 Graciela C. Rollán
Graciela C. Rollán Carla L. Gerez
Carla L. Gerez Jean G. LeBlanc
Jean G. LeBlanc- Centro de Referencia para Lactobacilos (CERELA) - CONICET, San Miguel de Tucumán, Argentina
One of the greatest challenges is to reduce malnutrition worldwide while promoting sustainable agricultural and food systems. This is a daunting task due to the constant growth of the population and the increasing demands by consumers for functional foods with higher nutritional values. Cereal grains are the most important dietary energy source globally; wheat, rice, and maize currently provide about half of the dietary energy source of humankind. In addition, the increase of celiac patients worldwide has motivated the development of gluten-free foods using alternative flour types to wheat such as rice, corn, cassava, soybean, and pseudocereals (amaranth, quinoa, and buckwheat). Amaranth and quinoa have been cultivated since ancient times and were two of the major crops of the Pre-Colombian cultures in Latin- America. In recent years and due to their well-known high nutritional value and potential health benefits, these pseudocereals have received much attention as ideal candidates for gluten-free products. The importance of exploiting these grains for the elaboration of healthy and nutritious foods has forced food producers to develop novel adequate strategies for their processing. Fermentation is one of the most antique and economical methods of producing and preserving foods and can be easily employed for cereal processing. The nutritional and functional quality of pseudocereals can be improved by fermentation using Lactic Acid Bacteria (LAB). This review provides an overview on pseudocereal fermentation by LAB emphasizing the capacity of these bacteria to decrease antinutritional factors such as phytic acid, increase the functional value of phytochemicals such as phenolic compounds, and produce nutritional ingredients such as B-group vitamins. The numerous beneficial effects of lactic fermentation of pseudocereals can be exploited to design novel and healthier foods or grain ingredients destined to general population and especially to patients with coeliac disease.
Introduction
According to the Food and Agriculture Organization (FAO), global hunger is in ascent again after constantly decreasing for over a decade (1). The number of chronically undernourished people in the world is estimated to have increased to 815 million (11% of the global population) in 2016, up from 777 million in 2015, as reported in the edition of the Annual United Nations on World Food Security and Nutrition published in September 2017, based on reports by five organizations [ONU, FAO (Food and Agriculture Organization), WHO (World Health Organization), IFAD (International Fund for Agricultural Development (IFAD), and World Food Program (WFP)] (1). At the same time, different forms of malnutrition are threatening the health of millions of people worldwide. Nearly 795 million people have eating disorders and do not carry out healthy and active lifestyles with an estimated 41 million children that are now overweight according to the World Food Program. Added to this serious situation, the world population is expected to reach nine billion persons in the coming decades, imposing the need for urgent solutions to increase food supplies (2). In addition, climate change is rapidly degrading the conditions of crop production, affecting the availability of water and arable land, increasing salinization and aridity, generating a serious problem in the yield of food. It is estimated that approximately one billion hectares or crop land will be affected worldwide due to these problems, especially those in the hottest and most arid regions of the world (3–7). In addition to climate change, global staple crop production is also threatened by restrictions such as accelerated erosion of soil and natural resources (8). Frison et al. (9) also reported that modern agriculture generates serious problems in the environment causing soil degradation and erosion, water pollution and biodiversity decline. Therefore, it is essential to increase food production for a growing population that uses low input regimes. The FAO urges to expand the response to climate change in agriculture. According to their 2017 document “A systemic approach that involves the relevant agricultural and food sectors and those interested in the adoption of agroecology, has the potential to greatly accelerate the transition to sustainable and resilient food systems, in line with the various international commitments assumed by the member countries.” Agroecology, in an integral manner, can support the execution of different social, economic, environmental, nutritional, and health objectives.
Diets throughout the world are based on two dozen crops with a dominant proportion of the “big three” cereals: wheat (Triticum aestivum), maize (Zea mays), and rice (Oryza sativa), which contribute to approximately 60% of the total caloric intake (10). However, these crops may not intrinsically be the best-suited species to face up to extreme weather events that are becoming more frequent due to climate change; thus, world grain production per capita is expected to decline by at least 14% between 2008 and 2030 (11). The rapid growth of the world population and per capita food consumption worldwide puts great pressure on the food industry to produce more food (12). The food supply must double by 2050 to counterbalance the effects of climate change and population pressure on global food systems and thus novel food sources must be found (6). Less than 0.6% of plant species that are suitable for human consumption have reached the world markets (13). The diversification of main crops and the systems in which they grow is essential for agriculture to be sustainable, resilient, and suitable for local environments and soils in the future. One critical measure to ensure future food availability for all is to provide more diverse food sources and develop agricultural systems that are resistant to climate change. Furthermore, the new challenge for the food industries and scientific areas such as chemistry, biology, medicine, pharmacology, and food technology is to obtain foods with a higher nutritional value that also possess functional properties which go beyond traditional health requirements. In response to this issue, one of the leading strategies is unlocking the potential of underutilized crops. Most of these crops have high nutritional value, resilience traits, with the ability to withstand drought, flooding, extreme temperatures, and pests and diseases better than current major staples and thus they should be investigated, developed, and now more than ever used (14). Current emphasis is now placed on the use of ancient cereals and pseudocereals that include amaranth, buckwheat, quinoa, teff, millets amongst others.
The aim of this review is to highlight certain nutritional and functional properties of pseudocereals and how lactic acid fermentation can be used as an advantageous biotechnological strategy to improve the natural potential of these grains. This review provides an overview on pseudocereal fermentation by lactic acid bacteria (LAB) emphasizing the capacity of these bacteria to decrease antinutritional factors such as phytic acid, increase the functional value of phytochemicals such as phenolic compounds, and produce nutritional ingredients such as B-group vitamins.
Crop Diversification: Pseudocereals
Crop diversification is an important strategy to protect global food supplies and to fight against malnutrition. Sustainable diets should provide nutritious food at affordable costs, while having a low impact on the environment (15). Effective analysis of sustainable plant resources is an important assignment for ensuring global food security in the future (16).
The need for the diversification of grains for human consumption and the consumer's demands for gluten-free and more nutritious products caused the resurgence and valoration of underutilized crops, so-called minor grains such as sorghum, millets, and pseudocereals through the world during the last several decades (17). In the “International AACC list of recognized grains” pseudocereals are also mentioned (18) where the most important species are quinoa (Chenopodium quinoa Willd), amaranth (Amaranthus sp.), and buckwheat (Fagopyrum esculentum). Pseudocereals are dicotyledonous species unlike true cereals (Poaceae family), that are monocotyledonous species. Pseudocereals are known as such since they are similar to cereals in their physical appearance and their seeds are edible with high starch content that can be milled into flour (19). Their high nutritional value is mostly due to their elevated content and quality of proteins (20, 21).
Celiac disease (CD) is one of the most common lifelong disorders worldwide with as estimated mean prevalence of 1% of the general population (22). The increase of celiac patients throughout the world has led to intensify the search for alternative flours to wheat (19). The development of gluten-free (GF) products is therefore essential and poses novel challenges for food producers (23). In the last decade, due to the pseudocereals characteristics, GF and good nutritional advantages, the use of these grains has increased for their addition in healthy diets especially for people allergic to cereals. Thus, the integration of these grains into GF diets could be a valuable contribution for improving the quality of the existing GF products, which have been mainly based on rice and maize flour (24). Despite the fact that the interest in pseudocereals due to its high nutritional value has increased, only a few products including these grains are available on the market [Figure 1; (25)].
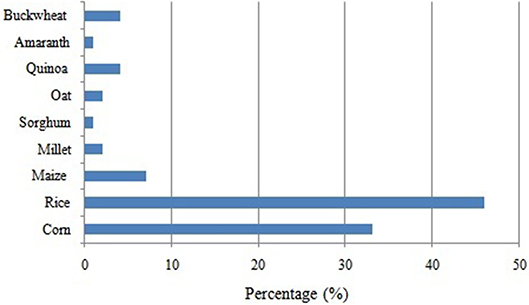
Figure 1. Principal grains used for gluten-free products on the market [adapted from (25)].
Pseudocereals Historical Background
Pseudocereals nourished the Native Americans populations and allowed them to increase their endurance and mental development; because of their properties, Mayans and Incas considered these grains sacred. The conquest of America meant not only a political and social domination of indigenous civilizations, but also a change in their feeding habits. Jacobsen (26) reported that quinoa is one of the oldest crops in the Andean region, having been grown for approximately 7000 years; it is considered the principal crop of the pre-Columbian cultures in Latin America (27, 28). The Incas called quinoa “the mother grain” for many reasons: (i) it is one of the few crops able to grow in high salt soils in Southern Bolivia and Northern Chile, (ii) it has high tolerance to abiotic stresses, (iii) it grows in soils with water scarcity, and (iv) it is resistant to extreme temperatures (−4 to 38°C) and harsh climate conditions (15, 27, 29, 30). The FAO (United Nations) declared, “quinoa has the balance of proteins and nutrients closest to the ideal food for humans.”
Amaranth (Amaranthus sp.) is an ancient crop consumed during the Mayan and Aztec periods. It was called “the Inca wheat” by the Spanish conquerors. Amaranth grain species are annual herbaceous plants native of America but they are also now distributed in Asia and Africa (31, 32). Amaranthus caudatus was discovered in the north of Argentina (Salta) 2000 years ago (33). When the Spaniards arrived, they decided to exterminate pseudocereals because of their religious implications. Ironically, it is now Europeans who teach us how to consume the grains that were used by the Native American civilizations. The interest in these Andean ancestral crops in the world has led to an increase in their cultivation and production in recent years.
Buckwheat (Fagopyrum esculentum Moench) has its origin in Asia and it is believed to have been cultivated in China during the fifth and sixth centuries. It came to Europe after some 800–900 years and to North America in seventeenth century (34).
Underutilized species by means of sustainable intensification, adaptation and mitigation can accelerate the process to obtain climate-smart agriculture [Figure 2; (35)].
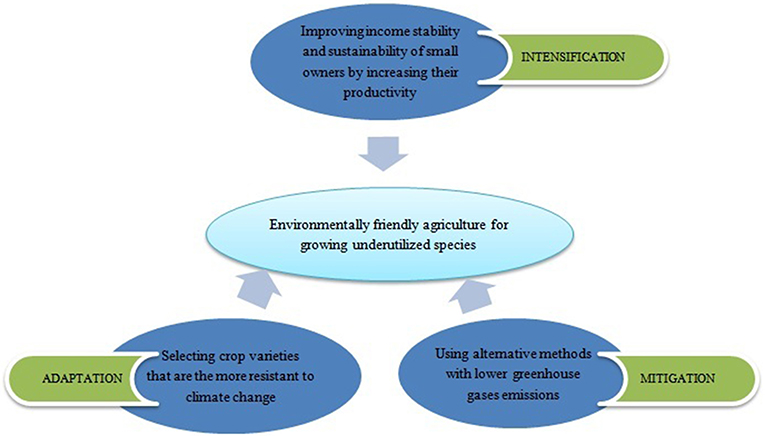
Figure 2. Strategies to use underutilized species and increase the crops biodiversification for a sustainable agriculture [adapted from (35)].
Nutritional Value
Charalampopoulos et al. (36) reported that 73% of the total world harvested area corresponds to cereal crops and contributes to more than 60% of the world food production, furnishing proteins, minerals, dietary fiber, and vitamins necessary for human health. Cereals contribute around 50% of the mean daily energy intake in most populations, and 70% in some developing countries, converting them into one of the most important sources of energy in the world (37). However, most grains are, to a greater or lesser extent, deficient in a number of elemental nutrients such as the essential amino acids threonine, lysine, and tryptophan. Their protein digestibility is also lower than that of animal origin, due partially to the presence of phytic acid, tannins, and polyphenols which bind to protein thus making them indigestible (38). Pseudocereals in turn have been described as “the grains of the twenty-first century” (39, 40). The FAO classified quinoa as one of humanity's promising crops destined to contribute to food security in the twenty-first century by its high nutritive potential and genetic diversity [Food and Agriculture Organization Regional Office for Latin America and PROINPA, (27, 41)]. Quinoa and amaranth have tender leaves that are used in food preparation; however, it is their grains that attract the most interest due to their high nutritional value. They are rich in proteins of excellent quality with a balanced essential amino acid composition that include abundant amounts of sulfur- rich amino acids (42). They are also a good source of minerals (calcium, iron and zinc), vitamins, and natural antioxidants (43). They are a significant source of compounds such as flavonoids, polyphenols and phytosterols with potential nutraceutical benefits. From a food provision perspective, pseudocereals are potentially important crops due to their properties (exceptional nutritional value, ability to grow in dry conditions, and their resilience to climatic conditions). Several reviews have reported the nutritional value of Andean grains (19, 43–49).
The General Assembly of the United Nations declared 2013 as the International Year of Quinoa (IYQ), with the goal of focalizing global attention on the role it can perform in contributing to food security, nutrition, and poverty eradication (50, 51). The rapid expansion of the harvested area, with a doubling of countries from 2013, is rapidly changing the perception and representation of quinoa from a minor to a potential major crop. The excellent properties of quinoa led to this ancestral grain to be considered a possible crop in NASA's Controlled Ecological Life Support System for long-duration manned space flights (39).
Vitamin in Pseudocereals
Vitamins are essential micronutrients since only small quantities are required for adequate growth and function of numerous metabolic reactions. Vitamins are divided based on their solubility in fat (A, D, E, and K), or in water (C and the B-group vitamins). Since there are no foods that contain all vitamins, there is a worldwide increase in their deficiencies due to unbalanced diets. Other causes of vitamin deficiencies are malabsorption that can be due to certain drug treatments or diseases, by the presence of antinutritional factors found in certain foods. Although most vitamins are present in cereals and pseudocereals, a large portion of water-soluble vitamins are lost during processing and cooking, especially when water is used for grain soaking. In this sense, many countries have adopted mandatory fortification programs with specific vitamins and minerals. Folic acid is frequently added in foods of mass consumption (such as different flours) in order to prevent deficiencies in the general population. However, the chemical form of the vitamin used in these programs is controversial. Folic acid, a chemical derivative of folates, is not found in nature and can cause many side effects, especially masking vitamin B12 deficiency and affecting the activity of certain liver enzymes, but also has been associated with increased risks of colon and prostate cancers (52). Natural folate, present in numerous different chemical forms in vegetables or produced by certain microorganisms does not cause these undesirable side effects. For this reason, more and more researchers are now searching for more natural methods to increase water soluble vitamins such as folate and riboflavin in foods to not only prevent deficiencies, but also to reduce the use of chemical additives in the food chain.
It was demonstrated that staple foods produced from amaranth contained total folate contents of 35.5 μg/100 g in bread, 36.3 μg/100 g in cookies, and 38.9 μg/100 g in noodles, whereas when wheat was used, breads contained only 12.0 μg/100 g (53). The riboflavin content in amaranth flour is in the range of 0.29–0.32 mg/100 g, which is about 10-fold higher than that of wheat (54). In general, significant reductions of all vitamins take place during processing, which affect the nutritional value of the products (55). The production of fermented food products with high levels of B-group vitamins increases their commercial and nutritional value and eliminates the need for fortification (56).
Phytochemical Profile of Pseudocereals
Pseudocereals are important phytochemical sources in the diet. Like cereal, these grains contain a great amount of functional phytochemicals including the phenolic compounds (PC) (57–60). The PC constitute a group of secondary metabolites with important functions in cereals and pseudocereals. The chemical structures of these compounds include an aromatic ring with one or more hydroxyl substituents, and vary from simple phenolic molecules to highly polymerised compounds (61). PC are broadly divided in four classes; phenolic acids (benzoic or hydroxycinnamic acid derivatives), flavonoids (flavonols, flavones, isoflavones, flavanones, and anthocyanidins), stilbenes and lignans. In addition to this diversity, polyphenols may be associated with various carbohydrates and organic acids (60). In general, ferulic, p-coumaric, caffeic, isoferulic, vanillic, sinapic, p-hydroxybenzoic, syringic, and protocatechuic acids are present in all grains; with ferulic acid the most abundant phenolic acid (62, 63). Gorinstein et al. (64, 65) reported a high content of polyphenols, anthocyanins and flavonoids in pseudocereals such as buckwheat, quinoa and amaranth. Likewise, the highest amount of PC was reported in quinoa (490.2 mg/kg DW), slightly lower in amaranth v. Aztek (464 mg/kg DW), and the lowest in amaranth v. Rawa (424.6 mg/kg DW) (66).
PC may provide health benefits to humans since they are associated with a reduced risk of chronic diseases such as anti-allergenic, anti-inflammatory, anti-microbial, antioxidant, anti-thrombotic, cardioprotective, and stimulates insulin secretion in diabetes mellitus type 2 (57, 67–71). The dietary PC contribute to the maintenance of a healthy gut by modulating the gut microbial balance (beneficial bacteria/pathogen bacteria). Metagenomic and metabolomic studies providing more insight into the health effects of PC in humans are needed to understand the dietary PC/gut microbiota relationship and their mechanisms of action. The PC effect on the modulation of the gut ecology and the two-way relationship “polyphenols ↔ microbiota” is currently being studied (72).
The biological effects of PC depend principally on their bioaccessibility (release of the food matrix in an absorbable form during digestion) and bioavailability (absorption and transference to the bloodstream), and both depend on their chemical structure, matrix interactions, antioxidant activity, and food processing (73–79). Natural PC usually occur as glycosides, esters or polymers that have no biological activity (80). Of the total PC intake, only 5–10% is absorbed in the small intestine and the remaining PC (90–95%) accumulate in the large intestinal lumen where they are subjected to the enzymatic activities of the gut microbial community (81). Food technologists need to find the operating conditions to increase bioaccessibility and bioavailability of PC from the matrix. The addition of purified enzymes such as feruloyl and p-coumaryl esterases, xylanase, β-glucanase, and α-amylase from natural sources has been proposed to increase the active PC content in cereals [i.e., wheat and rye; (82–84)]. However, these studies in pseudocereals are still missing.
Antinutritive Factors in Pseudocereals
Grains of cereals, pseudocereals, and legumes are of global importance in the feeding of monogastric animals (humans and domestic animals) since they are a good source of proteins, bioactive compounds and trace elements (85). However, they contain certain antinutrients compounds, such as phytic acid, saponins, tannins, polyphenols, and protease inhibitors (86). In this sense the bioavailability of minerals in whole grain foods is negatively affected by the presence of phytate (87). Since phytate is an antinutritional factor that is found in the highest quantities in pseudocereals and due to its important negative effect on malnutrition, this review will focus on this antinutritive factor.
Phytates
Phytic acid (PA) (myo-inositol 1,2,3,4,5,6-hexakisphosphate) is an abundant plant constituent, comprising 1–5% (w/w) of legumes, cereals, pseudocereals, oil seeds, pollen and nuts and represents the largest form of phosphorus storage (88, 89). Besides phytate, myo-inositol 1,2,3,4,5 pentaphosphate and myo-inositol 1,2,3,4 tetraphosphates are also present in seeds, but to a much lower extent (<15%) (90).
Phytic acid is negatively charged at physiological pH, which gives it an extraordinary chelating power with affinity for various components present in foods that are positively charged such as minerals and trace elements. The formed complexes are stable, insoluble and difficult to digest at physiological pH, thus decreasing their bioavailability in the human digestive tract (91).
In certain world populations where staples like wheat, maize and rice are the major or the only source of nutrition, PA as antinutritional factor attracts higher attention because the reduced bioavailability of minerals complexed by it can lead to significant deficiencies in humans (92). Also, Arendt et al. (93) reported that gluten free flours/ingredients have variable concentrations of phytate, i.e., rice, 0.12%; pearl millet, 0.25%; amaranth, 0.47%; teff, 0.70%; lupin, 0.77%; corn, 0.92%; oats,1.13%; quinoa, 1.18%; and soybean, 1.33%. Micronutrient deficiencies affect more than half of the world population, especially in developing countries where plants are the major source of food. Thus, improving the nutritional value of such type of food will improve the nutritional status of entire population (94). High content of phytates in the diet, especially of infants, children, elderly, and people in clinical situations, can significantly decrease the retention of calcium, iron and zinc (95). Reddy et al. (96) reported that PA also is present in the diets of non-ruminant animals, representing 50–80% of total phosphorus content in cereal grains and legumes frequently used in livestock animal feeds. However, phytate phosphorus present in food and feed has low bioavailability and is underutilized due to the lack or low levels of gastrointestinal phytases in monogastric animals (swine, poultry, and fishes) (97, 98). In order to meet the phosphorus requirements in these animals, inorganic phosphorus has to be added to the animal feedstuff as an additional nutrient, which in turn increases the feed cost and phosphorus pollution (99, 100). Undigested phytate and unabsorbed inorganic phosphate are excreted to a large extent (70%) and remains in manure and can lead to its accumulation in the soil and waters. This fact can generate the eutrophication of water, a serious phosphorus pollution problem in areas of intensive livestock production. The eutrophication of water surfaces can then generate cyanobacterial blooms, hypoxia and death of aquatic animals and nitrous oxide production, a potential green- house gas producing a severe environmental problem (101).
PA forms a strong complex with some proteins (the free portion of the basic amino acids such as Lys, Arg, His) and resists their proteolysis. PA negatively affects the absorption of proteins present in foods because inhibits enzymes that are necessary to their digestion such as pepsine. In general, the interaction of phytate with protein is dependent on pH (102).
Lee et al. (103) reported that dietary phytate forms complexes with carbohydrates, reducing their solubility and negatively affecting glucose absorption, leading to a decrease in the glycemic index (blood glucose response). In addition, it was postulated that phytate, by complexing with Ca2+ ion, inhibits amylase activity (104).
Functional Foods and Bioactive Compounds
The demand of consumers for healthier foods has led the food industry to formulate new products within the area of so-called functional foods. Functional foods were defined by Bech-Larsen and Grunert (105) as “Foods that may provide health benefits beyond basic nutrition” and “Food similar in appearance to conventional food that is intended to be consumed as part of a normal diet, but has been modified to subserve physiological roles beyond the provision of simple nutrient requirements.” According to these definitions, certain fruits and vegetables, rich in fiber and bioactive phytochemicals, can be considered functional products. Bioactive compounds are phytochemicals present in plants that can promote health but are not essential for life (106). In the last years, cereals have also been explored due to their potential utilization in developing functional foods (107–109). The key bioactive components of whole grain cereals provide health benefits, principally due to their content of flavonoid and dietary fiber. The covalent interactions between these two components increase their individual anti-inflammatory effects and their positive impact on the gut microbiome (67, 69, 110). In addition to their exceptional nutritional value, pseudocereals are characterized for being rich in many “health-promoting” phytochemicals, such as polyphenols and dietary fiber which exhibit anti-oxidant and free-radical scavenging activity (28, 64, 66, 111–115). Amaranth oil has high levels of tocotrienols and squalene, which are involved in the cholesterol metabolism and could play a significant role in lowering the low-density lipoprotein (LDL)—cholesterol in blood (116). Also, dietary fiber and polyphenols intake has been associated with reduced risk for a number of cardiovascular diseases including stroke, hypertension, and heart disease (117, 118). Pasko et al. (119) reported that the supplementation of a fructose-containing diet with quinoa in male Wistar rats reduced serum total cholesterol, triglycerides, glucose, LDL and plasma total protein and suggests the potential ability of this pseudocereal to prevent cardiovascular disease.
Furthermore, Berti et al. (120) reported that good glycemic control is especially important in CD, as there appears to be a higher incidence of type I diabetes among CD patients. Certain studies in vivo demonstrated that pseudocereals have hypoglycemic effects, for this reason they have been suggested as an alternative to habitual ingredients in the production of cereal-based GF products with low GI (120–123). Hence, the utilization of pseudocereals has increased not only in special diets for people allergic to cereals, but also as part of healthy diets (65).
Fermentation
Fermentation is a metabolic process in which carbohydrates are oxidized to liberate energy in the absence of external electron acceptor. This process is one of the oldest and most economical techniques applied in food preservation and processing (124). Fermented foods, produced and consumed since the development of human civilizations, form part of normal human diet (125). The original purpose of fermentation was the preservation effect. Subsequently, with the development of numerous available preservation technologies, plenty of fermented foods were therefore manufactured because of their unique flavors, aromas, and textures that are much appreciated by consumers. The fermentation of cereals plays a vital role in the production of compounds of great influence on the organoleptic characteristics (such as aroma, taste, and texture) and on the improvement of nutritional properties with a final positive impact on human health (126). Microorganisms are found in almost all ecological niches; cereals and pseudocereals are, in general, a good medium for microbial fermentations. They are rich in polysaccharides, which can be used as a source of carbon and energy by microorganisms during fermentation. Besides carbohydrates, they also contain minerals, vitamins, sterols, and other growth factors (127). Fermented products prepared from more common cereals (such as rice, wheat, corn, or sorghum) and pseudocereals are widespread around the world (128–130). In certain developing countries such as Asia and Africa, high consumption of cereals was reported where these grains are mixed with legumes to improve overall protein quality of the final fermented products (131). Cereal and pseudocereal grains normally have an indigenous microbiota composed by molds, LAB, enterobacteria, aerobic spore formers, etc., which compete for nutrients. The type of microbiota present in each fermented food depends on the pH value, water activity, salt concentration, temperature and composition of the food matrix (132).
Lactic Acid Fermentation
Lactic acid bacteria are Gram positive, non-sporulating, cytochrome deficient, catalase negative, aerotolerant, fastidious, acid-tolerant, and strictly fermentative microorganisms, produce lactic acid as the major metabolic end product of carbohydrate fermentation (133–135). LAB is a heterogeneous group of microorganisms with GRAS (Generally Recognized as Safe) status that have traditionally been associated with food fermentation (136). The effective carbohydrate fermentation coupled to substrate-level phosphorylation is essential characteristic of LAB; the ATP produced is then employed for biosynthetic functions. LAB are generally related with habitats rich in nutrients, for example different foods (milk, beverages, vegetables, meat, cereals); however, some LAB are members of normal flora of the intestine, mouth, and vagina of mammals (137, 138).
Hammes and Ganzle (139) reported that “Sourdough is a leavening agent traditionally obtained through a backslopping procedure, without the addition of starter microorganisms, whose use in bread making has a long history.” Likewise, according to Hammes et al. (140) “The concerted hydrolytic activities of the grain and microorganisms (LAB and yeasts) are the origin of all cereal fermentations and are best represented by the traditional sourdough fermentation.” The application of selected autochthonous LAB to ferment sourdough constitutes an adequate biotechnology to exploit the potential of cereals, non- wheat cereals and pseudocereals in breadmaking (141, 142). This criterion is of great importance when considering the different biochemical, technological, nutritional, and functional characteristics of different flours. The activity of LAB during cereal fermentation is well-documented. A wide variety of metabolites and compounds, such as organic acids, exopolysaccharides (EPS), antimicrobial compounds, and useful enzymes, among others, are produced by LAB (143–151). There are several different ways how the nutritional and functional quality of cereals and pseudocereals could be improved by their fermentation such as: production of bioactive peptides that may stimulate immune system (152); elimination of cereal gluten (153–158); production of gamma-aminobutyric acid (141); increasing total phenolic content and antioxidant capacity (159–162); improving antiproliferative activity (162); decreasing of antinutritional factors, such as phytic acid, tannins and enzyme inhibitors (163–167).
Traditional cereal- and pseudocereal-fermented products are made all over the world, mainly widespread in Asia and Africa (168). Innovative functional fermented foods were formulated using cereal matrices and LAB (169–172). Currently, there are many products derived from cereals fermented by LAB, however only a few are derived from pseudocereals. Fermented quinoa-based beverages were developed by Ludena Urquizo et al. (173) and Jeske et al. (174). Jeske et al. (175), reported the beneficial effect of fermentation by mannitol-producing LAB in combination with various exogenous enzymes in the reduction of sugar in a quinoa-based milk substitute.
Improving the Functional Phytochemical Value by Lactic Fermentation
Numerous commercial microbial enzymes have been used to increase the functional value of phytochemicals present in plant sources however, lactic acid fermentation is preferred to improve the nutraceutical value of these foods because it is relatively inexpensive and improve overall organoleptic and nutritional characteristics (84). Contradictorily, PCs are able to exert an inhibitory effect on LAB (176). In addition, the incidence of certain chemical and physical parameters, such as the lack of fermentable carbohydrates, osmotic stress, and the acidic environment, are adverse conditions for bacterial growth. However, several LAB can adapt and grow in these substrates, being L. plantarum the most isolated species (166, 167, 177–179). The adaptation and survival strategies of LAB during cereal fermentation by activation of specific metabolic pathways have been investigated through a panel of various interacting omics approaches (metabolomic, phenomic, and transcriptomic profile) (180–183). The study of these adaptation responses would allow the optimal design of fermentation strategies for cereals and others plant matrices; however, these “omics” studies were not reported in pseudocereals fermentation.
The effects of LAB on the release of PC and modification of phenolic profiles in both cereals and pseudocereals have been reported. They depend mainly on the grains types, species of microorganisms, fermentation conditions, particularly time, temperature, and pH values (159, 183–185). Some studies have highlighted the capacity of lactic fermentation of pseudocereals to enhance the PC in bread (159, 161, 186), beverages (173, 187, 188), tarhana soup (189) and pasta (190). The PC metabolism in LAB has two important physiological functions, it is an efficient mechanism to detoxify such compounds (191), and can have a role in the cellular energy balance because LAB employ hydroxycinnamic acids as external acceptors of electrons (192). The metabolism of PC by LAB was described principally in L. plantarum strains, and only few studies were reported in Weissella spp., Leuconostoc mesenteroides, L. paracollinoides, L. hilgardii, and Oenococcus oeni (192–194). The enzymes involved in the PC metabolism by LAB such as decarboxylases (PAD), reductases (PAR), esterases and/or glycosidases were reported [Figure 3; (176, 179, 196–198)]. The production of vinyl-phenol, vinyl-guaiacol and vinyl catechol from the p-coumaric, ferulic and caffeic acids, respectively, by PAD activities, are the most relevant (176, 177). Subsequently, these hydroxycinnamates by action of reductase are transformed to their corresponding phenylpropionic acids (199).
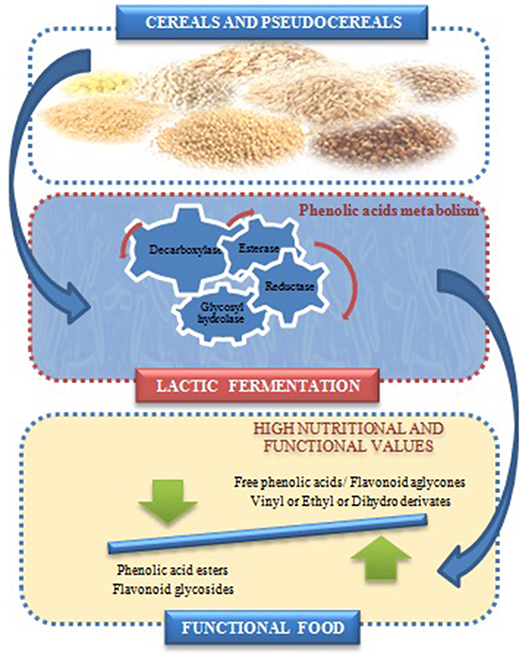
Figure 3. Effect of lactic acid fermentation on phenolic compounds profile of cereal and pseudocereals [adapted from (195)].
The pseudocereals (i.e., buckwheat and quinoa) have a higher content of flavonoids, mainly rutine, kaempferol, and quercetin, with respect to cereals such as rye and wheat (112, 200–204). The flavonoid aglycones are more potent in their functional action (i.e., antioxidant activity) than their corresponding glycosides. Shin et al. (205) showed that a strain of Enterococcus avium was able to metabolize rutine, a flavonol glycoside, in quercetin, a flavonol with many beneficial effects on health. Recently, Zielinski et al. (206) observed a decrease in rutin content in buckwheat flours fermented by different species of Lactobacillus. According to Yang et al. (207), quercetin has numerous biological and pharmacological effects, such as anticancer, antioxidative, antiviral, anti-inflammatory, and antiatherogenic activities. Fan et al. (208) reported the inhibition mechanism of quercetin on tyrosinase (rate-limiting enzyme in the melanogenesis pathway) and its potential use in the treatment of pigmentation disorders. In addition, Xiao (209) reviewed different biological benefits and pharmacokinetic behaviors between flavonoid glycosides and their aglycones. The elucidation of the metabolic pathways of these compounds will lead to obtain strains resistant to PC or adequate enzymes for cereals/pseudocereals processing and products with higher functional values, such as antioxidants.
Vitamins Produced by LAB
LAB and other vitamin-producing microorganisms can be used as an alternative to mandatory fortification in many countries to reduce deficiencies. Some LAB strains can produce elevated concentrations of the natural form of vitamins, which reduces the side-effects of chemically synthesized vitamins (masking of vitamin B12 deficiency, reduced enzyme activities in the liver, etc.) that are normally used (210, 211). Besides being a more natural alternative, vitamins producing LAB can also lower production costs by eliminating the need to add synthetic vitamins. The search for natural LAB strains from different ecological niches that can produce vitamins, such as folate, is essential in order to produce novel fermented foods that have high concentrations of this vitamin (212, 213). Vitamin producing strains have been able to revert and prevent vitamin deficiencies in animal models (52, 214).
LAB diversity is interesting not only at a species level, but also at the strain level, since most technological and nutritional properties are strain dependent. Raw cereals/pseudocereals constitute an interesting ecological niche to isolate new LAB strains with important characteristics to be used as a starter culture in the preparation of fermented cereal food. It was shown that folate producing LAB were isolated from wheat, sorghum and triticale (215). Previously, a few strains have been studied for this capacity in oat brans and rye sourdoughs (216, 217). In terms of vitamin-producing strains in pseudocereals, it has been shown that certain strains of LAB isolated from quinoa and amaranth sourdough have the capacity to produce elevated concentrations of riboflavin and folate in vitamin-free media (166, 167). These strains were used to obtain a B9 and B2 bio enriched pasta, which was able to prevent and revert vitamin deficiency in different rodent models (218).
In Africa, folate deficiency is related to the low dietary diversity and nutrient concentrations in complementary foods for infants (219, 220). In several African countries, cereal-based porridges are consumed as an alternative or in complement to breast feeding (221) but this product does not contain sufficient nutrients to prevent folate deficiencies (222). It has been suggested that porridges can also be consumed after fermentation with LAB, which can improve their overall nutritional quality, especially by increasing vitamin B9 concentrations (131, 223).
Several vitamin B2–producing LAB were isolated from durum wheat flour (224). Two L. plantarum over producer strains used as starter cultures were able to increase between 2 and 3 times the initial concentration of vitamin B2 in both, bread and pasta fermentations.
Russo et al. (225) reported that L. fermentum PBCC11 isolated from sourdough was able to produce riboflavin. Bread produced using the co-inoculum yeast and L. fermentum PBCC11.5 led to an approximately 2-fold increase of final vitamin B2 content compared to the wild-type strain (L. fermentum PBCC11). It was also reported that some LAB strains that are able to produce pseudo-vitamin B12 could also be used to increase the concentrations in cereal-based foods (226). These authors stated that the first pseudo-cobalamin producing strain of LAB was L. reuteri CRL 1098 that was isolated from sourdough (227). It has been suggested that the pseudo-cobalamin produced by LAB would not be biologically available since the intrinsic factor has a low affinity for this compound (228). However, it has been shown that soy fermented with this strain was able to prevent vitamin B12 deficiency in mice (229), demonstrating that pseudo cobalamins are bioavailable. The analysis of sequenced genomes of LAB will provide more insights and new potential candidates that could be used to ferment cereal-based foods.
Strategies to Decrease Phytates in Pseudocereals: Phytases
Different methods have been applied to reduce the PA content in grains and food to improve their nutritional value (230). Phytases, [myo-inositol (1–6) hexakisphosphate phosphohydrolase] constitute a particular subgroup of phosphatases capable of initiating the gradual dephosphorylation of phytate [myo-inositol (1–6) hexakisphosphate] forming myo-inositol phosphate intermediates decreasing or eliminating its antinutritional effect (231, 232). Phytases can be derived from different sources including plants, animals and microorganisms such as yeast and LAB; however, their structures are different (233).
One strategy to reduce phytate in cereals/pseudocereals includes treatments, such as soaking and malting (94) or germination (234, 235), that activate phytases present in plants (236). However, this activity is considered insufficient to eliminate the phytate present in these substrates (237). Recent research has shown that microbial sources are more promising for the production of phytases on a commercial level and in cereal based foods (238, 239).
Nowadays, phytase is one of the most important enzymes for non-ruminant animal production. The application of phytases is broad, can be used to eliminate phytates in the food and feed industries, protect the environment by reducing phosphorus contamination and the eutrophication of water surfaces [Figure 4; (100, 239, 241, 242)]. Phytases have been successfully used in monogastric feeds for about to 20 years. In the beginning, marketable phytases were of fungal origin, mostly from Aspergillus species. Different studies have shown that the bioavailability of phosphorus increases, and the amount of phosphorus excreted is reduced (30–50%) by supplementing animal feeds with phytases (243–245). Recently, Theodoropoulos et al. (246) reported that treatment with commercial phytase decreased the content of myo-inositol phosphates and improved the nutritional value of soy drink, by improving the solubility of Ca2+, Fe2+, and Zn2+. Currently, the significance of bacterial phytases as potential tools in biotechnology is increasing (247).
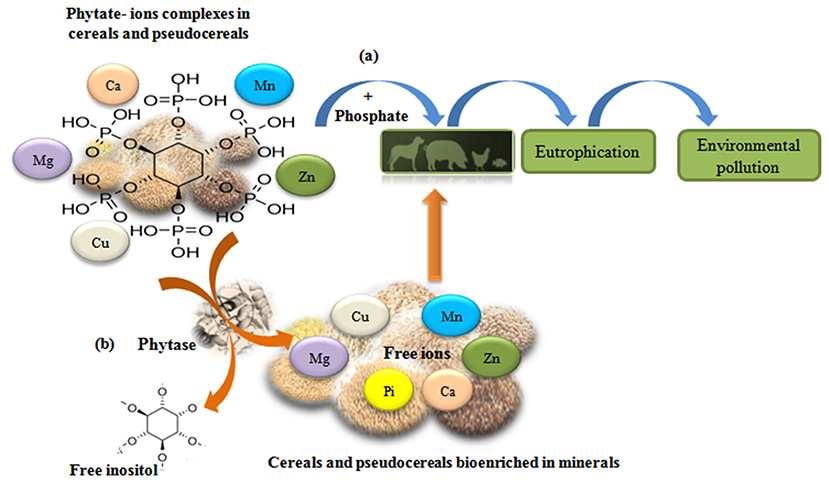
Figure 4. Phytate-ions complexes reduce bioavailability of minerals in cereal and pseudocereals. (a) Phosphorus supplementation in animal feedstuff cause pollution problems. (b) Phytase activity increase mineral biodisponibility and improve nutrition value of foods [adapted from (240)].
LAB phytase activity
The prevalence of LAB in cereal ecosystems and their contribution to the improvement of this particular fermentation processes could be due to their biochemical and metabolic characteristics (248). Studies of enzymes, like phytases involved in nutritional aspects in determined ecosystems, are important for the understanding of particular traits of LAB that are relevant for their right exploitation as starters (249). The PA contained in gluten free flours can be reduced by lactic acid fermentation, directly by LAB phytase activity or indirectly providing the optimal conditions to the endogenous phytase activity (250). Several studies were carried out on different aspects of LAB phytases in cereals fermentation (129, 130, 163, 238, 251–253), nevertheless, there are only few reports of these enzymes in autochthonous LAB isolated from pseudocereals. Carrizo et al. (166, 167) reported high phytase activities in LAB strains isolated from quinoa and amaranth (grains and sourdough), such as E. durans CRL 2122 (1,041 ± 48 U/mL), E. mundtii CRL 2007 (957 ± 25 U/ml) and L. plantarum CRL 2106 (730 ± 25 U/mL), among other. Afterwards, the minerals bioavailability present in pasta made with quinoa flour as a dietary matrix and fermented by selected LAB producing phytase was evaluated in an animal model. The animal group fed with the bio-enriched pasta fermented by LAB (L. plantarum CRL 2107 + CRL 1964) showed higher concentrations of minerals (P, Ca+2, Fe+2, and Mg+2) with respect to control animal group (218). Also, Rizzello et al. (186) reported that the use of quinoa sourdough with autochthonous LAB (L. plantarum T6B10 and L. rossiae T0A16) increased phytase activity during the fermentation respect to non-fermented flour. The results confirmed that quinoa fermented with selected starters had a phytase activity ca. 2.75- times higher than raw quinoa flour.
Recent studies showed that phytate degradation by recombinant probiotic LAB could provide a solution for phosphate utilization in humans (254, 255). Regarding this topic, Vasudevan et al. (247) reviewed the contributions of recombinant technology to phytase research during the last decade with specific emphasis on new generation phytases. These results are relevant in the design of new functional foods with improved nutritional quality by using food-grade strains expressing microbial phytases.
Conclusions
Throughout the world and especially in developing countries, the interest in pseudocereals has increased for both consumers and small businesses. Recent studies strongly suggest that non-essential nutrients like phytochemicals of pseudocereals can also have potential health beneficial effects. This fact has promoted different processing techniques that may enhance the biological value of pseudocereals. Despite the important nutritional and functional value of these grains, their commercialization is still quite limited. Lactic acid fermentation is an ancestral process of food preservation but with renewed interest over time. It has become an important strategy to exploit the bioactive potential of pseudocereals by hydrolysing anti-nutrients factors and increasing the level of health beneficial compounds. The multiple beneficial effects of pseudocereals fermented by selected LAB can be exploited in different ways leading to the design of novel plant-based foods that can target specific populations.
This review summarized recent research reporting some different beneficial effects of pseudocereals and contributes to increase the knowledge on LAB capacity to produce B-group vitamins, metabolize phytochemicals, and decrease phytates present in Andean grains. In this way, lactic acid fermentation can contribute to improve the nutritional and functional potential of fermented foods based on these grains for wide use throughout the world.
Author Contributions
GR: writing—original draft, conceptualization, funding acquisition, and project administration. CG and JL: writing—original draft.
Funding
The authors acknowledge the financial support of the Consejo Nacional de Investigaciones Científicas y Técnicas (PIP 0357-CONICET), Agencia Nacional de Promoción Científica y Tecnológica (PICT 2014 N° 1666-ANPCyT), and Fondo para la Investigación Científica y Tecnológica (FONCyT).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Food and Agriculture Organization, International Fund for Agricultural Development IFAD, UNICEF, World Food Program and World Health Organization (2017). The State of Food Security and Nutrition in the World 2017. Building resilience for peace and food security. Rome: FAO.
2. Jacobsen SJ, Sørensen M, Pedersen SM, Weiner J. Feeding the world: genetically modified crops versus agricultural biodiversity. Agron Sustain Dev. (2013) 33:651–62. doi: 10.1007/s13593-013-0138-9
3. Hasegawa PM. Sodium (Na+) homeostasis and salt tolerance of plants. Environ Exp Bot. (2013) 92:19–31. doi: 10.1016/j.envexpbot.2013.03.001
4. Dwivedi SL, Sahrawat KL, Upadhyaya HD, Ortiz R. Food, nutrition and agrobiodiversity under global climate change. Adv Agron. (2013) 120:1–128. doi: 10.1016/B978-0-12-407686-0.00001-4
5. Deryng D, Conway D, Ramankutty N, Price J, Warren R. Global crop yield response to extreme heat stress under multiple climate change futures. Environ Res Lett. (2014) 9:3. doi: 10.1088/1748-9326/9/3/034011
6. Massawe F, Mayes S, Cheng A. Crop diversity: an unexploited treasure trove for food security. Trends Plant Sci. (2016) 2:365–368. doi: 10.1016/j.tplants.2016.02.006
7. Ghatak A, Chaturvedi P, Weckwerth W. Cereal crop proteomics: systemic analysis of crop drought stress responses towards marker-assisted selection breeding. Front Plant Sci. (2017) 8:757. doi: 10.3389/fpls.2017.00757
8. Arzani A, Ashraf M. Smart engineering of genetic resources for enhanced salinity tolerance in crop plants. Cric Rev Plant Sci. (2016) 35:146–89. doi: 10.1080/07352689.2016.1245056
9. Frison E, Cherfas J, Hodgkin T. Agricultural biodiversity is essential for a sustainable improvement in food and nutrition security. Sustainability. (2011) 3:238–53. doi: 10.3390/su3010238
10. Hermann M. The impact of the European Novel Food Regulation on trade and food innovation based on traditional plant foods from developing coutries. Food Policy. (2009) 34:499–507. doi: 10.1016/j.foodpol.2009.08.005
11. Funk C, Brown ME. Declining Global Per Capita Agricultural Production and Warming Oceans Threaten Food Security. Lincoln, NE: NASA Publications; University of Nebraska (2009). doi: 10.1007/s12571-009-0026-y
12. Henry RJ, Rangan P, Furtado A. Functional cereals for production in new and variable climates. Curr Opin Plant Biol. (2016) 30:11–8. doi: 10.1016/j.pbi.2015.12.008
13. Jacques PJ, Jacques RC. Monocropping cultures into ruin: the loss of food varieties and cultural diversity. Sustain. (2012) 4:2974–97. doi: 10.3390/su4112970
14. Mayes S, Massawe FJ, Alderson PG, Roberts JA, Azam-Ali SN, Hermann M. The potential for underutilized crops to improve security of food production. J Exp Bot. (2012) 63:1075–9. doi: 10.1093/jxb/err396
15. Ruiz KB, Biondi S, Oses R, Acuña-Rodríguez IS, Antognoni F, Martínez-Mosqueira EA, et al. Quinoa biodiversity and sustainability for food security under climate change. A review. Agron Sustain Dev. (2014) 34:349–59. doi: 10.1007/s13593-013-0195-0
16. Hawkesworth S, Dangour AD, Johnston D, Lock K, Poole N, Rushton J, et al. Feeding the world healthily: the challenge of measuring the effects of agriculture on health. Philos Trans R Soc Lond B Biol Sci. (2010) 365:3083–97. doi: 10.1098/rstb.2010.0122
17. Alemayehu FR, Bendevis MA, Jacobsen SE. The potential for utilizing the seed crop amaranth (Amaranthus spp.) in East Africa as an alternative crop to support food security and climate change mitigation. J Agron Crop Sci. (2015) 201:321–9. doi: 10.1111/jac.12108
18. Gordon E. Analysing the versatility and complexity of cereal grains. Food Sci Newslettter. (2006) 88:1–5.
19. Alvarez-Jubete L, Arendt E, Gallagher E. Nutritive value of pseudocereals and their increasing use as functional gluten-free ingredients. Trends Food Sci Technol. (2010) 21:106–13. doi: 10.1016/j.tifs.2009.10.014
20. Das S. Amaranthus: A Promising Crop of Future. Singapore: Springer Nature (2016). doi: 10.1007/978-981-10-1469-7
21. Shewry PR. The major seed storage proteins of spelt wheat, sorghum, millets and pseudocereals. In: Belton PS, Taylor JRN, editors. Pseudocereals and Less Common Cereals, Grain Properties and Utilization Potential. Berlin; Heidelberg: Springer-Verlag (2002). p. 1–24. doi: 10.1007/978-3-662-09544-7_1
22. Rodrigo L. Celiac disease. World J Gastroenterol. (2006) 12:6585–93. doi: 10.3748/wjg.v12.i41.6585
23. Poutanen K, Sozer N, Della Valle G. How can technology help to deliver more of grain in cereal food for a healthy diet? J Cereal Sci. (2014) 59:327–36. doi: 10.1016/j.jcs.2014.01.009
24. Peñas E, Uberti F, di Lorenzo C, Ballabio C, Brandolini A, Restani P. Biochemical and immunochemical evidences supporting the inclusion of quinoa (Chenopodium quinoa Willd.) as a gluten-free ingredient. Plant Foods Human Nutr. (2014) 69:297–303. doi: 10.1007/s11130-014-0449-2
25. Schoenlechner R. Chapter 9: pseudocereals in gluten-free products. In: Haros CM, Schoenlechner R, editors. Pseudocereals: Chemistry and Technology. 1st, ed. Oxford: John Wiley & Sons, Ltd. (2017). p. 193–216. doi: 10.1002/9781118938256.ch9
26. Jacobsen SE. The worldwide potential for quinoa (Chenopodium quinoa Willd.). In: Bazile D, Bertero D, Nieto C, editors. Estado del Arte de la Quínoa en el Mundo en 2013. Montpellier: FAO, Santiago de Chile and CIRAD (2013). p. 13.
27. Jacobsen SE, Mujica A, Jensen CR. The resistance of quinoa (Chenopodium quinoa Willd.) to adverse abiotic factors. Food Rev Int. (2003) 19:99–109. doi: 10.1081/FRI-120018872
28. Repo-Carrasco-Valencia R, Hellström JK, Pihlava JM, Mattila PH. Flavonoids and other phenolic compounds in Andean indigenous grains: Quinoa (Chenopodium quinoa), kañiwa (Chenopodium pallidicaule) and kiwicha (Amaranthus caudatus). Food Chem. (2010) 120:128–33. doi: 10.1016/j.foodchem.2009.09.087
29. Ruiz Carrasco K, Antognoni F, Coulibaly AK, Lizardi S, Covarrubias A, Martínez E, et al. Variation in salinity tolerance of four lowand genotypes of quinoa (Chenopodium quinoa, Willd.) as assessed by growth, physiological traits, and sodium transporter gene expression. Plant Physiol. Biochem. (2011) 49:1333–41. doi: 10.1016/j.plaphy.2011.08.005
30. Miranda M, Vega-Gálvez A, Quispe-Fuentes I, Rodríguez MJ, Maureira H, Martínez EA. Nutritional aspects of six quinoa (Chenopodium quinoa Willd.). Ecotypes from three geographical areas of Chile. Chil J Agricul Res. (2012) 72:175–81. doi: 10.4067/S0718-58392012000200002
31. Repo-Carrasco-Valencia R, Peña J, Kallio H, Salminen S. Dietary fiber and other functional components in two varieties of crude and extruded kiwicha (Amaranthus caudatus). J Cereal Sci. (2009) 49:219–24. doi: 10.1016/j.jcs.2008.10.003
32. Sauer JD. Grain amaranths Amaranthus spp. (Amaranthaceae). In: Simmonds NW, editor. Evolution of Crop Plants. London: Longman (1976). p. 4–7.
33. Hunziker AT, Planchuelo AM. Sobre un nuevo hallazgo de Amaranthus caudatus en tumbas indígenas de Argentina. Kurtziana. (1971) 6:63–7.
34. Mazza G, Oomah BD. Buckwheat. In: Caballero B, editor. Encyclopedia of Food Sciences and Nutrition. Oxford: Academic Press (2003). p. 692–9. doi: 10.1016/B0-12-227055-X/00132-2
35. Cheng A. Review: shaping a sustainable food future by rediscovering long-forgotten ancient grains. Plant Sci. (2018) 269:136–42. doi: 10.1016/j.plantsci.2018.01.018
36. Charalampopoulos D, Wang R, Pandiella SS, Webb C. Application of cereals and cereal components in functional foods: a review. Int J Food Microbiol. (2002) 79:131–41. doi: 10.1016/S0168-1605(02)00187-3
37. De Anton Migliorati M, Bell M, Grace PR, Scheer C, Rowlings DW, Liu S. Legume pastures can reduce N2O emissions intensity in subtropical cereal cropping systems. Agric Ecosyst Environ. (2015) 204:27–39. doi: 10.1016/j.agee.2015.02.007
38. Oyewole OB. Lactic fermented foods in Africa and their benefits. Food Control. (1997) 8:289–97. doi: 10.1016/S0956-7135(97)00075-3
39. Schlick G, Bubenheim DL. Quinoa. In: Janick J, editor. Candidate Crop for NASA's Controlled Ecological Life Support Systems. Arlington, VA: ASITS Press (1996). p. 632–40.
40. Konishi Y. Nutritional characteristics of pseudocereal amaranth and quinoa: alternative foodstuff for patients with food al- lergy. J Jpn Soc Nutr Food Sci. (2002) 55:299–302. doi: 10.4327/jsnfs.55.299
41. Food and Agriculture Organization Regional Office for Latin America, and the Caribbean PROINPA (2011). Quinoa: An Ancient Crop to Contribute to World Food Security. Santiago: FAO Regional Office for Latin American and the Caribbean. Retrieved from: http://www.fao.org/alc/file/media/pubs/2011/cultivo_quinua_en.pdf
42. Koziol MJ. Chemical composition and nutritional value of quinoa (Chenopodium quinoa Willd.). J Food Compost Anal. (1992) 5:35–68. doi: 10.1016/0889-1575(92)90006-6
43. Repo-Carrasco R, Espinoza C, Jacobsen SE. Nutritional value and use of the Andean crops quinoa (Chenopodium quinoa) and kañiwa (Chenopodium pallidicaule). Food Rev Int. (2003) 19:179–89. doi: 10.1081/FRI-120018884
44. Abugoch James LE. Chapter 1: Quinoa (Chenopodium quinoa Willd.): composition, chemistry, nutritional, and functional properties. In: Taylor S, editor. Advances in Food and Nutrition Research. Vol. 58. Cambridge, MA: Academic Press (2009). p. 58, 1–31. doi: 10.1016/S1043-4526(09)58001-1
45. Alvarez-Jubete L, Arendt EK, Gallagher E. Nutritive value and chemical composition of pseudocereals as gluten-free ingredients. Int J Food Sci Nutr. (2009) 60:240–57. doi: 10.1080/09637480902950597
46. Vega-Gálvez A, Miranda M, Vergara J, Uribe E, Puente L, Martínez EA. Nutrition facts and functional potential of quinoa (Chenopodium quinoa Willd.), an ancient Andean grain: a review. J Sci Food Agricul. (2010) 90:2541–7. doi: 10.1002/jsfa.4158
47. Sanz-Penella JM, Wronkowska M, Soral-Smietana M, Haros M. Effect of whole amaranth flour on bread properties and nutritive value. LWT Food Sci Technol. (2013) 50:679–85. doi: 10.1016/j.lwt.2012.07.031
48. Caselato-Sousa VM, Amaya-Farfán J. State of knowledge on amaranth grain: a comprehensive review. J Food Sci. (2012) 77:93–104. doi: 10.1111/j.1750-3841.2012.02645.x
49. Venskutonis PR, Kraujalis P. Nutritional components of amaranth seeds and vegetables: a review on composition, properties, and uses. Compr Rev Food Sci Food Saf. (2013) 12:381–412. doi: 10.1111/1541-4337.12021
50. Burlingame B, Charrondiere UR, Dernini S, Stadlmayr B, Mondovì S. Food biodiversity and sustainable diets: implications of applications for food production and processing. In: Boye JI, Arcand Y, editors. Green Technologies in Food Production and Processing. Boston, MA: Springer, U.S. (2012). p. 643–57. doi: 10.1007/978-1-4614-1587-9_24
51. Food and Agriculture Organization. Home-International Year of Quinoa 2013. (2013). Available online at: http://www.fao.org/quinoa-2013/en/ (accessed February 21, 2014).
52. Laiño JE, Zelaya H, Juarez del Valle M, Savoy de Giori G, LeBlanc JG. Milk fermented with selected strains of lactic acid bacteria is able to improve folate status of deficient rodents and also prevent folate deficiency. J Funct Foods. (2015) 17:22–32. doi: 10.1016/j.jff.2015.04.055
53. Schoenlechner R, Wendner M, Siebenhandl-Ehn S, Berghofer E. Pseudocereals as alternative sources for high folate content in staple foods. J Cereal Sci. (2010) 52:475–9. doi: 10.1016/j.jcs.2010.08.001
54. McKevith B. Nutritional aspects of cereals. Brit Nutrit Found Nutr Bull. (2004) 29:111–42. doi: 10.1111/j.1467-3010.2004.00418.x
55. Dini C, García MA, Viña SZ. Non-traditional flours: frontiers between ancestral heritage and innovation. Food Funct. (2012) 3:606–20. doi: 10.1039/c2fo30036b
56. Burgess CM, Smid EJ, van Sinderen D. Bacterial vitamin B2, B11 and B12 overproduction: an overview. Int J Food Microbiol. (2009) 133:1–7. doi: 10.1016/j.ijfoodmicro.2009.04.012
57. Aguilar-Garcia C, Gavino G, Baragano-Mosqueda M, Hevia P, Gavino VC. Correlation of tocopherol, tocotrienol, g-oryzanol and total polyphenol content in rice bran with different antioxidant capacity assays. Food Chem. (2007) 102:1228–32. doi: 10.1016/j.foodchem.2006.07.012
58. Tang Y, Tsao R. Phytochemicals in quinoa and amaranth grains and their antioxidant, anti-inflammatory, and potential health beneficial effects: a review. Mol Nutr Food Res. (2017) 61:1600767. doi: 10.1002/mnfr.201600767
59. Carcea M, Narducci V, Turfani V, Giannini V. Polyphenols in raw and cooked cereals/pseudocereals/legume pasta and couscous. Foods. (2017) 6:E80. doi: 10.3390/foods6090080
60. Pang Y, Ahmed S, Xu Y, Beta T, Zhu Z, Shao Y, et al. Bound phenolic compounds and antioxidant properties of whole grain and bran of white, red and black rice. Food Chem. (2018) 240:212–21. doi: 10.1016/j.foodchem.2017.07.095
61. Bravo LP. Polyphenols: chemistry, dietary sources, metabolism, and nutritional significance. Nutr Rev. (1998) 56:317–33. doi: 10.1111/j.1753-4887.1998.tb01670.x
62. Peñarrieta JM, Alvarado JA, Akesson B, Bergenståhl B. Total antioxidant capacity and content of flavonoids and other phenolic compounds in canihua (Chenopodium pallidicaule): an Andean pseudocereal. Mol Nutr Food Res. (2008) 52:708–17. doi: 10.1002/mnfr.200700189
63. Zhang H, Shao Y, Bao J, Beta T. Phenolic compounds and antioxidant properties of breeding lines between the white and black rice. Food Chem. (2015) 172:630–9. doi: 10.1016/j.foodchem.2014.09.118
64. Gorinstein S, Medina Vargas O, Jaramillo N, Arnao Salas I, Martinez Ayala A, Arancibia-Avila P, et al. The total polyphenols and the antioxidant potentials of some selected cereals and pseudocereals. Eur Food Res Technol. (2007) 225:321–8. doi: 10.1007/s00217-006-0417-7
65. Gorinstein S, Lojek A, CíŽ M, Pawelzik E, Delgado-Licon E, Medina OJ, et al. Comparison of composition and antioxidant capacity of some cereals and pseudocereals. Int J Food Sci Technol. (2008) 43:629–37. doi: 10.1111/j.1365-2621.2007.01498.x
66. Drzewiecki J, Martinez-Ayala AL, Lozano-Grande MA, Leontowicz H, Leontowicz M, Jastrzebski Z, et al. In vitro screening of bioactive compounds in some gluten-free plants. Appl Biochem Biotechnol. (2018) 186:847–60. doi: 10.1007/s12010-018-2772-9
67. Manach C, Williamson G, Morand C, Scalbert A, Remesy C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am J Clin Nutr. (2005) 81:230S−42. doi: 10.1093/ajcn/81.1.230S
68. Hung PV, Hatcher DW, Barker W. Phenolic acid composition of sprouted wheats by ultra-performance liquid chromatography (UPLC) and their antioxidant activities. Food Chem. (2011) 126:1896–901. doi: 10.1016/j.foodchem.2010.12.015
69. Awika JM, Rose DJ, Simsek S. Complementary effects of cereal and pulse polyphenols and dietary fiber on chronic inflammation and gut health. Food Funct. (2018) 9:1389–409. doi: 10.1039/C7FO02011B
70. Lu Y, Shan S, Li H, Shi J, Zhang X, Li Z. Reversal effects of bound polyphenol from foxtail millet bran on multidrug resistance in human hct-8/fu colorectal cancer cell. J Agric Food Chem. (2018) 66:5190–9. doi: 10.1021/acs.jafc.8b01659
71. Zambrana S, Lundqvist LCE, Veliz V, Catrina SB, Gonzales E, Östenson CG. Amaranthus caudatus stimulates insulin secretion in goto-kakizaki rats, a model of diabetes mellitus type 2. Nutrients. (2018) 10:E94. doi: 10.3390/nu10010094
72. Pham T, Teoh KT, Savary BJ, Chen MH, McClung A, Lee SO. In vitro fermentation patterns of rice bran components by human gut microbiota. Nutrients. (2017) 9:E1237. doi: 10.3390/nu9111237
73. Manach C, Scalbert A, Morand C, Rémésy C, Jiménez L. Polyphenols: food sources and bioavailability. Am J Clin Nutr. (2004) 79:727–47. doi: 10.1093/ajcn/79.5.727
74. Ozdal T, Capanoglu E, Altay F. A review on protein–phenolic interactions and associated changes. Food Res Int. (2013) 51:954–70. doi: 10.1016/j.foodres.2013.02.009
75. Ktenioudaki A, Alvarez-Jubete L, Gallagher E. A Review of the process-induced changes in the phytochemical content of cereal grains: the breadmaking process. Crit Rev Food Sci Nutr. (2014) 55:611–9. doi: 10.1080/10408398.2012.667848
76. Jakobek L. Interactions of polyphenols with carbohydrates, lipids and proteins. Food Chem. (2015) 175:556–67. doi: 10.1016/j.foodchem.2014.12.013
77. Gutierrez-Grijalva EP, Ambriz-Pere DL, Leyva-Lopez N, Castillo-Lopez RI, Heiedia JB. Review: dietary phenolic compounds, health benefits and bioaccessibility. Arch Latinoam Nutr. (2016) 66:87–100.
78. Ribas-Agustí A, Martín-Belloso O, Soliva-Fortuny R, Elez-Martínez P. Food processing strategies to enhance phenolic compounds bioaccessibility and bioavailabilityin plant-based foods. Crit Rev Food Sci Nutr. (2017) 58:2531–254. doi: 10.1080/10408398.2017.1331200
79. Aborus NE, Šaponjac VT, Canadanović-Brunet J, Cetković G, Hidalgo A, Vulić J, et al. Sprouted and freeze-dried wheat and oat seeds - phytochemical profile and in vitro biological activities. Chem Biodivers. (2018) 15:e1800119. doi: 10.1002/cbdv.201800119
80. Hur SJ, Lee SY, Kim YC, Choi I, Kim GB. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. (2014) 160:346–56. doi: 10.1016/j.foodchem.2014.03.112
81. Cardona F, Andrés-Lacueva C, Tulipani S, Tinahones FJ, Queipo-Ortuño MI. Benefits of polyphenols on gut microbiota and implications in human health. J Nutr Biochem. (2013) 24:1415–22. doi: 10.1016/j.jnutbio.2013.05.001
82. Anson NM, Aura AM, Selinheimo E, Mattila I, Poutanen K, van den Berg R, et al. Bioprocessing of wheat bran in whole wheat bread increases the bioavailability of phenolic acids in men and exerts antiinflammatory effects ex vivo. J Nutr. (2011) 141:137–43. doi: 10.3945/jn.110.127720
83. Wang T, He F, Chen G. Improving bioaccessibility and bioavailability of phenolic compounds in cereal grains through processing technologies: a concise review. J Funct Food. (2014) 7:101–11. doi: 10.1016/j.jff.2014.01.033
84. Acosta-Estrada BA, Gutiérrez-Uribe JA, Serna-Saldívar SO. Bound phenolics in foods, a review. Food Chem. (2014) 152:46–55. doi: 10.1016/j.foodchem.2013.11.093
85. Katina K, Arendt E, Liukkonen KH, Autio K, Flander L, Poutanen K. Potential of sourdough for healthier cereal products. Trends Food Sci Technol. (2005) 16:104–12. doi: 10.1016/j.tifs.2004.03.008
86. Ruales J, Nair BM. Saponins, phytic acid, tannins and protease inhibitors in quinoa (Chenopodium quinoa, Willd) seeds. Food Chem. (1993) 48:137–43. doi: 10.1016/0308-8146(93)90048-K
87. Kumar V, Sinha AK, Makkar HP, Becker K. Dietary roles of phytate and phytase in human nutrition: a review. Food Chem. (2010) 120:945–59. doi: 10.1016/j.foodchem.2009.11.052
88. Gibson DM, Ullah ABJ. Phytase and their actions on phytic acid. In: Morre DJ, Bross WF, Loewus FA, editors. Inositol Metabolism in Plants. New York, NY: Wiley-Liss (1990). p. 77–92.
89. Loewus F. Biosynthesis of phytate in food grains and seeds. In: Reddy NR, Sathe SK, editors. Food Phytates. Boca Raton, FL: CRC Press (2002). p. 53–61. doi: 10.1201/9781420014419.ch4
90. Dorsch JA, Cook A, Young KA, Anderson JM, Bauman AT, Volkmann CJ, et al. Seed phosphorus and inositol phosphate phenotype of barley low phytic acid genotypes. Phytochemistry. (2003) 62:691–706. doi: 10.1016/S0031-9422(02)00610-6
91. Konietzny U, Greiner R. Phytic acid: nutritional impact. In: Caballero B, Trugo L, Finglas P, editors. Encyclopedia of Food Science and Nutrition. London: Elsevier (2003). p. 4555–63. doi: 10.1016/B0-12-227055-X/00923-8
92. Schlemmer U, Frolich W, Prieto RM, Grases F. Phytate in foods and significance for humans: food sources, intake, processing, bioavailability, protective role and analysis. Mol Nutr Food Res. (2009) 53:S330–75. doi: 10.1002/mnfr.200900099
93. Arendt EK, Moroni A, Zannini E. Medical nutrition therapy: use of sourdough lactic acid bacteria as a cell factory for delivering functional biomolecules and food ingredients in gluten free bread. Microb Cell Fact. (2011) 10:1–9. doi: 10.1186/1475-2859-10-S1-S15
94. Gupta RK, Gangoliya SS, Singh NK. Reduction of phytic acid and enhancement of bioavailable micronutrients in food grains. J Food Sci Technol. (2015) 52:676–84. doi: 10.1007/s13197-013-0978-y
95. Mendoza C. Effect of genetically modified low phytic acid plants on mineral absorption. J Food Sci Technol. (2002) 37:759–67. doi: 10.1046/j.1365-2621.2002.00624.x
96. Reddy NR, Sathe SK, Salunkhe DK. Phytates in legumes and cereals. Adv Food Res. (1982) 28:1–92. doi: 10.1016/S0065-2628(08)60110-X
97. Jongbloed AW, Mroz Z, Kemme PA. The effect of supplementary Aspergillus niger phytase in diets for pigs on concentration and apparent digestibility of dry-matter, total phosphorus, and phytic acid in different sections of the alimentary tract. J Anim Sci. (1992) 70:1159–68. doi: 10.2527/1992.7041159x
98. Selle PH, Ravindran V, Bryden WL, Scott T. Influence of dietary phytate and exogenous phytase on amino acid digestibility in poultry: a review. J Poult Sci. (2006) 43:89–103. doi: 10.2141/jpsa.43.89
99. Jorquera M, Martínez O, Maruyama F, Marschner P, de la Luz Mora M. Current and future biotechnological applications of bacterial phytases and phytase-producing bacteria. Microbes Environ. (2008) 23:182–91. doi: 10.1264/jsme2.23.182
100. Singh B, Satyanarayana T. Fungal phytases: characteristics and amelioration of nutritional quality and growth of non-ruminants. J Anim Physiol Anim Nutr. (2015) 99:646–60. doi: 10.1111/jpn.12236
101. Vats P, Bhattacharyya MS, Banerjee UC. Use of phytases (myo-inositolhexakisphosphate phosphohy- drolases) for combatting environmental pollution: a biological approach. Crit Rev Environ Sci Technol. (2005) 35:469–86. doi: 10.1080/10643380590966190
102. Cheryan M. Phytic acid interactions in food systems. Crit Rev Food Sci Nutr. (1980) 13:297. doi: 10.1080/10408398009527293
103. Lee S-H, Park H-J, Chun H-K, Cho S-Y, Cho S-M, Lillehoj HS. Dietary phytic acid lowers the blood glucose level in diabetic KK mice. Nutr Res. (2006) 26:474–9. doi: 10.1016/j.nutres.2006.06.017
104. Selle PH, Ravindran V, Caldwell RA, Bryden WL. Phytate and phytase: Consequences for protein utilisation. Nutr Res Rev. (2000) 13:255–78. doi: 10.1079/095442200108729098
105. Bech-Larsen T, Grunert KG. The perceived healthiness of functional foods- A conjoint study of Danish, Finnish and American consumers' perception of functional foods. Appetite. (2003) 40:9–14. doi: 10.1016/S0195-6663(02)00171-X
106. Arendt E, Zannini E. Cereal Grains for the Food and Beverage Industries. Cambridge: Woodhead (2013). p. 369–466. doi: 10.1533/9780857098924.369
107. Taylor JRN, Belton PS, Beta T, Duodu KG. Increasing the utilisation of sorghum, millets and pseudocereals: developments in the science of their phenolic phytochemicals, biofortification and protein functionality. J Cereal Sci. (2014) 59:257–75. doi: 10.1016/j.jcs.2013.10.009
108. Suriano S, Iannucci A, Codianni P, Fares C, Russo M, Pecchioni N, et al. Phenolic acids profile, nutritional and phytochemical compounds, antioxidant properties in colored barley grown in southern Italy. Food Res Int. (2018) 113:221–33. doi: 10.1016/j.foodres.2018.06.072
109. Filipčev B, Kojić J, Krulj J, BodroŽa-Solarov M, Ilić N. Betaine in cereal grains and grain-based products. Foods. (2018) 7:49. doi: 10.3390/foods7040049
110. Quiròs-Sauceda AE, Palafox-Carlos H, Sáyago-Ayerdi SG, Ayala-Zavala JF, Bello-Perez LA, Álvarez-Parrilla E, et al. Dietary fiber and phenolic compounds as functional ingredients: interaction and possible effect after ingestion. Food Funct. (2014) 5:1063–72. doi: 10.1039/C4FO00073K
111. Berti C, Riso P, Brusamolino A, Porrini M. Effect on appetite control of minor cereal and pseudocereal products. Br J Nutr. (2005) 94:850–8. doi: 10.1079/BJN20051563
112. Alvarez-Jubete L, Wijngaard H, Arendt EK, Gallagher E. Polyphenol composition and in vitro antioxidant activity of amaranth, quinoa buckwheat and wheat as affected by sprouting and baking. Food Chem. (2010) 119:770–8. doi: 10.1016/j.foodchem.2009.07.032
113. Inglett GE, Chen D, Liu SX. Antioxidant activities of selective gluten free ancient grains. Food Nutr Sci. (2015) 6:612–21. doi: 10.4236/fns.2015.67065
114. Carciochi RA, Manrique GD, Dimitrov K. Optimization of antioxidant phenolic compounds extraction from quinoa (Chenopodiumquinoa) sedes. J Food Sci Technol. (2015) 52:4396–404. doi: 10.1007/s13197-014-1514-4
115. Rocchetti G, Chiodelli G, Giuberti G, Masoero F, Trevisan M, Lucini L. Evaluation of phenolic profile and antioxidant capacity in gluten-free flours. Food Chem. (2017) 228:367–73. doi: 10.1016/j.foodchem.2017.01.142
116. Bodroza-Solarov M, Filiocev B, Kevresan Z, Mandic A, Simurina O. Quality of bread supplemented with popped Amaranthus cruentus grain. J Food Process Eng. (2008) 31:602–18. doi: 10.1111/j.1745-4530.2007.00177.x
117. Scalbert A, Manach C, Morand C, Remesy C, Jimenez L. Dietary polyphenols and the prevention of diseases. Crit Rev Food Sci Nutr. (2005) 45:287–306. doi: 10.1080/1040869059096
118. Kurth T, Moore SC, Gaziano JM, Kase CS, Stampfer MJ, Berger K, et al. Healthy lifestyle and the risk of stroke in women. Arch Intern Med. (2006) 166:1403–9. doi: 10.1001/archinte.166.13.1403
119. Pasko P, Zagrodzki P, Barton H, Chlopika J, Gorinstein S. Effect of quinoa seeds (Chenopodium quinoa) in diet on some biochemical parameters and essential elements in blood of high fructose-fed rats. Plant Foods Hum Nutr. (2010) 65:333–8. doi: 10.1007/s11130-010-0197-x
120. Berti C, Riso P, Brusamolino A, Porrini M. In vitro starch digestibility and in vivo glucose response of gluten-free foods and their gluten counterparts. Eur J Nutr. (2004) 43:198–204. doi: 10.1007/s00394-004-0459-1
121. Skrabanja V, Liljeberg Elmstahl HGM, Kreft I, Bjorck IME. Nutritional properties of starch in buckwheat products: studies in vitro and in vivo. J Agricul Food Chem. (2001) 49:490–6. doi: 10.1021/jf000779w
122. Kim HK, Kim MJ, Cho HY, Kim EK, Shin DH. Antioxidative and anti-diabetic effects of Amaranth (Amaranthus esculantus) in streptozotocin-induced diabetic rats. Cell Biochem Funct. (2006) 24:195–9. doi: 10.1002/cbf.1210
123. Wolter A, Hager AS, Zannini E, Arendt EK. In vitro starch digestibility and predicted glycaemic indexes of buckwheat, oat, quinoa, sorghum, teff and commercial gluten-free bread. J Cereal Sci. (2013) 58:431–36. doi: 10.1016/j.jcs.2013.09.003
124. Stiles ME. Biopreservation by lactic acid bacteria. Antonie Van Leeuwenhoek. (1996) 70:331–45. doi: 10.1007/BF00395940
125. Marco ML, Heeney D, Binda S, Cifelli CJ, Cotter PD, Foligné B, et al. Health benefits of fermented foods: microbiota and beyond. Curr Opin Biotechnol. (2017) 44:94–102. doi: 10.1016/j.copbio.2016.11.010
126. Bourdichon F, Casaregola S, Farrokh CH, Frisvad JC, Gerds ML, Hammes WP, et al. Food fermentations: microorganisms with technological beneficial use. Int J Food Microbiol. (2012) 154:87–97. doi: 10.1016/j.ijfoodmicro.2011.12.030
127. Gobbetti M, Minervini F, Pontonio E, Di Cagno R, De Angelis M. Drivers for the establishment and composition of the sourdough lactic acid bacteria biota. Int J Food Microbiol. (2016) 239:3–18. doi: 10.1016/j.ijfoodmicro.2016.05.022
128. Kocková M, Mendel J, Medvedová A, Šturdík E, Valík L. Cereals and pseudocereals as substrates for growth and metabolism of a probiotic strain Lactobacillus rhamnosus GG. J Food Nutr Res. (2013) 52:25–36.
129. Fischer MM, Egli IM, Aeberli I, Hurrell RF, Meile L. Phytic acid degrading lactic acid bacteria in tef-injera fermentation. Int J Food Microbiol. (2014) 190:54–60. doi: 10.1016/j.ijfoodmicro.2014.08.018
130. Mamhoud A, Nionelli L, Bouzaine T, Hamdi M, Gobbetti M, Rizzello CG. Selection of lactic acid bacteria isolated from Tunisian cereals and exploitation of the use as starters for sourdough fermentation. Int J Food Microbiol. (2016) 225:9–19. doi: 10.1016/j.ijfoodmicro.2016.03.004
131. Blandino A, Al-Aseeri ME, Pandiella SS, Cantero D, Webb C. Cereal based fermented foods and beverages. Food Res Int. (2003) 36:527–43. doi: 10.1016/S0963-9969(03)00009-7
132. Salovaara H. Lactic acid bacteria in cereal-based products, lactic acid bacteria. In: Salminen S, von Wright A, Ouwehand A, editors. Microbiological and Functional Aspects. 3rd ed. New York, NY: Marcel Dekker, Inc. (2004). p. 431–52. doi: 10.1201/9780824752033.ch15
133. Kandler O. Carbohydrate metabolism in lactic acid bacteria. Antonie Van Leeuwenhoek. (1983) 49:209–24. doi: 10.1007/BF00399499
134. Leroy F, De Vuyst L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci Technol. (2004) 15:67–78. doi: 10.1016/j.tifs.2003.09.004
135. Hugenholtz J. The lactic acid bacterium as a cell factory for food ingredient production. Int Dairy J. (2008) 18:466–75. doi: 10.1016/j.idairyj.2007.11.015
136. Carr FJ, Chill D, Maida N. The lactic acid bacteria: a literature survey. Crit Rev Microbiol. (2002) 28:281–370. doi: 10.1080/1040-840291046759
137. Axelsson L. Lactic acid bacteria: classification and physiology in lactic acid bacteria. In: Salminen S, Ouwehand A, editors. Microbiological and Functional Aspects, 3rd ed. New York, NY: Marcel Dekker Inc. (2004). p. 1–66. doi: 10.1201/9780824752033.ch1
138. Holzapfel WH, Wood BJB editors. Introduction to the lactic acid bacteria. In: Lactic Acid Bacteria: Biodiversity and Taxonomy. West Sussex: John Wiley & Sons, Ltd. (2014). p. 1–12. doi: 10.1002/9781118655252
139. Hammes WP, Ganzle MG. Sourdough breads and related products. In: Woods BJB, editor. Microbiology of Fermented Foods. Vol. 1. London: Blackie Academic Professional (1998). p. 199–216. doi: 10.1007/978-1-4613-0309-1_8
140. Hammes WP, Brandt MJ, Francis KL, Rosenheim J, Seitter MFH, Vogelmann SA. Microbial ecology of cereal fermentation. Trends Food Sci Technol. (2005) 16:4–11. doi: 10.1016/j.tifs.2004.02.010
141. Coda R, Rizzello CG, Gobbetti M. Use of sourdough fermentation and pseudo-cereals and leguminous flours for the making of a functional bread enriched of ∂-aminobutyric acid (GABA). Int J Food Microbiol. (2010) 137:236–45. doi: 10.1016/j.ijfoodmicro.2009.12.010
142. Coda R, Nionelli L, Rizzello CG, De Angelis M, Tossut P, Gobbetti M. Spelt and emmer flours: characterization of the lactic acid bacteria microbiota and selection of mixed autochthonous starters for bread making. J Appl Microbiol. (2010) 108:925–35. doi: 10.1111/j.1365-2672.2009.04497.x
143. Rollán G, Font de Valdez G. The peptide hydrolase system of Lactobacillus reuteri. Int J Food Microbiol. (2001) 70:303–7. doi: 10.1016/S0168-1605(01)00544-X
144. Katina K. Sourdough: A Tool for the Improved Flavour, Texture and Shelf-Life of Wheat Bread. Espoo: VTT Publications (2005). p. 569.
145. Gänzle MG, Loponen J, Gobbetti M. Proteolysis in sourdough fermentations: mechanisms and potential for improved bread quality. Trends Food Sci Technol. (2008) 19:513–21. doi: 10.1016/j.tifs.2008.04.002
146. Gerez CL, Cuezzo S, Rollán G, Font de Valdez G. Lactobacillus reuteri CRL 1100 as starter culture for wheat dough fermentation. Food Microbiol. (2008) 25:253–59. doi: 10.1016/j.fm.2007.10.011
147. Gerez CL, Torino MI, Rollán G, Font de Valdez G. Prevention of bread mould spoilage by using lactic acid bacteria with antifungal properties. Food Control. (2009) 20:144–8. doi: 10.1016/j.foodcont.2008.03.005
148. Gerez CL, Torres MJ, Font de Valdez G, Rollán G. Control of spoilage fungi by lactic acid bacteria. Biol Control. (2013) 64:231–7. doi: 10.1016/j.biocontrol.2012.10.009
149. Gobbetti M, Rizzello CG, Di Cagno R, De Angelis M. How the sourdough may affect the functional features of leavened baked goods. Food Microbiol. (2014) 37:30–40. doi: 10.1016/j.fm.2013.04.012
150. Celano G, De Angelis M, Minervini F, Gobbetti M. Different flour microbial communities drive to sourdoughs characterized by diverse bacterial strains and free amino acid profiles. Front Microbiol. (2016) 7:1770. doi: 10.3389/fmicb.2016.01770
151. Zhao CJ, Schieber A, Gänzle MG. Formation of taste-active amino acids, amino acid derivatives and peptides in food fermentations - a review. Food Res Int. (2016) 89:39–47. doi: 10.1016/j.foodres.2016.08.042
152. Wouters JTM, Ayad EHE, Hugenholtz J, Smit G. Microbes from raw milk for fermented dairy products. Int Dairy J. (2002) 12:91–109. doi: 10.1016/S0958-6946(01)00151-0
153. Rollán G, De Angelis M, Gobbetti M, Font de Valdez G. Proteolytic activity and reduction of gliadin-like fractions by sourdough lactobacilli. J Appl Microbiol. (2005) 99:1495–15. doi: 10.1111/j.1365-2672.2005.02730.x
154. Gerez CL, Rollán GC, Font de Valdez G. Gluten breakdown by lactobacilli and pediococci strains isolated from sourdough. Lett Appl Microbiol. (2006) 42:459–64. doi: 10.1111/j.1472-765X.2006.01889.x
155. Gerez CL, Dallagnol A, Rollán G, Font de Valdez G. A combination of two lactic acid bacteria improves the hydrolysis of gliadin during wheat dough fermentation. Food Microbiol. (2012) 32:427–30. doi: 10.1016/j.fm.2012.06.007
156. Gobbetti M, Rizzello CG, Di Cagno R, De Angelis M. Sourdough lactobacilli and celiac disease. Food Microbiol. (2007) 24:187–96. doi: 10.1016/j.fm.2006.07.014
157. De Angelis M, Cassone A, Rizzello CG, Gagliardi F, Minervini F, Calasso M, et al. Mechanism of degradation of immunogenic gluten epitopes from Triticum turgidum L. var durum by sourdough lactobacilli and fungal proteases. Appl Environ Microbiol. (2010) 76:508–18. doi: 10.1128/AEM.01630-09
158. Zannini E, Pontonio E, Waters DM, Arendt EK. Applications of microbial fermentations for production of gluten-free products and perspectives. Appl Microbiol Biotechnol. (2012) 93:473–85. doi: 10.1007/s00253-011-3707-3
159. Ðorđević TM, Šiler-Marinković SS, Dimitrijević-Branković SI. Effect of fermentation on antioxidant properties of some cereals and pseudo cereals. Food Chem. (2010) 119:957–63. doi: 10.1016/j.foodchem.2009.07.049
160. Ferri M, Serrazanetti DI, Tassoni A, Baldissarri M, Gianotti A. Improving the functional and sensorial profile of cereal-based fermented foods by selecting Lactobacillus plantarum strains via a metabolomics approach. Food Res Int. (2016) 89:1095–105. doi: 10.1016/j.foodres.2016.08.044
161. Rizzello CG, Lorusso A, Russo V, Pinto D, Marzani B, Gobbetti M. Improving the antioxidant properties of quinoa flour through fermentation with selected autochthonous lactic acid bacteria. Int J Food Microbiol. (2017) 241:252–61. doi: 10.1016/j.ijfoodmicro.2016.10.035
162. Ayyash M, Johnson SK, Liu SQ, Mesmari N, Dahmani S, Al Dhaheri AS, et al. In vitro investigation of bioactivities of solid-state fermented lupin, quinoa and wheat using Lactobacillus spp. Food Chem. (2019) 275:50–8. doi: 10.1016/j.foodchem.2018.09.031
163. De Angelis M, Gallo G, Corbo MR, McSweeney PL, Faccia M, Giovine M, et al. Phytase activity in sourdough lactic acid bacteria: purification and characterization of a phytase from Lactobacillus sanfranciscensis CB1. Int J Food Microbiol. (2003) 87:259–70. doi: 10.1016/S0168-1605(03)00072-2
164. Corsetti A, Settanni L. Lactobacilli in sourdough fermentation. Food Res Int. (2007) 40:539–58. doi: 10.1016/j.foodres.2006.11.001
165. LeBlanc JG, Garro M, Rollán G, Savoy de Giori G. Chapter 13: Removal of antinutritional factors in soy-based foods using lactic acid bacteria. In: Ahmad A, editor. Nutrition, Consumption and Health. New York, NY: Publicado por Nova Science Publishers, Inc. (2013). p. 267–81.
166. Carrizo LS, Montes de Oca C, Laiño JE, Suarez NE, Vignolo G, LeBlanc JG, et al. Ancestral Andean grain quinoa as a source of functional lactic acid bacteria. Food Res Int. (2016) 89:488–94. doi: 10.1016/j.foodres.2016.08.013
167. Carrizo SL, Montes de Oca CE, Hebert ME, Saavedra L, Vignolo G, LeBlanc JG, et al. Lactic acid bacteria from Andean grain amaranth: source of functional value enzymes and vitamins. J Mol Microbiol Biotechnol. (2017) 27:289–98. doi: 10.1159/000480542
168. Kohajdová Z, Karovicová J. Fermentation of cereals for specific purpose. J Food Nutr Res. (2007) 46:51–7.
169. Rathore S, Salmerón I, Pandiella SS. Production of potentially probiotic beverages using single and mixed cereal substrates fermented with lactic acid bacteria cultures. Food Microbiol. (2012) 30:239–44. doi: 10.1016/j.fm.2011.09.001
170. Waters DM, Mauch A, Coffey A, Arendt EK, Zannini E. Lactic acid bacteria as a cell factory for the delivery of functional biomolecules and ingredients in cereal-based beverages: a review. Crit Rev Food Sci Nutr. (2013) 55:503–20. doi: 10.1080/10408398.2012.660251
171. Nionelli L, Coda R, Curiel JA, Poutanen K, Gobbetti M, Rizzello CG. Manufacture and characterization of a yogurt-like beverage made with oat flakes fermented by selected lactic acid bacteria. Int J Food Microbiol. (2014) 185:17–26. doi: 10.1016/j.ijfoodmicro.2014.05.004
172. Salmerón I, Thomas K, Pandiella SS. Effect of potentially probiotic lactic acid bacteria on the physicochemical composition and acceptance of fermented cereal beverages. J Functional Foods. (2015) 15:106–15. doi: 10.1016/j.jff.2015.03.012
173. Ludena Urquizo FE, García Torres SM, Tolonen T, Jaakkola M, Pena-Niebuhr MG, von Wright A, et al. Development of a fermented quinoa-based beverage. Food Sci Nutr. (2017) 5:602–8. doi: 10.1002/fsn3.436
174. Jeske S, Zannini E, Cronin MF, Arendt EK. Impact of protease and amylase treatment on proteins and the product quality of a quinoa-based milk substitute. Food Funct. (2018) 9:3500–8. doi: 10.1039/C8FO00336J
175. Jeske S, Zannini E, Lynch KM, Coffey A, Arendt EK. Polyol-producing lactic acid bacteria isolated from sourdough and their application to reduce sugar in a quinoa-based milk substitute. Int J Food Microbiol. (2018) 286:31–6. doi: 10.1016/j.ijfoodmicro.2018.07.013
176. Rodríguez H, Landete JM, Rivas B, Muñoz R. Metabolism of food phenolic acids by Lactobacillus plantarum CECT 748T. Food Chem. (2009) 107:1393–8. doi: 10.1016/j.foodchem.2007.09.067
177. Barthelmebs L, Divies C, Cavin JF. Knockout of the p-coumarate decarboxylase gene from Lactobacillus plantarum reveals the existence of two other inducible enzymatic activities involved in phenolic acid metabolism. Appl Environ Microbiol. (2000) 66:3368–75. doi: 10.1128/AEM.66.8.3368-3375.2000
178. Rodríguez H, Landete JM Curiel JA, de las Rivas B, Mancheño JM, Muñoz R. Characterization of the p-coumaric acid decarboxylase from Lactobacillus plantarum CECT 748T. J Agric Food Chem. (2008) 56:3068–72. doi: 10.1021/jf703779s
179. Torres ME, Landete JM, Reverón I, Santamaría L, Rivas B, Muñoz R. A Lactobacillus plantarum esterase active on a broad range of phenolic esters. Appl Environ Microbiol. (2015) 81:3235–42. doi: 10.1128/AEM.00323-15
180. Filannino P, Cardinali G, Rizzello CG, Buchin S, De Angelis M, Gobbetti M, et al. Metabolic responses of Lactobacillus plantarum strains during fermentation and storage of vegetable and fruit juices. Appl Environ Microbiol. (2014) 80:2206–15. doi: 10.1128/AEM.03885-13
181. Filannino P, Di Cagno R, Crecchio C, De Virgilio C, De Angelis M, Gobbetti M. Transcriptional reprogramming and phenotypic switching associated with the adaptation of Lactobacillus plantarum C2 to plant niches. Sci Rep. (2016) 6:27392. doi: 10.1038/srep27392
182. Golomb BL, Marco ML. Lactococcus lactis metabolism and gene expression during growth on plant tissues. J Bacteriol. (2015) 197:371–81. doi: 10.1128/JB.02193-14
183. De Angelis M, Minervini F, Siragusa S, Rizzello CG, Gobbetti M. Wholemeal wheat flours drive the microbiome and functional features of wheat sourdoughs. Int J Food Microbiol. (2018) 302:35–46. doi: 10.1016/j.ijfoodmicro.2018.08.009
184. Katina K, Laitila A, Juvonen R, Liukkonen KH, Kariluoto S, Piironen V, et al. Bran fermentation as a means to enhance technological properties and bioactivity of rye. Food Microbiol. (2007) 24:175–86. doi: 10.1016/j.fm.2006.07.012
185. Katina K, Liukkonen KH, Kaukovirta-Norja A, Adlercreutz H, Heinonen SM, Lampi AM, et al. Fermentation-induced changes in the nutritional value of native or germinated rye. J Cereal Sci. (2007) 46:348–55. doi: 10.1016/j.jcs.2007.07.006
186. Rizzello CG, Lorusso A, Montemurro M, Gobbetti M. Use of sourdough made with quinoa (Chenopodium quinoa) flour and autochthonous selected lactic acid bacteria for enhancing the nutritional, textural and sensory features of white bread. Food Microbiol. (2016) 56:1–13. doi: 10.1016/j.fm.2015.11.018
187. Matejćeková Z, Liptá ková D, Valík Z. Evaluation of the potential of amaranth flour for lactic acid fermentation. J Pharm Pharm Nutr Sci. (2016) 6:1–6. doi: 10.6000/1927-5951.2016.06.01.1
188. Soltani M, Hekmat S, Ahmadi L. Microbial and sensory evaluation of probiotic yoghurt supplemented with cereal/pseudo-cereal grains and legumes. Int J Dairy Thecnol. (2017) 70:1–8. doi: 10.1111/1471-0307.12389
189. Karademir E, Yalçin E. Effect of fermentation on some quality properties of cornelian cherry tarhana produced from different cereal/pseudocereal flours. Qual Assur Saf Crops Foods. (2019) 11:127–35. doi: 10.3920/QAS2018.1389
190. Lorusso A, Verni M, Montemurro M, Coda R, Gobbetti M, Rizzello CG. Use of fermented quinoa flour for pasta making and evaluation of the technological and nutritional features. LWT Food Sci Technol. (2017) 78:215–21. doi: 10.1016/j.lwt.2016.12.046
191. Sánchez-Maldonado AF, Schieber A, Gänzle MG. Structure–function relationships of the antibacterial activity of phenolic acids and their metabolism by lactic acid bacteria. J Appl Microbiol. (2011) 111:1176–84. doi: 10.1111/j.1365-2672.2011.05141.x
192. Filannino P, Gobbetti M, De Angelis M, Di Cagno R. Hydroxycinnamic acids used as external acceptors of electrons: an energetic advantage for strictly hetero-fermentative lactic acid bacteria. Appl Environ Microbiol. (2014) 80:7574–82. doi: 10.1128/AEM.02413-14
193. Alberto MR, Gómez Cordovés C, Manca de Nadra MC. Metabolism of gallic acid and catechin by Lactobacillus hilgardii from wine. J Agricul Food Chem. (2004) 52:6465–9. doi: 10.1021/jf049239f
194. LoCascio R, Mills DA, Waterhouse AL. Reduction of catechin, rutin, and quercetin levels by interaction with food-related microorganisms in a resting state. J Sci Food Agric. (2006) 86:2105–12. doi: 10.1002/jsfa.2583
195. Gobbetti M, De Angelis M, Di Cagno R, Calasso M, Archetti G, Rizzello CG. Novel insights on the functional/nutritional features of the sourdough fermentation. Int J Food Microbiol. (2018) 302:103–113. doi: 10.1016/j.ijfoodmicro.2018.05.018
196. Rodríguez H, de las Rivas B, Gómez-Cordovés C, Muñoz R. Degradation of tannic acid by cell-free extracts of Lactobacillus plantarum. Food Chem. (2008) 107:664–70. doi: 10.1016/j.foodchem.2007.08.063
197. Jiménez N, Curiel AJ, Reverón I, Rivas B, Muñoz R. Uncovering the Lactobacillus plantarum WCFS1 gallate decarboxylase involved in tannin degradation. Appl Environ Microbiol. (2013) 79:4253–63. doi: 10.1128/AEM.00840-13
198. Huynh NT, Camp JV, Smagghe G, Raes K. Improved release and metabolism of flavonoids by steered fermentation processes: a review. Int J Mol Sci. (2014) 15:19369–88. doi: 10.3390/ijms151119369
199. Santamaría L, Reverón I, López F, De Las Rivas B, Muñoz R. Unravelling the reduction pathway as an alternative metabolic route to hydroxycinnamate decarboxylation in Lactobacillus plantarum. Appl Environ Microbiol. (2018) 84:e01123–18. doi: 10.1128/AEM.01123-18
200. Hirose Y, Fujita T, Ishii T, Ueno N. Antioxidative properties and flavonoid composition of Chenopodium quinoa seeds cultivated in Japan. Food Chem. (2010) 119:1300–6. doi: 10.1016/j.foodchem.2009.09.008
201. Lee CC, Shen SR, Lai YJ, Wu SC. Rutin and quercetin, bioactive compounds from tartary buckwheat, prevent liver inflammatory injury. Food Funct. (2013) 4:794–802. doi: 10.1039/c3fo30389f
202. Kocevar Glavac N, Stojilkovski K, Kreft S, Park CH, Kreft I. Determination of fagopyrins, rutin, and quercetin in Tartary buckwheat products. LWT Food Sci Technol. (2017) 79:423–7. doi: 10.1016/j.lwt.2017.01.068
203. Koistinen VM, Katina K, Nordlund E, Poutanen K, Hanhineva K. Changes in the phytochemical profile of rye bran induced by enzymatic bioprocessing and sourdough fermentation. Food Res Int. (2016) 89:1106–15. doi: 10.1016/j.foodres.2016.06.027
204. Koistinen VM, Mttila O, Katina K, Poutanen K, Aura AM, Hanhineva K. Metabolic profiling of sourdough fermented wheat and rye bread. Sci Rep. (2018) 8:5684. doi: 10.1038/s41598-018-24149-w
205. Shin NR, Moon JS, Shin SY, Li L, Lee YB, Kim TJ, et al. Isolation and characterization of human intestinal Enterococcus avium EFEL009 converting rutin to quercetin. Lett Appl Microbiol. (2015) 62:68–74. doi: 10.1111/lam.12512
206. Zielinski H, Szawara-Nowak D, Baczek N, Wronkowska M. Effect of liquid-state fermentation on the antioxidant and functional properties of raw and roasted buckwheat flours. Food Chem. (2019) 271:291–7. doi: 10.1016/j.foodchem.2018.07.182
207. Yang S, Cao C, Chen S, Hu LY, Bao W, Shi HD, et al. Serum metabolomics analysis of quercetin against acrylamide-induced toxicity in rats. J Agricul Food Chem. (2016) 64:9237–45. doi: 10.1021/acs.jafc.6b04149
208. Fan M, Zhang G, Hu X, Xu X, Gong D. Quercetin as a tyrosinase inhibitor: Inhibitory activity, conformational change and mechanism. Food Res Int. (2017) 100:226–33. doi: 10.1016/j.foodres.2017.07.010
209. Xiao J. Dietary flavonoid aglycones and their glycosides: which show better biological significance? Crit Rev Food Sci Nutr. (2017) 57:1874–905. doi: 10.1080/10408398.2015.1032400
210. LeBlanc JG, Sybesma W, Starrenburg M, Sesma F, de Vos WM, Hugenholtz J, et al. Supplementation with engineered Lactococcus lactis improves the folate status in deficient rats. Nutrition. (2010) 26:835–41. doi: 10.1016/j.nut.2009.06.023
211. LeBlanc JG, Milani C, Savoy de Giori G, Sesma F, van Sinderen D, Ventura M. Bacteria as vitamin suppliers to their host: a gut microbiota perspective. Curr Opin Biotechnol. (2013) 24:160–8. doi: 10.1016/j.copbio.2012.08.005
212. Laiño JE, LeBlanc JG, Savoy de Giori G. Selection of folate producing starter cultures of yogurt isolated from Northwestern Argentina. Can. J Microbiol. (2012) 58:581–8. doi: 10.1139/w2012-026
213. Laiño J, Juarez del Valle M, Savoy de Giori G, LeBlanc JG. Applicability of a Lactobacillus amylovorus strain as co-culture for obtaining of a novel naturally bio-enriched in folate fermented milk. Int J Food Microbiol. (2014) 191:10–6. doi: 10.1016/j.ijfoodmicro.2014.08.031
214. Juarez del Valle M, Laiño JE, de Moreno de LeBlanc A, Savoy de Giori G, LeBlanc JG. Soymilk fermented with riboflavin producing Lactobacillus plantarum CRL 2130 reverts and prevents ariboflavinosis in murine models. Br J Nutr. (2016) 116:1229–35. doi: 10.1017/S0007114516003378
215. Salvucci E, LeBlanc JG, Perez G. Technological characterization of lactic acid bacteria isolated from raw cereal material. LWT Food Sci Technol. (2016) 70:185–91. doi: 10.1016/j.lwt.2016.02.043
216. Kariluoto S, Aittamaa M, Korhola M, Salovaara H, Vahteristo L, Piironen V. Effects of yeasts and bacteria on the levels of folates in rye sourdoughs. Int J Food Microbiol. (2006) 106:137–43. doi: 10.1016/j.ijfoodmicro.2005.06.013
217. Herranen M, Kariluoto S, Edelmann M, Piironen V, Ahvenniemi K, Iivonen V, et al. Isolation and characterization of folate-producing bacteria from oat bran and rye flakes. Int J Food Microbiol. (2010) 142:277–85. doi: 10.1016/j.ijfoodmicro.2010.07.002
218. Carrizo SL. Aplicación de bacterias lácticas en el desarrollo de alimentos novedosos a base de seudocereales (Thesis). Universidad Nacional de Tucumán, Tucumán, Argentina (2018).
219. McNulty H. Folate requirements for health in different population groups. Br J Biomed Sci. (1995) 52:110–9.
220. Dewey KG, Brown KH. Update on technical issues concerning complementary feeding of young children in developing countries and implications for intervention programs. Food Nutr Bull. (2003) 24:5–28. doi: 10.1177/156482650302400102
221. Dop MC, Benbouzid D. Regional features of complementary feeding in Africa and middle east. In: MC Dop D, Benbouzid S, Trèche B, de Benoist A, Verster F, editors. Delpeuch Complementary Feeding of Young Children in Africa and the Middle East. Geneva: World Health Organization (1999). p. 43–58.
222. Mouquet-Rivier C, Icard-Vernière C, Guyot JP, Hassane Tou E, Rochette I, Trèche S. Consumption pattern, biochemical composition and nutritional value of fermented pearl-millet gruels in Burkina Faso. Int J Food Sci Nutr. (2008) 59:716–29. doi: 10.1080/09637480802206389
223. Singh A, Rehal J, Kaur A, Jyot G. Enhancement of attributes of cereals by germination and fermentation: a review. Crit Rev Food Sci Nutr. (2015) 55:1575–89. doi: 10.1080/10408398.2012.706661
224. Capozzi V, Menga V, Digesu AM, De Vita P, van Sinderen D, Cattivelli L, et al. Biotechnological production of vitamin B2-enriched bread and pasta. J Agric Food Chem. (2011) 59:8013–20. doi: 10.1021/jf201519h
225. Russo P, Capozzi V, Arena MP, Spadaccino G, Dueñas MT, López P, et al. Riboflavin-overproducing strains of Lactobacillus fermentum for riboflavin-enriched bread. Appl Microbiol Biotechnol. (2014) 98:3691–700. doi: 10.1007/s00253-013-5484-7
226. Capozzi V, Russo P, Dueñas MT, López P, Spano G. Lactic acid bacteria producing B-group vitamins: a great potential for functional cereals products. Appl Microbiol Biotechnol. (2012) 96:1383–94. doi: 10.1007/s00253-012-4440-2
227. Taranto MP, Vera JL, Hugenholtz J, Font de Valdez G, Sesma F. Lactobacillus reuteri CRL1098 produces cobalamin. J Bacteriol. (2003) 185:5643–7. doi: 10.1128/JB.185.18.5643-5647.2003
228. Varmanen P, Deptula P, Chamlagain B, Piironen V. Letter to the editor on Enhancing vitamin B12 content in soy-yogurt by Lactobacillus reuteri. Int J Food Microbiol. (2016) 206:56–9. doi: 10.1016/j.ijfoodmicro.2016.03.029
229. Molina V, Medici M, Font de Valdez G, Taranto MP. Soybean-based functional food with vitamin B12-producing lactic acid bacteria. J Funct Foods. (2012) 4:831–6. doi: 10.1016/j.jff.2012.05.011
230. Nout MJR. Processed weaning foods for tropical climates. Int J Food Sci Nutr. (1993) 43:13–22. doi: 10.3109/09637489309027545
231. Mullaney EJ, Daly CB, Kim T, Porres JM, Lei XG, Sethumadhavan K, et al. Site-directed mutagenesis of Aspergillus niger NRRL 3135 phytase at residue 300 to enhance catalysis at pH 4.0. Biochem Biophys Res Commun. (2002) 297:1016–20. doi: 10.1016/S0006-291X(02)02325-2
232. Vohra A, Satyanarayana T. Phytases: microbial sources, production, purification, and potential biotechnological applications. Crit Rev Biotechnol. (2003) 23:29–60. doi: 10.1080/713609297
233. Lei XG, Porres JM, Mullaney EJ, Brinch-Pedersen H. Phytase: source, structure and application. In: Polaina J, Mac-Cabe AP, editors. Industrial Enzymes-Structure, Function and Applications. Dordrecht: Springer (2007). p. 505–29. doi: 10.1007/1-4020-5377-0_29
234. Masud T, Mahmood T, Latif A, Sammi S, Hameed T. Influence of processing and cooking methodologies for reduction of phytic acid content in wheat (Triticum aestivum) varieties. J Food Process Pres. (2007) 31:583–94. doi: 10.1111/j.1745-4549.2007.00147.x
235. Egli I, Davidsson L, Juillerat MA, Barclay D, Hurrell RF. The influence of soaking and germination on the phytase activity and phytic acid content of grains and seeds potentially useful for complementary feeding. J Food Sci. (2002) 67:3484–8. doi: 10.1111/j.1365-2621.2002.tb09609.x
236. Greiner R, Konietzny U. Phytase for food application. Food Technol Biotechnol. (2006) 44:125–40.
237. Türk M, Sandberg AS. Phytate degradation during breadmaking - effect of phytase addition. J Cereal Sci. (1992) 15:281–94. doi: 10.1016/S0733-5210(09)80126-4
238. Lopez HW, Ouvry A, Bervas E, Guy C, Messager A, Demigne C, et al. Strains of lactic acid bacteria isolated from sour doughs degrade phytic acid and improve calcium and magne- sium solubility from whole wheat flour. J Agric Food Chem. (2000) 48:2281–5. doi: 10.1021/jf000061g
239. Singh B, Kunze G, Satyanarayana T. Developments in biochemical aspects and biotechnological applications of microbial phytases. Biotechnol Mol Biol Rev. (2011) 6:69–87.
240. Rebello S, Jose L, Sindhu R, Aneesh EM. Molecular advancements in the development of thermostable phytases. Appl Microbiol Biotechnol. (2017) 101:2677–89. doi: 10.1007/s00253-017-8195-7
241. Simons PCM, Versteegh HAJ, Jongbloed AW, Kemme PA, Slump P, Bos KD, et al. Improvement of phosphorus availability by microbial phytase in broilers and pigs. Br J Nutr. (1990) 64:525–40. doi: 10.1079/BJN19900052
242. Idriss E, Makarewicz O, Farouk A, Rosner K, Greiner R, Bochow HT, et al. Extracellular phytase activity of Bacillus amyloliquefaciens FZB45 contributes to its plant-growth-promoting effect. Microbiol. (2002) 148:2097–109. doi: 10.1099/00221287-148-7-2097
243. Yano F, Nakajima T, Matsuda M. Reduction of nitrogen and phosphorus from livestock waste: a major priority for intensive animal production. Asian-Aust, J Anim Sci. (1999) 12:651–6. doi: 10.5713/ajas.1999.651
244. Greiner R, Carlsson NG. Myo-Inositol phosphate isomers generated by the action of a phytate-degrading enzyme from Klebsiella terrigena on phytate. Can J Microbiol. (2006) 52:759–68. doi: 10.1139/w06-028
245. Selle PH, Ravindran V. Microbial phytase in poultry nutrition. Animal Feed Sci Technol. (2007) 135:1–41. doi: 10.1016/j.anifeedsci.2006.06.010
246. Theodoropoulos VCT, Turatti MA, Greiner R, Macedo GA, Azevedo Lima Pallone J. Effect of enzymatic treatment on phytate content and mineral bioacessability in soy drink. Food Res Int. (2018) 108:68–73. doi: 10.1016/j.foodres.2018.03.018
247. Vasudevan UM, Krishna S, Jalaja V, Pandey A. Microbial phytase: Impact of advances in genetic engineering in revolutionizing its properties and applications. Bioresource Technol. (2017) 245:1790–9. doi: 10.1016/j.biortech.2017.05.060
248. Vogel RF, Müller M, Stolz P, Ehrmann M. Ecology in sourdoughs produced by traditional and modern technologies. Adv Food Sci. (1996) 18:152–9.
249. Palacios MC, Haros M, Rosell CM, Sanz Y. Characterization of an acid phosphatase from Lactobacillus pentosus: regulation and biochemical properties. J Appl Microbiol. (2005) 98:229–37. doi: 10.1111/j.1365-2672.2004.02447.x
250. Rizzello CG, Nionelli L, Coda R, de Angelis M, Gobbetti M. Effect of sourdough fermentation on stabilisation, and chemical and nutritional characteristics of wheat germ. Food Chem. (2010) 119:1079–89. doi: 10.1016/j.foodchem.2009.08.016
251. Anastasio M, Pepe O, Cirillo T, Palomba S, Blaiotta G, Villani F. Selection and use of phytate-degrading LAB to improve cereal-based products by mineral solubilization during dough fermentation. J Food Sci. (2010) 75:28–35. doi: 10.1111/j.1750-3841.2009.01402.x
252. Cizeikiene D, Juodeikiene G, Bartkiene E, Damasius J, Paskevicius A. Phytase activity of lactic acid bacteria and their impact on the solubility of minerals from wholemeal wheat bread. Int J. Food Sci Nutr. (2015) 66:736–42. doi: 10.3109/09637486.2015.1088939
253. Nuobariene L, Cizeikiene D, Gradzeviciute E, Hansen ÅS, Rasmussen SK, Juodeikiene G, et al. Phytase-active lactic acid bacteria from sourdoughs: Isolation and identification. LWT Food Sci Technol. (2015) 63:766–72. doi: 10.1016/j.lwt.2015.03.018
254. Priyodip P, Balaji S. Microbial degradation of myo-inositol hexakisphosphate (IP6): specificity, kinetics, and simulation. Biotech. (2018) 8:268. doi: 10.1007/s13205-018-1302-3
Keywords: lactic acid bacteria, pseudocereals, vitamins, phytate, phytochemicals
Citation: Rollán GC, Gerez CL and LeBlanc JG (2019) Lactic Fermentation as a Strategy to Improve the Nutritional and Functional Values of Pseudocereals. Front. Nutr. 6:98. doi: 10.3389/fnut.2019.00098
Received: 31 January 2019; Accepted: 14 June 2019;
Published: 03 July 2019.
Edited by:
Rossana Coda, University of Helsinki, FinlandReviewed by:
Pasquale Russo, University of Foggia, ItalyFrancesca Valerio, Italian National Research Council (CNR), Italy
Copyright © 2019 Rollán, Gerez and LeBlanc. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Graciela C. Rollán, cm9sbGFuQGNlcmVsYS5vcmcuYXI=