 Alyssa J. Matz1
Alyssa J. Matz1 Lili Qu
Lili Qu Keaton Karlinsey
Keaton Karlinsey Anthony T. Vella
Anthony T. Vella Beiyan Zhou
Beiyan Zhou
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Immunol., 16 February 2023
Sec. Molecular Innate Immunity
Volume 14 - 2023 | https://doi.org/10.3389/fimmu.2023.1148188
This article is part of the Research TopicPlasticity and Metabolic Switching in Adipose Tissue MacrophagesView all 11 articles
Adipose tissue macrophages (ATMs) bolster obesity-induced metabolic dysfunction and represent a targetable population to lessen obesity-associated health risks. However, ATMs also facilitate adipose tissue function through multiple actions, including adipocyte clearance, lipid scavenging and metabolism, extracellular remodeling, and supporting angiogenesis and adipogenesis. Thus, high-resolution methods are needed to capture macrophages’ dynamic and multifaceted functions in adipose tissue. Herein, we review current knowledge on regulatory networks critical to macrophage plasticity and their multifaceted response in the complex adipose tissue microenvironment.
Obesity is a prevalent health risk to an expanding list of co-morbidities, increasing global rates of disability and mortality (1–3). Obesity induces white adipose tissue (WAT) dysfunction that significantly contributes to obesity-associated health risks through chronic, low-grade tissue inflammation, insulin resistance, hyperlipidemia, and hypertension (4). Studies have demonstrated that controlling obesity-associated WAT inflammation can improve tissue function and systemic health (5–9).
Within white adipose tissue, macrophages are the most abundant immune population. During obesity, the adipose tissue macrophage (ATM) population increases 10-fold in cell number and exacerbates local inflammation. ATMs originate from hematopoietic stem-cell-derived circulating monocytes and self-replicating tissue residents seeded during fetal development (10–12). Inhibiting macrophage expansion by limiting monocyte-derived macrophage recruitment during obesity lessens WAT inflammation (11, 13, 14). However, the ATM population is heterogeneous and critical to tissue function. Indeed, inhibition of specific ATM functions worsens systemic metabolic health (15). Thus, it is essential to understand the molecular signaling pathways that enrich beneficial ATM functions under obesity rather than eliminate them. Several ATM functions have been identified, including dead adipocyte clearance, lipid scavenging and metabolism, extracellular remodeling, and supporting angiogenesis and adipogenesis. In addition, the macrophage population in brown adipose tissue is less defined, but several studies demonstrate their importance in maintaining thermogenesis (16–18). This review summarizes ATM functions important to white adipose tissue and relevant regulatory pathways that promote these actions. Herein, we aim to demonstrate the power of high-resolution investigations to characterize diverse macrophage populations and the need for function-based analysis to deconvolute targetable networks to lessen obesity-induced comorbidity and mortality.
Macrophages can perform an array of functions to diverse stimuli, including pathogen- and damage-associated molecular patterns, cytokines, chemokines, metabolites, and extracellular vesicles (19, 20). Several models have been developed to classify macrophage features. One of the most widely utilized models is the M1/M2 paradigm, which delineates two central functional states: classically activated macrophages (M1) and alternatively activated macrophages (M2) (21). This and other in vitro models have allowed the characterization of regulatory mechanisms and signaling pathways crucial to several macrophage responses. It is now appreciated that these in vitro-based models do not recapitulate the complex stimuli experienced by tissue-resident macrophages but provide a basis to deconvolute the responses seen in vivo. Thus, mapped macrophage responses to stimuli experienced in adipose tissue and other physiologic and pathogenic states will be summarized in the following section (Figure 1A).
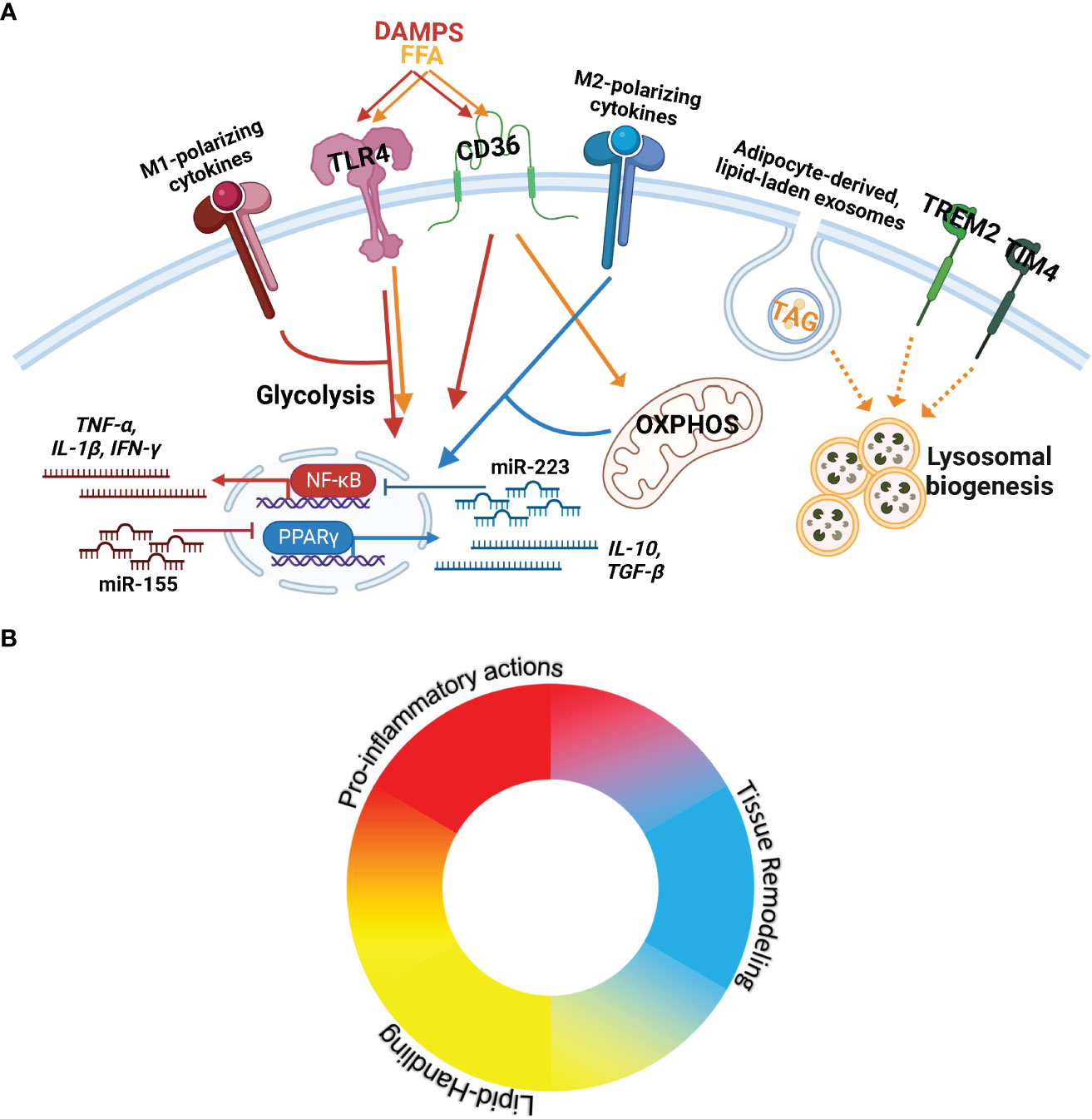
Figure 1 ATMs experience a complex microenvironment with opposing and compounding signals that results in a spectrum of activity. (A) Key stimuli, receptors, and signaling cascades known to control macrophage actions and implicated in ATM biology. (From Left to Right) M1-polarizing cytokines include TNF-α and IFN-γ. M1-polarizing chemokines are also present in AT and include CCL2, CCL4, and CXCL8. DAMPS= Danger-associated molecular patterns including exogenous (lipopolysaccharide) and endogenous signaling (e.g. apoptotic cells) are recognized through TLR4 to elicit pro-inflammatory signaling (JAK-STAT/NK-κB/MAPK) towards the production of type-1 cytokines (TNF-α, IL-1β, IFN-γ). miR-155 promotes M1-polarization through inhibition of signaling cascades towards PPARγ. FFA= Free Fatty Acids signaling through TLR4 elicits pro-inflammatory activation; CD36 transports FFAs, upregulating fatty acid oxidation and OXPHOS= Oxidative Phosphorylation. CD36 also recognizes DAMPs to activate pro-inflammatory activation. M2-polarizing cytokines (IL-4, IL-13, IL-10, IL-17A, IL-25) activate PPARγ to produce type-2 cytokines (IL-10, TGF-β). miR-223 promotes M2-polarization through the repression of pro-inflammatory signal cascade components. OXPHOS is required for M2 actions. Adipocyte-derived exosomes transport neutral lipids (including TAG=Triacylglycerols) into macrophages, which induce lysosomal biogenesis for lipid metabolism. Trem2 and Tim4 also elicit lysosomal biogenesis, but the ligands and downstream signaling cascades are unclear. (B) Macrophages are plastic, responding to diverse stimuli to provide appropriate action. Due to the plasticity of macrophages, their multifaceted capacity, and the complex microenvironment in AT, a single ATM can perform various actions. The ATM population is best represented as a spectrum of macrophage functions.
WAT is a source of adipokines, including immune modulatory cytokines, chemokines, and growth hormones. Many in vitro studies have detailed cytokines/chemokines that differentially contribute to the polarization of macrophages. Tumor necrosis factor alpha (TNF-α), interferon gamma (IFN-γ), C-C motif chemokine 2 (CCL2), CCL4, and interleukin 8 (CXCL8) can induce the classical activation of macrophages. M1-polarized macrophages provide acute pro-inflammatory effector functions by expressing reactive oxygen species, nitric oxide, and secretion of type-1 cytokines such as TNF-α, IFN-γ, and interleukin 1 beta (IL-1β). In contrast, macrophages are activated towards M2 polarization by IL-4, IL-13, IL-10, IL-17A, IL-25, and CCL5 to resolve acute inflammation and secrete type-2 cytokines, including IL-10 and arginase (21–25). The effect of cytokines on the polarization of macrophages has been investigated through in vivo studies in various contexts relevant to ATM functions (22, 26–28). Administration of IL-25 to obese mice mitigated weight gain through enhanced ATM M2-polarization, mitochondrial respiratory capacity, and lipolysis, demonstrating the therapeutic potential of targeting macrophage actions in obesity-associated disease (22).
Macrophages are armed with an extensive repertoire of germ-line encoded pattern recognition receptors (PRRs) that recognize conserved molecular patterns that stimulate rapid innate immunity. Macrophages are also phagocytic and respond to particulate ligands from engulfed vesicles. Macrophage PRRs and ligands are summarized elsewhere (29); herein, we focus on those implicated in adipose tissue function. Receptor ligand interactions regulate macrophage responses via the release of stored mediators, transcription activation, and metabolic reprogramming. An important family of PRRs is Toll-like receptors (TLRs), which elicit pro-inflammatory responses to exogenous and endogenous molecular targets. TLR4 can be activated by lipopolysaccharide to initiate classical macrophage polarization (M1), as well as saturated fatty acids (30, 31), which are rich in obese AT. Inhibiting TLR4 signaling in models of obesity improves systemic metabolic function (32). Macrophages also express phosphatidylserine recognition receptors, scavenger receptors, type 3 complement receptors (CR3), β-glucan receptors, Fc receptors, and mannose receptors. Phosphatidylserine recognition receptors are a diverse group of proteins apt for recognizing apoptotic cells (33). Their dynamics in ATMs warrant further investigation, given their role in adipocyte clearance. Within scavenger receptors, CD36 is elevated in ATMs under obese stress (34). CD36 is a receptor for long-chain fatty acid transport (35) and binds various danger-associated molecular patterns (DAMPS) (31, 36, 37). Ligand-dependent activation of signaling cascades through CD36 relies on specific co-receptors and partners (33). Fatty acid binding to CD36 upregulates fatty acid oxidation (38), while DAMP interactions initiate a pro-inflammatory cascade in macrophages (39). CD36-mediated triacylglycerol uptake is necessary to support M2 activation through stimulating lipolysis and elevated oxidative phosphorylation (40). In the case of circulating macrophages, CD36 binding to oxidized low-density lipoproteins initiates a cellular metabolic shift to activate Mitogen-activated protein kinases (MAPK) signaling towards M1-polarization (39, 41, 42).
Various stimuli elicit common downstream signaling pathways and metabolic programs, yielding similar macrophage responses. These signaling cascades’ strength is tuned by small non-coding RNA molecules termed microRNA (miRNAs). The effect of miRNA on the polarization of
macrophages and its impact on ATM function has been extensively researched and is reviewed elsewhere (43). Notably, miR-155 and miR-223 are key regulators of macrophage polarization and profoundly impact systemic metabolism in obesity.
M1 polarization relies on the Janus kinase-signal transducer and activator of transcription (JAK-STAT) pathway via nuclear factor kappa B (NF-κB and MAPK signaling (44). The microRNA miR-155 supports pro-inflammatory activation of macrophages (45, 46) by repressing the translation of potent anti-inflammatory mediators including Socs1, Ship1, and IL13Rα1 (47–50).
In contrast, M2 activation depends on the transcription factor Peroxisome proliferator-activated receptor γ (PPARγ), activated downstream of IL4-IL4R signaling. PPARγ stimulates miR-223 expression, supporting PPARγ in a positive feedback loop (51–53). miR-223 suppresses components of NF-κB signaling, including Nfat5, Rasa1, and PKNOX1, therein reducing pro-inflammatory cytokine production (51–53). Obese mice with systemic deletion of miR-223 had heightened WAT pro-inflammatory macrophage infiltration, inflammation, and worsened systemic metabolism (51).
Macrophage cellular metabolism is integral to activation and function. For pro-inflammatory activation, macrophages rely on HIF1α-mediated aerobic glycolysis enrichment (40, 54, 55), increasing glucose and oxygen consumption likely to increase the production of reactive oxygen species (40). M2-polarization requires lipolysis and oxidative phosphorylation (56); inhibiting metabolic reprogramming severely weakens M2-mediated responses, including clearance of parasite helminth infections (56). Macrophage lipid metabolism is regulated through lipid uptake, synthesis, and clearance through mediators Sterol regulatory element-binding protein 1 (SREBP) and L-xylulose reductase (LXR). Inhibition of SREBP signaling in ATMs reduces cholesterol efflux, activating M1-polarization cascades and increasing the proportion of pro-inflammatory ATMs (57). In addition, LXR-stimulated fatty acid synthesis represses pro-inflammatory cytokine production in ATMs and increases systemic insulin sensitivity in obesity (58). However, TLR4-mediated signaling inhibits LXR-mediated fatty acid synthesis and downstream Myeloid differentiation primary response protein (MyD88)- and TIR domain-containing adapter molecule (TRIF)- signaling pathways alters the lipid composition of macrophages to intensify inflammation (59). These opposing signaling cascades are likely responsible for the spectrum of ATMs activation states observed in vivo (Figure 1B).
ATM populations are heterogeneous, containing specialized subsets that can perform pro-inflammatory actions, lipid scavenging and metabolism, extracellular remodeling, and support angiogenesis and adipogenesis (Table 1). Functional characterization of tissue-resident macrophages is necessary to understand their regulation and identify modulatory pathways to promote metabolic health in obesity.
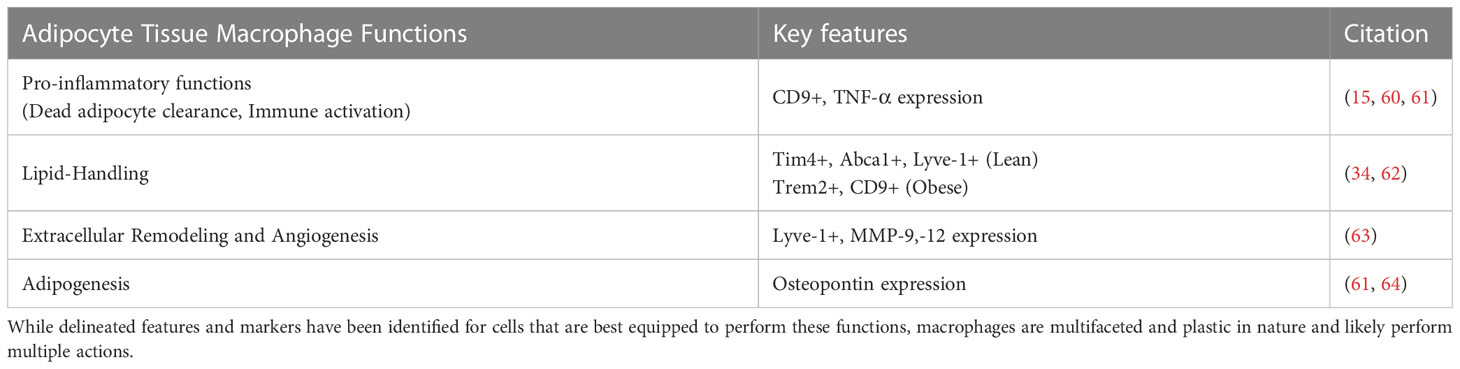
Table 1 Macrophage functions in adipose tissue and identified markers/features of specialized cells.
A primary function of recruited macrophages in WAT is immune activation for the removal of dead adipocytes, accomplished by pro-inflammatory ATMs. ATMs produce pro-inflammatory mediators that bolster tissue and systemic inflammation including TNF-α, IL-6, IL-1β, CCL2, Inducible nitric oxide synthase (iNOS), and others (65). Under obesity stress, WAT remodeling is continual, and the macrophage per adipocyte ratio increases significantly (10, 60, 66). The first characterization of ATM dynamics in lean and obese conditions found a shift from M2-like to M1-like cell predominance under obesity and secrete elevated TNF-α (60, 61). RNA-sequencing identified CD9-expressing, pro-inflammatory ATMs that surround dead adipocytes are abundant in obesity (15). ATMs also contribute to local tissue immune activation through antigen presentation to resident adaptive immune cells. During obesity, ATM express increased major histocompatibility complex II (MHCII) and co-stimulatory molecules to activate CD4+ T cells (67).
Notably, glucagon-like peptide-1 (GLP-1) agonists and GLP-1 analogs are a class of medication utilized in the treatment of obesity and type 2 diabetes (68, 69). Administration significantly reduces fat mass and macrophage per gram of fat in models of obesity (70). ATMs and peritoneal macrophages from treated mice also express less TNF-α and IL-6 (70), suggesting a possible direct role on macrophage activity. Given the impact of GLP-1 on obesity-induced health risk, further investigation into their impact on ATM actions is warranted.
Macrophages support adipose tissue storage capacity by metabolizing lipids through lysosomal lipolysis. While all ATMs upregulate surface expression of the CD36 after a high-fat meal (34), subsets of specialized lipid-handling ATMs have been identified in both lean and obese WAT. In lean WAT, a subset of ATMs delineated by phospholipid-transporting ATPase ABCA1 (Abca1), T-cell immunoglobulin and mucin domain-containing protein 4 (Tim4), and lymphatic vessel endothelial hyaluronic acid receptor 1 (Lyve1) expression are self-replicating and most apt for lipid uptake and metabolism (34). After a meal, Tim4+ ATMs increase their lipid uptake, lysosomal content, and release HDL as part of the reverse cholesterol pathway (34).
During obesity, ATM transcriptomes demonstrate lysosome biogenesis is significantly enriched (66). Lipid-laden ATMs are more abundant and associated with elevated lysosome content (15, 62, 66). While Tim4+ ATMs persist in obesity, a novel lipid-laden ATM subset predominates. High-resolution investigations first delineated obesity-associated lipid-laden ATMs with CD9 expression (15), which were further demarcated by Triggering receptor expressed on myeloid cells 2 (Trem2) expression in both mice and humans (62). Recruited monocyte-derived macrophages are programmed by Trem2 signaling into lipid-laden ATMs (15). While inhibiting infiltration of monocyte-derived macrophages into obese WAT lessened overall tissue inflammation (11, 13, 14), specific inhibition of the Trem2 lipid-laden program of recruited macrophages exacerbates dyslipidemia and adipocyte hypertrophy, worsening overall metabolic health (15). Interestingly, ATMs uptake neutral lipids from adipocyte-derived exosomes, a distinct means of lipid handling for macrophages (71). Adipocyte-derived exosomes can induce ATM features from monocytes in vitro and, during obesity, are released at a significantly higher rate (71). Tim4 and Trem2 are relevant for ATM lysosome biogenesis. However, their ligand and downstream signaling cascades have not been elucidated. Tim4 and Trem2 are receptors for phosphatidylserine, a major component of exosomes (72, 73). Further research into the mechanism of adipocyte exosomes in developing lipid-laden ATMs could yield translatable discoveries for metabolic disease.
WAT angiogenesis, adipogenesis, and extracellular remodeling are tightly linked processes to expand lipid storage capacity. ATMs regulate each process, but it is unclear if specialized subsets persist in WAT to perform these actions or if multifaceted macrophages contribute to these functions. While these cells do not recapitulate the M2 program, tissue remodeling and Transforming growth factor β (TGF-β) production have been described as an action of M2-polarized macrophages (74, 75). Whether important regulatory pathways towards M2 activation are responsible for ATM extracellular remodeling, angiogenesis, and adipogenesis remains unclear.
Angiogenesis is crucial to prevent hypoxia in expanding WAT. Increasing AT capillary density abrogates obesity-induced insulin resistance and metabolic dysfunction (76). A monocyte-derived ATMs expressing Lyve1 are recruited to hypoxic outgrowths of WAT in adolescent, lean mice. Lyve1+ ATMs express angiogenic matrix metalloproteinases (MMPs) and other angiogenic factors to form dense vascular networks that permit subsequent adipogenesis-mediated WAT development (63). MMPs degrade extracellular matrix components and are elevated in models of obesity. However, the extent of Lyve1+ ATM extracellular remodeling has yet to be explored. In addition, examples of macrophage-dependent angiogenesis in other contexts suggest M1 and M2 features are required for different stages of angiogenesis (77).
WAT requires extracellular remodeling to reduce stress on expanding and newly generated adipocytes and allow for proper vascularization. Unresolved remodeling and inflammation can lead to excessive extracellular matrix (ECM) component deposition, known as fibrosis. Obesity-induced WAT fibrosis has been linked to worsened metabolic dysfunction resistant to weight loss (78–80). In models of obesity-associated WAT fibrosis, macrophage depletion ameliorates fibrosis by reducing tissue inflammation and fibroblast activation (74). The primary mediator of macrophage-mediated activation of fibroblasts is TGF-β (63, 74, 81). In contrast, macrophages are also capable of ECM component uptake and degradation; however, these actions have not been explored in WAT.
Adipose tissue undergoes continual adipocyte turnover, refreshing the population through adipogenesis. In addition to adipogenesis within tissue outgrowths, ATMs initiate adipogenesis by expressing osteopontin to recruit pre-adipocytes towards a dying adipocyte (61, 64). In this way, the adipocyte is quickly replaced, and newly differentiated cells have space to develop; however, localization of the progenitor cell to the periphery of pro-inflammatory ATM clearance of a dead cell can be a double edge sword. Pro-inflammatory mediators such as TNF-α repress master adipogenic transcription factor PPARγ in pre-adipocytes (82, 83). During obesity, insufficient adipogenesis forces adipocytes to increase in size, or hypertrophy, and is correlated with increased inflammation, worsened metabolic health, and greater risk for co-morbidities (84–89).
Traditional methods to infer macrophage function rely on detecting a limited number of M1/M2 markers at the RNA or protein levels. Transcriptomic investigations, including microarrays and RNA-sequencing, have demonstrated the diversity of ATMs that cannot be neatly dichotomized into M1/M2 (15, 90) and revealed the importance of lipid-mediated reprogramming (15, 66). The advent of single-cell RNA-sequencing (scRNA-seq) has permitted unparalleled resolution to characterize the diversity in the immune compartment of adipose tissue.
ScRNA-seq generates highly dimensional data, and strategies to analyze begin with performing dimension reduction to facilitate downstream comparisons. Most strategies for dimension reduction utilize the whole transcriptome to establish similarities across cells, such as t-distributed stochastic neighbor embedding (tSNE) (91) and uniform manifold approximation and project (UMAP) (92). Following dimension reduction, cells are typically clustered into groups based on relative distances in the low-dimensional projection and overall transcriptomic similarity. These unsupervised algorithms delineate cells well based on major perturbations in the transcriptome to cluster cells based on lineage or for de novo subset identification. Researchers can then characterize clusters based on known cell markers and differential gene expression. However, due to frequent gene “dropouts” in scRNA-seq data, where expressed genes are not detected, and the variable turnover of marker proteins, traditional biomarker genes are not always reliable. To accommodate the disconnect observed between mRNA and protein levels, semi-supervised tools have been developed that use machine learning-based classifiers or manually curated lists of biomarkers to classify groups of cells based on their transcriptomic profiles (93–95). In addition, multiomic strategies combining scRNA-seq with targeted proteomic analysis (96) or spatial location (97) are becoming more popular. Application of scRNA-Seq in adipose tissue depicted the heterogeneity of ATMs, revealing the lipid-handling Tim4+ and obesity-associated Trem2+ population in mice (34, 62) and tissue immune cell dynamics across lineages in lean and obese humans (98).
Beyond identifying cell subsets, cell function is of great scientific interest in scRNA-seq studies. Several programs have been widely applied for the downstream cluster-based annotation of gene ontology from transcriptomics data, including DAVID (99, 100) and Qiagen Ingenuity Pathway Analysis (IPA) (101). However, these methods are not high-throughput and thus cannot represent the continuum of cell actions. In addition, while traditional whole-transcriptome dimension reduction and clustering techniques work well to distinguish cell lineages, differences in cell function, especially for cells such as macrophages, are often represented by much more subtle transcriptomic changes. Therefore, cell functional annotation in scRNA-seq data is a current technical challenge representing an area of active research.
Depicting the dynamic actions of macrophages in transcriptomics data has been challenging. Due to the whole-transcriptome input into unsupervised algorithms, functional distinctions are not intentionally utilized for clustering. Further, cells respond to diverse stimuli that induce divergent transcriptomes for multifaceted functions (102). Thus, capturing complex stimulation-induced signaling network changes towards multifaceted actions poses an important technical challenge in bioinformatics (103). To this end, we have designed two programs, MacSpectrum (90) and AtheroSpectrum (104), to depict three actions of macrophages along a spectrum of action intensity: monocyte maturation, macrophage inflammatory polarization, and atherosclerosis-related foaming. Applications of MacSpectrum in ATMs depicted the enrichment of pro-inflammatory ATMs in obesity (90), permitting investigation into obesity-associated macrophage inflammatory programs. Further, MacSpectrum characterized an ATM CD206+CD11c- subset enriched in diabetic obese humans as phenotypically distinct, with terminal differentiation and less pro-inflammatory than lipid-laden cells (105). AtheroSpectrum is tailored to atherosclerotic macrophages. Utilization of AtheroSpectrum revealed two novel macrophage foaming programs: homeostasis foaming and pathogenic foaming, the latter associated with cardiovascular disease. Depicting these distinct programs allowed for a focused investigation into pathogenic foaming that enabled leveraging program-specific genes to improve cardiovascular risk prediction models (104). These programs demonstrate the importance of depicting macrophage plasticity to parse out nuanced regulatory networks driving diversified macrophage function.
Obesity is a major health risk in part due to adipose tissue dysfunction. In the adipose tissue, macrophages potentiate local inflammation through pro-inflammatory cytokine production and immune activation. However, ATMs represent a heterogeneous population that support tissue function through dead adipocyte clearance, lipid-buffering, extracellular remodeling, and supporting angiogenesis and adipogenesis. High-resolution techniques have allowed the identification of ATM subsets best suited to these functions. Leveraging scRNA-seq techniques to capture the spectrum of multifaceted macrophage actions allows for important macrophage programs to emerge that are not evident using traditional macrophage categorization models or low-resolution techniques. Function-guided macrophage annotations are important to understanding tissue heterogeneity and investigating ATM programs correlated with metabolic health.
AM wrote and revised the manuscript. LQ, KK, AV and BZ contributed to manuscript design and revisions. All authors contributed to the article and approved the submitted version.
This research was funded by the American Heart Association grant 19TPA34910079 to BZ; and National Institutes of Health/National Institute of Diabetes and Digestive and Kidney Diseases R01DK121805 to BZ and AV; the American Heart Association grants 23PRE1023282 to AM, and 23PRE1023195 to KK.
All figures were created with BioRender.com.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Field AE, Coakley EH, Must A, Spadano JL, Laird N, Dietz WH, et al. Impact of overweight on the risk of developing common chronic diseases during a 10-year period. Arch Intern Med (2001) 161:1581–6. doi: 10.1001/archinte.161.13.1581
2. GBD 2015 Obesity Collaborators, Afshin A, Forouzanfar MH, Reitsma MB, Sur P, Estep K, et al. Health effects of overweight and obesity in 195 countries over 25 years. N Engl J Med (2017) 377:13–27. doi: 10.1056/NEJMoa1614362
3. Juonala M, Magnussen CG, Berenson GS, Venn A, Burns TL, Sabin MA, et al. Childhood adiposity, adult adiposity, and cardiovascular risk factors. N Engl J Med (2011) 365:1876–85. doi: 10.1056/NEJMoa1010112
4. Fruhbeck G. Overview of adipose tissue and its role in obesity and metabolic disorders. Methods Mol Biol (2008) 456:1–22. doi: 10.1007/978-1-59745-245-8_1
5. Xu H, Barnes GT, Yang Q, Tan G, Yang D, Chou CJ, et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J Clin Invest (2003) 112:1821–30. doi: 10.1172/JCI19451
6. Fleischman A, Shoelson SE, Bernier R, Goldfine AB. Salsalate improves glycemia and inflammatory parameters in obese young adults. Diabetes Care (2008) 31:289–94. doi: 10.2337/dc07-1338
7. Stanley TL, Zanni MV, Johnsen S, Rasheed S, Makimura H, Lee H, et al. TNF-alpha antagonism with etanercept decreases glucose and increases the proportion of high molecular weight adiponectin in obese subjects with features of the metabolic syndrome. J Clin Endocrinol Metab (2011) 96:146. doi: 10.1210/jc.2010-1170
8. Hauser TH, Salastekar N, Schaefer EJ, Desai T, Goldfine HL, Fowler KM, et al. Effect of targeting inflammation with salsalate: The TINSAL-CVD randomized clinical trial on progression of coronary plaque in overweight and obese patients using statins. JAMA Cardiol (2016) 1:413–23. doi: 10.1001/jamacardio.2016.0605
9. Oral EA, Reilly SM, Gomez AV, Meral R, Butz L, Ajluni N, et al. Inhibition of IKKvarepsilon and TBK1 improves glucose control in a subset of patients with type 2 diabetes. Cell Metab (2017) 26:157–170.e7. doi: 10.1016/j.cmet.2017.06.006
10. Weisberg SP, McCann D, Desai M, Rosenbaum M, Leibel RL, Ferrante AW Jr. Obesity is associated with macrophage accumulation in adipose tissue. J Clin Invest (2003) 112:1796–808. doi: 10.1172/JCI200319246
11. Kanda H, Tateya S, Tamori Y, Kotani K, Hiasa K, Kitazawa R, et al. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J Clin Invest (2006) 116:1494–505. doi: 10.1172/JCI26498
12. Amano SU, Cohen JL, Vangala P, Tencerova M, Nicoloro SM, Yawe JC, et al. Local proliferation of macrophages contributes to obesity-associated adipose tissue inflammation. Cell Metab (2014) 19:162–71. doi: 10.1016/j.cmet.2013.11.017
13. Odegaard JI, Ricardo-Gonzalez RR, Goforth MH, Morel CR, Subramanian V, Mukundan L, et al. Macrophage-specific PPARgamma controls alternative activation and improves insulin resistance. Nature (2007) 447:1116–20. doi: 10.1038/nature05894
14. Patsouris D, Li PP, Thapar D, Chapman J, Olefsky JM, Neels JG. Ablation of CD11c-positive cells normalizes insulin sensitivity in obese insulin resistant animals. Cell Metab (2008) 8:301–9. doi: 10.1016/j.cmet.2008.08.015
15. Hill DA, Lim HW, Kim YH, Ho WY, Foong YH, Nelson VL, et al. Distinct macrophage populations direct inflammatory versus physiological changes in adipose tissue. Proc Natl Acad Sci U.S.A. (2018) 115:E5096–105. doi: 10.1073/pnas.1802611115
16. Rosina M, Ceci V, Turchi R, Chuan L, Borcherding N, Sciarretta F, et al. Ejection of damaged mitochondria and their removal by macrophages ensure efficient thermogenesis in brown adipose tissue. Cell Metab (2022) 34:533–548.e12. doi: 10.1016/j.cmet.2022.02.016
17. Adachi Y, Ueda K, Nomura S, Ito K, Katoh M, Katagiri M, et al. Beiging of perivascular adipose tissue regulates its inflammation and vascular remodeling. Nat Commun (2022) 13:5117–6. doi: 10.1038/s41467-022-32658-6
18. Gallerand A, Stunault MI, Merlin J, Luehmann HP, Sultan DH, Firulyova MM, et al. Brown adipose tissue monocytes support tissue expansion. Nat Commun (2021) 12:5255–1. doi: 10.1038/s41467-021-25616-1
19. Locati M, Curtale G, Mantovani A. Diversity, mechanisms, and significance of macrophage plasticity. Annu Rev Pathol (2020) 15:123–47. doi: 10.1146/annurev-pathmechdis-012418-012718
20. Qu L, Matz AJ, Karlinsey K, Cao Z, Vella AT, Zhou B. Macrophages at the crossroad of meta-inflammation and inflammaging. Genes (Basel) (2022) 13:2074. doi: 10.3390/genes13112074
21. Mills CD, Kincaid K, Alt JM, Heilman MJ, Hill AM. M-1/M-2 macrophages and the Th1/Th2 paradigm. J Immunol (2000) 164:6166–73. doi: 10.4049/jimmunol.164.12.6166
22. Feng J, Li L, Ou Z, Li Q, Gong B, Zhao Z, et al. IL-25 stimulates M2 macrophage polarization and thereby promotes mitochondrial respiratory capacity and lipolysis in adipose tissues against obesity. Cell Mol Immunol (2018) 15:493–505. doi: 10.1038/cmi.2016.71
23. Guo M, Härtlova A, Gierliński M, Prescott A, Castellvi J, Losa JH, et al. Triggering MSR1 promotes JNK-mediated inflammation in IL-4-activated macrophages. EMBO J (2019) 38:e100299. doi: 10.15252/embj.2018100299
24. Ruytinx P, Proost P, Van Damme J, Struyf S. Chemokine-induced macrophage polarization in inflammatory conditions. Front Immunol (2018) 9:1930. doi: 10.3389/fimmu.2018.01930
25. Czimmerer Z, Halasz L, Daniel B, Varga Z, Bene K, Domokos A, et al. The epigenetic state of IL-4-polarized macrophages enables inflammatory cistromic expansion and extended synergistic response to TLR ligands. Immunity (2022) 55:2006–2026.e6. doi: 10.1016/j.immuni.2022.10.004
26. Seki E, de Minicis S, Inokuchi S, Taura K, Miyai K, van Rooijen N, et al. CCR2 promotes hepatic fibrosis in mice. Hepatology (2009) 50:185–97. doi: 10.1002/hep.22952
27. Karlmark KR, Weiskirchen R, Zimmermann HW, Gassler N, Ginhoux F, Weber C, et al. Hepatic recruitment of the inflammatory Gr1+ monocyte subset upon liver injury promotes hepatic fibrosis. Hepatology (2009) 50:261–74. doi: 10.1002/hep.22950
28. Götze AM, Schubert C, Jung G, Dörr O, Liebetrau C, Hamm CW, et al. IL10 alters peri-collateral macrophage polarization and hind-limb reperfusion in mice after femoral artery ligation. Int J Mol Sci (2020) 21:2821. doi: 10.3390/ijms21082821
29. Gordon S. Pattern recognition receptors: doubling up for the innate immune response. Cell (2002) 111:927–30. doi: 10.1016/s0092-8674(02)01201-1
30. Shi H, Kokoeva MV, Inouye K, Tzameli I, Yin H, Flier JS. TLR4 links innate immunity and fatty acid-induced insulin resistance. J Clin Invest (2006) 116:3015–25. doi: 10.1172/JCI28898
31. Kopp A, Gross P, Falk W, Bala M, Weigert J, Buechler C, et al. Fatty acids as metabolic mediators in innate immunity. Eur J Clin Invest (2009) 39:924–33. doi: 10.1111/j.1365-2362.2009.02185.x
32. Vila IK, Badin P, Marques M, Monbrun L, Lefort C, Mir L, et al. Immune cell toll-like receptor 4 mediates the development of obesity- and endotoxemia-associated adipose tissue fibrosis. Cell Rep (2014) 7:1116–29. doi: 10.1016/j.celrep.2014.03.062
33. Penberthy KK, Ravichandran KS. Apoptotic cell recognition receptors and scavenger receptors. Immunol Rev (2016) 269:44–59. doi: 10.1111/imr.12376
34. Magalhaes MS, Smith P, Portman JR, Jackson-Jones LH, Bain CC, Ramachandran P, et al. Role of Tim4 in the regulation of ABCA1(+) adipose tissue macrophages and post-prandial cholesterol levels. Nat Commun (2021) 12:4434–7. doi: 10.1038/s41467-021-24684-7
35. Harmon CM, Abumrad NA. Binding of sulfosuccinimidyl fatty acids to adipocyte membrane proteins: Isolation and amino-terminal sequence of an 88-kD protein implicated in transport of long-chain fatty acids. J Membr Biol (1993) 133:43–9. doi: 10.1007/BF00231876
36. Chen Y, Zhang J, Cui W, Silverstein RL. CD36, a signaling receptor and fatty acid transporter that regulates immune cell metabolism and fate. J Exp Med (2022) 219:e20211314. doi: 10.1084/jem.20211314
37. Greenberg ME, Sun M, Zhang R, Febbraio M, Silverstein R, Hazen SL. Oxidized phosphatidylserine-CD36 interactions play an essential role in macrophage-dependent phagocytosis of apoptotic cells. J Exp Med (2006) 203:2613–25. doi: 10.1084/jem.20060370
38. Samovski D, Sun J, Pietka T, Gross RW, Eckel RH, Su X, et al. Regulation of AMPK activation by CD36 links fatty acid uptake to β-oxidation. Diabetes (2015) 64:353–9. doi: 10.2337/db14-0582
39. Chen Y, Yang M, Huang W, Chen W, Zhao Y, Schulte ML, et al. Mitochondrial metabolic reprogramming by CD36 signaling drives macrophage inflammatory responses. Circ Res (2019) 125:1087–102. doi: 10.1161/CIRCRESAHA.119.315833
40. Liu L, Lu Y, Martinez J, Bi Y, Lian G, Wang T, et al. Proinflammatory signal suppresses proliferation and shifts macrophage metabolism from myc-dependent to HIF1α-dependent. Proc Natl Acad Sci USA (2016) 113:1564–9. doi: 10.1073/pnas.1518000113
41. Rahaman SO, Lennon DJ, Febbraio M, Podrez EA, Hazen SL, Silverstein RL. A CD36-dependent signaling cascade is necessary for macrophage foam cell formation. Cell Metab (2006) 4:211–21. doi: 10.1016/j.cmet.2006.06.007
42. Moore KJ, El Khoury J, Medeiros LA, Terada K, Geula C, Luster AD, et al. A CD36-initiated signaling cascade mediates inflammatory effects of beta-amyloid. J Biol Chem (2002) 277:47373–9. doi: 10.1074/jbc.M208788200
43. Matz A, Qu L, Karlinsey K, Zhou B. Impact of microRNA regulated macrophage actions on adipose tissue function in obesity. Cells (2022) 11:1336. doi: 10.3390/cells11081336
44. Mosser DM, Edwards JP. Exploring the full spectrum of macrophage activation. Nat Rev Immunol (2008) 8:958–69. doi: 10.1038/nri2448
45. Taganov KD, Boldin MP, Chang KJ, Baltimore D. NF-kappaB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc Natl Acad Sci USA (2006) 103:12481–6. doi: 10.1073/pnas.0605298103
46. O'Connell RM, Taganov KD, Boldin MP, Cheng G, Baltimore D. MicroRNA-155 is induced during the macrophage inflammatory response. Proc Natl Acad Sci USA (2007) 104:1604–9. doi: 10.1073/pnas.0610731104
47. Androulidaki A, Iliopoulos D, Arranz A, Doxaki C, Schworer S, Zacharioudaki V, et al. The kinase Akt1 controls macrophage response to lipopolysaccharide by regulating microRNAs. Immunity (2009) 31:220–31. doi: 10.1016/j.immuni.2009.06.024
48. O'Connell RM, Chaudhuri AA, Rao DS, Baltimore D. Inositol phosphatase SHIP1 is a primary target of miR-155. Proc Natl Acad Sci USA (2009) 106:7113–8. doi: 10.1073/pnas.0902636106
49. Rajaram MV, Ni B, Morris JD, Brooks MN, Carlson TK, Bakthavachalu B, et al. Mycobacterium tuberculosis lipomannan blocks TNF biosynthesis by regulating macrophage MAPK-activated protein kinase 2 (MK2) and microRNA miR-125b. Proc Natl Acad Sci USA (2011) 108:17408–13. doi: 10.1073/pnas.1112660108
50. Martinez-Nunez RT, Louafi F, Sanchez-Elsner T. The interleukin 13 (IL-13) pathway in human macrophages is modulated by microRNA-155 via direct targeting of interleukin 13 receptor alpha1 (IL13Ralpha1). J Biol Chem (2011) 286:1786–94. doi: 10.1074/jbc.M110.169367
51. Zhuang G, Meng C, Guo X, Cheruku PS, Shi L, Xu H, et al. A novel regulator of macrophage activation: miR-223 in obesity-associated adipose tissue inflammation. Circulation (2012) 125:2892–903. doi: 10.1161/CIRCULATIONAHA.111.087817
52. Ying W, Tseng A, Chang RC, Morin A, Brehm T, Triff K, et al. MicroRNA-223 is a crucial mediator of PPARγ-regulated alternative macrophage activation. J Clin Invest (2015) 125:4149–59. doi: 10.1172/JCI81656
53. Zhou H, Xiao J, Wu N, Liu C, Xu J, Liu F, et al. MicroRNA-223 regulates the differentiation and function of intestinal dendritic cells and macrophages by targeting C/EBPβ. Cell Rep (2015) 13:1149–60. doi: 10.1016/j.celrep.2015.09.073
54. El Kasmi KC, Stenmark KR. Contribution of metabolic reprogramming to macrophage plasticity and function. Semin Immunol (2015) 27:267–75. doi: 10.1016/j.smim.2015.09.001
55. Newsholme P, Curi R, Gordon S, Newsholme EA. Metabolism of glucose, glutamine, long-chain fatty acids and ketone bodies by murine macrophages. Biochem J (1986) 239:121–5. doi: 10.1042/bj2390121
56. Huang SC, Everts B, Ivanova Y, O'Sullivan D, Nascimento M, Smith AM, et al. Cell-intrinsic lysosomal lipolysis is essential for alternative activation of macrophages. Nat Immunol (2014) 15:846–55. doi: 10.1038/ni.2956
57. Lee J, Lee SH, Lee E, Cho J, Song D, Lee YJ, et al. SCAP deficiency facilitates obesity and insulin resistance through shifting adipose tissue macrophage polarization. J Adv Res (2022) S2090-1232(22)00124–2. doi: 10.1016/j.jare.2022.05.013
58. Li P, Spann NJ, Kaikkonen MU, Lu M, Oh DY, Fox JN, et al. NCoR repression of LXRs restricts macrophage biosynthesis of insulin-sensitizing omega 3 fatty acids. Cell (2013) 155:200–14. doi: 10.1016/j.cell.2013.08.054
59. Hsieh W, Zhou QD, York AG, Williams KJ, Scumpia PO, Kronenberger EB, et al. Toll-like receptors induce signal-specific reprogramming of the macrophage lipidome. Cell Metab (2020) 32:128–143.e5. doi: 10.1016/j.cmet.2020.05.003
60. Lumeng CN, Bodzin JL, Saltiel AR. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J Clin Invest (2007) 117:175–84. doi: 10.1172/JCI29881
61. Lumeng CN, DelProposto JB, Westcott DJ, Saltiel AR. Phenotypic switching of adipose tissue macrophages with obesity is generated by spatiotemporal differences in macrophage subtypes. Diabetes (2008) 57:3239–46. doi: 10.2337/db08-0872
62. Jaitin DA, Adlung L, Thaiss CA, Weiner A, Li B, Descamps H, et al. Lipid-associated macrophages control metabolic homeostasis in a Trem2-dependent manner. Cell (2019) 178:686–98.e14. doi: 10.1016/j.cell.2019.05.054
63. Cho CH, Koh YJ, Han J, Sung HK, Jong Lee H, Morisada T, et al. Angiogenic role of LYVE-1-positive macrophages in adipose tissue. Circ Res (2007) 100:47. doi: 10.1161/01.RES.0000259564.92792.93
64. Lee YH, Petkova AP, Granneman JG. Identification of an adipogenic niche for adipose tissue remodeling and restoration. Cell Metab (2013) 18:355–67. doi: 10.1016/j.cmet.2013.08.003
65. Varol C, Mildner A, Jung S. Macrophages: development and tissue specialization. Annu Rev Immunol (2015) 33:643–75. doi: 10.1146/annurev-immunol-032414-112220
66. Xu X, Grijalva A, Skowronski A, van Eijk M, Serlie MJ, Ferrante AW Jr. Obesity activates a program of lysosomal-dependent lipid metabolism in adipose tissue macrophages independently of classic activation. Cell Metab (2013) 18:816–30. doi: 10.1016/j.cmet.2013.11.001
67. Morris DL, Cho KW, Delproposto JL, Oatmen KE, Geletka LM, Martinez-Santibanez G, et al. Adipose tissue macrophages function as antigen-presenting cells and regulate adipose tissue CD4+ T cells in mice. Diabetes (2013) 62:2762–72. doi: 10.2337/db12-1404
68. Urva S, Coskun T, Loh MT, Du Y, Thomas MK, Gurbuz S, et al. LY3437943, a novel triple GIP, GLP-1, and glucagon receptor agonist in people with type 2 diabetes: a phase 1b, multicentre, double-blind, placebo-controlled, randomised, multiple-ascending dose trial. Lancet (2022) 400:1869–81. doi: 10.1016/S0140-6736(22)02033-5
69. Jastreboff AM, Aronne LJ, Ahmad NN, Wharton S, Connery L, Alves B, et al. Tirzepatide once weekly for the treatment of obesity. N Engl J Med (2022) 387:205–16. doi: 10.1056/NEJMoa2206038
70. Lee Y, Park M, Choung J, Kim S, Oh H, Choi C, et al. Glucagon-like peptide-1 inhibits adipose tissue macrophage infiltration and inflammation in an obese mouse model of diabetes. Diabetologia (2012) 55:2456–68. doi: 10.1007/s00125-012-2592-3
71. Flaherty SE, Grijalva A, Xu X, Ables E, Nomani A, Ferrante AWJ. A lipase-independent pathway of lipid release and immune modulation by adipocytes. Science (2019) 363:989–93. doi: 10.1126/science.aaw2586
72. Yoshida T, Hanayama R. TIM4-affinity methods targeting phosphatidylserine for isolation or detection of extracellular vesicles. Methods Mol Biol (2022) 2466:23–36. doi: 10.1007/978-1-0716-2176-9_2
73. Sudom A, Talreja S, Danao J, Bragg E, Kegel R, Min X, et al. Molecular basis for the loss-of-function effects of the alzheimer's disease-associated R47H variant of the immune receptor TREM2. J Biol Chem (2018) 293:12634–46. doi: 10.1074/jbc.RA118.002352
74. Martinez-Santibanez G, Singer K, Cho KW, DelProposto JL, Mergian T, Lumeng CN. Obesity-induced remodeling of the adipose tissue elastin network is independent of the metalloelastase MMP-12. Adipocyte (2015) 4:264–72. doi: 10.1080/21623945.2015.1027848
75. Rao L, Wang Y, Zhang L, Wu G, Zhang L, Wang F, et al. IL-24 deficiency protects mice against bleomycin-induced pulmonary fibrosis by repressing IL-4-induced M2 program in macrophages. Cell Death Differ (2021) 28:1270–83. doi: 10.1038/s41418-020-00650-6
76. Cao Y. Angiogenesis and vascular functions in modulation of obesity, adipose metabolism, and insulin sensitivity. Cell Metab (2013) 18:478–89. doi: 10.1016/j.cmet.2013.08.008
77. Spiller KL, Anfang RR, Spiller KJ, Ng J, Nakazawa KR, Daulton JW, et al. The role of macrophage phenotype in vascularization of tissue engineering scaffolds. Biomaterials (2014) 35:4477–88. doi: 10.1016/j.biomaterials.2014.02.012
78. Divoux A, Tordjman J, Lacasa D, Veyrie N, Hugol D, Aissat A, et al. Fibrosis in human adipose tissue: Composition, distribution, and link with lipid metabolism and fat mass loss. Diabetes (2010) 59:2817–25. doi: 10.2337/db10-0585
79. Lackey DE, Burk DH, Ali MR, Mostaedi R, Smith WH, Park J, et al. Contributions of adipose tissue architectural and tensile properties toward defining healthy and unhealthy obesity. Am J Physiol Endocrinol Metab (2014) 306:233. doi: 10.1152/ajpendo.00476.2013
80. Hasegawa Y, Ikeda K, Chen Y, Alba DL, Stifler D, Shinoda K, et al. Repression of adipose tissue fibrosis through a PRDM16-GTF2IRD1 complex improves systemic glucose homeostasis. Cell Metab (2018) 27:180–194.e6. doi: 10.1016/j.cmet.2017.12.005
81. Shen H, Huang X, Zhao Y, Wu D, Xue K, Yao J, et al. The hippo pathway links adipocyte plasticity to adipose tissue fibrosis. Nat Commun (2022) 13:6030–0. doi: 10.1038/s41467-022-33800-0
82. Meng L, Zhou J, Sasano H, Suzuki T, Zeitoun KM, Bulun SE. Tumor necrosis factor alpha and interleukin 11 secreted by malignant breast epithelial cells inhibit adipocyte differentiation by selectively down-regulating CCAAT/enhancer binding protein alpha and peroxisome proliferator-activated receptor gamma: Mechanism of desmoplastic reaction. Cancer Res (2001) 61:2250–5.
83. Cawthorn WP, Heyd F, Hegyi K, Sethi JK. Tumour necrosis factor-alpha inhibits adipogenesis via a beta-catenin/TCF4(TCF7L2)-dependent pathway. Cell Death Differ (2007) 14:1361–73. doi: 10.1038/sj.cdd.4402127
84. Pischon T, Boeing H, Hoffmann K, Bergmann M, Schulze MB, Overvad K, et al. General and abdominal adiposity and risk of death in Europe. N Engl J Med (2008) 359:2105–20. doi: 10.1056/NEJMoa0801891
85. Shungin D, Winkler TW, Croteau-Chonka DC, Ferreira T, Locke AE, Mägi R, et al. New genetic loci link adipose and insulin biology to body fat distribution. Nature (2015) 518:187–96. doi: 10.1038/nature14132
86. Wang QA, Tao C, Gupta RK, Scherer PE. Tracking adipogenesis during white adipose tissue development, expansion and regeneration. Nat Med (2013) 19:1338–44. doi: 10.1038/nm.3324
87. Jeffery E, Church CD, Holtrup B, Colman L, Rodeheffer MS. Rapid depot-specific activation of adipocyte precursor cells at the onset of obesity. Nat Cell Biol (2015) 17:376–85. doi: 10.1038/ncb3122
88. Wueest S, Rapold RA, Rytka JM, Schoenle EJ, Konrad D. Basal lipolysis, not the degree of insulin resistance, differentiates large from small isolated adipocytes in high-fat fed mice. Diabetologia (2009) 52:541–6. doi: 10.1007/s00125-008-1223-5
89. Gao H, Mejhert N, Fretz JA, Arner E, Lorente-Cebrián S, Ehrlund A, et al. Early b cell factor 1 regulates adipocyte morphology and lipolysis in white adipose tissue. Cell Metab (2014) 19:981–92. doi: 10.1016/j.cmet.2014.03.032
90. Li C, Menoret A, Farragher C, Ouyang Z, Bonin C, Holvoet P, et al. Single cell transcriptomics based-MacSpectrum reveals novel macrophage activation signatures in diseases. JCI Insight (2019) 5:e126453. doi: 10.1172/jci.insight.126453
92. Becht E, McInnes L, Healy J, Dutertre C, Kwok IWH, Ng LG, et al. Dimensionality reduction for visualizing single-cell data using UMAP. Nat Biotechnol (2018) 37:38–44. doi: 10.1038/nbt.4314
93. Abdelaal T, Michielsen L, Cats D, Hoogduin D, Mei H, Reinders MJT, et al. A comparison of automatic cell identification methods for single-cell RNA sequencing data. Genome Biol (2019) 20:194–z. doi: 10.1186/s13059-019-1795-z
94. Crowell HL, Soneson C, Germain P, Calini D, Collin L, Raposo C, et al. Muscat Detects subpopulation-specific state transitions from multi-sample multi-condition single-cell transcriptomics data. Nat Commun (2020) 11:6077–4. doi: 10.1038/s41467-020-19894-4
95. Pliner HA, Shendure J, Trapnell C. Supervised classification enables rapid annotation of cell atlases. Nat Methods (2019) 16:983–6. doi: 10.1038/s41592-019-0535-3
96. Lacar B. Generation of centered log-ratio normalized antibody-derived tag counts from Large single-cell sequencing datasets. Methods Mol Biol (2022) 2386:203–17. doi: 10.1007/978-1-0716-1771-7_14
97. Moncada R, Barkley D, Wagner F, Chiodin M, Devlin JC, Baron M, et al. Integrating microarray-based spatial transcriptomics and single-cell RNA-seq reveals tissue architecture in pancreatic ductal adenocarcinomas. Nat Biotechnol (2020) 38:333–42. doi: 10.1038/s41587-019-0392-8
98. Hildreth AD, Ma F, Wong YY, Sun R, Pellegrini M, O'Sullivan TE. Single-cell sequencing of human white adipose tissue identifies new cell states in health and obesity. Nat Immunol (2021) 22:639–53. doi: 10.1038/s41590-021-00922-4
99. Huang DW, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc (2009) 4:44–57. doi: 10.1038/nprot.2008.211
100. Huang DW, Sherman BT, Lempicki RA. Bioinformatics enrichment tools: paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res (2009) 37:1–13. doi: 10.1093/nar/gkn923
101. Krämer A, Green J, Pollard JJ, Tugendreich S. Causal analysis approaches in ingenuity pathway analysis. Bioinformatics (2014) 30:523–30. doi: 10.1093/bioinformatics/btt703
102. Murray PJ, Allen JE, Biswas SK, Fisher EA, Gilroy DW, Goerdt S, et al. Macrophage activation and polarization: nomenclature and experimental guidelines. Immunity (2014) 41:14–20. doi: 10.1016/j.immuni.2014.06.008
103. Holland CH, Tanevski J, Perales-Patón J, Gleixner J, Kumar MP, Mereu E, et al. Robustness and applicability of transcription factor and pathway analysis tools on single-cell RNA-seq data. Genome Biol (2020) 21:36–z. doi: 10.1186/s13059-020-1949-z
104. Li C, Qu L, Matz AJ, Murphy PA, Liu Y, Manichaikul AW, et al. AtheroSpectrum reveals novel macrophage foam cell gene signatures associated with atherosclerotic cardiovascular disease risk. Circulation (2022) 145:206–18. doi: 10.1161/CIRCULATIONAHA.121.054285
Keywords: obesity, adipose tissue, macrophage, immune response, single-cell RNA sequencing
Citation: Matz AJ, Qu L, Karlinsey K, Vella AT and Zhou B (2023) Capturing the multifaceted function of adipose tissue macrophages. Front. Immunol. 14:1148188. doi: 10.3389/fimmu.2023.1148188
Received: 19 January 2023; Accepted: 06 February 2023;
Published: 16 February 2023.
Edited by:
Junji Xing, Houston Methodist Research Institute, United StatesReviewed by:
Jingbo Pang, University of Illinois at Chicago, United StatesCopyright © 2023 Matz, Qu, Karlinsey, Vella and Zhou. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Beiyan Zhou, Ynpob3VAdWNoYy5lZHU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.