
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 05 February 2025
Sec. Marine Biology
Volume 12 - 2025 | https://doi.org/10.3389/fmars.2025.1490317
 Jaquelino Varela1,2*
Jaquelino Varela1,2* Catarina Pereira Santos1,2,3
Catarina Pereira Santos1,2,3 Emanuel Nunes1,4
Emanuel Nunes1,4 Vasco Pissarra1,2
Vasco Pissarra1,2 Stiven Pires5Bárbara P. Ribeiro6,7Eduarda Vieira6,7Tiago Repolho1,8Nuno Queiroz6,7
Stiven Pires5Bárbara P. Ribeiro6,7Eduarda Vieira6,7Tiago Repolho1,8Nuno Queiroz6,7 Rui Freitas9
Rui Freitas9 Rui Rosa1,2,8*
Rui Rosa1,2,8*The northeast Atlantic Ocean contains multiple habitats considered critical for shark conservation, including nursery areas, migratory corridors and aggregation sites. In this context, updating knowledge on shark diversity and the threats affecting them in this region is essential to defining priorities and implementing the right management and conservation measures. Here, we show that Cabo Verde, Canarias, Madeira and Azores islands are home to 78 shark species (comprising 26 families), and 56% are threatened with extinction. The Canary Islands revealed the greatest richness (with 56 species), followed by Cabo Verde (53), Madeira (52), and the Azores (45). Cabo Verde presents fewer similarities with the rest of the islands. We also found that: i) Azores share more species with the Canary Islands than Madeira (despite the greater geographical proximity with the latter), and ii) there are no oviparous species in the Cabo Verde archipelago, contrary to the Canary Islands (5), the Azores (4), and Madeira (3). Fishing and habitat degradation are the most relevant anthropogenic pressures for the region, with Cabo Verde having the highest number of endangered species (66%) and a greater magnitude and diversity of threats. As such, this archipelago presents the highest priority area for shark conservation due to the intense industrial fishing in its waters, poor management measures in combination with its greater vulnerability to climate change.
Sharks belong to the ancient Chondrichthyes group (Dulvy et al., 2017; Ebert et al., 2021). They appeared more than 400 million years ago on the planet and have survived profound transformations of the Earth (Davidson et al., 2016; Dulvy et al., 2014, 2017).With more than 500 species currently recognized, sharks are an ecologically important and functionally diverse group (Ebert et al., 2021). However, they are currently facing an unprecedented decline in their populations due mainly to increased anthropogenic pressures (Dulvy et al., 2014; Pacoureau et al., 2021). Their reproductive strategy, characterized by late sexual maturity, low fecundity, few offspring and low growth, renders this group more vulnerable to anthropogenic pressures (Frisk et al., 2001; Myers and Worm, 2005). It is estimated that around 100 million sharks are caught every year and, in the last 50 years some oceanic species have seen their populations decline by more than 70% (Pacoureau et al., 2021; Worm et al., 2013). As a result, one-third of sharks and rays are threatened with extinction mainly because of overfishing in targeted fisheries and incidental catches (Dulvy et al., 2021).
Overfishing is a major anthropogenic threat in the ocean, leading the decline in stocks of various organisms and disrupting marine ecosystems (Dulvy et al., 2021; Jackson et al., 2001; Myers et al., 2007; Roff et al., 2018). Industrial fishing in the Northeast Atlantic began in the 1950s and has recorded one of the highest initial catches per unit shelf area (Ferretti et al., 2010). Initially, sharks were caught as bycatch or by small-scale artisanal fishing for subsistence. However, with the global increase in their demand, mainly for their meat and fins, and the associated high yields, sharks are now the subject of targeted fishing (Dulvy et al., 2017). The North Atlantic, in specific, is one of the regions with the greatest overlap between industrial fishing and shark habitat use (Queiroz et al., 2016), representing one of the most heavily fished regions of the world, with the blue shark (Prionace glauca) and the shortfin mako shark (Isurus oxyrinchus) among the elasmobranchs most often captured by industrial fisheries (Camhi et al., 2008; Torres et al., 2016).
On the other hand, the northeast Atlantic Ocean represents an important area for the conservation of marine megafauna (Afonso et al., 2020). It is one of the most important cetacean biodiversity hotspots in the world (Afonso et al., 2020; Wenzel et al., 2023), hosts one of the largest sea turtle nesting grounds (Marco et al., 2012), and nursery habitats for endemic and critically endangered shark species (Jiménez‐Alvarado et al., 2020; Rosa et al., 2023). Within the region, a group of volcanic islands historically known as the Macaronesia biogeographic region stands out for its diversity of marine habitats, biodiversity, and richness of endemism (Afonso et al., 2020; Das and Afonso, 2017; Freitas et al., 2019; Roberts et al., 2002; Wirtz et al., 2013). While traditionally said to include the archipelagos of the Azores, Madeira, Canary Islands and Cabo Verde, there is no consensus on the inclusion of the latter in the Macaronesia biogeographic region based on the differences in diversity and endemism of marine organisms between the archipelagos (Freitas et al., 2019). However, the comparison of elasmobranchs species, particularly sharks, among archipelagos has seldom been explored, limiting its contribution to this debate. Although many sharks are highly migratory, ocean currents and climatic gradients can limit the distribution of less mobile species and segregate them according to their biological characteristics. In fact, these archipelagos are influenced by different climatic zones, with the Azores, Madeira and Canaries Islands being located in temperate or sub-tropical latitudes, while Cabo Verde falls under tropical latitudes (Spalding et al., 2007). Furthermore, the archipelagos are influenced by a complex system of ocean currents and sea surface temperature gradients ranging from 15 to 25 °C (Freitas et al., 2019; Reverdin et al., 2003).
Within this context, the main goal of the present study is to compare shark diversity patterns among the different archipelagos of the Cabo Verde, Canarias, Madeira and Azores islands, evaluate the main threats, and discuss where conservation strategies should be prioritized.
Cabo Verde, Canarias, Madeira and Azores archipelagos consist of islands of volcanic origin located in the Northeast Atlantic Ocean, spanning from 39°N 31°W to 15°N 23°W (Figure 1). It encompasses, from north to south, the Azores, Madeira, the Canary Islands, and Cabo Verde archipelagos (Illera et al., 2012). The archipelagos occupy a latitudinal extension of almost 3000 km and a great climatic gradient with the Azores in the temperate zone, Madeira and the Canary Islands under the Mediterranean climate and Cabo Verde in the tropics (Fernández-Palacios et al., 2011; Florencio et al., 2021). Due to their common volcanic origin, the islands have similar coastal features surrounded by small continental shelves, abyssal depths near the coastlines and various seamount habitats (Fernández-Palacios et al., 2023). The great variations in geological age, size, latitudinal location and climate are the main drivers in the difference in the marine species assemblages of each archipelago (Florencio et al., 2021; Freitas et al., 2019; Whittaker, 1998). Moreover, while these archipelagos are relatively similar in terms of habitat composition, the Canary Islands are isolated in featuring extensive seagrass meadows (Barberá et al., 2005; Creed et al., 2016; McKenzie et al., 2020; Schäfer et al., 2021; Bishop et al., 2022a) and being strongly influenced by the Canary Current upwelling system, promoting marine productivity and supporting species diversity (Sambe et al., 2016; Watermeyer et al., 2022). On the other hand, sandy shores are not as ubiquitous in the Azores (Bishop et al., 2022b) while Cabo Verde has relatively few artificial shorelines and submerged structures (Suthers et al., 2022a; Suthers et al., 2022b) and is outside of present-day rhodolith distribution (Fragkopoulou et al., 2021; Keith et al., 2022).
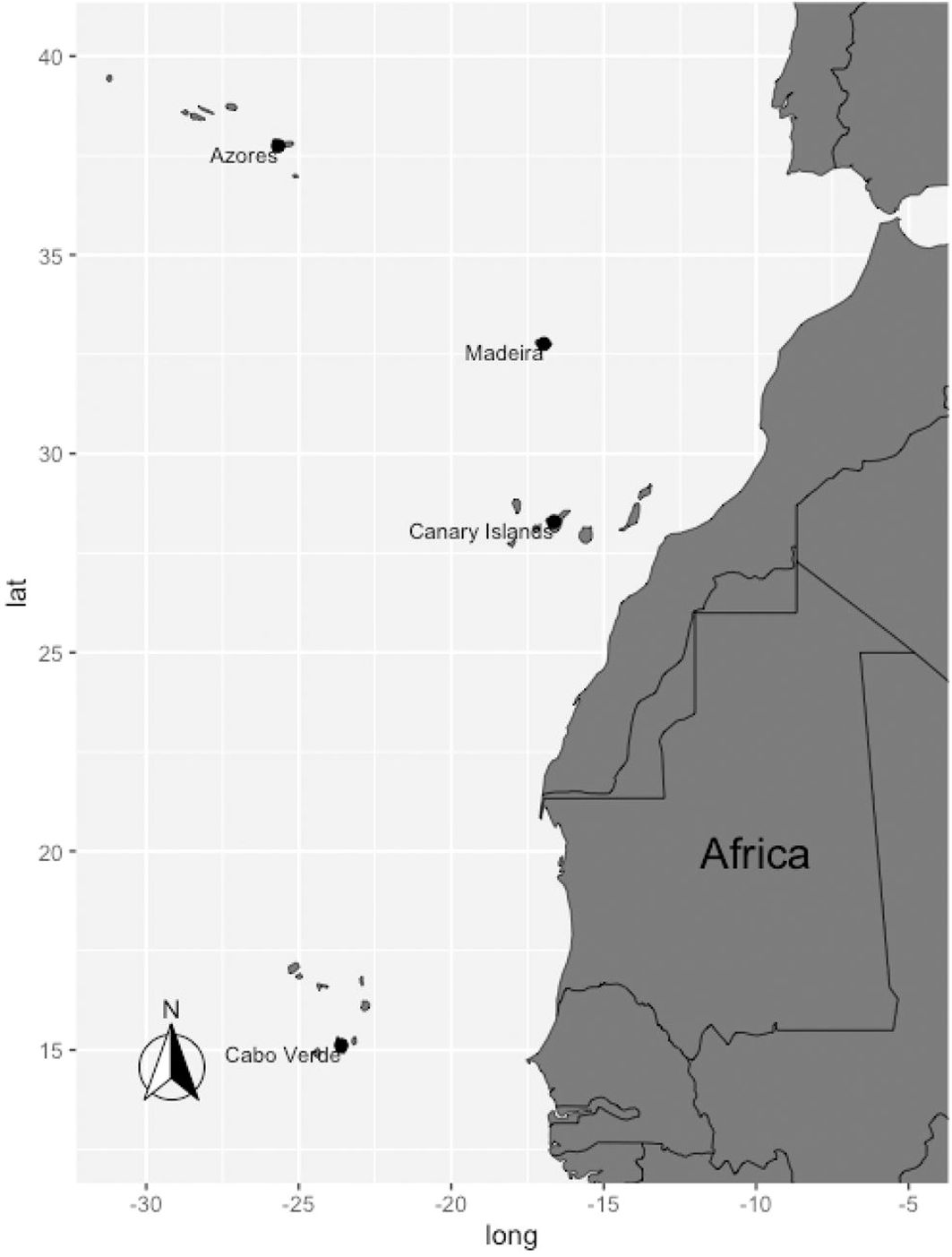
Figure 1. Geographical location of Cabo Verde, Canarias, Madeira and Azores archipelagos.
The Azores is the northernmost archipelago of Macaronesia, composed of nine islands located 1,400 km from Europe (Afonso et al., 2020; Sousa et al., 2021). The Madeira archipelago comprises two main islands located about 840 km southeast of the Azores (Sousa et al., 2021). The Canary Islands comprise seven main islands, located 400 km south of Madeira and 115 km off the west coast of Africa (Meyers et al., 2017; Sousa et al., 2021). Cabo Verde is the southernmost archipelago, consisting of 10 main islands situated around 500 km off the west coast of Africa. All archipelagos are characterised by complex oceanography and topography and are separated by depths of over 1,500 m (Freitas et al., 2019; Sousa et al., 2021). From a marine point of view, the region is mainly influenced by the Azores and Canary currents, along with the Gulf Stream (Barton, 2001; Cropper, 2013).
For Cabo Verde, we used a checklist previously elaborated by one of the authors (RF), as a starting point (Supplementary Table 4). Additions and deletions were made using peer-reviewed scientific publications, books, records from national investigation authorities and official documents. For the Canary archipelago, we retrieved the species from the International Union for Conservation of Nature (IUCN) database (https://www.iucnredlist.org/; last accessed in July 2024) and Ebert et al. (2021) and supplemented them with other scientific peer-reviewed publications (Supplementary Table 3). For the other archipelagos, we used recent checklists published in peer-reviewed journals, namely Santos et al. (2020) for the Azores and Biscoito et al. (2018) for Madeira, and updated them with Ebert et al. (2021) and IUCN database (Supplementary Tables 1, 2). Based on the type of habitat each species is known to use, sharks were categorized as neritic, oceanic and deep-sea. Neritic species use both shore (depth ≤ 30m) and continental shelf (30 m ≤ depth ≤ 200 m) while oceanic are found in offshore pelagic environments at depths ranging from 200 m to 1000 m (Santos et al., 2024b). Sharks found at depths greater than 1000 m were considered deep-sea users, as were benthic species that use environments greater than 200 m (Santos et al., 2024b). We used the IUCN’s map of the geographical range of species to classify each shark as boreal, temperate and tropical. Boreal species use climatic zones at latitudes above 60° north and south, temperate between 30° and 60° north and south while tropical occur between 0° and 30° degrees north and south. We followed Ebert et al. (2021) to assign each species to family level and mode of reproduction (namely lecithotrophic viviparity, matrotrophic viviparity, oviparity). The trophic position of each species was classified as apex predator, mesopredator and planktivore based on its diet and maximum body size (Cortes, 1999; Ebert et al., 2021; Weigmann, 2016). After compiling all the shark species recorded in each archipelago, we built a presence-absence table (Supplementary Tables). We then used a dissimilarity matrix to investigate biogeographical similarities among the archipelagos. To this end, the Jaccard distance metric was applied.
The threats facing each species were collected and adapted from the Threats Classification Scheme of the IUCN (version 3.3, 2024, last accessed in July 2024). For this study, we categorized the threats into 5 groups that best mirror the anthropogenic pressures that sharks are facing in today’s oceans: (i) “Fisheries”, which correspond to IUCN threat 5, more specifically 5.4; (ii) “Pollution”, which correspond to IUCN threat 9; (iii) “Direct human intrusion” witch correspond to IUCN threat 6; (iv) “Climate change” which correspond to IUCN threat 11; and (v) “Habitat degradation” which includes to IUCN threats 1-4, 7,8, and 12. Next, we collected the fishing vulnerability index score from FishBase (www.fishbase.org/, last accessed in November 2024) based on Cheung et al. (2005). It reflects the susceptibility of species to fishing pressure, classified on a scale of 1 to 100, categorized as Low (0-30), Moderate (30-50), High (50-70) and Very High (70-100) (Cheung et al., 2005). The climate change risk of each shark species was taken from a recent global assessment and categorized as Very-low, Low, Medium-low, Medium, Medium-high, High, and Very-high (Santos et al., 2024b). We obtained the FUSE (Functionally Unique, Specialized, and Endangered) values for each species from Pimiento et al. (2023). This conservation metric combines each species’ contribution to functional diversity and its IUCN extinction risk (Pimiento et al., 2020). FUSE index identifies the species with the highest conservation priority whose extinction leads to greater functional loss (Pimiento et al., 2023).
Around the archipelagos of Cabo Verde, Canarias, Madeira and Azores islands, there are 78 species of sharks belonging to 26 families, with Carcharhinidae, Somniosidae, and Centrophoridae as the most speciose (Figure 2). Within the region, the Canary Islands (56) present the highest shark richness, followed by Cabo Verde (53), Madeira (52), and the Azores (45).
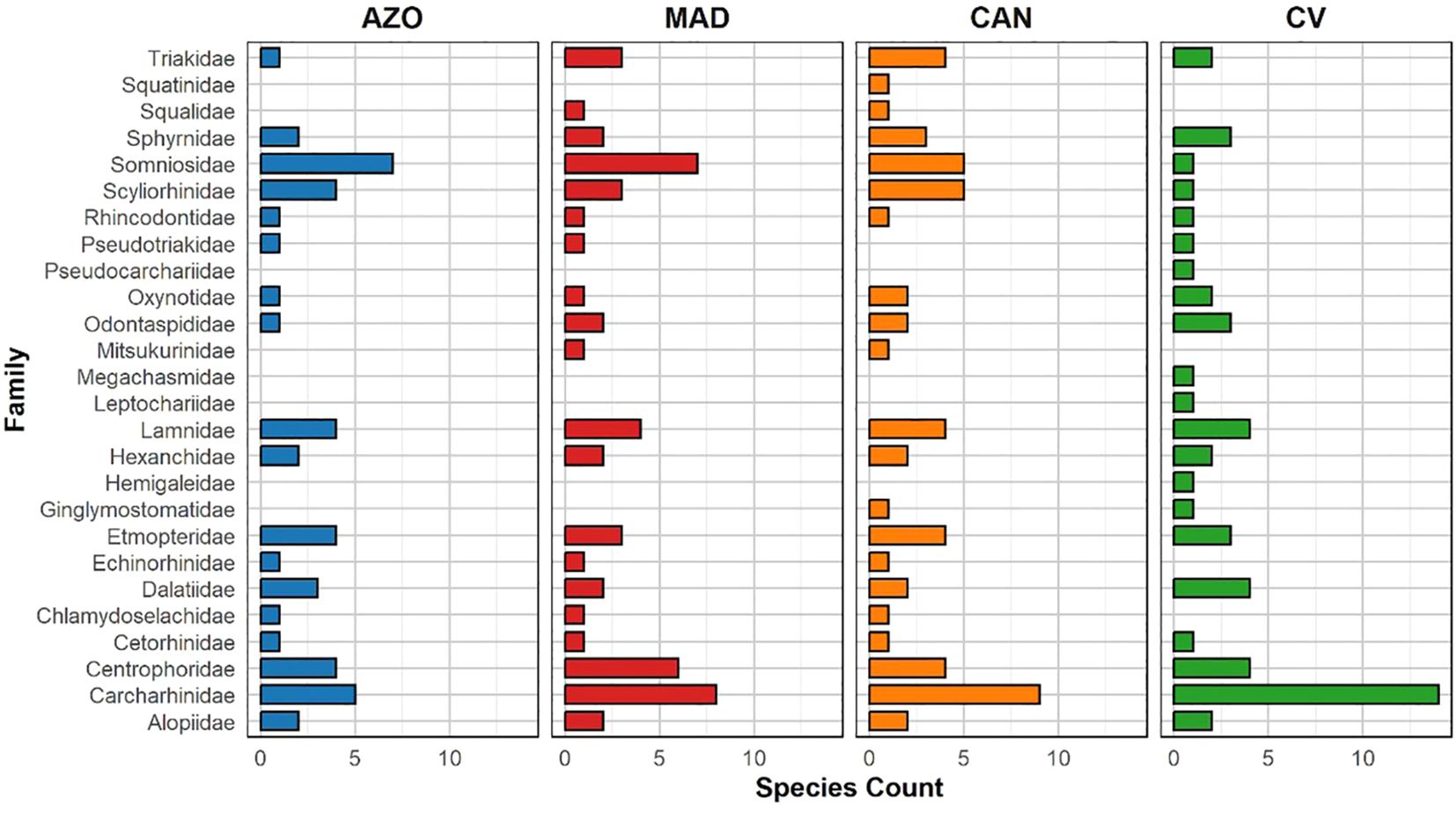
Figure 2. Species richness of sharks, by families, across Cabo Verde, Canarias, Madeira and Azores archipelagos. AZO, Azores; MAD, Madeira; CAN, Canary Islands; CV, Cabo Verde.
Most of the species found in the Azores, Madeira and the Canary Islands prefer temperate zones, while in Cabo Verde the majority prefer tropical climates (Figure 3A). Cabo Verde has fewer species that use boreal zones compared to the other three archipelagos further north (Figure 3A). The majority of species use neritic habitats in all the archipelagos, followed by deep-sea in the Azores, Madeira and the Canary Islands, while in Cabo Verde there is a greater preference for pelagic environments (Figure 3B). The species assemblage of all four archipelagos is dominated by mesopredators, followed by apex predators increasing from the Azores to Cabo Verde (Figure 3C). The three planktivorous species are found around all four archipelagoes, namely the basking shark (Cetorhinus maximus), megamouth shark (Megachasma pelagios), and whale shark (Rhincodon typus). In terms of mode of reproduction, six oviparous species of the family Scyliorhinidae occur in Macaronesia. Cabo Verde is the only archipelago where no oviparous shark species have been recorded, contrary to the Canary Islands (5), the Azores (4), and Madeira (3) (Figure 3D).
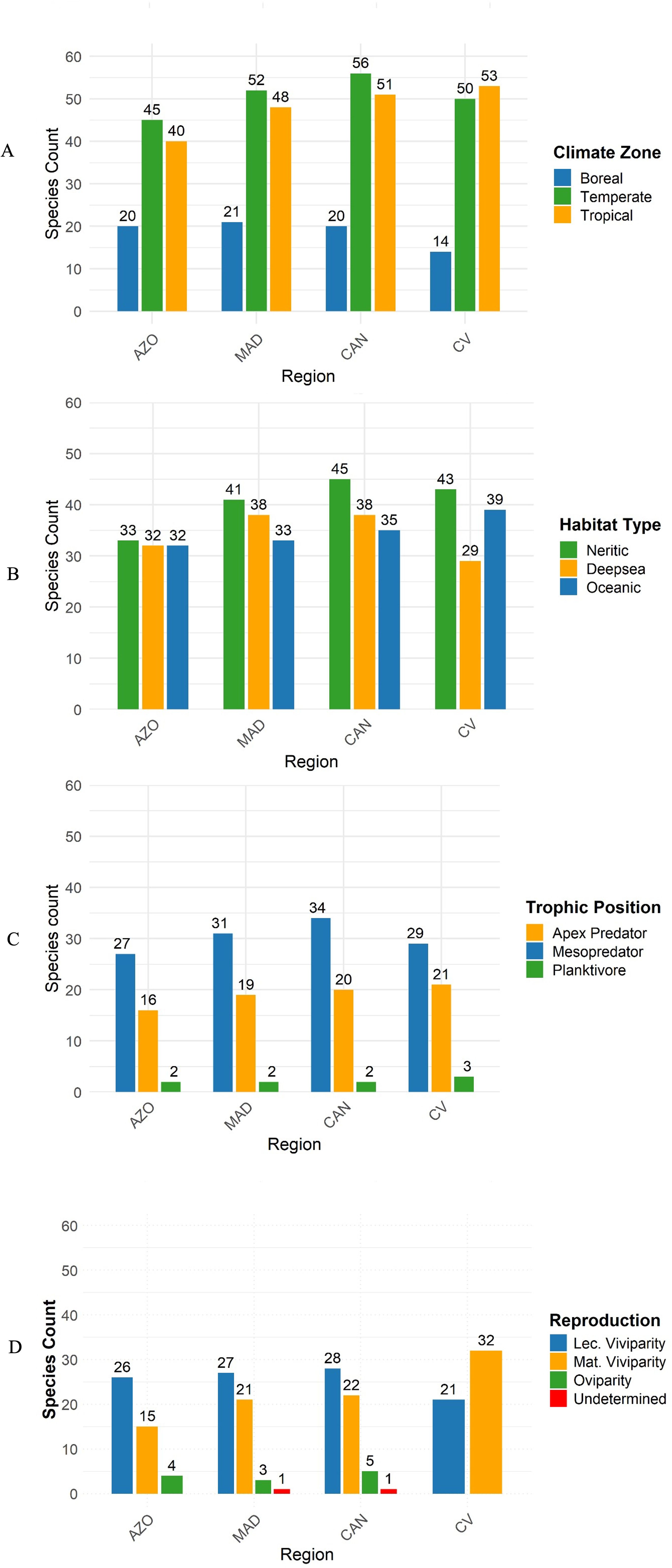
Figure 3. FFrequency of climate zones (A) boreal, temperate, and tropical), habitat types (B) neritic, deep-sea, and oceanic) used, trophic position (C) apex predator, mesopredator and planktivore), and mode of reproduction (D) lecithotrophic viviparity, matrotrophic viviparity, and oviparity) per archipelago. AZO, Azores; MAD, Madeira; CAN, Canary Islands; CV, Cabo Verde.
In terms of similarity, the archipelagos are grouped into 3 clusters (Figure 4A). The Canary Islands and Madeira form the group with the highest number of species in common, dominated by the families Carcharhinidae and Somniosidae, respectively. The Azores form a separate cluster, but they are more closely related to the Canary Islands than to Madeira, despite their greater proximity to the latter. There is a clear separation of Cabo Verde from the rest of the archipelagos, leaving it isolated and dominated by migratory species of the family Carcharhinidae. Cabo Verde has the highest number of exclusive species (10), followed by the Canary Islands (4), the Azores (3) and Madeira (1) (Figure 4B).
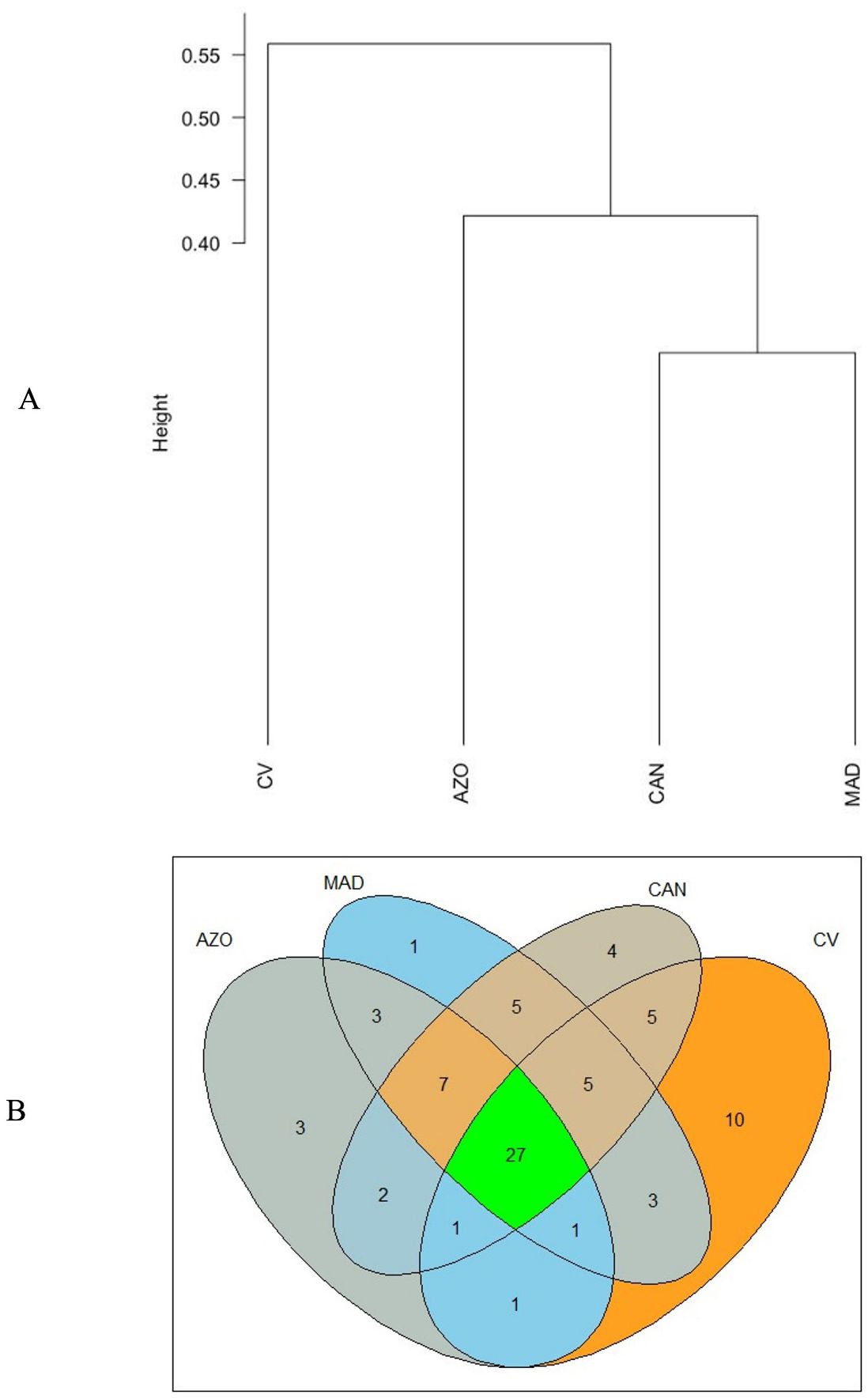
Figure 4. Similarities (A) and number of exclusive shark species (B) in Cabo Verde, Canarias, Madeira and Azores archipelagos. AZO, Azores; MAD, Madeira; CAN, Canary Islands; CV, Cabo Verde.
Of the 78 shark species recorded in Macaronesia and Cabo Verde, 56% are listed in one of the IUCN’s endangered categories as Critically Endangered, Endangered, or Vulnerable (Figure 5A). Carcharhinidae and Lamnidae are the families with the highest number of species at the greatest risk of extinction. Looking at the individual archipelagos, Cabo Verde (66%) has the highest number of endangered species, followed by the Canary Islands (61%), Madeira (52%), and the Azores (49%) (Figure 5A). The four archipelagos share the same top six FUSE species, namely longfin mako (Isurus paucus), shortfin mako (Isurus oxyrinchus), scalloped hammerhead (Sphyrna lewini), basking shark (Cetorhinus maximus), oceanic whitetip shark (Carcharhinus longimanus) and whale shark (Rhincodon typus) (Figure 5B). The remaining four species vary according to archipelago, with the angelshark (Squatina squatina) present only in the Canary Islands with the highest FUSE, followed by great hammerhead (Sphyrna mokarran) and sandtiger shark (Carcharias taurus), both only present in Cabo Verde and the Canary Islands (Figure 5B). Next with the highest FUSE is tope shark (Galeorhinus galeus) (in all archipelagos) followed by sandbar shark (Carcharhinus plumbeus) (Cabo Verde), white shark (Carcharodon carcharias) (Azores and Madeira), rough longnose dogfish (Deania hystricosa) and dusky shark (Carcharhinus obscurus) (Madeira), bramble shark (Echinorhinus brucus) and gulper shark (Centrophorus granulosus) (Azores).
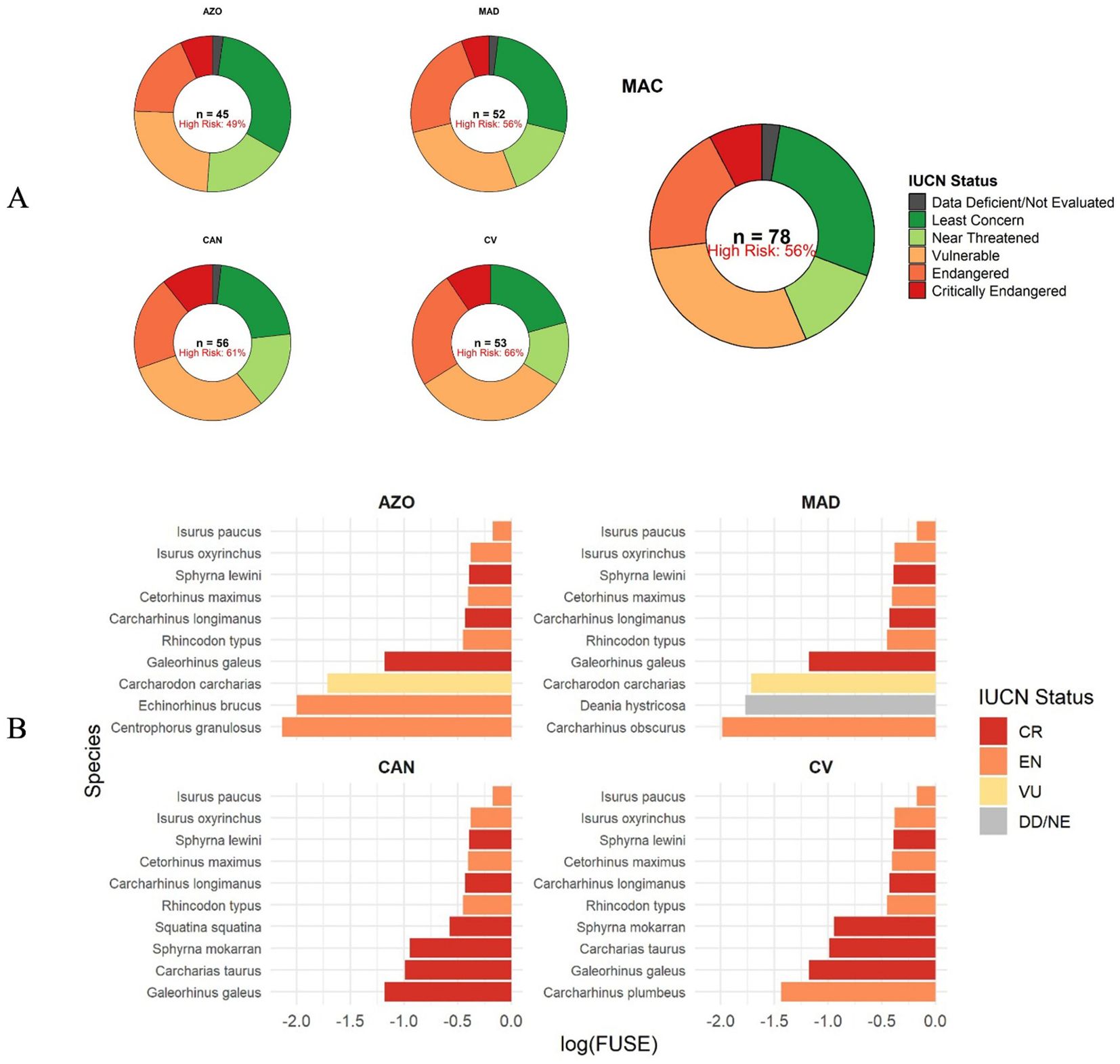
Figure 5. IUCN Red List Status (A) and top ten FUSE (Functionally Unique, Specialized, and Endangered) sharks in Cabo Verde, Canarias, Madeira and Azores archipelagos (B). AZO, Azores; MAD, Madeira; CAN, Canary Islands; CV, Cabo Verde.
Fishing (FIS) is the main threat to sharks in Macaronesia and Cabo Verde and affects all species in the four archipelagos (Figure 6A). Within the region, Cabo Verde has the highest number of species threatened by fishing within one of the IUCN threatened categories (Vulnerable, Endangered, and Critically Endangered) (Figure 6A). Shark species in the Canary Islands are most susceptible to fishing, followed by Cabo Verde, Madeira and the Azores (Figure 6B).
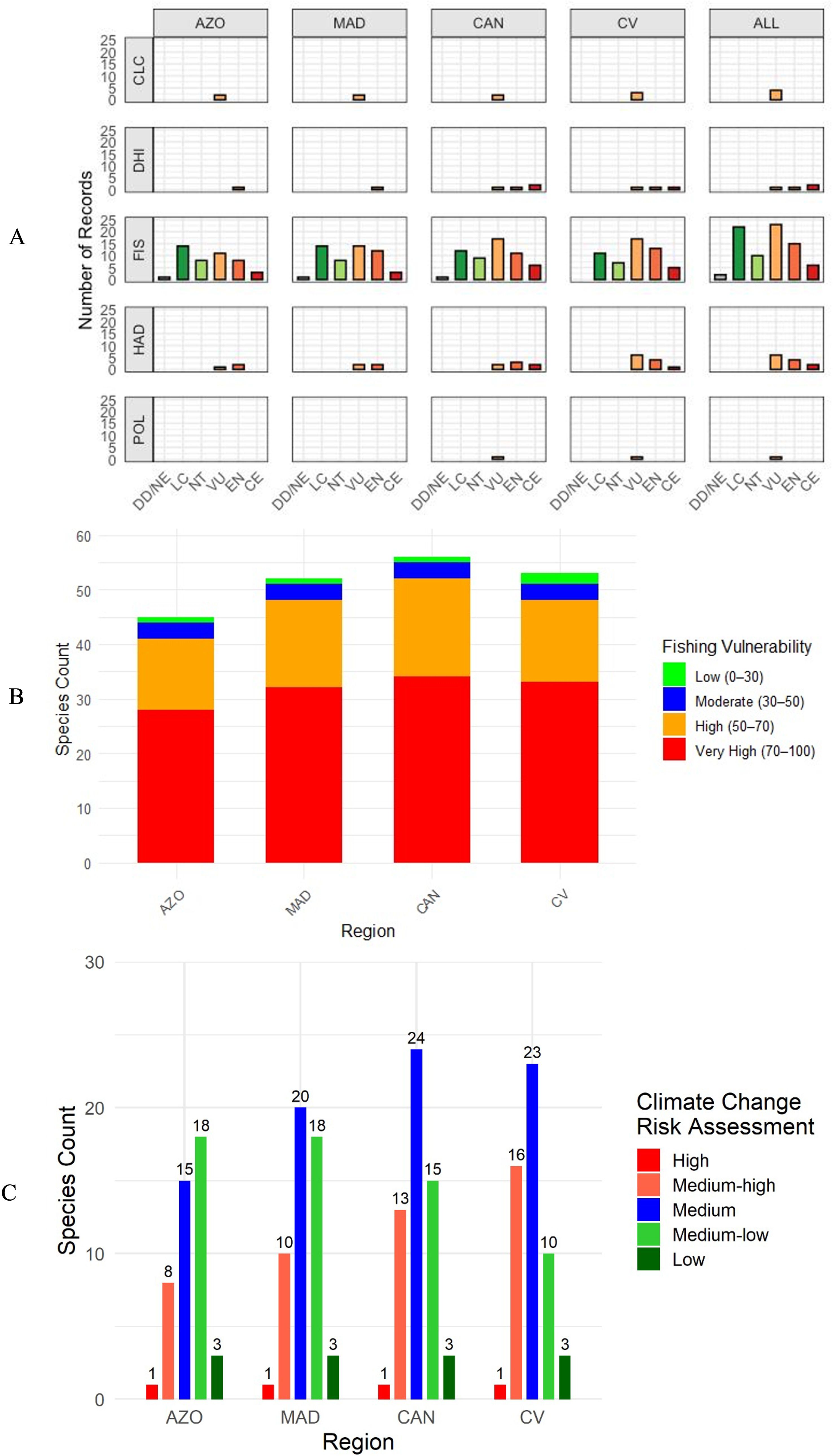
Figure 6. Frequency of shark species affected by each of the main threats per archipelago and their respective risk of extinction according to the IUCN (A), Fishing Vulnerability Index (B), and Climate Change Risk Assessment (C). DD, Data Deficient; NE, Not Evaluated; LC, Least Concern; NT, Near Threatened; VU, Vulnerable; EN, Endangered; CE, Critically Endangered. AZO, Azores; MAD, Madeira; CAN, Canary Islands; CV, Cabo Verde; CLC, Climate Change; DHI, Direct Human Intrusion; FIS, Fisheries; HAD, Habitat Degradation; POL, Pollution.
Regarding habitat degradation, Cabo Verde once again has the highest number of species considered to be affected by this anthropogenic pressure included in the IUCN threatened categories, followed by the Canary Islands (Figure 6A). Four species, namely the bull shark (Carcharhinus leucas), blacktip shark (Carcharhinus limbatus), nurse shark (Ginglymostoma cirratum), and Greenland shark (Somniosus microcephalus) are noted to be affected by climate change according to IUCN (Figure 6A). One species, the oceanic whitetip shark, classified as highly vulnerable to climate change is present in all archipelagos (Figure 6C). Cabo Verde (16) has the highest number of species considered to be at medium-high risk due to climate change, followed by the Canary Islands (13), Madeira (10) and the Azores (8) (Figure 6C). Nurse shark occurs in Cabo Verde and Canary Islands and is the only one considered to be affected by all five threats according to the IUCN. Direct human intrusion affects three endangered species, nurse shark, whale shark, and angel shark (Figure 6A). Taken all together, Cabo Verde and the Canary Islands are the archipelagos where sharks are under the greatest magnitudes and diversity of anthropogenic pressures.
The diversity of marine habitats surrounding the oceanic islands makes them important areas for species assemblages (Pimiento et al., 2023). Together, the archipelagos of Cabo Verde, Canarias, Madeira and Azores islands represent a hotspot of shark diversity, hosting around 15% of all shark species. Within the region, the Canary Islands stand out with the greatest species richness, consistent with a previous study that found a bimodal latitudinal gradient for sharks peaking in temperate region between latitudes 30 and 40 in both hemispheres (Lucifora et al., 2011). A similar latitudinal pattern has previously been observed in other marine taxa such as cephalopods (Rosa et al., 2008, 2019), tuna, billfish, and other fishes associated with optimal temperatures in the mid-latitudes (Boyce et al., 2008; Chaudhary et al., 2016). A global analysis to determine the main factors related to the richness of all existing elasmobranchs found that temperature was the main factor, particularly for sharks, with the Canary Islands standing out as one of diversity hot spot (Guisande et al., 2013). On the other hand, the Canary Islands are the archipelago closest to the mainland, Morocco to be precise, an area of high elasmobranch richness (Guisande et al., 2013). When comparing the richness of sharks among the archipelagos, there is less similarity between Cabo Verde and the rest. This supports the increasing evidence that excludes Cabo Verde from Macaronesia due to significant differences in the composition of marine assemblages. By comparing the differences between six marine groups with different dispersal capacities, Freitas et al. (2019) found less similarity between Cabo Verde and the rest of the archipelagos of Macaronesia, and even suggested that the biogeographic unit should be redefined.
Most of the shared species are oceanic sharks, whose migratory nature gives them greater latitudinal distribution capacity (Camhi et al., 2009; Queiroz et al., 2016). Water temperature is one of the main drivers of elasmobranch migration, with pelagic species preferring warmer tropical waters (Arrowsmith et al., 2021; Lee et al., 2019; Spurgeon et al., 2022). This corroborates our finding with Cabo Verde, the only archipelago located in the tropical region with the highest number of oceanic migratory pelagic species. Archipelagos with greater similarity and geographical proximity share a higher number of less mobile species. However, the Azores share more species with the Canary Islands than Madeira, despite its closer proximity to the latter. In a previous study, a greater similarity was found between the Azores and Madeira when comparing elasmobranch assemblage, and the Canary Islands were clustered with Portugal’s mainland (Das and Afonso, 2017). As deep-sea and non-migratory species are the main contributors to the difference in assemblage composition, the authors emphasize the role of the barriers imposed by the abyssal planes and the seamounts in the region (Das and Afonso, 2017). However, it should be noted here that that study also included rays and chimaeras and had a smaller number of shark species (39) (Das and Afonso, 2017) than the present study (45) for the Azores.
In terms of mode of reproduction, Cabo Verde is the only archipelago where there is no record of oviparous shark species. Six oviparous species, namely Iceland catshark (Apristurus laurussonii), ghost catshark (Apristurus manis), blackmouth catshark (Galeus melastomus), mouse catshark (Galeus murinus), smallspotted catshark (Scyliorhinus canicula) and nursehound (Scyliorhinus stellaris) have been recorded around the temperate archipelagos of Macaronesia. Although this needs further investigation, this trend seems to apply to rays as well. Rays and chimaeras are among the least studied chondrichthyans in the region, particularly in the archipelagos of Madeira and Cabo Verde (McIvor et al., 2022) and were not included in the present study due to the lack of data from these groups to support a solid comparison. Here, we argue that Cabo Verde’s great geographical isolation from the rest of the archipelagos and the mainland in combination with great depths and seamounts serve as a barrier to the colonization of oviparous species in this archipelago. In fact, oviparous sharks are generally small benthic and poor swimmers dependent on continuous habitats, which limit their long migrations (Ebert et al., 2021).
The North Atlantic Ocean is heavily exploited by industrial fishing (Queiroz et al., 2012, 2016). In this region, the longline fishing fleet is mainly made up of European Union countries, particularly Spain, France, the United Kingdom, and Portugal (Oceana, 2009). The fleets from these countries target areas that pelagic sharks use as their preferred habitats (Queiroz et al., 2016). Blue shark and the shortfin mako are reported to be the species most caught by industrial fishing in all the archipelagos of Macaronesia and Cabo Verde (Coelho et al., 2020; Pham et al., 2013). The highly migratory nature of these species increases the chance of interaction with industrial fisheries at high seas, both in targeted fisheries and as bycatch (Torres et al., 2016). The global annual catch of blue sharks has been estimated at 350000 tons, making it the most caught shark species in the world and one of the main elasmobranchs caught in the North Atlantic (Clarke et al., 2006; Rogan and Mackey, 2007). Stock status indicator for blue shark and shortfin mako, including analysis of size frequency distributions and standardized catch-per-unit-of-efforts indexes from pelagic longline fleets fishery of European Union in the Cabo Verde EEZ between 2006 and 2015, was performed by Coelho et al. (2020). They found that the blue shark catch is composed mainly of adults, which can be a sign of a stable population, contrary to shortfin mako, which consists mainly of juveniles (Coelho et al., 2020). The authors argue that, together with a decrease in the mean size of shortfin mako caught by the European Union fleet, it may indicate overfishing of the species in the region (Coelho et al., 2020). As our results show, the species that occur in the Canary Islands and Cabo Verde have a higher vulnerability index to fishing than the Azores and Madeira. This is partly due to the greater number of pelagic species in the Canary Islands and Cabo Verde, favoring their greater interaction with industrial fishing. On the other hand, the overexploitation of species in Cabo Verde (Coelho et al., 2020; Seymour et al., 2024) can be explained by poor governance in insular tropical countries, as noted in a previous study (MacNeil et al., 2020). While in the Azores, Madeira and the Canary Islands operate only their national and EU fleets, Cabo Verde has fishing agreements with several other countries and poor enforcement of fishing regulations in its waters (Carneiro, 2011; Seymour et al., 2024). In addition to the agreement with the European Union, which currently allows access to 56 vessels from Spain, France and Portugal, Cabo Verde also has fishing agreements with Senegal, Mauritania, São Tomé and Principe, Japan and China (Ministério do Mar, 2024). On top of this, Cabo Verde’s national fleet has seen an increase in both artisanal and semi-industrial vessels in recent years (Ministério do Mar, 2024), while the other Macaronesian archipelagos show the opposite trend (Fernández-Palacios et al., 2023).
Marine protected areas are widely recognized as one of the most effective tools for protecting marine habitats and species, including sharks (Albano et al., 2021). Its effectiveness depends on the extent of the protected area and the level of protection (Bonnin et al., 2021; Ferreira et al., 2022; Lara-Lizardi et al., 2022; Murie et al., 2023). The current state of governance of marine protected areas in the Azores offers greater protection to sharks than the archipelagos further south. The Azores has the largest EEZ (approximately 980,000 km2) in Macaronesia and was one of the pioneers in establishing marine protected areas in the 1980s (Abecasis et al., 2015; Maestro et al., 2020). The extent of its marine protected area covers 112,635 km2 (Maestro et al., 2020). More recently, the Azorean regional government passed legislation extending its network of marine protected areas to cover 30% of its EEZ, 15% of which is fully protected and 15% highly protected. Like the archipelagos of Madeira and the Canary Islands, the Azores’ marine protected areas cover a diversity of coastal, open sea, seamounts and deep-sea habitats, offering greater protection for sharks in the region (Afonso et al., 2022; Vandeperre et al., 2014). Madeira and the Canary Islands have an exclusive economic zone (around 450,000 km2) and a marine protected area of approximately the same size of around 12,000 km2. The Selvagens islands of Madeira have one of the largest fully protected marine areas in Europe, covering an area of approximately 2700 km2 (Alves et al., 2022). Cabo Verde has the second largest EEZ (approximately 730,000 km2) and the smallest MPA in the region (only 1321.28 km2) (Sena et al., 2023). In addition, its marine protected areas are all exclusively coastal, established as extensions of terrestrial protected areas, focusing mainly on protecting of sea turtles and seabirds (Seymour et al., 2024).
Habitat degradation is the second most significant pressure on sharks in Macaronesia and Cabo Verde, affecting many species, most of them coastal and all threatened with extinction. This finding is very relevant to the region, given that due to its tourist attractions, there is a growing pressure on coastal areas (Morey et al., 2019). However, Das and Afonso (2017) state that there is no clear evidence of habitat degradation, pollution, or other factors affecting elasmobranchs in the Azores. The angel shark is a critically endangered coastal species found in Canarian waters (Morey et al., 2019; Jiménez‐Alvarado et al., 2020). The rapid development of new infrastructure, accommodation, and facilities to meet the demands of increased tourism is known to potentially increase habitat loss and degradation, water pollution, and altered sediment transport, thus affecting the fitness of the angelshark species in the Canary Islands (Morey et al., 2019; Barker et al., 2016). In Cabo Verde, for example, there has been increased pressure from tourists visiting a potential nursery area of lemon sharks (Negaprion brevirostris), in the island of Sal. This is a very shallow area that favors the presence of people, which can consequently alter the bottom, the behavior of the animals, and potentially their abundance.
Of the 78 shark species that occur in Macaronesia and Cabo Verde, only four are recognized by the IUCN as being affected by climate change. These species are bull shark (Carcharhinus leucas), blacktip shark (Carcharhinus limbatus), nurse shark (Ginglymostoma cirratum), and Greenland shark (Somniosus microcephalus). They are all threatened with extinction and are indirectly affected by climate change through impacts on their habitats, prey, or changes to their range. However, many studies have shown that several other species already suffer some direct or indirect impact from climate change (Santos et al., 2021). In fact, the North Atlantic will experience significant warming, loss of oxygen content (Bates and Johnson, 2020), and changes in salinity and ocean currents (Ditlevsen and Ditlevsen, 2023; Manabe and Stouffer, 1995) depending on the different greenhouse gas emission scenarios. Many pelagic shark species in the North Atlantic are highly migratory (Coelho et al., 2020; Queiroz et al., 2016) and their movements correlate with sea water temperature (Hammerschlag et al., 2022; Vedor et al., 2021a). Evidence shows that sharks may respond to climate change by shifting their latitudinal range or moving to deeper, cooler waters to enhance their physiological processes (Bangley et al., 2018; Crear et al., 2023). In the context of climate change, the tropical region (where many species live at their thermal limit) will lose habitat suitability while the higher latitudes will gain for many sharks (Diaz-Carballido et al., 2022; Rodriguez-Burgos et al., 2022; Santos et al., 2024a). Cabo Verde has fewer species that use boreal and temperate zones, compared to the other archipelagos, which makes it more vulnerable to climate change, and therefore with more species under medium-high risk (Santos et al., 2024b). On the other hand, in the archipelagos with higher latitudes, species have a greater latitudinal range and there is a gain in habitat suitability. For example, in recent years there has been an increase in whale shark sightings in the Azores associated with increasingly warmer water in the region (Afonso et al., 2014). Hammerschlag et al. (2022) used combined analysis of animal tracking, remotely sensed environmental data, habitat modeling, and capture data to investigate the impacts of climate variability and change on the distributional range and migratory phenology of tiger shark (Galeocerdo cuvier) in the North Atlantic. They found that tiger sharks extended their migrations farther poleward, arriving earlier in the northern latitude in response to anomalously high sea-surface temperature (Hammerschlag et al., 2022). In the same study, they analyzed about four decades of tiger shark captures in the region and found that areas of highest catch densities have continuously increased towards the north, and catches are occurring earlier (Hammerschlag et al., 2022). In another study, researchers using satellite tagging of blue sharks and environmental modelling in the oxygen minimum zone around Cabo Verde’s Exclusive Economic Zone, showed that sharks are pushed to the surface and thus potentially increase the chance of interaction with industrial fishing and their capture (Vedor et al., 2021b).
Considering FUSE, we found that all four archipelagos share the same top six priority sharks for protection. In addition to all being listed as endangered or critically endangered, they all exhibit very high vulnerability to fishing. With the exception of basking shark and scalloped hammerhead, which are at medium risk from climate change, longfin mako, shortfin mako and whale shark are at medium-high risk, while oceanic whitetip is at high risk (Santos et al., 2024b). The critically endangered Galeorhinus galeus, found throughout the archipelagos, is also a species of conservation concern because of its high FUSE value. Due to their highly migratory nature, the protection of species in the region requires a joint effort between the archipelagos to ensure effective species conservation. This finding supports the idea previously suggested of the need to declare a marine corridor to protect elasmobranchs in the region (Tuya et al., 2022). A shared marine protected area in the region would protect not only sharks but also other taxa such as sea turtles and cetaceans that use the region for their migratory routes.
This study shows that Cabo Verde, Canarias, Madeira and Azores archipelagos faces similar challenges for shark conservation. However, the archipelago of Cabo Verde presents greater challenges due to the existence of a greater number of threatened species and fragility due to poor monitoring, conservation and research. Faced with these challenges, better control of the species caught in its waters is essential to ensure that only species permitted under the agreements are caught and to collect scientific data that can be used for decision-making. The next reviews of laws and management measures should look at other species (e.g. sand tiger shark) that are not migratory but are highly impacted by local fisheries (e.g. nurse shark). Local fisheries should also be the target of scientific studies, better involvement in the decision-making and management measures, as well as training in the identification of prohibited species and release mechanisms. There is an urgent need to expand and create more marine protected areas (both coastal and pelagic environments) in Cabo Verde that guarantee the protection of critical areas, such as Sal Rei Bay, Boa Vista Island, the first shark nursery described in Cabo Verde and the only one of multiple species in the whole of Atlantic Africa (Rosa et al., 2023). While for Cabo Verde it is essential to boost research to define new science-based marine protected areas to ensure the inclusion of key habitats and endangered species, for the Azores, Madeira and the Canary Islands their main priorities are effective management and enforcement of their waters.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
JV: Conceptualization, Investigation, Writing – original draft, Writing – review & editing. CS: Conceptualization, Methodology, Writing – review & editing. EN: Formal analysis, Writing – review & editing. VP: Writing – review & editing. SP: Writing – review & editing. BR: Data curation, Investigation, Methodology, Validation, Visualization, Writing – review & editing. EV: Data curation, Investigation, Methodology, Validation, Visualization, Writing – review & editing. TR: Writing – review & editing. NQ: Data curation, Investigation, Methodology, Validation, Visualization, Writing – review & editing. RF: Writing – review & editing. RR: Conceptualization, Funding acquisition, Methodology, Supervision, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research is part of project NGANDU funded by the Portuguese Foundation for Science and Technology (FCT) and the Aga Khan Development Network (AKDN) under grant agreement FCT AGA-KHAN/541746579/2019. The first author would like to thank the Prince Albert II of Monaco Foundation, Intergovernmental Panel on Climate Change (IPCC), and Camões-Instituto da Cooperação e da Língua, I.P. All authors acknowledge funding from FCT under the strategic project UIDB/04292/2020 granted to MARE, project LA/P/0069/2020 granted to the Associate Laboratory ARNET. FCT further supports this work through PhD grants to CS (SFRH/BD/117890/2016 and project NGANDU). EN acknowledges funding from FCT PhD grant SFR/BD/135438/2017. TR was funded through an FCT researcher contract (DL57/2016/CP1479/CT0023).
The authors declare that the research was conducted without any commercial or financial relationships that could potentially create a conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1490317/full#supplementary-material
Abecasis R. C., Afonso P., Colaço A., Longnecker N., Clifton J., Schmidt L., et al. (2015). Marine conservation in the azores: evaluating marine protected area development in a remote island context. Front. Mar. Sci. 2. doi: 10.3389/fmars.2015.00104
Afonso P., Fontes J., Giacomello E., Magalhães M. C., Martins H. R., Morato T., et al. (2020). The azores: A mid-atlantic hotspot for marine megafauna research and conservation. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00826
Afonso P., Gandra M., Graça G., Macena B., Vandeperre F., Fontes J. (2022). The multi-annual residency of juvenile smooth hammerhead shark in an oceanic island nursery. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.844893
Afonso P., McGinty N., Machete M. (2014). Dynamics of whale shark occurrence at their fringe oceanic habitat. PloS One 9, e102060. doi: 10.1371/journal.pone.0102060
Albano P. S., Fallows C., Fallows M., Schuitema O., Bernard A. T. F., Sedgwick O., et al. (2021). Successful parks for sharks: No-take marine reserve provides conservation benefits to endemic and threatened sharks off South Africa. Biol. Conserv. 261, 109302. doi: 10.1016/j.biocon.2021.109302
Alves F., Monteiro J. G., Oliveira P., Canning-Clode J. (2022). Portugal leads with Europe’s largest marine reserve. Nature 601, 318–318. doi: 10.1038/d41586-022-00093-8
Arrowsmith L., Sequeira A., Pattiaratchi C., Meekan M. (2021). Water temperature is a key driver of horizontal and vertical movements of an ocean giant, the whale shark Rhincodon typus. Mar. Ecol. Prog. Ser. 679, 101–114. doi: 10.3354/meps13899
Bangley C. W., Paramore L., Shiffman D. S., Rulifson R. A. (2018). Increased abundance and nursery habitat use of the bull shark (Carcharhinus leucas) in response to a changing environment in a warm-temperate estuary. Sci. Rep. 8, 6018. doi: 10.1038/s41598-018-24510-z
Barberá C., Tuya F., Boyra A., Sanchez-Jerez P., Blanch I., Haroun R. J. (2005). Spatial variation in the structural parameters of Cymodocea nodosa seagrass meadows in the Canary Islands: A multiscaled approach. Botanica Marina 48, 122–126. doi: 10.1515/BOT.2005.021
Barker J., Bartolí Á., Clark M., Dulvy N., Gordon C., Hood A., et al. (2016). Angelshark Action Plan for the Canary Islands (London, UK: Zoological Society of London).
Barton E. D. (2001). “Canary and Portugal Currents,” in Encyclopedia of Ocean Sciences (San Diego, USA: Elsevier), 330–339. doi: 10.1016/B978-0-12-813081-0.00360-8
Bates N. R., Johnson R. J. (2020). Acceleration of ocean warming, salinification, deoxygenation and acidification in the surface subtropical North Atlantic Ocean. Commun. Earth Environ. 1, 33. doi: 10.1038/s43247-020-00030-5
Biscoito M., Ribeiro C., Freitas M. (2018). Annotated checklist of the fishes of the archipelago of Madeira (NE Atlantic): I—Chondrichthyes. Zootaxa 4429, 459–494. doi: 10.11646/zootaxa.4429.3.2
Bishop M. J., Altieri A. H., Porter S. N., Orth R. J., Keith D. A. (2022a). M1.1 Seagrass meadows. In: Keith D.A., Ferrer-Paris J.R., Nicholson E., et al. (2022). A function-based typology for Earth’s ecosystems – Appendix S4. The IUCN Global Ecosystem Typology v2.1: Descriptive profiles for Biomes and Ecosystem Functional Groups. Nature 610, 1295 513–518. doi: 10.1038/s41586-022-05318-4
Bishop M. J., McSweeney S. L., Keith D. A. (2022b). MT1.3 Sandy Shorelines. In: Keith D. A., Ferrer-Paris J.R., Nicholson E., et al (2022). A function-based typology for Earth’s ecosystems – Appendix S4. The IUCN Global Ecosystem Typology v2.1: Descriptive profiles for Biomes and Ecosystem Functional Groups. Nature 610, 1295 513–518. doi: 10.1038/s41586-022-05318-4
Bonnin L., Mouillot D., Boussarie G., Robbins W. D., Kiszka J. J., Dagorn L., et al. (2021). Recent expansion of marine protected areas matches with home range of grey reef sharks. Sci. Rep. 11, 14221. doi: 10.1038/s41598-021-93426-y
Boyce D., Tittensor D., Worm B. (2008). Effects of temperature on global patterns of tuna and billfish richness. Mar. Ecol. Prog. Ser. 355, 267–276. doi: 10.3354/meps07237
Camhi M. D., Lauck E., Pikitch E. K., Babcock E. A. (2008). “A Global Overview of Commercial Fisheries for Open Ocean Sharks,” in Sharks of the Open Ocean, 1st ed. Eds. Camhi M. D., Pikitch E. K., Babcock E. A. (Oxford, UK: Wiley), 166–192. doi: 10.1002/9781444302516.ch14
Camhi M. D., Valenti S. V., Fordham S. V., Fowler S. L., Gibson C. (2009). The conservation status of pelagic sharks and rays: Report of the IUCN Shark Specialist Group: Pelagic Shark Red List Workshop, Tubney House, University of Oxford, UK, 19–23 February 2007 (IUCN Species Survival Commission’s Shark Specialist Group).
Carneiro G. (2011). They come, they fish, and they go:“ EC fisheries agreements with cape verde and são tomé e príncipe. Mar. Fisheries Review 73, 1–25.
Chaudhary C., Saeedi H., Costello M. J. (2016). Bimodality of latitudinal gradients in marine species richness. Trends Ecol. Evol. 31, 670–676. doi: 10.1016/j.tree.2016.06.001
Cheung W. W. L., Pitcher T. J., Pauly D. (2005). A fuzzy logic expert system to estimate intrinsic extinction vulnerabilities of marine fishes to fishing. Biol. Conserv. 124, 97–111. doi: 10.1016/j.biocon.2005.01.017
Clarke S. C., McAllister M. K., Milner-Gulland E. J., Kirkwood G. P., Michielsens C. G. J., Agnew D. J., et al. (2006). Global estimates of shark catches using trade records from commercial markets. Ecol. Lett. 9, 1115–1126. doi: 10.1111/j.1461-0248.2006.00968.x
Coelho R., Macías D., Ortiz De Urbina J., Martins A., Monteiro C., Lino P. G., et al. (2020). Local indicators for global species: Pelagic sharks in the tropical northeast Atlantic, Cabo Verde islands region. Ecol. Indic. 110, 105942. doi: 10.1016/j.ecolind.2019.105942
Cortes E. (1999). Standardized diet compositions and trophic levels of sharks. ICES J. Mar. Sci. 56, 707–717. doi: 10.1006/jmsc.1999.0489
Crear D. P., Curtis T. H., Hutt C. P., Lee Y. (2023). Climate-influenced shifts in a highly migratory species recreational fishery. Fisheries Oceanography 32, 327–340. doi: 10.1111/fog.12632
Creed J. C., Engelen A. H., D'Oliveira E. C., Bandeira S., Serrão E. A. (2016). First record of seagrass in Cape Verde, eastern Atlantic. Mar. Biodiversity Records 9, 57. doi: 10.1186/s41200-016-0067-9
Cropper T. (2013). The weather and climate of Macaronesia: Past, present and future. Weather 68, 300–307. doi: 10.1002/wea.2155
Das D., Afonso P. (2017). Review of the diversity, ecology, and conservation of elasmobranchs in the azores region, Mid-North Atlantic. Front. Mar. Sci. 4. doi: 10.3389/fmars.2017.00354
Davidson L. N. K., Krawchuk M. A., Dulvy N. K. (2016). Why have global shark and ray landings declined: Improved management or overfishing? Fish Fisheries 17, 438–458. doi: 10.1111/faf.12119
Diaz-Carballido P. L., Mendoza-González G., Yañez-Arenas C. A., Chiappa-Carrara X. (2022). Evaluation of shifts in the potential future distributions of carcharhinid sharks under different climate change scenarios. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.745501
Ditlevsen P., Ditlevsen S. (2023). Warning of a forthcoming collapse of the Atlantic meridional overturning circulation. Nat. Commun. 14, 4254. doi: 10.1038/s41467-023-39810-w
Dulvy N. K., Fowler S. L., Musick J. A., Cavanagh R. D., Kyne P. M., Harrison L. R., et al. (2014). Extinction risk and conservation of the world’s sharks and rays. eLife 3, e00590. doi: 10.7554/eLife.00590
Dulvy N. K., Pacoureau N., Rigby C. L., Pollom R. A., Jabado R. W., Ebert D. A., et al. (2021). Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Curr. Biol. 31, 4773–4787.e8. doi: 10.1016/j.cub.2021.08.062
Dulvy N. K., Simpfendorfer C. A., Davidson L. N. K., Fordham S. V., Bräutigam A., Sant G., et al. (2017). Challenges and priorities in shark and ray conservation. Curr. Biol. 27, R565–R572. doi: 10.1016/j.cub.2017.04.038
Ebert D. A., Dando M., Fowler S. L. (2021). Sharks of the world: A complete guide (New Jersey, USA: Princeton University Press).
Fernández-Palacios J. M., De Nascimento L., Otto R., Delgado J. D., García-del-Rey E., Arévalo J. R., et al. (2011). A reconstruction of Palaeo-Macaronesia, with particular reference to the long-term biogeography of the Atlantic island laurel forests: Palaeo-Macaronesia and the Atlantic island laurel forests. J. Biogeography 38, 226–246. doi: 10.1111/j.1365-2699.2010.02427.x
Fernández-Palacios Y., Kaushik S., Abramic A., Cordero-Penín V., García-Mendoza A., Bilbao-Sieyro A., et al. (2023). Status and perspectives of blue economy sectors across the Macaronesian archipelagos. J. Coast. Conserv. 27, 39. doi: 10.1007/s11852-023-00961-z
Ferreira H. M., Magris R. A., Floeter S. R., Ferreira C. E. L. (2022). Drivers of ecological effectiveness of marine protected areas: A meta-analytic approach from the Southwestern Atlantic Ocean (Brazil). J. Environ. Manage. 301, 113889. doi: 10.1016/j.jenvman.2021.113889
Ferretti F., Worm B., Britten G. L., Heithaus M. R., Lotze H. K. (2010). Patterns and ecosystem consequences of shark declines in the ocean. Ecol. Lett. 13, 1055–1071. doi: 10.1111/j.1461-0248.2010.01489.x
Florencio M., Patiño J., Nogué S., Traveset A., Borges P. A. V., Schaefer H., et al. (2021). Macaronesia as a fruitful arena for ecology, evolution, and conservation biology. Front. Ecol. Evol. 9. doi: 10.3389/fevo.2021.718169
Fragkopoulou E., Serrão E. A., Horta P. A., Koerich G., Assis J. (2021). Bottom trawling threatens future climate refugia of rhodoliths globally. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.594537
Freitas R., Romeiras M., Silva L., Cordeiro R., Madeira P., González J. A., et al. (2019). Restructuring of the ‘Macaronesia’ biogeographic unit: A marine multi-taxon biogeographical approach. Sci. Rep. 9, 15792. doi: 10.1038/s41598-019-51786-6
Frisk M. G., Miller T. J., Fogarty M. J. (2001). Estimation and analysis of biological parameters in elasmobranch fishes: A comparative life history study. Can. J. Fisheries Aquat. Sci. 58, 969–981. doi: 10.1139/f01-051
Guisande C., Patti B., Vaamonde A., Manjarrés-Hernández A., Pelayo-Villamil P., García-Roselló E., et al. (2013). Factors affecting species richness of marine elasmobranchs. Biodiversity Conserv. 22, 1703–1714. doi: 10.1007/s10531-013-0507-3
Hammerschlag N., McDonnell L. H., Rider M. J., Street G. M., Hazen E. L., Natanson L. J., et al. (2022). Ocean warming alters the distributional range, migratory timing, and spatial protections of an apex predator, the tiger shark (Galeocerdo cuvier). Global Change Biol. 28, 1990–2005. doi: 10.1111/gcb.16045
Illera J. C., Rando J. C., Richardson D. S., Emerson B. C. (2012). Age, origins and extinctions of the avifauna of Macaronesia: A synthesis of phylogenetic and fossil information. Quaternary Sci. Rev. 50, 14–22. doi: 10.1016/j.quascirev.2012.07.013
Jackson J. B. C., Kirby M. X., Berger W. H., Bjorndal K. A., Botsford L. W., Bourque B. J., et al. (2001). Historical overfishing and the recent collapse of coastal ecosystems. Science 293, 629–637. doi: 10.1126/science.1059199
Jiménez-Alvarado D., Meyers E. K. M., Caro M. B., Sealey M. J., Barker J. (2020). Investigation of juvenile angelshark ( Squatina squatina ) habitat in the Canary Islands with recommended measures for protection and management. Aquat. Conservation: Mar. Freshw. Ecosyst. 30, 2019–2025. doi: 10.1002/aqc.3337
Keith D. A., Ferrer-Paris J. R., Nicholson E., Bishop M. J., Polidoro B. A., Ramirez-Llodra E., et al. (2022). A function-based typology for Earth’s ecosystems. Nature 610, 513–518. doi: 10.1038/s41586-022-05318-4
Lara-Lizardi F., Hoyos-Padilla E. M., Klimley A. P., Grau M., Ketchum J. T. (2022). Movement patterns and residency of bull sharks, Carcharhinus leucas, in a marine protected area of the Gulf of California. Environ. Biol. Fishes 105, 1765–1779. doi: 10.1007/s10641-022-01223-x
Lee K., Smoothey A., Harcourt R., Roughan M., Butcher P., Peddemors V. (2019). Environmental drivers of abundance and residency of a large migratory shark, Carcharhinus leucas, inshore of a dynamic western boundary current. Mar. Ecol. Prog. Ser. 622, 121–137. doi: 10.3354/meps13052
Lucifora L. O., García V. B., Worm B. (2011). Global diversity hotspots and conservation priorities for sharks. PloS One 6, e19356. doi: 10.1371/journal.pone.0019356
MacNeil M. A., Chapman D. D., Heupel M., Simpfendorfer C. A., Heithaus M., Meekan M., et al. (2020). Global status and conservation potential of reef sharks. Nature 583, 801–806. doi: 10.1038/s41586-020-2519-y
Maestro M., Chica-Ruiz J. A., Pérez-Cayeiro M. L. (2020). Analysis of marine protected area management: The Marine Park of the Azores (Portugal). Mar. Policy 119, 104104. doi: 10.1016/j.marpol.2020.104104
Manabe S., Stouffer R. J. (1995). Simulation of abrupt climate change induced by freshwater input to the North Atlantic Ocean. Nature 378, 165–167. doi: 10.1038/378165a0
Marco A., Abella E., Liria-Loza A., Martins S., López O., Jiménez-Bordón S., et al. (2012). Abundance and exploitation of loggerhead turtles nesting in Boa Vista island, Cape Verde: The only substantial rookery in the eastern Atlantic: Loggerhead turtles of Cape Verde. Anim. Conserv. 15, 351–360. doi: 10.1111/j.1469-1795.2012.00547.x
McIvor A. J., Williams C. T., Alves F., Dinis A., Pais M. P., Canning-Clode J. (2022). The status of marine megafauna research in macaronesia: A systematic review. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.819581
McKenzie L. J., Nordlund L. M., Jones B. L., Cullen-Unsworth L. C., Roelfsema C., Unsworth R. K. F. (2020). The global distribution of seagrass meadows. Environ. Res. Lett. 15, 074041. doi: 10.1088/1748-9326/ab7d06
Meyers E. K. M., Tuya F., Barker J., Jiménez Alvarado D., Castro-Hernández J. J., Haroun R., et al. (2017). Population structure, distribution and habitat use of the Critically Endangered Angelshark, Squatina squatina, in the Canary Islands. Aquat. Conservation: Mar. Freshw. Ecosyst. 27, 1133–1144. doi: 10.1002/aqc.2769
Ministério do Mar (2024). Relatório Anual FiTI pelo Grupo Multissetorial Nacional (GMN) de Cabo Verde Verde à Iniciativa de Transparência nas Pescas. (Mindelo, Cabo Verde: Ministério do Mar).
Morey G., Barker J., Hood A., Gordon C., Bartolí A., Meyers E.K.M., et al (2019). Squatina squatina. The IUCN Red List of Threatened Species 2019: e.T39332A117498371. doi: 10.2305/IUCN.UK.2019-1.RLTS.T39332A117498371.en (Accessed January 23, 2025).
Murie C. J. G., Lebrato M., Lawrence A., Brown J., Gavard L., Bowles K. R., et al. (2023). A Mozambican marine protected area provides important habitat for vulnerable pelagic sharks. Sci. Rep. 13, 6454. doi: 10.1038/s41598-023-32407-9
Myers R. A., Baum J. K., Shepherd T. D., Powers S. P., Peterson C. H. (2007). Cascading effects of the loss of apex predatory sharks from a coastal ocean. Science 315, 1846–1850. doi: 10.1126/science.1138657
Myers R. A., Worm B. (2005). Extinction, survival or recovery of large predatory fishes. Philos. Trans. R. Soc. B: Biol. Sci. 360, 13–20. doi: 10.1098/rstb.2004.1573
Oceana F. (2009). Description of the European Union surface longline fleet operating in the Atlantic Ocean and compilation of detailed EUROSTAT data on shark catches by EU fleets in the Atlantic. ICCAT Coll. Vol. Sci. Pap 64, 1746–1754.
Pacoureau N., Rigby C. L., Kyne P. M., Sherley R. B., Winker H., Carlson J. K., et al. (2021). Half a century of global decline in oceanic sharks and rays. Nature 589, 567–571. doi: 10.1038/s41586-020-03173-9
Pham C. K., Canha A., Diogo H., Pereira J. G., Prieto R., Morato T. (2013). Total marine fishery catch for the Azores, (1950-2010). ICES J. Mar. Sci. 70, 564–577. doi: 10.1093/icesjms/fst024
Pimiento C., Albouy C., Silvestro D., Mouton T. L., Velez L., Mouillot D., et al. (2023). Functional diversity of sharks and rays is highly vulnerable and supported by unique species and locations worldwide. Nat. Commun. 14, 7691. doi: 10.1038/s41467-023-43212-3
Pimiento C., Leprieur F., Silvestro D., Lefcheck J. S., Albouy C., Rasher D. B., et al. (2020). Functional diversity of marine megafauna in the Anthropocene. Sci. Adv. 6, eaay7650. doi: 10.1126/sciadv.aay7650
Queiroz N., Humphries N. E., Mucientes G., Hammerschlag N., Lima F. P., Scales K. L., et al. (2016). Ocean-wide tracking of pelagic sharks reveals extent of overlap with longline fishing hotspots. Proc. Natl. Acad. Sci. 113, 1582–1587. doi: 10.1073/pnas.1510090113
Queiroz N., Humphries N. E., Noble L. R., Santos A. M., Sims D. W. (2012). Spatial dynamics and expanded vertical niche of blue sharks in oceanographic fronts reveal habitat targets for conservation. PloS One 7, e32374. doi: 10.1371/journal.pone.0032374
Reverdin G., Niiler P. P., Valdimarsson H. (2003). orth Atlantic Ocean surface currents. J. Geophysical Research: Oceans 108, 2-1-2-21. doi: 10.1029/2001JC001020
Roberts C. M., McClean C. J., Veron J. E. N., Hawkins J. P., Allen G. R., McAllister D. E., et al. (2002). Marine biodiversity hotspots and conservation priorities for tropical reefs. Science 295, 1280–1284. doi: 10.1126/science.1067728
Rodriguez-Burgos A. M., Briceño-Zuluaga F. J., Ávila Jiménez J. L., Hearn A., Peñaherrera-Palma C., Espinoza E., et al. (2022). The impact of climate change on the distribution of Sphyrna lewini in the tropical eastern Pacific. Mar. Environ. Res. 180, 105696. doi: 10.1016/j.marenvres.2022.105696
Roff G., Brown C. J., Priest M. A., Mumby P. J. (2018). Decline of coastal apex shark populations over the past half century. Commun. Biol. 1, 223. doi: 10.1038/s42003-018-0233-1
Rogan E., Mackey M. (2007). Megafauna bycatch in drift nets for albacore tuna (Thunnus alalunga) in the NE Atlantic. Fisheries Res. 86, 6–14. doi: 10.1016/j.fishres.2007.02.013
Rosa R., Dierssen H. M., Gonzalez L., Seibel B. A. (2008). Large-scale diversity patterns of cephalopods in the atlantic open ocean and deep sea. Ecology 89, 3449–3461. doi: 10.1890/08-0638.1
Rosa R., Nunes E., Pissarra V., Santos C. P., Varela J., Baptista M., et al. (2023). Evidence for the first multi-species shark nursery area in Atlantic Africa (Boa Vista Island, Cabo Verde). Front. Mar. Sci. 10. doi: 10.3389/fmars.2023.1077748
Rosa R., Pissarra V., Borges F. O., Xavier J., Gleadall I. G., Golikov A., et al. (2019). Global patterns of species richness in coastal cephalopods. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00469
Sambe B., Tandstad M., Caramelo A. M., Brown B. E. (2016). Variations in productivity of the Canary Current Large Marine Ecosystem and their effects on small pelagic fish stocks. Environ. Dev. 17, 105–117. doi: 10.1016/j.envdev.2015.11.012
Santos C. P., Borges F. O., Guerreiro M., Pissarra V., Varela J., Frazão-Santos C., et al. (2024a). Shifts in the habitat suitability for large hammerhead sharks under climate change. Mar. Biol. 171, 226. doi: 10.1007/s00227-024-04512-4
Santos R., Novoa-Pabon A., Silva H., Pinho M. (2020). Elasmobranch species richness, fisheries, abundance and size composition in the Azores archipelago (NE Atlantic). Mar. Biol. Res. 16, 103–116. doi: 10.1080/17451000.2020.1718713
Santos C. P., Rosa R., Frazão-Santos C. (2024b). Global risk assessment of sharks to climate change. Sci. Total Environ. 954, 176361. doi: 10.1016/j.scitotenv.2024.176361
Santos C. P., Sampaio E., Pereira B. P., Pegado M. R., Borges F. O., Wheeler C. R., et al. (2021). Elasmobranch responses to experimental warming, acidification, and oxygen loss—A meta-analysis. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.735377
Schäfer S., Monteiro J., Castro N., Gizzi F., Henriques F., Ramalhosa P., et al. (2021). Lost and found: A new hope for the seagrass Cymodocea nodosa in the marine ecosystem of a subtropical Atlantic Island. Regional Stud. Mar. Sci. 41, 101575. doi: 10.1016/j.rsma.2020.101575
Sena N., Veiga A., Semedo A., Abu-Raya M., Semedo R., Fujii I., et al. (2023). Co-designing protected areas management with small island developing states’ Local stakeholders: A case from coastal communities of Cabo Verde. Sustainability 15, 15178. doi: 10.3390/su152015178
Seymour Z. T. A., Monteiro Z. L., Monteiro A., Baremore I. E., Garzon F., Graham R. T. (2024). Baseline assessment of the coastal elasmobranch fauna of Eastern Cabo Verde, West Africa. Aquat. Conservation: Mar. Freshw. Ecosyst. 34, e4206. doi: 10.1002/aqc.4206
Sousa A., Alves F., Arranz P., Dinis A., Fernandez M., González García L., et al. (2021). Climate change vulnerability of cetaceans in Macaronesia: Insights from a trait-based assessment. Sci. Total Environ. 795, 148652. doi: 10.1016/j.scitotenv.2021.148652
Spalding M. D., Fox H. E., Allen G. R., Davidson N., Ferdaña Z. A., Finlayson M., et al. (2007). Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. BioScience 57, 573–583. doi: 10.1641/B570707
Spurgeon E., Anderson J. M., Liu Y., Barajas V. L., Lowe C. G. (2022). Quantifying thermal cues that initiate mass emigrations in juvenile white sharks. Sci. Rep. 12, 19874. doi: 10.1038/s41598-022-24377-1
Suthers I. M., Bishop M. J., Firth L. B., and Dafforn K. A., Keith D. A. (2022a). MT3.1 Artificial shorelines. In: Keith D. A., Ferrer-Paris J.R., Nicholson E., et al (2022). A function-based typology for Earth’s ecosystems – Appendix S4. The IUCN Global Ecosystem Typology v2.1: Descriptive profiles for Biomes and Ecosystem Functional Groups. Nature 610, 1295 513–518. doi: 10.1038/s41586-022-05318-4
Suthers I. M., Claisse J. T., Crowe T. P., Keith D. A. (2022b). M4.1 Submerged artificial structures. In: Keith D. A., Ferrer-Paris J.R., Nicholson E., et al (2022). A function-based typology for Earth’s ecosystems – Appendix S4. The IUCN Global Ecosystem Typology v2.1: Descriptive profiles for Biomes and Ecosystem Functional Groups. Nature 610, 1295 513–518. doi: 10.1038/s41586-022-05318-4
Torres P., Tristão Da Cunha R., Santos Rodrigues A. D. (2016). The elasmobranch fisheries of the Azores. Mar. Policy 73, 108–118. doi: 10.1016/j.marpol.2016.07.027
Tuya F., Pérez-Neira F., Espino F., Bosch N. E., Meyers E. K. M., Jiménez-Alvarado D., et al. (2022). Coastal sharks and rays in the Northeastern Atlantic: From an urgent call to collect more data to the declaration of a marine corridor. Global Ecol. Conserv. 38, e02261. doi: 10.1016/j.gecco.2022.e02261
Vandeperre F., Aires-da-Silva A., Fontes J., Santos M., Serrão Santos R., Afonso P. (2014). Movements of Blue Sharks (Prionace glauca) across Their Life History. PloS One 9, e103538. doi: 10.1371/journal.pone.0103538
Vedor M., Mucientes G., Hernández-Chan S., Rosa R., Humphries N., Sims D. W., et al. (2021a). Oceanic diel vertical movement patterns of blue sharks vary with water temperature and productivity to change vulnerability to fishing. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.688076
Vedor M., Queiroz N., Mucientes G., Couto A., Costa I. D., Santos A. D., et al. (2021b). Climate-driven deoxygenation elevates fishing vulnerability for the ocean’s widest ranging shark. eLife 10, e62508. doi: 10.7554/eLife.62508
Weigmann S. (2016). Annotated checklist of the living sharks, batoids and chimaeras (Chondrichthyes) of the world, with a focus on biogeographical diversity. J. Fish Biol. 88, 837–1037. doi: 10.1111/jfb.12874
Watermeyer K. E., Gregr E. J., Rykaczewski R. R., Shannon L. J., Suthers I. M., Keith D. A. (2022). M1.9 Upwelling zones. In: Keith D.A., Ferrer-Paris J.R., Nicholson E., et al (2022). A function-based typology for Earth’s ecosystems – Appendix S4. The IUCN Global Ecosystem Typology v2.1: Descriptive profiles for Biomes and Ecosystem Functional Groups. Nature 610, 1295 513–518. doi: 10.1038/s41586-022-05318-4
Wenzel F. W., Jann B., Allen J., Carrillo M., Hanquet S., Katona S. K., et al. (2023). Migration of a humpback whale (Megaptera novaeangliae) between the Cape Verde Islands and Iceland. J. Cetacean Res. Manage. 5, 125–129. doi: 10.47536/jcrm.v5i2.812
Whittaker R. J. (1998). Island biogeography: Ecology, evolution, and conservation (Oxford, UK: Oxford University Press).
Wirtz P., Brito A., Falcón J. M., Freitas R., Fricke R., Monteiro V., et al. (2013). The coastal fishes of the Cape Verde Islands – new records and an annotated check-list. Spixiana 36, 113–142.
Keywords: elasmobranch, northeast Atlantic, fishery, habitat degradation, climate change
Citation: Varela J, Santos CP, Nunes E, Pissarra V, Pires S, Ribeiro BP, Vieira E, Repolho T, Queiroz N, Freitas R and Rosa R (2025) Sharks in Cabo Verde, Canarias, Madeira and Azores islands: species richness, conservation status and anthropogenic pressures. Front. Mar. Sci. 12:1490317. doi: 10.3389/fmars.2025.1490317
Received: 03 September 2024; Accepted: 14 January 2025;
Published: 05 February 2025.
Edited by:
Pierluigi Carbonara, COISPA Tecnologia & Ricerca, ItalyReviewed by:
Carlos Eduardo Leite Ferreira, Fluminense Federal University, BrazilCopyright © 2025 Varela, Santos, Nunes, Pissarra, Pires, Ribeiro, Vieira, Repolho, Queiroz, Freitas and Rosa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jaquelino Varela, amFxdWVsaW5vdmFyZWxhMTk4N0BnbWFpbC5jb20=; Rui Rosa, cnJvc2FAZmMudWwucHQ=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.