kernels, flour, and flour-based products. Food Microbiol. (2019) 84:103269. doi: 10.1016/j.fm.2019.103269
24. Maldonado J, Gil A, Narbona E, Molina JA. Special formulas in infant nutrition: a review. Early Hu Dev. (1998) 53:23–32. doi: 10.1016/S0378-3782(98)00062-0
25. Masum AKM, Chandrapala J, Huppertz T, Adhikari B, Zisu B. Production and characterization of infant milk formula powders: a review. Drying Technol. (2020). doi: 10.1080/07373937.2020.1767645. [Epub ahead of print].
26. World Health Organization. Safe Preparation, Storage and Handling of Powdered Infant Formula Guidelines. Geneva: World Health Organization (2017). Available online at: https://www.who.int/foodsafety/publications/powdered-infant-formula/en/ (accessed October 26, 2020).
27. Kent RM, Fitzgerald GF, Hill C, Stanton C, Ross RP. Novel approaches to improve the intrinsic microbiological safety of powdered infant milk formula. Nutrients. (2015) 7:1217–44. doi: 10.3390/nu7021217
28. Parra-Flores J, Maury-Sintjago E, Rodriguez-Fernández A, Acuña S, Cerda F, Aguirre J, et al. Microbiological quality of powdered infant formula in Latin America. J Food Prot. (2020) 83:534–41. doi: 10.4315/0362-028X.JFP-19-399
29. Reich F, König R, Von Wiese W. Klein G. Prevalence of Cronobacter spp. in a powdered infant formula processing environment. Int J Food Microbiol. (2010) 140:214–7. doi: 10.1016/j.ijfoodmicro.2010.03.031
30. Jacobs C, Braun P, Hammer P. Reservoir and routes of transmission of Enterobacter sakazakii (Cronobacter spp.) in a milk powder-producing plant. J Dairy Sci. (2011) 94:3801–10. doi: 10.3168/jds.2011-4318
31. Yan Q, Power KA, Cooney S, Fox E, Gopinath G, Grim CJ, et al. Complete genome sequence and phenotype microarray analysis of Cronobacter sakazakii SP291: a persistent isolate cultured from a powdered infant formula production facility. Front Microbiol. (2013) 4:256. doi: 10.3389/fmicb.2013.00256
32. Pei X, Li Y, Zhang H, Zhan L, Yu X, Lan G, et al. Surveillance and characterisation of Cronobacter in powdered infant formula processing factories. Food Control. (2019) 96:318–23. doi: 10.1016/j.foodcont.2018.09.009
33. Lu Y, Liu P, Li C, Sha M, Fang J, Gao J, Xu X, Matthews KR. Prevalence and genetic diversity of Cronobacter species isolated from four infant formula production factories in China. Front Microbiol. (2019) 10:1938. doi: 10.3389/fmicb.2019.01938
34. Gurtler JB, Beuchat LR. Survival of Enterobacter sakazakii in powdered infant formula as affected by composition, water activity, and temperature. J Food Prot. (2007) 70:1579–86. doi: 10.4315/0362-028X-70.7.1579
35. Caubilla-Barron J, Forsythe S. Dry stress and survival time of Enterobacter sakazakii and other Enterobacteriaceae. J Food Prot. (2007) 70:2111–7. doi: 10.4315/0362-028X-70.9.2111
36. Breeuwer P, Lardeau A, Peterz M, Joosten HM. Desiccation and heat tolerance of Enterobacter sakazakii. J Appl Microbiol. (2010) 95:967–73. doi: 10.1046/j.1365-2672.2003.02067.x
37. Nazarowec-White M, Farber JM. Thermal resistance of Enterobacter sakazakii in reconstituted dried infant formula. Lett Appl Microbiol. (1997) 24:9–13. doi: 10.1046/j.1472-765X.1997.00328.x
38. Osaili TM, Shaker RR, Al-Haddaq MS, Al-Nabulsi AA, Holley RA. Heat resistance of Cronobacter species (Enterobacter sakazakii) in milk and special feeding formula. J Appl Microbiol. (1989) 107:928–35. doi: 10.1111/j.1365-2672.2009.04271.x
39. Arroyo C, Condon S, Pagan R. Thermobacteriological characterization of Enterobacter sakazakii. Int J Food Microbiol. (2009) 136:110–8. doi: 10.1016/j.ijfoodmicro.2009.09.013
40. Arku B, Fanning S, Jordan K. Heat adaptation and survival of Cronobacter spp. (formerly Enterobacter sakazakii). Foodborne Pathog Dis. (2011) 8:975–81. doi: 10.1089/fpd.2010.0819
41. Huertas JP, Álvarez-Ordóñez A, Morrissey R, Ros-Chumillas M, Esteban MD, Maté J, et al. Heat resistance of Cronobacter sakazakii DPC 6529 and its behavior in reconstituted powdered infant formula. Food Res Int. (2015) 69:401–9. doi: 10.1016/j.foodres.2015.01.010
42. World Health Organization. WHO/FAO Guidelines for the Safe Preparation, Storage and Handling of Powdered Infant Formula. (2006). Available online at: www.ennonline.net/infantformulaguidelines (accessed October 26, 2020).
43. Osaili TM, Al-Nabulsi AA, Shaker RR, Ayyash MM, Olaimat AN, Abu Al-Hasan AS, et al. Effects of extended dry storage of powdered infant milk formula on susceptibility of Enterobacter sakazakii to hot water and ionizing irradiation. J Food Prot. (2008) 71:934–9. doi: 10.4315/0362-028X-71.5.934
44. Turck D. Safety aspects in preparation and handling of infant food. Ann Nutr Metab. (2012) 60:211–4. doi: 10.1159/000338215
45. Cho TJ, Hwang JY, Kim HW, Kim YK, Kwon JI, Kim YJ, et al. Underestimated risks of infantile infectious disease from the caregiver's typical handling practices of infant formula. Sci Rep. (2019) 9:9799. doi: 10.1038/s41598-019-46181-0
46. Iversen C, Lane M, Forsythe SJ. The growth profile, thermotolerance and biofilm formation of Enterobacter sakazakii grown in infant formula milk. Lett Appl Microbiol. (2004) 38:378–82. doi: 10.1111/j.1472-765X.2004.01507.x
47. Lehner A, Riedel K, Eberl L, Breeuwer P, Diep B. Biofilm formation, extracellular polysaccharide production, and cell-to-cell signaling in various Enterobacter sakazakii strains: aspects promoting environmental persistence. J Food Prot. (2005) 6:2287–94. doi: 10.4315/0362-028X-68.11.2287
48. Hurrell E, Kucerova E, Loughlin M, Caubilla-Barron J, Forsythe SJ. Biofilm formation on enteral feeding tubes by Cronobacter sakazakii, Salmonella serovars and other Enterobacteriaceae. Int J Food Microbiol. (2009) 136:227–31. doi: 10.1016/j.ijfoodmicro.2009.08.007
49. Ling N, Forsythe S, Wu Q, Ding Y, Zhang J, Zeng H. Insights into Cronobacter sakazakii biofilm formation and control strategies in the food industry. Engineering. (2020) 6:393–405. doi: 10.1016/j.eng.2020.02.007
50. Pina-Pérez MC, Rodrigo D, Martínez A. Nonthermal inactivation of Cronobacter sakazakii in infant formula milk: a review. Crit Rev Food Sci Nutr. (2016) 56:1620–29. doi: 10.1080/10408398.2013.781991
51. Ahern GJ, Hennessy AA, Ryan CA, Ross R, Stanton C. Advances in infant formula science. Ann Rev Food Sci Technol. (2019) 10:75–102. doi: 10.1146/annurev-food-081318-104308
52. Roman S, Sanchez-Siles LM, Siegrist M. The importance of food naturalness for consumers: results of a systematic review. Trends Food Sci Technol. (2017) 67:44–57. doi: 10.1016/j.tifs.2017.06.010
53. Gurtler JB, Fan X, Jin T, Niemira BA. Influence of antimicrobial agents on the thermal sensitivity of foodborne pathogens: a review. J Food Prot. (2019) 82:628–44. doi: 10.4315/0362-028X.JFP-18-441
54. Quinto EJ, Caro I, Villalobos-Delgado LH, Mateo J, De-Mateo-Silleras B, Redondo-Del-Río MP. Food safety through natural antimicrobials. Antibiotics. (2019) 8:208. doi: 10.3390/antibiotics8040208
55. Tajkarimi M, Ibrahim S, Cliver D. Antimicrobial herb and spice compounds in food. Food Control. (2010) 21:1199. doi: 10.1016/j.foodcont.2010.02.003
56. Silva JL, Weng WL, Chen WW, Corbitt M, Jung YS, Chen YS. Inactivation of Enterobacter sakazakii by water-soluble muscadine seed extracts. Int J Food Microbiol. (2009) 129:3295–9. doi: 10.1016/j.ijfoodmicro.2008.12.014
57. Kim TJ, Weng WL, Silva JL, Jung YS, Marshall D. Identification of natural antimicrobial substances in red muscadine juice against Cronobacter sakazakii. J Food Sci. (2010) 75:150–4. doi: 10.1111/j.1750-3841.2010.01531.x
58. Pina-Pérez MC, Rodrigo D, Martinez A. Bacteriostatic effect of cocoa powder rich in polyphenols to control Cronobacter sakazakii proliferation in infant milk formula. Science and Technology against Microbial Pathogens. Research, Development and Evaluation. In: Proceedings of the International Conference on Antimicrobial research (ICAR 2010). Valladolid: World Scientific Publishing Co. Pte. Ltd (2010). p. 85–8.
59. Joshi SS, Howell AB, D'Souza DH. Cronobacter sakazakii reduction by blueberry proanthocyanidins. Food Microbiol. (2014) 39:127–31. doi: 10.1016/j.fm.2013.11.002
60. Singh N, Patil A, Prabhune A, Goel G. Inhibition of quorum-sensing mediated biofilm formation in Cronobacter sakazakii strains. Microbiology. (2016) 162:1708–14. doi: 10.1099/mic.0.000342
61. Li R, Fei P, Man CX, Lou BB, Niu JT, Feng J, et al. Tea polyphenols inactivate Cronobacter sakazakii isolated from powdered infant formula. J Dairy Sci. (2016) 99:1019–28. doi: 10.3168/jds.2015-10039
62. Fei P, Ali MA, Gong S, Sun Q, Bi X, Liu S, et al. Antimicrobial activity and mechanism of action of olive oil polyphenols extract against Cronobacter sakazakii. Food Control. (2018) 94:289–94. doi: 10.1016/j.foodcont.2018.07.022
63. Polat Yemiş G, Bach S, Delaquis P. Antibacterial activity of polyphenol-rich pomegranate peel extract against Cronobacter sakazakii. Int J Food Prop. (2019) 22:985–93. doi: 10.1080/10942912.2019.1622564
64. Franková A, Marounek M, Mozrová V, Weber J, Klouček P, Lukešová D. Antibacterial activities of plant-derived compounds and essential oils toward Cronobacter sakazakii and Cronobacter malonaticus. Foodborne Pathog Dis. (2014). 11:795–7. doi: 10.1089/fpd.2014.1737
65. Al-Nabulsi AA, Awaisheh SS, Osaili TM, Olaimat AN, Rahahaleh RJ, Al-Dabbas FM, et al. Inactivation of Cronobacter sakazakii in reconstituted infant milk formula by plant essential oils. J Appl Bot Food Qual. (2015) 88:97–101. doi: 10.5073/JABFQ.2015.088.013
66. Abu-Ghazaleh BM. Antimicrobial activity of Thymus vulgaris extract, Syzygium aromaticum extract and Zingiber officinale extract on Cronobacter spp. as compared with common preservatives. Pharmacologyonline Arch. (2019) 2:1–11.
67. Berthold-Pluta A, Stasiak-Rózańska L, Pluta A, Garbowska M. Antibacterial activities of plant-derived compounds and essential oils against Cronobacter strains. Eur Food Res Technol. (2019) 245:1137–47. doi: 10.1007/s00217-018-3218-x
68. Lee SY, Jin HH. Inhibitory activity of natural antimicrobial compounds alone or in combination with nisin against Enterobacter sakazakii. Lett Appl Microbiol. (2008) 47:315–21. doi: 10.1111/j.1472-765X.2008.02432.x
69. Amalaradjou MA, Hoagland TA, Venkitanarayanan K. Inactivation of Enterobacter sakazakii in reconstituted infant formula by trans-cinnamaldehyde. Int J Food Microbiol. (2009) 129:146–9. doi: 10.1016/j.ijfoodmicro.2008.11.016
70. Amalaradjou MAR, Venkitanarayanan K. Effect of trans-cinnamaldehyde on reducing resistance to environmental stresses in Cronobacter sakazakii. Foodborne Pathog Dis. (2011) 8:403–9. doi: 10.1089/fpd.2010.0691
71. Polat Yemiş GP, Pagotto F, Bach S, Delaquis P. Effect of vanillin, ethyl vanillin, and vanillic acid on the growth and heat resistance of Cronobacter species. J Food Prot. (2011) 74:2062–9. doi: 10.4315/0362-028X.JFP-11-230
72. Polat Yemiş GP, Pagotto F, Bach S, Delaquis P. Thermal tolerance and survival of Cronobacter sakazakii in powdered infant formula supplemented with vanillin, ethyl vanillin, and vanillic acid. J Food Sci. (2012) 77:523–7. doi: 10.1111/j.1750-3841.2012.02834.x
73. Jang HI, Rhee MS. Inhibitory effect of caprylic acid and mild heat on Cronobacter spp. (Enterobacter sakazakii) in reconstituted infant formula and determination of injury by flow cytometry. Int J Food Microbiol. (2009) 133:113–20. doi: 10.1016/j.ijfoodmicro.2009.05.009
74. Choi MJ, Kim SA, Lee NY, Rhee MS. New decontamination method based on caprylic acid in combination with citric acid or vanillin for eliminating Cronobacter sakazakii and Salmonella enterica serovar Typhimurium in reconstituted infant formula. Int J Food Microbiol. (2013). 166:499–507. doi: 10.1016/j.ijfoodmicro.2013.08.016
75. Shi C, Song K, Zhang X, Sun Y, Sui Y, Chen Y, et al. Antimicrobial activity and possible mechanism of action of citral against Cronobacter sakazakii. PLoS ONE. (2016) 11:e0159006. doi: 10.1371/journal.pone.0159006
76. Shi C, Sun Y, Zheng Z, Zhang X, Song K, Jia Z, et al. Antimicrobial activity of syringic acid against Cronobacter sakazakii and its effect on cell membrane. Food Chem. (2016) 197:100–6. doi: 10.1016/j.foodchem.2015.10.100
77. Shi C, Zhang X, Sun Y, Yang M, Song K, Zheng Z, et al. Antimicrobial activity of ferulic acid against Cronobacter sakazakii and possible mechanism of action. Foodborne Pathog Dis. (2016) 13:196–204. doi: 10.1089/fpd.2015.1992
78. Shi C, Jia Z, Chen Y, Yang M, Liu X, Sun Y, et al. Inactivation of Cronobacter sakazakii in reconstituted infant formula by combination of thymoquinone and mild heat. J Appl Microbiol. (2015) 119:1700–6. doi: 10.1111/jam.12964
79. Shi C, Yan C, Sui Y, Sun Y, Guo D, Chen Y, et al. Thymoquinone inhibits virulence related traits of Cronobacter sakazakii ATCC 29544 and has anti-biofilm formation potential. Front Microbiol. (2017) 8:2220. doi: 10.3389/fmicb.2017.02220
80. Chen Y, Wen Q, Chen S, Guo D, Xu Y, Liang S, et al. Effect of thymoquinone on the resistance of Cronobacter sakazakii to environmental stresses and antibiotics. Food Control. (2020) 109:106944. doi: 10.1016/j.foodcont.2019.106944
81. Cao Y, Zhoua A, Zhou D, Xiao X, Yua Y, Li X. Cronobacter sakazakii CICC 21544 responds to the combination of carvacrol and citral by regulating proton motive force. LWT Food Sci Technol. (2020) 122:109040. doi: 10.1016/j.lwt.2020.109040
82. Guo D, Wang S, Li J, Bai F, Yang Y, Xu Y, et al. The antimicrobial activity of coenzyme Q0 against planktonic and biofilm forms of Cronobacter sakazakii. Food Microbiol. (2020) 86:103337. doi: 10.1016/j.fm.2019.103337
83. Bouarab-Chibane L, Forquet V, Lantéri P, Clément Y, Léonard-Akkari L, Oulahal N, et al. Antibacterial properties of polyphenols: characterization and QSAR (Quantitative Structure-Activity Relationship) models. Front Microbiol. (2019) 10:829. doi: 10.3389/fmicb.2019.00829
84. Turek C, Stintzing FC. Stability of essential oils: a review. Compr Rev Food Sci Food Saf. (2013) 12:40–53. doi: 10.1111/1541-4337.12006
85. Rao J, Chen B, McClements DJ. Improving the efficacy of essential oils as antimicrobials in foods: mechanisms of action. Annu Rev Food Sci Technol. (2019) 10:365–87. doi: 10.1146/annurev-food-032818-121727
86. Maes C, Bouquillon S, Fauconnier ML. Encapsulation of essential oils for the development of biosourced pesticides with controlled release: a review. Molecules. (2019) 24:2539. doi: 10.3390/molecules24142539
87. Hayes M, Barrett E, Ross RP, Fitzgerald GF, Hill C, Stanton C. Evaluation of an antimicrobial ingredient prepared from a Lactobacillus acidophilus casein fermentate against Enterobacter sakazakii. J Food Prot. (2009) 72:340. doi: 10.4315/0362-028X-72.2.340
88. Awaisheh SS, Al-Nabulsi AA, Osaili TM, Ibrahim S, Holley R. Inhibition of Cronobacter sakazakii by heat labile bacteriocins produced by probiotic LAB isolated from healthy infants. J Food Sci. (2013) 78:1416–20. doi: 10.1111/1750-3841.12209
89. Charchoghlyan H, Kwon H, Hwang DJ, Lee JS, Lee J, Kim M. Inhibition of Cronobacter sakazakii by Lactobacillus acidophilus n.v. Er2 317/402. Korean J Food Sci Anim Resour. (2016) 36:635–40. doi: 10.5851/kosfa.2016.36.5.635
90. Kim DH, Jeong D, Song KY, Kang IB, Kim H, Seo KH. Culture supernatant produced by Lactobacillus kefiri from kefir inhibits the growth of Cronobacter sakazakii. J Dairy Res. (2018) 85:98–103. doi: 10.1017/S0022029917000802
91. Campana R, Federici S, Ciandrini E, Mani A, Buffone W. Lactobacillus spp. inhibit the growth of Cronobacter sakazakii ATCC 29544 by altering its membrane integrity. J Food Sci Technol. (2019) 56:3962–7. doi: 10.1007/s13197-019-03928-x
92. Campion A, Morrissey R, Field D, Cotter PD, Hill C, Ross RP. Use of enhanced nisin derivatives in combination with food-grade oils or citric acid to control Cronobacter sakazakii and Escherichia coli O157: H7. Food Microbiol. (2017) 65:254–63. doi: 10.1016/j.fm.2017.01.020
93. Collado MC, Isolauri E, Salminen S. Specific probiotic strains and their combinations counteract adhesion of Enterobacter sakazakii to intestinal mucus. FEMS Microbiol Lett. (2008) 285:58–64. doi: 10.1111/j.1574-6968.2008.01211.x
94. Kapourchali FR, Cresci GAM. Early-life gut microbiome-the importance of maternal and infant factors in its establishment. Nutr Clin Pract. (2020) 35:386–405. doi: 10.1002/ncp.10490
95. Navarro-Tapia E, Sebastiani G, Sailer, S Almeida, Toledano L, Serra-Delgado M, et al. Probiotic supplementation during the perinatal and infant period: effects on gut dysbiosis and disease. Nutrients. (2020) 12:2243. doi: 10.3390/nu12082243
96. Deshpande G, Athalye-Jape G, Patole S. Para-probiotics for preterm neonates - the next frontier. Nutrients. (2018)10:871. doi: 10.3390/nu10070871
97. Aguilar-Toalá JE, Garcia-Varela R, Garcia HS, Mata-Haro V, González-Córdova AF, Vallejo-Cordoba B, et al. Postbiotics: an evolving term within the functional foods field. Trends Food Sci Technol. (2018) 75:105–14. doi: 10.1016/j.tifs.2018.03.009
98. Ibarra-Sánchez LA, El-Haddad N, Mahmoud D, Miller MJ, Karam L. Invited review: advances in nisin use for preservation of dairy products. J Dairy Sci. (2020) 103:2041–52. doi: 10.3168/jds.2019-17498
99. Al-Nabulsi A, Osaili TM, Al-Holy MA, Shaker RR, Ayyash MM, Olaimat AN, Holley RA. Influence of desiccation on the sensitivity of Cronobacter spp. to lactoferrin or nisin in broth and powdered infant formula. Int J Food Microbiol. (2009) 136:221–6. doi: 10.1016/j.ijfoodmicro.2009.08.008
100. O'Connor PM, Kuniyoshi TM, Oliveira RP, Hill C, Ross RP, Cotter PD. Antimicrobials for food and feed; a bacteriocin perspective. Curr Opin Biotechnol. (2020) 61:160–7. doi: 10.1016/j.copbio.2019.12.023
101. Gurtler JB, Beuchat LR. Inhibition of growth of Enterobacter sakazakii in reconstituted infant formula by the lactoperoxidase system. J Food Prot. (2007) 70:2104–10. doi: 10.4315/0362-028X-70.9.2104
102. Harouna S, Carramiñana JJ, Navarro F, Pérez MD, Calvo M, Sánchez L. Antibacterial activity of bovine milk lactoferrin on the emerging foodborne pathogen Cronobacter sakazakii: effect of media and heat treatment. Food Control. (2015) 47:520–5. doi: 10.1016/j.foodcont.2014.07.061
103. Harouna S, Franco I, Carramiñana JJ, Blázquez A, Abad I, Pérez MD, et al. Effect of hydrolysis and microwave treatment on the antibacterial activity of native bovine milk lactoferrin against Cronobacter sakazakii. Int J Food Microbiol. (2020) 319:108495. doi: 10.1016/j.ijfoodmicro.2019.108495
104. Téné N, Roche-Chatain V, Rifflet A, Bonnafe E, Lefranc B, Leprince J, et al. Potent bactericidal effects of bicarinalin against strains of the Enterobacter and Cronobacter genera. Food Control. (2014) 42:202–6. doi: 10.1016/j.foodcont.2014.02.026
105. Abusheliabi A, Al-Rumaithi HO, Olaimat AN, Al-Nabulsi AA, Osaili T, Shaker R, et al. Inhibitory effect of camel milk on Cronobacter sakazakii. J Food Saf. (2017) 37:e12343. doi: 10.1111/jfs.12343
106. Willemin ME, Lumen A. Thiocyanate: a review and evaluation of the kinetics and the modes of action for thyroid hormone perturbations. Crit Rev Toxicol. (2017) 47:537–63. doi: 10.1080/10408444.2017.1281590
107. Ellison RT, Giehl TJ, LaForce FM. Damage of the outer membrane of enteric Gram-negative bacteria by lactoferrin and transferrin. Infect Immunol. (1988) 56:2774–81. doi: 10.1128/IAI.56.11.2774-2781.1988
108. Kell DB, Heyden EL, Pretorius E. The biology of lactoferrin, an iron-binding protein that can help defend against viruses and bacteria. Front Immunol. (2020)11:1221. doi: 10.3389/fimmu.2020.01221
109. Gao Y, Hou L, Lu C, Wang Q, Pan B, Wang Q, et al. Enteral lactoferrin supplementation for preventing sepsis and necrotizing enterocolitis in preterm infants: a meta analysis with trial sequential analysis of randomized controlled trials. Front Pharmacol. (2020) 11:1186. doi: 10.3389/fphar.2020.01186
110. Hoek KS, Milne J, Grieve PA, Dionysius DA, Smith R. Antibacterial activity of bovine lactoferrin-derived peptides. Antimicrob Agents Chemother. (1997) 41:54–9. doi: 10.1128/AAC.41.1.54
111. Nielsen SD, Beverly RL, Qu Y, Dallas DC. Milk bioactive peptide database: a comprehensive database of milk protein-derived bioactive peptides and novel visualization. Food Chem. (2017) 232:673–82. doi: 10.1016/j.foodchem.2017.04.056
112. Wang G, Li X, Wang Z. APD3: the antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. (2016) 44:1087–93. doi: 10.1093/nar/gkv1278
113. Montagna MT, Diella G, Triggiano F, Caponio GR, De Giglio O, Caggianon G, et al. Chocolate, “food of the gods”: history, science, and human health. Int J Environ Res Public Health. (2019) 16:4960. doi: 10.3390/ijerph16244960
114. Tarka SM, Morrissey RB, Apgar JL, Hostetler KA, Shively CA. Chronic toxicity/carcinogenicity studies of cocoa powder in rats. Food Chem Toxic. (1991) 29:7–19. doi: 10.1016/0278-6915(91)90057-E
115. Ballotey-babington L, Kwapong A, N'Guessan B, Amponsah S, Asiedu-Gyekye I. Unsweetened natural cocoa powder: a potent nutraceutical in perspective. IntechOpen. (2019). Available online at: https://www.intechopen.com/books/theobroma-cacao-deploying-science-for-sustainability-of-global-cocoa-economy/unsweetened-natural-cocoa-powder-a-potent-nutraceutical-in-perspective (accessed October 26, 2020).
116. Zhang H, Qi R, Mine Y. The impact of oolong and black tea polyphenols on human health. Food Biosci. (2019) 29:55–61. doi: 10.1016/j.fbio.2019.03.009
117. Yang CS, Zhang JS. Studies on the prevention of cancer and cardiometabolic diseases by tea: issues on mechanisms, effective doses, and toxicities. J Agri. Food Chem. (2019) 67:5446–56. doi: 10.1021/acs.jafc.8b05242
118. Sergi C. Epigallocatechin-3-gallate toxicity in children: a potential and current toxicological event in the differential diagnosis with virus-triggered fulminant hepatic failure. Front Pharmacol. (2020) 10:1563. doi: 10.3389/fphar.2019.01563
119. United States Food and Drug Administration (US FDA). CFR – Code of Federal Regulations Title 21. (2019). Available online at: https://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/CFRSearch.cfm?CFRPart=184&showFR=1 (accessed October 26, 2020).
120. United States Food and Drug Administration (USFDA). Dietary Supplements. (2019). Available online at: https://www.fda.gov/food/dietary-supplements (accessed October 26, 2020).
121. Zhang C, Fan L, Fan S, Wang J, Luo T, Tang Y, et al. Cinnamomum cassia Presl: a review of its traditional uses, phytochemistry, pharmacology and toxicology. Molecules. (2019) 24:3473. doi: 10.3390/molecules24193473
122. Singletary K. Cinnamon. Update of potential health benefits. Nutr Today. (2019) 54:42–52. doi: 10.1097/NT.0000000000000319
123. Hajimonfarednejad M, Ostovar M, Raee MJ, Hashempur MH, Mayer JG, Heydari M. Cinnamon: a systematic review of adverse events. Clin Nutr. (2019) 38:594–602. doi: 10.1016/j.clnu.2018.03.013
124. Singletary K. Vanilla. Potential health benefits. Nutr Today. (2020) 55:186–96. doi: 10.1097/NT.0000000000000412
125. van Assendelft A. Bronchospasm induced by vanillin and lactose. Eur J Respir Dis. (1984) 65:468–72.
126. Wang XS, Xue YS, Jiang Y, Ni HL, Zhu H, Luo BG, et al. Occupational contact dermatitis in manufacture of vanillin. Chin Med J. (1987) 100:250–4.
127. World Health Organization (WHO). Evaluations of the Joint FAO/WHO Expert Committee on Food Additives (JECFA). (2019). Available online at: https://apps.who.int/food-additives-contaminants-jecfa-database/search.aspx (accessed October 26, 2020).
128. Deshpande GC, Cai W. Use of lipids in neonates requiring parenteral nutrition. J Parenter Enteral Nutr. (2020) 44:45–54. doi: 10.1002/jpen.1759
129. Maher T, Clegg ME. A systematic review and meta-analysis of medium-chain triglycerides effects on acute satiety and food intake. Crit Rev Food Sci Nutr. (2020) 26:1–13. doi: 10.1080/10408398.2020.1742654
130. EFSA ANS Panel (EFSA Panel on Food Additives and Nutrient Sources added to Food) Mortensen A Aguilar F Crebelli R Di Domenico A Dusemund B Frutos MJ. Scientific opinion on the re-evaluation of fatty acids (E 570) as a food additive. EFSA J. (2017) 15:4785. doi: 10.2903/j.efsa.2017.4785
131. Samarghandian S, Azimi-Nezhad M, Farkhondeh T. Thymoquinone-induced antitumor and apoptosis in human lung adenocarcinoma cells. J Cell Physiol. (2019) 234:10421–31. doi: 10.1002/jcp.27710
132. Khader M, Bresgen N, Eckl PM. In vitro toxicological properties of thymoquinone. Food Chem Toxicol. (2009) 47:129–33. doi: 10.1016/j.fct.2008.10.019
133. Dollah HMA, Parhizkar S, Latiff LA, Hassan MHB. Toxicity effect of Nigella sativa on the liver function of rats. Adv Pharm Bull. (2013) 3:97–102. doi: 10.5681/apb.2013.016
134. Rahmani AH, Alzohairy MA, Khan MA, Aly SM. Therapeutic implications of black seed and its constituent TQ in the prevention of cancer through inactivation and activation of molecular pathways. Evid Based Complement Alternat Med. (2014) 2014:724658. doi: 10.1155/2014/724658
135. Lin CC, Kumar KJS, Liao JW, Kuo YH, Wang SY. Genotoxic, teratotoxic and oral toxic assessments of Antrodia cinnamomea health food product [Leader Deluxe Antrodia cinnamomea (R)]. Toxicol Rep. (2015) 2:1409–17. doi: 10.1016/j.toxrep.2015.10.007
136. Nguyen T, Brody H, Lin G-H, Rangé H, Kuraji R, Ye C, et al. Probiotics, including nisin-based probiotics, improve clinical and microbial outcomes relevant to oral and systemic diseases. Periodontol. (2000) 82:173–85. doi: 10.1111/prd.12324
137. Rodrigues G, Silva GGO, Buccini DF, Duque HM, Dias SC, Franco OL. Bacterial proteinaceous compounds with multiple activities toward cancers and microbial infection. Front Microbiol. (2019) 10:1690. doi: 10.3389/fmicb.2019.01690
138. Kitagawa N, Otani T, Inai T. Nisin, a food preservative produced by Lactococcus lactis, affects the localization pattern of intermediate filament protein in HaCaT cells. Anat Sci Int. (2019) 94:163–71. doi: 10.1007/s12565-018-0462-x
139. Gorlewska-Roberts KM, Teitel CH, Lay JO Jr, Roberts DW, Kadlubar FF. Lactoperoxidase-catalyzed activation of carcinogenic aromatic and heterocyclic amines. Chem Res Toxicol. (2004) 17:1659–66. doi: 10.1021/tx049787n
140. Abu-Serie MM, El-Fakharany EM. Efficiency of novel nanocombinations of bovine milk proteins (lactoperoxidase and lactoferrin) for combating different human cancer cell lines. Sci Rep. (2017) 7:16769. doi: 10.1038/s41598-017-16962-6
141. Morita Y, Ono A, Serizawa A, Yogo K, Ishida-Kitagawa N, Takeya T, et al. Purification and identification of lactoperoxidase in milk basic proteins as an inhibitor of osteoclastogenesis. J Dairy Sci. (2011) 94:2270–9. doi: 10.3168/jds.2010-4039
142. Gibbons JA, Kanwar JR, Kanwar RK. Iron-free and iron-saturated bovine lactoferrin inhibit surviving expression and differentially modulate apoptosis in breast cancer. BMC Cancer. (2015) 15:1–16. doi: 10.1186/s12885-015-1441-4
143. Manzoni P. Clinical benefits of lactoferrin for infants and children. J Pediatr. (2016) 173:43–52. doi: 10.1016/j.jpeds.2016.02.075
144. Telang S. Lactoferrin: a critical player in neonatal host defense. Nutrients. (2018) 10:1228. doi: 10.3390/nu10091228
145. Motoki N, Mizuki M, Tsukahara T, Miyakawa M, Kubo S, Oda H, et al. Effects of lactoferrin-fortified formula on acute gastrointestinal symptoms in children aged 12-32 months: a randomized, double-blind, placebo-controlled trial. Front Pediatr. (2020) 8:233. doi: 10.3389/fped.2020.00233
146. Yamauchi K, Toida T, Nishimura S, Nagano E, Kusuoka O, Teraguchi S, et al. 13-week oral repeated administration toxicity study of bovine lactoferrin in rats. Food Chem Toxicol. (2000) 38:503–12. doi: 10.1016/S0278-6915(00)00036-3
147. Cutone A, Rosa L, Ianiro G, Lepanto MS, Bonaccorsi di Patti MC, Valenti P, et al. Lactoferrin's anti-cancer properties: safety, selectivity, and wide range of action. Biomolecules. (2020) 10:456. doi: 10.3390/biom10030456
148. Piqué N, Berlanga M, Miñana-Galbis D, Piqué N, Berlanga M, Miñana-Galbis D. Health benefits of heat-killed (Tyndallized) probiotics: an overview. Int J Mol Sci. (2019) 20:2534. doi: 10.3390/ijms20102534
149. Lee JE, Lee J, Kim JH, Cho N, Lee SH, Park SB, et al. Characterization of the anti-cancer activity of the probiotic bacterium Lactobacillus fermentum using 2D vs. 3D culture in colorectal cancer cells. Biomolecules. (2019) 9:557. doi: 10.3390/biom9100557
150. Constable A, Mahadevan B, Pressman P, Garthoff JA, Meunier L, Schrenk D, et al. An integrated approach to the safety assessment of food additives in early life. Toxicol Res Appl. (2017) 1:1–26. doi: 10.1177/2397847317707370
151. Katz DL, Doughty K, Ali A. Cocoa and chocolate in human health and disease. Antioxid Redox Signal. (2011)15:2779–811. doi: 10.1089/ars.2010.3697
152. Martins TF, Palomino OM, Álvarez-Cilleros D, Martín MA, Ramos S, Goya L. Cocoa flavanols protect human endothelial cells from oxidative stress. Plant Foods Hum Nutr. (2020)75:161–8. doi: 10.1007/s11130-020-00807-1
153. Cory H, Passarelli S, Szeto J, Tamez M, Mattei J. The role of polyphenols in human health and food systems: a mini-review. Front Nutr. (2018) 5:1–9. doi: 10.3389/fnut.2018.00087
154. Liu X, Li FF, Xie JH, Huang DF, Xie MY. Fetal and neonatal genistein exposure aggravates to interfere with ovarian follicle development of obese female mice induced by high-fat diet. Food Chem Toxicol. (2020) 135:110982. doi: 10.1016/j.fct.2019.110982
155. Adgent AA, Umbach DM, Zemel BS, Kelly A, Schall JI, Ford EG, et al. Longitudinal study of estrogen-responsive tissues and hormone concentrations in infants fed soy formula. J Clin Endocrinol Metab. (2018) 103:1899–909. doi: 10.1210/jc.2017-02249
156. Testa I, Salvatori C, Di Cara G, Latini A, Frati F, Troiani S, et al. Soy-based infant formula: are phyto-oestrogens still in doubt? Front Nutr. (2018) 5:110. doi: 10.3389/fnut.2018.00110
157. Jensen RG, Clark RM, Ferris AM. Composition of the lipids in human milk: a review. Lipids. (1980) 15:345–55. doi: 10.1007/BF02533550
158. Hageman JH, Danielsen M, Nieuwenhuizen AG, Feitsma AL, Dalsgaard TK. Comparison of bovine milk fat and vegetable fat for infant formula: implications for infant health. Int Dairy J. (2019) 92:37–49. doi: 10.1016/j.idairyj.2019.01.005
159. Schreiner L, Bauer J, Ortner E, Buettner A. Structure–odor activity studies on derivatives of aromatic and oxygenated monoterpenoids synthesized by modifying p-cymene. J Nat Prod. (2020) 83:834–42. doi: 10.1021/acs.jnatprod.9b00339
160. Younes M, Aggett P, Aguilar F, Crebelli R, Dusemund B, Filipič M, et al. Safety of nisin (E 234) as a food additive in the light of new toxicological data and the proposed extension of use. EFSA J. (2017) 15:5063. doi: 10.2903/j.efsa.2017.5063
161. Banks J G, Board RG. Preservation by the lactoperoxidase system (LP-S) of a contaminated infant milk formula. Lett Appl Microbiol. (1985) 1:81–5. doi: 10.1111/j.1472-765X.1985.tb01495.x
162. Kawaguchi S, Hayashi T, Masano H, Okuyama K, Suzuki T, Kawase K. Effect of lactoferrin-enriched infant formula on low birth weight infants. Shuusankiigaku. (1989)19:125–30.
163. Wakabayashi H, Yamauchi K, Abe F. Quality control of commercial bovine lactoferrin. BioMetals. (2018) 31:313–9. doi: 10.1007/s10534-018-0098-2
164. Mennella JA, Beauchamp GK. The human infants' responses to vanilla flavors in human milk and formula. Infant Behav Dev. (1996) 19:13–9. doi: 10.1016/S0163-6383(96)90040-5
165. Beauchamp GK, Mennella JA. Early flavor learning and its impact on later feeding behavior. J Pediatr Gastr Nutr. (2009) 48:25–30. doi: 10.1097/MPG.0b013e31819774a5
166. Valencia GA, Zare EN, Makvandi P, Gutiérrez TJ. Self-assembled carbohydrate polymers for food applications: a review. Compr Rev Food Sci Food Saf. (2019) 18:2009–24. doi: 10.1111/1541-4337.12499
167. Sampathkumar K, Tan KX, Loo SCJ. Developing nano-delivery systems for agriculture and food applications with nature-derived polymers. iScience. (2020) 23:101055. doi: 10.1016/j.isci.2020.101055
Keywords: natural, antimicrobials, Cronobacter, safety, powdered infant formula
Citation: Polat Yemiş G and Delaquis P (2020) Natural Compounds With Antibacterial Activity Against Cronobacter spp. in Powdered Infant Formula: A Review. Front. Nutr. 7:595964. doi: 10.3389/fnut.2020.595964
Received: 18 August 2020; Accepted: 08 October 2020;
Published: 23 November 2020.
Edited by:
Maria João Fraqueza, University of Lisbon, PortugalReviewed by:
Ben Davies Tall, United States Food and Drug Administration, United StatesDejan S. Stojkovic, University of Belgrade, Serbia
Copyright © 2020 Her Majesty the Queen in Right of Canada, as represented by the Minister of Agriculture and Agri-Food Canada. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pascal Delaquis, cGFzY2FsLmRlbGFxdWlzQGNhbmFkYS5jYQ==