 Xiangbin Sun1†
Xiangbin Sun1† Zhanpeng Xie1†‡
Zhanpeng Xie1†‡ Zhen Wu1†
Zhen Wu1† Meiyang Song1Youxian Zhang1Zezhan Zhang1
Meiyang Song1Youxian Zhang1Zezhan Zhang1 Xinxin Cui1Aodi Liu1
Xinxin Cui1Aodi Liu1 Ke Li2*
Ke Li2*- 1Medical School of Shihezi University, Shihezi, China
- 2Department of Preventive Medicine, Medical School of Shihezi University, Shihezi, China
With the increasing number of people with HIV (PWH) and the use of antiretroviral treatment (ART) for PWH, HIV has gradually become a chronic infectious disease. However, some infected individuals develop issues with immunologic non-responses (INRs) after receiving ART, which can lead to secondary infections and seriously affect the life expectancy and quality of life of PWH. Disruption of the gut microbiota is an important factor in immune activation and inflammation in HIV/AIDS, thus stabilizing the gut microbiota to reduce immune activation and inflammation and promoting immune reconstitution may become a direction for the treatment of HIV/AIDS. This paper, based on extensive literature review, summarizes the definition, mechanisms, and solutions for INRs, starting from the perspective of gut microbiota.
1 Introduction
Acquired immunodeficiency syndrome (AIDS) is an infectious disease with immune deficiency caused by infection with HIV (1, 2). CD4+T cells are the target cells of HIV, the intestinal tract is a reservoir of HIV, Without using ART, resulting in low immune function of patients and subsequent death of various diseases (3, 4), for example, tumors, opportunistic infections. Recently, in ART used for PWH, HIV replication is suppressed, even HIV load below the detection level, but still about 15% -30% of patients in INRs (5), seriously affect the quality of life and life expectancy, INRs patients have higher non-AIDS events, such as the occurrence of malignant tumor (6, 7). With the development of research on INRs, more and more mechanisms of INRs have been discovered, treatments for INRs have been proposed, and better therapeutic results have been achieved in recent years. Therefore, this paper aims to provide direction for future solutions of INRs in PWH.
2 Definition of the INRs
Since the introduction of ART for PWH, the condition of PWH has been effectively controlled, resulting in improved quality of life and increased life expectancy (8). Additionally, there has been a significant reduction in viral load and a restoration or near normalization of CD4+ T cell counts, approaching levels seen in healthy individuals. However, there is still a subset of individuals in whom the CD4+ T cell count fails to increase or increases at a slow pace. This phenomenon is referred to as INRs (9, 10).
Currently, it is generally recognized that individuals who have been receiving ART for one year and have achieved undetectable viral RNA copies in plasma, but still have a CD4+ T cell count below 200 cells/ul, or have a CD4+ T cell count that has not increased by at least 20% from baseline, are classified as INRs. Additionally, there are individuals who, after receiving ART for 4-7 years, still have a CD4+ T cell count below 350 cells/ul, also considered as INRs (11, 12). However, the precise definition of INRs remains unclear.
3 The mechanism of the INRs
3.1 Time of infection and timing of treatment
Numerous studies have indicated that compared to younger patients, elderly patients are more likely to experience INRs (13). Research by Kiros et al. (14) demonstrated that the probability of INRs occurring in patients aged 50 and above is 1.97 times higher. This may be due to the thymic atrophy and function decline in elderly individuals, resulting in reduced immune function and diminished production of CD4+ T cells (15). Moreover, individuals with larger thymic volume are more likely to experience a recovery in CD4+ T cell counts, contributing to a lower likelihood of INRs. Additionally, the timing of initiating ART also impacts the occurrence of INRs. Thus, early initiation of ART is beneficial in reducing the incidence of INRs.
Zhang et al. (16) conducted a large-scale retrospective cohort study, which indicated that early initiation of ART is more advantageous in reducing the occurrence of INRs in PWH. Furthermore, the World Health Organization (WHO) recommended in 2016 that once diagnosed with HIV/AIDS, immediate initiation of ART is essential (17). The rationale behind this recommendation is that early initiation of ART is advantageous in reducing inflammatory responses and disturbances in gut microbiota, thereby lowering CD4+ T cell depletion and promoting immune restoration. Therefore, immediate treatment upon diagnosis of HIV/AIDS is crucial to prevent the occurrence of INRs.
3.2 Translocation of the gut microbiota
In individuals infected with the HIV, there is a significant increase in apoptosis of T cells in the intestines, leading to damage to the intestinal mucosal barrier and increased permeability. This imbalance ultimately results in dysbiosis of the gut microbiota and abnormalities in metabolite production, potentially causing increased intestinal permeability, chronic inflammation, and overactivation of the immune system (18). Studies indicate that following HIV infection, there is a shift in the predominant bacterial species in the gut from Bacteroidetes to Prevotella (19), influenced more by sexual behavior patterns (such as men who have sex with men) (MSM)rather than HIV infection itself (20). Additionally, HIV leads to decreased microbial diversity, enrichment of γ-β-Proteobacteria, and reduction in members of the Clostridia genus (21).
Overall, the gut microbiota in HIV-infected individuals shows a reduction in beneficial bacteria and an increase in harmful bacteria. Furthermore, HIV attacks the intestinal mucosa, further compromising the intestinal barrier and resulting in dysregulated microbial distribution, exacerbating inflammation and immune activation. Harmful bacteria such as Catenibacterium, Prevotellaceae, and Enterobacteriaceae can metabolize tryptophan, with metabolites such as indole and quinolinic acid associated with adverse health outcomes (22). Conversely, beneficial bacteria enhance intestinal barrier function by producing short-chain fatty acids, thereby reducing intestinal permeability and inflammation levels. Additionally, L-methylsulfonylmethane helps to reduce oxidative stress in the intestines (22). Beneficial bacteria also modulate host immune responses by affecting tryptophan metabolism rate and availability, producing beneficial metabolites like indole-3-acetic acid, which protects the host from the negative effects of excessive tryptophan on health (23) (Figure 1).
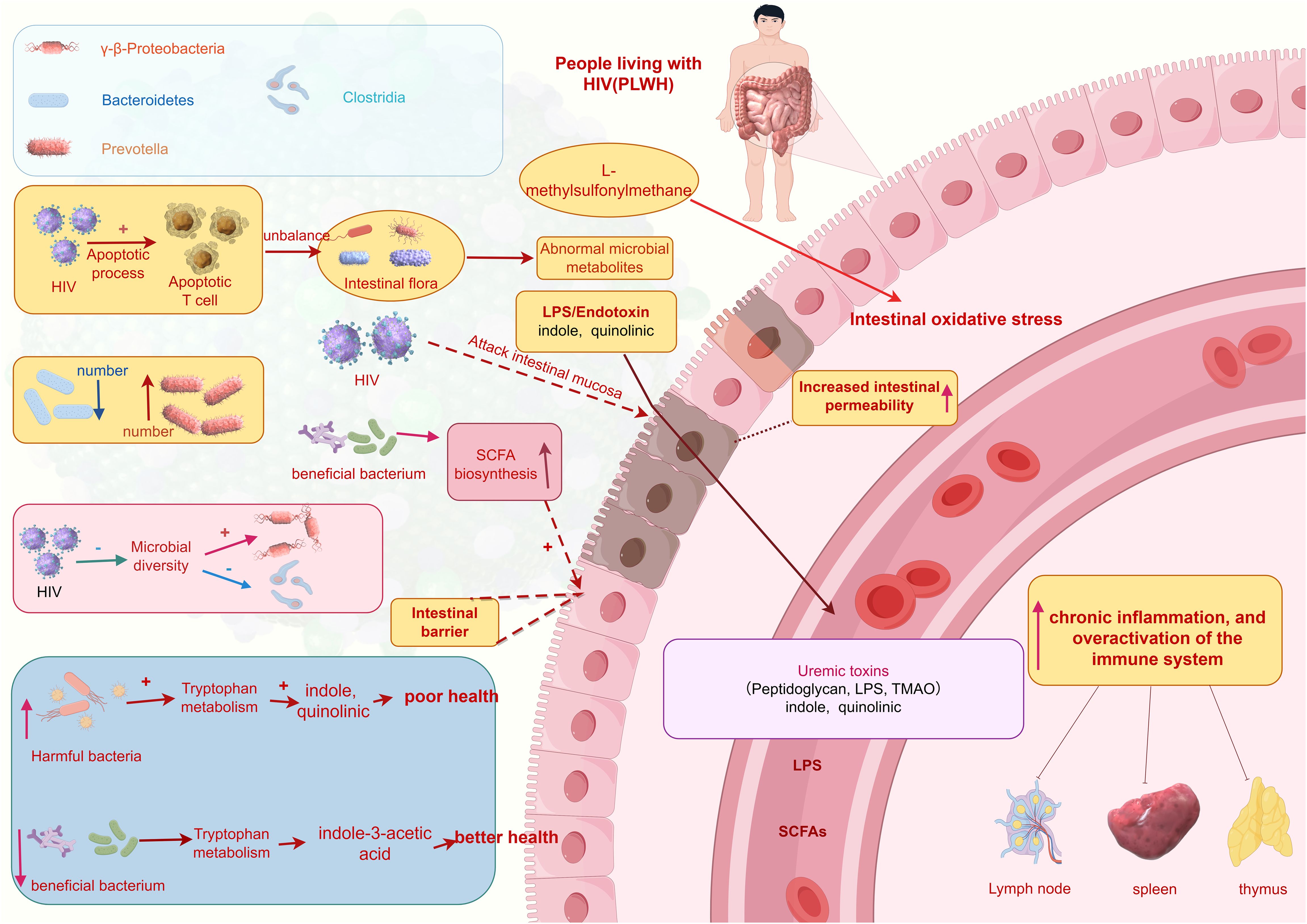
Figure 1. Translocation of the gut microbiota. (This figure is made by Figdraw.) (The patterns in the figure are only for effective differentiation and do not represent the actual appearance of the things referred to).
Heterotopic gut microbiota is an important factor affecting the degree of immune recovery after ART. Prevotellaceae is enriched in INRs, and its abundance is positively correlated with the activation of mucosal T cells (24). In addition, Ruminococcus is significantly reduced in INR, positively correlated with CD4+ T cell counts, and negatively correlated with serum pro-inflammatory cytokine levels (25). In addition, gut microbiota is also involved in the differentiation of T cell subsets (Th17 and higher regulatory T cells (Tregs)), and INRs has higher regulatory Tregs and lower Th17 percentage compared to immunological responders (IRs). The Th17/Treg ratio is negatively correlated with the levels of intestinal fatty acid binding proteins. Due to the continuous entry of bacterial lipopolysaccharides from the intestine into the bloodstream, immune activation and inflammation are exacerbated, leading to poor immune reconstitution.
3.3 Hyperactivation of the immune system
The disordered gut microbiota is characterized by an increase in pro-inflammatory bacterial species and a decrease in anti-inflammatory bacterial species (25, 26). Furthermore, bacterial lipopolysaccharides and inflammatory factors can both promote excessive immune system activation, virus replication, and the progression of HIV/AIDS, leading to damage to CD4+ T cells and the onset of non-AIDS defining illnesses (NADIs), thereby accelerating the course of HIV/AIDS. Excessive immune system activation is closely associated with mortality in HIV infection, and systemic chronic immune activation is considered to be a driving force behind CD4+ T cell depletion (27). Therefore, inhibiting excessive immune system activation may represent a new approach to addressing NADIs.
3.4 Platelet-T cell complexes
Dai et al. utilized flow cytometry analysis and immunofluorescence microscopy to observe that during HIV-1 infection, platelet-CD4+ T cell aggregates increase in treatment-naïve HIV-1-infected individuals (TNs) and INRs compared to healthy controls. However, the aggregation of platelet-CD4+ T cells in the INRs group did not decrease significantly compared to TNs and was associated with severe immune dysfunction. Platelet-CD4+ T cell aggregation positively correlated with HIV-1 viral load and negatively correlated with CD4+ T cell count and CD4+ T cell/CD8+ T cell ratio. CD45RO, HIV co-receptors, high levels of caspase-1 and caspase-3, and low levels of anti-apoptotic protein Bcl-2 were highly expressed in platelet-CD4+ T cell aggregates, which may contribute to CD4+ T cell depletion and sustained chronic inflammation in INRs (28). Zhu et al. recently demonstrated that platelets containing HIV in INRs can induce metabolic changes in CD4+ T cells through non-infectious mechanisms, specifically by forming platelet-CD4+ T cell aggregates (29–31). Enhanced glycolysis in immune cells and its contribution to ATP production are directly correlated with poor immune reconstitution. This effect specifically occurs in CD4+ T cells interacting with INRs platelets containing HIV. However, the mechanism behind this correlation may be due to INRs having a greater capacity for platelets to form aggregates with CD4+ T cells compared to IRs (28). Increased glycolysis is a crucial hallmark of T cell activation (32). Increased energy metabolism induced by platelets on T cells via INRs may contribute to the hyperactivation of CD4+ T cells, leading to CD4+ T cell exhaustion and triggering immune senescence associated with immune failure. Several mechanisms have been proposed to explain the immune modulation caused by platelet interaction with CD4+ T cells. Gerdes et al. suggest that platelet interactions with lymphocytes inhibit T cell proliferation and drive initial or memory CD4+ T cells towards regulatory (Treg: FoxP3+) or inflammatory (e.g., Th17) differentiation, thereby leading to immune failure (33). Platelets can induce polarization and/or secretion of chemotactic factors in lymphocytes through direct contact and interaction (34). Platelets can also release extracellular vesicles (microparticles) that directly interact with these lymphocytes, as well as myeloid cells and epithelial cells (35, 36). Functionally, these platelet-derived microvesicles can transfer active mRNA and microRNA (miRNA) to target cells, promoting the differentiation of Treg (34, 37) (Supplementary Figure S1).
3.5 Co-infection
Negash, Hadush et al. (38) have shown that the rate of immune reconstitution failure in HIV-infected individuals co-infected with tuberculosis is significantly higher compared to those without co-infection. In addition to Mycobacterium tuberculosis, studies have also indicated that co-infection with hepatitis viruses is a contributing factor to NADIs in PWH (39). The mechanisms through which co-infections result in NADIs involve inflammatory cytokines, which trigger excessive immune system activation. Moreover, elevated plasma lipopolysaccharides lead to dysbiosis and translocation of gut microbiota, reducing the quantity and diversity of anti-inflammatory bacteria, which compromises the integrity of the intestinal mucosa. As a result, this reduction in intestinal T cell populations contributes to the development of NADIs.
3.6 Adipose factor
In obese PWH, inflammation, fat accumulation, and dysfunction interact with each other. In recent years, adipose tissue has been found to play a crucial role in regulating immune function in the body (40, 41). In fact, adipose tissue is considered a potential reservoir for HIV, and the persistent presence of the virus in adipose tissue may be related to metabolic and immune dysfunction in adipose tissue cells (42, 43). Elena Yeregui et al. conducted a multicenter prospective study evaluating the association of adipose factors such as apelin, apelin receptor (APLNR), and zinc-alpha-2-glycoprotein (ZAG) with poor immune recovery in PWH undergoing ART. The study found that concentrations of APLNR and ZAG were significantly lower in immunological INRs compared to responders, and these lower levels persisted during the treatment follow-up period. Levels of ZAG were positively correlated with levels of retinol-binding protein 4 (RBP4), and low circulating RBP4 concentrations were associated with poor CD4+ T cell recovery (44). ZAG is a novel adipokine primarily expressed in visceral and subcutaneous adipose tissue (45). Its role in the immune system may be mediated through its anti-inflammatory effects on T cells and macrophages (46). In addition, pro-inflammatory adipokines like leptin and resistin released into circulation may suppress the expression of anti-inflammatory ZAG protein through activation of inflammatory pathways such as TNF signaling. Therefore, the imbalance between circulating anti-inflammatory and pro-inflammatory adipokines in untreated and treated PWH may be crucial for the reconstruction of CD4+ T cell counts and subsequent immune responses. Apelin is the endogenous ligand for the apelin receptor (APLNR), which is a protein receptor secreted by adipocytes. Interestingly, apelin inhibits HIV entry into human cells by binding to the co-receptor APLNR in both T-tropic and HIV-1 strains (44). In this context, both apelin and APLNR are positively correlated with CD4+ T cell counts, supporting their association with HIV replication. Therefore, lower circulating concentrations of apelin and apelin receptor in PWH with immune failure may suggest a lack of inhibition of HIV entry into cells, thereby indirectly linking immune failure to the “reservoir” of persistent HIV in adipose tissue (47, 48).
3.7 Host metabolic levels
Although viral load decreases to a certain extent after ART, incomplete immune reconstitution and the associated chronic non-AIDS-related diseases remain a focal point in the treatment and recovery of PWH. Changes in host metabolic levels accompany HIV infection, raising questions about whether they can return to normal after ART and their relationship with immune recovery, which is a critical research area in immunoreconstitution. Previous studies have observed high accumulation of plasma acylcarnitines in INRs (49), and a persistent decline in the activity of sphingosine-1-phosphate phosphatase 1 (50). Lu et al. conducted a metabolomics and machine learning analysis comparing the metabolic profiles of healthy controls and PWH, both before and after long-term ART. They found that disruptions in lipid and nucleotide metabolism observed during HIV infection did not return to normal levels post-treatment. Only three metabolites—maltose, N,N-dimethyl-5-aminovalerate, and decadienoic acid—showed significant differences between IRs and INRs. Additionally, Qian S et al. discovered significant increases in medium-chain acylcarnitine, palmitoylcarnitine, stearoylcarnitine, and oleoylcarnitine levels in INRs (49). Another study found that high-density lipoprotein cholesterol and larger-sized low-density lipoprotein particles contribute to better immune recovery post-treatment (51). However, the sample sizes of identified metabolites in both studies were less than 20, and their conclusions were inconsistent, lacking generalizability. Additionally, the pathogenic reasons remain unclear. Therefore, the potential biological mechanisms of the metabolites described in the above studies in immune rebuilding still require further robust validation.
3.8 Other factors
Ge, Y et al. (52) conducted a retrospective cohort study and found that the mode of sexual transmission can influence the immune reconstitution after ART. It was observed that heterosexual men are more likely to experience NADIs compared to men who have sex with men. The bone marrow and thymus are vital for T cell production, and their impaired functionality can result in impaired T cell output, thereby affecting immune reconstitution (12).
4 The solution to the INRs
4.1 Gut microbiota intervention
As mentioned earlier, obesity plays a significant role in inflammation and immune regulation. The use of ART has aligned the obesity rates among PWH with those of the general population (53). Additionally, research by Gogokhia et al. indicates that regardless of the ART regimen used, the incidence of obesity continues to rise (54). However, different ART drugs have varying impacts on weight gain due to their effects on the gut microbiota (55). Therefore, investigating the influence of different drugs on the gut microbiota is crucial.
Imahashi et al. demonstrated that NRTIs significantly increase the β-diversity of the gut microbiota while reducing its α-diversity in PWH. Long-term use of NRTIs also leads to an enrichment of Prevotella species bacteria and a reduction in Bacteroides species bacteria (56). Prevotella species are known as pro-inflammatory bacteria and currently dominate the gut microbiota of PWH. The increase in Prevotella abundance raises the pH of the gut, creating a more favorable environment for HIV infection and replication, thereby enhancing the potential for bacterial colonization and transmission amplification (57). On the other hand, Bacteroides species are typical anti-inflammatory bacteria. Additionally, PWH treated with NNRTIs or INSTIs also exhibit reduced α-diversity and increased β-diversity of their gut microbiota (58, 59).
Interestingly, current studies on the effects of Protease Inhibitors (PIs) on gut microbiota yield inconsistent results. Some suggest that compared to NNRTIs or INSTIs, PIs have minimal impact on gut microbiota (60), while others indicate that PI treatment significantly affects microbiota, correlating with higher plasma levels of soluble CD14 and I-FABP (61). These differences may be influenced by confounding factors such as diet, antibiotic use, lifestyle habits, and ethnicity affecting gut microbiota. Moreover, as mentioned earlier, the predominant gut microbiota shifts from Bacteroides to Prevotella in adult infection cases linked to sexual behavior (MSM), whereas in children with HIV infection, Prevotella enrichment is a microbiological feature (62). Therefore, future studies need to correct for these confounding factors to determine how microbiota influences HIV capability and transmission.
In conclusion, ART significantly reduces HIV viral load and restores CD4+ T cell counts, but immune function recovery remains limited in some patients. Thus, addressing immune restoration strategies from the perspective of gut microbiota is crucial for optimizing ART (Figure 2A).
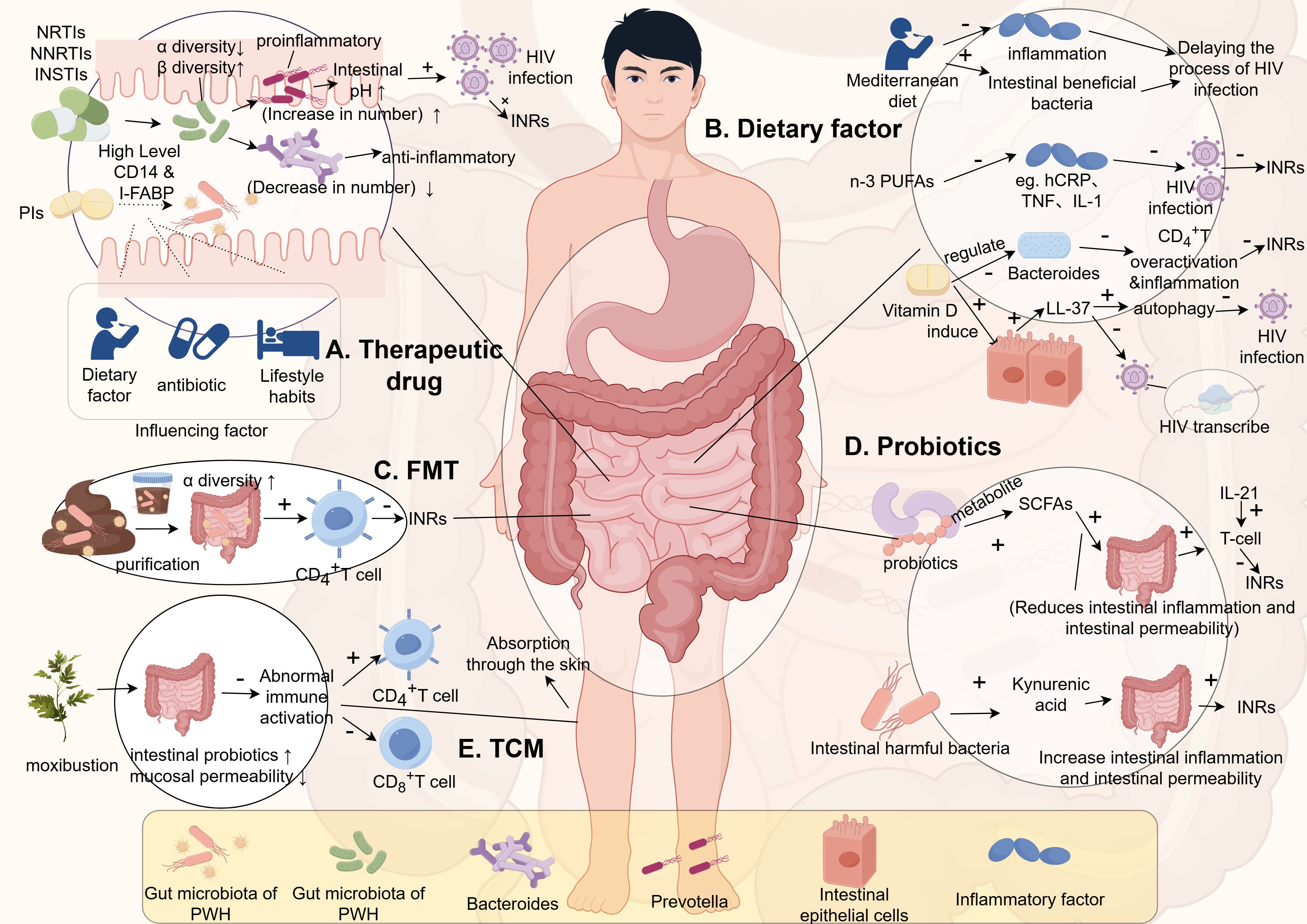
Figure 2. The relationship between gut microbiota and INRs. (This figure is made by Figdraw.) (The patterns in the figure are only for effective differentiation and do not represent the actual appearance of the things referred to). INRs, Immunologic non-responses; n-3 PUFAs, n-3 polyunsaturated fatty acids; FMT, fecal microbiota transplantation; SCFAs, short-chain fatty acids; TCM, Traditional Chinese Medicine; PWH, people living with HIV.
The gut is a target organ for HIV infection as it contains a large number of CD4+ T cells, making it a reservoir for the virus. HIV alters the abundance and composition of the gut microbiota, characterized by a reduction in beneficial bacteria, such as Bifidobacteria, and an increase in pathogenic bacteria. The gut microbiota plays a crucial role in maintaining the integrity of the gut mucosa and the functionality of the gut mucosal barrier, which is closely associated with the development of Th17 helper T cells. Accordingly, improving the gut microbiota by increasing the abundance of anti-inflammatory bacterial species and reducing the quantity of pro-inflammatory bacteria may enhance CD4+ T cell count and improve immune reconstitution.
In a clinical randomized controlled trial, d’Ettorre et al. (63) administered probiotics to HIV-infected patients twice daily for 48 weeks, resulting in a decrease in T lymphocyte immune activation and inflammatory markers. Additionally, in a randomized double-blind study by Mortezazadeh et al. (64), the consumption of probiotic yogurt was found to increase T cell levels. However, the impact of probiotic supplementation on T cell count remains inconsistent in different meta-analyses, with some studies suggesting a positive effect while others demonstrating contradictory results (65, 66).
Although the supplementation of probiotics or prebiotics alone cannot reduce HIV viral load, they can contribute to the reduction of immune system overactivation and inflammation caused by gut permeability. Therefore, supplementing with probiotics or prebiotics is beneficial for restoring gut barrier function (67), thereby slowing down the progression of HIV, reducing the occurrence of NADIs, increasing patient survival expectancy, and improving overall quality of life (68–73).
Some studies have indicated that the effects on T cells vary between the use of individual probiotics and combination therapies. Concurrent use of probiotics and IL-21 during probiotic supplementation has been shown to be more favorable for immune reconstitution. Hence, future research may consider exploring the ratio of relevant probiotics or prebiotics and their combination, as well as the effects of different types of probiotics and interleukins on immune reconstitution (Figure 2D).
It is important to note that different probiotics have varying effects, and the aforementioned meta-analyses did not classify the types of probiotics used, which may introduce bias. Moreover, current research standards for probiotics are not yet well-defined, such as intervention durations, which have varied from weeks to years across different studies. Thus, future research should establish clear guidelines regarding probiotic types, intervention durations, and other relevant criteria.
Some studies have suggested that metabolites produced by the gut microbiota can have different effects on the host’s immune system. Short-chain fatty acids (such as butyrate) produced by probiotics can help maintain gut mucosal barrier integrity, decrease inflammatory responses, and reduce immune system activation. This, in turn, can help prevent the loss of CD4+ T cells and contribute to immune reconstitution (74). On the other hand, uric acid produced by the gut microbiota can disrupt the gut mucosa, impair gut immunity, and contribute to INRs. Therefore, increasing the levels of short-chain fatty acids in the body, as an adjunct to ART therapy, may potentially reduce the occurrence of INRs (Figure 2D).
Currently, sequencing techniques for the gut microbiota mainly rely on genomics and metabolomics, with limited use of proteomics. Future research may require the integration of multiple “omics” approaches to determine the roles of different metabolites in HIV immune reconstitution. This comprehensive approach could provide new insights and potential solutions for addressing INRs.
In recent years, fecal microbiota transplantation (FMT) has shown promising results in improving the gut microbiota. FMT has been found to significantly increase the alpha diversity of the gut microbiota, which is decreased in HIV-infected individuals (75). Furthermore, individuals with lower CD4+ T cell counts also exhibit reduced alpha diversity of their gut microbiota, indicating a significant correlation between gut microbiota diversity and immune status. Moreover, studies have demonstrated that FMT can significantly increase CD4+ T cell counts (76), suggesting that transplanting probiotics into the gut of HIV-infected individuals may reduce the occurrence of INRs. Plasma levels of inflammation-associated proteins in INRs were significantly higher than those in IRs. Moreover, these biomarkers were negatively correlated with the CD4+ cell count and positively correlated with the HIV viral load (77). A recent authoritative omics study published by Diaz-Garcia, C. et al. demonstrated that FMT could change the gut microbiota by targeting (the Ruminococcaceae, Succinivibrionaceae, Prevotellaceae families, and the Clostridium genus, etc.) to effectively reduce the levels of 46 inflammatory proteins such as IL6 in PWH (78). Repeated oral FMT significantly enhanced the levels of intestinal fatty acid-binding protein (IFABP), which has been demonstrated to be an independent predictor of mortality as a biomarker of intestinal damage (79). Therefore, the more stable intestinal barrier and the lower inflammation level formed by FMT might further decrease the level of INRs in PWH or reduce the early occurrence of INRs. With a stricter and more standardized study design and a more innovative proteomic assay, this research constitutes an important step in the exploration of longitudinal associations between fecal bacteria and plasma-associated proteins. However, considering the complex immune system regulation and the unclear influence mechanism of FMT on PWH, we should further clarify the regulatory mechanism and functional level of the targeted gut microbiome in PWH inflammation through transcriptomics and metabolomics in the future. However, there are still many selective and controversial issues that need to be addressed regarding FMT. For example, it is essential to investigate whether the newly formed microbiota after transplantation can be maintained in a stable state for the long term, rather than just in the short term. Additionally, it is important to determine if the transplanted microbiota is highly compatible with the host, rather than exacerbating immune reactions and stimulating immune system activation. Factors influencing the stability of the transplanted microbiota also need to be studied. Therefore, large-scale longitudinal studies are needed in the future to explore the role of FMT in immune reconstitution in HIV-infected individuals. However, neither FMT nor the intervention of probiotics and prebiotics has provided clear evidence for reducing chronic inflammation and immune system activation in HIV. Further research is still required to explore these areas. Additionally, the gut microbiota consists not only of bacteria and fungi but also of viral communities. Compared with non-pathogenic SIV, enteric virome expansion has been found in pathogenic SIV, and plays a role in immune deficiency (80). Transplanting the gut viral community from healthy individuals has shown benefits in treating recurrent Clostridium difficile infection, alleviating diet-induced obesity, and preventing necrotizing enterocolitis in premature infants (81). Therefore, whether immune deficiency is involved in PWH by enteric virome expansion. However, there are currently no reports on the transplantation of the viral community in PWH. Therefore, investigating the effects of viral community transplantation on the immune deficiencies of PWH should be explored in the future (Figure 2C).
4.2 Traditional Chinese medicine intervention
TCM can enhance patients’ immune function, providing a new potential solution for individuals with INRs (82). The gastrointestinal tract serves not only as an immune organ but also as a digestive organ. Following absorption in the gastrointestinal tract, TCM can impact the metabolism of the gut microbiota. Simultaneously, the gut microbiota influences the bioavailability of TCM, promoting the proliferation of beneficial bacteria while reducing the abundance of harmful bacteria (83, 84).
Mechanistic studies on TCM therapy for PWH primarily focus on alterations in the gut microbiota, damage to the intestinal mucosal barrier, and changes in CD4+ T lymphocytes (85, 86). TCM has the capacity to enhance patients’ immune function, reduce complications, and improve survival rates and quality of life (87).
The combined use of traditional Chinese medicine and ART for PWH can offer complementary advantages (88). Adopting an integrated approach to treatment involving both TCM and ART can provide valuable insights into immune reconstitution and viral reservoir clearance in PWH (89).
Moxibustion therapy mainly achieves its therapeutic effects through the warmth and stimulation produced by the burning of moxa, as well as the absorption of moxa particles and aroma by the skin or through inhalation. Moxibustion offers the advantage of multi-targeted and bidirectional immune regulation in the human body. It can correct abnormal immune responses in the intestines, increase the population of beneficial probiotics in the gut microbiota, and reduce the permeability of the intestinal mucosa through the metabolic byproducts of probiotics (90). Additionally, it can lower the occurrence of abnormal immune responses, maintain intestinal stability, and prevent excessive T-cell activation. Therefore, moxibustion has a direct or indirect impact on the intestines of PWH. Moxibustion can increase the CD4+ T cell count in patients, while also reducing the number of CD8+ T cells, restoring the normal ratio of CD4+ T cells to CD8+ T cells, enhancing immune function, reducing the production of inflammatory factors, and improving INRs. Furthermore, moxibustion is a non-invasive treatment that can effectively prevent healthcare-associated infections caused by bloodborne pathogens in the context of HIV (Figure 2E).
In recent years, research on the use of Artesunate for PWH has become increasingly frequent. Chen et al. (91) conducted a clinical randomized controlled trial, administering Artesunate orally to PWH. After 48 weeks, the study found a reduction in T-cell activation markers and a decrease in T-cell apoptosis levels. However, there was no improvement in T-cell count. On the other hand, Artesunate also improved the gut microbiota of infected individuals, increasing the abundance of probiotics such as Actinobacteria and Bifidobacterium. Moreover, a higher level of CD4+ T cells in peripheral blood was associated with an increased level of Actinobacteria in the gut microbiota. Additionally, Artesunate appeared to have no significant clinical effect on immune reconstitution in younger individuals, but had a more pronounced effect in middle-aged and older individuals, suggesting that Artesunate may improve thymic atrophy and therefore contribute to the improvement of INRs.
Liu et al. (92) administered Tripterygium wilfordii to PWH with immune non-response for 17 months, and observed an increase in CD4+ T cell count along with a decrease in T cell activation. Through multi-omics studies, the mechanism was found to involve the inhibition of the interferon signaling pathway. Genomics is currently widely used in research on the role of the gut microbiota in HIV/AIDS. However, genomics has focused excessively on microbiota differences, and genomics and transcriptomics are primarily used for functional predictions. Nevertheless, predicted functions do not necessarily translate into actual expression. Therefore, future research should incorporate transcriptomics and proteomics to specifically investigate their roles and underlying mechanisms (93).
4.3 Dietary intervention
Diet is also an important pathway for altering the gut microbiota. Manzano et al. (94) compared the differential effects of the Mediterranean diet and the Western diet for PWH treatment and found that the Mediterranean diet can reduce inflammation levels while preserving beneficial bacteria. Although no specific dietary pattern has been found to increase CD4+ T cell counts, dietary changes can lower inflammation levels and slow the progression of HIV/AIDS. Currently, there is a lack of specific dietary interventions for the treatment of PWH, and further research is needed to explore this area in the future.
4.3.1 Vitamin D
In recent years, multiple studies have demonstrated that nutrient compounds such as vitamin D, which possess immune-regulatory properties, play a significant role in maintaining intestinal homeostasis through their effects on both innate and adaptive immunity (95, 96). Vitamin D can induce the antimicrobial peptide LL-37 from epithelial cells and immune cells (97). LL-37 exhibits multifaceted protective effects that can be enhanced by vitamin D (98). In the intestinal mucosal barrier, the production of LL-37 serves as an important natural defense mechanism primarily by activating autophagy to delay the progression of HIV disease. Autophagy reduces intracellular HIV replication (99). Alternatively, it can delay infection by inhibiting HIV-1 transcription (100, 101). Additionally, vitamin D can stabilize the tight junction structure of intestinal epithelial cells (102). And may also regulate the composition of human gut microbiota. Recent studies suggest that vitamin D may modulate the relative abundance of pro-inflammatory Bacteroides species in the gut (103, 104). Raftery et al. and Ponda et al. separately found that vitamin D treatment is associated with reduced systemic inflammation levels and disease activity in inflammatory bowel disease and chronic kidney disease (105). However, Missailidis et al., using randomized controlled trials, found that vitamin D + phenylbutyrate supplementation did not improve markers of intestinal inflammation or gut microbiota composition in treatment-naive individuals with active HIV-1 replication. This lack of effect may be partly attributed to ongoing viral replication in untreated HIV. Future studies should investigate the supplementation of vitamin D in individuals receiving ART for HIV infection more extensively, to better assess its regulatory role in a controlled viral environment (106) (Figure 2B).
4.3.2 Reduction of the refined sugars and saturated fatty acids
The progression of HIV infection is often associated with various metabolic and cardiovascular complications (107). These complications may be related to the side effects of ART, but HIV infection itself, even during successful ART, can induce metabolic changes through mechanisms such as chronic low-grade systemic inflammation (108, 109). Fatty acids (FAs), whether free or as part of other lipids such as triglycerides, phospholipids, sphingolipids, and sterol lipids, play critical roles in cellular energy metabolism and are essential components of cell membranes. FAs also have the ability to signal through peroxisome proliferator-activated receptors (110, 111). Disruption in the composition of FAs has been recognized to impact the development of various metabolic, cardiovascular, and inflammatory diseases (112). For example, certain polyunsaturated fatty acids (PUFAs) such as arachidonic acid (AA; C20:4n-6), eicosapentaenoic acid (EPA; C20:5n-3), and docosahexaenoic acid (DHA; C22:6n-3) serve as precursors for the synthesis of biologically active lipid mediators (e.g., prostaglandins [PGs], leukotrienes [LTs], lipoxins, and resolvins) (113, 114), mediating both inflammatory and anti-inflammatory effects (114, 115). There is also significant focus on the central role of fatty acids in regulating immune cell function, emphasizing their direct impact on many cellular processes involved in T cell responses and antigen presentation (116–118). There is evidence suggesting that FA metabolism is disrupted in PWH. Additionally, a recent meta-analysis indicated that supplementing n-3 polyunsaturated fatty acids (n-3 PUFA) may alleviate inflammation in PWH, as assessed by levels of C-reactive protein in patients receiving ART (119). In an earlier meta-analysis, it was shown that supplementation with n-3 polyunsaturated fatty acids can lower triglyceride levels in PWH undergoing ART (120). Furthermore, several studies suggest that n-3 polyunsaturated fatty acids (n-3 PUFA) may inhibit classical inflammatory cytokines such as tumor necrosis factor (TNF), interleukin (IL)-1, and IL-6 (121–123), and are associated with HIV-related pathogenic mechanisms involving these cytokines (124). Therefore, PWH at any stage should consume foods rich in unsaturated fatty acids and minimize intake of refined sugars and saturated fats commonly found in Western diets (125) (Figure 2B).
5 Summary
In recent years, the number of PWH has been increasing year after year, and research in the area of INRs has gradually intensified. In this paper, we formulated the definition of INRs, expounded the mechanism underlying the association between gut microbiota translocation, platelet-CD4+ T cell coupling, and host metabolism with immune failure in INRs, and further explored potential therapeutic interventions for INRs gut microbiota. In the future, we will endeavor to provide a more distinct and comprehensive definition of INRs. However, at present, there is a dearth of large-scale data and evidence from evidence-based medical research to support approaches targeting INRs. Therefore, future studies should focus on the mechanism of the gut microbiota for immune reconstitution in INRs and make use of advanced technologies such as multi-omics analysis to conduct in-depth research in all aspects of this field.
Author contributions
XS: Investigation, Writing – original draft, Writing – review & editing. ZX: Writing – original draft, Writing – review & editing, Conceptualization, Methodology, Supervision, Investigation. ZW: Investigation, Writing – original draft. MS: Investigation, Writing – review & editing. YZ: Conceptualization, Methodology, Supervision, Writing – review & editing. ZZ: Investigation, Writing – review & editing. XC: Investigation, Writing – review & editing. AL: Conceptualization, Methodology, Supervision, Writing – review & editing. KL: Conceptualization, Methodology, Supervision, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was supported by Xinjiang Production and Construction Corps Social Science Project (21YB08) and Shihezi University (Philosophy and Social Science) Fund (ZZZC202147).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2024.1378431/full#supplementary-material
References
1. Chen X, Lai Y. Knowledge domain and emerging trends in HIV pre-exposure prophylaxis: A visualization analysis via CiteSpace. Front Microbiol. (2023) 14:1099132. doi: 10.3389/fmicb.2023.1099132
2. Du X, Zhang Q, Hao J, Gong X, Liu J, Chen J. Global trends in depression among patients living with HIV: A bibliometric analysis. Front Psychol. (2023) 14:1125300. doi: 10.3389/fpsyg.2023.1125300
3. Wang D, Zhou M, Wang P, Zhang J, Mi Y, Cheng F, et al. Treatment-naïve people living with HIV aged 50 years or older in Beijing, China, 2010-2020: joinpoint regression model analysis of surveillance data. J Int AIDS Soc. (2023) 26:e26193. doi: 10.1002/jia2.26193
4. Sponaugle A, Weideman AMK, Ranek J, Atassi G, Kuruc J, Adimora AA, et al. Dominant CD4+T cell receptors remain stable throughout antiretroviral therapy-mediated immune restoration in people with HIV. Cell rep Medicine. (2023) 4:101268. doi: 10.1016/j.xcrm.2023.101268
5. Wan LY, Huang HH, Zhen C, Chen SY, Song B, Cao WJ, et al. Distinct inflammation-related proteins associated with T cell immune recovery during chronic HIV-1 infection. Emerging Microbes infect. (2023) 12:2150566. doi: 10.1080/22221751.2022.2150566
6. Chen C, Wang J, Xun J, Zhang X, Liu L, Song Z, et al. Role of thymosin α1 in restoring immune response in immunological nonresponders living with HIV. BMC Infect Dis. (2024) 24:97. doi: 10.1186/s12879-024-08985-y
7. Guo XY, Guo YT, Wang ZR, Jiao YM, Hu Y, Fan LN, et al. Severe intestinal barrier damage in HIV-infected immunological non-responders. Heliyon. (2023) 9:e20790. doi: 10.1016/j.heliyon.2023.e20790
8. Wen CH, Lu PL, Lin CY, Lin YP, Chen TC, Chen YH, et al. Effect of immunological non-response on incidence of Non-AIDS events in people living with HIV: A retrospective multicenter cohort study in Taiwan. J microbiol immunol infect = Wei mian yu gan ran za zhi. (2023) 56:977–87. doi: 10.1016/j.jmii.2023.06.005
9. Wang YT, Ji SX, Xia XY, Wan TJ, Zou JX, Li BX, et al. Proteomic investigation and biomarker identification of lung and spleen deficiency syndrome in HIV/AIDS immunological nonresponders. J Thorac Dis. (2023) 15:1460–72. doi: 10.21037/jtd-23-322
10. Geng ST, Zhang JB, Wang YX, Xu Y, Lu D, Zhang Z, et al. Pre-digested protein enteral nutritional supplementation enhances recovery of CD4+ T cells and repair of intestinal barrier in HIV-infected immunological non-responders. Front Immunol. (2021) 12:757935. doi: 10.3389/fimmu.2021.757935
11. Fan L, Li P, Yu A, Liu D, Wang Z, Wu Y, et al. Prevalence of and prognosis for poor immunological recovery by virally suppressed and aged HIV-infected patients. Front Med. (2023) 10:1259871. doi: 10.3389/fmed.2023.1259871
12. Zhang W, Ruan L. Recent advances in poor HIV immune reconstitution: what will the future look like? Front Microbiol. (2023) 14:1236460. doi: 10.3389/fmicb.2023.1236460
13. Tanaka H, Wada T, Ohyama KO, Ishii T. Analysis of the time-to-onset and factors affecting clinical outcomes of immune reconstitution inflammatory syndrome in people living with HIV using data from the Japanese spontaneous reporting database. J Pharm Pharm sci: Publ Can Soc Pharm Sci Societe Can Des Sci Pharm. (2021) 24:153–60. doi: 10.18433/jpps31675
14. Kiros T, Taye A, Workineh L, Eyayu T, Damtie S, Hailemichael W, et al. Immuno-virological status and its associated factors among HIV-positive patients receiving highly active antiretroviral therapy at delgi primary hospital, northwest Ethiopia, 2020/2021: A cross-sectional study. Heliyon. (2022) 8:e10169. doi: 10.1016/j.heliyon.2022.e10169
15. Scherpenisse M, Kootstra NA, Bakker M, Berkhout B, Pasternak AO. Cell-associated HIV-1 unspliced-to-multiply-spliced RNA ratio at 12 weeks of ART predicts immune reconstitution on therapy. mBio. (2021) 12:e00099-21. doi: 10.1128/mBio.00099-21
16. Zhang W, Yan J, Luo H, Wang X, Ruan L. Incomplete immune reconstitution and its predictors in people living with HIV in Wuhan, China. BMC Public Health. (2023) 23:1808. doi: 10.1186/s12889-023-16738-w
17. Guideline on When to Start Antiretroviral Therapy and on Pre-Exposure Prophylaxis for HIV. World Health Organization (2015).
18. Ashuro AA, Lobie TA, Ye DQ, Leng RX, Li BZ, Pan HF, et al. Review on the alteration of gut microbiota: the role of HIV infection and old age. AIDS Res Hum Retroviruses. (2020) 36:556–65. doi: 10.1089/AID.2019.0282
19. Russo E, Nannini G, Sterrantino G, Kiros ST, Di Pilato V, Coppi M, et al. Effects of viremia and CD4 recovery on gut “microbiome-immunity” axis in treatment-naïve HIV-1-infected patients undergoing antiretroviral therapy. World J Gastroenterol. (2022) 28:635–52. doi: 10.3748/wjg.v28.i6.635
20. Satish S, Abu Y, Gomez D, Kumar Dutta R, Roy S. HIV, opioid use, and alterations to the gut microbiome: elucidating independent and synergistic effects. Front Immunol. (2023) 14:1156862. doi: 10.3389/fimmu.2023.1156862
21. Vujkovic-Cvijin I, Sortino O, Verheij E, Sklar J, Wit FW, Kootstra NA, et al. HIV-associated gut dysbiosis is independent of sexual practice and correlates with noncommunicable diseases. Nat Commun. (2020) 11:2448. doi: 10.1038/s41467-020-16222-8
22. Singh S, Giron LB, Shaikh MW, Shankaran S, Engen PA, Bogin ZR, et al. Distinct intestinal microbial signatures linked to accelerated systemic and intestinal biological aging. Microbiome. (2024) 12:31. doi: 10.1186/s40168-024-01758-4
23. MacCann R, Landay AL, Mallon PWG. HIV and comorbidities - the importance of gut inflammation and the kynurenine pathway. Curr Opin HIV AIDS. (2023) 18:102–10. doi: 10.1097/COH.0000000000000782
24. Wang Z, Zhen C, Guo X, Qu M, Zhang C, Song J, et al. Landscape of gut mucosal immune cells showed gap of follicular or memory B cells into plasma cells in immunological non-responders. Clin Trans Med. (2024) 14:e1699. doi: 10.1002/ctm2.1699
25. Lu D, Zhang JB, Wang YX, Geng ST, Zhang Z, Xu Y, et al. Association between CD4+ T cell counts and gut microbiota and serum cytokines levels in HIV-infected immunological non-responders. BMC Infect Dis. (2021) 21:742. doi: 10.1186/s12879-021-06491-z
26. Shukla S, Kumari S, Bal SK, Monaco DC, Ribeiro SP, Sekaly RP, et al. Go”, “No Go,” or “Where to Go”; does microbiota dictate T cell exhaustion, programming, and HIV persistence? Curr Opin HIV AIDS. (2021) 16:215–22. doi: 10.1097/COH.0000000000000692
27. Lu W, Feng Y, Jing F, Han Y, Lyu N, Liu F, et al. Association between gut microbiota and CD4 recovery in HIV-1 infected patients. Front Microbiol. (2018) 9:1451. doi: 10.3389/fmicb.2018.01451
28. Dai XP, Wu FY, Cui C, Liao XJ, Jiao YM, Zhang C, et al. Increased platelet-CD4+ T cell aggregates are correlated with HIV-1 permissiveness and CD4+ T cell loss. Front Immunol. (2021) 12:799124. doi: 10.3389/fimmu.2021.799124
29. Zhu A, Real F, Zhu J, Greffe S, de Truchis P, Rouveix E, et al. HIV-sheltering platelets from immunological non-responders induce a dysfunctional glycolytic CD4+ T-cell profile. Front Immunol. (2022) 12:781923. doi: 10.3389/fimmu.2021.781923
30. Real F, Capron C, Sennepin A, Arrigucci R, Zhu A, Sannier G, et al. Platelets from HIV-infected individuals on antiretroviral drug therapy with poor CD4+ T cell recovery can harbor replication-competent HIV despite viral suppression. Sci Trans Med. (2020) 12:eaat6263. doi: 10.1126/scitranslmed.aat6263
31. Palmer CS, Henstridge DC, Yu D, Singh A, Balderson B, Duette G, et al. Emerging role and characterization of immunometabolism: relevance to HIV pathogenesis, serious non-AIDS events, and a cure. J Immunol (Baltimore Md.: 1950). (2016) 196:4437–44. doi: 10.4049/jimmunol.1600120
32. Soto-Heredero G, Gomez de Las Heras MM, Gabande-Rodriguez E, Oller J, Mittelbrunn M. Glycolysis - a key player in the inflammatory response. FEBSJ. (2020) 287:3350–69. doi: 10.1111/febs.15327
33. Zhu A, Real F, Zhu J, Greffe S, de Truchis P, Rouveix E, et al. HIV-sheltering platelets from immunological non-responders induce a dysfunctional glycolytic CD4 T-cell profile. Front Immunol. (2022) 12:781923+. doi: 10.3389/fimmu.2021.781923+
34. D’Ambrosi S, Nilsson RJ, Wurdinger T. Platelets and tumor-associated RNA transfer. Blood. (2021) 137:3181–91. doi: 10.1182/blood.2019003978
35. Yan C, Wu H, Fang X, He J, Zhu F. Platelet, a key regulator of innate and adaptive immunity. Front Med. (2023) 10:1074878. doi: 10.3389/fmed.2023.1074878
36. El Filaly H, Mabrouk M, Atifi F, Guessous F, Akarid K, Merhi Y, et al. Dissecting platelet’s role in viral infection: A double-edged effector of the immune system. Int J Mol Sci. (2023) 24:2009. doi: 10.3390/ijms24032009
37. Leng Q, Ding J, Dai M, Liu L, Fang Q, Wang DW, et al. Insights into platelet-derived microRNAs in cardiovascular and oncologic diseases: potential predictor and therapeutic target. Front Cardiovasc Med. (2022) 9:879351. doi: 10.3389/fcvm.2022.879351
38. Negash H, Legese H, Tefera M, Mardu F, Tesfay K, Gebresilasie S, et al. The effect of tuberculosis on immune reconstitution among HIV patients on highly active antiretroviral therapy in Adigrat general hospital, eastern Tigrai, Ethiopia; 2019: a retrospective follow up study. BMC Immunol. (2019) 20:45. doi: 10.1186/s12865-019-0327-7
39. Bloch M, John M, Smith D, Rasmussen TA, Wright E. Managing HIV-associated inflammation and ageing in the era of modern ART. HIV Med. (2020) 21:2–16. doi: 10.1111/hiv.12952
40. Blüher M, Mantzoros CS. From leptin to other adipokines in health and disease: facts and expectations at the beginning of the 21st century. Metabolism: Clin Exp. (2015) 64:131–45. doi: 10.1016/j.metabol.2014.10.016
41. Maximus PS, Al Achkar Z, Hamid PF, Hasnain SS, Peralta CA. Adipocytokines: are they the theory of everything? Cytokine. (2020) 133:155144. doi: 10.1016/j.cyto.2020.155144
42. Couturier J, Suliburk JW, Brown JM, Luke DJ, Agarwal N, Yu X, et al. Human adipose tissue as a reservoir for memory CD4+ T cells and HIV. AIDS. (2015) 29:667–74. doi: 10.1097/QAD.0000000000000599
43. Damouche A, Lazure T, Avettand-Fènoël V, Huot N, Dejucq-Rainsford N, Satie AP, et al. Adipose tissue is a neglected viral reservoir and an inflammatory site during chronic HIV and SIV infection. PloS Pathog. (2015) 11:e1005153. doi: 10.1371/journal.ppat.1005153
44. Yeregui E, Masip J, Viladés C, Domingo P, Pacheco YM, Blanco J, et al. Adipokines as new biomarkers of immune recovery: apelin receptor, RBP4 and ZAG are related to CD4+ T-cell reconstitution in PLHIV on suppressive antiretroviral therapy. Int J Mol Sci. (2022) 23:2202. doi: 10.3390/ijms23042202
45. Lei L, Li K, Li L, Fang X, Zhou T, Zhang C, et al. Circulating zinc-α2-glycoprotein levels are low in newly diagnosed patients with metabolic syndrome and correlate with adiponectin. Nutr Metab. (2017) 14:53. doi: 10.1186/s12986-017-0210-6
46. Noh JY, Shin JU, Kim JH, Kim SH, Kim BM, Kim YH, et al. ZAG regulates the skin barrier and immunity in atopic dermatitis. J Invest Dermatol. (2019) 139:1648–1657.e7. doi: 10.1016/j.jid.2019.01.023
47. Tincati C, Merlini E, Braidotti P, Ancona G, Savi F, Tosi D, et al. Impaired gut junctional complexes feature late-treated individuals with suboptimal CD4+ T-cell recovery upon virologically suppressive combination antiretroviral therapy. AIDS (London England). (2016) 30:991–1003. doi: 10.1097/QAD.0000000000001015
48. Zhang LX, Song JW, Zhang C, Fan X, Huang HH, Xu RN, et al. Dynamics of HIV reservoir decay and naïve CD4 T-cell recovery between immune non-responders and complete responders on long-term antiretroviral treatment. Clin Immunol (Orlando Fla.). (2021) 229:108773. doi: 10.1016/j.clim.2021.108773
49. Qian S, Chen X, Wu T, Sun Y, Li X, Fu Y, et al. The accumulation of plasma acylcarnitines are associated with poor immune recovery in HIV-infected individuals. BMC Infect Dis. (2021) 21:808. doi: 10.1186/s12879-021-06525-6
50. Scarpellini B, Zanoni M, Sucupira MC, Truong HM, Janini LM, Segurado ID, et al. Plasma metabolomics biosignature according to HIV stage of infection, pace of disease progression, viremia level and immunological response to treatment. PloS One. (2016) 11:e0161920. doi: 10.1371/journal.pone.0161920
51. Rodriguez-Gallego E, Gomez J, Pacheco YM, Peraire J, Vilades C, Beltran-Debon R, et al. A baseline metabolomic signature is associated with immunological CD4+ T-cell recovery after 36 months of antiretroviral therapy in HIV-infected patients. AIDS. (2018) 32:565–73. doi: 10.1097/QAD.0000000000001730
52. Ge Y, Zhou Y, Liu Y, Lu J, Qiu T, Shi LE, et al. Immune reconstitution efficacy after combination antiretroviral therapy in male HIV-1 infected patients with homosexual and heterosexual transmission. Emerging Microbes infect. (2023) 12:2214250. doi: 10.1080/22221751.2023.2214250
53. Cook RR, Fulcher JA, Tobin NH, Li F, Lee D, Woodward C, et al. Combined effects of HIV and obesity on the gastrointestinal microbiome of young men who have sex with men. HIV Med. (2020) 21:365–77. doi: 10.1111/hiv.12838
54. Gogokhia L, Taur Y, Juluru K, Yagan N, Zhu YS, Pamer E, et al. Intestinal dysbiosis and markers of systemic inflammation in viscerally and generally obese persons living with HIV. J acquired Immune deficiency syndromes (1999). (2020) 83:81–9. doi: 10.1097/QAI.0000000000002229
55. Baltazar-Díaz TA, Amador-Lara F, Andrade-Villanueva JF, González-Hernández LA, Cabrera-Silva RI, Sánchez-Reyes K, et al. Gut bacterial communities in HIV-infected individuals with metabolic syndrome: effects of the therapy with integrase strand transfer inhibitor-based and protease inhibitor-based regimens. Microorganisms. (2023) 11:951. doi: 10.3390/microorganisms11040951
56. Imahashi M, Ode H, Kobayashi A, Nemoto M, Matsuda M, Hashiba C, et al. Impact of long-term antiretroviral therapy on gut and oral microbiotas in HIV-1-infected patients. Sci Rep. (2021) 11:960. doi: 10.1038/s41598-020-80247-8
57. Meng P, Zhang G, Ma X, Ding X, Song X, Dang S, et al. Traditional Chinese medicine (Xielikang) reduces diarrhea symptoms in acquired immune deficiency syndrome (AIDS) patients by regulating the intestinal microbiota. Front Microbiol. (2024) 15:1346955. doi: 10.3389/fmicb.2024.1346955
58. Ray S, Narayanan A, Giske CG, Neogi U, Sönnerborg A, Nowak P. Altered gut microbiome under antiretroviral therapy: impact of efavirenz and zidovudine. ACS Infect Dis. (2021) 7:1104–15. doi: 10.1021/acsinfecdis.0c00536
59. Baltazar-Díaz TA, Andrade-Villanueva JF, Sánchez-Álvarez P, Amador-Lara F, Holguín-Aguirre T, Sánchez-Reyes K, et al. A two-faced gut microbiome: butyrogenic and proinflammatory bacteria predominate in the intestinal milieu of people living with HIV from Western Mexico. Int J Mol Sci. (2024) 25:4830. doi: 10.3390/ijms25094830
60. Ancona G, Merlini E, Tincati C, Barassi A, Calcagno A, Augello M, et al. Long-term suppressive cART is not sufficient to restore intestinal permeability and gut microbiota compositional changes. Front Immunol. (2021) 12:639291. doi: 10.3389/fimmu.2021.639291
61. Pinto-Cardoso S, Lozupone C, Briceño O, Alva-Hernández S, Téllez N, Adriana A, et al. Fecal Bacterial Communities in treated HIV infected individuals on two antiretroviral regimens. Sci Rep. (2017) 7:43741. doi: 10.1038/srep43741
62. Sainz T, Gosalbes MJ, Talavera-Rodríguez A, Jimenez-Hernandez N, Prieto L, Escosa L, et al. Effect of a nutritional intervention on the intestinal microbiota of vertically HIV-infected children: the pediabiota study. Nutrients. (2020) 12:2112. doi: 10.3390/nu12072112
63. d’Ettorre G, Ceccarelli G, Giustini N, Serafino S, Calantone N, De Girolamo G, et al. Probiotics reduce inflammation in antiretroviral treated, HIV-infected individuals: results of the “Probio-HIV” Clinical trial. PloS One. (2015) 10:e0137200. doi: 10.1371/journal.pone.0137200
64. Mortezazadeh M, Kalantari S, Abolghasemi N, Ranjbar M, Ebrahimi S, Mofidi A, et al. The effect of oral probiotics on CD4 count in patients with HIV infection undergoing treatment with ART who have had an immunological failure. Immunity Inflammation Dis. (2023) 11:e913. doi: 10.1002/iid3.913
65. Sachdeva M, Sra HK, Agarwal A, Chauhan A, Pradhan P, Singh M, et al. Effect of probiotics on the frequency of CD4+ T-cells in HIV-infected children and adolescents: A systematic review and meta-analysis of randomized controlled trials. J Trop Pediatr. (2022) 68:fmac006. doi: 10.1093/tropej/fmac006
66. Zhang XL, Chen MH, Geng ST, Yu J, Kuang YQ, Luo HY, et al. Effects of probiotics on diarrhea and CD4 cell count in people living with HIV: A systematic review and meta-analysis. Front Pharmacol. (2021) 12:570520. doi: 10.3389/fphar.2021.570520
67. Arnbjerg CJ, Vestad B, Hov JR, Pedersen KK, Jespersen S, Johannesen HH, et al. Effect of lactobacillus rhamnosus GG supplementation on intestinal inflammation assessed by PET/MRI scans and gut microbiota composition in HIV-infected individuals. J acquired Immune deficiency syndromes (1999). (2018) 78:450–7. doi: 10.1097/QAI.0000000000001693
68. Villar-Garcia J, Hernandez JJ, Guerri-Fernandez R, Gonzalez A, Lerma E, Guelar A, et al. Effect of probiotics (Saccharomyces boulardii) on microbial translocation and inflammation in HIV-treated patients: a double-blind, randomized, placebo-controlled trial. J Acquir Immune Defic Syndr. (2015) 68:256–63. doi: 10.1097/QAI.0000000000000468
69. Serrano-Villar S, Vazquez-Castellanos JF, Vallejo A, Latorre A, Sainz T, Ferrando-Martinez S, et al. The effects of prebiotics on microbial dysbiosis, butyrate production and immunity in HIV-infected subjects. Mucosal Immunol. (2017) 10:1279–93. doi: 10.1038/mi.2016.122
70. Brenchley JM, Serrano-Villar S. From dysbiosis to defense: harnessing the gut microbiome in HIV/SIV therapy. Microbiome. (2024) 12:113. doi: 10.1186/s40168-024-01825-w
71. D’Ettorre G, Rossi G, Scagnolari C, Andreotti M, Giustini N, Serafino S, et al. Probiotic supplementation promotes a reduction in T-cell activation, an increase in Th17 frequencies, and a recovery of intestinal epithelium integrity and mitochondrial morphology in ART-treated HIV-1-positive patients. Immun Inflamm Dis. (2017) 5:244–60. doi: 10.1002/iid3.160
72. Falasca K, Vecchiet J, Ucciferri C, Di Nicola M, D’angelo C, Reale M. Effect of probiotic supplement on cytokine levels in HIV-infected individuals: a preliminary study. Nutrients. (2015) 7:8335–47. doi: 10.3390/nu7105396
73. Ishizaki A, Bi X, Nguyen LV, Matsuda K, Pham HV, Phan CTT, et al. Effects of short-term probiotic ingestion on immune profiles and microbial translocation among HIV-1-infected Vietnamese children. Int J Mol Sci. (2017) 18:2185. doi: 10.3390/ijms18102185
74. Hishiya N, Uno K, Nakano A, Konishi M, Higashi S, Eguchi S, et al. Association between the gut microbiome and organic acid profiles in a Japanese population with HIV infection. J infect chemother. (2024) 30:58–66. doi: 10.1016/j.jiac.2023.09.013
75. Geng ST, Zhang ZY, Wang YX, Lu D, Yu J, Zhang JB, et al. Regulation of gut microbiota on immune reconstitution in patients with acquired immunodeficiency syndrome. Front Microbiol. (2020) 11:594820. doi: 10.3389/fmicb.2020.594820
76. Ouyang J, Isnard S, Lin J, Fombuena B, Peng X, Nair Parvathy S, et al. Treating from the inside out: relevance of fecal microbiota transplantation to counteract gut damage in GVHD and HIV infection. Front Med. (2020) 7:421. doi: 10.3389/fmed.2020.00421
77. Guo X, Wang Z, Qu M, Guo Y, Yu M, Hong W, et al. Abnormal blood microbiota profiles are associated with inflammation and immune restoration in HIV/AIDS individuals. mSystems. (2023) 8:e0046723. doi: 10.1128/msystems.00467-23
78. Díaz-García C, Moreno E, Talavera-Rodríguez A, Martín-Fernández L, González-Bodí S, Martín-Pedraza L, et al. Fecal microbiota transplantation alters the proteomic landscape of inflammation in HIV: identifying bacterial drivers. Microbiome. (2024) 12:214. doi: 10.1186/s40168-024-01919-5
79. Serrano-Villar S, Talavera-Rodríguez A, Gosalbes MJ, Madrid N, Pérez-Molina JA, Elliott RJ, et al. Fecal microbiota transplantation in HIV: A pilot placebo-controlled study. Nat Commun. (2021) 12:1139. doi: 10.1038/s41467-021-21472-1
80. Handley SA, Thackray LB, Zhao G, Presti R, Miller AD, Droit L, et al. Pathogenic simian immunodeficiency virus infection is associated with expansion of the enteric virome. Cell. (2012) 151:253–66. doi: 10.1016/j.cell.2012.09.024
81. Shah SA, Deng L, Thorsen J, Pedersen AG, Dion MB, Castro-Mejía JL, et al. Expanding known viral diversity in the healthy infant gut. Nat Microbiol. (2023) 8:986–98. doi: 10.1038/s41564-023-01345-7
82. Wan T, Liu X, Su Y, Zou J, Wu X, Jiang C, et al. Biological differentiation of traditional Chinese medicine from excessive to deficient syndromes in AIDS: Comparative microRNA microarray profiling and syndrome-specific biomarker identification. J Med Virol. (2021) 93:3634–46. doi: 10.1002/jmv.26704
83. Zhang S, Wang Y, Lu F, Mohammed SAD, Liu H, Ding S, et al. Mechanism of action of Shenerjiangzhi formulation on hyperlipidemia induced by consumption of a high-fat diet in rats using network pharmacology and analyses of the gut microbiota. Front Pharmacol. (2022) 13:745074. doi: 10.3389/fphar.2022.745074
84. Li K, Jiang Y, Wang N, Lai L, Xu S, Xia T, et al. Traditional Chinese medicine in osteoporosis intervention and the related regulatory mechanism of gut microbiome. Am J Chin Med. (2023) 51:1957–81. doi: 10.1142/S0192415X23500866
85. Su LL, Li X, Guo ZJ, Xiao XY, Chen P, Zhang JB, et al. Effects of different steaming times on the composition, structure and immune activity of Polygonatum Polysaccharide. J ethnopharmacol. (2023) 310:116351. doi: 10.1016/j.jep.2023.116351
86. Wu X, Chen H, Gao X, Gao H, He Q, Li G, et al. Natural herbal remedy wumei decoction ameliorates intestinal mucosal inflammation by inhibiting th1/th17 cell differentiation and maintaining microbial homeostasis. Inflammatory bowel Dis. (2022) 28:1061–71. doi: 10.1093/ibd/izab348
87. Ding X, Meng P, Ma X, Cui W, Li L, Song X, et al. Integrated traditional Chinese medicine intervention for delaying HIV morbidity: study protocol for a multicentre randomised controlled trial. Trials. (2022) 23:665. doi: 10.1186/s13063-022-06625-x
88. Liu Z. Treating older patients with AIDS using Traditional Chinese Medicine Combined with Conventional Western Medicine in China. Aging Dis. (2021) 12:1872–8. doi: 10.14336/AD.2021.0925
89. Tao Z, Huang XJ, Liu Y, Wang R, Dong JP, Liang BY, et al. Efficacy of integrated Traditional Chinese Medicine and anti-retroviral therapy on immunological nonresponse in patients with human immunodeficiency virus/acquired immunodeficiency syndrome: a Meta-analysis of randomized controlled trial. J tradit Chin Med = Chung i tsa chih ying wen pan. (2021) 41:669–76. doi: 10.19852/j.cnki.jtcm.2021.05.002
90. Jia YJ, Li TY, Han P, Chen Y, Pan LJ, Jia CS. Effects of different courses of moxibustion treatment on intestinal flora and inflammation of a rat model of knee osteoarthritis. J Integr Med. (2022) 20:173–81. doi: 10.1016/j.joim.2022.01.004
91. Chen S, Xu Q, Wang J, Tan X. Effects of artesunate tablet on immune activation and reconstitution among highly active antiretroviral therapy-treated patients with incomplete immune responses. AIDS Res Hum Retroviruses. (2022) 38:100–10. doi: 10.1089/AID.2020.0254
92. Liu X, Lin L, Lv T, Lu L, Li X, Han Y, et al. Combined multi-omics and network pharmacology approach reveals the role of Tripterygium Wilfordii Hook F in treating HIV immunological non-responders. Phytomedicine. (2022) 101:154103. doi: 10.1016/j.phymed.2022.154103
93. Nganou-Makamdop K, Douek DC. The gut and the translocated microbiomes in HIV infection: current concepts and future avenues. Pathog Immun. (2024) 9:168–94. doi: 10.20411/pai.v9i1.693
94. Manzano M, Talavera-Rodríguez A, Moreno E, Madrid N, Gosalbes MJ, Ron R, et al. Relationship of diet to gut microbiota and inflammatory biomarkers in people with HIV. Nutrients. (2022) 14:1221. doi: 10.3390/nu14061221
95. Pitman MC, Meagher N, Price DJ, Rhodes A, Chang JJ, Scher B, et al. Effect of high dose vitamin D3 on the HIV-1 reservoir: A pilot randomised controlled trial. J Virus eradication. (2023) 9:100345. doi: 10.1016/j.jve.2023.100345
96. Wang J, Mei L, Hao Y, Xu Y, Yang Q, Dai Z, et al. Contemporary perspectives on the role of vitamin D in enhancing gut health and its implications for preventing and managing intestinal diseases. Nutrients. (2024) 16:2352. doi: 10.3390/nu16142352
97. Wang TT, Nestel FP, Bourdeau V, Nagai Y, Wang Q, Liao J, et al. Cutting edge: 1,25-dihydroxyvitamin D3 is a direct inducer of antimicrobial peptide gene expression. J Immunol (Baltimore Md.: 1950). (2004) 173:2909–12. doi: 10.4049/jimmunol.173.5.2909
98. Vandamme D, Landuyt B, Luyten W, Schoofs L. A comprehensive summary of LL-37, the factotum human cathelicidin peptide. Cell Immunol. (2012) 280:22–35. doi: 10.1016/j.cellimm.2012.11.009
99. Campbell GR, Spector SA. Autophagy induction by vitamin D inhibits both Mycobacterium tuberculosis and human immunodeficiency virus type 1. Autophagy. (2012) 8:1523–5. doi: 10.4161/auto.21154
100. Wang G, Watson KM, Buckheit RW Jr. Anti-human immunodeficiency virus type 1 activities of antimicrobial peptides derived from human and bovine cathelicidins. Antimicrobial Agents chemother. (2008) 52:3438–40. doi: 10.1128/AAC.00452-08
101. Aguilar-Jimenez W, Zapata W, Rugeles MT. Antiviral molecules correlate with vitamin D pathway genes and are associated with natural resistance to HIV-1 infection. Microbes infect. (2016) 18:510–6. doi: 10.1016/j.micinf.2016.03.015
102. Kong J, Zhang Z, Musch MW, Ning G, Sun J, Hart J, et al. Novel role of the vitamin D receptor in maintaining the integrity of the intestinal mucosal barrier. Am J Physiol Gastrointestinal liver Physiol. (2008) 294:G208–16. doi: 10.1152/ajpgi.00398.2007
103. Kanhere M, He J, Chassaing B, Ziegler TR, Alvarez JA, Ivie EA, et al. Bolus weekly vitamin D3 supplementation impacts gut and airway microbiota in adults with cystic fibrosis: A double-blind, randomized, placebo-controlled clinical trial. J Clin Endocrinol Metab. (2018) 103:564–74. doi: 10.1210/jc.2017-01983
104. Bashir M, Prietl B, Tauschmann M, Mautner SI, Kump PK, Treiber G, et al. Effects of high doses of vitamin D3 on mucosa-associated gut microbiome vary between regions of the human gastrointestinal tract. Eur J Nutr. (2016) 55:1479–89. doi: 10.1007/s00394-015-0966-2
105. Raftery T, Martineau AR, Greiller CL, Ghosh S, McNamara D, Bennett K, et al. Effects of vitamin D supplementation on intestinal permeability, cathelicidin and disease markers in Crohn's disease: Results from a randomised double-blind placebo-controlled study. United Eur Gastroenterol J. (2015) 3:294–302. doi: 10.1177/2050640615572176
106. Missailidis C, Sørensen N, Ashenafi S, Amogne W, Kassa E, Bekele A, et al. Vitamin D and phenylbutyrate supplementation does not modulate gut derived immune activation in HIV-1. Nutrients. (2019) 11:1675. doi: 10.3390/nu11071675
107. Ramos SR, Reynolds H, Johnson C, Melkus G, Kershaw T, Thayer JF, et al. Perceptions of HIV-related comorbidities and usability of a virtual environment for cardiovascular disease prevention education in sexual minority men with HIV: formative phases of a pilot randomized controlled trial. J Med Internet Res. (2024) 26:e57351. doi: 10.2196/57351
108. Ueland T, Waagsbø B, Berge RK, Trøseid M, Aukrust P, Damås JK. Fatty acids composition and HIV infection: altered levels of n-6 polyunsaturated fatty acids are associated with disease progression. Viruses. (2023) 15:1613. doi: 10.3390/v15071613
109. Flynn JK, Ortiz AM, Vujkovic-Cvijin I, Welles HC, Simpson J, Castello Casta FM, et al. Translocating bacteria in SIV infection are not stochastic and preferentially express cytosine methyltransferases. Mucosal Immunol. (2024) 17:1089–101. doi: 10.1016/j.mucimm.2024.07.008
110. Zhang H, Xie Y, Cao F, Song X. Gut microbiota-derived fatty acid and sterol metabolites: biotransformation and immunomodulatory functions. Gut Microbes. (2024) 16:2382336. doi: 10.1080/19490976.2024.2382336
111. Galli G, Corsetto PA, Proto C, Lo Russo G, Ganzinelli M, Rulli E, et al. Circulating fatty acid profile as a biomarker for immunotherapy in advanced non-small cell lung cancer. Clin Lung Cancer. (2022) 23:e489–99. doi: 10.1016/j.cllc.2022.07.010
112. Lankinen MA, Stančáková A, Uusitupa M, Ågren J, Pihlajamäki J, Kuusisto J, et al. Plasma fatty acids as predictors of glycaemia and type 2 diabetes. Diabetologia. (2015) 58:2533–44. doi: 10.1007/s00125-015-3730-5
113. Kotlyarov S, Kotlyarova A. Anti-inflammatory function of fatty acids and involvement of their metabolites in the resolution of inflammation in chronic obstructive pulmonary disease. Int J Mol Sci. (2021) 22:12803. doi: 10.3390/ijms222312803
114. Das UN. Cell membrane theory of senescence" and the role of bioactive lipids in aging, and aging associated diseases and their therapeutic implications. Biomolecules. (2021) 11:241. doi: 10.3390/biom11020241
115. Kaur P, Gao J, Wang Z. Liposomal formulations enhance the anti-inflammatory effect of eicosapentaenoic acid in HL60 cells. Pharmaceutics. (2022) 14:520. doi: 10.3390/pharmaceutics14030520
116. Lee J, Moraes-Vieira PM, Castoldi A, Aryal P, Yee EU, Vickers C, et al. Branched fatty acid esters of hydroxy fatty acids (FAHFAs) protect against colitis by regulating gut innate and adaptive immune responses. J Biol Chem. (2016) 291:22207–17. doi: 10.1074/jbc.M115.703835
117. Whelan J, Gowdy KM, Shaikh SR. N-3 polyunsaturated fatty acids modulate B cell activity in pre-clinical models: Implications for the immune response to infections. Eur J Pharmacol. (2016) 785:10–7. doi: 10.1016/j.ejphar.2015.03.100
118. Garcia AM, Bishop EL, Li D, Jeffery LE, Garten A, Thakker A, et al. Tolerogenic effects of 1,25-dihydroxyvitamin D on dendritic cells involve induction of fatty acid synthesis. J Steroid Biochem Mol Biol. (2021) 211:105891. doi: 10.1016/j.jsbmb.2021.105891
119. Morvaridzadeh M, Sepidarkish M, Yavari M, Tahvilian N, Heydarian A, Khazdouz M, et al. The effects of omega-3 fatty acid supplementation on inflammatory factors in HIV-infected patients: A systematic review and meta-analysis of randomized clinical trials. Cytokine. (2020) 136:155298. doi: 10.1016/j.cyto.2020.155298
120. Oliveira JM, Rondó PH. Omega-3 fatty acids and hypertriglyceridemia in HIV-infected subjects on antiretroviral therapy: systematic review and meta-analysis. HIV Clin trials. (2011) 12:268–74. doi: 10.1310/hct1205-268
121. Delgado Dolset MI, Pablo-Torres C, Contreras N, Couto-Rodríguez A, Escolar-Peña A, Graña-Castro O, et al. Severe allergy as a chronic inflammatory condition from a systems biology perspective. Clin Exp allergy: J Br Soc Allergy Clin Immunol. (2024) 54:550–84. doi: 10.1111/cea.14517
122. Zhang X, Han Y, Tian Q, Du L, Chen L, Zhang Y, et al. The association between n-3 polyunsaturated fatty acid intakes and asthma in US children and adolescents: A cross-sectional study from NHANES. Pediatr Allergy Immunol. (2023) 34:e14024. doi: 10.1111/pai.14024
123. Osuna E, Symington EA, Malan L, Ricci C, Zandberg L, Smuts CM, et al. Higher n-3 polyunsaturated fatty acid status during early pregnancy is associated with lower risk for depression at 12 months postpartum: The NuPED study. Prostaglandins leukotrienes essential Fatty Acids. (2023) 190:102528. doi: 10.1016/j.plefa.2022.102528
124. Lv T, Cao W, Li T. HIV-related immune activation and inflammation: current understanding and strategies. J Immunol Res. (2021) 2021:7316456. doi: 10.1155/2021/7316456
Keywords: HIV/AIDS, INRs, ART, gut microbiota, traditional Chinese medicine (TCM)
Citation: Sun X, Xie Z, Wu Z, Song M, Zhang Y, Zhang Z, Cui X, Liu A and Li K (2024) Mechanisms of HIV-immunologic non-responses and research trends based on gut microbiota. Front. Immunol. 15:1378431. doi: 10.3389/fimmu.2024.1378431
Received: 29 January 2024; Accepted: 12 December 2024;
Published: 26 December 2024.
Edited by:
Elena Moreno, Ramón y Cajal University Hospital, SpainReviewed by:
Erick De La Torre Tarazona, Ramón y Cajal University Hospital, SpainGuo Mu, Zigong Fourth People’s Hospital, China
Texca Tatevari Méndez López, Laboratorio Estatal de Salud Publica de Michoacan Vigilancia Epidemiologica, Mexico
Copyright © 2024 Sun, Xie, Wu, Song, Zhang, Zhang, Cui, Liu and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ke Li, NDY0MzQyMTg3QHFxLmNvbQ==
†These authors share first authorship
‡ORCID: Zhanpeng Xie, orcid.org/0009-0000-5719-6823