 Carina R. Hartmann1†
Carina R. Hartmann1† Robin Khan1†Jennifer Schöning1Maximilian Richter1Maike Willers2
Robin Khan1†Jennifer Schöning1Maximilian Richter1Maike Willers2 Sabine Pirr2,3Julia Heckmann1
Sabine Pirr2,3Julia Heckmann1 Johannes Dirks1,4
Johannes Dirks1,4 Henner Morbach1,5Monika Konrad1Elena Fries1Magdalene Winkler6Johanna Büchel6Silvia Seidenspinner1Jonas Fischer1Claudia Vollmuth1Martin Meinhardt1Janina Marissen1
Henner Morbach1,5Monika Konrad1Elena Fries1Magdalene Winkler6Johanna Büchel6Silvia Seidenspinner1Jonas Fischer1Claudia Vollmuth1Martin Meinhardt1Janina Marissen1 Mirco Schmolke7,8Sibylle Haid9Thomas Pietschmann3,9,10Simone Backes11Lars Dölken3,12
Mirco Schmolke7,8Sibylle Haid9Thomas Pietschmann3,9,10Simone Backes11Lars Dölken3,12 Ulrike Löber13,14,15,16Thomas Keil17,18,19Peter U. Heuschmann18,20,21Achim Wöckel6
Ulrike Löber13,14,15,16Thomas Keil17,18,19Peter U. Heuschmann18,20,21Achim Wöckel6 Sagar22Thomas Ulas23,24,25
Sagar22Thomas Ulas23,24,25 Sofia K. Forslund-Startceva13,14,15,26
Sofia K. Forslund-Startceva13,14,15,26 Christoph Härtel1‡
Christoph Härtel1‡ Dorothee Viemann1,2,3,27*‡
Dorothee Viemann1,2,3,27*‡- 1Department of Pediatrics, University Hospital Würzburg, Würzburg, Germany
- 2Department of Pediatric Pneumology, Allergology and Neonatology, Hannover Medical School, Hannover, Germany
- 3Cluster of Excellence RESIST (EXC 2155), Hannover Medical School, Hannover, Germany
- 4German Center for Infection Research, Site Hamburg-Lübeck-Borstel-Riems, Hamburg, Germany
- 5Center for Primary Immunodeficiencies and Autoinflammatory Diseases (CIDA), University Hospital Würzburg, Würzburg, Germany
- 6Department of Gynecology and Obstetrics, University Hospital Würzburg, Würzburg, Germany
- 7Department of Microbiology and Molecular Medicine, University of Geneva, Geneva, Switzerland
- 8Center for Inflammation Research, Faculty of Medicine, University of Geneva, Geneva, Switzerland
- 9Institute for Experimental Virology, Centre for Experimental and Clinical Infection Research (TWINCORE), a joint venture between the Helmholtz Centre for Infection Research and The Hannover Medical School, Hannover, Germany
- 10German Centre for Infection Research, Partner Site Braunschweig-Hannover, Braunschweig, Germany
- 11Institute for Virology and Immunobiology, University of Würzburg, Würzburg, Germany
- 12Institute of Virology, Hannover Medical School, Hannover, Germany
- 13Max-Delbrück-Center for Molecular Medicine in the Helmholtz Association, Berlin, Germany
- 14Experimental and Clinical Research Center, a cooperation of Charité - Universitätsmedizin Berlin and Max-Delbrück-Center for Molecular Medicine, Berlin, Germany
- 15Charité - Universitätsmedizin Berlin, Corporate Member of Freie Universität Berlin, Humboldt-Universität zu Berlin, and Berlin Institute of Health, Berlin, Germany
- 16German Centre for Cardiovascular Research (DZHK), Partner site Berlin, Berlin, Germany
- 17Institute of Social Medicine, Epidemiology and Health Economics, Charité - Universitätsmedizin Berlin, Berlin, Germany
- 18Institute of Clinical Epidemiology and Biometry, University of Würzburg, Würzburg, Germany
- 19State Institute of Health I, Bavarian Health and Food Safety Authority, Erlangen, Germany
- 20Institute for Medical Data Science, University Hospital Würzburg, Würzburg, Germany
- 21Clinical Trial Centre Würzburg, University Hospital Würzburg, Würzburg, Germany
- 22Department of Medicine II (Gastroenterology, Hepatology, Endocrinology, and Infectious Diseases), Freiburg University Medical Center, University of Freiburg, Freiburg, Germany
- 23Systems Medicine, German Center for Neurodegenerative Diseases Deutsches Zentrum für Neurodegenerative Erkrankungen (DZNE), Bonn, Germany
- 24PRECISE Platform for Single Cell Genomics and Epigenomics, Deutsches Zentrum für Neurodegenerative Erkrankungen (DZNE) and University of Bonn, Bonn, Germany
- 25Genomics and Immunoregulation, Life and Medical Sciences (LIMES) Institute, University of Bonn, Bonn, Germany
- 26Structural and Computational Biology Unit, European Molecular Biology Laboratory (EMBL), Heidelberg, Germany
- 27Center for Infection Research, University Würzburg, Würzburg, Germany
Introduction: Respiratory viral infections (RVIs) are a major global contributor to morbidity and mortality. The susceptibility and outcome of RVIs are strongly age-dependent and show considerable inter-population differences, pointing to genetically and/or environmentally driven developmental variability. The factors determining the age-dependency and shaping the age-related changes of human anti-RVI immunity after birth are still elusive.
Methods: We are conducting a prospective birth cohort study aiming at identifying endogenous and environmental factors associated with the susceptibility to RVIs and their impact on cellular and humoral immune responses against the influenza A virus (IAV), respiratory syncytial virus (RSV) and severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). The MIAI birth cohort enrolls healthy, full-term neonates born at the University Hospital Würzburg, Germany, with follow-up at four defined time-points during the first year of life. At each study visit, clinical metadata including diet, lifestyle, sociodemographic information, and physical examinations, are collected along with extensive biomaterial sampling. Biomaterials are used to generate comprehensive, integrated multi-omics datasets including transcriptomic, epigenomic, proteomic, metabolomic and microbiomic methods.
Discussion: The results are expected to capture a holistic picture of the variability of immune trajectories with a focus on cellular and humoral key players involved in the defense of RVIs and the impact of host and environmental factors thereon. Thereby, MIAI aims at providing insights that allow unraveling molecular mechanisms that can be targeted to promote the development of competent anti-RVI immunity in early life and prevent severe RVIs.
Clinical trial registration: https://drks.de/search/de/trial/, identifier DRKS00034278.
Introduction
Several epidemiological studies demonstrate strong age-dependent differences for the susceptibility and outcome of respiratory viral infections (RVIs) (1). Prime examples are infections with the respiratory syncytial virus (RSV), the influenza A virus (IAV) and the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2).
RSV is the most common cause of severe respiratory infection in infants, leading to over 3 million hospitalizations and around 66,000 deaths worldwide each year (2–4). Remarkable age-related differences in innate cytokine responses following recognition of RSV have been observed (3, 5), suggesting a critical role of the host response in the disease pathology and clinical outcome. RSV infects virtually all children by the age of three years and then repeatedly infects throughout life. Therefore, host factors of the epithelial barrier and the innate immune system might define the susceptibility to RSV infections, while adaptive immunity, which underlies profound maturational changes during early life, might rather contribute to the severity of the course of established RSV infections, e.g. by early-life Th-2 skewed T-cell responses (6, 7) promoted by neonatally high epithelial-derived interleukin-33 levels (8, 9), but does not necessarily acquire long-lasting protective memory.
In contrast to the age profile of RSV infections, work from the Global Burden of Disease Study showed that the population attributable fraction of IAV-caused mortality as well as morbidity are lowest in children under 5 years of age and highest in mid-aged adults (30-40 years) (10). The age-dependent differences in the clinical outcome of IAV infection are not fully explained by pre-existing comorbidities or maternal transfer of adaptive anti-influenza immunity to infants and point to less understood functional distinctions that protect infants against IAV. Possibly, the difference lies particularly at the level of the airway epithelial response to IAV. It likely differs from that against RSV and might be more effective in infants compared to adults in directly preventing the establishment of IAV infection or indirectly initiating a recruitment of less inflammatory immune cells according the tolerance concept of neonatal immunity (11–13). Another important aspect might be the influenza vaccine coverage, which is highest in children aged 0-4 years (about 60% in the U.S., 40% in the U.K.) and lowest in young adults aged 18-49 years (about 40% in the U.S., 7% in the U.K.) (14, 15).
Children have also less severe symptoms when infected with SARS-CoV-2 (16, 17). Several hypotheses for the age-related difference in the severity of coronavirus disease 2019 (COVID-19) are discussed, such as more robust type I interferon (IFN) response, more effective T cell immunity (18), and higher expression of SARS-CoV-2 sensing receptors and inflammatory baseline activation of pediatric compared to adult airway epithelial cells (AECs) (19–21). However, the causes and biological meaning of such differential programming of anti-viral immunity at different ages and in dependency of the type of virus remains elusive.
Apart from the age dependency there is also strong heterogeneity in the course and outcome of RVIs among individuals of same age, pointing to considerable genetic variability or differential development of immunity in concert with their environment. Understanding inter-individual variation and its causes has important implications for targeting patients for escalation of care, inclusion in clinical trials, and individualized medical therapy including vaccination. Besides clear biological differences such as age, sex, race, presence of comorbidities, and genetic variation, differential maturation of immunity during environmental adaptation after birth might result in a different education and regulation of immune responses against RVIs. In humans, the key factors influencing the reprogramming of immunity towards RVIs are still poorly defined and comparison of their effect sizes is long overdue.
A myriad of data has demonstrated that the immune system of the fetus and infant is characterized by a high plasticity, which comes with a high susceptibility to lifelong imprinting effects from environmental cues (12, 13, 22–24). Over the past two decades strong evidence accumulated that the composition and function of the human microbiome play a particularly critical role for the development of immunity and overall health (25, 26). Several longitudinal studies revealed aberrant trajectories of upper respiratory microbiota during infancy that are associated with an increased susceptibility to respiratory tract infections. These particularly include delayed and low abundance of Corynebacterium and Dolosigranulum species and high abundance of Moraxella, Haemophilus or Streptococcus species (27–31). However, our knowledge on how exactly these strains impede respective promote immunity against respiratory viruses remains largely elusive.
Furthermore, the gut microbiota seems to have impact on the development of immunity against RVIs. Though hitherto only shown in mice, gut microbiota-derived short chain fatty acids protected against RSV infections by improving type 1 IFN responses and increasing IFN-stimulated gene expression in lung epithelial cells (32), while the presence of segmented filamentous bacteria protected against IAV, RSV as well as SARS-CoV-2 infection by priming resident alveolar macrophages (33). In humans, only an association between specific gut microbial profiles and RSV disease severity in hospitalized infants has been detected so far (34). However, the molecular pathways of this gut-lung axis and their delicate dependency on age and interactions with environmental exposures (e.g., diet, passive smoke, infections, pollutants) remain unclear. Several clinical trials contributed to an increasing body of evidence that probiotics might be an option to lower the susceptibility to RVIs (35–37); however, many questions on how exactly (e.g., strain dependency), to what extent individualized (e.g., age dependency, host microbial background) and when (e.g., neonatal window) to use such interventions are still open (38–40). Addressing these remaining knowledge gaps could enable modulation of airway and gut microbial communities and their metabolic patterns, presenting a potential therapeutic strategy to prevent or ameliorate severe RVIs.
In summary, at specific developmental stages opposite infectious susceptibility can be observed depending on the causative viral strain. Moreover, different trajectories of immune maturation after birth can lead to strong inter-population differences in the susceptibility to severe RVIs. To understand which specific age- and microbiota-related immunological differences and temporal changes are most relevant across various RVIs, we need a comprehensive view of the longitudinal development of anti-RVI immunity and the impact of and interplay with key factors involved. This requires the establishment of a tailored birth cohort study that investigates longitudinally and in-depth the development of the function of the airway epithelial barrier site and professional innate and adaptive immune cells in newborns against different respiratory viruses. Thereby, clinical and environmental factors including the status of the respiratory and gut microbiota will be documented and analyzed to assess their influence on these developmental processes. Such comprehensive datasets are imperative for the development of novel interventions that either target the host or the microbiota to early prevent or successfully treat severe RVIs in the future.
Aim
The MIAI study hypothesizes that: (i) the course and outcome of RVIs among individuals is related to the large variability in maturational trajectories of anti-viral immunity and (ii) environmental, lifestyle and dietary factors influence the early imprinting of anti-viral immunity in ways that either promote an increased susceptibility to RVIs and their complicated course or support protective patterns (Figure 1). To verify these hypotheses, the MIAI birth cohort has been set up and includes healthy newborn infants born at term and their families. We aim at a comprehensive characterization of age-related differences and changes in the human immune system of the participants with a focus on the function of the cellular key players involved in the establishment of infections with RSV, IAV and SARS-CoV-2. Detailed phenotyping including factors potentially promoting and confounding the individual immune trajectory will be established; in particular, we are interested in the role of the developing microbiome on priming and regulating anti-RVI immunity. The specific aims of our study are:
● Aim 1. To characterize the developmental changes of innate and adaptive immune responses to RVIs during the first year of life.
● Aim 2. To determine the impact of sociodemographic background, clinical histories and environmental factors like lifestyle, vaccinations and dietary factors on the trajectories of immune responses toward RVIs.
● Aim 3. To elucidate whether there is a link between the microbiome establishment and anti-RVI immunity developing after birth.
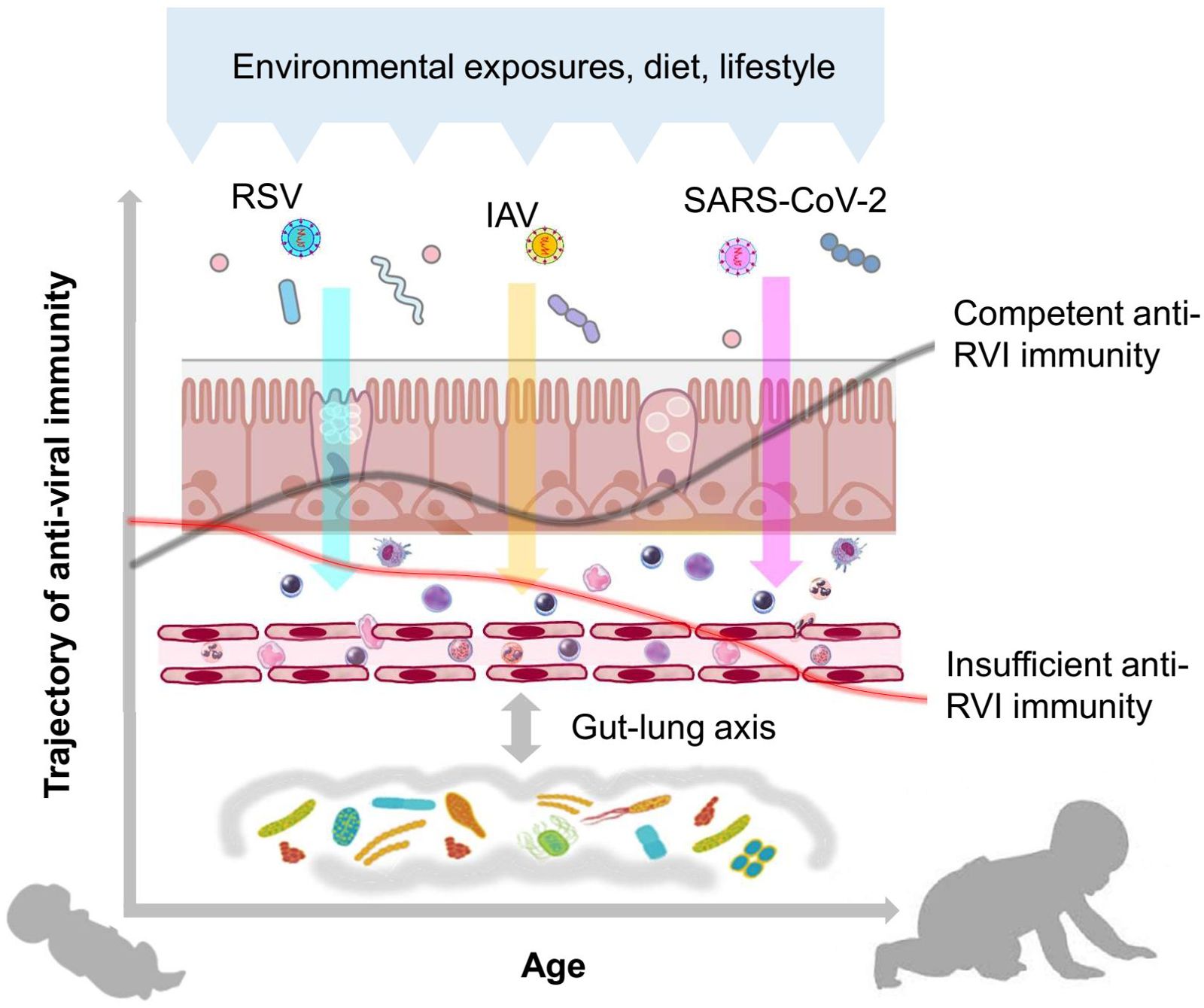
Figure 1. Hypothesis of the MIAI study. The hypothesis of the MIAI study is that the maturation of anti-viral immunity toward RVIs in early life underlies a large developmental variability, which is imprinted by clinical and environmental factors such as dietary habits, lifestyle, and particularly the composition and function of the co-evolving respiratory and gut microbiota.
In brief, we aim to provide insights into age-dependent programming and early-life changes of immunity toward RVIs in humans, highlighting training, priming and acquisition of tolerance, by collecting granular clinical metadata and performing comprehensive unbiased immune phenotyping using multi-omics approaches (Figure 2).
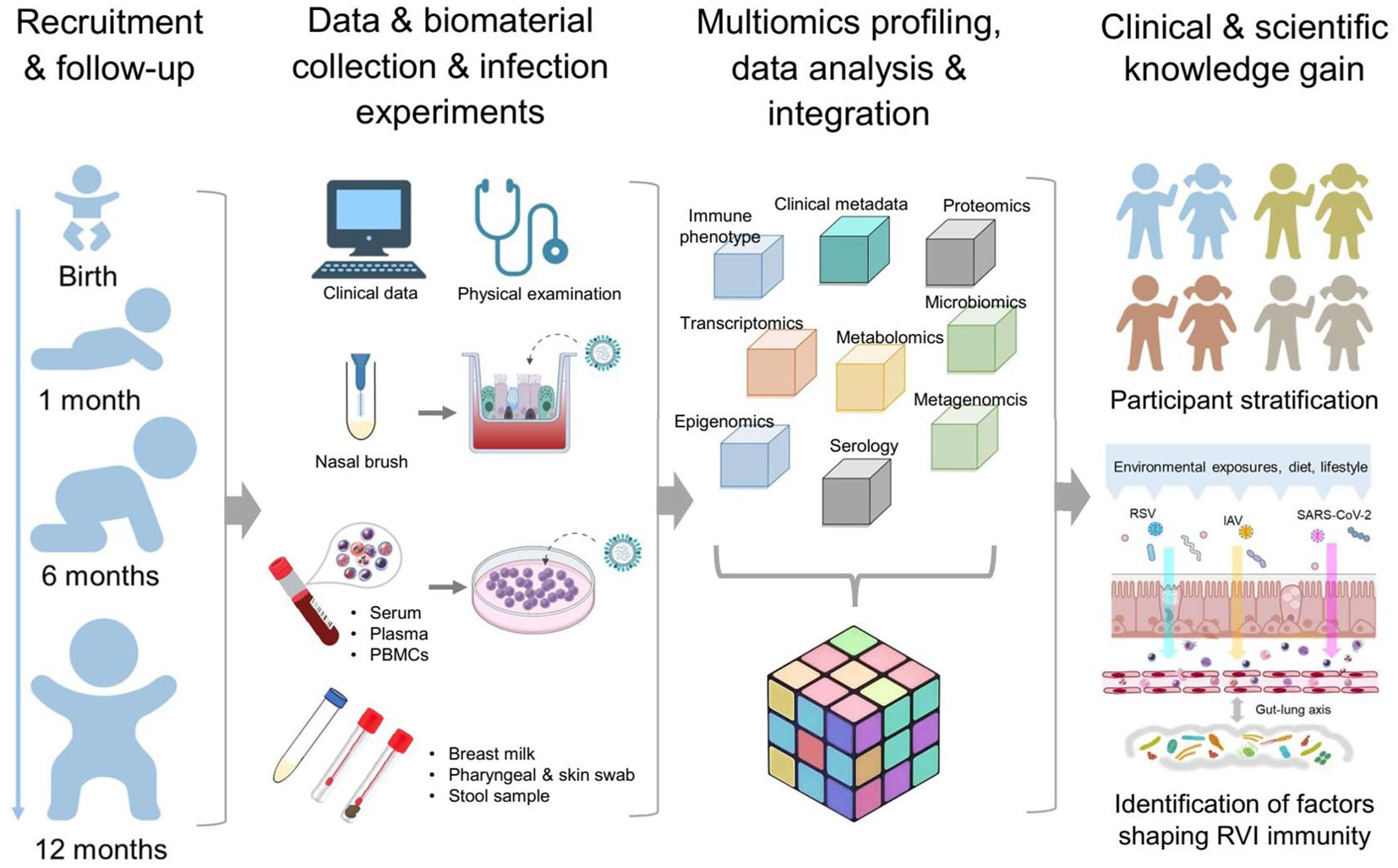
Figure 2. Design MIAI study. The MIAI study encompasses four phases starting with recruitment of term healthy infants and follow-up visits at defined timepoints. At each appointment, clinical data and biomaterials are collected for storage and/or usage in ex vivo respiratory viral infection models. Multi-omics datasets will be generated including immune profiles, transcriptomics, epigenomics, proteomics, metabolomics and microbiomics. To gain clinical and scientific knowledge, overall integration of mixed omics data shall enable participant stratification based on marker patterns linking to RVI susceptibility and reveal host and environmental factors imprinting anti-RVI immunity in the first year of life.
Methods and analysis
Study design, cohort recruitment and follow-up visits
The MIAI study is a single center, population-based birth cohort study at the University Hospital Würzburg. MIAI enrolls healthy newborn infants and their families and follows them throughout the first year of life (Figure 2). The ongoing recruitment started in February 2022 reaching about 170 infants up to date (with a 2-month interruption of recruitment due to an interim shortage of staff) (Table 1). Inclusion criteria encompass full-term, healthy neonates (37 + 0 to 41 + 6 gestational weeks) born at the perinatal center of the University Hospital Würzburg, Germany, whose families´ residency is located in the city of Würzburg or its administrative district. Exclusion criteria are amniotic infection syndrome, early-onset neonatal sepsis, congenital malformations, primary immunodeficiency or metabolic diseases, and remarkable language barriers. We will however do an earnest attempt to provide understandable information on our study in the native language of parents whose infants would be eligible thus aiming for a population-representative sample. The study recruits a convenience sample cohort; as such, the sample size is not calculated.
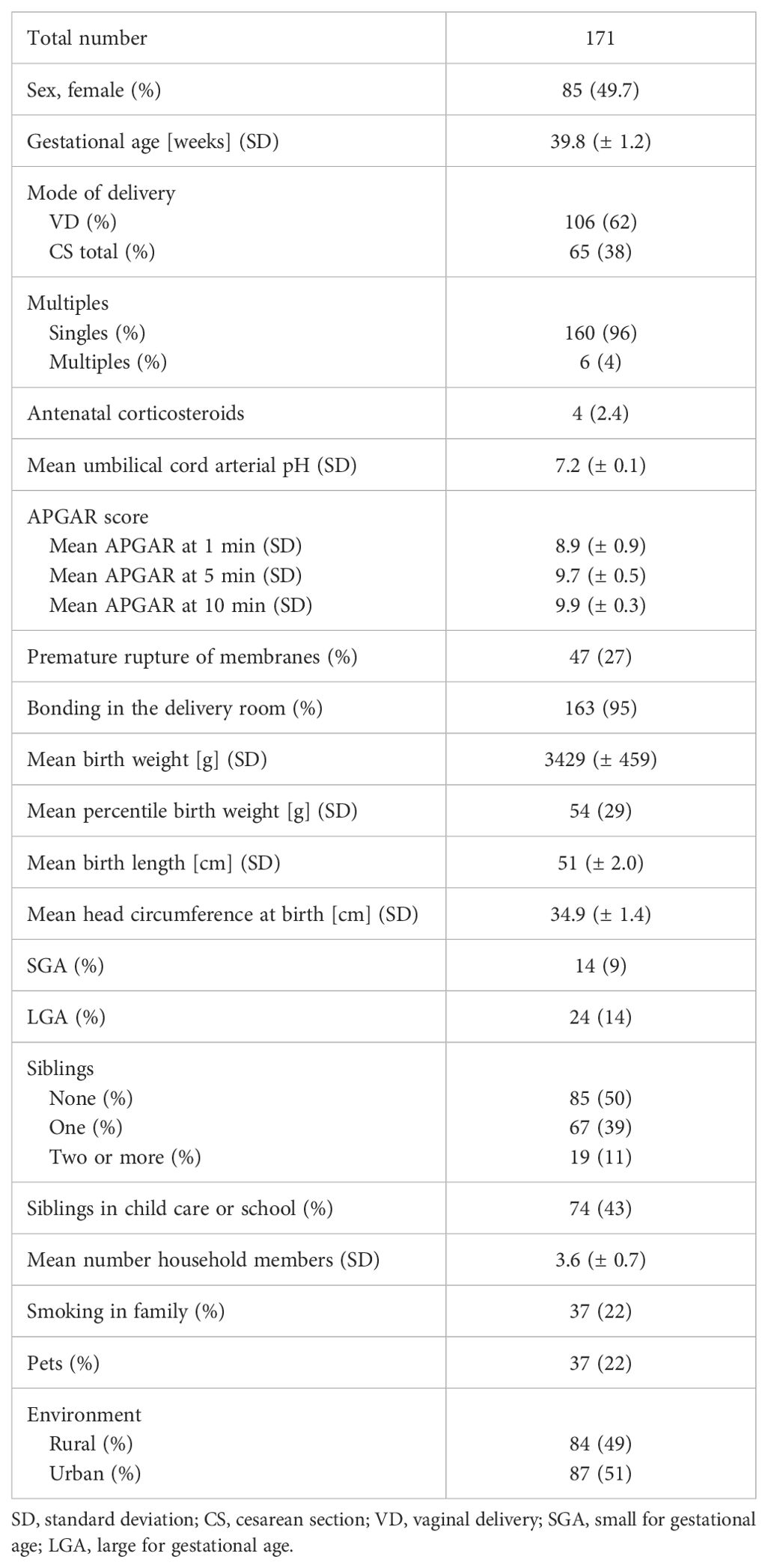
Table 1. Baseline characteristics of the MIAI cohort.
The first study visit is performed at the University Hospital for Women`s health and Obstetrics Würzburg. Mothers of eligible infants are approached in the first three days after birth. After obtaining written informed consent from the parents, the baseline assessment takes place during the hospital stay of mother and infant. The follow-up appointments are performed one month, six months and one year after birth at the dedicated Pediatric Clinical Study Outpatient Service at the University Children’s Hospital Würzburg. We are planning to track the study participants for long-term follow-up during childhood.
Overall, the acceptance of our study design with its follow-up appointments and sample collection is high with a relatively low drop-out rate of about 8% after one year as of May 2024, including families that relocated outside of Würzburg and its administrative district. The baseline characteristics of infants enrolled so far are presented in Table 1. Overall, the proportions of female and male infants are equal and the mean gestational age and birth weight appropriate and in line with the inclusion criterium of full-term birth. The largest number of neonates was born vaginally (VD, 62%). Cesarean sections (CS, 38%) were in the majority of CS secondary (60%), i.e. after rupture of membranes, and only a minor part elective (37%) or due to emergency (3%). Half of the MIAI infants have older siblings which of 86% are visiting child care or school. The exposure to nicotine due to smoking family members affects 22% of the MIAI infants and 22% of the families have pets. The distribution of rural compared to urban living environments is even.
Characteristic values of pregnancy histories from infants enrolled in MIAI including maternal factors potentially impacting on the child’s RVI immunity are listed in Table 2. The mean weight gain during pregnancies was 14 kg with a mean body mass index (BMI) of 29.4 (± 5.8) at birth, not exceeding the limits recommended by obstetricians (41). The smaller number of mothers were primigravida (38%). During routine prenatal screenings, gestational diabetes mellitus was detected in 12% and gestational hypertension in 1.8%. The proportions of mothers vaccinated and/or recovered from IAV and SARS-CoV-2 infections are 48% and 97%, respectively. For the Influenza status, 52% of mothers did not know whether they have experienced an IAV infection, while only 3% of mothers did not know their SARS-CoV-2 status.
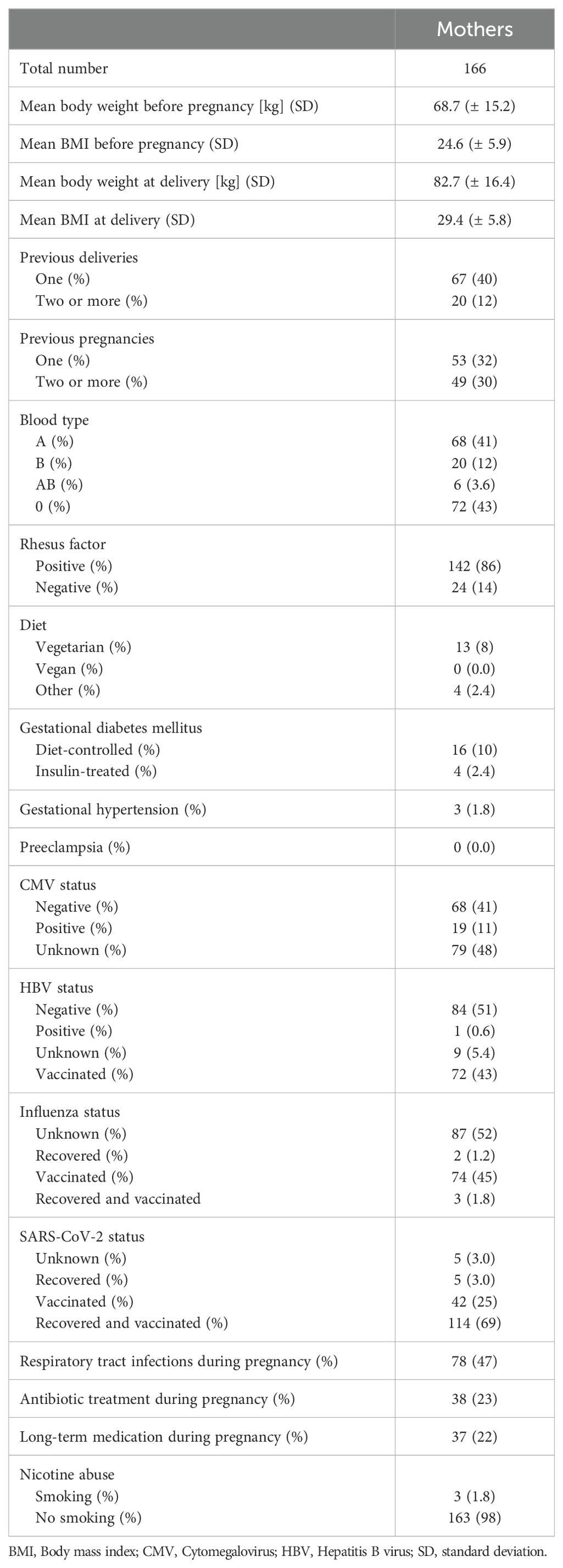
Table 2. Baseline characteristics of pregnancy in the MIAI cohort.
Table 3 summarizes anthropometric, medical and sociodemographic characteristics of participating parents. The mean age of mothers and fathers from the entire cohort is 32.8 (± 4.5) years and 35.3 (± 5.4) years, respectively. The medical history data of the parents showed that 52% of the mothers and 24% of the fathers reported somatic disorders including allergies, psoriasis, neurodermatitis, cardiovascular disorders, obesity, thyroid disorders, as well as benign and malignant tumors. Mental disorders were indicated by 8.4% of the mothers and 6.0% of the fathers. About 84% of the participating parents reported a Western European ethnicity and 79% of the mothers and 71% of the fathers were higher educated.
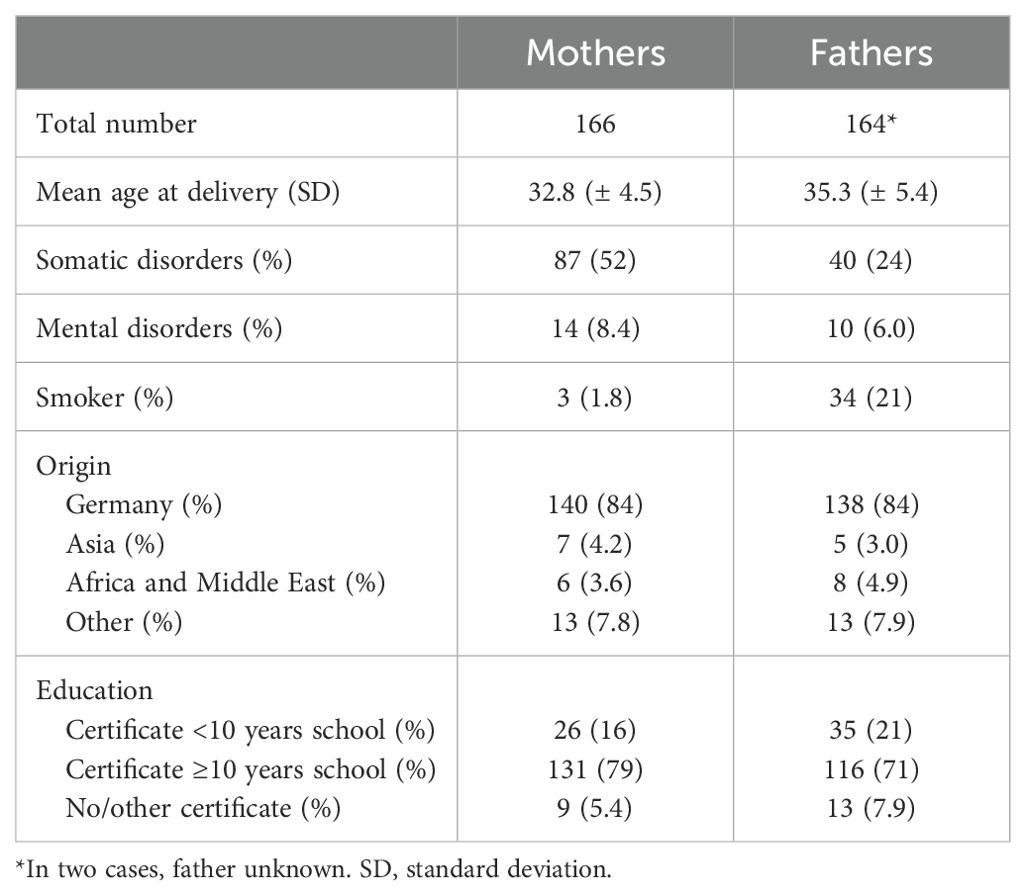
Table 3. Baseline characteristics of the parents in the MIAI cohort.
Clinical data collection and data management
Clinical data collection in MIAI is accomplished using tailored case report forms (CRFs) at each study timepoint (Table 4). At baseline, the histories of pregnancy and perinatal events are documented upon enrollment. From mothers and fathers, detailed information on demographic and psychosocial background, family histories and lifestyles are requested. Particular attention is paid on maternal respiratory infections and vaccinations, before and during pregnancy. For the infants, we document at each study visit body measurements, the histories of vaccinations, infections, antibiotics including days of treatment, medications including inhalations, episodes of wheezing, skin rashes, allergies, hospital stays, and surgical procedures, diets including dietary supplements, lifestyle including the number of siblings and further household members, pets, residence (rural/urban), and stays abroad. In addition to the questionnaires, the MIAI pediatrician performs a comprehensive physical examination of the infant at each study appointment to monitor the overall health status including growth, physical and psychomotor development (Table 4).
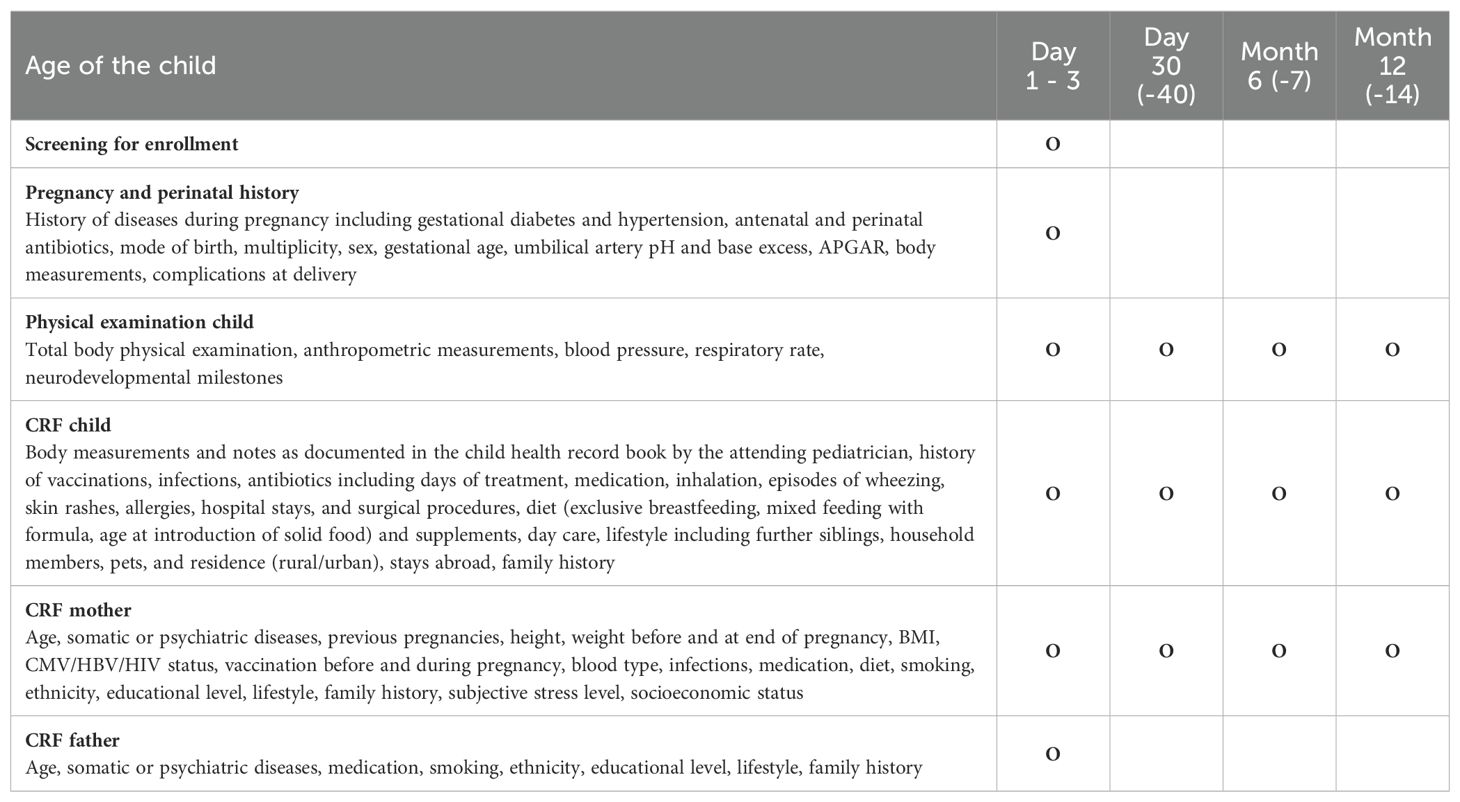
Table 4. Overview of study appointments and clinical data collected in MIAI.
To align with the FAIR data principles (Findable, Accessible, Interoperable, Reusable), the collected clinical data is managed to ensure high standards of data quality and usability. For data protection reasons, the participant’s data is pseudonymized and assigned a unique study ID, which is documented in a separate and password-secured data file accessible by authorized MIAI staff only. A separate external data file is used as a contact database for tracking study participations and invitations to follow-up. The pseudonymized clinical data are double-entered into a password-secured REDCap database hosted at the Institute of Clinical Epidemiology and Biometry, University of Würzburg (IKE-B). Local password-secured copies of the database are generated. The database provides a comprehensive log/audit feature to record data input and changes based on user and time/date. By adhering to these FAIR principles, MIAI enhances the value of its clinical data, promoting its use in further research while maintaining high standards of data security and participant confidentiality.
Collection and storage of biomaterials
At each appointment, biomaterials from mothers (breast milk) and child (nasal brushes, pharyngeal swabs, skin swabs, stool, blood) are collected according to the schedule illustrated in Figure 3. We have chosen these timepoints to afford serial profiling while minimizing burden. Collection and storage of biomaterials are performed according to established standard operating procedures (SOPs). After collection, biomaterials are immediately transferred to the adjacent laboratory for processing and biobanking.
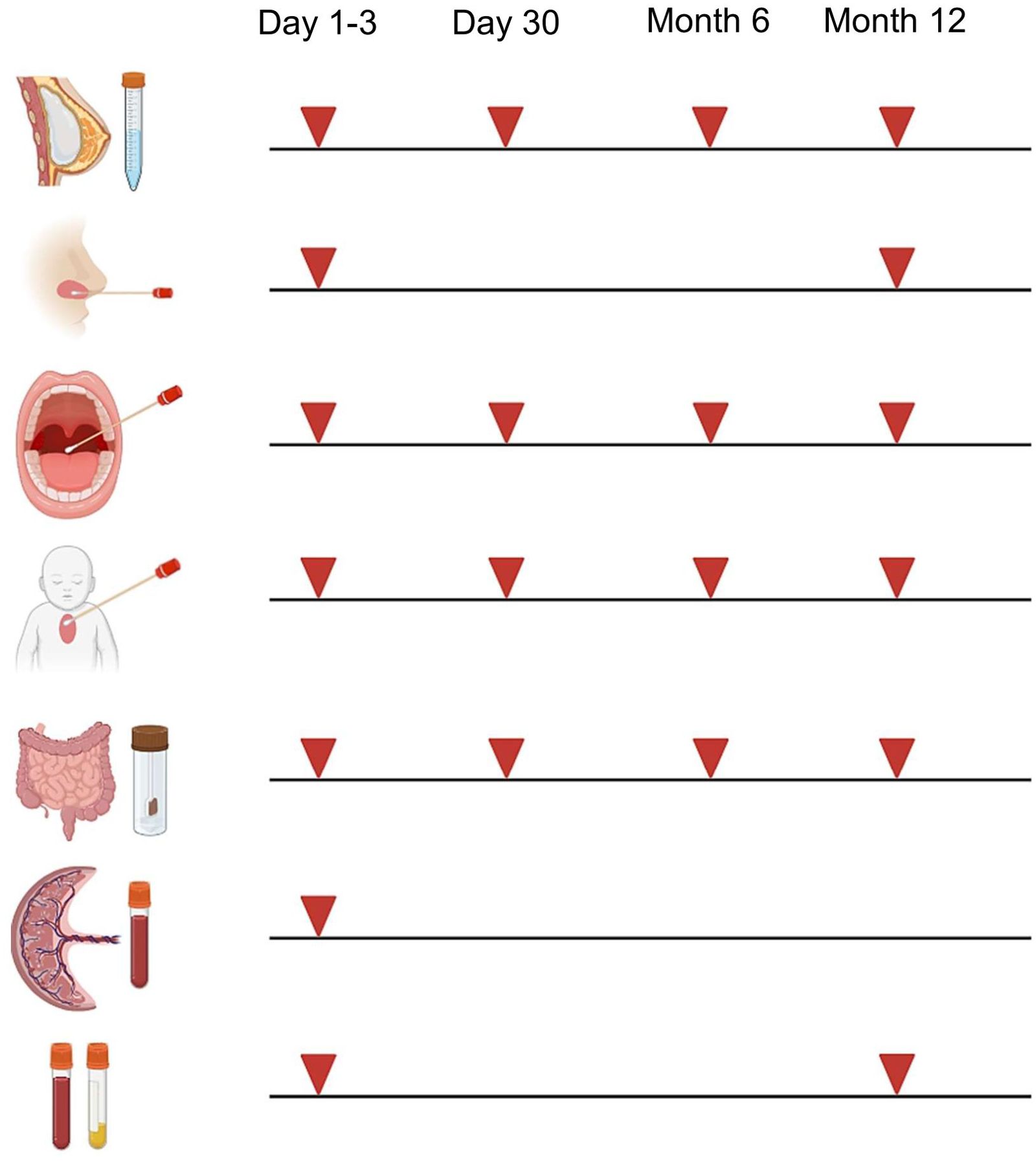
Figure 3. Collection of biomaterials in MIAI. Biomaterials collected in the MIAI cohort include breast milk, nasal brushes and secretions, pharyngeal swabs, skin swabs, stool, cord blood and peripheral blood (EDTA and serum). They are obtained at the defined study time points, i.e. either at day 1-3 and/or day 30 of life and/or at 6 months and/or 1 year following birth. (Created with BioRender.com).
Breast milk samples
As long as the mother is breastfeeding, breast milk/colostrum are sampled at each appointment. First, cells are pelleted by centrifugation and the skim milk and the fat layer are removed. The skim milk and fat layer are once more sharply centrifuged and aliquoted for storage at -80°C for later proteomic and microbial studies. The cells are cryopreserved at -152°C in RPMI 1640 with 20% fetal calf serum (FCS) and 10% dimethyl sulfoxide (DMSO) (all Sigma-Aldrich, Darmstadt, Germany) following multiple washing steps for later single cell studies.
Nasal brushes
Nasal brushing is an established method in pediatric pulmonology, e.g. essential for diagnosing primary ciliary dyskinesia, and provides a robust technique to generate airway epithelial cell (AEC) cultures in term and even preterm infants (21, 42). Brushings are taken from the inferior nasal turbinate using sterile interdental brushes (Rossmann, ISO sizes 4 or 5). After sampling, brushes are immediately placed in sterile tubes with transport medium (MEM without additives, antibiotics and antimycotics) and processed within 1 h of collection. Brushes are carefully moved up and down to scrape the cells off and collect them into the transport medium. Next, cells are centrifuged and supernatants stored as “nasal secretions” at -80°C for later proteomic, cytokine and microbiota profiling. The AECs are first expanded in submersion according to established protocols (43, 44) and then cryopreserved at -152°C in BEGM medium (Lonza, Basel, Schweiz, Switzerland) with 20% FCS and 10% DMSO. Some of the expanded AECs are directly differentiated at the air-liquid-interface (ALI) into a mature pseudostratified respiratory epithelium as described previously (44) and used for infection experiments with RSV (RSV-A-ON1, HRSV/A/DEU/H1/2013) (45) and IAV (H1N1, A/Netherlands/602/2009). Infections with SARS-CoV-2 (SARS-CoV-2/human/DEU/REGS-200701-CA/2020) are done later under biosafety level 3 conditions using cryopreserved AECs for differentiation at ALI. After infection, AECs are harvested and subjected to immediate flow cytometry studies capturing cell composition and cell quality, generation of total RNA lysates for the analysis of transcriptomes (bulk RNASeq) and viral loads (qRT-PCR), and standardized cryopreservation (46) for later single cell (scRNASeq) and epigenetic (ATACSeq) studies. Apical and basolateral supernatants are stored at -80°C for later analyses of proteome (mass spectrometry) and cytokine (FACS-based multiplex assays) profiles and virus plaque assays assessing viral progeny. This approach aims at generating data from each MIAI participant regarding individual airway epithelial immune responsiveness and function toward RVIs at birth and at 1 year of life (Figure 3). The strategy is based on previous reports that ALI-AEC models established from human primary AECs recapitulate well the in vivo phenotype of the airway epithelium (21, 47, 48).
Pharyngeal and skin swabs
Three pharyngeal swabs and two skin swabs are collected, respectively. One of the respective swabs is transferred to a sterile tube containing sterile PBS with 10% BSA (for protein stabilization) and stored at -80°C for protein and cytokine measurements. The other two respective swabs are stored at -80°C without additives for later DNA isolation and microbiota studies (16S rRNA sequencing and shotgun metagenomics).
Stool samples
Stool samples are collected by the MIAI staff from diapers and immediately transferred to -80°C. In cases where stool collection at the respective appointments not succeeds, we use samples collected and stored at 4°C by the parents at the evening before or the morning of the follow-up appointment for biobanking at -80°C. Three 1.5 ml tubes are filled with stool, respectively, one of which contains 600 µl DNA/RNA shield (Zymo Research, California, USA) to preserve nucleic acids for 16S rRNA sequencing and metagenomics studies. The other two stool aliquots are stored without additives and used for the extraction and analysis of immunoactive proteins such as S100-alarmins (49) and cytokine profiles according to established protocols (50).
Blood samples
Cord blood is collected preemptively from all eligible neonates but only processed when parents agree on participation in the MIAI study. If no consent is given, cord blood samples are discarded. The volumes of drawn peripheral venous blood samples align to the pediatric guidelines of the European Medical Agency, which recommends that blood collection for research purposes should be at maximum 1% of total blood volume (i.e., 0.8 ml/kg body weight). At the first sampling timepoint, we seek to combine blood collection in MIAI with the routine newborn screening taken between 36 and 72 hours of life. Blood samples are processed within 1 h after collection. After blood clotting, serum samples collected in serum separator tubes are centrifuged at room temperature 1,800 x g for 10 minutes and stored in two aliquots at -80°C and used for cytokine and metabolome studies (high-resolution mass spectrometry) and serological studies including quantification of antibody titers against RSV, IAV and SARS-CoV-2. From EDTA samples, peripheral blood mononuclear cells (PBMCs) are isolated by Ficoll-Paque PLUS (Sigma-Aldrich) density gradient centrifugation and profiled using established multidimensional flow cytometry panels (51–55). One part of isolated PBMCs is cryopreserved at -152°C in RPMI 1640 with 20% FCS and 10% DMSO for later single cell, epigenetic and experimental studies (e.g., analysis of T and B cell subsets and receptor repertoires including virus-specific effector T cells). To avoid functional impairment of the myeloid cells due to cryopreservation, monocytes are directly isolated from the other part of isolated PBMCs as described previously (11, 56) and infected with RSV and IAV. After infection, total RNA lysates are generated and culture supernatants harvested and both stored at -80°C for the analysis of transcriptomes, viral loads, proteome and cytokine profiles, and viral progeny.
Sample analysis strategy
Due to the longitudinal nature of the study, exceeding differences in the sample storage times that could give rise to data variability and bias ideally should be avoided. Moreover, exceeding storage times in principle need to be prevented due to potential degradation over time depending on the nature of the biological material and the intended downstream application. Our sample analysis strategy adheres to the standards of the International Society for Biological and Environmental Repositories (ISBER) and the Biobanking and BioMolecular resources Research Infrastructure (BBMRI-ERIC). Samples stored at -80°C that will be subjected to proteomic and cytokine studies (skim milk supernatants, nasal and pharyngeal secretions, skin swabs, stool and serum samples, and cell culture supernatants from AEC and monocyte infection models) are usually stable for several years to decades. At least two aliquots are prepared to avoid freeze-thaw cycles. Likewise, samples collected for microbiome studies (skim milk supernatants, nasal and pharyngeal secretions, skin swabs, and stool samples) can be stored for several years at -80°C. After DNA isolation, samples are directly subjected to library preparation and sequencing. Likewise, RNA lysates in RNAlater generated from AEC and monocyte cell culture infection models can be stored for years at -80°C while RNA is only isolated and libraries prepared shortly before bulk RNAseq is appointed. Cryopreserves (breast milk cells, AECs, and PBMCs) are stored at -152°C in a highly standardized manner ensuring cell viability for several years and high quality for later single cell studies like scRNAseq.
In principle, samples stored for subsequent generation of big omics datasets in batches (e.g., transcriptomics, proteomics, metabolomics, microbiomics) are processed when full series from n=100 individuals have been collected and when the storage time of the most recently stored samples is 6 months. With a current recruitment rate of 6 children per month this strategy ensures the analysis of full series from n=100 children within 2.8 years (calculated from the date on which the first patient of each batch has been recruited and sampled). This comes with differences in storage times between d1-3 samples and 1-year samples but needed to be weighed against the batch effects that result from analyzing samples at too many different time points. Based on our experiences in multiple different cohorts we evaluated the latter more considerable. The 6-months storage quarantine is a strategy to mitigate the storage time differences by subjecting 1-year samples at least to a significant storage time to become more comparable to d1-3 samples. Nevertheless, the sample storage time is definitely handled as potential covariate and is as such integral part of each data analysis pipeline used in MIAI.
Data analysis and statistics
Omics data analysis in MIAI will be performed on different levels as results from flow cytometrics, transcriptomics including lymphocyte receptor repertoires, epigenomics, 16S rRNA gene and shot-gun metagenomics sequencing, breast milk and serum metabolomics and proteomics including cytokine and serologic profiles. We aim at subjecting full longitudinal sample series from 400 infants to study the omics profiles. Given the longitudinal nature of our study set up, stored samples previewed for sequencing, proteomic and metabolomic studies will be randomized to reduce batch effects. Initially, each omics dataset will undergo individual analysis to elucidate its unique characteristics. Flow cytometry data are acquired with an Attune CytPix Flow Cytometer using the Attune Cytometric Software version 6.0 (Thermo Fisher Scientific) and then subjected in batches to analyses using the Pytometry (57) and cyCONDOR (58) pipeline. Subsequent exploratory analyses will determine the necessity of batch correction for each dataset. If required, corrections will be implemented using ComBat batch method to normalize across multiple analytical runs and WithinVariation function implemented in the R mixOmics package and to normalize features as change over time within each child. Post-correction, we will validate that the biological variability is preserved and that no artifacts were introduced. We will perform omics-specific statistical analyses and co-expression network analyses using hCoCena (59, 60) to identify and characterize differential subgroups with greater precision. All data generated including standardized meta-data will be quality-controlled and gathered within a unified framework following standards for omics and clinical measurement representation, allowing within-study integration and comparative analysis with previous studies according to FAIR data principles.
Differential trajectorial endotypes will be identified using KmL (k-means for longitudinal data) clustering. To determine the optimal number of clusters (k), we will run 1000 permutations, utilizing the Harabasz criterion (Calinski-Harabasz index) to evaluate clustering validity. This methodology will facilitate a nuanced, continuous classification of patients, transcending binary categorization and allowing also for a more detailed representation of endotypic variation. Specialized software packages for dataset-specific imputation will be employed to address any missing data points, ensuring robust classification of immune and microbiome trajectories across the cohort. Advanced statistical methods and machine learning algorithms will be integrated into the analysis pipeline to enhance the precision and reliability of the endotype classification.
To elucidate the relationships between specific immunotypes, respiratory or gut microbiome profiles, environmental factors, and clinical outcomes, we have created two specialized software tools, metadeconfoundR (61) and longdat (62). These tools enable comprehensive mixed omics data analysis while managing the complex interplay of numerous demographic, therapeutic, and disease-related variables. MetadeconfoundR is designed to facilitate the identification of biomarkers within cross-sectional multi-omics medical datasets, while carefully accounting for confounders. Initially, it identifies significant associations between features and metadata using nonparametric tests such as the Mann-Whitney U test. Subsequently, it assesses and adjusts for confounding effects among these metadata variables through post-hoc nested linear model comparison. Importantly, metadeconfoundR can integrate previously established knowledge of confounders, such as drug associations from the MetaCardis cohort, into its analysis. This inclusion aids in maintaining robust conclusions even when new datasets lack the statistical power to independently identify these confounders. LongDat, meanwhile, is an R package tailored for analyzing longitudinal multivariable data, effectively handling a wide array of covariates. It distinguishes direct from indirect intervention effects and identifies key covariates that may act as mechanistic intermediates. Although primarily focused on longitudinal microbiome data, LongDat’s versatility allows application to various data types including binary, categorical, and continuous data. Comparative testing against other analytical tools like MaAsLin2, ANCOM, lgpr, and ZIBR has demonstrated LongDat’s superior performance in terms of accuracy, runtime, and memory efficiency, making it an ideal choice for high-dimensional longitudinal studies where multiple covariates are a consideration.
Next steps and outlook
To gain a comprehensive picture of how immune profile development including cellular responses to RVIs varies amongst individuals, we are in the process to analyze the biomaterials by a multi-omics approach. This encompasses transcriptomic, epigenomic, metagenomic, and metabolomic layers. These data are extended by cellular phenotyping using high-dimensional flow cytometry and the innate and adaptive immune responses to IAV, RSV and SARS-CoV-2. Next, the omic phenotypes will be integrated with large clinical datasets.
As most of the one-year-old infants have not yet attended a day care center, which is an important contributor to microbiome maturation (63, 64), we continue the longitudinal assessment of MIAI participants by the means of annual questionnaires and plan regular follow-ups including biomaterial sampling and further omics profiling at the age of 3, 6, 9 and 12 years. Our long-term vision is to disentangle the role of early programming of anti-viral immunity for the susceptibility to RVIs and also non-communicable diseases such as asthma, obesity or autoimmune disorders. Future plans include data collection of participants until 12 years of participants’ age, which would allow to investigate how differences in omics immune and microbiome endotypes translate into population differences in clinical outcomes and susceptibility to RVIs.
Discussion
Considering the impact of the recent SARS-CoV-2 pandemic (65) or previous pandemics and epidemics due to IAV and RSV on human health (10, 66, 67), our knowledge about the ontogeny of immunity towards RVIs and the factors determining the developmental variability is little. Except for pre-existing co-morbidities and primary immunodeficiencies, hitherto, only the acquisition of autoantibodies against type I IFNs (68) and genetic variants, particularly in cytokines and pattern recognition receptors (69), could be linked to severe RVI cases, though only in a minor proportion of RVIs, and explain inter-population variability of anti-RVI immunity.
We recently initiated the recruitment of a birth cohort of healthy newborn infants in Germany that has been designed to investigate in particular the development of immunity towards RVIs in early childhood. The data shown in the present manuscript reflect an initial characterization of the MIAI cohort. Comparison of the structure of the current MIAI cohort with those reported from other recruiting maternal or term infant birth cohorts in Europe show similar distributions and rates of e.g. sex, gestational age, birth weight, multiple births, mode of delivery, first-time mothers, number of siblings, maternal age and BMI, gestational diabetes and hypertension, infections during pregnancy, and parental education and smoking (70–73). It is gratifying that in MIAI the drop-out rate of 8% is low in comparison to other birth cohorts which also perform repeated collection of biomaterials, e.g., the SweMaMi cohort (72) and KUNO-Kids cohort (70) that recorded 40-50% of lost 1-year follow-ups.
The common immunological finding in severe IAV, RSV and SARS-CoV-2 related infectious diseases is an exceeding cytokine release (74–77), suggesting a strong involvement of a systemic inflammatory response due to a lack of viral clearance at the epithelial barrier site or its breakage. Therefore, our first research focus in MIAI will lay on the age-dependent immunological profiling of the airway epithelium and blood monocytes, as key mediators of systemic cytokine storms during severe RVIs (78, 79), and the identification of demographic, clinical, environmental, lifestyle and dietary factors on the trajectories of their immune responsiveness toward RVIs. Interestingly, a previous study revealed a link between a low susceptibility of monocytes to IAV infections and high basal monocyte activation, which the authors assumed to be driven by environmental factors and/or weak-effect genetic variants that remain to be identified (80).
A plethora of studies report on distinct gut and respiratory microbiota compositions that are associated with an increased risk of RVIs with either IAV, RSV or SARS-CoV-2 (25, 27, 28, 30–32, 34, 81–83). How exactly such reported microbiota profiles influence anti-RVI immunity remains largely elusive. To what extent the developmental changes of the microbiota shape the postnatal ontogeny of the development of anti-RVI immunity is even less understood. The MIAI study will help to characterize differential trajectories of anti-RVI immunity and improve our knowledge about influencing factors, in particular the influence of the gut and respiratory microbiota.
The strength of the MIAI study is the population-based approach and the standardized in-depth longitudinal characterization of healthy individuals at birth and the comprehensive analysis of preserved biomaterials. The usage of cutting-edge multi-omics technology for the generation of longitudinal immune-microbiome profiles combined with functional profiling of the cellular immune response against RVIs make this cohort study a unique approach. Finally, by ensuring that the multi-omics-based profiles are integrated with highly granular clinical information, we are confident to identify factors that either promote or impede the development of immunity against RVIs. As in other population-based studies, we observe a bias toward high socioeconomic status among MIAI participants. Technically, the study has its limitation in the standardization of stool sample collection as sometimes home sampling is required, which precludes immediate freezing at -80°C as warranted by on-site visit sampling through the staff. With respect to clinical information, the study would benefit from an app-based monitoring that allows immediate reporting of acute infections that in case of respiratory symptoms then would prompt viral diagnostics. We aim at a timely implementation of this important add-on.
In conclusion, the MIAI birth cohort study expects to generate data that give a holistic picture on the variability of the maturation of immunity towards RVIs in early life and identify host and environmental factors that influence the trajectory. By linking differential trajectories along with clinical information on incidences and outcomes of RVIs to specific immune features, MIAI will pave the way for in-depth elucidation of molecular mechanisms, which is the basis for the development of age-specific host-directed treatment strategies against severe courses of RVIs (e.g., (anti-)cytokine biologicals, tailored cell- or RNA-technology-based therapies). Of particular importance will be the identification of factors shaping anti-RVI immunity, as this is essential to design personalized interventions such as metabolic, probiotic and/or dietary measures that promote developing competent anti-RVI immunity in early life thus preventing severe RVIs in the long term.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethics statement
The studies involving humans were approved by Ethical Review Committee, University of Würzburg, Würzburg, Germany. The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent for participation in this study was provided by the participants’ legal guardians/next of kin.
Author contributions
CRH: Conceptualization, Data curation, Formal Analysis, Project administration, Supervision, Writing – original draft, Writing – review & editing. RK: Conceptualization, Data curation, Writing – original draft, Writing – review & editing, Investigation. JS: Investigation, Writing – review & editing, Methodology, Supervision. MR: Investigation, Methodology, Writing – review & editing. MKW: Investigation, Methodology, Writing – review & editing, Conceptualization, Data curation, Supervision. SP: Conceptualization, Data curation, Methodology, Writing – review & editing. JH: Investigation, Methodology, Writing – review & editing. JD: Data curation, Investigation, Methodology, Writing – review & editing. HM: Writing – review & editing, Data curation, Investigation, Methodology. MK: Data curation, Investigation, Writing – review & editing. EF: Data curation, Investigation, Writing – review & editing. MW: Investigation, Writing – review & editing. JB: Investigation, Writing – review & editing. SS: Investigation, Methodology, Writing – review & editing. JF: Data curation, Investigation, Writing – review & editing. CV: Data curation, Investigation, Writing – review & editing. MM: Data curation, Investigation, Software, Writing – review & editing. JM: Data curation, Investigation, Writing – review & editing. MS: Methodology, Writing – review & editing. SH: Methodology, Writing – review & editing. TP: Methodology, Writing – review & editing. SB: Methodology, Writing – review & editing. LD: Methodology, Writing – review & editing. UL: Formal Analysis, Methodology, Software, Writing – review & editing. TK: Conceptualization, Software, Writing – review & editing. PH: Conceptualization, Software, Writing – review & editing. AW: Investigation, Writing – review & editing. S: Formal Analysis, Investigation, Methodology, Writing – review & editing. TU: Formal Analysis, Investigation, Methodology, Software, Supervision, Writing – review & editing. SF-S: Formal Analysis, Investigation, Methodology, Software, Supervision, Writing – review & editing. CH: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Writing – original draft, Writing – review & editing. DV: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work is supported by grants from Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) to DV (VI 538-9-1), CH (HA-6409-5-1) and TU (UL 521/1-1). The following other funding sources support the project: the Federal Ministry of Education and Research (BMBF) to SP, CH, and DV (PROSPER; 01EK2103B and 01EK2103A, respectively); the DFG to SP (PI 1512/1-3) and DV (VI 538/6-3); the DFG SFB 1583/1 (“DECIDE”) project number 492620490 to DV and SB; the DFG TRR 359 (“PILOT”) project number 491676693 to DV; the DFG Germany`s Excellence Strategy – EXC 2155 ‘RESIST’ – Project ID 390874280 to DV and TP; the BMBF Advanced Clinician Scientist Program (“INTERACT”, project number 01EO2108) to HM. We acknowledge the financial support of SKFS and UL from the German Research Foundation (DFG) under grants FO 1279/7-1 (Fetale Programmierung der Entwicklung des Mikrobioms und Konsequenzen für die Gehirnentwicklung), FO 1279/2-2, and FO 1279/1-2 as part of the Clinical Research Group “Nahrungsmittelallergie und Toleranz (Food@)” for subproject TP 09 - C2 “Data Analysis, Data Integration, and Rule-Based Modeling of Food Allergies.” Further support was provided by the European Union under the Horizon 2020 Research and Innovation Action (RIA) 101095540 IMMEDIATE for the project “Imminent Disease Prediction and Prevention at the Environment Host Interface,” and by the German Federal Ministry of Education and Research (BMBF) under grant 01EK2103C PROSPER for the project “Prevention of Sepsis through Personalized Nutritional Supplementation of S100A8/A9 in Vulnerable Neonates.
Acknowledgments
We would like to extend our gratitude towards families including the mothers, fathers and their infants included in the study, the sample collection team and recruitment team. This study would not have been possible without the work of a large group of staff members from the different departments. In particular, we acknowledge the contributions of the student assistants Lucia Baumgartner, Theresa Wurm and Leon Fagner, the entire team of midwives and the maternity unit including the lactation specialists.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2024.1443665/full#supplementary-material
References
1. Zhu G, Xu D, Zhang Y, Wang T, Zhang L, Gu W, et al. Epidemiological characteristics of four common respiratory viral infections in children. Virol J. (2021) 18:10. doi: 10.1186/s12985-020-01475-y
2. Nair H, Nokes DJ, Gessner BD, Dherani M, Madhi SA, Singleton RJ, et al. Global burden of acute lower respiratory infections due to respiratory syncytial virus in young children: a systematic review and meta-analysis. Lancet. (2010) 375:1545–55. doi: 10.1016/S0140-6736(10)60206-1
3. Openshaw PJ, Chiu C, Culley FJ, Johansson C. Protective and harmful immunity to RSV infection. Annu Rev Immunol. (2017) 35:501–32. doi: 10.1146/annurev-immunol-051116-052206
4. Shi T, McAllister DA, O’Brien KL, Simoes EA, Madhi SA, Gessner BD, et al. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in young children in 2015: a systematic review and modelling study. Lancet. (2017) 390:946–58. doi: 10.1016/S0140-6736(17)30938-8
5. Smyth RL, Openshaw PJ. Bronchiolitis. Lancet. (2006) 368:312–22. doi: 10.1016/S0140-6736(06)69077-6
6. Kristjansson S, Bjarnarson SP, Wennergren G, Palsdottir AH, Arnadottir T, Haraldsson A, et al. Respiratory syncytial virus and other respiratory viruses during the first 3 months of life promote a local TH2-like response. J Allergy Clin Immunol. (2005) 116:805–11. doi: 10.1016/j.jaci.2005.07.012
7. Legg JP, Hussain IR, Warner JA, Johnston SL, Warner JO. Type 1 and type 2 cytokine imbalance in acute respiratory syncytial virus bronchiolitis. Am J Respir Crit Care Med. (2003) 168:633–9. doi: 10.1164/rccm.200210-1148OC
8. Saravia J, You D, Shrestha B, Jaligama S, Siefker D, Lee GI, et al. Respiratory syncytial virus disease is mediated by age-variable IL-33. PloS Pathog. (2015) 11:e1005217. doi: 10.1371/journal.ppat.1005217
9. de Kleer IM, Kool M, de Bruijn MJW, Willart M, van Moorleghem J, Schuijs MJ, et al. Perinatal activation of the interleukin-33 pathway promotes type 2 immunity in the developing lung. Immunity. (2016) 45:1285–98. doi: 10.1016/j.immuni.2016.10.031
10. GBD 2017 Influenza Collaborators. Mortality, morbidity, and hospitalisations due to influenza lower respiratory tract infections, 2017: an analysis for the Global Burden of Disease Study 2017. Lancet Respir Med. (2019) 7:69–89. doi: 10.1016/S2213-2600(18)30496-X
11. Ulas T, Pirr S, Fehlhaber B, Bickes MS, Loof TG, Vogl T, et al. S100-alarmin-induced innate immune programming protects newborn infants from sepsis. Nat Immunol. (2017) 18:622–32. doi: 10.1038/ni.3745
12. Brook B, Harbeson D, Ben-Othman R, Viemann D, Kollmann TR. Newborn susceptibility to infection vs. disease depends on complex in vivo interactions of host and pathogen. Semin Immunopathol. (2017) 39:615–25. doi: 10.1007/s00281-017-0651-z
13. Henneke P, Kierdorf K, Hall LJ, Sperandio M, Hornef M. Perinatal development of innate immune topology. Elife. (2021) 10:e67793. doi: 10.7554/eLife.67793
14. CDC - Centers for Disease Control and Prevention. Influenza vaccination coverage for persons 6 months and older(2021). Available online at: https://www.cdc.gov/flu/fluvaxview/interactive-general-population.htm. (Accessed August 8, 2024).
15. Hoang U, Delanerolle G, Fan X, Aspden C, Byford R, Ashraf M, et al. A profile of influenza vaccine coverage for 2019-2020: database study of the english primary care sentinel cohort. JMIR Public Health Surveill. (2024) 10:e39297. doi: 10.2196/39297
16. Castagnoli R, Votto M, Licari A, Brambilla I, Bruno R, Perlini S, et al. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection in children and adolescents: A systematic review. JAMA Pediatr. (2020) 174:882–9. doi: 10.1001/jamapediatrics.2020.1467
17. Zhang C, Gu J, Chen Q, Deng N, Li J, Huang L, et al. Clinical and epidemiological characteristics of pediatric SARS-CoV-2 infections in China: A multicenter case series. PloS Med. (2020) 17:e1003130. doi: 10.1371/journal.pmed.1003130
18. Chou J, Thomas PG, Randolph AG. Immunology of SARS-CoV-2 infection in children. Nat Immunol. (2022) 23:177–85. doi: 10.1038/s41590-021-01123-9
19. Loske J, Röhmel J, Lukassen S, Stricker S, Magalhães VG, Liebig J, et al. Pre-activated antiviral innate immunity in the upper airways controls early SARS-CoV-2 infection in children. Nat Biotechnol. (2021) 40:319–24. doi: 10.1038/s41587-021-01037-9
20. Yoshida M, Worlock KB, Huang N, Lindeboom RG, Butler CR, Kumasaka N, et al. Local and systemic responses to SARS-CoV-2 infection in children and adults. Nature. (2022) 602:321–7. doi: 10.1038/s41586-021-04345-x
21. Woodall MN, Cujba A-M, Worlock KB, Case K-M, Masonou T, Yoshida M, et al. Age-specific nasal epithelial responses to SARS-CoV-2 infection. Nat Microbiol. (2024) 9:1293–311. doi: 10.1038/s41564-024-01658-1
22. Kollmann TR, Kampmann B, Mazmanian SK, Marchant A, Levy O. Protecting the newborn and young infant from infectious diseases: lessons from immune ontogeny. Immunity. (2017) 46:350–63. doi: 10.1016/j.immuni.2017.03.009
23. Torow N, Hand TW, Hornef MW. Programmed and environmental determinants driving neonatal mucosal immune development. Immunity. (2023) 56:485–99. doi: 10.1016/j.immuni.2023.02.013
24. Zhang X, Zhivaki D, Lo-Man R. Unique aspects of the perinatal immune system. Nat Rev Immunol. (2017) 17:495–507. doi: 10.1038/nri.2017.54
25. Lynch SV, Pedersen O. The human intestinal microbiome in health and disease. N Engl J Med. (2016) 375:2369–79. doi: 10.1056/NEJMra1600266
26. Di Simone SK, Rudloff I, Nold-Petry CA, Forster SC, Nold MF. Understanding respiratory microbiome-immune system interactions in health and disease. Sci Transl Med. (2023) 15:eabq5126. doi: 10.1126/scitranslmed.abq5126
27. Biesbroek G, Tsivtsivadze E, Sanders EA, Montijn R, Veenhoven RH, Keijser BJ, et al. Early respiratory microbiota composition determines bacterial succession patterns and respiratory health in children. Am J Respir Crit Care Med. (2014) 190:1283–92. doi: 10.1164/rccm.201407-1240OC
28. Bosch AATM, de Steenhuijsen Piters WAA, van Houten MA, Chu MLJN, Biesbroek G, Kool J, et al. Maturation of the infant respiratory microbiota, environmental drivers, and health consequences. A prospective cohort study. Am J Respir Crit Care Med. (2017) 196:1582–90. doi: 10.1164/rccm.201703-0554OC
29. de Steenhuijsen Piters WAA, Binkowska J, Bogaert D. Early life microbiota and respiratory tract infections. Cell Host Microbe. (2020) 28:223–32. doi: 10.1016/j.chom.2020.07.004
30. de Steenhuijsen Piters WAA, Watson RL, de Koff EM, Hasrat R, Arp K, Chu MLJN, et al. Early-life viral infections are associated with disadvantageous immune and microbiota profiles and recurrent respiratory infections. Nat Microbiol. (2022) 7:224–37. doi: 10.1038/s41564-021-01043-2
31. Teo SM, Mok D, Pham K, Kusel M, Serralha M, Troy N, et al. The infant nasopharyngeal microbiome impacts severity of lower respiratory infection and risk of asthma development. Cell Host Microbe. (2015) 17:704–15. doi: 10.1016/j.chom.2015.03.008
32. Antunes KH, Fachi JL, de Paula R, Da Silva EF, Pral LP, Dos Santos AÁ, et al. Microbiota-derived acetate protects against respiratory syncytial virus infection through a GPR43-type 1 interferon response. Nat Commun. (2019) 10:3273. doi: 10.1038/s41467-019-11152-6
33. Ngo VL, Lieber CM, Kang H-J, Sakamoto K, Kuczma M, Plemper RK, et al. Intestinal microbiota programming of alveolar macrophages influences severity of respiratory viral infection. Cell Host Microbe. (2024) 32:335–348.e8. doi: 10.1016/j.chom.2024.01.002
34. Harding JN, Siefker D, Vu L, You D, DeVincenzo J, Pierre JF, et al. Altered gut microbiota in infants is associated with respiratory syncytial virus disease severity. BMC Microbiol. (2020) 20:140. doi: 10.1186/s12866-020-01816-5
35. Zhao Y, Dong BR, Hao Q. Probiotics for preventing acute upper respiratory tract infections. Cochrane Database Syst Rev. (2022) 8:CD006895. doi: 10.1002/14651858.CD006895.pub4
36. Du T, Lei A, Zhang N, Zhu C. The beneficial role of probiotic lactobacillus in respiratory diseases. Front Immunol. (2022) 13:908010. doi: 10.3389/fimmu.2022.908010
37. Shi HY, Zhu X, Li WL, Mak JW, Wong SH, Zhu ST, et al. Modulation of gut microbiota protects against viral respiratory tract infections: a systematic review of animal and clinical studies. Eur J Nutr. (2021) 60:4151–74. doi: 10.1007/s00394-021-02519-x
38. Nesbitt H, Burke C, Haghi M. Manipulation of the upper respiratory microbiota to reduce incidence and severity of upper respiratory viral infections: A literature review. Front Microbiol. (2021) 12:713703. doi: 10.3389/fmicb.2021.713703
39. Harper A, Vijayakumar V, Ouwehand AC, Haar JT, Obis D, Espadaler J, et al. Viral infections, the microbiome, and probiotics. Front Cell Infect Microbiol. (2020) 10:596166. doi: 10.3389/fcimb.2020.596166
40. Suez J, Zmora N, Segal E, Elinav E. The pros, cons, and many unknowns of probiotics. Nat Med. (2019) 25:716–29. doi: 10.1038/s41591-019-0439-x
41. Rasmussen KM, Catalano PM, Yaktine AL. New guidelines for weight gain during pregnancy: what obstetrician/gynecologists should know. Curr Opin Obstet Gynecol. (2009) 21:521–6. doi: 10.1097/GCO.0b013e328332d24e
42. Martens A, Amann G, Schmidt K, Gaupmann R, Böhm B, Dehlink E, et al. An optimized, robust and reproducible protocol to generate well-differentiated primary nasal epithelial models from extremely premature infants. Sci Rep. (2019) 9:20069. doi: 10.1038/s41598-019-56737-9
43. Jonsdottir HR, Dijkman R. Characterization of human coronaviruses on well-differentiated human airway epithelial cell cultures. Methods Mol Biol. (2015) 1282:73–87. doi: 10.1007/978-1-4939-2438-7_8
44. Schreiber A, Viemann D, Schöning J, Schloer S, Mecate Zambrano A, Brunotte L, et al. The MEK1/2-inhibitor ATR-002 efficiently blocks SARS-CoV-2 propagation and alleviates pro-inflammatory cytokine/chemokine responses. Cell Mol Life Sci. (2022) 79:65. doi: 10.1007/s00018-021-04085-1
45. Sake SM, Zhang X, Rajak MK, Urbanek-Quaing M, Carpentier A, Gunesch AP, et al. Drug repurposing screen identifies lonafarnib as respiratory syncytial virus fusion protein inhibitor. Nat Commun. (2024) 15:1173. doi: 10.1038/s41467-024-45241-y
46. de Domenico E, Bonaguro L, Schulte-Schrepping J, Becker M, Händler K, Schultze JL. Optimized workflow for single-cell transcriptomics on infectious diseases including COVID-19. STAR Protoc. (2020) 1:100233. doi: 10.1016/j.xpro.2020.100233
47. Essaidi-Laziosi M, Brito F, Benaoudia S, Royston L, Cagno V, Fernandes-Rocha M, et al. Propagation of respiratory viruses in human airway epithelia reveals persistent virus-specific signatures. J Allergy Clin Immunol. (2018) 141:2074–84. doi: 10.1016/j.jaci.2017.07.018
48. Iverson E, Kaler L, Agostino EL, Song D, Duncan GA, Scull MA. Leveraging 3D model systems to understand viral interactions with the respiratory mucosa. Viruses. (2020) 12:1425. doi: 10.3390/v12121425
49. Viemann D. S100-alarmins are essential pilots of postnatal innate immune adaptation. Front Immunol. (2020) 11:688. doi: 10.3389/fimmu.2020.00688
50. Willers M, Ulas T, Völlger L, Vogl T, Heinemann AS, Pirr S, et al. S100A8 and S100A9 are important for postnatal development of gut microbiota and immune system in mice and infants. Gastroenterology. (2020) 159:2130–2145.e5. doi: 10.1053/j.gastro.2020.08.019
51. Dirks J, Andres O, Paul L, Manukjan G, Schulze H, Morbach H. IgD shapes the pre-immune naïve B cell compartment in humans. Front Immunol. (2023) 14:1096019. doi: 10.3389/fimmu.2023.1096019
52. Dirks J, Viemann D, Beyersdorf N, Härtel C, Morbach H. Insights into B-cell ontogeny inferred from human immunology. Eur J Immunol. (2023) 53:e2250116. doi: 10.1002/eji.202250116
53. León-Lara X, Fichtner AS, Willers M, Yang T, Schaper K, Riemann L, et al. γδ T cell profiling in a cohort of preterm infants reveals elevated frequencies of CD83+ γδ T cells in sepsis. J Exp Med. (2024) 221:e20231987. doi: 10.1084/jem.20231987
54. Morbach H, Eichhorn EM, Liese JG, Girschick HJ. Reference values for B cell subpopulations from infancy to adulthood. Clin Exp Immunol. (2010) 162:271–9. doi: 10.1111/j.1365-2249.2010.04206.x
55. Ravens S, Fichtner AS, Willers M, Torkornoo D, Pirr S, Schöning J, et al. Microbial exposure drives polyclonal expansion of innate γδ T cells immediately after birth. Proc Natl Acad Sci U.S.A. (2020) 117:18649–60. doi: 10.1073/pnas.1922588117
56. Austermann J, Friesenhagen J, Fassl SK, Petersen B, Ortkras T, Burgmann J, et al. Alarmins MRP8 and MRP14 induce stress tolerance in phagocytes under sterile inflammatory conditions. Cell Rep. (2014) 9:2112–23. doi: 10.1016/j.celrep.2014.11.020
57. Büttner M, Hempel F, Ryborz T, Theis FJ, Schultze JL. Pytometry: Flow and mass cytometry analytics in Python. bioRxiv preprint. (2022). doi: 10.1101/2022.10.10.511546
58. Kroeger C, Müller S, Leidner J, Kröber T, Warnat-Herresthal S, Spintge JB, et al. Unveiling the Power of High-Dimensional Cytometry Data with cyCONDOR. bioRxiv preprint (2024). doi: 10.1101/2024.02.29.582727
59. Holsten L, Dahm K, Oestreich M, Becker M, Ulas T. hCoCena: A toolbox for network-based co-expression analysis and horizontal integration of transcriptomic datasets. STAR Protoc. (2024) 5:102922. doi: 10.1016/j.xpro.2024.102922
60. Oestreich M, Holsten L, Agrawal S, Dahm K, Koch P, Jin H, et al. hCoCena: horizontal integration and analysis of transcriptomics datasets. Bioinformatics. (2022) 38:4727–34. doi: 10.1093/bioinformatics/btac589
61. Forslund SK, Chakaroun R, Zimmermann-Kogadeeva M, Markó L, Aron-Wisnewsky J, Nielsen T, et al. Combinatorial, additive and dose-dependent drug-microbiome associations. Nature. (2021) 600:500–5. doi: 10.1038/s41586-021-04177-9
62. Chen C-Y, Löber U, Forslund SK. LongDat: an R package for covariate-sensitive longitudinal analysis of high-dimensional data. Bioinform Adv. (2023) 3:vbad063. doi: 10.1093/bioadv/vbad063
63. Amir A, Erez-Granat O, Braun T, Sosnovski K, Hadar R, BenShoshan M, et al. Gut microbiome development in early childhood is affected by day care attendance. NPJ Biofilms Microbiomes. (2022) 8:2. doi: 10.1038/s41522-021-00265-w
64. Stewart CJ, Ajami NJ, O’Brien JL, Hutchinson DS, Smith DP, Wong MC, et al. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature. (2018) 562:583–8. doi: 10.1038/s41586-018-0617-x
65. Msemburi W, Karlinsky A, Knutson V, Aleshin-Guendel S, Chatterji S, Wakefield J. The WHO estimates of excess mortality associated with the COVID-19 pandemic. Nature. (2023) 613:130–7. doi: 10.1038/s41586-022-05522-2
66. Hall CB, Weinberg GA, Iwane MK, Blumkin AK, Edwards KM, Staat MA, et al. The burden of respiratory syncytial virus infection in young children. N Engl J Med. (2009) 360:588–98. doi: 10.1056/NEJMoa0804877
67. Li Y, Wang X, Blau DM, Caballero MT, Feikin DR, Gill CJ, et al. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in children younger than 5 years in 2019: a systematic analysis. Lancet. (2022) 399:2047–64. doi: 10.1016/S0140-6736(22)00478-0
68. Zhang Q, Bastard P, Cobat A, Casanova J-L. Human genetic and immunological determinants of critical COVID-19 pneumonia. Nature. (2022) 603:587–98. doi: 10.1038/s41586-022-04447-0
69. Forbester JL, Humphreys IR. Genetic influences on viral-induced cytokine responses in the lung. Mucosal Immunol. (2021) 14:14–25. doi: 10.1038/s41385-020-00355-6
70. Brandstetter S, Toncheva AA, Niggel J, Wolff C, Gran S, Seelbach-Göbel B, et al. KUNO-Kids birth cohort study: rationale, design, and cohort description. Mol Cell Pediatr. (2019) 6:1. doi: 10.1186/s40348-018-0088-z
71. Pangratz-Fuehrer S, Genzel-Boroviczény O, Bodensohn WE, Eisenburger R, Scharpenack J, Geyer PE, et al. Cohort profile: the MUNICH Preterm and Term Clinical study (MUNICH-PreTCl), a neonatal birth cohort with focus on prenatal and postnatal determinants of infant and childhood morbidity. BMJ Open. (2021) 11:e050652. doi: 10.1136/bmjopen-2021-050652
72. Fransson E, Gudnadottir U, Hugerth LW, Itzel EW, Hamsten M, Boulund F, et al. Cohort profile: the Swedish Maternal Microbiome project (SweMaMi) - assessing the dynamic associations between the microbiome and maternal and neonatal adverse events. BMJ Open. (2022) 12:e065825. doi: 10.1136/bmjopen-2022-065825
73. Pandolfini C, Clavenna A, Cartabia M, Campi R, Bonati M. National, longitudinal NASCITA birth cohort study to investigate the health of Italian children and potential influencing factors. BMJ Open. (2022) 12:e063394. doi: 10.1136/bmjopen-2022-063394
74. Bohmwald K, Gálvez NM, Canedo-Marroquín G, Pizarro-Ortega MS, Andrade-Parra C, Gómez-Santander F, et al. Contribution of cytokines to tissue damage during human respiratory syncytial virus infection. Front Immunol. (2019) 10:452. doi: 10.3389/fimmu.2019.00452
75. Dunning J, Blankley S, Hoang LT, Cox M, Graham CM, James PL, et al. Progression of whole-blood transcriptional signatures from interferon-induced to neutrophil-associated patterns in severe influenza. Nat Immunol. (2018) 19:625–35. doi: 10.1038/s41590-018-0111-5
76. Itoh Y, Shinya K, Kiso M, Watanabe T, Sakoda Y, Hatta M, et al. In vitro and in vivo characterization of new swine-origin H1N1 influenza viruses. Nature. (2009) 460:1021–5. doi: 10.1038/nature08260
77. To KK, Hung IF, Li IW, Lee K-L, Koo C-K, Yan W-W, et al. Delayed clearance of viral load and marked cytokine activation in severe cases of pandemic H1N1 2009 influenza virus infection. Clin Infect Dis. (2010) 50:850–9. doi: 10.1086/650581
78. Iwasaki A, Pillai PS. Innate immunity to influenza virus infection. Nat Rev Immunol. (2014) 14:315–28. doi: 10.1038/nri3665
79. Merad M, Martin JC. Pathological inflammation in patients with COVID-19: a key role for monocytes and macrophages. Nat Rev Immunol. (2020) 20:355–62. doi: 10.1038/s41577-020-0331-4
80. O’Neill MB, Quach H, Pothlichet J, Aquino Y, Bisiaux A, Zidane N, et al. Single-cell and bulk RNA-sequencing reveal differences in monocyte susceptibility to influenza A virus infection between Africans and Europeans. Front Immunol. (2021) 12:768189. doi: 10.3389/fimmu.2021.768189
81. Reyman M, van Houten MA, van Baarle D, Bosch AATM, Man WH, Chu MLJN, et al. Impact of delivery mode-associated gut microbiota dynamics on health in the first year of life. Nat Commun. (2019) 10:4997. doi: 10.1038/s41467-019-13014-7
82. Alba C, Aparicio M, González-Martínez F, González-Sánchez MI, Pérez-Moreno J, Del Toledo Castillo B, et al. Nasal and fecal microbiota and immunoprofiling of infants with and without RSV bronchiolitis. Front Microbiol. (2021) 12:667832. doi: 10.3389/fmicb.2021.667832
Keywords: birth cohort, respiratory viral infection, IAV, RSV, SARS-CoV-2, multi-omics data, anti-viral immunity, microbiota
Citation: Hartmann CR, Khan R, Schöning J, Richter M, Willers M, Pirr S, Heckmann J, Dirks J, Morbach H, Konrad M, Fries E, Winkler M, Büchel J, Seidenspinner S, Fischer J, Vollmuth C, Meinhardt M, Marissen J, Schmolke M, Haid S, Pietschmann T, Backes S, Dölken L, Löber U, Keil T, Heuschmann PU, Wöckel A, Sagar, Ulas T, Forslund-Startceva SK, Härtel C and Viemann D (2024) A clinical protocol for a German birth cohort study of the Maturation of Immunity Against respiratory viral Infections (MIAI). Front. Immunol. 15:1443665. doi: 10.3389/fimmu.2024.1443665
Received: 05 June 2024; Accepted: 29 August 2024;
Published: 17 September 2024.
Edited by:
Joann Diray-Arce, Harvard Medical School, United StatesReviewed by:
Lance Kendall Blevins, Michigan State University, United StatesSimon Daniel Van Haren, Boston Children’s Hospital and Harvard Medical School, United States
Copyright © 2024 Hartmann, Khan, Schöning, Richter, Willers, Pirr, Heckmann, Dirks, Morbach, Konrad, Fries, Winkler, Büchel, Seidenspinner, Fischer, Vollmuth, Meinhardt, Marissen, Schmolke, Haid, Pietschmann, Backes, Dölken, Löber, Keil, Heuschmann, Wöckel, Sagar, Ulas, Forslund-Startceva, Härtel and Viemann. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dorothee Viemann, dmllbWFubl9kQHVrdy5kZQ==
†These authors have contributed equally to this work and share first authorship
‡These authors have contributed equally to this work and share senior authorship