 Mariantonia Maglio*
Mariantonia Maglio* Riccardo Troncone
Riccardo Troncone- Department of Medical Translational Sciences and European Laboratory for the Investigation of Food-Induced Diseases, University Federico II, Naples, Italy
Celiac disease (CD) is a systemic disease that primarily affects the small intestine. The presence of anti-tissue transglutaminase 2 (anti-TG2) antibodies in the serum, as well as the presence of autoimmune phenomena, account for the inclusion of CD among autoimmune diseases. Anti-TG2 autoantibodies are produced at intestinal level, where they are deposited even before they appear in circulation. The pathogenic events that lead to their production are still not completely defined, but a central role seems to be played by gliadin-specific T cells. Interestingly, limited somatic mutations have been observed in VH and VL genes in TG2-specific plasma cells, another important aspect being the biased use of a heavy chain encoded by the VH5 gene. Conflicting data have been produced over the years on the effect of anti-TG2 antibodies on TG2 function. Although the presence of anti-TG2 antibodies in serum is considered a hallmark of CD and relevant from a clinical viewpoint, the role of these autoantibodies in the development of the celiac lesion remains to be defined. In the years, different technical approaches have been implemented to detect and measure intestinal CD-associated autoantibody production. Two aspects can make intestinal anti-TG2 antibodies relevant: from a clinical viewpoint: the first is their proposed ability in potential coeliac patients to predict the development of a full-blown enteropathy; the second is their possible role in revealing a condition of reactivity to gluten in patients with no circulating CD-associated autoantibodies. In fact, the detection of CD-specific autoantibodies production in the intestine, in the absence of serum positivity for the same antibodies, could be suggestive of a very early condition of gluten reactivity; alternatively, it could be not specific for CD and merely attributable to intestinal inflammation. In conclusion, the role of mucosal anti-TG2 antibodies in pathogenesis of CD is unknown. Their presence, the modalities of their production, their gluten dependence render them a unique model to study autoimmunity.
Introduction
Celiac disease (CD) is an immune-mediated systemic disorder elicited in genetically susceptible individuals by the ingestion of gluten contained in wheat and related prolamines in rye and barley. The disorder is characterized by a variable combination of clinical manifestations depending on dietary gluten exposure, the presence in the serum of CD-specific antibodies (anti-tissue transglutaminase and anti-endomysium antibodies), and different degree of enteropathy. CD patients show the presence of alleles of class II major histocompatibility complex DQA1*0501-DQB1*02 (HLA-DQ2 haplotype) and/or DQA1*0301-DQB1*0302 (HLA-DQ8 haplotype) (1). The prevalence of CD in European Countries varies considerably from 0.2 to 3% (2, 3). Comparable prevalence data have been reported also in South America and the USA (4, 5). CD is a common disorder in North Africa, Middle East Countries and India as well (6, 7). Taken together, such rates of prevalence establish celiac disease as one of the most common genetically-based chronic diseases.
The expression of autoimmune phenomena (8, 9), first of all the presence in the serum of anti-tissue transglutaminase 2 (anti-TG2) antibodies (10), and the strict association with autoimmune disorders suggested ESPGHAN to define CD as a systemic autoimmune disease (1).
In this short review we focus our attention on intestinal anti-TG2 antibodies discussing the mechanisms of their production, their possible role in the pathogenesis of the disease and the clinical implications of their detection.
T Cells in CD
The current concept of CD pathogenesis is that it involves components of both innate and adaptive immunity. Gluten is highly resistant to proteolytic degradation by mammalian and microbial intraluminal enzymes for its content of repetitive glutamine- and proline-rich sequences (11, 12). The incomplete degradation results in the persistence of gluten-derived gliadin peptides which access the lamina propria, either actively through the transepithelial way or passively by paracellular flux through damaged epithelial barrier (13, 14), inducing the activation of the mucosal immune system. It is still unclear if there is a primary defect of the barrier. The role played by some gliadin peptides, such as p31–43, is unclear too. They are thought to induce epithelial stress and proinflammatory events, which pave the way to the activation of the adaptive immune response (15–17). Other gliadin peptides, the immunodominant ones such as the 33-mer, activate in lamina propria gluten-specific CD4+ T cell responses presented by HLA-DQ2 or DQ8 molecules (18, 19). A key role in this process is played by tissue transglutaminase 2 (TG2) (20). The enzyme converts particular glutamine residues in gluten peptides to glutamic acid during a deamidation reaction. This results in higher affinity of these gliadin peptides for HLA-DQ2 or DQ8, thereby promoting the activation of T cells (21–23). Once activated, gluten-specific CD4+ T cells produce a pattern of pro-inflammatory cytokines dominated by interferon (IFN)-γ (Th1 skewed) and IL-21 (24, 25). Other cytokines, expression of the innate immune response, are also overproduced in the CD mucosa, such as IL15, IL18 and type 1 interferons. They are thought to be produced by stressed intestinal epithelial cells and/or dendritic cells (26–28). The mechanisms responsible of their recruitment are still unknown, but it is commonly accepted that cytotoxic intraepithelial lymphocytes are the key effector T cells mediating villous atrophy in celiac disease (29). They require complementary signals generated by adaptive anti-gluten immunity and epithelial stress to become pathogenic licensed killer cells (30). For the T lymphocytes intraepithelial recruitment, a key role seems to be carried out by IL-15 implicated in the expression of activating natural killer receptors CD94 (31) and NKG2D (32, 33), as well as in the expression of stress molecules at epithelial level (16). A complex remodeling of the mucosa takes place downstream of T-cell activation which leads to the classical “flat mucosa” of celiac disease. This process involve metalloproteinases (34) and growth factors (35), (Figure 1).
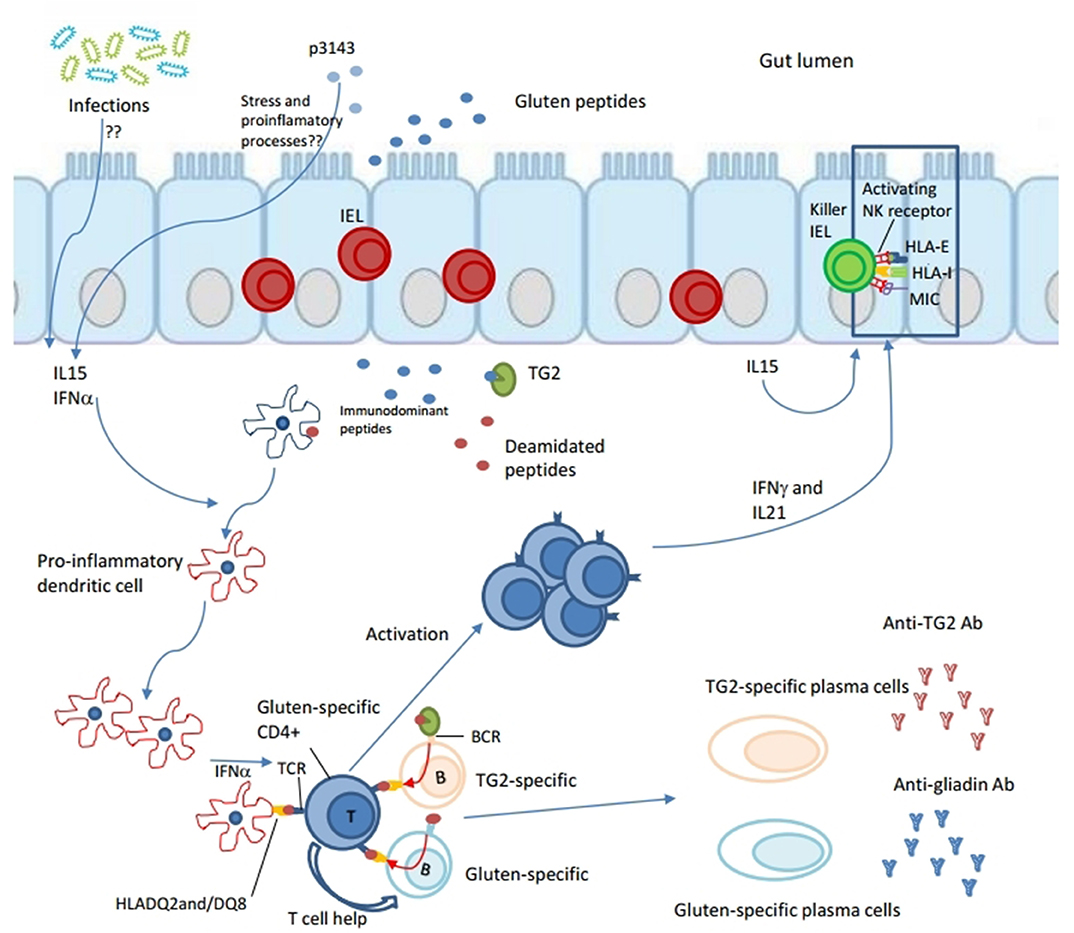
Figure 1. Both innate and adaptive immune responses are induced in the pathogenesis of celiac disease. Gliadin peptides resulting from the partial degradation of gluten in the intestinal lumen cross the epithelial barrier through the transepithelial way or passively by paracellular flux gaining access to lamina propria. Some gliadin peptides, such as p31–43, are thought to induce epithelial stress and inflammation. Other, the immunodominant ones such as the 33-mer, are deamidated by tissue transglutaminase 2 (TG2), resulting in higher affinity for HLA-DQ2 or DQ8 molecules. Deamidated gliadin-peptides are taken up by antigen presenting cells (APCs), such as pro-inflammatory dendritic cells, which promote activation of gluten-specific CD4+ T cell responses in lamina propria. Because of an already inflamed environment (by virus, gliadin? eliciting IL15 and type 1 interferons response), gluten-specific CD4+ T cells express a Th1 phenotype dominated by interferon (IFN)-γ and IL-21. Furthermore, gluten-specific T cells are thought to give help to both TG2-specific and gluten-specific B cells to differentiate into secreting anti-TG2 and anti-gliadin antibodies plasma cells, respectively. In particular, B cell recognizes its antigen (TG2-gliadin complex or gliadin peptides) by BCR, internalizes it, processes and presents gliadin in the context of HLA-DQ2 or DQ8 molecules to gliadin-specific CD4+ T cell. Because of their interaction both T cell and B cell would be activated producing pro-inflammatory cytokines, the first one, and differentiating in secreting plasma cell, the second. Other cytokines, such as IL15 and INF α are thought to be produced by stressed intestinal epithelial cells and/or dendritic cells. They are involved into innate immune responses. IL15 seems to have a central role in the recruitment of intraepithelial lymphocytes (IELs) and in their licensing to become pathogenic killer cells. In fact, IL15 seems involved in the expression of activating natural killer (NK) receptors CD94 and NKG2D, as well as in epithelial expression of stress molecules such as HLA-E and MIC. Finally, a complex remodeling of small intestinal mucosa takes place downstream of T-cell activation that leads to the classical “flat mucosa” of celiac disease.
Tissue Transglutaminase 2 (TG2) as Autoantigen
The first autoantibodies reported in CD were reticulin antibodies (36, 37) directed vs. collagen fibers (36, 38, 39). Subsequently, antibodies against endomysium, connective tissue that covers smooth muscle fibers, were identified (40). Anti-reticulin and anti-endomysium antibodies recognize in fact tissue transglutaminase 2 as proved by Dieterich et al. (10). TG2, the main autoantigen in CD, is a calcium dependent ubiquitous enzyme which exerts various functions. This enzyme has a wide distribution being localized in intracellular compartments and being released from cells in extracellular matrix during inflammation or as a consequence of tissue damage; moreover in extracellular space it is rapidly inactivated by oxidation (41). TG2 is involved in cellular processes of proliferation, of differentiation and apoptosis (42), of recycling endosomes (43), of cell adhesion and fibronectin assembly (44).
TG2 is believed to have at least two crucial roles in celiac disease: as a deamidating enzyme, that can enhance the immunostimulatory effect of gluten, and as a target autoantigen in the immune response. TG2 catalyzes post-translational modification of proteins forming isopeptide bonds; it catalyzes crosslinking between a glutamine residue (glutamine donor) and a lysine residue (glutamine acceptor) of a protein, resulting in the formation of an ε-(γ-glutamyl)–lysine (isopeptidyl) bond, process named transamidation (45). At a relatively low pH can also hydrolyse peptide-bound glutamine to glutamic acid introducing negatively charged residues, processed termed deamidation (45). This process generates negatively charged T cell epitopes in gluten peptides rising their bound affinity to CD-associated HLA molecules (21, 22). As known, the gliadin peptides are excellent substrates for this enzyme as up to 36% of their glutamine residues are accessible to deamidation by TG2 (46). Deamidation activity is believed to be a key step in the pathogenesis of celiac disease; it is a Ca++ dependent process, while it is inhibited when GDP binds to protein (47).
Tissue Transglutaminase2 Antibodies: Mechanisms of Production
The adaptive immune response in celiac disease is characterized by the activation of gluten-specific CD4+ T cells in small intestinal mucosa. In the process leading to the antibody responses to both wheat gliadin and TG2 in the celiac small intestine, gluten-specific CD4+ T cells have a central role. In fact, no TG2 specific T cells have been identified. As known, production of anti-TG2 antibodies can develop only in HLA-DQ2 (48, 49) or HLA-DQ8 individuals (50) and recedes when gluten is excluded from the diet (51, 52). This observation led Sollid et al. (53) to formulate the so-called hapten-carrier hypothesis as a mechanism to explain how TG2-specific B cells get help from gliadin-specific CD4+ T cells to differentiate into IgA and IgG anti-TG2 plasma cells. B cells specific for TG2 recognize their antigens (TG2–gliadin complexes) via surface B cell receptors (BCRs), internalize them and present the processed gluten peptides to gluten-specific CD4+ T cells. Upon the interaction of HLA- DQ2 or HLA- DQ8, gliadin peptides and distinct T cell receptors (TCRs), both the T cells and the B cells would be activated. Once activated, gluten-specific CD4+ T cells start secreting inflammatory cytokines, thereby creating an inflamed environment in the small intestinal lamina propria and, on the other hand, the activated B cells can differentiate into plasma cells that secrete IgA and IgG antibodies against TG2 and deamidated gluten peptides. In this system, complexes formation between gliadin and TG2 makes gliadin a carrier protein for TG2. When gliadin is removed from the diet, T cell help for anti-TG2 specific B cells cease and the serum titers of anti-TG2 antibodies decline.
Site of Production
The presence of anti-TG2 antibodies in serum is a hallmark of CD, being a very sensitive and specific marker to be used as diagnostic tool (54) and in the follow-up of treated patients (55). For years, it has been thought that antibody response in celiac disease originates in the intestinal mucosa. The first experimental evidence came from organ culture studies in which it was observed anti-endomysium antibody production by intestinal fragments after gliadin treatment (56). Conclusive in this regard are the studies by Marzari et al. (57) based on antibody libraries from the gut. They showed that the anti-TG2 autoantibodies are primarily synthesized by specific activated B lymphocytes in small intestinal mucosa of celiac disease patients, and their presence in the serum is probably attributable to the spillover from this source into the blood compartment. Furthermore, this study (57) showed that the heavy chain variable regions of these autoantibodies are primarily derived from the IGHV5-51 gene from the VH5 antibody variable gene family, indicating a preferential usage of this gene in the gluten-dependent autoimmune response to TG2 as subsequently confirmed by Di Niro et al. (58) using single-cell PCR on intestinal B cells.
The abundant local antibody production is confirmed by the observation that TG2 specific antibodies target the antigen in the extracellular matrix and the endothelium of small bowel vessels. A positive correlation between the percentage of TG2 specific plasma cells and serum levels and the intensity of mucosal IgA deposits was observed (59).
Plasma Cells in the Celiac Mucosa
Approximately 1 and 10% of the plasma cells in small intestinal mucosa of celiac patients in the active phase of disease are specific for gliadin or TG2, respectively (58, 60). More recently, TG2 specific plasma cells have been found also in the lamina propria of 79% of potential CD patients before development of mucosal lesion (59). The density of TG2-specific plasma cells is considerably reduced in the celiac intestinal mucosa within 6 months since the start of gluten-free diet, but it remains elevated in comparison to no-CD patients (61). In fact, plasma cells specific for TG2 have been also found in subjects on a gluten free diet for several years (58). These cells could potentially live in vivo for months or years producing antibodies thanks to a favorable microenvironment. Di Niro et al. (58) with a cytometry-based method proved the intestinal production of anti-TG2 antibodies by plasma cells specific for TG2. They reported a very high density of plasma cells secreting TG2-specific IgA autoantibodies with limited somatic hypermutation in the damaged intestinal mucosa of active CD.
However, plasma cell function is not restricted to the production of antibodies. In fact, these cells appear to have a functional B cell receptor (BCR), together with expression of MHCII, as well as the co-stimulatory CD40, CD80, and CD86 molecules, which together may give them ability to activate T cells (62). In this context, recent evidence showed that both gluten-specific and TG2-specific plasma cells, but not dendritic cells and macrophages, were the most abundant cells presenting the immunodominant gluten peptide DQ2.5-glia-α1a in the lamina propria compartment of celiac mucosa (62). Plasma cells loading deamidated gliadin peptides will expand gliadin specific T cell clones. The amplification will provide further help to gliadin specific and TG2-specific B cells. The role played in the amplification of the T cell response renders plasma cells and B cells a target for future therapeutic strategies, even if antibodies do not seem to exert a direct pathogenic effect.
Preferential Epitope Recognition and Usage of VH Genes
Another consequence of the T-B collaboration on the production of anti-TG2 antibodies is the definition of the enzyme epitopes. The autoantibody response against the TG2 in celiac disease patients has been investigated by generating recombinant antibodies from single gut plasma cells reactive with discrete antigen domains (58) and by undertaking proteomic analysis of anti-TG2 serum antibodies (63). Studying a panel of anti-TG2 antibodies produced by single plasma cells from damaged duodenal mucosa of active CD patients was clear that majority of them recognized epitopes in the N-terminal domain of TG2 (64). These antibodies did not interfere with enzymatic activity of TG2 (58) and did not impair antigen presentation to T cells, but it was not the same for the antibodies binding to C-terminal domain of TG2 (65). As proved, antibodies recognizing C-terminal epitopes interfered with TG2 cross-linking activity, and B cells specific for C-terminal epitopes were inefficient at taking up TG2-gluten complexes for presentation to gluten-specific T cells (65).
The intestinal autoimmune response to TG2 is characterized by an extensive switch to IgA occurring in the majority of plasma cells specific for TG2. Interestingly limited somatic mutations have been observed and this has led to the suggestion that their production takes place at extrafollicular sites more than at germinal centers (58). Another important aspect is the biased use of a heavy chain encoded by the VH5 gene. The reason for the preferential expansion of VH5 in the CD repertoire is not known. B cells expressing the IgH variable gene segment IgH V 5-51 have their B cell receptor (BCR) crosslinked to gliadin peptides by TG2 (66). This could account for the overrepresentation of IGHV 5-51 among TG2-specific plasma cells. This phenomenon could be specific for celiac disease. In fact, Maglio et al. (67) showed immunofluorescent detection of anti-TG2 autoantibodies mucosal deposits in 78% of type 1 diabetic children with serum positivity for the same antibodies. In these patients, the phage display analysis of intestinal antibodies libraries showed anti-TG2 antibodies made with a preferential use of the VH5 antibody gene family. At the same time, anti-TG2 antibodies belonging to other VH gene families, such as VH3 and VH1, were shown to be produced in the intestine of type 1 children with normal serum titers of the same antibodies. In the latter subjects high percentage was observed of phage antibodies belonging to VH3 gene family (67).
Pathogenicity of TG2 Antibodies
The role of anti-TG2 and anti-gluten antibodies in the development of the celiac lesion remains to be defined. These antibodies are thought to increase the permeability of the epithelial barrier (68), allowing gliadin peptides to access the lamina propria and affecting epithelial cell biology (69, 70). Moreover, they may amplify the inflammatory immune response to gluten by increasing gluten uptake (71). Conflicting data have been produced over the years on the effect of anti-TG2 antibodies on TG2 function. Several research groups have investigated this topic with different results as to the degree of inhibition (72–76). Evidence have shown that IgA and IgG anti-TG2 autoantibodies from serum of CD patients as well as commercial monoclonal anti-TG2 perform a dose dependent and partial inhibitory effect on the transamidating activity of TG2 of gluten peptides. It has been shown that anti-TG2 antibodies interacting with the extra cellular bound TG2 induce relevant cytoskeleton changes with actin redistribution (70). Furthermore, interaction of anti-TG2 antibodies with extracellular TG2 triggers a rapid mobilization of calcium and the increased intracellular levels of calcium were able to activate intracellular TG2 (77).
Transglutaminase2 is part of a numerous enzyme family. The autoantibody responses targeting other members of this family are of great interest due to their association with specific clinical manifestations of celiac disease. In particular, anti-TG3 and anti-TG6 antibodies, which occur in the context of dermatitis herpetiformis and gluten ataxia, respectively, have been considered as potential contributors in the pathogenesis of these extra-intestinal manifestations (78, 79). Furthermore, anti-TG3 and anti-TG6 autoantibodies are the result of specific immune responses against TG3 and TG6, respectively, and are not the result of cross-reactivity to TG2 (64).
Intestinal Anti-Tissue Transglutaminase 2 Antibodies: Tools for Detection
In the last 40 years different technical approaches have been used to detect and measure intestinal CD-associated autoantibody production: measurement of anti-reticulin antibodies in jejunal juice (80) or of anti-endomysium antibodies (EMA) and anti-TG2 IgA antibodies in feces (81), search of the same antibodies in cultured biopsy supernatants (56, 82–85) or detection of deposited anti-TG2 antibodies in duodenal fragments (86, 87), or of plasma cells secreting them (58) or, lastly, expression in intestinal phage libraries of RNA coding for the celiac autoantibodies (57), (Table 1).
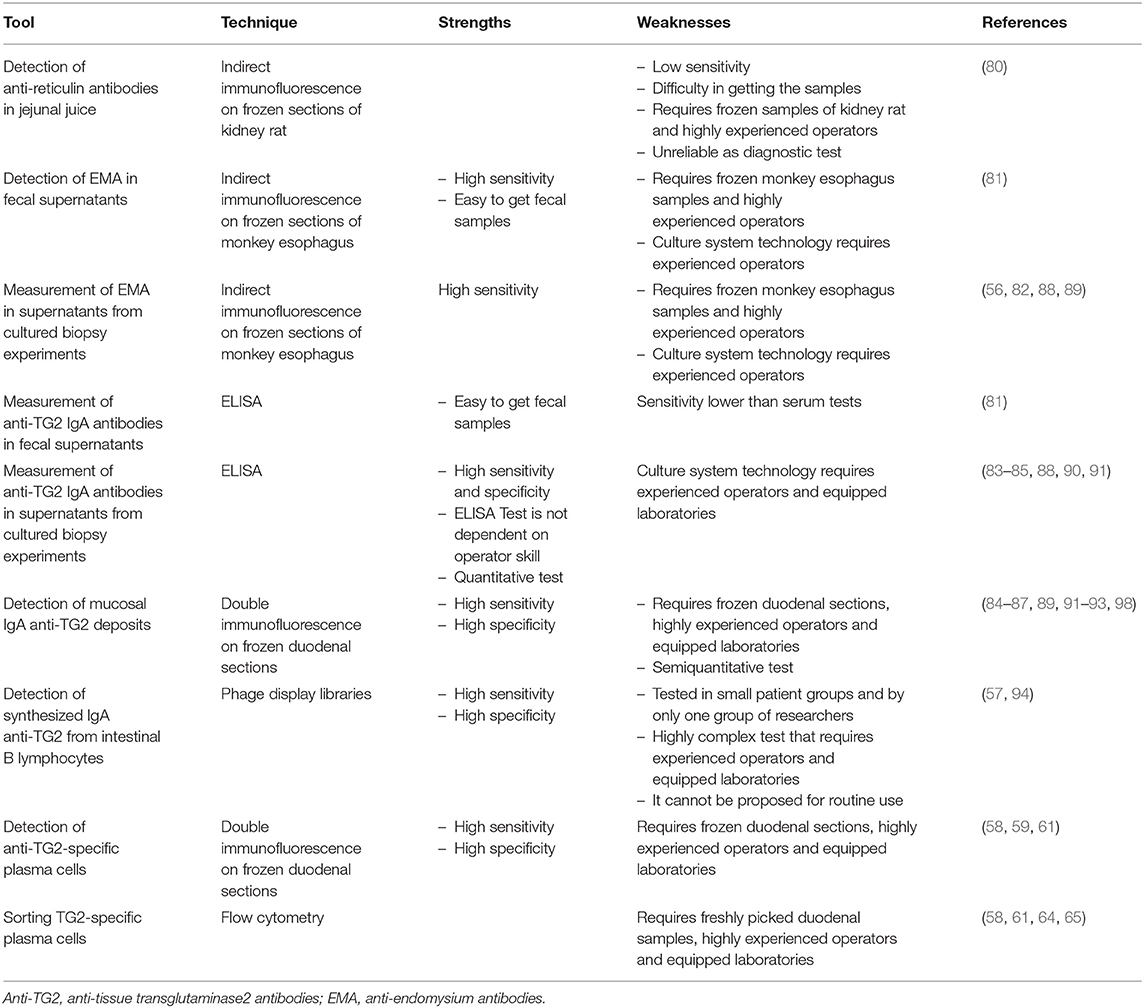
Table 1. Tools employed to detect intestinal production of anti-TG2 autoantibodies.
The first evidence of antibody production in the celiac intestine dates back to the seventies, Mawhinney and Love (80), have demonstrated, recurring to indirect immunofluorescence on frozen sections of kidney rat, the presence of anti-reticulin antibodies in the jejunal juice of CD patients suggesting this feature as a result of stimulation of the secretory immune system. Afterwards, detection of EMA and anti-TG2 IgA antibodies in fecal supernatants, by indirect immunofluorescence analysis on cryostat sections of monkey esophagus and by ELISA test, respectively, represented one of the first proofs that intestinal mucosa is a site of celiac auto-antibody production (81). The authors proposed to recur to this measurement, together with the histological examination of multiple biopsies, as a useful diagnostic tool of CD in mucosae characterized by a patchy lesion; however, at a later time it was determined to be unreliable as a diagnostic test with a sensitivity lower than serum tests (95).
Further evidence in favor of the hypothesis of the intestinal production of the celiac-specific autoantibodies came from in vitro cultured small intestinal biopsy experiments. The search for EMA and/or anti-TG2 antibodies in supernatants collected after 24 h of CD small intestine culture (56, 82–85), recurring to indirect immunofluorescence and ELISA tests, respectively, revealed very high sensitivity and specificity. In the pediatric population ELISA test, that is objective and not influenced by the operator ability, showed intestinal production of anti-TG2 autoantibodies in 100% of CD patients in the active phase of disease with a specificity of 93% (84, 91). In potential celiac disease, when there are circulating CD-specific autoantibodies, but no signs of morphological mucosal injury, this assay proved that in 96% of (84) or in 100% of cases (89) CD intestinal mucosa produced and secreted autoantibodies. It was proposed in particular to improve the accuracy of CD diagnosis in subjects with mild enteropathy (84, 88) and in patients negative for serum antibodies (88–90). Moreover, while the measurement of secreted anti-TG2 in culture supernatants shows high sensitivity and specificity, currently organ cultures are used for only research purposes; it requires expertise and advanced techniques, but might be useful in special situations when other diagnostic methods are inconclusive (88, 89).
Using double immunofluorescence and frozen sections of duodenal mucosa, Korponay-Szabò et al. (86) highlighted thick bundles of mucosal IgA anti-TG2 deposits, secreted at intestinal level and deposited under surface epithelium and around crypts small intestinal mucosa of CD patients. This assay has been validated in several pathology laboratories for a diagnostic use, showing a sensitivity ranging from 73 to 100% (84, 87, 92, 93, 98) and a specificity ranging from 82 to 100% (87, 91, 92, 98). In potential CD patients deposits of IgA anti-TG2 are found less often (84, 87, 96) in about 68% of cases. The assay, while having high sensitivity and specificity, presents some limitations: requires frozen intestinal samples, equipped laboratory and highly experienced operators, (Figure 2). Tosco et al. (84) comparing the diagnostic efficiency of the detection of intestinal deposits of anti-TG2 antibodies and their measurement in biopsy culture supernatants in patients with potential CD, showed a higher diagnostic sensitivity and specificity for the measurement of secreted anti-TG2 antibody into culture supernatants than for immunofluorescence detection of mucosal deposits of the same autoantibodies.
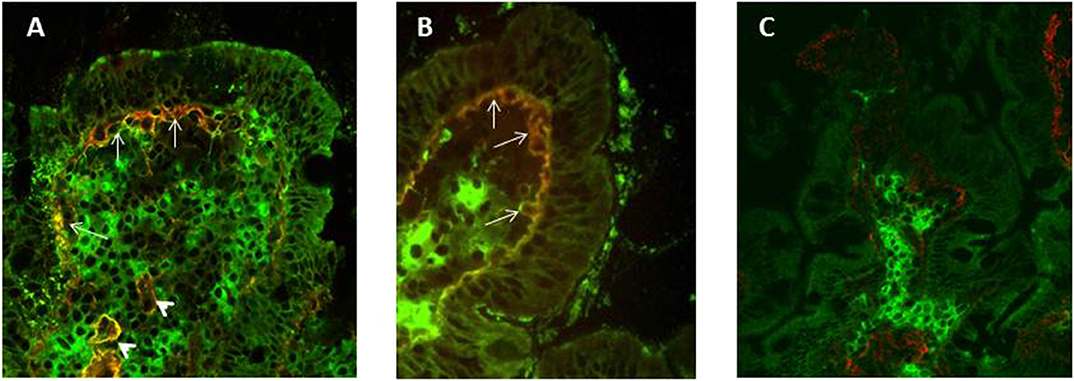
Figure 2. Double Immunofluorescence to detect mucosal deposits of immunoglobulin (Ig)A anti-tissue transglutaminase 2 (TG2). Duodenal mucosa sections from an active CD (A) and a potential CD (B) patient show bundles of IgA anti-TG2 antibody deposits in yellow, which are located under surface epithelium (white arrows) and around blood vessels (white arrowheads). IgAs secreted by plasma cells are detected in green (A–C), TG2 with a sub-epithelial localization is shown in red (C). In panel (C), a duodenal section from a no-CD subject does not show mucosal deposits of anti-TG2 antibodies.
Finally, by means of phage display libraries, Marzari et al. (57) showed that anti-TG2 IgA antibodies are synthesized primarily by specific B lymphocytes in the CD small intestinal mucosa and that in antibodies from celiac patients there is a preferential use of heavy chain variable regions belonging to the VH5 gene family. The phage display libraries technique seems to be more sensitive than other assays, such as detection of mucosal deposits of anti-TG2 antibodies using double immunofluorescence assay, in identifying patients with gluten-dependent intestinal or extra-intestinal symptoms, such as relatives of CD patients who show the presence of normal intestinal mucosa and the absence of serum anti-TG2 antibodies (94). However, this technology has been tested in small groups of patients and only from one research group; further studies are needed for validation. At the same time, it must be emphasized that, because of its complexity, this test cannot be proposed for routine use.
More recently, Di Niro et al. (58) by double immunofluorescence and flow cytometry methods identified TG2-specific plasma cells in duodenal intestine of CD patients in the active phase of disease and characterized their antibody response. Recurring to double immunofluorescence on duodenal cryosections it has been possible to detect TG2 specific plasma cells in all stages of CD but not in controls (58, 59, 61).
Clinical Implications
Intestinal Anti-TG2 in Active CD
Many studies investigated production of anti-TG2 autoantibodies in the intestine of patients with overt CD at diagnosis, by the tools described above. In adults with untreated CD anti-TG2 autoantibodies deposited in the intestine were detected in 100% of cases (92, 97, 98). In pediatric population more variability was reported. Mucosal deposits were identified in 96 to 100% of untreated CD patients (86, 87, 89, 93, 99). Considering the subgroup of children younger than 2 years of age this percentage is reduced (73%) (93). In this age group the search for mucosal deposits of anti-TG2–IgA did not improve the diagnostic performance in comparison to serum detection, (Table 2).
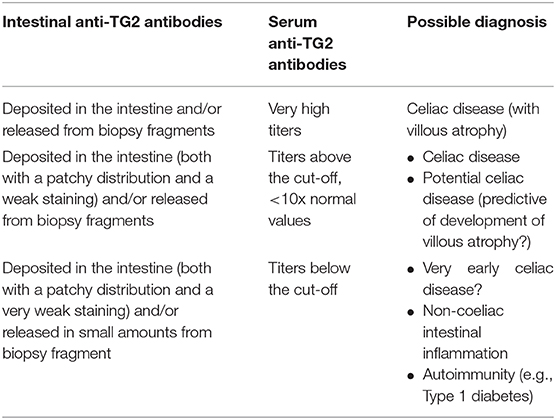
Table 2. Clinical implications of IgA anti TG2 antibodies.
Evidence showed the presence of anti-TG2 autoantibodies deposited in the small intestine of patients with dermatitis herpetiformis (DH) (86), a common extraintestinal manifestation of CD. Mucosal deposits were detectable also in DH patients with normal duodenal morphology and negative for serum specific-CD autoantibodies (86, 100) as well as in DH patients on a gluten-free diet (100).
Another gluten-related disorder investigated for the presence of mucosal deposits in the intestine is gluten ataxia. Anti-TG2 autoantibodies were showed in the gut and brain of patients with gluten ataxia with or without an enteropathy; these mucosal deposits were similar to those detected in the intestine of celiac disease and dermatitis herpetiformis patients but were not detected in subjects with other causes of ataxia. This finding strengthens the idea that gluten ataxia is immune mediated and belongs to the spectrum of gluten related disorders (101).
Intestinal Anti-TG2 in Potential CD
Anti-TG2 autoantibodies production in the intestine has been investigated also in potential CD, a condition characterized by presence of anti-TG2 in the serum, but normal mucosa architecture. Several studies performed in adult and pediatric populations found the presence of mucosal deposits of anti-TG2 ranging from 68 to 100% (87, 89, 92, 97, 99, 102, 103). Tosco et al. (96) speculated that in a group of potential CD patients followed-up with repeated biopsy the detection of mucosal deposits of autoantibodies in the first biopsy could predict the later development of a full-blown villous atrophy, (Table 2).
Intestinal Anti-TG2 in Subjects With Selective IgA Deficiency
The presence of mucosal deposits of anti-TG2 antibodies was also investigated in patients with selective immunoglobulin (Ig)A deficiency (SIgAD), that is a condition frequently associated with celiac disease. A 10–20-fold increased risk of CD may be observed in subjects affected by SIgAD. For this reason, screening for CD is mandatory in SIgAD patients and is performed searching serum IgG anti-TG2 autoantibodies. At intestinal level lack of secretory-IgA is replaced by a compensatory increase in secretory-IgM (104). Borrelli et al. (105) investigated by immunofluorescence the presence of IgM anti-TG2 antibodies in the intestine of SIgAD patients. The authors showed IgM anti-TG2 antibody deposits being present only in CD active patients with SIgAD. However, intestinal IgM anti-TG deposits did not discriminate between SIgAD and potential CD with SIgAD. Therefore, they concluded that the finding of serum IgG CD-associated autoantibodies remains crucially important in the diagnosis of CD in SIgAD.
Intestinal Anti-TG2 in Patients on GFD
The disappearance of serum anti-TG2 antibodies is observed progressively after the beginning of a GFD. It has been considered a useful tool to evaluate the compliance with the diet, although it is clear that it is not able to reveal minor transgressions. It is also not yet established if it is helpful to assess complete histological recovery (106). Several studies were performed in CD patients on a gluten free diet to investigate the effect of the diet on anti-TG2 antibodies intestinal production. Studies on the follow-up of CD patients on GFD (107, 108) showed significant decrease of circulating anti-TG2 antibodies during the first 12 months of the diet and their disappearance in the next 12 months. Moreover, the disappearance from serum of specific CD autoantibodies, following a strict GFD, does not mean the end of their intestinal production. Many studies, involving pediatric or adult patients, showed the presence of intestinal anti-TG2 deposits both after a short term (1 year) and long term (>2 years) GFD (85, 98, 109). After the disappearance from serum, anti-TG2 antibodies as deposits would disappear slowly from the intestine, and only after a long period of GFD small intestinal mucosa would cease to produce them. Even when the mucosal deposits were not detectable in GFD patients, their intestine continue to produce low amounts of these antibodies in culture supernatants as recently showed (85). Further evidence of intestinal anti-TG2 production, even after years of gluten free diet, came from flow cytometry cell sorting of TG2 specific plasma cells from duodenal samples of treated CD patients (58, 61). More recently, Risnes et al. (110) recurring to HLA-DQ:gluten tetramers technology proved the persistence for decades of gluten-specific T cells, which are thought to play a central role in antibody production process, in the intestine of treated CD. Interestingly, during a GFD the CD markers normalize in a sequential order: serum anti-TG2, morphology of small intestine, density of CD3+ intraepithelial lymphocytes and then mucosal deposits. The only marker that remained positive also very late is the high density of TCR-γδ+ intraepithelial lymphocytes (98).
Intestinal Anti-TG2 in Subjects Seronegative: Gluten-related or False Positive?
Intestinal anti-TG2 antibodies may be present early, before they appear in the serum. Borrelli et al. (111), having available very early biopsies from at-risk infants, enrolled in the context of the European multicenter project Prevent Celiac Disease (PreventCD, www.preventcd.com) (112), investigated the appearance of intestinal anti-TG2 deposits and their predictive value for villous atrophy. The authors showed anti-TG2 antibodies deposited in duodenal mucosa of all subjects who had diagnosis of CD. However, mucosal IgA anti-TG2 deposits were also present before the mucosal damage and even before they appeared in serum.
Identifying intestinal production of anti-TG2 becomes important in support of a diagnosis of celiac disease in borderline cases, mainly in patients with absence of serum CD-associated autoantibodies (87, 89, 91, 113) which include relatives of CD patients that Not et al. have defined as affected by cryptic genetic gluten intolerance (94). The production of anti-TG2 antibodies was showed in the intestine of DQ2- or DQ8-positive relatives of CD patients even in the presence of a normal intestinal mucosa and without circulating anti-TG2 antibodies by detection of mucosal deposits (89, 91, 114) and by mucosal phage display antibody assay (94). These patients have been shown produce only at intestinal level IGVH5-51 anti-TG2 antibodies, in other words using the same VH genes found in overt celiac patients. Interestingly the presence of symptoms in these subjects responsive to the gluten free diet, placed them in the spectrum of gluten-related disorders.
Several investigators have shown that intestinal anti-TG2 IgA deposits were also detectable in subjects with diagnosis other than CD (87, 97, 98). The data were confirmed by measuring anti-TG2 antibodies spontaneously secreted into supernatants from organ culture of small intestinal biopsy and by the phage display technologies (91). Indeed, the construction of antibody libraries from biopsy samples showed a production of an anti-TG2 specific antibody repertoire dominated by the usage of VH5-51 gene segment and by the VH3 gene segment. The presence of intestinal anti-TG2 have been reported, in most of such cases associated with other autoimmune disorders such as type 1 diabetes (T1D). In all these conditions, the relation of intestinal anti-TG2 autoantibodies to CD is difficult to establish, the detection of mucosal deposits might be a very early sign of gluten sensitivity or simply a “false-positive” staining. It is possible that intestinal anti-TG2 is an expression of mucosa inflammation related to the autoimmunity state. As already mentioned, a high prevalence of anti-TG2 intestinal deposits has been reported in patients with T1D on a normal diet irrespective of the presence of serum anti-TG2 antibodies (67). However, only T1D patients with elevated serum levels of anti-TG2 antibodies showed the VH5-51 gene usage typical of the anti-TG2 antibodies produced in the intestine of CD patients, suggesting a gluten-dependent phenomenon. In serum anti-TG2 negative T1D patients, who produced and deposited intestinal anti-TG2 antibodies belonging to the VH-1 and VH3- gene families (67, 115), the relationship with dietary gluten is uncertain, (Table 2).
Intestinal Anti-TG2 in Relation With Other Gluten-Related Bio-markers
Many evidences related the presence of mucosal anti-TG2 deposits with other CD markers. The density of TCR-γδ+ intraepithelial lymphocytes is considered highly sensitive and specific for CD (116). In fact, high density of this marker has been shown to be indicative of early developing CD (103). Mucosal deposits of anti-TG2 antibodies have been reported in children who were negative for serum CD-associated autoantibodies but with an increased number of TCR-γδ+ intraepithelial lymphocytes, with mucosal deposits of anti-TG2 antibodies (117). It remains to be established if these subjects belong to the spectrum of gluten related disorders.
Conclusions
Anti-TG2 autoantibodies are a key feature of celiac disease; their intestinal production as well as the deposition in the mucosa, even before their appearance in serum, have been extensively proved. Intestinal anti-TG2 antibodies can be considered to be relevant from a clinical viewpoint for two reasons: the first is their supposed capability to predict the development of a full blown enteropathy in a early phase disease condition such as potential celiac disease; the second is their possible role in revealing a condition of gluten reactivity in patients with absence of circulating CD-associated autoantibodies. Alternatively, the detection of specific CD autoantibodies production in the intestine, in the absence of serum positivity for the same antibodies, could be not specific for celiac disease, but a phenomenon merely attributable to intestinal inflammation.
As previously mentioned, the pathogenic role of mucosal anti-TG2 antibodies in CD is unknown. However, their presence, the modalities of their production, their gluten dependence render them a unique model to study mechanisms of autoimmunity.
Author Contributions
MM and RT contributed equally to the planning and drafting of the manuscript.
Conflict of Interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling Editor declared a past co-authorship with the authors RT.
References
1. Husby S, Koletzko S, Korponay-Szabo IR, Mearin ML, Phillips A, Shamir R, et al. European society for pediatric gastroenterology, hepatology, and nutrition guidelines for the diagnosis of coeliac disease. J Pediatr Gastroenterol Nutr. (2012) 54:136–60. doi: 10.1097/MPG.0b013e31821a23d0
2. Myléus A, Ivarsson A, Webb C, Danielsson L, Hernell O, Högberg L, et al. Celiac disease revealed in 3% of Swedish 12-year-olds born during an epidemic. J Pediatr Gastroenterol Nutr. (2009) 49:170–6. doi: 10.1097/MPG.0b013e31818c52cc
3. Mustalahti K, Catassi C, Reunanen A, Fabiani E, Heier M, McMillan S, et al. The prevalence of celiac disease in Europe: results of a centralized, international mass screening project. Ann Med. (2010) 42:587–95. doi: 10.3109/07853890.2010.505931
4. Rubio-Tapia A, Ludvigsson JF, Brantner TL, Murray JA, Everhart JE. The prevalence of celiac disease in the United States. Am J Gastroenterol. (2012) 107:1538–44. doi: 10.1038/ajg.2012.219
5. Parra-Medina R, Molano-Gonzalez N, Rojas-Villarraga A, Agmon-Levin N, Arango MT, Shoenfeld Y, et al. Prevalence of celiac disease in Latin America: a systematic review and meta-regression. PLoS ONE. (2015) 10:e0124040. doi: 10.1371/journal.pone.0124040
6. Shamir R, Lerner A, Shinar E, Lahat N, Sobel E, Bar-or R, et al. The use of a single serological marker underestimates the prevalence of celiac disease in Israel: a study of blood donors. Am J Gastroenterol. (2002) 97:2589–94. doi: 10.1111/j.1572-0241.2002.06028.x
7. Ramakrishna BS, Makharia GK, Chetri K, Dutta S, Mathur P, Ahuja V, et al. Prevalence of adult celiac disease in India: regional variations and associations. Am J Gastroenterol. (2016) 111:115–23. doi: 10.1038/ajg.2015.398
8. Ventura A, Magazzù G, Greco L. Duration of exposure to gluten and risk for autoimmune disorders in patients with coeliac disease SIGEP study group for autoimmune disorders in coeliac disease. Gastroenterology. (1999) 117:297–303. doi: 10.1053/gast.1999.0029900297
9. Hadjivassiliou M, Sanders DS, Grunewald RA, Woodroofe N, Boscolo S, Aeschlimann D. Gluten sensitivity: from gut to brain. Lancet Neurol. (2010) 9:318–30. doi: 10.1016/S1474-4422(09)70290-X
10. Dieterich W, Ehnis T, Bauer M, Donner P, Volta U, Riecken EO, et al. Identification of tissue transglutaminase as the autoantigen of celiac disease. Nat Med. (1997) 3:797–801. doi: 10.1038/nm0797-797
11. Shan L, Molberg Ø, Parrot I, Hausch F, Filiz F, Gray GM, et al. Structural basis for gluten intolerance in celiac sprue. Science. (2002) 297:2275–9. doi: 10.1126/science.1074129
12. Sollid LM. Coeliac disease: dissecting a complex inflammatory disorder. Nat Rev Immunol. (2002) 2:647–55. doi: 10.1038/nri885
13. Schulzke JD, Schulzke I, Fromm M, Riecken EO. Epithelial barrier and ion transport in coeliac sprue: electrical measurements on intestinal aspiration biopsy specimens. Gut. (1995) 37:777–82. doi: 10.1136/gut.37.6.777
14. Schulzke JD, Bentzel CJ, Schulzke I, Riecken EO, Fromm M. Epithelial tight junction structure in the jejunum of children with acute and treated celiac sprue. Pediatr Res. (1998) 43:435–41. doi: 10.1203/00006450-199804000-00001
15. Maiuri L, Ciacci C, Ricciardelli I, Vacca L, Raia V, Auricchio S, et al. Association between innate response to gliadin and activation of pathogenic T cells in coeliac disease. Lancet. (2003) 362:30–7. doi: 10.1016/S0140-6736(03)13803-2
16. Hüe S, Mention JJ, Monteiro RC, Zhang S, Cellier C, Schmitz J, et al. A direct role for NKG2D/MICA interaction in villous atrophy during celiac disease. Immunity. (2004) 21:367–77. doi: 10.1016/j.immuni.2004.06.018
17. Barone MV I, Gimigliano A, Castoria G, Paolella G, Maurano F, Paparo F, et al. Growth factor-like activity of gliadin, an alimentary protein: implications for coeliac disease. Gut. (2007) 56:480–8. doi: 10.1136/gut.2005.086637
18. Sollid LM. Molecular basis of celiac disease. Annu Rev Immunol. (2000) 18:53–81. doi: 10.1146/annurev.immunol.18.1.53
19. Arentz-Hansen H, McAdam SN, Molberg Ø, Fleckenstein B, Lundin KE, Jørgensen TJ, et al. Celiac lesion T cells recognize epitopes that cluster in regions of gliadins rich in proline residues. Gastroenterology. (2002) 123:803–9. doi: 10.1053/gast.2002.35381
20. Arentz-Hansen H, Körner R, Molberg Ø, Quarsten H, Vader W, Kooy YM, et al. The intestinal T cell response toα-gliadin in adult celiac disease is focused on a single deamidated glutamine targeted by tissue transglutaminase. J Exp Med. (2000) 191:603–12. doi: 10.1084/jem.191.4.603
21. Molberg O, Mcadam SN, Körner R, Quarsten H, Kristiansen C, Madsen L, et al. Tissue transglutaminase selectively modifies gliadin peptides that are recognized by gut-derived T cells in celiac disease. Nat Med. (1998) 4:713–7. doi: 10.1038/nm0698-713
22. van de Wal Y, Kooy Y, van Veelen P, Peña S, Mearin L, Papadopoulos G, et al. Selective deamidation by tissue transglutaminase strongly enhances gliadin-specific T cell reactivity. J Immunol. (1998) 161:1585–8.
23. Dørum S, Qiao SW, Sollid LM, Fleckenstein B. A quantitative analysis of transglutaminase 2-mediated deamidation of gluten peptides: implications for the T-cell response in celiac disease. J Proteome Res. (2009) 8:1748–55. doi: 10.1021/pr800960n
24. Nilsen EM, Jahnsen FL, Lundin KE, Johansen FE, Fausa O, Sollid LM, et al. Gluten induces an intestinal cytokine response strongly dominated by interferon gamma in patients with celiac disease. Gastroenterology. (1998) 115:551–63. doi: 10.1016/S0016-5085(98)70134-9
25. Fina D, Sarra M, Caruso R, Del Vecchio Blanco G, Pallone F, MacDonald TT, et al. Interleukin 21 contributes to the mucosal T helper cell type 1 response in coeliac disease. Gut. (2008) 57:887–92. doi: 10.1136/gut.2007.129882
26. Maiuri L, Ciacci C, Auricchio S, Brown V, Quaratino S, Londei M. Interleukin 15 mediates epithelial changes in celiac disease. Gastroenterology. (2000) 119:996–1006. doi: 10.1053/gast.2000.18149
27. Monteleone G, Pender SL, Alstead E, Hauer AC, Lionetti P, McKenzie C, et al. Role of interferon alpha in promoting T helper cell type 1 responses in the small intestine in coeliac disease. Gut. (2001) 48:425–9. doi: 10.1136/gut.48.3.425
28. Salvati VM, MacDonald TT, Bajaj-Elliott M, Borrelli M, Staiano A, Auricchio S, et al. Interleukin 18 and associated markers of T helper cell type 1 activity in coeliac disease. Gut. (2002) 50:186–90. doi: 10.1136/gut.50.2.186
29. Setty M, Discepolo V, Abadie V, Kamhawi S, Mayassi T, Kent A, et al. Distinct and synergistic contributions of epithelial stress and adaptive immunity to functions of intraepithelial killer cells and active celiac disease. Gastroenterology. (2015) 149:681–91.e10. doi: 10.1053/j.gastro.2015.05.013
30. Meresse B, Curran SA, Ciszewski C, Orbelyan G, Setty M, Bhagat G, et al. Reprogramming of CTLs into natural killer-like cells in celiac disease. J Exp Med. (2006) 203:1343–55. doi: 10.1084/jem.20060028
31. Jabri B, de Serre NP, Cellier C, Evans K, Gache C, Carvalho C, et al. Selective expansion of intraepithelial lymphocytes expressing the HLA-E-specific natural killer receptor CD94 in celiac disease. Gastroenterology. (2000) 118:867–79. doi: 10.1016/S0016-5085(00)70173-9
32. Meresse B, Chen Z, Ciszewski C, Tretiakova M, Bhagat G, Krausz TN, et al. Coordinated induction by IL15 of a TCR-independent NKG2D signaling pathway converts CTL into lymphokine activated killer cells in celiac disease. Immunity. (2004) 21:357–66. doi: 10.1016/j.immuni.2004.06.020
33. Roberts AI, Lee L, Schwartz E, Groh V, Spies T, Ebert EC, et al. NKG2Dreceptors induced by IL-15 costimulate CD28-negative effector CTL in the tissue microenvironment. J Immunol. (2001) 167:5527–30. doi: 10.4049/jimmunol.167.10.5527
34. Daum S, Bauer U, Foss HD, Schuppan D, Stein H, Riecken EO, et al. Increased expression of mRNA for matrix metalloproteinases-1 and−3 and tissue inhibitor of metalloproteinases-1 in intestinal biopsy specimens from patients with coeliac disease. Gut. (1999) 44:17–25. doi: 10.1136/gut.44.1.17
35. Salvati VM, Bajaj-Elliott M, Poulsom R, Mazzarella G, Lundin KE, Nilsen EM, et al. Keratinocyte growth factor and coeliac disease. Gut. (2001) 49:176–81. doi: 10.1136/gut.49.2.176
36. Seah PP, Fry L, Hoffbrand AV, Holborow EJ. Tissue antibodies in dermatitis herpetiformis and adult coeliac disease. Lancet. (1971) 1:834–6. doi: 10.1016/S0140-6736(71)91499-1
37. Alp MH, Wright R. Autoantibodies to reticulin in patients with idiopathic steatorrhoea, coeliac disease, and Crohn's disease, and their relation to immunoglobulins and dietary antibodies. Lancet. (1971) 2:682–5. doi: 10.1016/S0140-6736(71)92249-5
38. Rizzetto M, Doniach D. Types of 'WF' antibodies detected in human sera by immunofluorescence. J. Clin. Pathol. (1973) 26:841–51. doi: 10.1136/jcp.26.11.841
39. Eade OE, Lloyd RS, Lang C, Wright R. IgA and IgG reticulin antibodies in coeliac and non-coeliac patients. Gut. (1977) 18:991–3. doi: 10.1136/gut.18.12.991
40. Chorzelski TP, Beutner EH, Sulej J, Tchorzewska H, Jablonska S, Kumar V, et al. IgA anti-endomysium antibody. A new immunological marker of dermatitis herpetiformis and coeliac disease. Br J Dermatol. (1984) 111:395–402. doi: 10.1111/j.1365-2133.1984.tb06601.x
41. Stamnaes J, Pinkas DM, Fleckenstein B, Khosla C, Sollid LM. Redox regulation of transglutaminase 2 activity. J Biol Chem. (2010) 285:25402–9. doi: 10.1074/jbc.M109.097162
42. Fésüs L, Szondy Z. Transglutaminase 2 in the balance of cell death and survival. FEBS Lett. (2005) 579:3297–302. doi: 10.1016/j.febslet.2005.03.063
43. Zemskov EA, Mikhailenko I, Hsia RC, Zaritskaya L, Belkin AM. Unconventional secretion of tissue transglutaminase involves phospholipid-dependent delivery into recycling endosomes. PLoS ONE. (2011) 6:e19414. doi: 10.1371/journal.pone.0019414
44. Akimov SS, Belkin AM. Cell surface tissue transglutaminase is involved in adhesion and migration of monocytic cells on fibronectin. Blood. (2001) 98:1567–76. doi: 10.1182/blood.V98.5.1567
45. Folk JE. Mechanism and basis for specificity of transglutaminase-catalyzed epsilon-(gamma-glutamyl) lysine bond formation. Adv Enzymol Relat Areas Mol Biol. (1983) 54:1–56. doi: 10.1002/9780470122990.ch1
46. Ciccocioppo R, Di Sabatino A, Corazza GR. The immune recognition of gluten in coeliac disease. Clin Exp Immunol. (2005) 140:408–16. doi: 10.1111/j.1365-2249.2005.02783.x
47. Di Venere A, Rossi A, De Matteis F, Rosato N, Agrò AF, Mei G. Opposite effects of Ca(2+) and GTP binding on tissue transglutaminase tertiary structure. J Biol Chem. (2000) 275:3915–21. doi: 10.1074/jbc.275.6.3915
48. Petterson A, Sjöberg K, Lernmark Å, Eriksson S. HLA genotypes in coeliac disease and in healthy individuals carrying gliadin antibodies. Eur J Gastroenterol Hepatol. (1993) 5:445–50. doi: 10.1097/00042737-199306000-00009
49. Mäki M. The humoral immune system in coeliac disease. Baillieres Clin Gastroenterol. (1995) 9:231–49.
50. Björck S, Brundin C, Lörinc E, Lynch KF, Agardh D. Screening detects a high proportion of celiac disease in young HLA-genotyped children. J Pediatr Gastroenterol Nutr. (2010) 50:49–53. doi: 10.1097/MPG.0b013e3181b477a6
51. Sulkanen S, Halttunen T, Laurila K, Kolho KL, Korponay-Szab? IR, Sarnesto A, et al. Tissue transglutaminase autoantibody enzyme-linked immunosorbent assay in detecting celiac disease. Gastroenterology. (1998) 115:1322–8. doi: 10.1016/S0016-5085(98)70008-3
52. Basso D, Guariso G, Plebani M. Serologic testing for celiac disease. Clin Chem. (2002) 48:2082–3. doi: 10.1093/clinchem/48.11.2082
53. Sollid LM, Molberg O, McAdam S, Lundin KE. Autoantibodies in coeliac disease: tissue transglutaminase–guilt by association? Gut. (1997) 41:851–2. doi: 10.1136/gut.41.6.851
54. Giersiepen K, Lelgemann M, Stuhldreher N, Ronfani L, Husby S, Koletzko S, et al. Accuracy of diagnostic antibody tests for coeliac disease in children: summary of an evidence report. J Pediatr Gastroenterol Nutr. (2012) 54:229–41. doi: 10.1097/MPG.0b013e318216f2e5
55. Sugai E, Nachman F, Váquez H, González A, Andrenacci P, Czech A, et al. Dynamics of celiac disease-specific serology after initiation of a gluten-free diet and use in the assessment of compliance with treatment. Dig Liver Dis. (2010) 42:352–8. doi: 10.1016/j.dld.2009.07.011
56. Picarelli A, Maiuri L, Frate A, Greco M, Auricchio S, Londei M. Production of antiendomysial antibodies after in-vitro gliadin challenge of small intestine biopsy samples from patients with coeliac disease. Lancet. (1996) 348:1065–7. doi: 10.1016/S0140-6736(96)03060-7
57. Marzari R, Sblattero D, Florian F, Tongiorgi E, Not T, Tommasini A, et al. Molecular dissection of the tissue transglutaminase autoantibody response in celiac disease. J Immunol. (2001) 166:4170–6. doi: 10.1016/0950-3528(95)90030-6
58. Di Niro R, Mesin L, Zheng NY, Stamnaes J, Morrissey M, Lee JH, et al. High abundance of plasma cells secreting transglutaminase 2-specific IgA autoantibodies with limited somatic hypermutation in celiac disease intestinal lesions. Nat Med. (2012) 18:441–5. doi: 10.1038/nm.2656
59. Hietikko M, Koskinen O, Kurppa K, Laurila K, Saavalainen P, Salmi T, et al. Small-intestinal TG2-specific plasma cells at different stages of coeliac disease. BMC Immunol. (2018) 19:36. doi: 10.1186/s12865-018-0275-7
60. Steinsbø Ø, Henry Dunand CJ, Huang M, Mesin L, Salgado-Ferrer M, Lundin KE, et al. Restricted VH/VL usage and limited mutations in gluten-specific IgA of coeliac disease lesion plasma cells. Nat Commun. (2014) 5:4041. doi: 10.1038/ncomms5041
61. Di Niro R, Snir O, Kaukinen K, Yaari G, Lundin KE, Gupta NT, et al. Responsive population dynamics and wide seeding into the duodenal lamina propria of transglutaminase-2-specific plasma cells in celiac disease. Mucosal Immunol. (2016) 9:254–64. doi: 10.1038/mi.2015.57
62. Høydahl LS, Richter L, Frick R, Snir O, Gunnarsen KS, Landsverk OJB, et al. Plasma cells are the most abundant gluten peptide mhc-expressing cells in inflamed intestinal tissues from patients with celiac disease. Gastroenterology. (2019) 156:1428–1439.e10. doi: 10.1053/j.gastro.2018.12.013
63. Iversen R, Snir O, Stensland M, Kroll JE, Steinsbø Ø, Korponay-Szabó IR, et al. Strong clonal relatedness between serum and gut IgA despite different plasma cell origins. Cell Rep. (2017) 20:2357–67. doi: 10.1016/j.celrep.2017.08.036
64. Iversen R, Di Niro R, Stamnaes J, Lundin KE, Wilson PC, Sollid LM. Transglutaminase 2-specific autoantibodies in celiac disease target clustered, N-terminal epitopes not displayed on the surface of cells. J Immunol. (2013) 190:5981–91. doi: 10.4049/jimmunol.1300183
65. Iversen R, Roy B, Stamnaes J, Høydahl LS, Hnida K, Neumann RS, et al. Efficient T cell-B cell collaboration guides autoantibody epitope bias and onset of celiac disease. Proc Natl Acad Sci USA. (2019) 116:15134–9. doi: 10.1073/pnas.1901561116
66. Iversen R, Fleur du Pré M, Di Niro R, Sollid LM. Igs as substrates for transglutaminase 2: implications for autoantibody production in celiac disease. J Immunol. (2015) 195:5159–68. doi: 10.4049/jimmunol.1501363
67. Maglio M, Florian F, Vecchiet M, Auricchio R, Paparo F, Spadaro R, et al. Majority of children with type 1 diabetes produce and deposit anti-tissue transglutaminase antibodies in the small intestine. Diabetes. (2009) 58:1578–84. doi: 10.2337/db08-0962
68. Zanoni G, Navone R, Lunardi C, Tridente G, Bason C, Sivori S, et al. In celiac disease, a subset of autoantibodies against transglutaminase binds toll-like receptor 4 and induces activation of monocytes. PLoS Med. (2006) 3:e358. doi: 10.1371/journal.pmed.0030358
69. Halttunen T, Mäki M. Serum Immunoglobulin A from patients with celiac disease inhibit human T84 intestinal crypt epithelial cell differentiation. Gastroenterology. (1999) 116:566–72. doi: 10.1016/S0016-5085(99)70178-2
70. Barone MV II, Caputo I, Ribecco MT, Maglio M, Marzari R, Sblattero D, et al. Humoral immune response to tissue transglutaminase is related to epithelial cell proliferation in celiac disease. Gastroenterology. (2007) 132:1245–53. doi: 10.1053/j.gastro.2007.01.030
71. Caputo I, Barone MV, Lepretti M, Martucciello S, Nista I, Troncone R, et al. Celiac anti-tissue transglutaminase antibodies interfere with the uptake of alpha gliadin peptide 31-43 but not of peptide 57-68 by epithelial cells. Biochim Biophys Acta. (2010) 1802:717–27. doi: 10.1016/j.bbadis.2010.05.010
72. Esposito C, Paparo F, Caputo I, Rossi M, Maglio M, Sblattero D, et al. Anti-tissue transglutaminase antibodies from coeliac patients inhibit transglutaminase activity both in vitro and in situ. Gut. (2002) 51:177–81. doi: 10.1136/gut.51.2.177
73. Dieterich W, Trapp D, Esslinger B, Leidenberger M, Piper J, Hahn E, et al. Autoantibodies of patients with coeliac disease are insufficient to block tissue transglutaminase activity. Gut. (2003) 52:1562–6. doi: 10.1136/gut.52.11.1562
74. Roth EB, Sjöberg K, Stenberg P. Biochemical and immuno-pathological aspects of tissue transglutaminase in celiac disease. Autoimmunity. (2003) 36:221–6. doi: 10.1080/0891693031000118974
75. Király R, Vecsei Z, Deményi T, Korponay-Szabó IR, Fésüs L. Coeliac autoantibodies can enhance transamidating and inhibit GTPase activity of tissue transglutaminase: dependence on reaction environment and enzyme fitness. J Autoimmun. (2006) 26:278–87. doi: 10.1016/j.jaut.2006.03.002
76. Byrne G, Ryan F, Jackson J, Feighery C, Kelly J. Mutagenesis of the catalytic triad of tissue transglutaminase abrogates celiac disease serum IgA autoantibody binding. Gut. (2007) 56:336–41. doi: 10.1136/gut.2006.092908
77. Caputo I, Lepretti M, Secondo A, Martucciello S, Paolella G, Sblattero D, et al. Anti-tissue transglutaminase antibodies activate intracellular tissue transglutaminase by modulating cytosolic Ca2+ homeostasis. Amino Acids. (2013) 44:251–60. doi: 10.1007/s00726-011-1120-y
78. Reunala T, Salmi TT, Hervonen K, Laurila K, Kautiainen H, Collin P, et al. IgA antiepidermal transglutaminase antibodies in dermatitis herpetiformis: a significant but not complete response to a gluten-free diet treatment. Br J Dermatol. (2015) 172:1139–41. doi: 10.1111/bjd.13387
79. Hadjivassiliou MI, Aeschlimann P, Sanders DS, Mäki M, Kaukinen K, Grünewald RA, et al. Transglutaminase 6 antibodies in the diagnosis of gluten ataxia. Neurology. (2013) 80:1740–5. doi: 10.1212/WNL.0b013e3182919070
80. Mawhinney H, Love AH. Anti-reticulin antibody in jejunal juice in coeliac disease. Clin Exp Immunol. (1975) 21:394–8.
81. Picarelli A, Sabbatella L, Di Tola M, Di CT, Vetrano S, Anania MC. Antiendomysial antibody detection in fecal supernatants: in vivo proof that small bowel mucosa is the site of antiendomysial antibody production. Am J Gastroenterol. (2002) 97:95–8. doi: 10.1111/j.1572-0241.2002.05426.x
82. Carroccio A, Iacono G, D'Amico D, Cavataio F, Teresi S, Caruso C, et al. Production of antiendomysial antibodies in cultured duodenal mucosa: usefulness in coeliac disease diagnosis. Scand J Gastroenterol. (2002) 37:32–8. doi: 10.1080/003655202753387329
83. Picarelli A, Libanori V, De Nitto D, Saponara A, Di Tola M, Donato G. Organ culture system as a means to detect celiac disease. Ann Clin Lab Sci. (2010) 40:85–7.
84. Tosco A, Aitoro R, Auricchio A, Ponticelli D, Miele E, Paparo F, et al. Intestinal anti-tissue transglutaminase antibodies in potential coeliac disease. Clin Exp Immunol. (2013) 171:69–75. doi: 10.1111/j.1365-2249.2012.04673.x
85. Tosco A, Auricchio R, Aitoro R, Ponticelli D, Primario M, Miele E, et al. Intestinal titres of anti-tissue transglutaminase 2 antibodies correlate positively with mucosal damage degree and inversely with gluten-free diet duration in coeliac disease. Clin Exp Immunol. (2014) 177:611–7. doi: 10.1111/cei.12366
86. Korponay-Szabo IR, Halttunen T, Szalai Z, Laurila K, Király R, Kovács JB, et al. In vivo targeting of intestinal and extraintestinal transglutaminase 2 by coeliac autoantibodies. Gut. (2004) 53:641–8. doi: 10.1136/gut.2003.024836
87. Maglio M, Tosco A, Auricchio R, Paparo F, Colicchio B, Miele E, et al. Intestinal deposits of anti-tissue transglutaminase IgA in childhood celiac disease. Dig Liver Dis. (2011) 43:604–8. doi: 10.1016/j.dld.2011.01.015
88. Picarelli A, Di Tola M, Marino M, Libanori V, Borghini R, Salvi E, et al. Usefulness of the organ culture system when villous height/crypt depth ratio, intraepithelial lymphocyte count, or serum antibody tests are not diagnostic for celiac disease. Transl Res. (2013) 161:172–80. doi: 10.1016/j.trsl.2012.10.010
89. De Leo L, Bramuzzo M, Ziberna F, Villanacci V, Martelossi S, Leo GD, et al. Diagnostic accuracy and applicability of intestinal auto-antibodies in the wide clinical spectrum of coeliac disease. EBioMedicine. (2020) 51:102567. doi: 10.1016/j.ebiom.2019.11.028
90. Carroccio A, Di Prima L, Pirrone G, Scalici C, Florena AM, Gasparin M, et al. Anti-transglutaminase antibody assay of the culture medium of intestinal biopsy specimens can improve the accuracy of celiac disease diagnosis. Clin Chem. (2006) 52:1175–80. doi: 10.1373/clinchem.2005.061366
91. Maglio M, Ziberna F, Aitoro R, Discepolo V, Lania G, Bassi V, et al. Intestinal production of anti-tissue transglutaminase 2 antibodies in patients with diagnosis other than celiac Disease. Nutrients. (2017) 9:1050. doi: 10.3390/nu9101050
92. Salmi TT, Collin P, Reunala T, Mäki M, Kaukinen K. Diagnostic methods beyond conventional histology in celiac disease diagnosis. Dig Liver Dis. (2010) 42:28–32. doi: 10.1016/j.dld.2009.04.004
93. Maglio M, Tosco A, Paparo F, Auricchio R, Granata V, Colicchio B, et al. Serum and intestinal celiac disease-associated antibodies in children with celiac disease younger than 2 years of age. J Pediatr Gastroenterol Nutr. (2010) 50:43–8. doi: 10.1097/MPG.0b013e3181b99c8f
94. Not T, Ziberna F, Vatta S, Quaglia S, Martelossi S, Villanacci V, et al. Cryptic genetic gluten intolerance revealed by intestinal antitransglutaminase antibodies and response to gluten-free diet. Gut. (2011) 60:1487–93. doi: 10.1136/gut.2010.232900
95. Kappler M, Krauss-Etschmann S, Diehl V, Zeilhofer H, Koletzko S. Detection of secretory IgA antibodies against glidin and human tissue transglutaminase in stool to screen for coeliac disease in children: validation study. BMJ. (2006) 332:213–4. doi: 10.1136/bmj.38688.654028.AE
96. Tosco A, Salvati VM, Auricchio R, Maglio M, Borrelli M, Coruzzo A, et al. Natural history of potential celiac disease in children. Clin Gastroenterol Hepatol. (2011) 9:320–5. doi: 10.1016/j.cgh.2010.09.006
97. Koskinen O, Collin P, Korponay-Szabo I, Salmi T, Iltanen S, Haimila K, et al. Gluten-dependent small bowel mucosal transglutaminase 2-specific IgA deposits in overt and mild enteropathy coeliac disease. J Pediatr Gastroenterol Nutr. (2008) 47:436–42. doi: 10.1097/MPG.0b013e31817b6dec
98. Koskinen O, Collin P, Lindfors K, Laurila K, Mäki M, Kaukinen K. Usefulness of small-bowel mucosal transglutaminase-2 specific autoantibody deposits in the diagnosis and follow-up of celiac disease. J Clin Gastroenterol. (2010) 44:483–8. doi: 10.1097/MCG.0b013e3181b64557
99. Kurppa K, Ashorn M, Iltanen S, Koskinen LL, Saavalainen P, Koskinen O, et al. Celiac disease without villous atrophy in children: a prospective study. J Pediatr. (2010) 157:373–80.380.e1. doi: 10.1016/j.jpeds.2010.02.070
100. Salmi TT, Hervonen K, Laurila K, Collin P, Mäki M, Koskinen O, et al. Small bowel transglutaminase 2-specific IgA deposits in dermatitis herpetiformis. Acta Derm Venereol. (2014) 94:393–7. doi: 10.2340/00015555-1764
101. Hadjivassiliou M, Mäki M, Sanders DS, Williamson CA, Grünewald RA, Woodroofe NM, et al. Autoantibody targeting of brain and intestinal transglutaminase in gluten ataxia. Neurology. (2006) 66:373–7. doi: 10.1212/01.wnl.0000196480.55601.3a
102. Kaukinen K, Peräaho M, Collin P, Partanen J, Woolley N, Kaartinen T, et al. Small-bowel mucosal transglutaminase 2-specific IgA deposits in coeliac disease without villous atrophy: a prospective and randomized clinical study. Scand J Gastroenterol. (2005) 40:564–72. doi: 10.1080/00365520510023422
103. Salmi TT I, Collin P, Jarvinen O, Haimila K, Partanen J, Laurila K, et al. Immunoglobulin A autoantibodies against transglutaminase 2 in the small intestinal mucosa predict forthcoming celiac disease. Aliment Pharmacol Ther. (2006) 24:541–52. doi: 10.1111/j.1365-2036.2006.02997.x
104. Natvig IB, Johansen FE, Nordeng TW, Haraldsen G, Brandtzaeg P. Mechanism for enhanced external transfer of dimeric IgA over pentameric IgM: studies of diffusion, binding to the human polymeric Ig receptor, and epithelial transcytosis. J Immunol. (1997) 159:4330–40.
105. Borrelli M, Maglio M, Agnese M, Paparo F, Gentile S, Colicchio B, et al. High density of intraepithelial gammadelta lymphocytes and deposits of immunoglobulin (Ig)M anti-tissue transglutaminase antibodies in the jejunum of coeliac patients with IgA deficiency. Clin Exp Immunol. (2010) 160:199–206. doi: 10.1111/j.1365-2249.2009.04077.x
106. Tursi A, Brandimarte G, Giorgetti GM. Lack of usefulness of antitransglutaminase antibodies in assessing histologic recovery after gluten-free diet in celiac disease. J Clin Gastroenterol. (2003) 37:387–91. doi: 10.1097/00004836-200311000-00007
107. Fabiani E, Catassi C, International Working Group. The serum IgA class anti-tissue transglutaminase antibodies in the diagnosis and follow up of coeliac disease. Results of an international multicenter study. International working group on Eu-tTG. Eur J Gastroenterol Hepatol. (2001) 13:659–65. doi: 10.1097/00042737-200106000-00008
108. Vahedi K, Mascart F, Mary JY, Laberenne JE, Bouhnik Y, Morin MC, et al. Reliability of antitransglutaminase antibodies as predictors of gluten-free diet compliance in adult celiac disease. Am J Gastroenterol. (2003) 98:1079–87. doi: 10.1111/j.1572-0241.2003.07284.x
109. Koskinen O II, Lindfors K, Collin P, Peräaho M, Laurila K, Woolley N, et al. Intestinal transglutaminase 2 specific antibody deposits in non-responsive coeliac disease. Dig Liver Dis. (2010) 42:692–7. doi: 10.1016/j.dld.2010.03.008
110. Risnes LF, Christophersen A, Dahal-Koirala S, Neumann RS, Sandve GK, Sarna VK, et al. Disease-driving CD4+ T cell clonotypes persist for decades in celiac disease. J Clin Invest. (2018) 128:2642–2265. doi: 10.1172/JCI98819
111. Borrelli M, Maglio M, Korponay-Szabó IR, Vass V, Mearin ML, Meijer C, et al. Intestinal anti-transglutaminase 2 immunoglobulin A deposits in children at risk for coeliac disease (CD): data from the PreventCD study. Clin Exp Immunol. (2018) 191:311–7. doi: 10.1111/cei.13078
112. Vriezinga SL, Auricchio R, Bravi E, Castillejo G, Chmielewska A, Crespo Escobar P, et al. Randomized feeding intervention in infants at high risk for celiac disease. N Engl J Med. (2014) 371:1304–15. doi: 10.1056/NEJMoa1404172
113. Salmi TT I, Collin P, Korponay-Szabó IR, Laurila K, Partanen J, Huhtala H, et al. Endomysial antibody-negative coeliac disease: clinical characteristics and intestinal autoantibody deposits. Gut. (2006) 55:1746–53. doi: 10.1136/gut.2005.071514
114. Quaglia S, De Leo L, Ziberna F, Vatta S, Villanacci V, Granzotto M, et al. Intestinal-mucosa anti-transglutaminase antibody assays to test for genetic gluten intolerance. Cell Mol Immunol. (2014) 11:617–20. doi: 10.1038/cmi.2014.32
115. Sblattero D, Ventura A, Tommasini A, Cattin L, Martelossi S, Florian F, et al. Cryptic gluten intolerance in type 1 diabetes: identifying suitable candidates for a gluten free diet. Gut. (2006) 55:133. doi: 10.1136/gut.2005.077511
116. Iltanen S, Holm K, Ashorn M, Ruuska T, Laippala P, Mäki M. Changing jejunal gamma/delta+ T cells receptor (TCR)-bearing intraepithelial lymphocyte density in celiac disease. Clin Exp Immunol. (1999) 117:51–5. doi: 10.1046/j.1365-2249.1999.00948.x
Keywords: intestinal anti-TG2 antibodies, autoimmunity, gluten, celiac disease, intestinal production of anti-TG2
Citation: Maglio M and Troncone R (2020) Intestinal Anti-tissue Transglutaminase2 Autoantibodies: Pathogenic and Clinical Implications for Celiac Disease. Front. Nutr. 7:73. doi: 10.3389/fnut.2020.00073
Received: 24 January 2020; Accepted: 28 April 2020;
Published: 29 May 2020.
Edited by:
Gianfranco Mamone, Italian National Research Council, ItalyReviewed by:
Shuo-Wang Qiao, University of Oslo, NorwayLei Shi, Georgia State University, United States
Copyright © 2020 Maglio and Troncone. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mariantonia Maglio, bWFyaWFudG9uaWEubWFnbGlvQHVuaW5hLml0