
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 19 February 2025
Sec. Microbe and Virus Interactions with Plants
Volume 15 - 2024 | https://doi.org/10.3389/fmicb.2024.1501057
This article is part of the Research Topic Plant Microbiome: Interactions, Mechanisms of Action, and Applications, Volume III View all 22 articles
 Kuan Yan1,2*Xinyi Li1,2Yu Cai1,2Lina Meng1,2Qin Wei1,2Xianming Zhao1,2Rania M. Y. Heakel3Amr M. Atif4Mohamed A. Abd Elhamid3Salma A. Soaud3Ahmed H. El-Sappah1,2,3*
Kuan Yan1,2*Xinyi Li1,2Yu Cai1,2Lina Meng1,2Qin Wei1,2Xianming Zhao1,2Rania M. Y. Heakel3Amr M. Atif4Mohamed A. Abd Elhamid3Salma A. Soaud3Ahmed H. El-Sappah1,2,3*Introduction: Bamboo is a sustainable and degradable resource for sustenance, high-strength cellulose microfibers, and synthetic fiber in China. Endophytic bacteria enhance root development and ethylene levels, benefiting the host plants’ physiology.
Methods: We investigated the population, diversity, and abundance of endophytic bacteria in the leaves of three bamboo species—Phyllostachys edulis, Bambusa rigida, and Pleioblastus amarus—using high-throughput 16S rDNA sequencing.
Results and discussion: A total of 1,159 operational taxonomic units (OTUs) were obtained and further classified into 26 phyla, 64 classes, 158 orders, 270 families, 521 genera, and 811 species. The phyla with the highest abundance were Proteobacteria, Actinobacteria, and Myxococcota, and the highest genera included 1,174–901-12, Sphingomonas, and unclassified_f__Enterobacteriaceae. The relative richness of endophytic bacteria in the three species was in the following order: B. rigida > P. amarus > Ph. edulis. The PICRUSt functional richness analysis of endophytic bacteria indicated their involvement in six biological pathways: “cellular processes,” “environmental information processing,” “genetic information processing,” “human diseases,” “metabolism,” and “organic systems.” Among the 41 sub-functions, the most common were “amino acid metabolism,” “carbohydrate metabolism,” “cell motility,” “cellular signaling,” “energy metabolism,” and “membrane transport.” Our results provide precise knowledge for better managing bamboo forests and pave the way for isolating secondary metabolites and potential bioactive compounds.
Microbial endophytes are microorganisms that live within the tissues of plants and form symbiotic relationships with their hosts (Gond et al., 2015). Anti-pathogenic bacteria, plant endophytic bacteria, abiotic stress tolerance, and physiology enhancement in host plants are all outcomes of their interactions (Hardoim et al., 2008). Plant endophytic bacteria are significant biological resources commonly employed across various industries (Donoso et al., 2017; Yan et al., 2021a). The species richness and diverse community structure of endophytes in plants are primarily determined by various factors, including plant species, seed vector endophytes, life cycle, adjacent environment, and plant tissues (Yan et al., 2022). Diverse endophytes are found in greater abundance in leaves than in any other tissue due to their susceptibility to airborne damage, stomatal openings, and transportability (Cui et al., 2017).
Generally, distinct endophytic bacterial species inhabit a variety of plant species (Liu et al., 2021). A comprehensive examination unveiled that even variations of identical plant species support a wide array of endophyte community structures and species diversity (Massimo et al., 2015). An investigation by Yuan et al. (2014) revealed a notably heterogeneous endophytic microbiome in rhizomes procured from different varieties of moso bamboo across distinct regions. A total of 129 species and 54 genera of endophytic bacteria have been identified in diverse plant taxa (White et al., 2015). Proteus, Bacillus, Enterobacter, and Azospirillum are notably abundant genera. In addition, the diversity of the endophytic bacterial community is influenced by the surrounding environment and the diversity of the endophyte host plants. Factors such as plant species, life span, location, and various plant tissues are regulated in endophytic bacterial diversity regulation. Many endophytic bacterial strains have been identified in the aerial portions of plants, specifically the leaves, except for strains specific to particular tissues. This is because leaves are more exposed to the environment and are more delicate (Cui et al., 2017).
Bambusoideae, a member of the Poaceae family, is renowned for its rapid growth, adaptability, and versatility. Its abundant species diversity spans over 70 genera and 1,200 species across the globe (Liu et al., 2017). Bamboo is a forest resource that contributes significantly to the regional economy and contemporary forestry by incorporating ecological, social, and economic benefits. The paper and pulp industries extensively use bamboo cellulose. It is a renewable primary material utilized in the construction and furniture industries. Recently, bamboo microfibers with a strength of 1.26 ± 0.21 GPa cm−3 g−1 have been extracted; these fibers are more powerful than steel and will bring about a paradigm shift in the construction sector (Li et al., 2022). Because of its adaptability to a wide range of environmental conditions and the abundance of bamboo species, it is capable of harboring a greater variety of microbial species (Shen et al., 2014; Zhou et al., 2017). The investigation of endophytic bacteria in Phyllostachys praecox and Phyllostachys edulis has extended beyond the Shunan bamboo sea (Zhang et al., 2019). Bamboo is being developed extensively as an ecological and economic forest, but most research focuses on material quality and variety propagation. For example, the growth, composition, and mechanics of Ph. edulis have been the subject of numerous studies (Xu et al., 2011; Wang et al., 2012). The majority of studies on Pleioblastus amarus focus on its medicinal and edible properties (Ying et al., 2022). Bambusa rigida, a native bamboo species in Sichuan, has significant timber and papermaking resource development (Wen et al., 2011; Liu et al., 2014). However, research on B. rigida is currently limited and primarily concerns itself with lignocellulose. Therefore, three bamboo species with entirely different genetic characteristics and uses were selected to study the diversity of endophytic bacteria, which can help in better understanding the differences in the endophytic bacterial community structure and function between them.
Endophyte research is crucial for the development of the bamboo-based industry, the maintenance of bamboo forest ecosystems, and the cultivation of bamboo forests. Numerous microorganisms continue to be uncultivable, rendering conventional cultivation methods disappointing. The utilization of next-generation sequencing techniques has enabled a comprehensive examination of the microbiome of any given species. Therefore, three representative bamboos, Phyllostachys edulis, Bambusa rigida, and Pleioblastus amarus, from the Shunan Bamboo Sea, Yibin, China, were selected as the research objects. The endophytic bacteria in the leaves of the three bamboo species were analyzed by molecular biotechnology, and their community structure and diversity characteristics were revealed.
The methods of this study follow relevant institutional, national, and international guidelines and legislation. This study protocol also adheres to the IUCN Policy Statement on Research Involving Species at Risk of Extinction and the Convention on the Trade in Endangered Species of Wild Fauna and Flora.
To investigate the density and diversity of endophytic bacteria leaves, the study material was collected from Ph. edulis (MZ), B. rigida (YT), and Pl. amarus (KZ) located in the Shunan Bamboo Sea (Yibin, Sichuan, 28°30′2″N, 105°04′7″E), China. The sample plot of each bamboo species was set up into three blocks. In each block, five bamboos with vigorous growth, far away from the edge zone, free from pests and diseases, and with an interval of about 15 m were selected. The leaves of these bamboos were collected and mixed into a sample, which was quickly maintained at 4°C. Upon arriving at the laboratory, all appendages were trimmed away, and dirt areas on the surface of leaves were thoroughly cleaned with running tap water. Leaves of each growth stage were weighed (10 g), packed in sterile bags, and stored at 4°C for future use as research material. Surface sterilization of leaves was performed via soaking in 75% ethanol for 30 s and 2% NaClO solution for 2 min with gene shaking, followed by three washings with sterile ddH2O. To ensure proper surface sterilization of leaves, 20 mL of ddH2O from the last rinse of each sample was streaked on Potato Dextrose Agar (PDA) medium in three biological replicates and incubated at 28°C under dark conditions.
To extract DNA, 5 g of sample was weighed, chopped into small pieces, and immediately soaked in 50 mL sterile Tween-NaCl buffer [0.9% NaCl (w/v), 0.05% Tween 20 (v/v), and 2% polyvinylpolypyrrolidone (w/v)], homogenized, and sonicated at 4°C for 30 min, as described by Yan et al. (2021b). Subsequently, the solution was filtered through a three-layered sterile gauze to remove coarse particles, followed by centrifugation at 12,298 × g for 10 min at 4°C. The supernatant was discarded, and the residue was shifted into a new sterile tube for genomic DNA (gDNA) extraction. We employed an E.Z.N.A™ HP Plant DNA Kit (OMEGA®, USA) by following the manufacturer’s standard protocol to isolate bacterial genomic DNA. To avoid noise during polymerase chain reaction (PCR), impurities in bacterial genomic DNA, such as polyphenol, were removed with the help of the E.Z.N.A™. Soil Kit. The quality of the bacterial genomic DNA was confirmed by running 2 μL of extracted DNA of each sample on 1% agarose gel and visualized under a UV light-installed gel documentation system (iBright 1,500 imaging system, ThermoFisher, USA) (El-Sappah et al., 2017). Finally, DNA concentrations were recorded on a nanodrop spectrophotometer (NanoDrop 2000) at OD 260/280, which were 2.1–42.5 ng/μl in each sample (El-Sappah et al., 2021).
In order to design primers to use in PCR to amplify bacterial rRNA, we used the Premier 5 tool (Wang et al., 2018). The following primer pair, 799F(5’-AACMGGATTAGATACCCKG-3′) and 1193R(5’-ACGTCATCCCCACCTTCC-3′), was used to amplify the V5–V7 regions of the bacterial 16S rRNA gene (Chen et al., 2018). The PCR system consisted of 5 μL of 10x KOD buffer, 5 μL of 2 mmol/L dNTPs, 2 μL of MgSO4, 1 μL of KOD Plus enzyme, 1.5 μL of each primer pair (799F and 1193R), DNA template (total DNA of leaves and endophytes 50 ng), and the total volume was adjusted to 50 μL by adding sterile ddH2O. The following PCR conditions were used: initial denaturation at 94°C for 2 min; denaturation at 94°C for 30 s; primer binding at 63°C for 30 s; amplification at 68°C for 30 s, with a total of 30 cycles; final amplification at 68°C for 5 min; and cooling at 4°C indefinitely. PCR for each sample was performed in three biological replicates, and the total amplification product of each sample was the sum of the three replicate PCR products. Finally, high-quality PCR products were further used in library construction and subsequent sequencing analysis using robust Illumina MiSeq™ (PE300).
To identify operational taxonomic units (OTUs) in sequencing reads, phylogenetic clustering analysis was performed using a Uparse v7.0.10901 OTU clustering tool at a 97% identity threshold level (Ye et al., 2017). Chimeric reads in sequencing data were identified with the help of UCHIME v4.2 software and discarded (Edgar et al., 2011). A group of eight bases was called “word.” The combinations were calculated and compared to the reference library at 100 times the bootstrap cutoff value. The RDP classifier Bayesian algorithm was employed at an 80% confidence threshold level to measure the confidence score of taxa assignment at each taxonomic level. The composition of endophytic bacterial communities in each sample of bamboo leaves was classified into kingdom, phylum, class, order, family, genus, and species (Chen et al., 2016). The common and unique OTU numbers in each sample were counted and illustrated by constructing a Venn diagram (Fouts et al., 2012).
The diversity of endophytic bacteria in bamboo leaves was analyzed. α-Diversity represents sequencing depth, coverage, and comparison of abundance and diversity of endophytes in a microbial community. Good’s coverage metric (C = 1-n1/N) was employed to estimate sequence coverage. Herein, “n1” is the number of OTUs containing only one sequence read, and “N” is the total number of sequence reads obtained from one sample (He et al., 2018). α-Diversity of three biological repeats of each sample was analyzed by employing Chao 1, ACE, Shannon, and Simpson indices (Gihring et al., 2012; Rogers et al., 2016). The linear discriminant analysis (LDA) effect size (LEfSe)1 was used to perform the LDA on samples with different grouping conditions according to taxonomic composition. Euclidean distances, dissimilarity indices, and diversity in OTUs at a 97% identity threshold were calculated in each sample by principal coordinate analysis (PCoA) (Calderón et al., 2017). In order to investigate the composition of dominant species in each sample and their distribution among different samples, a Circos sample–species relationship map was constructed with the help of Circos-0.67-72 software (Sui et al., 2016). PICRUSt package was employed for KEGG functional prediction of each 16S amplicon in an OTU (Tang et al., 2018). Comparative taxonomic analyses of the endophytic bacterial community were performed at each classification level in all samples. The R tool was used to construct a community structure diagram and histogram (Lu et al., 2016; Zhou et al., 2017).
The analysis was performed using a one-way analysis of variance (ANOVA), with all data points represented as means. Additionally, at a significance level of p < 0.05, Tukey’s HSD test was employed to examine differences in means of Shannon, Simpson, ACE, Chao, and diversity indices. All correlation and path coefficient analyses were conducted using Excel 2019 and SPSS Statistics 20.0 (SPSS Inc., Chicago, IL, United States).
In triplicate, the 16S rDNA gene was used to analyze the sequence readings of all three bamboo leaf samples, MZ, KZ, and YT (Table 1).
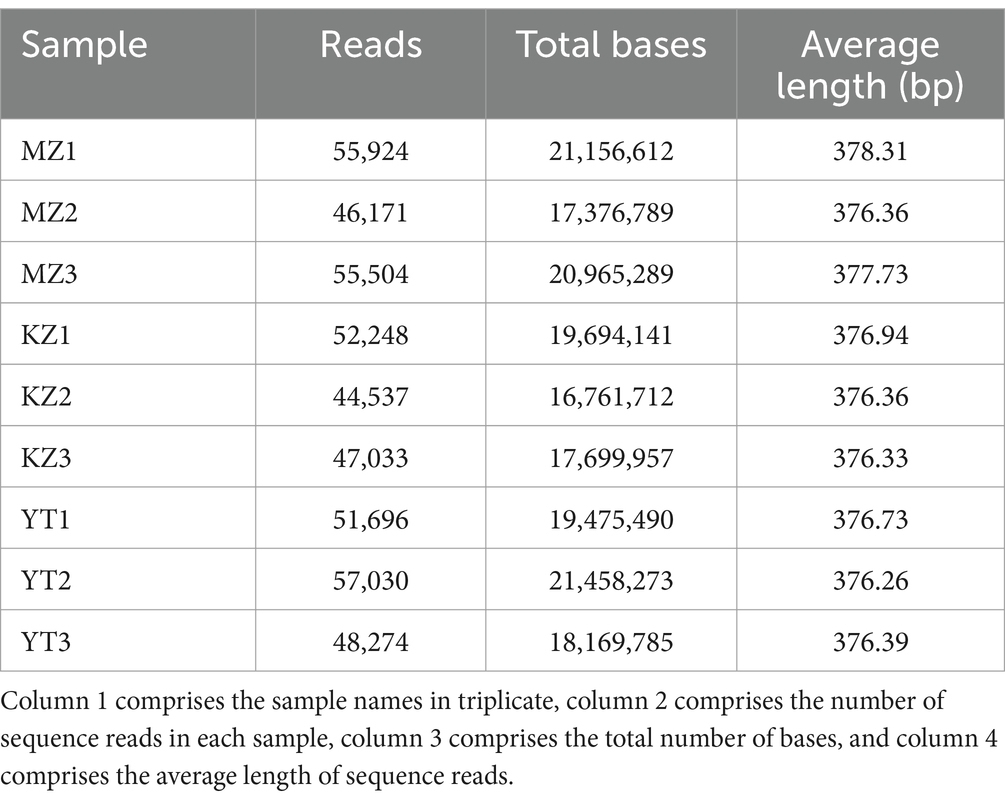
Table 1. Sequencing reads analysis of endophytic bacteria in the three samples of bamboo leaves in triplicate.
All reads that were incomplete or redundant were eliminated. Full-length recovered reads from MZ bamboo leaf samples were 55,924, 46,171, and 55,504 bp; total number of bases were 21,156,612, 17,376,789, and 20,965,289 bp, and average sequence lengths were 378.31, 376.36, and 377.73 bp. Full-length reads from KZ samples were 52,248, 44,537, and 47,033; the total base count was 19,694,141, 16,761,712, and 17,699,957 bp, and the average read length was 376.94, 376.36, and 376.33 bp. Similarly, in YT samples, the total length sequence reads were 51,696, 57,030, and 48,274, the total number of bases was 19,475,490, 21,458,273, and 18,169,785 bp, and the average sequence length was 376.73, 376.26, and 376.39 bp.
The rarefaction curves show the coverage, sampling depth, richness, and uniformity of the endophytic bacterial species present in each bamboo leaf sample. We saw a log phase in all three samples followed by an abrupt flat trend, except for the MZ sample, which had a reasonably short span and a fast decline. The flatness of all three curves demonstrated the abundance of bacterial communities in each sample and sufficient sampling (Figure 1).
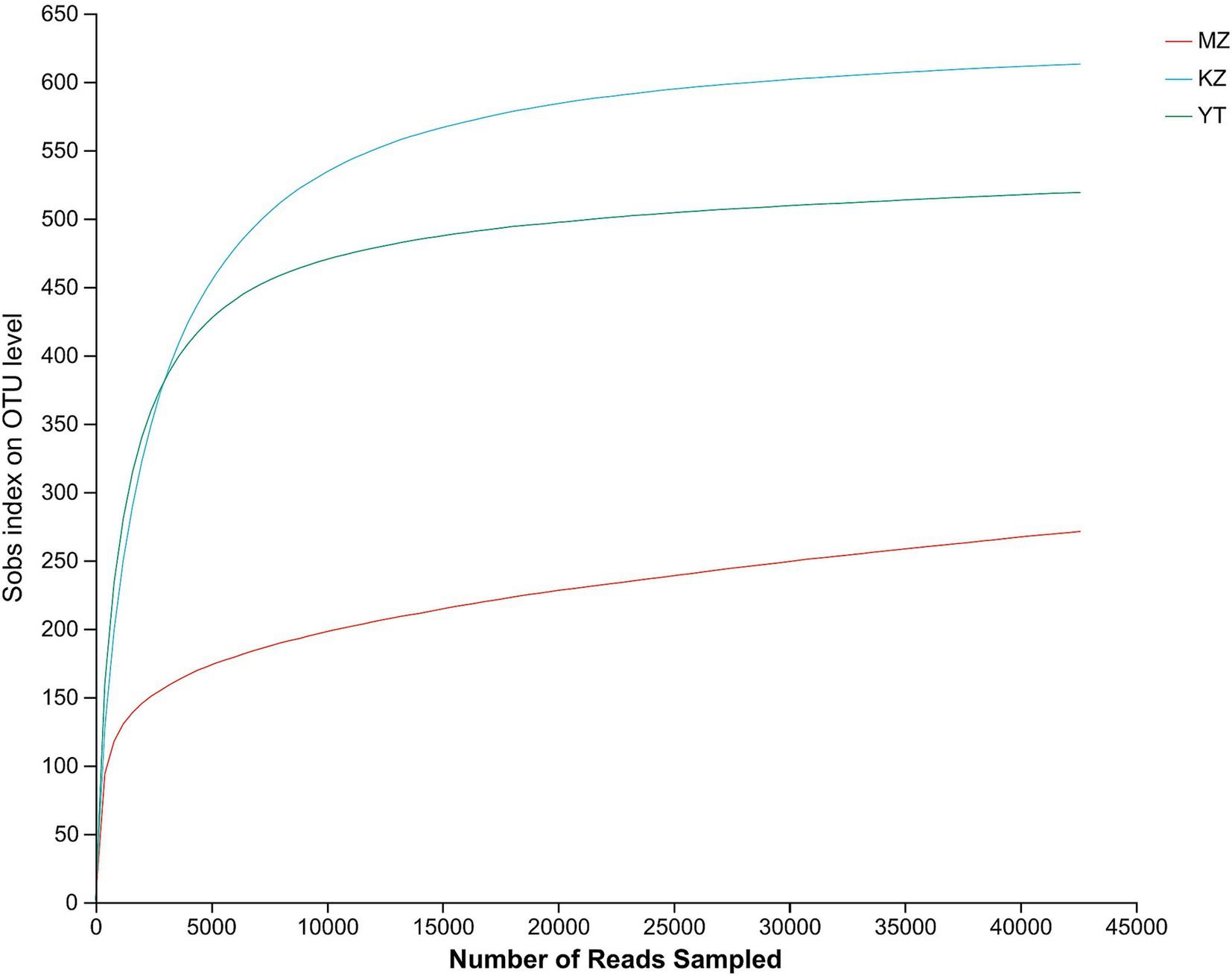
Figure 1. Rarefaction curves of all three samples of bamboo leaves. The abscissa shows the data randomly selected by the Illumina MeSeq™ platform, and the ordinate represents the relative numbers of species. The position of the abscissa of the extension end point of the sample curve represents the number of species in each sample. The flatness of curves indicates a sufficient amount and diversity of bacterial strains, while a steep decline suggests a high proportion of bacterial strains and low bacterial diversity.
In order to illustrate compositional similarity and overlap among OTUs of all three samples, MZ, KZ, and YT, a Venn diagram was constructed (Figure 2).
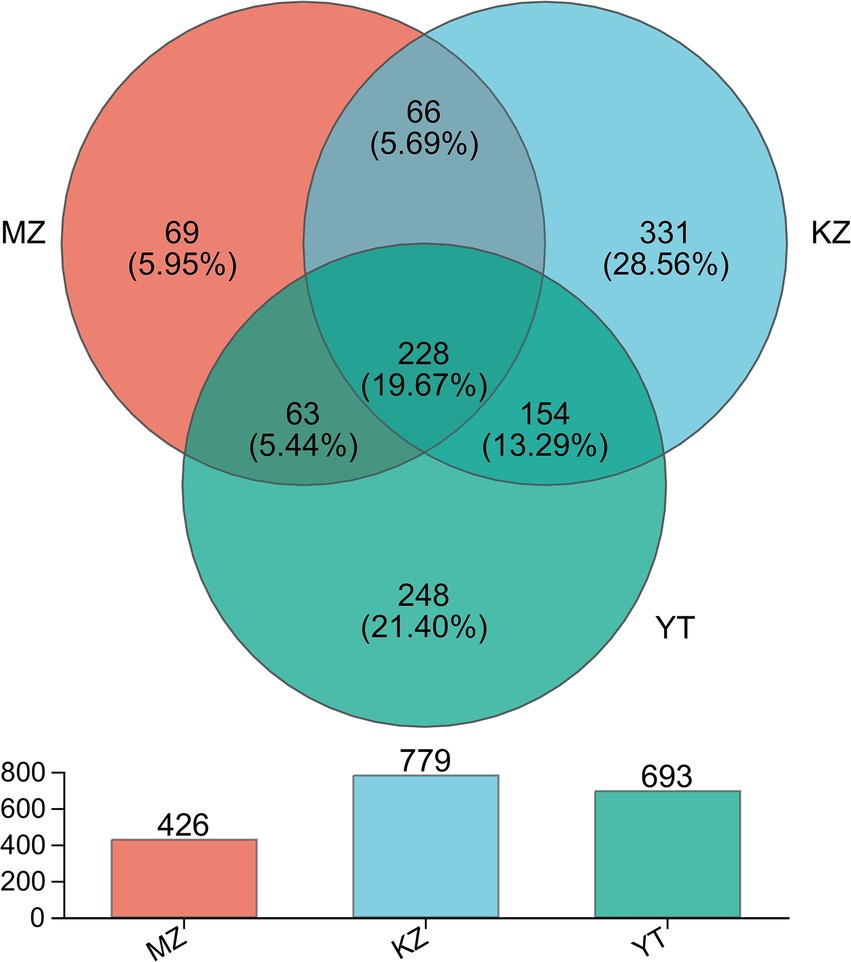
Figure 2. Venn diagram illustrating the unique and common OTUs among all three samples of bamboo leaves. Different colors are assigned to each group. Common and unique OTUs are shown with their numbers. The total number of OTUs in each group, excluding chimeric sequences, is also represented with a bar chart.
In silico analysis, combined with statistical analysis, revealed 1,159 OTUs via clustering of high-fidelity data free of chimeric reads at a 97% similar index. Furthermore, established OTUs were divided into 26 phyla, 64 classes, 158 orders, 270 families, 521 genera, and 811 species. More precisely, established OTUs in each sample, MZ, KZ, and YT, were 426, 779, and 693, respectively (Figure 2). The Venn diagram exhibited 69 (5.95%) unique OTUs in the MZ sample (peach), 331 (28.56%) in the KZ sample (Tiffany blue), and 248 (21.40%) in the YT sample (green). Furthermore, a total of 66 common OTUs between the MZ and KZ samples (teal), 154 common OTUs between the KZ and YT samples (blue-green), 63 common OTUs between the MZ and YT samples (mint) were observed, and a total of 228 (19.7%) common OTUs were observed among all three samples (Victorian teal). These findings proved that community structures of endophytic bacteria in all three samples of bamboo leaves were significantly different (Figure 2).
The abundance and diversity of endophytic bacterial communities were investigated in bamboo leaves. The average sequencing depths of bamboo leaf samples MZ, KZ, and YT were 0.9990, 0.9986, and 0.998, respectively, demonstrating the real-time coverage of endophytic bacterial communities (Table 2).
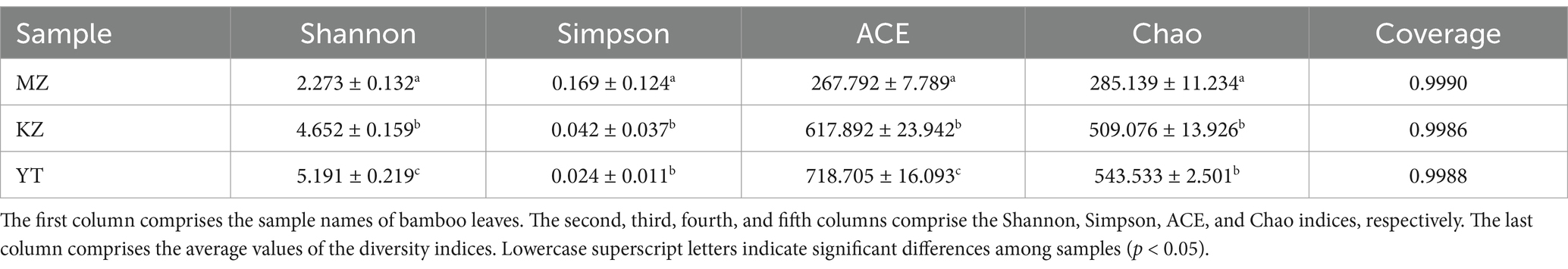
Table 2. Endophytic bacterial community richness and diversity indices in Phyllostachys edulis, Bambusa rigida, and Pleioblastus amarus leaves.
Generally, higher Shannon and lower Simpson indices are indicators of localization of the highest endophytic bacterial diversity index in any biological sample. The highest diversity of the endophytic bacterial community was observed in YT, followed by KZ, and lowest in the MZ group. In order to investigate the richness of communities of bacterial endophytes in bamboo leaf samples, the Chao and ACE indices were also measured. The highest richness of endophytic bacterial communities was observed in YT, followed by KZ, and lowest in the MZ group (Table 2). These results indicate that the richness and diversity of endophytic bacteria in the leaves of all three bamboo species were not adequate.
Relative percentages of community abundance of endophytic bacterium at phylum (Figure 3) and genus levels (Figure 4) have been shown in all three samples of bamboo leaves.
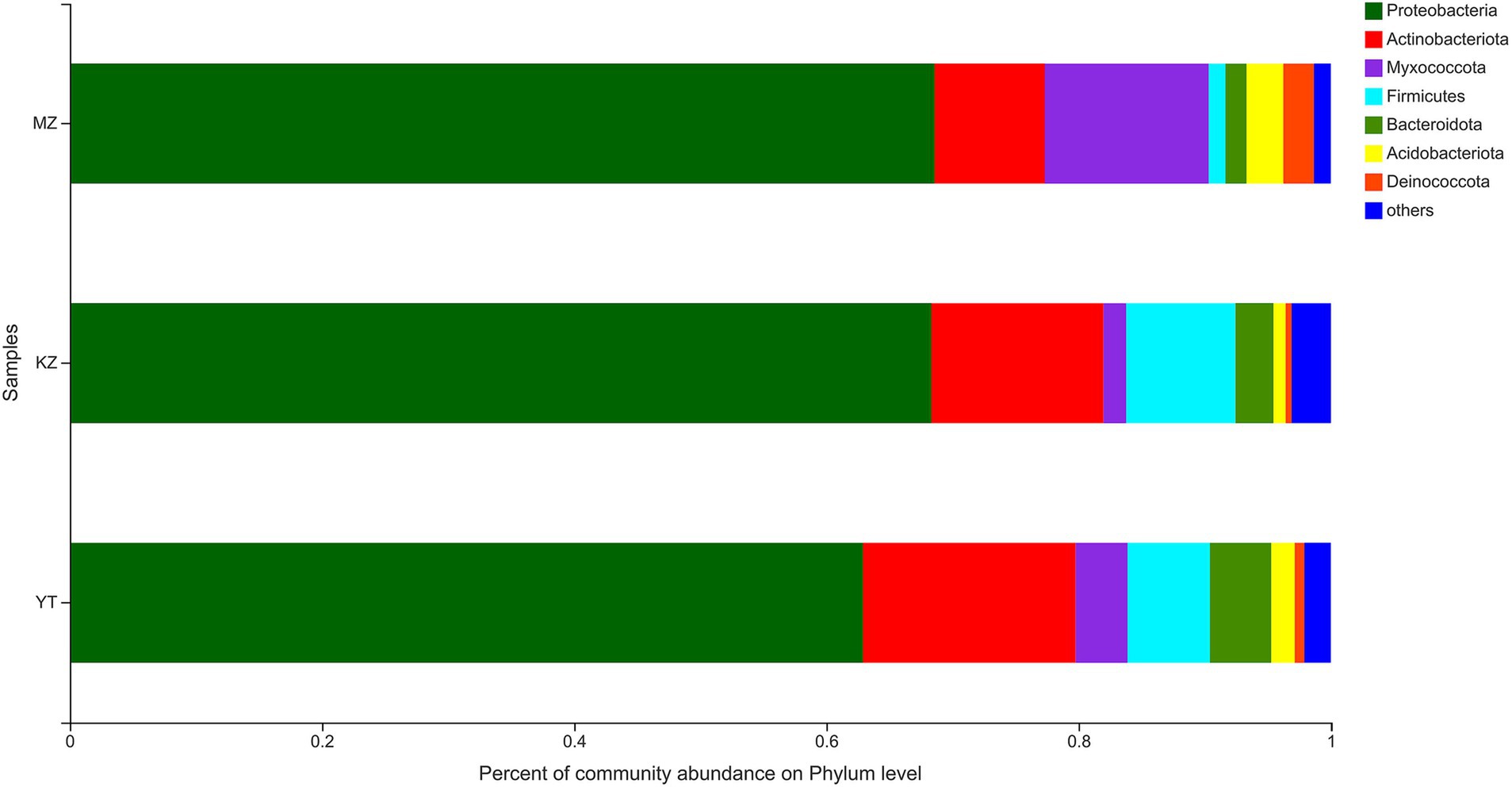
Figure 3. Bar plot analysis of the relative percentage of the endophytic bacterial community at the phylum level. The ordinate represents the names of different samples, and the abscissa represents the relative percentage of different phyla, expressed by columns with different colors and sizes. Phyla with <1% were merged as others.
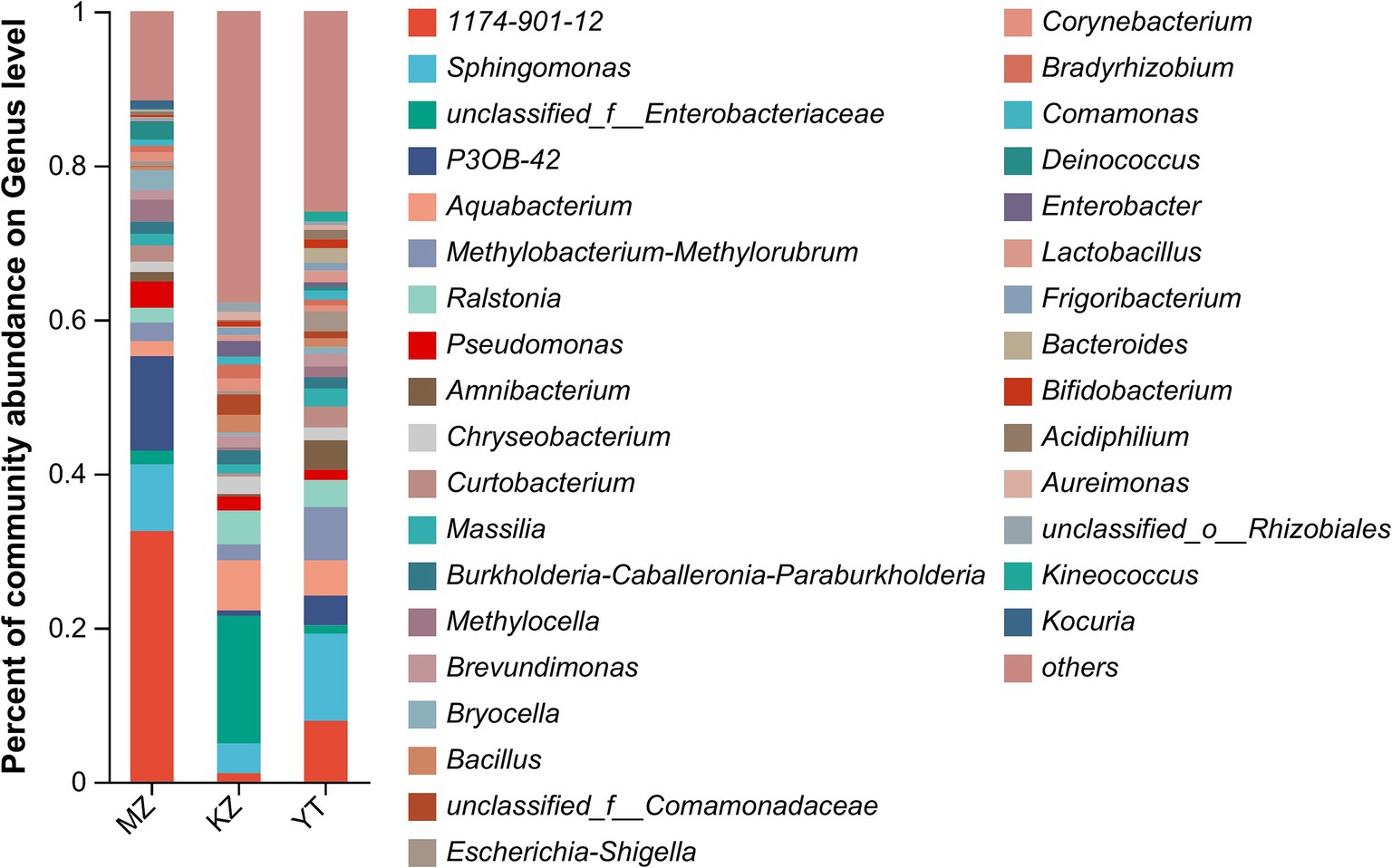
Figure 4. Bar plot analysis of the relative percentage of the endophytic bacterial community at the genus level. The ordinate represents the relative percentage of different genera, and the abscissa represents the names of different samples, expressed by columns with different colors and sizes. Genera with <1% were merged as others.
The highly abundant seven phyla were precisely annotated, while <1% relative abundant phyla were merged into “others.” Dominant phyla include Proteobacteria, Actinobacteria, Myxococcota, Firmicutes, Bacteroidota, Acidobacteriota, and Deinococcota. Notably, the proportion of only three phyla, Proteobacteria, Actinobacteria, and Myxococcota, reached more than 90%. The proportion of Proteobacteria was highly abundant in all three samples of bamboo leaves, with MZ, KZ, and YT accounting for 68.58, 68.30, and 62.88%, respectively. Contrarily, the lowest proportion of the Deinococcota phylum was noticed in all three bamboo leaf samples, with MZ, KZ, and YT accounting for 2.42, 0.48, and 0.78%, respectively. The community abundance percentage of Actinobacteria, Firmicutes, and Bacteroidota in the KZ and YT samples was significantly higher than that in MZ. At the same time, Myxococcota and Acidobacteriota were substantially lower in the KZ and YT samples than in the MZ sample (Figure 3).
Notably, an abundance of endophytic bacterial genera was identified in all three samples of bamboo leaves, and genera with abundances <1% were placed under the “others” category (Figure 4). Furthermore, the proportion of dominant endophytic bacterial taxa in each sample and the distribution ratio of each dominant genus across all three samples of bamboo leaves were visually represented using a circular Circos graph (Figure 5). The dominant genera in the MZ sample were 1,174–901-12 (32.54%), P3OB-42 (12.28%), Sphingomonas (8.66%), and Pseudomonas (3.39%), while those in the KZ sample were unclassified_f_Enterobacteriaceae (16.58%), Aquabacterium (6.51%), Ralstonia (4.38%), and Sphingomonas (3.87%). In contrast, the composition of the YT sample varied, with Sphingomonas (11.29%), 1,174–901-12 (7.89%), Methylobacterium-Methylorubrum (6.89%), and Aquabacterium (4.58%), although their proportions varied among different bamboo species. Additional research is necessary to determine the precise causes for the fluctuating abundances of various genera in all three bamboo species. In summary, we noted that the abundance of different genera of endophytic bacteria varied among the three bamboo species. The LDA value quantifies species’ impact on the difference effect, while LEfSe assesses multi-level differences between species.
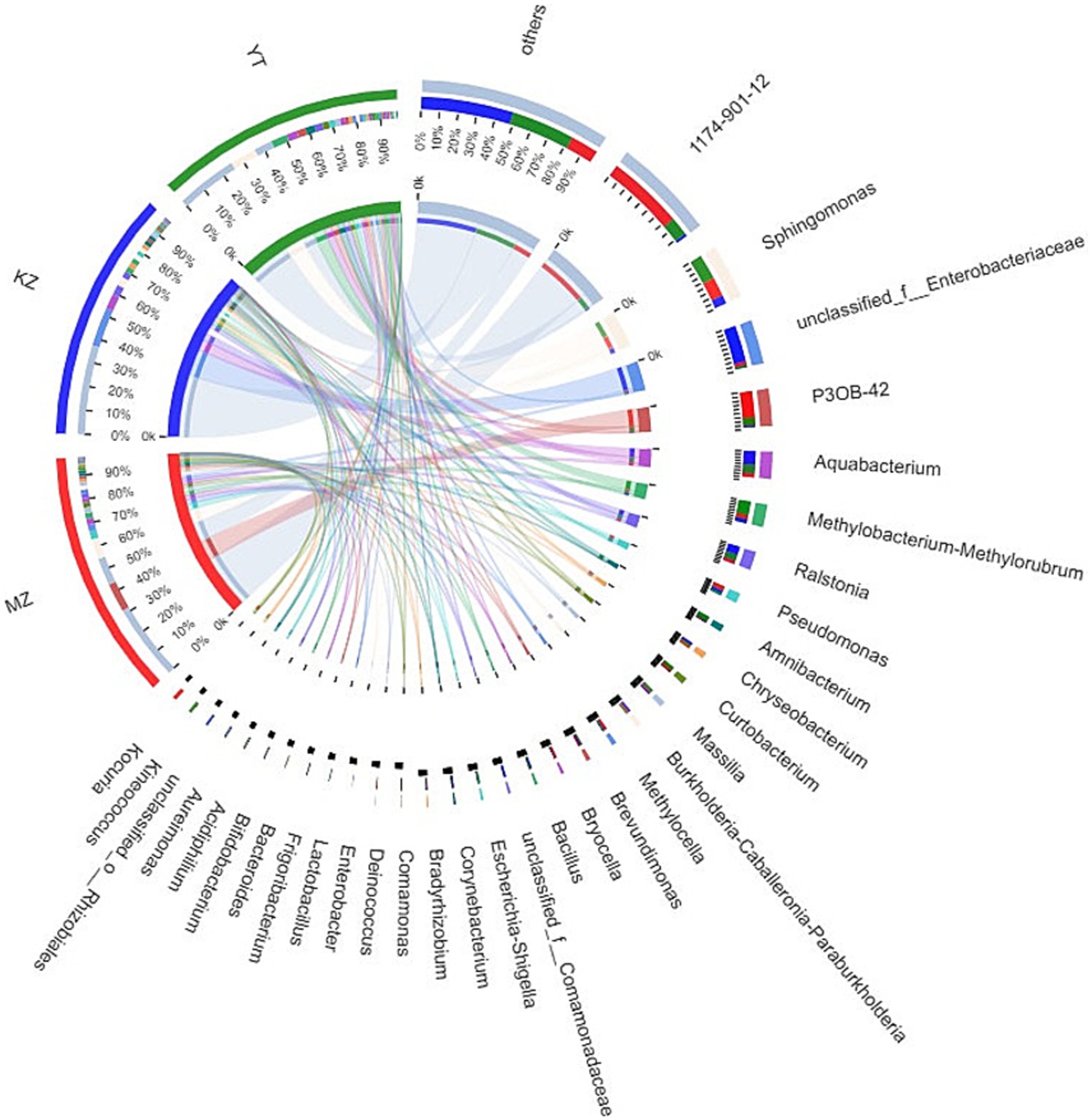
Figure 5. Sample–species relationship-based Circos diagram. The small semicircle (left half circle) represents the composition of species in a sample, the color of the outer ribbon represents different groups, the color of the inner ribbon represents species, and the length represents the relative abundance of the genus in a sample. The large semicircle (right half circle) represents the proportion of genera in different samples.
This indicates that the aforementioned species potentially significantly influence the environmental change process (Huhe et al., 2017). A bar graph was constructed using the genus with the “LDA value >2,” which represented statistically significant differences among the three bamboo endophytic bacteria as determined by LEfSe analysis (Figure 6). Unclassified_Rhizobiales obtained the highest LDA score in the KZ group, followed by norank_Hyphomicrobiaceae. This suggests that the community structures of the three bamboo endophytic bacteria differed significantly.
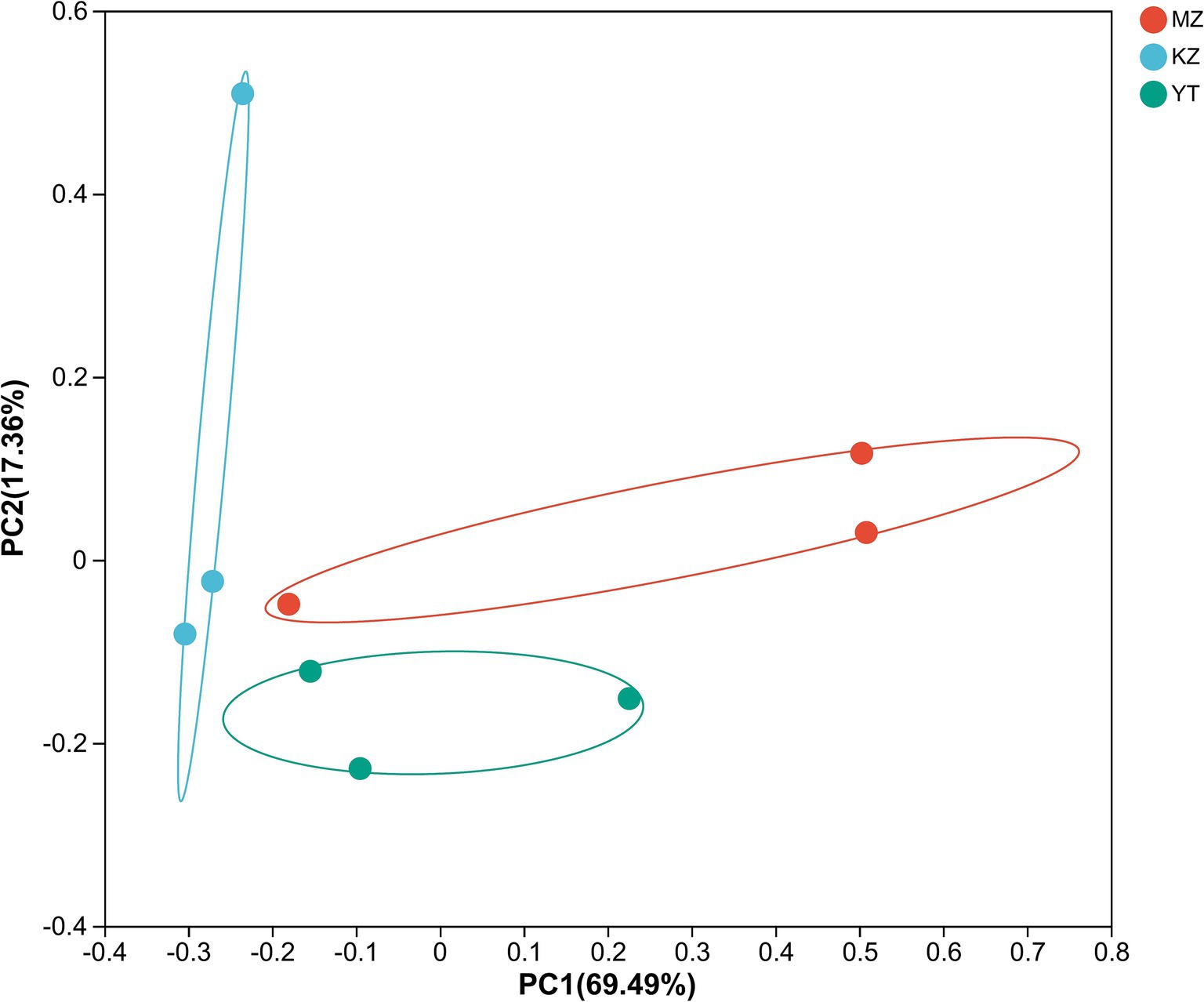
Figure 6. Genus-level discriminant histogram of flora using LDA. The bar graph represents the LDA values for various species. The magnitude of the effect of species abundance on the difference effect increases with the LDA score. In distinct categories, bars of varying colors are used to represent the samples.
The variation index and Euclidean distances among all three samples were calculated and shown using PCoA (Figure 7).
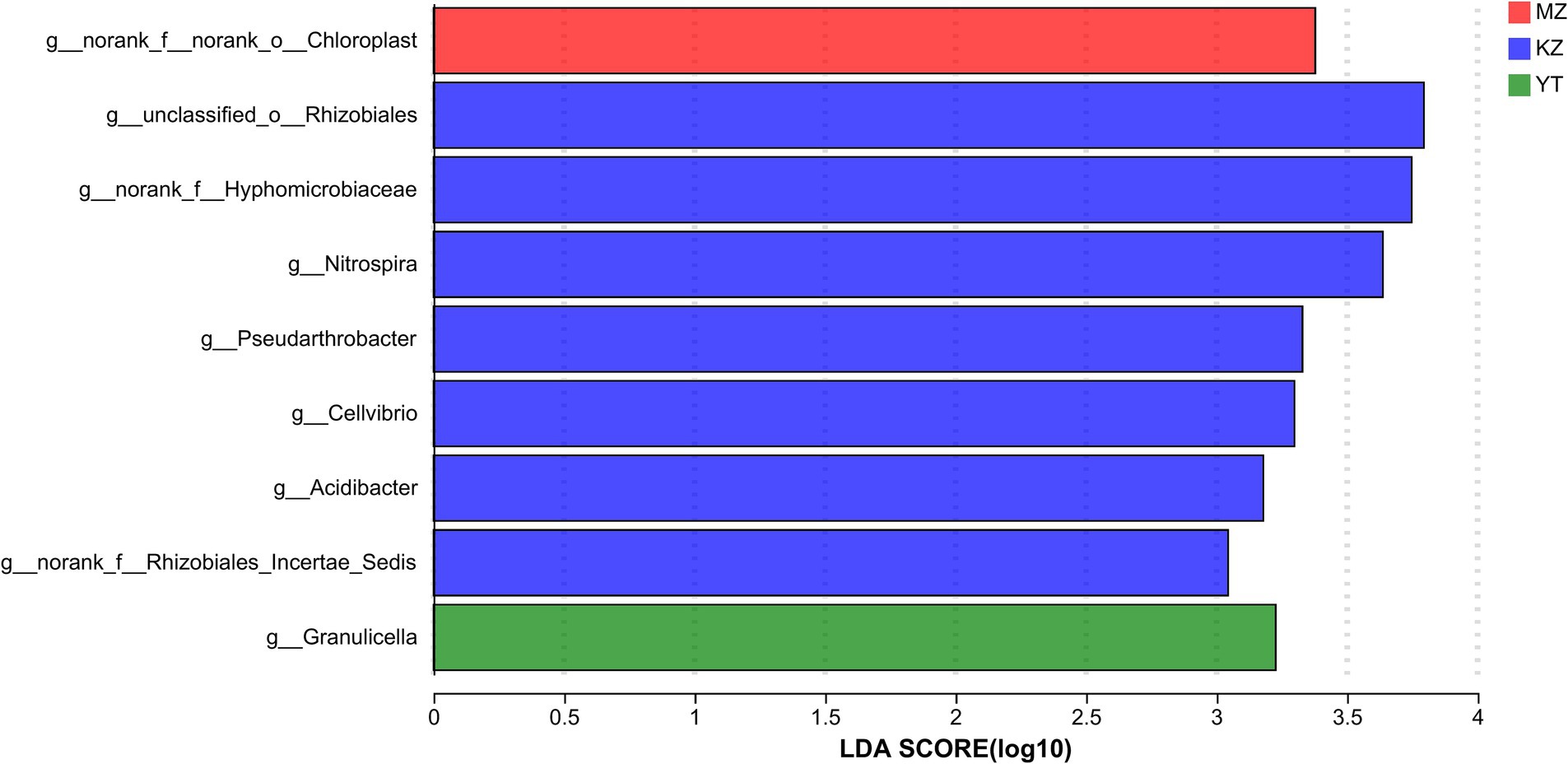
Figure 7. Principal coordinate analysis (PCoA) of multiple samples at the OTU level. The x- and y-axes represent the selected principal component axes, respectively, and the percentage indicates the variation in sample composition by the principal component; the relative distances were denoted on the x- and y-axis scales. In distinct groupings, samples are represented by different color points or shapes. Compositional similarity between bacterial species indicates the proximity of points or structures.
To create PCoA maps, extraordinarily abundant endophytic bacterial species were identified in each sample. PC1 and PC2 contributed 86.85% to PCoA (Figure 7). The distances between all three bamboo samples were remarkably more significant and dispersed, suggesting that the bacterial population structure was highly diverse.
In order to predict functional annotation of Illumina MiSeq™-generated endophytic bacterial 16S amplicons of all three samples of bamboo leaves, PICRUSt software was employed to standardize all OTUs followed by KEGG database, KO, Pathway, and EC information (Table 3).
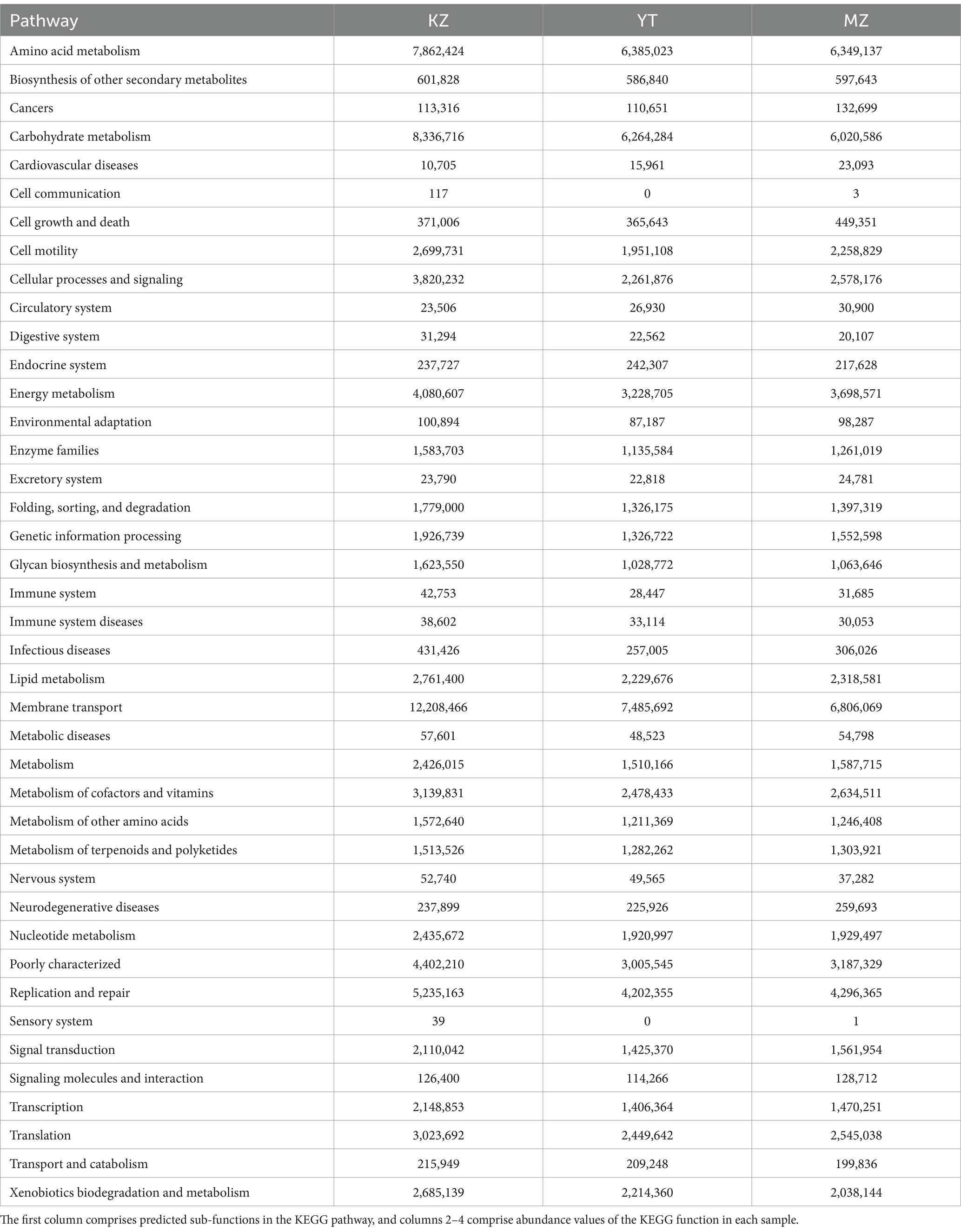
Table 3. Analysis of 16S amplicons using KEGG pathway functions.
In total, 41 sub-functions were annotated by PICRUSt software and KEGG database. Functional annotations of all three samples were similar and predominantly involved in metabolic pathways of cellular processes, environmental information processing, genetic information processing, human diseases, metabolism, and organic systems. Dominant sub-functions were relevant to amino acid metabolism, carbohydrate metabolism, cell motility, cellular processes and signaling, energy metabolism, and membrane transport. Significant variations in sub-functional abundances have been observed among all three samples; that is, carbohydrate metabolism was more abundant in KZ than in YT and MZ. Similarly, cell growth and death, cardiovascular diseases, signaling molecules, and interaction were abundant in MZ compared to KZ and YT (Table 3).
Genetic composition, germination, growth time, growth potential, and localization to a specific region primarily influence endophytic bacterial diversity, variable community structure, and abundance in various bamboo species. This study determined that the diversity of endophytic fungi in the leaves of the three bamboo species differed, among which B. rigida was the most diverse and abundant endophytic bacteria, followed by Pl. amarus and Ph. edulis. This can be seen in the disparities between Populus varieties concerning endophytic microorganisms’ diversity, as reported by Yuan et al. (2014). Comparable occurrences were noted in the diversity of endophytic fungi across various Rhodiola varieties (Cui et al., 2015). High-throughput gene sequencing is a robust and leading technology for precise and quick exploration of the community structure and abundance analysis of endophytic bacteria in a given sample compared to traditional culturing techniques, which are laborious, time-consuming, and even unable to culture numerous endophytes. We observed significant variations in the number of OTUs, the composition of endophytic bacterial communities, and their diversity and richness among different bamboo species. The highest endophytic bacterial community richness was observed in YT and the lowest in MZ, while KZ and YT harbor more OTUs than MZ. These variations could be due to different physiological characteristics and the organizational structure of B. rigida, compared to Ph. edulis and Pl. amarus. Specific tissue structure and more extended localization of B. rigida clumps resulted in a higher accumulation of endophytic bacteria. The tissue structure of B. rigida (YT) is sympodial, and its genetic makeup is also unique as compared to monopodial and mixed bamboo (Gao et al., 2019), which facilitates the invasion and transmission of microbes (Les, 2020). PCoA revealed that the differences in community structure among the three bamboo endophytic bacteria provided further support for the above-stated hypothesis. The distance was close, and the three samples of B. rigida had good repeatability, whereas the Ph. edulis and Pl. amarus samples were poorly repeatable and relatively dispersed.
Numerous endophytic bacteria are localized inside bamboo leaves, and their genes complement specific biological functions (Suryanarayanan et al., 2022). For example, Proteobacteria, Actinobacteria, and Firmicutes significantly tolerate heavy metal stress and are adaptive to acidic soil, while bamboo also grows well in acidic and heavy metal contaminant soil (Chao et al., 2016; Liu et al., 2021). So, there would be a positive correlation between endophytes and bamboo, which further needs to be investigated. Studies have shown that Proteobacteria positively correlate with soil nutrients and nitrogen metabolism, denitrification under anaerobic conditions in a nitrate-rich environment, and nitrogen fixation in a nitrogen-deficient climate (Gao et al., 2019). Additionally, Proteobacteria can habitat water, sediments, soil, and alkaline and saline environments to support bamboo growth in all habitats (Deng et al., 2020). Actinobacteria produce and secrete secondary metabolites to regulate the carbon cycle of other microbes and promote the growth rate of bamboo plants (Jose et al., 2021). Firmicutes produce spores that can adapt to various environments and play a vital role in development of the bamboo ecosystem (Govil et al., 2021). This study determined the abundance of Proteobacteria, Actinobacteria, and Myxococcota in three kinds of bamboo, indicating an interaction between these bamboo species and endophytes. These endophytes can help bamboo adapt to the local acidic soil environment and humid climate and promote the growth of bamboo and the biosynthesis of secondary metabolites.
Many endophytes have been identified in various bamboo species, and bamboo endophytes are extensively distributed within the host. In their study, Liu et al. (2014) isolated 27 isolates of endophytic bacteria from Ph. edulis, the predominant genera of which were Bacillus, Arthrobacter, Staphylococcus, and Ochrobactrum. Bacillus and Pseudomonas were the most prevalent genera of endophytic bacteria in the leaves of six bamboo species, as compared and analyzed by Xu et al. (2014). From Ph. edulis, Yuan et al. (2014) isolated 34 endophytic bacteria strains. The prevailing genera were Arthrobacter, Bacillus, Curtobacterium, Alcaligenes, and Staphylococcus. Forty endophytic bacterial isolates were isolated from Bambusa blumeana by Hou et al. (2019), most of which belonged to the taxa Azospirillum, Aquaspirillum, Escherichia, and Pseudomonas. However, the community structure of endophytic bacteria in this study differs from that reported in previous studies. The most abundant species identified were P3OB-42, Aquabacterium, Methylobacterium-Methylorubrum, Sphingomonas, unclassified_f__Enterobacteriaceae, 1,174–901-12, Sphingomonas, Ralstonia, Pseudomonas, and Bacillus. Sphingomonas harbors particular cell membrane-localized components called glycosphingolipids, which can tolerate nutrient-deficient environments and have strong vitality. Sphingomonas also degrades macromolecular organic pollutants to control environmental pollution (Li et al., 2021). Enterobacteriaceae are harmless and widely distributed in soil, water, plants, insects, and animals (Ito et al., 2019). Pseudomonas is an opportunistic pathogen, and knowledge about its specific role in bamboo leaves is scarce (Williams et al., 2010). Bacillus is another widely distributed endophytic bacterium that can be isolated in many plant species. Members of the Bacillus genus are mostly probiotics, with numerous beneficial effects in the intestine, fecal frequency and characteristics, and skin properties of animals and humans. For example, Bacillus coagulans is a probiotic that can improve the flavor, taste, and shelf life of bamboo-based food (Endres et al., 2011).
In the past, the primary focus of high-throughput sequencing was only to explore the community structure of endophytic bacteria in plants (Ding et al., 2016). In metagenomic research, high-throughput sequencing-based PICRUSt function prediction is highly efficient, convenient, cost-effective, and reliable (Langille et al., 2013). In recent years, functional annotation of various habitats’ microbes has been investigated via PICRUSt (Zhu et al., 2016; Dong et al., 2018). This research predicted key endophytic bacterial species and their roles in biological functions in the leaves of three different bamboo species. The dominant sub-functions were amino acid metabolism, carbohydrate metabolism, cell motility, cellular processes and signaling, energy metabolism, and membrane transport, which are poorly characterized in all three experimental groups. These findings suggest that genetic characteristics, environmental conditions, and community structure are jointly affected by the metabolic functions of endophytes in bamboo. However, due to the limitations of PICRUSt function prediction, combining it with metagenomic sequencing is suggested to understand the microbial community functions of various bamboo species accurately.
Regarding bamboo distribution from the Shunan Bamboo Sea, the three bamboo species have extremely diverse endophytic bacteria, which provides a basis for screening bioactive substances of bamboo endophytic bacteria and developing bamboo resources. The above conclusion may have limitations as microorganisms have a high degree of functional redundancy and cannot accurately predict their functional characteristics. In contrast, functional genes can better predict the response of microbial communities to habitat changes. In the future, more bamboo samples from different regions will be collected for in-depth research using metagenomics methods to reveal the characteristics and diversity of endophytic bacteria in bamboo and their relationship with microbial function.
Bamboo has numerous ecological, commercial, ornamental, and industrial uses and is the fastest-growing forest resource. Endophytic microorganisms coexist symbiotically with an extensive variety of plant species. The composition of endophytic bacterial populations in the leaves of three different bamboo species was investigated using high-throughput sequencing. We determined that bamboo species have a particular impact on the structure and diversity of endophytic bacterial communities. Endophytic bacterial phyla that were exceptionally abundant included Proteobacteria, Actinobacteria, and Myxococcota. The dominant genera of the MZ sample were P3OB-42, Sphingomonas, and Pseudomonas, while the dominant genera of YT were Sphingomonas, 1,174–901-12, and Methylobacterium-Methylorubrum. B. rigida was the most diverse and abundant endophytic bacteria among the three bamboo species, followed by Pl. amarus and Ph. edulis. According to the PICRUSt function prediction analysis, endophytic bacteria in bamboo leaves are predominately enriched in the six biological pathways: human disease, metabolism, cellular processes, environmental information processing, genetic information processing, and organic systems. This indicates that the metabolic functions of endophytic bacteria in bamboo jointly affect bamboo’s genetic characteristics, environmental conditions, and community structure. In conclusion, the most advantageous microorganisms discovered in this study are beneficial, as they can regulate ecological pollution, perform biological control, and enhance food flavor. These results will guide the cultivation and management of bamboo forests, as well as a reference for the development of efficient and beneficial microorganisms with the potential to solve environmental pollution problems caused by climate change.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material.
KY: Writing – original draft, Writing – review & editing. XL: Writing – review & editing. YC: Writing – review & editing. LM: Writing – review & editing. QW: Writing – review & editing. XZ: Writing – review & editing. RH: Writing – review & editing. AA: Writing – review & editing. ME: Writing – review & editing. SS: Writing – review & editing. AE-S: Writing – original draft, Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This research was funded by the Project of Yibin Science and Technology Bureau (Nos. 2022NY010 and 2022NY011), Scientific Research Project of Yibin University, grant number “2022YY07,” and Key Lab of Aromatic Plant Resources Exploitation and Utilization in Sichuan Higher Education(No. 2018XLZ002).
The authors are grateful to and acknowledge the Sichuan Province Government for providing such a well-equipped platform to do research work, the management of Yibin University for their support and for providing us with a pleasant environment of research, the Chinese Government, and Chinese public in particular for their love of Science and Research.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2024.1501057/full#supplementary-material
Calderón, K., Spor, A., Breuil, M.-C., Bru, D., Bizouard, F., Violle, C., et al. (2017). Effectiveness of ecological rescue for altered soil microbial communities and functions. ISME J. 11, 272–283. doi: 10.1038/ismej.2016.86
Chao, Y., Liu, W., Chen, Y., Chen, W., Zhao, L., Ding, Q., et al. (2016). Structure, variation, and co-occurrence of soil microbial communities in abandoned sites of a rare earth elements mine. Environ. Sci. Technol. 50, 11481–11490. doi: 10.1021/acs.est.6b02284
Chen, B., Teh, B. S., Sun, C., Hu, S., Lu, X., Boland, W., et al. (2016). Biodiversity and activity of the gut microbiota across the life history of the insect herbivore Spodoptera littoralis. Sci. Rep. 6:29505. doi: 10.1038/srep29505
Chen, L., Zhang, Y., Chen, X., Fang, R., and Zhang, L. (2018). Identification of endophytic bacteria selectively enriched in Camellia sinensis leaf. Acta Microbiol Sin. 10, 1776–1785. doi: 10.13343/j.cnki.wsxb.20170592
Cui, J. L., Guo, T. T., Ren, Z. X., Zhang, N. S., and Wang, M. L. (2015). Diversity and antioxidant activity of culturable endophytic fungi from alpine plants of rhodiola crenulata, R. Angusta, and R. sachalinensis. PLoS One 10:e0118204. doi: 10.1371/journal.pone.0118204
Cui, J., Guo, S., and Xiao, P. (2017). Interaction between endophytes and host plant and the role of endophytes in genuineness analysis of medicinal plant. Acta Pharm. Sin. 52, 214–221. doi: 10.16438/j.0513-4870.2016-0726
Deng, J., Bai, X., Zhou, Y., Zhu, W., and Yin, Y. (2020). Variations of soil microbial communities accompanied by different vegetation restoration in an open-cut iron mining area. Sci. Total Environ. 704:135243. doi: 10.1016/j.scitotenv.2019.135243
Ding, C., Zheng, Y., Ren, X., and Chen, Z. (2016). Changes in bacterial community composition during the remediation of cd-contaminated soils of bioenergy crops. Acta Sci. Circumst. 36, 3009–3016. doi: 10.13671/j.hjkxxb.2015.0729
Dong, Z., Hong, M., Hu, H., Wang, Y., Zhang, D., and Wang, K. (2018). Effect of excess nitrogen loading on the metabolic potential of the bacterial community in oligotrophic coastal water. Acta Sci. Circumst. 38, 457–466. doi: 10.13671/j.hjkxxb.2017.0405
Donoso, R., Leiva-Novoa, P., Zúñiga, A., Timmermann, T., Recabarren-Gajardo, G., and González, B. (2017). Biochemical and genetic bases of indole-3-acetic acid (auxin phytohormone) degradation by the plant-growth-promoting rhizobacterium Paraburkholderia phytofirmans PsJN. Appl. Environ. Microbiol. 83, e01991–e01916. doi: 10.1128/AEM.01991-16
Edgar, R. C., Haas, B. J., Clemente, J. C., Quince, C., and Knight, R. (2011). UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27, 2194–2200. doi: 10.1093/bioinformatics/btr381
El-Sappah, A. H., Elrys, A. S., Desoky, E.-S. M., Zhao, X., Bingwen, W., El-Sappah, H. H., et al. (2021). Comprehensive genome wide identification and expression analysis of MTP gene family in tomato (Solanum lycopersicum) under multiple heavy metal stress. Saudi J. Biol. Sci. 28, 6946–6956. doi: 10.1016/j.sjbs.2021.07.073
El-Sappah, A. H., Shawky, A., Sayed-Ahmad, M. S., and Youssef, M. (2017). Estimation of heat shock protein 70 (hsp 70) gene expression in Nile tilapia (Oreochromis niloticus) using quantitative real-time PCR. Zagazig J. Agric. Res. 44, 1003–1015. doi: 10.21608/zjar.2017.52300
Endres, J., Qureshi, I., Farber, T., Hauswirth, J., Hirka, G., Pasics, I., et al. (2011). One-year chronic oral toxicity with combined reproduction toxicity study of a novel probiotic, Bacillus coagulans, as a food ingredient. Food Chem. Toxicol. 49, 1174–1182. doi: 10.1016/j.fct.2011.02.012
Fouts, D. E., Brinkac, L., Beck, E., Inman, J., and Sutton, G. (2012). PanOCT: automated clustering of orthologs using conserved gene neighborhood for pan-genomic analysis of bacterial strains and closely related species. Nucleic Acids Res. 40:e172. doi: 10.1093/nar/gks757
Gao, J., Feng, G., Yan, S., Qin, H., Chen, J., Liang, C., et al. (2019). Comparison on structure and diversity of endophytic bacteria between three species of bamboo. J. Bamboo Res. 38, 39–46. doi: 10.19560/j.cnki.issn1000-6567.2019.03.006
Gihring, T. M., Green, S. J., and Schadt, C. W. (2012). Massively parallel rRNA gene sequencing exacerbates the potential for biased community diversity comparisons due to variable library sizes. Environ. Microbiol. 14, 285–290. doi: 10.1111/j.1462-2920.2011.02550.x
Gond, S. K., Bergen, M. S., Torres, M. S., and White, J. F. Jr. (2015). Endophytic Bacillus spp. produce antifungal lipopeptides and induce host defence gene expression in maize. Microbiol. Res. 172, 79–87. doi: 10.1016/j.micres.2014.11.004
Govil, T., Paste, M., Samanta, D., David, A., Goh, K. M., Li, X., et al. (2021). Metagenomics and culture dependent insights into the distribution of Firmicutes across two different sample types located in the black hills region of South Dakota, USA. Microorganisms 9:113. doi: 10.3390/microorganisms9010113
Hardoim, P. R., van, L., and Elsas, J. (2008). Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 16, 463–471. doi: 10.1016/j.tim.2008.07.008
He, Q., Song, Q., Zhang, S., Zhang, W., and Wang, H. (2018). Simultaneous nitrification, denitrification and phosphorus removal in an aerobic granular sequencing batch reactor with mixed carbon sources: reactor performance, extracellular polymeric substances and microbial successions. Chem. Eng. J. 331, 841–849. doi: 10.1016/j.cej.2017.09.060
Hou, Z., Yin, J., Lu, Y., Song, J., Wang, S., Wei, S., et al. (2019). Transcriptomic analysis reveals the temporal and spatial changes in physiological process and gene expression in common buckwheat (Fagopyrum esculentum Moench) grown under drought stress. Agronomy 9:569. doi: 10.3390/agronomy9100569
Huhe, C., Chen, X., Hou, F., Wu, Y., and Cheng, Y. (2017). Bacterial and fungal community structures in loess plateau grasslands with different grazing intensities. Front. Microbiol. 8:606. doi: 10.3389/fmicb.2017.00606
Ito, K., Honda, H., Yoshida, M., Aoki, K., Ishii, Y., Miyokawa, S., et al. (2019). A metallo-beta-lactamase producing Enterobacteriaceae outbreak from a contaminated tea dispenser at a children's hospital in Japan. Infect. Control Hosp. Epidemiol. 40, 217–220. doi: 10.1017/ice.2018.331
Jose, P. A., Maharshi, A., and Jha, B. (2021). Actinobacteria in natural products research: Progress and prospects. Microbiol. Res. 246:126708. doi: 10.1016/j.micres.2021.126708
Langille, M. G., Zaneveld, J., Caporaso, J. G., McDonald, D., Knights, D., Reyes, J. A., et al. (2013). Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 31, 814–821. doi: 10.1038/nbt.2676
Li, Z., Chen, C., Xie, H., Yao, Y., Zhang, X., Brozena, A., et al. (2022). Sustainable high-strength macrofibres extracted from natural bamboo. Nat. Sustain. 5, 235–244. doi: 10.1038/s41893-021-00831-2
Li, Q., Li, J., Jiang, L., Sun, Y., Luo, C., and Zhang, G. (2021). Diversity and structure of phenanthrene degrading bacterial communities associated with fungal bioremediation in petroleum contaminated soil. J. Hazard. Mater. 403:123895. doi: 10.1016/j.jhazmat.2020.123895
Liu, J., Liu, W., Zhang, Y., Chen, C., Wu, W., and Zhang, T. C. (2021). Microbial communities in rare earth mining soil after in-situ leaching mining. Sci. Total Environ. 755:142521. doi: 10.1016/j.scitotenv.2020.142521
Liu, F., Yuan, Z. S., Zhang, G. F., and Wei, C. (2014). Diversity of endophytic bacterial isolated from Moso bamboo. Fujian J. Agric. Sci. 29, 1236–1239.
Liu, F., Yuan, Z., Zhang, X., Zhang, G., and Xie, B. (2017). Characteristics and diversity of endophytic bacteria in moso bamboo (Phyllostachys edulis) based on 16S rDNA sequencing. Arch. Microbiol. 199, 1259–1266. doi: 10.1007/s00203-017-1397-7
Lu, Y., Chen, J., Zheng, J., Hu, G., Wang, J., Huang, C., et al. (2016). Mucosal adherent bacterial dysbiosis in patients with colorectal adenomas. Sci. Rep. 6:26337. doi: 10.1038/srep26337
Massimo, N. C., Nandi Devan, M., Arendt, K. R., Wilch, M. H., Riddle, J. M., Furr, S. H., et al. (2015). Fungal endophytes in aboveground tissues of desert plants: infrequent in culture, but highly diverse and distinctive symbionts. Microb. Ecol. 70, 61–76. doi: 10.1007/s00248-014-0563-6
Rogers, M. B., Firek, B., Shi, M., Yeh, A., Brower-Sinning, R., Aveson, V., et al. (2016). Disruption of the microbiota across multiple body sites in critically ill children. Microbiome 4, 1–10. doi: 10.1186/s40168-016-0211-0
Shen, X.-Y., Cheng, Y.-L., Cai, C.-J., Fan, L., Gao, J., and Hou, C.-L. (2014). Diversity and antimicrobial activity of culturable endophytic fungi isolated from moso bamboo seeds. PLoS One 9:e95838. doi: 10.1371/journal.pone.0095838
Sui, Q., Liu, C., Zhang, J., Dong, H., Zhu, Z., and Wang, Y. (2016). Response of nitrite accumulation and microbial community to free ammonia and dissolved oxygen treatment of high ammonium wastewater. Appl. Microbiol. Biotechnol. 100, 4177–4187. doi: 10.1007/s00253-015-7183-z
Suryanarayanan, T., Ayesha, M., and Shaanker, R. U. (2022). Leaf photosynthesis: do endophytes have a say? Trends Plant Sci. 27, 968–970. doi: 10.1016/j.tplants.2022.07.009
Tang, R., Wei, Y., Li, Y., Chen, W., Chen, H., Wang, Q., et al. (2018). Gut microbial profile is altered in primary biliary cholangitis and partially restored after UDCA therapy. Gut 67, 534–541. doi: 10.1136/gutjnl-2016-313332
Wang, J., Abbas, M., Wen, Y., Niu, D., Wang, L., Sun, Y., et al. (2018). Selection and validation of reference genes for quantitative gene expression analyses in black locust (Robinia pseudoacacia L.) using real-time quantitative PCR. PLoS One 13:e0193076. doi: 10.1371/journal.pone.0193076
Wang, X., Zhang, Q., and Yao, H. (2012). Soil carbon, nitrogen and microbiological characteristics during bamboo highspeed growth. Shengtai Xuebao 32, 1412–1418. doi: 10.5846/stxb201101110047
Wen, J.-L., Xiao, L.-P., Sun, Y.-C., Sun, S.-N., Xu, F., Sun, R.-C., et al. (2011). Comparative study of alkali-soluble hemicelluloses isolated from bamboo (Bambusa rigida). Carbohydr. Res. 346, 111–120. doi: 10.1016/j.carres.2010.10.006
White, J. F., Chen, Q., Torres, M. S., Mattera, R., Irizarry, I., Tadych, M., et al. (2015). Collaboration between grass seedlings and rhizobacteria to scavenge organic nitrogen in soils. AoB Plants 7:lu093. doi: 10.1093/aobpla/plu093
Williams, K. P., Gillespie, J. J., Sobral, B. W., Nordberg, E. K., Snyder, E. E., Shallom, J. M., et al. (2010). Phylogeny of gammaproteobacteria. J. Bacteriol. 192, 2305–2314. doi: 10.1128/JB.01480-09
Xu, X., Bai, Y., Liu, H., Yang, C. L., and Ling, Y. G. (2014). Microflora analysis on six kinds of bamboo leaves in Sichuan Province. J. Huazhong Agric. Univ. 33, 53–59. doi: 10.13300/j.cnki.hnlkxb.2014.04.010
Xu, X., Du, H., Zhou, G., Ge, H., Shi, Y., Zhou, Y., et al. (2011). Estimation of aboveground carbon stock of Moso bamboo (Phyllostachys heterocycla var. pubescens) forest with a Landsat thematic mapper image. Int. J. Remote Sens. 32, 1431–1448. doi: 10.1080/01431160903551389
Yan, K., Abbas, M., Meng, L., Cai, H., Peng, Z., Li, Q., et al. (2021a). Analysis of the fungal diversity and community structure in Sichuan dark tea during pile-fermentation. Front. Microbiol. 12:706714. doi: 10.3389/fmicb.2021.706714
Yan, K., Pei, Z., Meng, L., Zheng, Y., Wang, L., Feng, R., et al. (2022). Determination of community structure and diversity of seed-vectored endophytic fungi in Alpinia zerumbet. Front. Microbiol. 13:814864. doi: 10.3389/fmicb.2022.814864
Yan, K., Yan, L., Meng, L., Cai, H., Duan, A., Wang, L., et al. (2021b). Comprehensive analysis of bacterial community structure and diversity in Sichuan dark tea (Camellia sinensis). Front. Microbiol. 12:735618. doi: 10.3389/fmicb.2021.735618
Ye, J., Joseph, S. D., Ji, M., Nielsen, S., Mitchell, D. R., Donne, S., et al. (2017). Chemolithotrophic processes in the bacterial communities on the surface of mineral-enriched biochars. ISME J. 11, 1087–1101. doi: 10.1038/ismej.2016.187
Ying, Y., Chunbao, L., Shuanglin, C., and Ziwu, G. (2022). Effect of habitats on the morphological and structural characteristic of rhizome roots of Pleioblastus amarus and its allometric growth. Acta Bot. Boreal. Occident. Sin. 42, 1583–1590.
Yuan, Z., Liu, F., and Zhang, G. (2014). Characteristics of endophytic bacteria isolated from mosobamboo rhizome and their 16S rDNA diversity. Genom. Appl. Biol. 33, 1019–1024.
Zhang, X., Zhong, Z., Gai, X., Du, X., Bian, F., Yang, C., et al. (2019). Changes of root endophytic bacterial community along a chronosequence of intensively managed lei bamboo (Phyllostachys praecox) forests in subtropical China. Microorganisms 7:616. doi: 10.3390/microorganisms7120616
Zhou, Y. K., Shen, X. Y., and Hou, C. L. (2017). Diversity and antimicrobial activity of culturable fungi from fishscale bamboo (Phyllostachys heteroclada) in China. World J. Microbiol. Biotechnol. 33, 104–107. doi: 10.1007/s11274-017-2267-9
Keywords: bamboo, Phyllostachys edulis, Bambusa rigida, Pleioblastus amarus, endophytic bacteria, 16 s rDNA sequencing
Citation: Yan K, Li X, Cai Y, Meng L, Wei Q, Zhao X, M. Y. Heakel R, M. Atif A, A. Abd Elhamid M, A. Soaud S and H. El-Sappah A (2025) High-throughput sequencing explores the genetic variability of endophytic bacteria in three Sichuan bamboo species (Phyllostachys edulis, Bambusa rigida, and Pleioblastus amarus). Front. Microbiol. 15:1501057. doi: 10.3389/fmicb.2024.1501057
Edited by:
Gustavo Santoyo, Universidad Michoacana de San Nicolás de Hidalgo, MexicoReviewed by:
Vishal Srivastava, Cleveland Clinic, United StatesCopyright © 2025 Yan, Li, Cai, Meng, Wei, Zhao, M. Y. Heakel, M. Atif, A. Abd Elhamid, A. Soaud and H. El-Sappah. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ahmed H. El-Sappah, ahmed_elsappah2006@yahoo.com; Kuan Yan, kuanyan-029@hotmail.com
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.