
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 31 May 2016
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 7 - 2016 | https://doi.org/10.3389/fmicb.2016.00817
This article is part of the Research TopicNew frontiers in the search of antimicrobials agents from natural productsView all 54 articles
 Ahmed K. Al Atya1Yanath Belguesmia1Gabrielle Chataigne1Rozenn Ravallec1Anne Vachée2Sabine Szunerits3
Ahmed K. Al Atya1Yanath Belguesmia1Gabrielle Chataigne1Rozenn Ravallec1Anne Vachée2Sabine Szunerits3 Rabah Boukherroub3
Rabah Boukherroub3 Djamel Drider1*
Djamel Drider1*Methicillin-resistant Staphylococcus aureus (MRSA) has become a worrisome superbug. This work aimed at studying the effects of two class IIb bacteriocins, enterocins DD28 and DD93 as anti-MRSA agents. Thus, these bacteriocins were purified, from the cultures supernatants of Enterococcus faecalis 28 and 93, using a simplified purification procedure consisting in a cation exchange chromatography and a reversed-phase high-performance liquid chromatography. The anti-Staphylococcal activity was shown in vitro by the assessment of the minimal inhibitory concentration (MIC), followed by a checkerboard and time-kill kinetics experiments. The data unveiled a clear synergistic effect of enterocins DD28 and DD93 in combination with erythromycin or kanamycin against the clinical MRSA-S1 strain. Besides, these combinations impeded as well the MRSA-S1 clinical strain to setup biofilms on stainless steel and glace devices.
Staphylococcus aureus is among the five top pathogens found as normal resident of the skin and nasal flora in at least 25–30% of healthy humans, and it is associated with hospital acquired (HA-MRSA) and community acquired (CA-MRSA) infections ranging from superficial wound infections to life-threatening deep infections such as septicemia, endocarditis, and toxic shock syndrome (David and Daum, 2010). Antibiotic resistance and biofilm-forming capabilities contribute to the success of S. aureus as a harsh human pathogen in the healthcare as well as in the community settings. The last decade has seen a welcome increase in the number of agents available for the treatment of MRSA including antibiotics such as fluoroquinolones, linezolid, rifampin, and antimicrobial peptides (AMPs) such as daptomycin, tigecycline, and mainly vancomycin. Resistance to methicillin was observed in 1961, 1 year after the commercial availability of this antibiotic. Susceptibility to vancomycin was first reported in 1996 in Japan, leading to emergence of heterogeneous resistance to vancomycin phenotype (Spagnolo et al., 2014). MRSA with reduced susceptibility to vancomycin was reported in ocular infections, and there was a rise in S. aureus resistance to new and old generation fluoroquinolones that were commonly used for prophylaxis after intravitreal injections and intraocular surgeries (Sadaka et al., 2015). Daptomycin which is considered drug of last resort after vancomycin breakdown for the treatment of MRSA infections (Claeys et al., 2015) has shown non-inferiority to vancomycin in the treatment of MRSA bacteremia (Holmes et al., 2015) was threatened because of the emergence of daptomycin resistance, especially in the deep-seated infections (Claeys et al., 2015). MRSA are responsible of diverse infections, especially in the healthcare structures. The increasing resistance of Gram-positive bacteria to the broad-spectrum antibiotics and the lack of new molecules expected to become available in the near future, advocates the need of novel anti-MRSA agents and therapeutic options (Tängdén, 2014).
Antimicrobial peptides were largely admitted as potential alternatives to traditional antibiotics in order to combat the scaring and increasing bacterial infections. AMPs are produced by all the living cells but also gathered by chemical synthesis and controlled enzymatic digestion of proteins. Bacteria are known as great sources of AMPs such as lipopeptides and bacteriocins. Conversely to lipopeptides, the bacteriocins are AMPs of proteinaceous nature, ribosomally synthesized mainly by lactic acid bacteria (LAB) (Drider and Rebuffat, 2011). LAB of Enterococcus genus produce a great number of bacteriocins designed as enterocins. Enterocins-producing strains were isolated from a wide range of sources, including fermented food, environmental, and clinical (Ishibashi et al., 2012). Enterocins resulted to be mainly produced by Enterococcus faecalis and E. faecium species (Goto and Yan, 2011). Enterocins produced by E. muntii, E. avium, E. durans, and E. hirae strains were also reported in the literature (Saavedra et al., 2004; Batdorj et al., 2006; Sánchez et al., 2007; Birri et al., 2010). Multiple enterocins-producing strains were characterized for their large range of activities, inhibiting the growth of many undesirable bacteria (Ishibashi et al., 2012). Cintas et al. (2000) underlined the potential of E. faecium L50 to produce three different enterocins named enterocins L50A and L50B, enterocin P, and enterocin Q which act synergistically and inhibit the growth of many Gram-positive bacteria. Remarkably, enterocins were also produced by enterococci from the gastrointestinal tract origins of humans, animals, human infection sites and healthy babies feces (Tomita et al., 1996; Kang and Lee, 2005; Sawa et al., 2012) A compilation of studies underpinning the inhibitory activities of enterocins, pointed out the capabilities of enterocin E-760 to inhibit the growth of Salmonella enterica, Escherichia coli, Pseudomonas aeruginosa, Campylobacter jejuni, and Staphylococcus aureus (Line et al., 2008). These data show that bacteriocins are sustainable antimicrobials that could be used alone or in combination with antibiotics.
Related to that, the combinations of antibiotics were presented as a promising option in the management of antibiotics use and treatment of life-threatening infections (Bassetti and Righi, 2015; Leone et al., 2015). The antibiotics were successfully combined to chemical or physical agents (Sawa et al., 2012; Kullar et al., 2015). In direct line, the combinations of antibiotics and bacteriocins could offer novel therapeutic options as supported by different and successful in vitro studies (Naghmouchi et al., 2010, 2011, 2012, 2013; Hassan et al., 2012; Mataraci and Dosler, 2012; Wolska et al., 2012; Singh et al., 2014).
In this study, enterocins DD28 and DD93 produced by E. faecalis 28 and E. faecalis 93 were purified and their DNA sequences were determined. The anti-MRSA activity was determined against the MRSA-S1 clinical strain. In spite of their relatively high MICs values against the aforementioned target, enterocins DD28 and DD93 were able to synergize with kanamycin or erythromycin and switch from resistance to susceptibility the phenotype of MRSA-S1 strain. Furthermore, these combinations hampered biofilm formation of the MRSA-S1 strain on AISI 304L stainless steel and glass devices. Overall, the combinations of antibiotics and bacteriocins offer a novel strategy to fight against pathogenic bacteria.
Enterococcus faecalis DD28 and E. faecalis DD93 recently isolated from meconium (Al-Atya et al., 2015) were grown in MRS (de Man, Rogosa, and Sharpe) medium (De Man et al., 1960) at 37°C for 16 to 18 h. S. epidermidis (Kindly provided by Dr. Anne Vachée, Roubaix hospital, France), S. aureus ATCC 33862, and Methicillin Resistant S. aureus (MRSA) strains including MRSA ATCC 43300, MRSA-S1, and MRSA-2 strains (kindly provided by Dr. Gilles Prévost, Strasbourg University, France) were grown in Brain Heart Infusion (BHI) broth at 37°C for appropriate experimental time. Listeria innocua CIP80.11 was grown at 37°C for overnight in BHI or Muller Hinton media according to the experiment purposes.
Antibiotic susceptibility was performed by VITEK 2 system (Bio-Mérieux, France), encompassing nearly all important antibiotics. Antibiotic susceptibility and MICs were determined and analyzed according to the French Committee on Antimicrobial Susceptibility Testing (French Committee on Antimicrobial Susceptibility Testing [FCAST], 2015).
Purification of enterocins DD28 and DD93 was performed at room temperature using two-step methods adapted from Abriouel et al. (2003). Briefly 1,000 ml of the cell-free culture supernatant of E. faecalis 28 and E. faecalis 93 were adjusted to pH 6.3 using 5 M NaOH. Then Carboxymethyl Sephadex CM-25 (GE Healthcare, Sweden) gel slurry equilibrated in distilled water was added to culture supernatants (1:40, v/v). The obtained mixture was held under stirring for 30 min and decanted for additional 30 min. The supernatants were removed, and the sediment gel slurry containing the bacteriocin activity, captured by cation interactions with the matrix was loaded onto a 10 cm × 50 cm glass column. The gels was washed with one volume (1V) of distilled water and 2V of 0.5 M NaCl, followed by 2V of 1.5 M NaCl to elute the adsorbed bacteriocin. The obtained fractions were filtrated through 0.22 μm pore size low protein binding filters (Millex GV; Millipore Corp., Belford, MA, USA) and tested for bacteriocin activity.
The active fractions from the cation exchange chromato-graphy were applied onto a reversed-phase high-performance liquid chromatography (RP-HPLC) by using a column C-18 (5 μm, 250 mm × 3 mm, VYDAC 218 TP53; Grace, Deerfield, IL, USA) previously equilibrated in solvent A (10 mM trifluoroacetic acid, TFA; Fluka), at a flow rate of 5 ml/min. Non-adsorbed material was eliminated by washing the column with solvent A until the UV absorbance of the effluent at 210 nm reached baseline. The material retained in the column was eluted with a gradient ranging from 0 to 40% of solvent B [isopropyl alcohol/acetonitrile (2:1, v/v) in 40 mM TFA] in over 5 min, then followed by increasing of solvent B gradient from 40 to 100% for over 25 min at a flow rate of 2.5 ml/min. Fractions of the column effluent were collected according to their UV light absorbance, lyophilized and dissolved again in distilled water before being tested for bacteriocin activity.
Antimicrobial activity was tested along the purification procedure on the cell-free supernatant (CFS), partly purified peptide (upon cation exchange step) and purified peptide (upon Reversed Phase-HPLC step). Arbitrary units (AU) were calculated based on the spot method (Naghmouchi et al., 2011). Thus, the resulting sample was serially diluted twofold with filter-sterilized phosphate buffer. Ten micro liter of each diluted sample was spotted onto the plate of BHI medium containing L. innocua CIP80.11 as indictor strain. The plates were then incubated at 37oC overnight, and the titer was defined as the reciprocal of the highest dilution (2n) that resulted in inhibition of the indicator lawn. The AU of antibacterial activity per milliliter was defined as 2n × 1,000 μl/10μl. The protein concentration was determined by BCA assay using Sigma–Aldrich BCA kit (USA).
Purified enterocins were analyzed with an Ultraflex MALDI-ToF/ToF mass spectrometer (Bruker, Bremen, Germany) equipped with a smart beam laser. Samples were analyzed using an accelerating voltage of 25 kV and matrix suppression in deflexion mode at m/z 1000. The laser power was set to just above the threshold of ionization (around 60%). Spectra were acquired in reflector positive mode in the range of 3 000 at 10 000 Da. Each spectrum was the result of 1 000 laser shots per m/z segment per sample delivered in 10 sets of 50 shots distributed in random locations on the surface of the matrix spot. The instrument was externally calibrated in positive reflector mode using Bradykinin (1–7) [M+H]+ 757.3991, Angiotensin II [M+H]+ 1 046.5418, Angiotensin I [M+H]+ 1 296.6848, Substance P [M+H]+ 1347.7354, Bombesin [M+H]+ 1619.8223, ACTH (1–17) [M+H]+ 2 093,0862. For analysis, a mixture of 1 μl of purified enterocin and 1 μl of α-cyano-4-hydroxycinnamic acid (10 mg/ml 70:30 water/acetonitrile with 0.1% TFA) was spotted onto a MALDI-TOF MTP 384 target plate (Bruker Daltonik GmbH, Leipzig, Germany) according to the procedure of the dried-droplet preparation.
A pure colony of each Staphylococcus strains used in this study was grown overnight in BHI medium at 37°C. Afterward 10 μl of each overnight culture were added to the wells of bioassay microplates of 96 well cell culture plate (Cellstar) containing different concentrations of enterocins DD28 and DD93, ranging from 50 to 800 mg/l of each bacteriocin. The minimal inhibitory concentration (MIC) is defined as the lowest concentration of an antibiotic that will inhibit the visible growth of a microorganism after overnight incubation.
Antimicrobial agent interactions were determined using checkerboard assay. The concentrations used for enterocins DD28 and DD93 were comprised between 25 and 400 mg/l, while those used for erythromycin and kanamycin were ranging from 0.25 to 64 mg/l. Microplates were inoculated with MRSA-S1 strain at about 106 CFU/ml, in a final volume of 200 μl per well, and incubated overnight at 37°C. The fractional inhibitory concentration index (FICI) was calculated for each combination using the following formula: FICA + FICB = FICI, where FICA = MIC of drug A in combination/MIC of drug A alone, and FICB = MIC of drug B in combination/MIC of drug B alone. The FICI was interpreted as follow: synergism = FICI ≤ 0.5; indifference = 0.5 < FICI ≤ 4; antagonism = FICI > 4 (Petersen et al., 2006).
This experiment was realized on the MRSA strain. Tubes containing BHI supplemented with enterocin DD28, enterocin DD93, erythromycin, kanamycin or combination of bacteriocin and antibiotic, at previously defined concentrations during checkboard assay, were inoculated with MRSA-S1 strain to a density of about 5 × 105 CFU/ml in a final volume of 5 ml and incubated at 37°C for 24 h. The killing kinetics of the enterocin DD28 alone, enterocin DD93 alone, erythromycin alone, kanamycin alone and enterocin DD28 or enterocin DD93 in combination with erythromycin or kanamycin was assessed against MRSA-S1 strain using standard time-killing experiments and viable bacterial counts on agar plates. The final concentration of enterocin DD28 and enterocin DD93 was 50 mg/l, erythromycin 1 mg/l, kanamycin 4 mg/l and for combination of bacteriocins with antibiotics the concentrations used were 50 mg/l for enterocin DD28 and DD93 and 1 mg/l or 4 mg/l, respectively, for erythromycin and kanamycin.
Aliquots were removed at different times 0, 3, 6, 9, and 24 h of incubation, and then serially diluted in saline solution for determination of viable counts. Diluted samples (100 μl) were plated on Tryptone Soya Agar (TSA) plates and colonies were counted after overnight incubation at 37°C. Bactericidal activity was determined as 3 log10 CFU/ml reduction in the colony count relative to the initial inoculums (Pankuch et al., 1994).
Total DNA from E. faecalis 28 and E. faecalis 93 were extracted following the same procedure recently described by Al-Atya et al. (2015). The following forward: ATGGGAGCAATCGCAAAATTAGTAG and reverse: TTAATGTCTTTTTAGCCATTTTTCAATTTG primers (Liu et al., 2011) were used to amplify the genes encoding enterocin L50A and L50B. The PCR conditions consisted in: initial denaturing step of 5 min at 95°C, followed by 30 cycles of 1 min denaturing at 95°C, 1 min annealing at a temperature specific for the primers for each of the known enterocin gene, and 10 min extension at 72°C. Polymerase chain reaction (PCR) was done using the PCR Master Mix (2X) (Thermo Scientific Fermentas, Villebon sur Yvette, France) as a mixture of Taq DNA polymerase. DNA extraction was performed using the Wizard® Genomic DNA Purification Kit (Promega Corp., Madison, WI, USA). Ligation of PCR products was done into pGEM-T Easy vector (Promega Corp., Madison, WI, USA). Plasmid extraction was carried out using GeneJET Plasmid DNA Purification Kit (Thermo Scientific Fermentas). Restriction endonucleases were supplied by Thermo Scientific Fermentas. Ligation of inserts to different vectors was effected using the DNA Ligation Kit < Mighty Mix > from Takara (Ozyme, Saint Quentin en Yvelines, France). Recovery of DNA from agarose gels was performed with GeneJET Gel Extraction kit (Thermo Scientific Fermentas).
In all cases, the instructions of the suppliers were followed. All the construction sequences were checked by DNA sequencing performed at Eurofins MWG operon (Ebersberg, Germany).
To study the effect of treatment of stainless steel slide, reproducing abiotic surfaces used in healthcare units and industries environment, we used the protocol adapted from Ait Ouali et al. (2014). Briefly 2 ml of antimicrobials at their MIC values: enterocin DD28 (200 mg/l), enterocin DD93 (200 mg/l), erythromycin (8 mg/l), ofloxacin (0.5 mg/l), vancomycin (1 mg/l), rifampin (0.03 mg/l), and combination of enterocin DD28 with erythromycin (50/1 mg/l) were added on the surface of each AISI 304L slide placed in sterile Petri plates, and incubated for 2 h at 37°C. During this step a conditioning film on the AISI surface may be formed. Sterile Tryptone Soy Broth – Yeast Extract (TSB-YE) was added in an additional AISI304L slide as a control for this conditioning step. After this step the antimicrobial compounds and the TSB-YE were removed and replaced by 2 ml of MRSA-S1 strain suspension at 107 CFU/ml. After 1 h of incubation time, the supernatant containing non-adherent MRSA-S1 strain cells was removed and replaced by 2 ml of sterile TSB-YE medium on the surface of each slide, and the incubation was conducted for 0, 3, 6, and 24 h to survey the installation of MRSA-S1 strain biofilm. After each time of incubation, the slides were washed twice with 30 ml of Phosphate Buffered Saline (PBS) (pH 7.0). Finally, the AISI 304L stainless steel slides were immersed individually in 30 ml phosphate buffer and sonicated. The detached MRSA-S1 cells were enumerated by plating the bacteria on TSA after growth at 37 °C for 24 h. Additional slides were prepared and served for epifluorescence observation after their staining with live/dead components as explained below.
For scanning electron microscope (SEM) analysis, sterilized glass slides of 1 cm2 were deposited in wells of sterile 24-well tissue culture plates (BD Falcon, USA). Then, as previously, the glass slides were treated with 2 ml of these following antimicrobial compounds : enterocin DD28 (200 mg/l), vancomycin (1 mg/l) and combination of enterocin DD28 with erythromycin (50/1 mg/l), or TSB-YE medium as control, during 2 h at 37°C. After this conditioning step of the glass slides, the antimicrobial compounds solutions were removed and replaced by 2 ml of MRSA suspension at 107 CFU/ml. The following steps are exactly the same applied on the AISI304L stainless steel slides; however, incubation was conducted for 5 consecutive days, changing the TSB-YE medium each 24 h. Once the incubation ended, the glass slides were prepared for SEM analysis.
Biofilm cells were stained with a BacLight LIVE/DEAD bacterial viability staining kit according to the manufacturer’s instructions (Molecular Probes, Invitrogen, France). After dilution of 1.5 μl of each reagent with 1 ml of physiological water (0.85% m/v NaCl), the obtained mixture was gently deposited on the upper face of the slide where the biofilm development may occurred. Following the incubation (15 min in the dark) of the slides, the staining solution was aspirated and biofilms were observed using an epifluorescence microscope (Nikon Optiphot-2 EFD3, Japan).
The first step of the purification procedure permitted to enhance the specific activity recovered from the CFS, from 86.20 to 3,902.44 AU/mg (enterocin DD28) and 96.15 to 2,318.84 AU/mg (enterocin DD93) (Table 1). The second purification step involving a separation on RP-HPLC column permitted to purify these enterocins to homogeneity and evaluate their runtime to 42.5 min. Importantly, the specific activity has increased to 67,368.42 AU/mg for enterocin DD28 and 77,108.43 AU/mg for enterocin DD93 (Table 1). The purified enterocins DD28 and DD93 were analyzed by mass spectrometry and appeared to have very close molecular masses of, respectively, 5,205.21 Da and 5,204.89 Da (Figure 1).
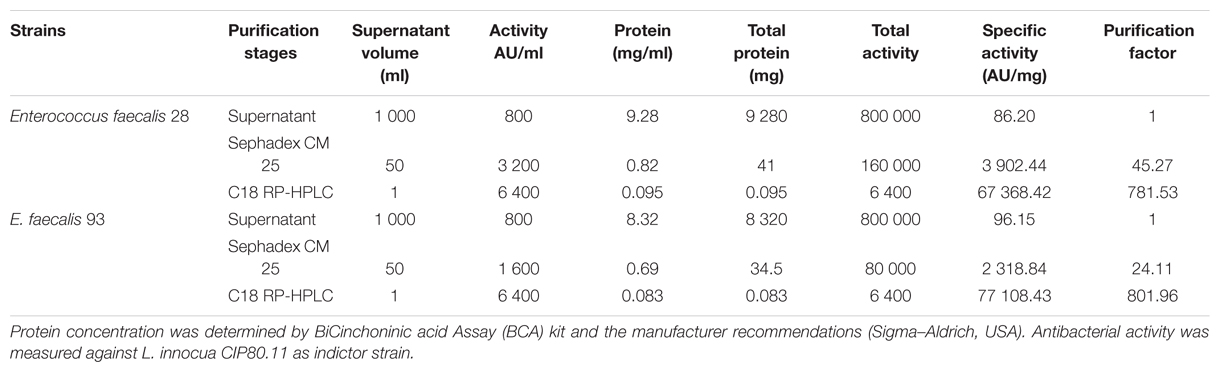
TABLE 1. Purification of Enterocins DD 28 and DD93.
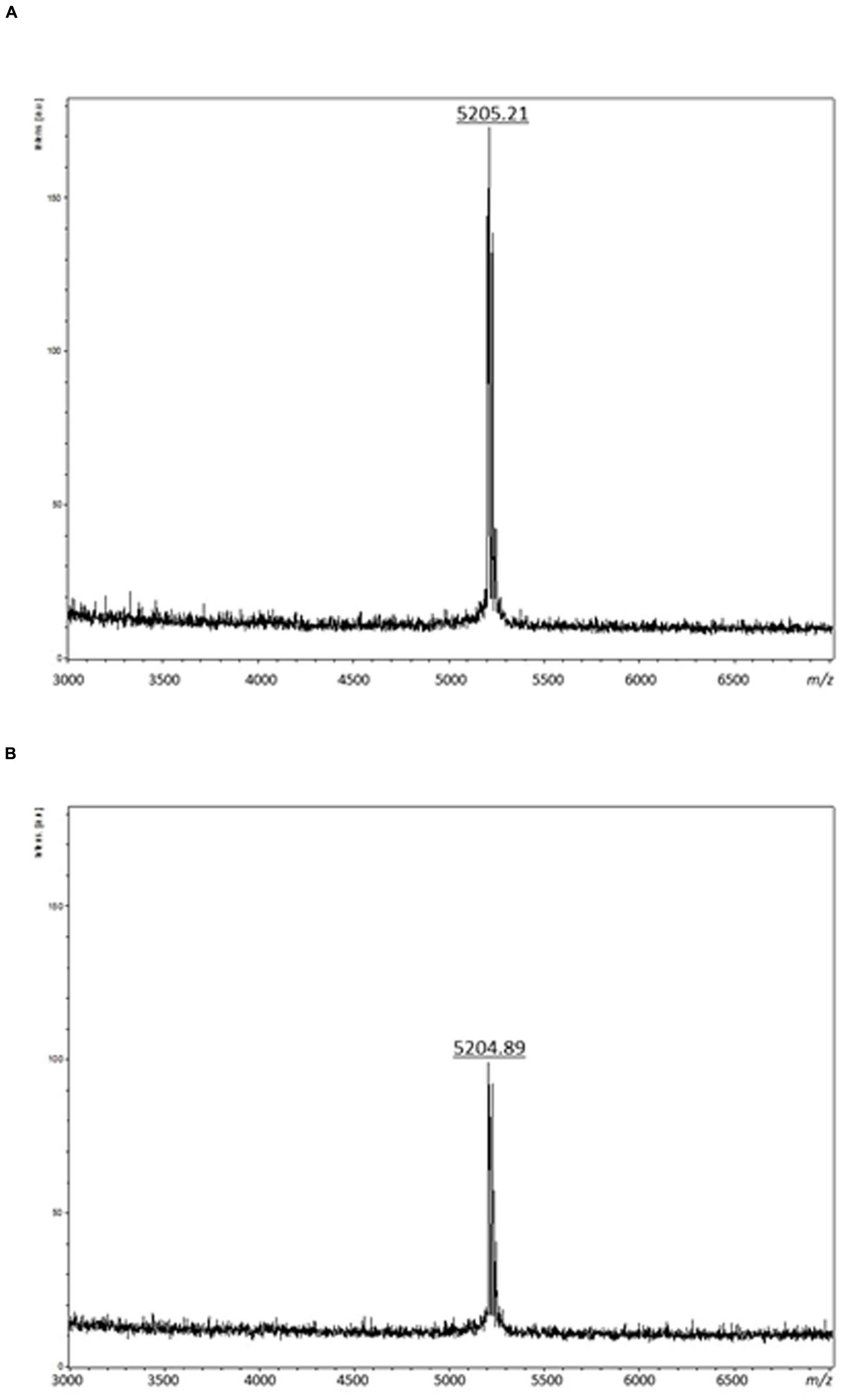
FIGURE 1. Mass spectrometry analyses of purified enterocin DD28 (A) and enterocin DD93 (B).
Different S. aureus strains were used to demonstrate the enterocins antibacterial activity. Thus strains S. epidermidis, S. aureus ATCC 33862, MRSA ATCC 43300, MRSA-S1, and MRSA-S2 were used as target strains. The MICs values were similar for S. epidermidis and S. aureus ATCC 33862, but they were twice higher for MRSA ATCC 43300, MRSA-S1 and MRSA-S2 strains. For this study, we have then proceeded with MRSA-S1 strain that was previously isolated from the blood of an 83 years old patient and studied for its susceptibility to chromagranin-derived peptides (Aslam et al., 2013).
The susceptibility of MRSA-S1 strain as determined by VITEK2 method showed resistance to erythromycin and kanamycin with MICs values of ≥8 mg/l and 32 mg/l, respectively. In turns, this strain exhibited sensibility to ofloxacin (0.5 mg/l), rifampin (0.03 mg/l), and vancomycin (1 mg/l) (Table 2). Notably, both enterocins DD28 and DD93 showed inhibition against MRSA-S1 strain and S. aureus ATCC 33862 used as controls (Table 3). The MICs values were 100 mg/l for S. aureus ATCC 33862, and 200 μg/ml for MRSA-S1 strain (Table 3).
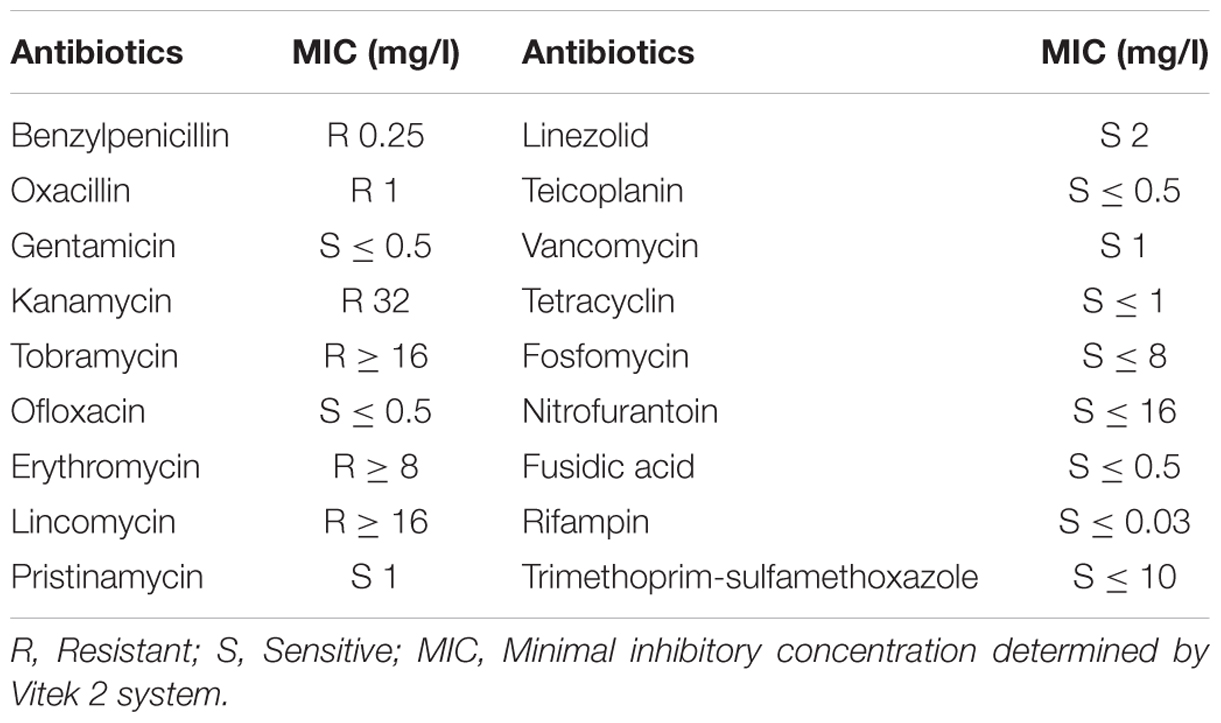
TABLE 2. Antibiotic susceptibility of MRSA-S1 strain.
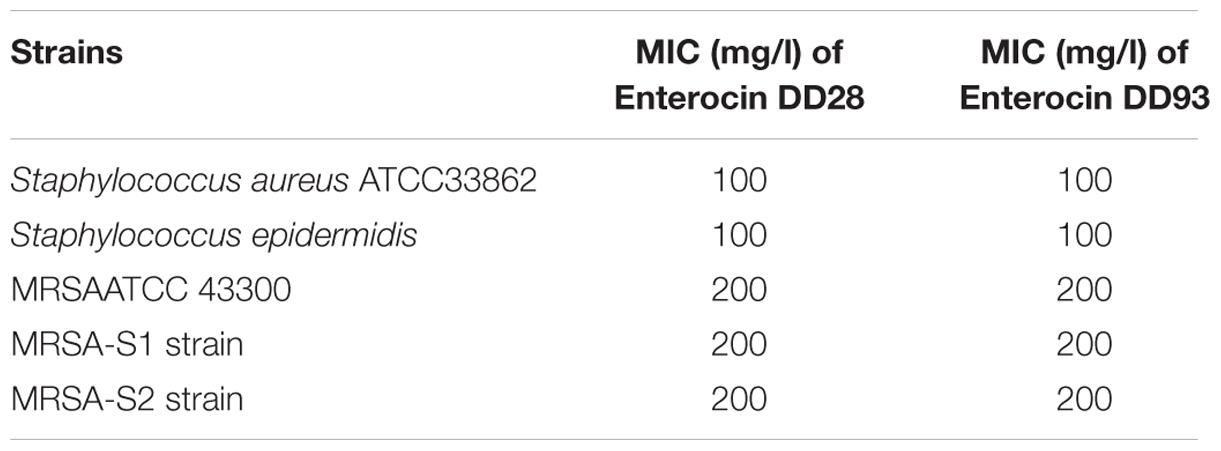
TABLE 3. Determination of minimum inhibitory concentration (MIC).
When erythromycin and kanamycin were used in combination with enterocin DD28 or enterocin DD93, the MICs values were lower than those obtained when these molecules were tested individually. These combinations permitted a synergetic effect regarding the FIC values of 0.31 registered for both enterocins DD28 and DD93, in combination with erythromycin and kanamycin on MRSA (Table 4). As evidenced from this assay, the MICs values of erythromycin and kanamycin have dropped under their breakpoint points (EUCAST, 2016) when they were associated to enterocins DD28 and DD93.
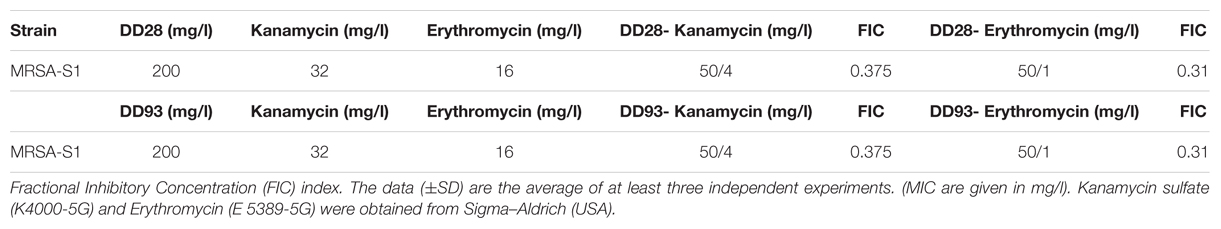
TABLE 4. Effects of antimicrobials combinations against MRSA-S1 strain.
The killing curves experiments realized on MRSA-S1 strain treated with erythromycin and kanamycin and their combinations with enterocins DD28 or DD93 have confirmed the synergistic effect anticipated by the FIC values. As expected, the aforementioned antibiotics are devoid of any inhibitory activity against the planktonic MRSA-S1 culture. The population number as means of CFU/ml remained identical in the untreated samples, as well as in those treated with erythromycin and kanamycin alone portraying this lack of activity. Nevertheless, the combination of enterocins DD28 and DD93 with erythromycin and kanamycin reduced the CFU/ml counts of MRSA-S1 by at least 2–3 logs during 3–24 h of incubation, leading to a killing of 99–99.9% of the CFU/ml of the initial bacterial populations (Figure 2; Table 5).
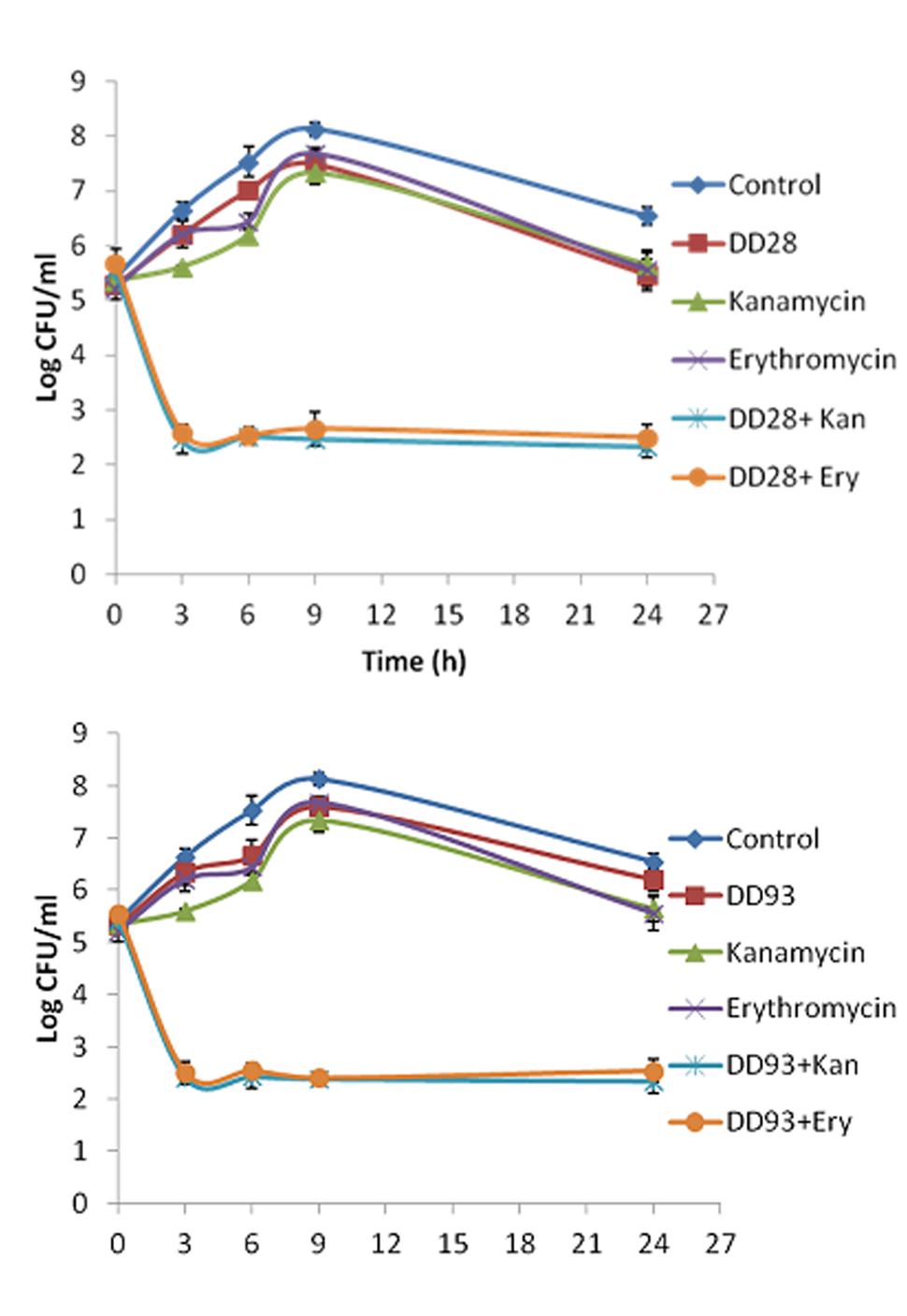
FIGURE 2. Time-killing kinetics on planktonic cultures of MRSA-S1 strain at 0, 3, 6, 9, and 24 h in the presence of enterocins DD28 and DD93 (50 mg/l) alone or combined with kanamycin (4 mg/l) and erythromycin (1 mg/l) against SARM-S1. (A) Shows the effect of enterocin DD28+Kanamycin at 50 mg/l-4 mg/l and enterocin DD28+erythromycin at 50-1 mg/l against MRSA-S1. (B) Shows the effect of enterocin DD93+kanamycin (50-4 mg/l) and enterocin DD93+erythromycin (50-1 mg/l) against MRSA-S1. The data (±SD) are the average of at least three independent experiments. In each experiment, three measures were performed.
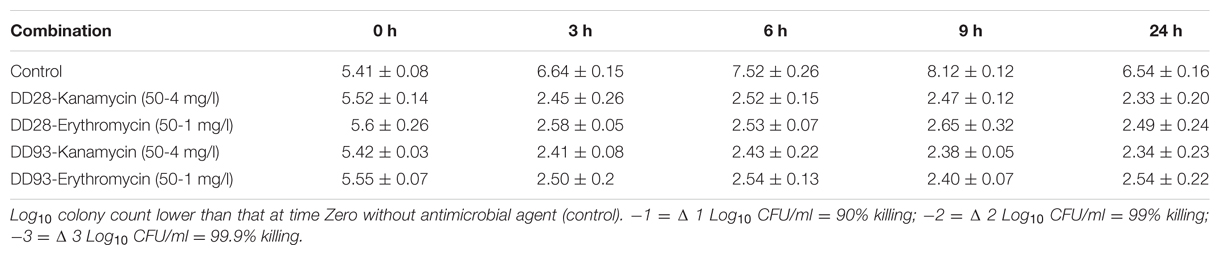
TABLE 5. Effect of DD28-Kanamycin, DD28-Erythromycin, DD93-Kanamycin, and DD93-Kanamycin combination against MRSA-S1.
Amplification of total DNA from E. faecalis 28 and E. faecalis 93 with the reverse and forward primers previously used to amplify DNA coding for enterocins L50A and L50B (Cintas et al., 1998) permitted in the present study to obtain 287 bp DNA amplicons. Afterward, these amplicons were successfully cloned into the pGEM-T plasmid and sequenced using 7T and SP6 primers. The resulting sequences were blasted on blastn pubmed database and showed complete alignment with the sequences of enterocin MR10A and MR10B (Figure 3), which were two class IIb bacteriocins produced by E. faecalis MRR 10-3 (Martín-Platero et al., 2006).

FIGURE 3. Alignment of putative enterocins DD98 and DD93 DNA sequences using ClustalO (http://www.ebi.ac.uk/Tools/msa/) software. (.) Sign indicates same nucleotide in the considered position for all the sequences aligned.
MRSA-S1 exhibited resistance to erythromycin. Thus, when the AISI 304L stainless steel slides were conditioned with 8 mg/l erythromycin, biofilm formation, as supported by epifluorescence microscopy, occurred normally. These results were comparable to those obtained with the untreated stainless steel slide (Figure 4; Table 6), advocating that MRSA-S1 biofilm formation was not affected by this antibiotic. Enterocins DD 28 or DD93 alone, at 200 mg/l, delayed the MRSA-S1 biofilm formation compared to the control test (Figure 4; Table 6). Remarkably, this result is similar to those obtained with vancomycin (1 mg/l) and rifampin (0.03 mg/l), for which the MRSA-S1 strain exhibited susceptibility (Table 1), and used as positive controls. Further, the combination of erythromycin and enterocin DD28 at 1 mg/l and 50 mg/l, respectively, has led to similar feature as that obtained with antibiotics.
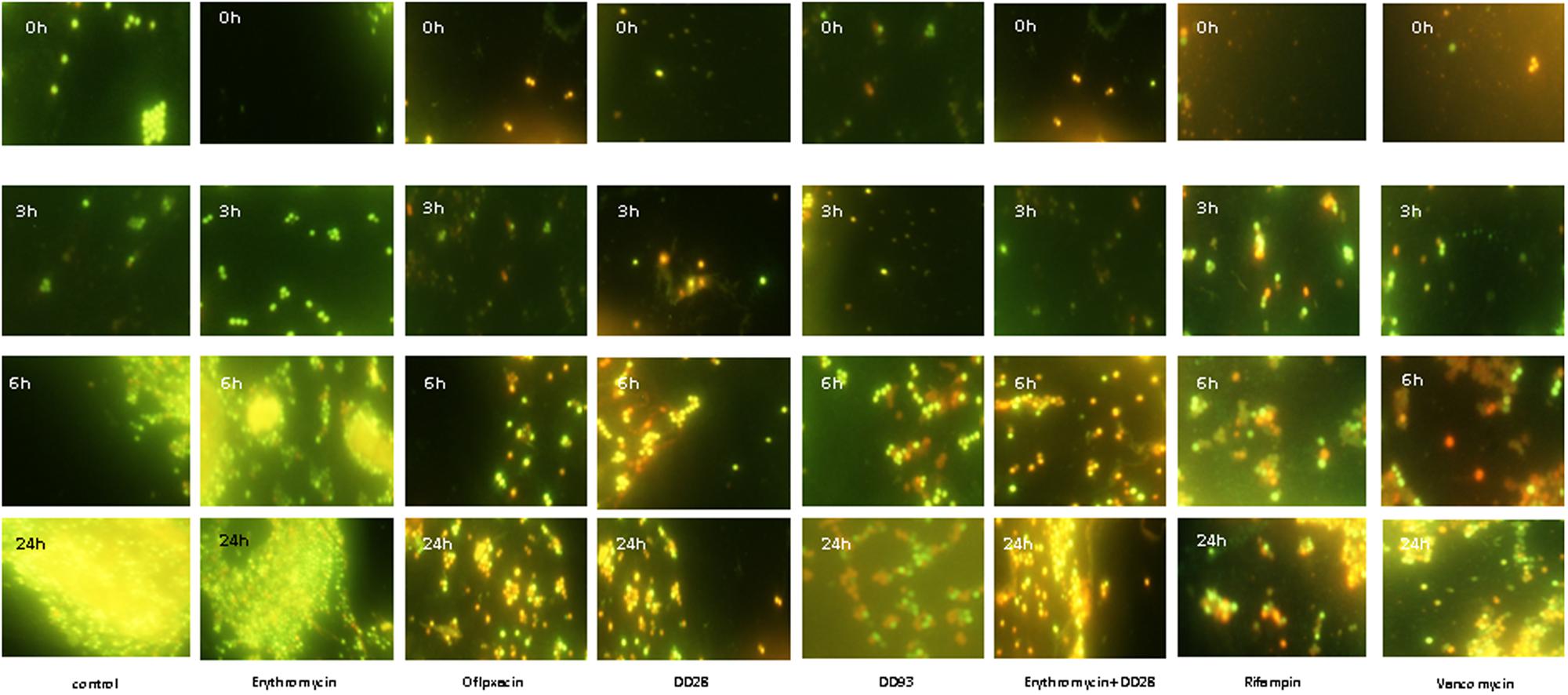
FIGURE 4. Biofilms formation by MRSA-S1 strain on AISI 304L stainless steel slides conditioned with antimicrobial agents for 2 h and then washed and inoculated with 2 ml of 107 CFU/ml MRSA-S1 culture before removing, washing and adding TSB-YE medium for 0, 3, 6, and 24 h of incubation at 37°C. Concentrations of antimicrobial agents used were 8 mg/l for erythromycin, 0.03 mg/l for rifampin, 200 mg/L for enterocin DD28, 50 mg/l + 1 mg/l for enterocin DD28 + erythromycin, respectively. Biofilms were stained with the BacLight Live/Dead Viability Kit and imaged by epifluorescence microscopy after staining pattern for live cells (green) and dead cells (red). The experiments were performed at least twice and representative images are shown.
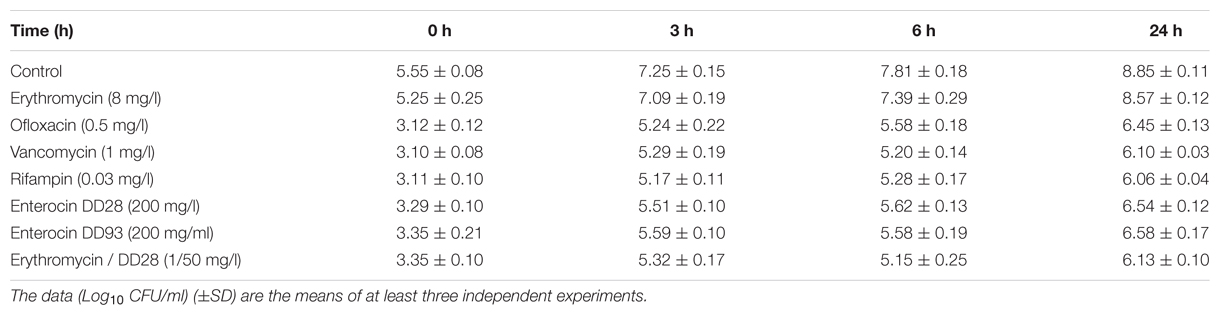
TABLE 6. Effect of antimicrobials on MRSA-S1 biofilm formation at different incubation times on AISI 304L stainless steel slides.
Scanning electron microscope microscopy analysis showed lower number of adherent MRSA-S1 cells on glass devices after conditioning with vancomycin (1 mg/l) and a combination of enterocin DD28+erythromycin (50/1 mg/l), comparatively to the unconditioned glass slide (Figure 5). However, the biofilm formation on a glass slide treated with enterocin DD28 alone (200 mg/l) showed more limited effect on the colonization of the surface compared to an untreated glass slide (Figure 5).

FIGURE 5. Scanning electron microscope (SEM) of biofilms formation by MRSA-S1 strain on glass slides. (A) Corresponds to the untreated sample (control) carrying 5 days old biofilm. (B–D) Show the samples conditionned with enterocin DD28 (200 mg/l), Erythromicin + enterocin DD28 (1–50 mg/l) and vancomycin (1 mg/l), respectively.
Staphylococcus aureus is the most often species encountered in the infectious diseases, generally as benign ones, but it can be involved in more serious infections as pneumonia, bacterial meningitis, and gastroenteritis. Moreover, this pathogen was also implied in the nosocomial infections, sometimes affecting bloodstream, joints, bones, lungs, or heart, resulting in very complicated infections, even with fatal epilog (Falcone et al., 2015). The methicillin-resistance was associated with a resistance to all β-lactams and some other antibiotics such as aminoglycosides and related macrolides, synergistines, fluoroquinolones, and fosfomycin. Some molecules including glycopeptides, rifampin, and fusidic acid remain active against MRSA (Hamdad et al., 2006; Falcone et al., 2015).
Staphylococci were shown to form biofilms on abiotic surfaces such as stainless steel, catheters and polystyrene material (Otto, 2008), and the biofilm lifestyle stands as a hurdle to antibiotic treatments that need to be overcome. The icaADBC genes coding for synthesis of exopolysaccharide of polymer matrix were shown to be involved in the biofilm formation (Tremblay et al., 2014). Biofilms act as a physical barrier, limiting drugs penetration. The ability of biofilm formation was also observed for other staphylococci species as S. epidermidis (Tremblay et al., 2014; Agarwal et al., 2015).
The present study permitted to shed light on the potential of enterocins DD28 and DD93 to help treating the MRSA burden. Furthermore, MRSA from hospital settings, are known to be resistant to erythromycin and kanamycin but remain sensitive to vancomycin and rifampin (Dumitrescu et al., 2010; Bhattacharya et al., 2015; Parhizgari et al., 2016).
Enterocins DD28 and DD93, two class IIb bacteriocins produced by E. faecalis DD28 and E. faecalis DD93 strains recently isolated from meconium were purified using the protocol described by Abriouel et al. (2003) and characterized for their masses and DNA sequences. Enterocins DD28 and DD93 masses were perfectly matching with those reported for enterocins enterocins L50A and L50B (Cintas et al., 2000), enterocins MR10A and MR10B (Abriouel et al., 2003) and enterocins A5–11A and A5–11B (Batdorj et al., 2006).
The anti-MRSA activity obtained with the semi-purified enterocins DD28 and DD93 appeared to be strain dependent. The highest MIC value was observed for the clinical MRSA-S1 strain, whilst the lowest ones were observed for S. epidermidis and S. aureus ATCC 33862. The anti-Staphylococcal activities obtained with enterocins DD28 and DD93 were less pronounced than the treatment afflicted by the antibiotics alone. Nevertheless, the combinations of these antibiotics with enterocins have revealed very promising therapeutic options. Indeed, bacteriocins were able to synergize, as supported by the FICI values and the killing curves experiments, the effects of antibiotics. The important drop of MRSA-S1 strain growth, when treated with enterocin DD28 or DD93 combined to erythromycin or kanamycin, occurred during the first 3 h of growth. Related to this, a drop of about 3 Log10 CFU/ml, was registered upon treatment of MRSA-S1 strain with the aforementioned antibacterial combinations. Based on this data, we assume that bacteriocins inputs for the anti-MRSA-S1 activity happened in the beginning of the treatment before their possible but plausible degradation. As the anti-MRSA-S1 activity was improved by the addition of the bacteriocins, and this strain could switch from a resistant to sensitive phenotype for the tested antibiotics, we support that anti-MRSA activity could be improved by incorporation of bacteriocins and the problem of their stability could be resolved by using encapsulated bacteriocins or nanoparticles coated bacteriocins.
Staphylococci were also reported to form biofilms on different biomaterials (Nan et al., 2015) designing them as guilty of recurrent infections taking place in the healthcare units (Amalaradjou and Venkitanarayanan, 2014). Mature biofilms are extremely difficult to eradicate (Dosler and Mataraci, 2013) complicating thereof treatment of this pathogen. The conditioning of AISI 304L stainless steel and glass slides, with enterocin DD28 and antibiotics impeded the biofilm formation by the MRSA-S1 strain. The combinations of these antimicrobials has not only impacted the growth of the MRSA-S1 strain under planktonic culture but hampered the biofilm formation. The MRSA-S1 strain population was reduced of about 2 Log10 CFU/ml, on AISI 304L stainless steel slides. This data was logically correlated to observations resulting from the epifluorescence and SEM analyses.
This study permitted to shed light on the anti-MRSA activity of enterocins DD28 and DD93. Importantly, these bacteriocins were able to synergize with erythromycin and kanamycin, two antibiotics used in the MRSA treatment. The data gathered in the frame of this work enabled us to confirm the role of bacteriocins as a novel class of antibiotics to assist or replace the fading antibiotics. The perfect similarities exhibited by DNA and amino-acids sequences of enterocins DD28 and DD93 vs. enterocins MR10A and MR10B let us to think that these bacteriocins are most probably similar.
SS and RB performed the SEM analysis of MRSA-S1 strain grown on glass (treated or not with antimicrobials). AV realized the Vitek-2antibiograms and provided us with the S. epidermidis strain used as well in this work. AA realized most of the experiments suchas MIC values, Killing curves, biofilms assays on steel slides and glass. AA did also the epifluorescence imaging YB contributed to MIC determinations and DNA sequences analyses GC realized the mass analysis of enterocins DD28 and DD93, RR and DD supervised this work. All authors read and approved the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
AA received a Ph.D. fellowship from French and Iraqi governments. The authors are indebted to Dr. Gilles Prévost (Strasbourg, France) and Prof. Giuseppe Spano (Foggia, Italy) for the gift of the clinical MRSA-S1 and MRSA-S2 strains, and critical reading of the manuscript, respectively. The authors are also indebted to ANR for the financial support on alternatives to antibiotics in the animal health. The authors thank also Nicolas Barrois from the Center for Infection and Immunity of Lille (UMR 8204-INSERM U1019) for helping with SEM analysis.
Abriouel, H., Valdivia, E., Martínez-Bueno, M., Maqueda, M., and Gálvez, A. (2003). A simple method for semi-preparative-scale production and recovery of enterocin AS-48 derived from Enterococcus faecalis subsp. liquefaciens A-48-32. J. Microbiol. Methods 55, 599–605. doi: 10.1016/S0167-7012(03)00202-1
Agarwal, S., Sharma, G., Dang, S., Gupta, S., and Gabrani, R. (2015). Antimicrobial peptides as anti-infectives against Staphylococcus epidermidis. Med. Princ. Pract. doi: 10.1159/000443479 [Epub ahead of print].
Ait Ouali, F., Al Kassaa, I., Cudennec, B., Abdallah, M., Bendali, F., Sadoun, D., et al. (2014). Identification of lactobacilli with inhibitory effect on biofilm formation by pathogenic bacteria on stainless steel surfaces. Int. J. Food. Microbiol. 191, 116–124. doi: 10.1016/j.ijfoodmicro.2014.09.011
Al-Atya, A. K., Drider-Hadiouche, K., Ravallec, R., Silvain, A., Vachee, A., and Drider, D. (2015). Probiotic potential of Enterococcus faecalis strains isolated from meconium. Front. Microbiol. 6:227. doi: 10.3389/fmicb.2015.00227
Amalaradjou, M. A., and Venkitanarayanan, K. (2014). Antibiofilm effect of octenidine hydrochloride on Staphylococcus aureus, MRSA and VRSA. Pathogens 3, 404–416. doi: 10.3390/pathogens3020404
Aslam, R., Marban, C., Corazzol, C., Jehl, F., Delanade, F., Van Dorsselaer, A., et al. (2013). Cateslytin, a chromogranin A derived peptide is active against Staphylococcus aureus and resistant to degradation by its proteases. PLoS ONE 8:e68993. doi: 10.1371/journal.pone.0068993
Bassetti, M., and Righi, E. (2015). Development of novel antibacterial drugs to combat multiple resistant organisms. Langenbecks Arch. Surg. 400, 153–165. doi: 10.1007/s00423-015-1280-4
Batdorj, B., Dalgalarrondo, M., Choiset, Y., Pedroche, J., Métro, F., Prévost, H., et al. (2006). Purification and characterization of two bacteriocins produced by lactic acid bacteria isolated from Mongolian airag. J. Appl. Microbiol. 101, 837–848. doi: 10.1111/j.1365-2672.2006.02966.x
Bhattacharya, S., Bir, R., and Majumdar, T. (2015). Evaluation of multidrug resistant Staphylococcus aureus and their association with biofilm production in a Tertiary Care Hospital, Tripura, Northeast India. J. Clin. Diagn. Res. 9, DC01–DC04. doi: 10.7860/JCDR/2015/13965.6417
Birri, D. J., Brede, D., Forberg, T., Holo, H., and Nes, I. F. (2010). Molecular and genetic characterization of a novel bacteriocin locus in Enterococcus avium isolates from infants. Appl. Environ. Microbiol. 76, 483–492. doi: 10.1128/AEM.01597-09
Cintas, L. M., Casaus, P., Herranz, C., Hâvarstein, L. S., Holo, H., Hernández, P. E., et al. (2000). Biochemical and genetic evidence that Enterococcus faecium L50 produces enterocins L50A and L50B, the sec-dependent enterocin P, and a novel bacteriocin secreted without an N-terminal extension termed enterocin Q. J. Bacteriol. 182, 6806–6814. doi: 10.1128/JB.182.23.6806-6814.2000
Cintas, L. M., Casaus, P., Holo, H., Hernandez, P. E., Nes, I. F., and Håvarstein, L. S. (1998). Enterocins L50A and L50B, two novel bacteriocins from Enterococcus faecium L50, are related to staphylococcal hemolysins. J. Bacteriol. 180, 1988–1994.
Claeys, K. C., Smith, J. R., Casapao, A. M., Mynatt, R. P., Avery, L., Shroff, A., et al. (2015). Impact of the combination of daptomycin and trimethoprim-sulfamethoxazole on clinical outcomes in methicillin-resistant Staphylococcus aureus infections. Antimicrob. Agents Chemother. 59, 1969–1976. doi: 10.1128/AAC.04141-14
David, M. Z., and Daum, R. S. (2010). Community-associated methicillin-resistant Staphylococcus aureus: epidemiology and clinical consequences of an emerging epidemic. Clin. Microbiol. Rev. 23, 616–687. doi: 10.1128/CMR.00081-09
De Man, J. C., Rogosa, M., and Sharpe, E. (1960). A medium for the cultivation of lactobacilli. J. Appl. Bacteriol. 23, 130–135. doi: 10.1111/j.1365-2672.1960.tb00188.x
Dosler, S., and Mataraci, E. (2013). In vitro pharmacokinetics of antimicrobial cationic peptides alone and in combination with antibiotics against methicillin resistant Staphylococcus aureus biofilms. Peptides 49, 53–58. doi: 10.1016/j.peptides.2013.08.008
Drider, D., and Rebuffat, S. (2011). Prokaryotic Antimicrobial Peptides: From Genes to Applications. New York, NY: Springer.
Dumitrescu, O., Dauwalder, O., Boisset, S., Reverdy, M. E., Tristan, A., and Vandenesch, F. (2010). Résistance aux antibiotiques chez Staphylococcus aureus: les points-clés en 2010. Med. Sci. 26, 943–949. doi: 10.1051/medsci/20102611943
EUCAST (2016). European Committee on Antimicrobial Susceptibility Testing Breakpoint Tables for Interpretation of MICs and Zone Diameters Version 6.0, Valid From 2016-01-01. Available at: http://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_6.0_Breakpoint_table.pdf
Falcone, M., Russo, A., and Venditti, M. (2015). Optimizing antibiotic therapy of bacteremia and endocarditis due to staphylococci and enterococci: new insights and evidence from the literature. J. Infect. Chemother. 21, 330–339. doi: 10.1016/j.jiac.2015.02.012
French Committee on Antimicrobial Susceptibility Testing [FCAST] (2015). du Comité Français de L’Antibiogramme de la Société Française de Microbiologie. Available at: http://www.sfm-microbiologie.org/UserFiles/files/casfm/CASFM_EUCAST_V1_2015.pdf
Goto, D. K., and Yan, T. (2011). Effects of land uses on fecal indicator bacteria in the water and soil of a tropical watershed. Microbes Environ. 26, 254–260. doi: 10.1264/jsme2.ME11115
Hamdad, F., Donda, F., Lefebvre, J. F., Laurans, G., Biendo, M., Thomas, D., et al. (2006). Detection of methicillin/oxacillin resistance and typing in aminoglycoside-susceptible methicillin-resistant and kanamycin-tobramycin-resistant methicillin-susceptible Staphylococcus aureus. Microb. Drug Resist. 12, 177–185. doi: 10.1089/mdr.2006.12.177
Hassan, M., Kjos, M., Nes, I. F., Diep, D. B., and Lotfipour, F. (2012). Natural antimicrobial peptides from bacteria: characteristics and potential applications to fight against antibiotic resistance. J. Appl. Microbiol. 113, 723–736. doi: 10.1111/j.1365-2672.2012.05338.x
Holmes, N. E., Tong, S. Y., Davis, J. S., and van Hal, S. J. (2015). Treatment of methicillin-resistant Staphylococcus aureus: vancomycin and beyond. Semin. Respir. Crit. Care Med. 36, 17–30. doi: 10.1055/s-0034-1397040
Ishibashi, N., Himeno, K., Fujita, K., Masuda, Y., Perez, R. H., Zendo, T., et al. (2012). Purification and characterization of multiple bacteriocins and an inducing peptide produced by Enterococcus faecium NKR-5-3 from Thai fermented fish. Biosci. Biotechnol. Biochem. 76, 947–953. doi: 10.1271/bbb.110972
Kang, J. H., and Lee, M. S. (2005). Characterization of a bacteriocin produced by Enterococcus faecium GM-1 isolated from an infant. J. Appl. Microbiol. 98, 1169–1176. doi: 10.1111/j.1365-2672.2005.02556.x
Kullar, R., Sakoulas, G., Deresinski, S., and van Hal, S. J. (2015). When sepsis persists: a review of MRSA bacteraemia salvage therapy. J. Antimicrob. Chemother. 71, 576–586. doi: 10.1093/jac/dkv368
Leone, S., Noviello, S., Boccia, G., De Caro, F., and Esposito, S. (2015). Methicillin-resistant Staphylococcus aureus infections: role of daptomycin/β-lactams combination. Infez. Med. 23, 99–104.
Line, J. E., Svetoch, E. A., Eruslanov, B. V., Perelygin, V. V., Mitsevich, E. V., Mitsevich, I. P., et al. (2008). Isolation and purification of enterocin E-760 with broad antimicrobial activity against gram-positive and gram-negative bacteria. Antimicrob. Agents Chemother. 52, 1094–1100. doi: 10.1128/AAC.01569-06
Liu, X., Vederas, J. C., Whittal, R. M., Zheng, J., Stiles, M. E., Carlson, D., et al. (2011). Identification of an N-terminal formylated, two-peptide bacteriocin from Enterococcus faecalis 710C. J. Agric. Food Chem. 59, 5602–5608. doi: 10.1021/jf104751v
Martín-Platero, A. M., Valdivia, E., Ruíz-Rodríguez, M., Soler, J. J., Martín-Vivaldi, M., Maqueda, M., et al. (2006). Characterization of antimicrobial substances produced by Enterococcus faecalis MRR 10-3, isolated from the uropygial gland of the hoopoe (Upupa epops). Appl. Environ. Microbiol. 72, 4245–4249. doi: 10.1128/AEM.02940-05
Mataraci, E., and Dosler, S. (2012). In vitro activities of antibiotics and antimicrobial cationic peptides alone and in combination against methicillin-resistant Staphylococcus aureus biofilms. Antimicrob. Agents Chemother. 56, 6366–6371. doi: 10.1128/AAC.01180-12
Naghmouchi, K., Baah, J., Hober, D., Jouy, E., Rubrecht, C., Sané, F., et al. (2013). Synergistic effect between colistin and bacteriocins in controlling Gram-negative pathogens and their potential to reduce antibiotic toxicity in mammalian epithelial cells. Antimicrob. Agents Chemother. 57, 2719–2725. doi: 10.1128/AAC.02328-12
Naghmouchi, K., Belguesmia, Y., Baah, J., Teather, R., and Drider, D. (2011). Antibacterial activity of class I and IIa bacteriocins combined with polymyxin E against resistant variants of Listeria monocytogenes and Escherichia coli. Res. Microbiol. 162, 99–107. doi: 10.1016/j.resmic.2010.09.014
Naghmouchi, K., Drider, D., Baah, J., and Teather, R. (2010). Nisin A and polymyxin B as synergistic inhibitors of Gram-positive and Gram-negative bacteria. Probiotics Antimicrob. Proteins 2, 513–517. doi: 10.1007/s12602-009-9033-8
Naghmouchi, K., Le Lay, C., Baah, J., and Drider, D. (2012). Antibiotic and antimicrobial peptide combinations: synergistic inhibition of Pseudomonas fluorescens and antibiotic-resistant variants. Res. Microbiol. 163, 101–108. doi: 10.1016/j.resmic.2011.11.002
Nan, L., Yang, K., and Ren, G. (2015). Anti-biofilm formation of a novel stainless steel against Staphylococcus aureus. Mater. Sci. Eng. C Mater. Biol. Appl. 51, 356–361. doi: 10.1016/j.msec.2015.03.012
Pankuch, G. A., Jacobs, M. R., and Appelbaum, P. C. (1994). Study of comparative antipneumococcal activities of penicillin G, RP 59500, erythromycin, sparfloxacin, ciprofloxacin, and vancomycin by using time-kill methodology. Antimicrob. Agents Chemother. 38, 2065–2072. doi: 10.1128/AAC.38.9.2065
Parhizgari, N., Khoramrooz, S. S., Malek Hosseini, S. A., Marashifard, M., Yazdanpanah, M., Emaneini, M., et al. (2016). High frequency of multidrug-resistant Staphylococcus aureus with SCCmec type III and Spa types t037 and t631 isolated from burn patients in southwest of Iran. APMIS 124, 221–228. doi: 10.1111/apm.12493
Petersen, P. J., Labthavikul, P., Jones, C. H., and Bradford, P. A. (2006). In vitro antibacterial activities of tigecycline in combination with other antimicrobial agents determined by chequerboard and time-kill kinetic analysis. J. Antimicrob. Chemother. 57, 573–576. doi: 10.1093/jac/dki477
Saavedra, L., Minahk, C., de Ruiz Holgado, A. P., and Sesma, F. (2004). Enhancement of the enterocin CRL35 activity by a synthetic peptide derived from the NH2-terminal sequence. Antimicrob. Agents Chemother. 48, 2778–2781. doi: 10.1128/AAC.48.7.2778-2781.2004
Sadaka, A., Durand, M. L., Sisk, R., and Gilmore, M. S. (2015). Staphylococcus aureus and its bearing on ophthalmic disease. Ocul. Immunol. Inflamm. 17, 1–11. doi: 10.3109/09273948.2015.1075559
Sánchez, J., Diep, D. B., Herranz, C., Nes, I. F., Cintas, L. M., and Hernández, P. E. (2007). Amino acid and nucleotide sequence, adjacent genes, and heterologous expression of hiracin JM79, a sec-dependent bacteriocin produced by Enterococcus hirae DCH5, isolated from Mallard ducks (Anas platyrhynchos). FEMS Microbiol. Lett. 27, 227–236. doi: 10.1111/j.1574-6968.2007.00673.x
Sawa, N., Wilaipun, P., Kinoshita, S., Zendo, T., Leelawatcharamas, V., Nakayama, J., et al. (2012). Isolation and characterization of enterocin W, a novel two-peptide lantibiotic produced by Enterococcus faecalis NKR-4-1. Appl. Environ. Microbiol. 78, 900–903. doi: 10.1128/AEM.06497-11
Singh, A. P., Preet, S., and Rishi, P. (2014). Nisin/β-lactam adjunct therapy against Salmonella enterica serovar Typhimurium: a mechanistic approach. J. Antimicrob. Chemother. 69, 1877–1887. doi: 10.1093/jac/dku049
Spagnolo, A. M., Orlando, P., Panatto, D., Amicizia, D., Perdelli, F., and Cristina, M. L. (2014). Staphylococcus aureus with reduced susceptibility to vancomycin in healthcare settings. J. Prev. Med. Hyg. 55, 137–144.
Tängdén, T. (2014). Combination antibiotic therapy for multidrug-resistant Gram-negative bacteria. Ups. J. Med. Sci. 119, 149–153. doi: 10.3109/03009734.2014.899279
Tomita, H., Fujimoto, S., Tanimoto, K., and Ike, Y. (1996). Cloning and genetic organization of the bacteriocin 31 determinant encoded on the Enterococcus faecalis pheromone-responsive conjugative plasmid pYI17. J. Bacteriol. 178, 3585–3593.
Tremblay, Y. D., Hathroubi, S., and Jacques, M. (2014). Bacterial biofilms: their importance in animal health and public health. Can. J. Vet. Res. 78, 110–116.
Keywords: enterocins, antibiotics, MRSA, synergism, bacteriocins
Citation: Al Atya AK, Belguesmia Y, Chataigne G, Ravallec R, Vachée A, Szunerits S, Boukherroub R and Drider D (2016) Anti-MRSA Activities of Enterocins DD28 and DD93 and Evidences on Their Role in the Inhibition of Biofilm Formation. Front. Microbiol. 7:817. doi: 10.3389/fmicb.2016.00817
Received: 20 February 2016; Accepted: 13 May 2016;
Published: 31 May 2016.
Edited by:
Maria Tereza Dos Santos Correia, Universidade Federal de Pernambuco, BrazilReviewed by:
Polpass Arul Jose, Madurai Kamaraj University, IndiaCopyright © 2016 Al Atya, Belguesmia, Chataigne, Ravallec, Vachée, Szunerits, Boukherroub and Drider. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Djamel Drider, ZGphbWVsLmRyaWRlckB1bml2LWxpbGxlMS5mcg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.