
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol., 15 December 2021
Sec. Vaccines and Molecular Therapeutics
Volume 12 - 2021 | https://doi.org/10.3389/fimmu.2021.778302
This article is part of the Research TopicImmune responses and immune mechanisms triggered by snake and scorpion venomsView all 7 articles
 Paulo Sérgio Bernarde1*
Paulo Sérgio Bernarde1* Manuela Berto Pucca2,3
Manuela Berto Pucca2,3 Ageane Mota-da-Silva4
Ageane Mota-da-Silva4 Wirven Lima da Fonseca1
Wirven Lima da Fonseca1 Marllus Rafael Negreiros de Almeida1
Marllus Rafael Negreiros de Almeida1 Isadora Sousa de Oliveira5
Isadora Sousa de Oliveira5 Felipe Augusto Cerni3
Felipe Augusto Cerni3 Felipe Gobbi Grazziotin6
Felipe Gobbi Grazziotin6 Marco A. Sartim7,8,9
Marco A. Sartim7,8,9 Jacqueline Sachett10,11
Jacqueline Sachett10,11 Fan Hui Wen12
Fan Hui Wen12 Ana Maria Moura-da-Silva7,13
Ana Maria Moura-da-Silva7,13 Wuelton M. Monteiro7,10*
Wuelton M. Monteiro7,10*The two-striped forest-pitviper (Bothrops bilineatus) is an arboreal snake that is currently represented by two subspecies (B. b. bilineatus and B. b. smaragdinus) that comprise a species complex, and its distribution is in the Amazon and the Atlantic Forest. The rarity of encounters with this snake is reflected in the low occurrence of cases of snakebites throughout its geographic distribution and the resulting low number of published clinical reports. However, in some areas, B. bilineatus proves to be more frequent and causes envenomations in a greater proportion. Herein, we review the main aspects of the species complex B. bilineatus, including its biology, ecology, taxonomy, morphology, genetic and molecular studies, geographic distribution, conservation status, venom, pathophysiology and clinical aspects, and epidemiology. In addition, the different antivenoms available for the treatment of envenomations caused by B. bilineatus are presented along with suggestions for future studies that are needed for a better understanding of the snakebites caused by this snake.
The two-striped forest-pitviper (Bothrops bilineatus) is an arboreal snake that is currently represented by two subspecies (B. b. bilineatus and B. b. smaragdinus) that comprise a species complex, and its distribution is in the Amazon and Atlantic Forest, though it is absent in intermediate open and dry habitats (1) (Figures 1 and 2). This viperid has adaptations for arboreal life that include a relatively long, prehensile tail and green coloration that makes it hard to spot in vegetation (3). Throughout its geographical distribution it is considered a relatively uncommon snake in studies of snake fauna [e.g. (4, 5)]; however, in the region of Alto Juruá in the state of Acre, it is one of the most common species to be found in some lowland forests (6–8). Bothrops bilineatus is also present in cocoa plantations that are contiguous to the forests in Bahia, where it is not considered to be a rare snake in this environment (9). Since it is characteristic of forested environments, its presence does not go unnoticed by residents in the communities, who report the encounters and the snakebites caused by this snake (7, 10–13).

Figure 1 Specimens of Bothrops bilineatus from Brazil: (A) lower Moa River forest, Cruzeiro do Sul (Acre); (B) Tapauá state forest (Amazonas state); (C) Murici ecological station (Alagoas state); (D) Municipality of Elísio Medrado (Bahia). Photos (A, B) by Paulo Bernarde, (C, D) by Marco Antonio Freitas.

Figure 2 Distribution map of Bothrops bilineatus in relation to the ecoregions of South America [Level I, II and III, adapted from Griffith et al., (2)]. See the map with the locations and their bibliographic references in Supplementary Material.
Throughout its geographical distribution, it receives different popular names, which are usually associated with its predominantly greenish coloring and the yellow coloring of the labial region (mainly in the populations of the Atlantic Forest and the eastern Amazon). Known mainly as jararaca-verde in the scientific community, it has various regional names in Brazil according to the state in which it occurs, such as papagaia (Acre), bico-de-papagaio (Mato Grosso and Rondônia), cobra-papagaio (Amazonas and Pará), jararaca-pinta-de-ouro (Pará), paraamboia (Pará), surucucu-de-patioba (Pará) and surucucu-pingo de ouro, surucucu-de-ouricana, ouricana, patioba (Bahia) (4, 9, 10, 14). In other Amazonian countries, it is known as víbora-loro (Bolivia), loro and palo verde (Ecuador), loro machaco (Peru), mapanare verde and mapanare rayada (Venezuela) (3).
Bothrops bilineatus has been classified as belonging to the B. taeniatus species group by all recent studies that are focused on the systematic of Bothrops [e.g. (15, 16)]. The species group is composed of six species of arboreal pit vipers that include B. bilineatus, B. chloromelas, B. oligolepis, B. medusa, B. pulchra, and B. taeniatus. In the last 20 years, the monophyly of the species group has been frequently recovered by molecular phylogenetic analyses (15, 17–24).
However, the relationship between the B. taeniatus group and the other species group of Bothrops has been a constant matter of debate (15, 21, 22, 24). The contrasting opinions about the classification of this group are reflected in its taxonomic history. The B. taeniatus group was considered to be an independent genus, i.e., Bothriopsis Peters (1861), for more than 30 years.
Although the species included in the B. taeniatus group have been described in several different genera, during the 70’s, all Neotropical diversity of pitvipers was classified as belonging to Bothrops [except Lachesis ssp.; see Peters and Orejas-Miranda, (25)]. At the end of the 80’s, Campbell and Lamar (26) based on the unpublished dissertation of W.L. Burger (27) split Bothrops (sensu lato) in five genera: Bothriechis, Bothriopsis, Bothrops (sensu stricto), Ophryacus and Porthidium. From then, until the second decade of this century, all species of the B. taeniatus group were referred as belonging to the genus Bothriopsis. They were synonymized again with Bothrops only after the widespread acceptance of the proposal suggested by Salomão et al. (17), which indicated the nested position of the B. taeniatus group within Bothrops (15). Therefore, more than 30 years of scientific literature on B. bilineatus (and other species of the B. taeniatus group) made reference to this species as Bothriopsis bilineatus.
B. bilineatus was described by Wied-Neuwied (1821) as Cophias bilineatus, based on two individuals from Brazil (“Brasilien”). Although undefined in the original description, the type’s location was restricted by Hoge and Lancini (28) to Vila Viçosa (Marobá), Rio Peruhype, Bahia, currently the municipality of Nova Viçosa. Some years later, Hoge (29) evaluated the morphological diversity within the species and described the subspecies, B. b. smaragdinus, based on specimens from the region of Alto Rio Purus in the Brazilian state of Acre. In the same study, he suggested the latter was also present in other areas in the western Amazon, in countries like Bolivia, Colombia, Ecuador and Peru.
After the description of B. b. smaragdinus, it has been suggested that the nominal subspecies B. b. bilineatus is only distributed in two disjunct areas, the northeastern Amazon and the Brazilian Atlantic Forest (1, 3). Although the taxonomic scheme and the geographic understanding of B. bilineatus presenting two subspecies is currently accepted, some studies have indicated the possible existence of intermediate morphotypes (30) and distinct genetic lineages (1).
Compared to other species of Bothrops, B. bilineatus is easily identifiable by its distinctive green color, slender body, and prehensile tail (3). The characteristic pholidosis of the species ranges from 190 to 218 ventral scales in males and from 192 to 220 in females; from 65 to 76 subcaudals in males and 55 to 73 in females; between 23 and 25 scales rows at mid-body; between 5 and 9 intersupraoculars; from 7 to 9 supralabials and 8 to 12 infralabials (3).
The two subspecies of B. bilineatus differ mainly in their coloration pattern, in which B. b. bilineatus presents vertical dark bars on the supralabials and yellowish or reddish spots on the dorsal region of the body, and B. b. smaragdinus lacks dark vertical bars on the supralabials and the dorsal spots, presenting instead a dorsum punctuated with small black dots (3, 29).
In a study based on the phylogeographic analysis of B. bilineatus, Dal Vechio et al. (1) show a deep genetic intraspecific structure, which does not exactly mirror the subspecies definition. They recovered a Bayesian phylogenetic tree topology that indicates the presence of four main clades within B. bilineatus, thus suggesting that B. bilineatus probably represents a complex that includes putative undescribed species. Dal Vechio et al. (1) suggest that these clades are genetic lineages that represent the following geographically restricted populations: 1) Atlantic Forest clade; 2) Guiana Shield clade; 3) Western Amazonian clade; and 4) Central Amazonian clade. The Atlantic Forest and Guiana Shield clades are sister groups, indicating a close relationship between these two lineages. On the other hand, the Western and Central Amazonian clades are relatively more divergent, and have been recovered as successive sister groups of Atlantic Forest and Guiana Shield.
The distribution of the Atlantic Forest and Guiana Shield lineages seems to be easily defined by the limits of the biogeographic regions where they occur. The Atlantic Forest lineage is restricted to the northern portions of the Brazilian Atlantic Forest, occurring in the Brazilian states of Minas Gerais, Espírito Santo, Bahia, Alagoas and Pernambuco (1, 31). While, the distribution of the Guiana Shield lineage seems to be restricted to the Guiana Shield formation, including the territories of Brazil, French Guyana, Guyana, Suriname and probably parts of Venezuela (east of the Orinoco River). In Brazil, this lineage is distributed north of the Amazon River, in the Brazilian states of Amapá, Roraima, northern Amazonas and northern Pará (1, 31).
The Western Amazonian lineage is relatively widespread, occurring in the western regions of the Brazilian Amazon (states of Acre and Rondônia) and in the Amazonian regions of Colombia and Ecuador (probably also occurring in Peru and Bolivia). The eastern limits of the distribution of the Western Amazonian lineage seem to be represented by the Orinoco and Negro Rivers (1). Presenting a similar broad distribution, the Central Amazonian lineage occurs south of the Amazon River, in the Brazilian states of Amazonas, Rondônia, Mato Grosso, Pará (1), and probably also in Maranhão (31).
Although Dal Vechio (2014) (32) in his unpublished Master’s thesis has studied the morphological variability of these genetically defined lineages, he did not formally propose the suggested changes on the taxonomy of B. b. bilineatus in any subsequent publication. Based on his results, we can suppose that the Atlantic Forest lineage represents B. b. bilineatus, but the allocation of the Guiana Shield lineage into this subspecies or in a new currently undescribed taxon is still open for investigation. The Western Amazonian lineages probably represent the subspecies B. b. smaragdinus, and the morphological limits between this lineage and the Central Amazonian lineage seem to be unclear, as well as the limits between the distribution of both lineages. He also suggested that there is evidence to support a possible split of the Western Amazonian lineage into two taxa, one in the north and the other in the south. The Central Amazonian lineage likely represents an undescribed taxon for the B. bilineatus complex, which is morphologically and genetically distinct (32).
The taxonomy for B. bilineatus will certainly change in the next years. The subspecies will be probably elevated to the species level and at least two new taxa will be described for the complex (32). However, the monophyly of the B. bilineatus complex seems to be stable (1), and thus the ecological and toxinological inferences already proposed for B. bilineatus will not be affected.
The maximum sizes reported for Bothrops bilineatus in the literature are 1230 mm (33) and 1200 mm (34), for B. b. smaragdinus in Venezuela. Harvey et al. (35) recorded 948 mm as the maximum size for B. b. smaragdinus in Bolivia. In the Alto Jurá region, west of the Brazilian Amazon, Turci et al. (6) reported two males of 530 (40 g) and 670 (55 g) mm and one female of 780 mm (100 g). Also in Alto Juruá, Fonseca et al. (8) recorded males with 499 to 668 mm (mean 588 mm) and females with 316 to 758 mm (mean 578 mm). In Rondônia, southwest of the Brazilian Amazon, Jorge-da-Silva Jr (36). reported two males (469 to 762 mm) and two females (358 to 825 mm). In Pará, in the eastern Amazon, Cunha and Nascimento (4) and Almeida et al. (37) reported maximum sizes of 822 mm and 980 mm (198 g) for female specimens, respectively. In the Atlantic Forest of Bahia, Argôlo (9) recorded the maximum size of 900 mm for B. b. bilineatus. According to literature records, female individuals tend to be larger (3, 6, 8, 36, 37).
The size of young was reported for B. b. smaragdinus in Rondônia, with males presenting from 255 to 270 mm (both with 4.1 g) and females with 260 mm (3.7 g), 265 mm (3.7 g) and 265 mm (4.1 g) (38). In Pará, male young with 240 mm (4 g) and 250 mm (5 g) and three females with 232 to 250 mm (4 to 6 g) were recorded (37). Although the smallest individual reported here presented 232 mm, Smalligan et al. (39) reported the shortest length of B. b. smaragdinus, which involved a specimen causing envenomation in Peru and which presented 150 mm.
Bothrops bilineatus occurs in lowland rainforests, especially those associated with watercourses (3), and is also present in terra firme and lowland rainforests (4, 5, 8, 36). This snake can also be present in ancient secondary forests near secondary forests (40) and cocoa plantations contiguous to forests (9). Although it is considered infrequent or even rare throughout its geographical distribution (4, 5, 36). In some locations, B. bilineatus has been shown to be one of the most abundant snakes (7, 9). In the Atlantic Forest, B. b. bilineatus was one of the most abundant snakes in southeastern Bahia (9) and, in Upper Juruá, in a lowland forest, B. b. smaragdinus was the most commonly found species during a night search.
In species surveys and snake community ecology studies, B. bilineatus is often infrequently recorded, with one to six specimens collected (5, 36, 41–47).
In some locations within its range, this species of snake has not been recorded, denoting that in some regions it is less frequent or even absent (48–50). In the region of Manaus (north of the Amazon River, west of the Negro River, east of the Uatumã River and south of President Figueiredo), despite the snakes sampling efforts carried out by some studies (48, 51–53), B. bilineatus was never recorded, and this may reflect a true absence (31).
In some published studies in which snakes were collected in some regions and in which specimen abundance data were provided, B. bilineatus corresponded to 0.16 to 28.46% of the specimens in the communities (Table 1). Of the 16 studies analyzed, half recorded three or fewer B. bilineatus specimens. In four studies in which there were higher numbers of snakes collected (988 to 4680 specimens (9, 36, 40, 57), B. bilineatus represented 0.16 to 1.34% of the specimens, denoting its low occurrence in these locations. In only one study carried out in the forest of the lower Moa River (8), B. bilineatus was the most abundant snake species and represented 28.46% of the snake specimens recorded in the location. This intriguing greater abundance of B. bilineatus in this location of lowland forest when compared to its lower frequency with other studies developed in terra firme areas may indicate that this snake may be less frequent or more difficult to detect by visual searches in other types of vegetation structures (8). In addition, the availability and interactions with their prey should be considered, especially with the abundance and occurrence of Osteocephalus amphibians in this location (8).
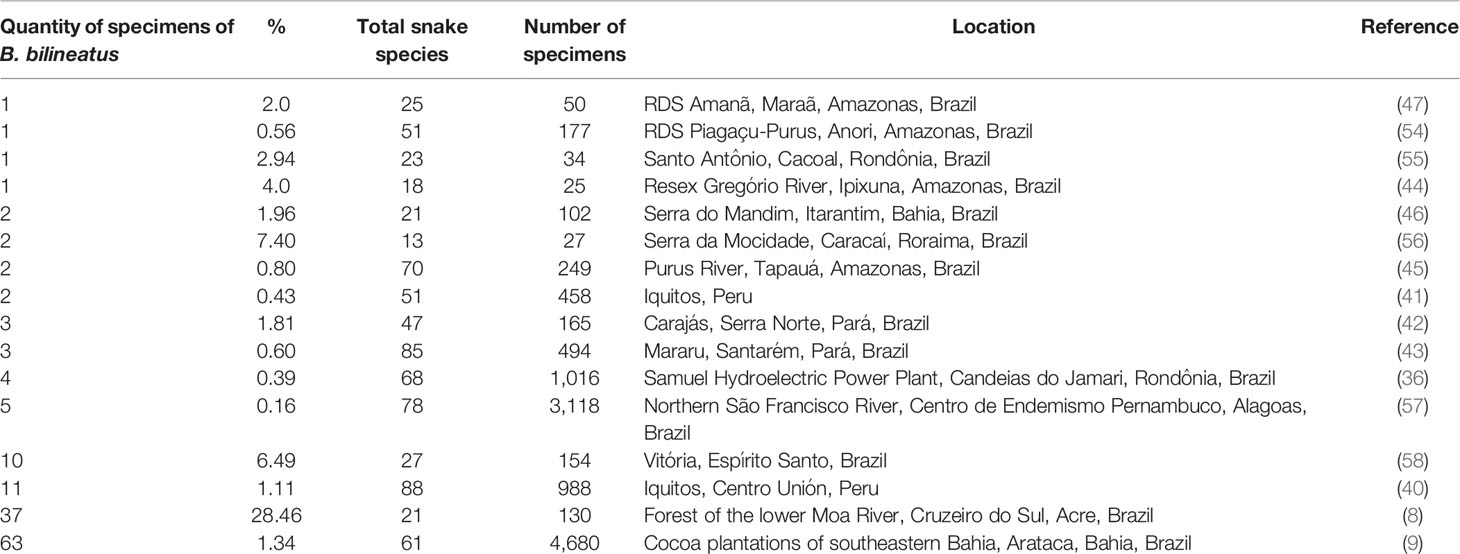
Table 1 Abundance and proportion of Bothrops bilineatus in snake communities.
Most adult specimens of Bothrops species are nocturnal and hunt mainly on the ground and some have a more specialized diet that includes small mammals (e.g., B. alternatus and B. cotiara), while others are more generalist, feeding on various animal groups (centipedes, anurous amphibians, lizards, other snakes, birds, small mammals) (e.g., B. atrox and B. moojeni) (59). Bothrops bilineatus is a species of nocturnal habits and that hunts predominantly while on vegetation, predating anurous amphibians, lizards, birds, rodents and bats (4, 6, 8, 40, 60–65). Despite it being predominantly nocturnal, Fonseca et al. (8) observed three specimens in standby hunting activity during the early morning hours. During the day, they rest on the vegetation, with the green color providing camouflage among the foliage (Figure 3B) (6, 8), usually in the same place that they were hunting at night.

Figure 3 Specimens of Bothrops bilineatus in the forest of the lower Moa River (Acre, Brazil): (A) specimen performing the caudal decoy tactic; (B) specimen resting during the day on a palm frond; (C) male individual found together with a female (Note the volume of stomach contents); (D) arrival of a second male. Photos by (A) Paulo Bernarde, (B) Saymon de Albuquerque, and (C, D) Luiz Carlos Turci.
The hunting tactic used by B. bilineatus is that of waiting, and the snake often uses the caudal decoy behavior (Figure 3A), which consists of performing sinuous movements with the tip of the tail that has a distinct color from the rest of the body, thus being able to attract potential prey close to the range of its predatory strike (8, 64). Caudal decoy behavior is performed by juveniles of some species of Bothrops (e.g., B. atrox, B. jararaca, B. jararacussu and B. moojeni) that feed preferentially on anurous amphibians and, during their development, the adult individuals lose this light coloration at the tip of the tail and cease to perform this behavior (59). Adult specimens of B. bilineatus retain the distinct coloration of the tip of the tail (white or brown) and continue to use the caudal decoy tactic during hunting activity, probably because ectothermic prey (anurous amphibians and lizards) continue to be important in their diet (59, 64, 66).
Among the species of Bothrops that present hunting activity on the vegetation, whether juveniles or eventually adults (e.g., B. atrox and B. jararaca) and arboreal species (e.g., B. insularis and B. taeniatus) (3, 59, 67, 68), B. bilineatus is the one that has been recorded at higher heights (up to 18 m high) (8). In the state of Alagoas (Atlantic Forest), B. bilineatus was observed in hunting activity at 9.4 m, and remained in the same place for three days (69). In a lowland forest in the region of Alto Juruá (State of Acre), western Amazon, B. bilineatus was recorded in hunting activity at heights of 30 cm to 18 m (average height of 6.4 m) (8). In the study by Fonseca et al. (8), the authors recorded that individuals of B. bilineatus usually stay only one night at the same site (17 observations), with some records of them staying longer; two (9 times) and three (6 times) days, with two specimens that were observed for 15 and 17 days.
Studies of communities and populations of snakes in the Amazon and the Atlantic Forest have shown that these animals present more activity and are thus more frequent during the wettest and warmest months (5, 48, 70), and more snakebites were also recorded during this period [e.g. (71–73)]. Unlike other snake species, B. bilineatus was more frequent during the dry season in a lowland forest in the western Amazon, while its congeneric B. atrox was more abundant in the rainy season (7, 8). Fonseca et al. (8) suggested that during the dry season a decrease in humidity occurs in the treetops and some amphibian species (Osteocephalus leprieuri and O. taurinus) migrate vertically to lower heights in search of more humid environments and B. bilineatus would also probably be more active in this forest strata in search of this type of prey.
Bothrops bilineatus is a viviparous snake, with data recording mating between July and August when in captivity, and gestation from 191 to 208 days and parturition in February (74). The number of young in each litter varies from 4 to 15, with a total length of 20 to 27 cm (Starace 1998) (37, 38). Pregnant females in the wild have been collected in September and October and gave birth after about a month in captivity (37, 38). In the wild, Campbell and Lamar (3) reported finding four young in January in Loreto (Peru), and Fonseca et al. (8) observed four young in August (1 young specimen), October (1) and November (2) in Acre (Brazil). Turci et al. (6) observed two males in unsuccessful courting behavior with a resting female (100 g) that had fed on a rodent (60 g) and remained indifferent to the attempts of the males (Figures 3C, D). In this observation by Turci et al. (6), a male of smaller size (53 cm of total length and 40 g), moved away from the site with the arrival of another larger male (67 cm; 55 g).
The national Red List of Threatened Fauna categorizes B. bilineatus with the status of “Least Concern” (75) and it is not listed on the IUCN Red List (76). However, in the Atlantic Forest, reptile species have suffered a considerable decline, mainly due to the loss and degradation of habitats in this biome and there is still a lack of information regarding the conservation status of the species in several locations (77). In the state of Rio de Janeiro, B. bilineatus was considered to be extinct, since it was last recorded in 1963 (73). In the state of the Espírito Santo, B. bilineatus was considered vulnerable (78) and, in Minas Gerais, Feio and Caramaschi (79, 80) proposed that this species be included in the Red List of Threatened Species for the state. Throughout its geographical distribution in the Atlantic Forest, it may have had a reduction in its populations and even local extinctions, since it is a forest and arboreal snake, which is probably being harmed by the deforestation that this biome has suffered.
The characterization of the composition of snake venoms is a key step for understanding the evolution and adaptive advantages of each phenotype. It allows us to predict the essential biological events that are triggered by each toxin family and that are responsible for subduing the prey and result in the fitness of the snake species (81). The presence and abundance of different toxins in the venoms also affect the clinical manifestations of patients afflicted by snakebites and indicate the presence of cross-reactive antigens responsible for the efficacy of the antivenom therapy (82). Thus, several studies have attempted to characterize the protein composition of the venoms and their correlation with the mechanisms by which the venoms are capable of inducing the biological effects. Among the main activities evaluated, neurotoxicity, hemostatic disturbances, tissue or cell cytotoxicity, and inflammation are the most studied and use either the whole venoms or isolated toxins. The data relating to B. bilineatus venom are highlighted in this section.
Only a few reports on the characterization of B. bilineatus venom-derived components are available. However, two recent venomics studies have been published, and contributed with important evidence about the composition of venom from B. bilineatus subspecies bilineatus and smaragdinus (83, 84). The authors analyzed samples of pooled venoms from Peruvian B. b. smaragdinus specimens, and also pooled venoms of two B. b. smaragdinus specimens originally from Rondonia state, Brazil, and the venom from a single specimen of B. b. bilineatus from Rondonia state that had been kept frozen and crystalized for more than 10 years. Interestingly, despite being of different subspecies, geographical origin or conservation conditions, the venom samples exhibited very similar proteomic profiles that comprised components belonging to the most frequent venom protein classes. Snake venom metalloproteinases (SVMPs) were the most abundant toxins with the predominance of the PIII-class; snake venom serine proteinases (SVSP), C-type lectin-like proteins (CTL), phospholipases A2 (PLA2), cysteine-rich secretory proteins (CRISP), and L-amino acid oxidases (LAAO) were also detected in all samples, representing the major components of their venom proteomes. Bradykinin-potentiating-like peptides (BPPs) and the tripeptide inhibitors of SVMPs were also abundant in the three samples analyzed in one of the studies (83, 84). Snake venom vascular endothelial growth factor, nerve growth factor, 5′-nucleotidase, phosphodiesterase, phospholipase B, and others were also detected in minor amounts in venom samples (83, 84). The relative abundance and differences of the major protein families in these venoms are represented in Table 2.
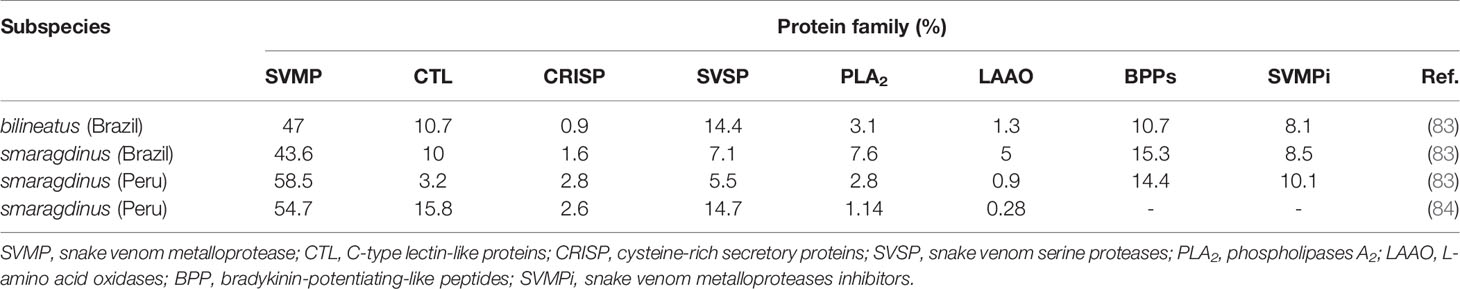
Table 2 Relative abundance (%) of the major toxin groups in venoms of Bothrops bilineatus subspecies.
Considering the most abundant and functionally related proteins, SVMPs comprise a group of zinc-dependent proteases, with a multi-domain organization including catalytic domains bearing the zinc-binding motif and adhesive domains recognized as disintegrin, disintegrin-like, or cysteine-rich domains (85). In the venoms, the two predominant forms are SVMPs from P-I and P-III classes and they are the major toxins in venoms from most species of Bothrops snakes (86). Accordingly, in B. bilineatus spp, the major toxin group was the SVMPs (83, 84), particularly of the PIII-class (83). These enzymes contain catalytic, disintegrin-like, and cysteine-rich domains. As described for the structurally related toxins present in other Bothrops venoms, PIII-class SVMPs are functionally versatile and are involved in most of the local and systemic effects induced by such venoms (87), including the local and systemic hemorrhages (88, 89), inhibition of collagen-induced platelet-aggregation (90–92) and activation of coagulation factors (93–95). SVMPs are also able to induce potent activation of inflammatory mediators (87) and release bioactive cryptides from the extracellular matrix or plasma proteins that act as VAMPs (venom-associated molecular patterns), which activate inflammatory cell Toll-like receptors (96, 97).
SVSPs, are the second most abundant enzymes in B. bilineatus venom samples (83, 84). Unlike the SVMPs, most SVSP activities are concentrated on the coagulation dysfunctions induced by Bothrops venoms. As reported by Serrano (2013), SVSPs are catalytically active proteins that are able to catalyze the polymerization of fibrinogen into fibrin and are commonly termed thrombin-like enzymes. SVSPs also activate coagulation factors and induce platelet aggregation by cleavage of PAR1-receptors. Anti-coagulant activities of SVSPs have also been described including the activation of plasminogen and protein C, as well as the kallikrein-like activity (98). Disorders of hemostasis are probably enhanced by the presence of considerable amounts of CTLs in B. bilineatus samples. CTLs are non-enzymatic components that are functionally relevant in the hemostatic disturbances, which are mostly due to their action on different platelet receptors, and induce aggregation (99) that may lead to the thrombocytopenia observed in some snakebite patients (100). CTLs are also inhibitors of thrombin activity (101, 102).
Different than other venoms of Bothrops snakes, the samples of B. bilineatus spp venom analyzed by proteomics (83, 84) contained minor amounts of PLA2s. PLA2s occur in a large variety of venoms, including the venoms of Bothrops snakes in basic and acidic isoelectric points (86). Considering the physiopathology that results from Bothrops venoms, basic PLA2s appear to be the most relevant due to their myotoxicity, while the acidic PLA2s are mostly anti-coagulant (103, 104). Interestingly, B. bilineatus venom proteomics evidenced low amounts of PLA2s, mostly in the acidic form (83, 84).
In addition, although the composition of venoms may vary depending on the region where the snake is found, other studies have shown very similar median lethal dose (LD50) values for Brazilian (102.92 μg) and Peruvian (92.74 μg) B. bilineatus in mice (18-20 g) (83, 84).
Bothrops bilineatus is classified as Highest Medical Importance (Category 1) in Ecuador, Peru, Colombia and Guyana and of Secondary Medical Importance (Category 2) in Brazil, Bolivia and Venezuela. Although they are highly venomous snakes, exact epidemiological or clinical data are lacking and they are less frequently implicated because of their behavior, habitat preferences, or occurrence in areas remote from large human populations.
The abundance of the above venom-derived toxins in B. bilineatus venoms thus explains the signs of envenomations presented by human patients bitten by these snakes, including the local inflammatory reaction, local tissue damage, and coagulation dysfunctions (14, 105, 106). The few reports of snakebites confirmed as being by B. bilineatus demonstrate that these envenomations lead to clinical manifestations similar to those caused by other Bothrops species, as evidenced in Table 3.

Table 3 Clinical aspects of envenomations by Bothrops bilineatus.
Regarding the pathophysiological effects resulting from the additive or synergistic action of the toxins, local effects represent a hallmark event during Bothrops snakebite. The event consists of the local action of venom toxins that are capable of inducing tissue damage, intense local inflammatory reaction, and muscular impairment, in which SVMPs and PLA2 are important components in this sense (107). Associated with the low capacity of antivenom neutralization in the region of the bite, the consequence of local effects can cause longer hospitalization periods and, in some cases, the impairment of limb function (108, 109). After being bitten by B. bilineatus, the main local manifestations are characterized by edema, increase local temperature, blister formation and secondary infection (Table 3).
During the feeding/capture process, the action of snake venom toxins to block the neuromuscular function of animals is one of the venom strategies to promote prey paralysis and involves myotoxic and neuro-blocking action. In this manner, the group of PLA2, found in the venoms as catalytically active or without enzymatic function, represents the main components responsible for these effects (110). The presence of phospholipase activity in B. bilineatus venom was reported in early studies conducted by Cadilio and colleagues (1991), though it displays lower activity when compared to other venoms from Bothrops, Crotalus, and Micrurus snakes, which is possibly due to the low amounts observed in B. bilineatus venom proteomics (83, 84, 111). Carregari and colleagues isolated the first PLA2 from B. bilineatus venom, named Bbil-TX, which consists of a calcium-dependent basic PLA2 (Asp49) (112). A series of studies have been performed since then in order to understand the action of both crude B. bilineatus venom and isolated Bbil-TX on myotoxicity and neuromuscular impairment. From a damage perspective, both venom and toxin were capable of inducing ultrastructural muscular alterations in mouse isolated nerve-phrenic diaphragm preparations in vitro, such as hypercontraction of myofilaments, disorganization of sarcomeres, and sarcoplasmic reticulum and mitochondrial damage (113, 114). There are no reports on in vivo myotoxicity using crude B. bilineatus venom; however, Bbil-TX has induced local myotoxicity in mice after IM administration (represented by increased levels of serum CK peaking at 2 h) with neglected systemic myotoxicity when administrated IV (112). Although the direct myotoxicity to muscle fibers has been reported, studies focused on the neuromuscular impairment caused by B. bilineatus venom and Bbil-TX indicate that these alterations were not enough to promote disorder in the contractile mechanism of the muscle. Crude B. bilineatus venom and Bbil-TX have also been reported to induce in vitro neuromuscular blockade using neuromuscular preparations. Moreover, they did not alter the muscle membrane resting potential or the response of endogenous and exogenous agonists, thus suggesting a presynaptic action and not muscle contraction impairment (113, 115, 116). Moreover, the presynaptic action of Bbil-TX involves modulation of potassium channel activity and presynaptic protein expression, as well as ultrastructural nerve alterations characterized by detachment of the axon from the myelin sheath and formation of periaxonal vacuoles (114, 116).
The inflammatory response involving Bothrops venom is an intriguing event during envenomation. The systemic and local immune responses are responsible for triggering several alterations and are considered one of the most important aspects involving local clinical manifestations such as edema and tissue damage. The mechanism involves the direct recognition of venom toxins named VAMPS (117) by immune cells receptors (such as toll-like receptors) and the release of soluble mediators (complement system, cytokines, and chemokines) by the direct action of toxins as well as by the hydrolysis products released from toxins by tissue degradation (extracellular matrix and cell damage products).
B. bilineatus venom was found to promote neutrophil migration to the peritoneal cavity, mediated mainly by metalloproteases. Moreover, dexamethasone and zileuton (a 5-lipoxygenase inhibitor) were capable of reducing cell infiltration, indicating that lipid mediators are involved in the venom-induced response (97, 118). As demonstrated for other snake toxins (119–121), the PLA2 Bbil-TX was found to induce paw edema and increased serum levels of TNF-α, IL-6, and IL-1 after IM administration (112). In addition to the main pro-inflammatory cytokines studied, Setubal et al. were the first to demonstrate that B. bilineatus venom induces IL-8 and PGE2 release by neutrophils. In addition, the research group showed that the referred venom stimulates neutrophil extracellular traps (NETs) (122). Indeed, the production of inflammatory mediators, phagocytosis, and the formation of NETS contribute to the clearance of necrotic material, which are important to the reparative and regenerative process after snakebite envenomation. On the other hand, the same components and formation of NETs could be responsible for the pathogenesis of local tissue damage (123). Thus, the role of neutrophils after B. bilineatus envenomation needs to be further elucidated.
In regards to systemic effects, B. bilineatus venom is capable of promoting hemostatic alterations. This is reported for venoms from other Bothrops species and is due to the abundance of SVMPs, SVSPs, and CTLs, which are hemostatically active toxins that are capable of interfering in several events such as the coagulation cascade, platelet function, and fibrinolysis. Moreover, SVMPs are potent hemorrhagins that consist of a class of toxins capable of acting on extravascular targets, such as matrix extracellular components of endothelial basal membranes, in order to induce vessel disruption and consequently local and systemic hemorrhage (124, 125). Rodriguez and colleagues reported that crude B. bilineatus venom presented a procoagulant activity in human plasma and fibrinogen; however, it was less potent than that of the Peruvian bushmaster Lachesis muta (126, 127). Bilinearin is a 23kDa glycosylated PI-metalloprotease isolated from B. bilineatus venom and is capable of inducing prothrombin activation in calcium and phospholipid-independent manner (127). The toxin did not induce isolated fibrinogen clots and was capable to induce human plasma coagulation through prothrombin activation (127). Therefore, the above-mentioned results demonstrate that B. bilineatus venom is responsible for a procoagulant activity mediated by the presence of prothrombin activator toxin(s), as well as the presence of thrombin-like toxin(s). As a result, B. bilineatus venom was found to promote hemostatic alterations, as cited by Silva (128). The author observed thrombus formation in medium-caliber vessels (such as the pulmonary artery) after histopathological analysis of the lungs of rabbits administered 0.5 mg/Kg of crude B. bilineatus venom via both IM and IV routes. As a consequence of Bothrops envenomations, the venom triggers blood clot formation, and leads to two important possible consequences: (1) the development of an unclottable state, associated with the consumption of hemostatic factors, and culminating in hemorrhagic events; and (2) the formation of intravascular thrombus, resulting in the reduction/cessation of tissue permeability and causing end-organ failure (125). Based on the results of in vitro and in vivo studies of B. bilineatus venom, the procoagulant activity of the venom, mediated by prothrombin activation and thrombin-like activities, is responsible for intravascular coagulation and, consequently, the deposition of thrombus. Although the consumption of coagulation factors has never been reported, it is plausible that the venom interferes in this manner and promotes an unclottable state, which is observed in B. bilineatus patients. More clinical and pre-clinical studies on hemostasis involving B. bilineatus envenomations are needed to improve our knowledge regarding this important event.
Within the geographical distribution of Bothrops bilineatus, other species of the genus that are more abundant locally, such as B. atrox in the Amazon and B. jararaca in the Atlantic Forest, are more common causes of snakebites (3). As such, its rarity or the difficulty in finding this snake in much of its distribution should reflect in the low occurrence of cases of snakebite and also of published clinical reports [e.g. (105, 106)] however, in some locations, B. bilineatus is shown to be more frequent: 15% of cases in Letícia (Amazonas, Colombia) (129), 36% in the Pastaza region (Ecuador) (39) and 5.3% in Alto Juruá (Acre, Brazil) (72).
In the Alto Juruá region, Bothrops bilineatus was the most commonly found species during night searches, and corresponded to 54% of the snakes, which was more than B. atrox (20.7% of the encounters) (7). However, in areas of terra firme in this region, although B. bilineatus is present, it is less easy to find and in some studies of species surveys it has not been recorded [e.g. (50, 130)]. Despite its greater abundance in lowland forests in Alto Juruá, snakebites caused by B. bilineatus still occur less frequently compared to B. atrox (72), which is probably because it is a species that is associated with forested environments and also due to its arboreal habits (8). The average height in which B. bilineatus is found in hunting and resting activity is 6.4 m (8), well above the height of a human being, which should contribute to the lower frequency of encounters and snakebites by this snake. Nonetheless, B. bilineatus is also found on vegetation at lower heights (less than 2 m in height) and even close to the ground (heights of 30 to 40 cm) (6, 8), which can result in a snakebite when someone accidently bumps into it or approaches it. The occurrence of the bite in the upper regions of the body (fingers of the hand, hand, arm, scapular region, chest and head) is a feature usually associated with envenomation by this snake (13, 105, 106, 129).
Snakebites caused by B. bilineatus occur mainly during activities within forests, such as during the opening up of new trails [e.g. (41)] and also extractivism of acai palm (Euterpe precatoria) (13). The green coloration of B. bilineatus makes it relatively more difficult to spot since it is well camouflaged in this environment (Figure 3). When walking in forests, people can bump into this snake on the vegetation and receive a defensive strike, thus envenomation occurs. Wallace (131) mentioned that indigenous people reported that a snake of the genus Craspedocephalus (possibly C. bilineatus, which was later synonymized with B. bilineatus) was very often found on the “piaçaba” (Leopoldinia piassaba) and that they were often bitten during the extractivism of this palm. In two cases of snakebite caused by B. bilineatus during the acai extractivism, the extractivists were 4 and 8 m high climbing the acai palm and the snake was on a neighboring tree (13). Silva et al. (7) interviewed 100 people who develop some type of activity (hunting, fishing, extractivism) in lowland forests in Alto Juruá and eleven of them reported having already found the “papagaia” (regional, popular name of B. bilineatus) on the acai palm or on a neighboring tree. The evidence indicates that B. bilineatus usually hunts waiting at the top of the acai trees or in the neighboring trees, waiting for small mammals (rodents and marsupials) that will feed on the fruits of acai, and may eventually bite the extractivists (13).
The specific treatment for the victims of B. bilineatus snakebite is the use of animal-derived antivenoms. So far, there are three different antivenoms (AV) that can be used to treat B. bilineatus envenomation: (1) Bothrops AV (2) Bothrops-Crotalus AV and (3) Bothrops-Lachesis AV (132). Although these three types of antivenoms exist, none are manufactured from the immunization of horses with the venoms of Amazonian Bothrops species (Figure 4 and Table 4).
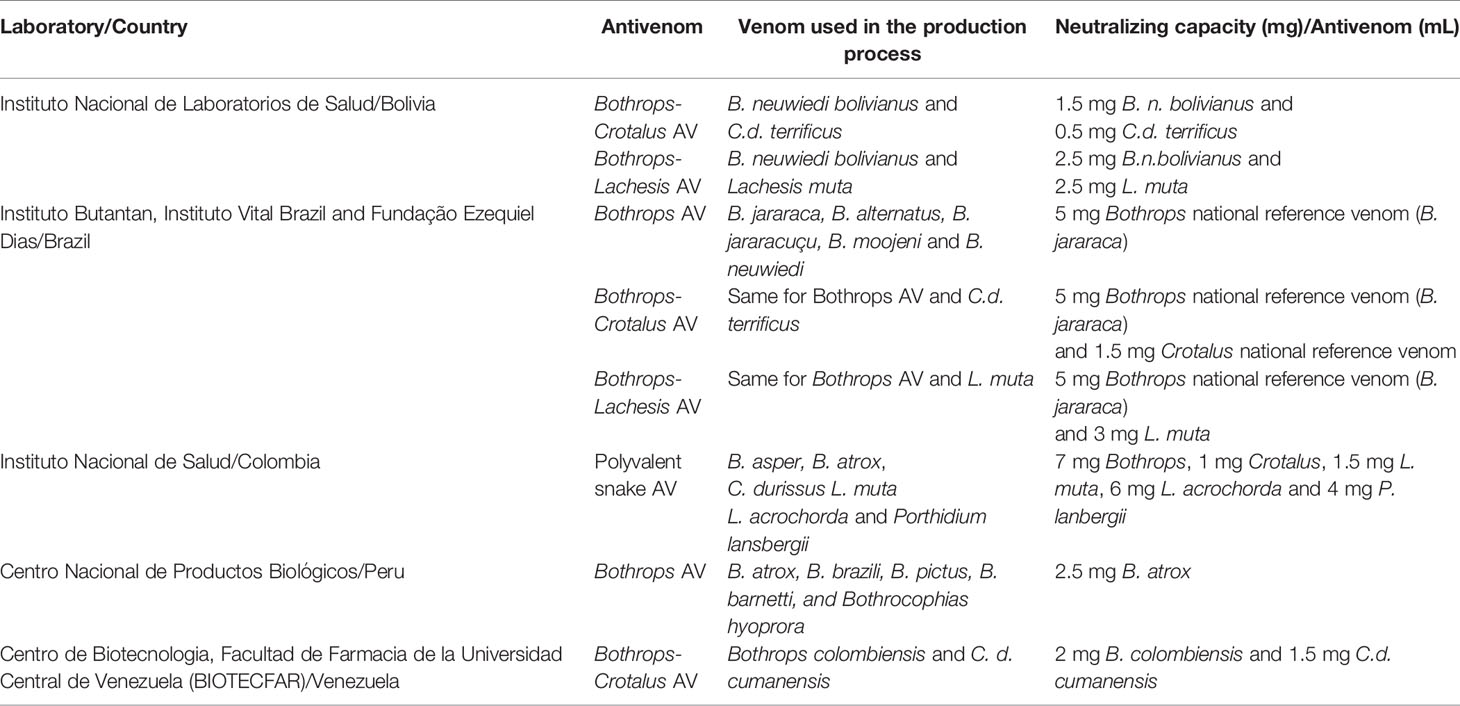
Table 4 South American antivenom-manufacturing laboratories and antivenoms manufactured by them.
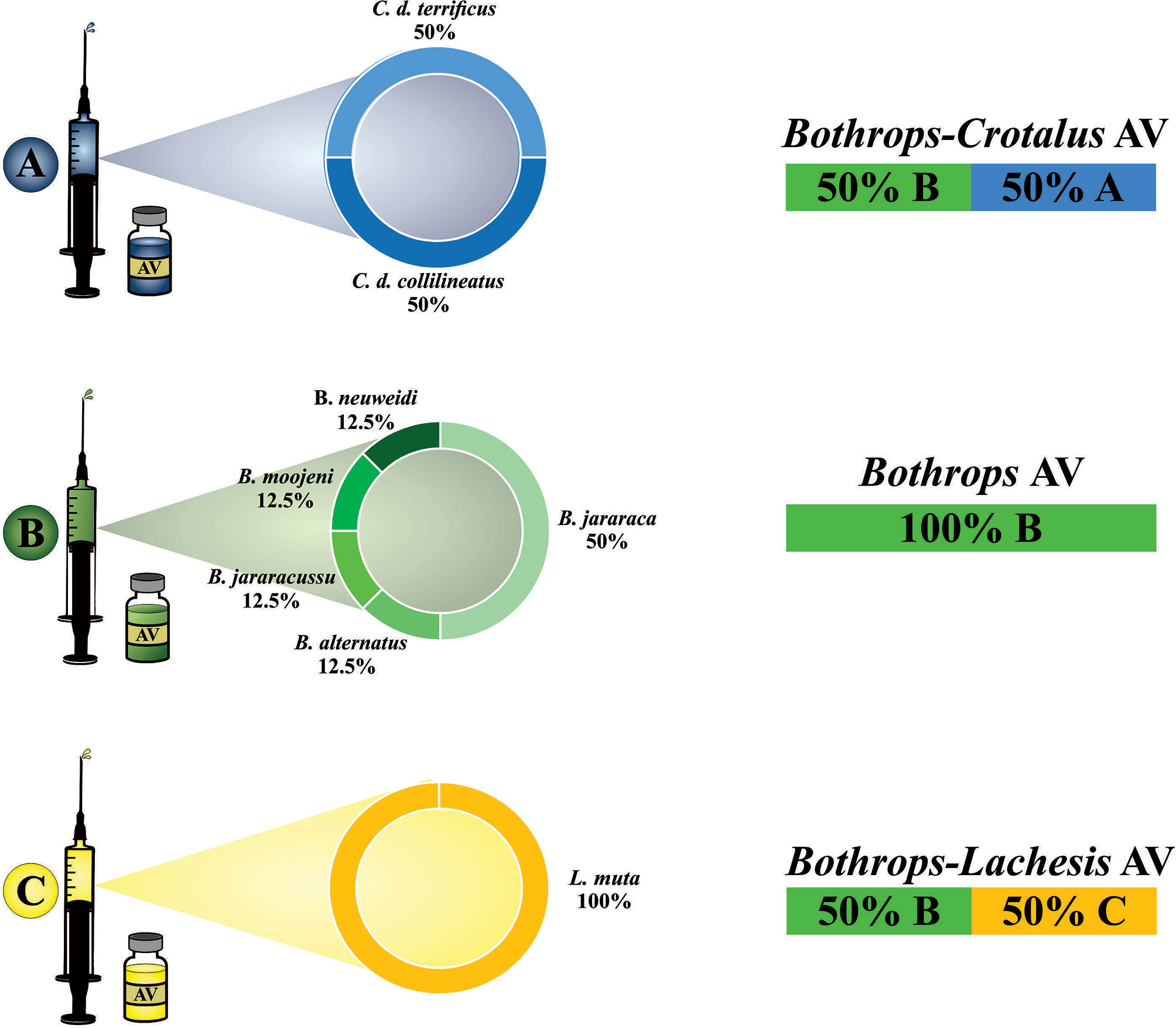
Figure 4 Brazilian antivenoms: venoms used as antigens for horse immunization to obtain specific neutralizing antibodies, and antivenoms currently available in Brazil. These are manufactured from the three different hyperimmune plasma: (A) Bothrops-Crotalus AV, (B) Bothrops AV, and (C) Bothrops-Lachesis AV (right), which can be used to treat B. bilineatus envenomations.
A previous study has already shown that Bothrops AV, although specific to Bothrops genus, demonstrated to recognize and neutralize Bothrops venoms from the Amazon Rainforest when compared to B. jararaca venom (133), including the venoms of B. atrox and B. bilineatus. Indeed, an antivenom study demonstrated that the pentavalent antivenom produced by Butantan Institute (Bothrops AV) was able to recognize B. bilineatus venom by cross-reactivity; however, when B. b. bilineatus proteins were analyzed individually, it demonstrated low reactivity for some of them (83).
None of the antivenom-manufacturing laboratories in South America include B. bilineatus venom as an antigen (Table 4). Clinical studies are scarce and tend to confirm the efficacy of the standard doses recommended in the national guidelines. In one large study performed in the Ecuadorian Amazon, blood coagulation disorders were reverted in almost 68 patients envenomed by B. bilineatus (identified by a herpetologist when the snake was brought or by immunoassay determination) within 24 hours after starting treatment with Colombian, Brazilian and Ecuadorian antivenoms (39). Likewise, a case report from the Brazilian Atlantic Forest shows a good clinical response in B. bilineatus envenomations when treated with the Brazilian antivenom (105). In contrast, a case reported in the Brazilian Amazon basin revealed a delay in the normalization of the coagulation test, which occurred on day 4-9 of hospitalization, and platelet counts that returned to normal almost 2 weeks after administration of the specific antivenom (106).
The diversity among antivenoms may be evidenced by the different antigens used in the manufacturing process of antivenoms and the values of their neutralizing capacities. As a consequence, treatment schedules vary from country to country. For instance, the Brazilian Ministry of Health establishes 2-4, 4-8 and 12 vials for mild, moderate, and severe Bothrops cases, regardless of the species responsible for the envenoming, while the package insert of the Colombian antivenom recommends 2, 4, and 8 vials, according to the severity of the envenomation. Although both liquid antivenoms are supplied in vials of 10 mL, antigens and neutralizing capacities are different, and the ability of these antivenoms to reverse clinical manifestations of B. bilineatus envenomations should be confirmed.
Besides the use of specific antivenoms, envenomation caused by Bothrops snakes also requires other clinical approaches. Blood pressure monitoring and hydration should be carried out to avoid hypovolemia (134–136). For the management of compartmental syndrome, the conservative attitude is the best approach, and fasciotomy should be indicated with caution (137). Surgical debridement of necrotic tissue in the affected limb may require long-term follow-up to avoid functional or permanent disability.
The use of broad-spectrum antibiotics, such as ampicillin, ceftriaxone, ciprofloxacin, and clindamycin, should be considered, as well as tetanus prevention measures, except for the administration of the tetanus vaccine, which should be avoided within 48 hours after the bite and/or if coagulopathies persist (136). To avoid acute kidney injury (AKI), the use of vasoactive and diuretic substances must be judicious, as well as hydroelectrolytic correction (134, 138).
Future molecular and taxonomic studies should reveal the status of the species within the complex Bothrops bilineatus, which will be the basis for research on the variation of toxins and for the comparison of interspecific bioecology. Regarding bioecology, further research is needed to better elucidate the abundance, seasonal patterns and living area of B. bilineatus in the locations where it is most frequent, a lowland forest in the western Brazilian Amazon.
Some species of Bothrops that present ontogenetic changes in the diet, whose juveniles predate mainly ectothermic prey (e.g., anurous amphibians and lizards) and endothermic adults (e.g., rodents), are characterized by differences in the composition of their toxins that are related to the animal groups on which they feed. The hunting tactic of caudal decoy to attract amphibians that is widely used also by adult individuals instigates the conduct of studies on possible ontogenetic variation of toxins.
The scarcity of clinical reports of poisoning by B. bilineatus makes the knowledge about the therapy relatively incipient, which when associated with the peculiar epidemiological circumstances of the snakebite usually involving higher regions of the body and associated with human activities within forests, provides the need to expand the clinical and epidemiological studies with this snake.
The snake Bothrops bilineatus is a species complex encountered in forests of the Amazon and the Atlantic forest, which presents arboreal habits and different population densities throughout its geographical distribution, being generally uncommon in some areas and relatively more abundant in others. These differences in the abundance of B. bilineatus probably reflect in the proportion of cases of snakebites in each region and their rarity in much of its geographical distribution.
The arboreal and forest habits of B. bilineatus, usually occurring at heights greater than the height of a human being, tends to link the snakebites with human activities within the forest (hunting, fishing) when it is on the lower strata of the vegetation or when the person is climbing palm trees during extractivism. Its green coloration and sedentary behavior make B. bilineatus well camouflaged in the environment, which increases the chances of it not being seen and, when approached or touched by a human, its defensive response often ends in an envenomation. Since extractivism often requires climbing palm trees and other trees, care and attention are essential in order to prevent envenomations by this snake.
PS and WM conceived the main idea of this work. All authors designed and wrote most of this review’s topics. FC, MA, and WF elaborated the figures of this review article. All authors corrected the manuscript and provided important contributions during the development of this work. All authors contributed to the article and approved the submitted version.
We thank Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, The National Council for Scientific and Technological Development, scholarship to MP no. 307184/2020-0, WM n. 309207/2020-7, to AMM-d-S no. 303958/2018-9, and PS 311509/2020-7) and Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP, São Paulo Research Foundation; research grant 2016/50127-5; scholarship to IO no. 2020/13176-3). WM acknowledges funding support from Fundação de Amparo à Pesquisa do Estado do Amazonas (PAPAC ̧ 005/2019, PRO-ESTADO and Posgrad calls). MP (Snakebite Roraima project coordinator) acknowledges funding support from Hamish Ogston Foundation - Global Snakebite Initiative.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2021.778302/full#supplementary-material
1. Dal Vechio F, Prates I, Grazziotin FG, Zaher H, Rodrigues MT. Phylogeography and Historical Demography of the Arboreal Pit Viper Bothrops Bilineatus (Serpentes, Crotalinae) Reveal Multiple Connections Between Amazonian and Atlantic Rain Forests. J Biogeography (2018) 45:2415–26. doi: 10.1111/jbi.13421
2. Griffith GE, Omernik JM, Azevedo SH. Ecological classification of the Western Hemisphere. Unpublished report. Corvallis: Environmental Protection Agency, Western Ecology Division (1998). p. 49.
3. Campbell JA, Lamar WW. The Venomous Reptiles of the Western Hemisphere. New York, USA: Cornell Univ. Press, Ithaca (2004).
4. Cunha OR, Nascimento FP. Ofídios Da Amazônia. As Cobras Da Região Leste do Pará. Bol Mus Para Emílio Goeldi Sér Zool (1993) 9:1–191.
5. Bernarde PS, Abe AS. A Snake Community at Espigão do Oeste, Rondônia, Southwestern Amazon, Brazil. South Am J Herpetology (2006) 1:102–13. doi: 10.2994/1808-9798(2006)1[102:ASCAED]2.0.CO;2
6. Turci LCB, de Albuquerque S, Bernarde PS, Miranda DB. Uso do Hábitat, Atividade E Comportamento De Bothriopsis Bilineatus E De Bothrops Atrox (Serpentes: Viperidae) Na Floresta do Rio Moa, Acre, Brasil. Biota Neotrop (2009) 9:197–206. doi: 10.1590/S1676-06032009000300020
7. da Silva JL, da Fonseca WL, Mota da Silva A, do Amaral GLG, Ortega GP, Oliveira A deS, et al. Venomous Snakes and People in a Floodplain Forest in the Western Brazilian Amazon: Potential Risks for Snakebites. Toxicon (2020) 187:232–44. doi: 10.1016/j.toxicon.2020.09.007
8. da Fonseca WL, Correa RR, Oliveira A deS, de Oliveira IS, Bernarde PS. Habitat Use and Activity of Bothrops Bilineatus Smaragdinus Hoge, 1966 in the Western Brazilian Amazon (Serpentes: Viperidae). Herpetology Notes (2021) 14:567–80.
9. Argôlo AJS. As Serpentes Dos Cacauais do Sudeste Da Bahia. Ilhéus, Bahia, Brazil: Editora da UESC (2004).
10. da Cunha OR. Ofídios Da Amazônia I - A Ocorrência De Bothrops Bilineatus Bilineatus (Wied) Nas Matas Dos Arredores De Belém, Pará (Ophidia, Crotalidae). Boletim do Museu Paraense Emílio Goeldi Nova série zoologia (1967) 66:1–12.
11. Pierini SV, Warrell DA, De Paulo A, Theakston RDG. High Incidence of Bites and Stings by Snakes and Other Animals Among Rubber Tappers and Amazonian Indians of the Juruá Valley, Acre State, Brazil. Toxicon (1996) 34:225–36. doi: 10.1016/0041-0101(95)00125-5
12. Waldez F, Vogt RC. Aspectos Ecológicos E Epidemiológicos De Acidentes Ofídicos Em Comunidades Ribeirinhas do Baixo Rio Purus, Amazonas, Brasil. Acta Amaz (2009) 39:681–92. doi: 10.1590/S0044-59672009000300025
13. Mota-da-Silva A, Sachett J, Monteiro WM, Bernarde PS. Extractivism of Palm Tree Fruits: A Risky Activity Because of Snakebites in the State of Acre, Western Brazilian Amazon. Rev Soc Bras Med Trop (2019) 52:e20180195. doi: 10.1590/0037-8682-0195-2018
14. Bernarde PS. Serpentes Peçonhentas E Acidentes Ofídicos No Brasil Anolis Books. São Paulo (2014).
15. Carrasco PA, Mattoni CI, Leynaud GC, Scrocchi GJ. Morphology, Phylogeny and Taxonomy of South American Bothropoid Pitvipers (Serpentes, Viperidae). Zoologica Scripta (2012) 41:109–24. doi: 10.1111/j.1463-6409.2011.00511.x
16. Carrasco PA, Grazziotin FG, Farfán RSC, Koch C, Ochoa JA, Scrocchi GJ, et al. A New Species of Bothrops (Serpentes: Viperidae: Crotalinae) From Pampas Del Heath, Southeastern Peru, With Comments on the Systematics of the Bothrops Neuwiedi Species Group. Zootaxa (2019) 4565:301. doi: 10.11646/zootaxa.4565.3.1
17. Salomão MG, Wüster W, Thorpe RS, Touzet JM. DNA Evolution of South American Pitvipers of the Genus Bothrops (Reptilia: Serpentes: Viperidae). Ann Symp Zool Soc London (1997) 70:89–98.
18. Salomão MG, Wüster W, Thorpe RS. MtDNA Phylogeny of Neotropical Pitvipers of the Genus Bothrops (Squamata: Serpentes: Viperidae). Kaupia (1999) 8:127–234.
19. Vidal N, Lecointre G, Vié JC, Gasc JP. Molecular Systematics of Pitvipers: Paraphyly of the Bothrops Complex. Comptes Rendus l’Académie Des Sci - Ser III - Sci la Vie (1997) 320:95–101. doi: 10.1016/S0764-4469(99)80091-2
20. Parkinson CL. Molecular Systematics and Biogeographical History of Pitvipers as Determined by Mitochondrial Ribosomal DNA Sequences. Copeia (1999) 1999:576. doi: 10.2307/1447591
21. Parkinson CL, Campbell JA, Chippindale PT. Multigene Phylogenetic Analysis of Pitvipers, With Comments on Their Biogeography. In: Schuett GW, Höggren M, Douglas ME, Greene HW, editors. Biology of the Vipers. Salta Lake City: Eagle Mountain Publishing (2002). p. 93–110.
22. Wüster W, Salomão MG, Quijada-Mascareñas JA, Thorpe RS. Origin and Evolution of the South American Pitviper Fauna: Evidence From Mitochondrial DNA Sequence Analysis. In: Schuett GW, Höggren M, Douglas ME, Greene HW, editors. Biology of the Vipers. Eagle Mountain: Eagle Mountain Publishing (2002). p. 111–28.
23. Castoe TA, Parkinson CL. Bayesian Mixed Models and the Phylogeny of Pitvipers (Viperidae: Serpentes). Mol Phylogenet Evol (2006) 39:91–110. doi: 10.1016/j.ympev.2005.12.014
24. Fenwick AM, Gutberlet RL, Evans JA, Parkinson CL. Morphological and Molecular Evidence for Phylogeny and Classification of South American Pitvipers, Genera Bothrops, Bothriopsis, and Bothrocophias (Serpentes: Viperidae). Zoological J Linn Soc (2009) 156:617–40. doi: 10.1111/j.1096-3642.2008.00495.x
25. Peters JA, Orejas-Miranda B. Catalogue of the Neotropical Squamata. Part I. Snakes. Bull United States Natl Museum (1970) 297:1–347. doi: 10.5479/si.03629236.297.1
26. Campbell JA, Lamar WW. The Venomous Reptiles of Latin America. Ithaca: Comstock Pub. Associates (1989).
27. Burger WL. Genera of Pitvipers. Unpublished PhD thesis. University of Kansas Lawrence, Kansas, USA (1971).
28. Hoge AR, Lancini AR. Sinopsis De Las Serpientes Venenosas De Venezuela. In: Grafos E, editor. Publicaciones Ocasionales Del Museo De Ciencias Naturales. Caracas, Venezuela (1962).
29. Hoge AR. Preliminary Account on Neotropical Crotalinae (Serpentea Viperidae). Memorias do Instituto Butantan (1965) 32:109–84.
30. Bernarde PS, Costa HC, Machado RA, São-Pedro VA. Bothriopsis Bilineata Bilineata (Wied, 1821) (Serpentes: Viperidae): New Records in the States of Amazonas, Mato Grosso and Rondônia, Northern Brazil. Check List (2011) 7:343–7. doi: 10.15560/7.3.343
31. Nogueira CC, Argôlo AJS, Arzamendia V, Azevedo JA, Barbo FE, Bérnils RS, et al. Atlas of Brazilian Snakes: Verified Point-Locality Maps to Mitigate the Wallacean Shortfall in a Megadiverse Snake Fauna. South Am J Herpetology (2019) 14:1–274. doi: 10.2994/SAJH-D-19-00120.1
32. Dal Vechio FH. Revisão Sistemática De Bothrops Bilineatus (Serpentes: Viperidae) Com Base Em Caracteres Moleculares E Morfológicos. (2014). doi: 10.11606/D.41.2015.tde-24032015-083805
33. Sandner–Montilla F. Mini Información Sobre Serpientes Venezolanas. Caracas: Instituto Venezolano de Ofidiologia (1994).
35. Harvey MB, Aparicio J, Gonzales L. Revision of the Venomous Snakes of Bolivia. II: The Pit Viper (Serpentes: Viperidae). Cara (2005) 74:1–37. doi: 10.2992/0097-4463(2005)74[1:ROTVSO]2.0.CO;2
36. da-Silva NJ Jr. The Snakes From Samuel Hydroelectric Power Plant and Vicinity, Rondônia, Brazil. Herpetol Nat Hist (1993) 1:37–86.
37. Almeida BJL, Almeida MSM, Cavalcante CS, Bernarde PS. Bothrops Bilineatus Bilineatus (Two-Striped Forest Pitviper) Reproduction. Herpetological Rev (2019) 50:385–6.
38. Grego KF, Fernandes W, Croce AP, Vasconcellos DR, Sant’Anna SS, Coragem JT. Bothriopsis Bilineatus Smaragdinus (Green Jararaca) Reproduction. Herpetological Rev (2012) 43:492.
39. Smalligan R, Cole J, Brito N, Laing GD, Mertz BL, Manock S, et al. Crotaline Snake Bite in the Ecuadorian Amazon: Randomised Double Blind Comparative Trial of Three South American Polyspecific Antivenoms. BMJ (2004) 329:1129. doi: 10.1136/bmj.329.7475.1129
40. Dixon JR, Soini P. The Reptiles of the Upper Amazon Basin, Iquitos Region, Peru. 2nd ed. Milwaukee, WI, USA: Milwaukee Public Museum (1986).
41. Duellman WE. The Biology of an Equatorial Herpetofauna in Amazonian Ecuador. Miscellaneous Publications, Museum of Natural History Vol. 65. USA: University of KansasLawrence: University of Kansas (1978) p. 1–352.
42. Cunha OR da, Nascimento FP do, Ávila-Pires TCS de. Os Répteis Da Área De Carajás, Pará, Brasil (Testudines E Squamata). Publicações Avulsas do Museu Paraense Emilio Goeldi Belém (1985) 40:9–92.
43. Frota JG, Santos-Jr AP, Menezes-Chalkidis H, Guedes AG. As Serpentes Da Região do Baixo Rio Amazonas, Oeste do Estado do Pará, Brasil (Squamata). Biociências (2005) 13:211–20.
44. Pantoja DL, de Fraga R. Herpetofauna of the Reserva Extrativista do Rio Gregório, Juruá Basin, Southwest Amazonia, Brazil. cl (2012) 8:360. doi: 10.15560/8.3.360
45. Frazão L, Oliveira ME, Menin M, Campos J, Almeida A, Kaefer IL, et al. Species Richness and Composition of Snake Assemblages in Poorly Accessible Areas in the Brazilian Amazonia. Biota Neotrop (2020) 20:e20180661. doi: 10.1590/1676-0611-bn-2018-0661
46. Souza-Costa CA, Mira-Mendes CVD, Iuri Ribeiro D, Kaique Brito S, Argôlo AJS, Solé M. Squamate Reptiles From Seasonal Semi-Deciduous Forest Remnantsin Southwestern Bahia, Brazil. Bonn Zoological Bull (2020) 69:85–94. doi: 10.20363/BZB-2020.69.1.085
47. Debien IV, Waldez F, Menin M. Diversity of Reptiles in Flooded and Unflooded Forests of the Amanã Sustainable Development Reserve, Central Amazonia. Herpetology Notes (2019) 12:1051–65.
48. Martins M, Oliveira ME. Natural History of Snakes in Forests of the Manaus Region, Central Amazonia, Brazil. Herpetological Natural History (1998) 6:78–150.
49. Prudente AL da C, Maschio GF, dos Santos-Costa MC, Feitosa DT. Serpentes Da Bacia Petrolífera De Urucu, Município De Coari, Amazonas, Brasil. Acta Amaz (2010) 40:381–6. doi: 10.1590/S0044-59672010000200016
50. Bernarde PS, Machado RA, Turci LCB. Herpetofauna Da Área do Igarapé Esperança Na Reserva Extrativista Riozinho Da Liberdade, Acre - Brasil. Biota Neotrop (2011) 11:117–44. doi: 10.1590/S1676-06032011000300010
51. Zimmerman BL, Rodrigues MT. Frogs, Snakes, and Lizards of the INPA/WWF Reserves Near Manaus, Brazil. In: Gentry AH, editor. Four Neotropical Rainforests. New Haven: Yale University Press (1990). p. 426–54.
52. Fraga R, Lima AP, Prudente ALC, Magnusson WE. Guide to the Snakes of the Manaus Region, Central Amazonia. Manaus: Editora INPA (2013).
53. Masseli GS, Bruce AD, Santos JGD, Vincen T, Kaefer IL. Composition and Ecology of a Snake Assemblage in an Upland Forest From Central Amazonia. Acad Bras Ciênc (2019) 91:e20190080. doi: 10.1590/0001-3765201920190080
54. Waldez F, Menin M, Vogt RC. Diversity of Amphibians and Squamata Reptilians From Lower Purus River Basin, Central Amazonia, Brazil. Biota Neotrop (2013) 13:300–16. doi: 10.1590/S1676-06032013000100029
55. Turci LCB, Bernarde PS. Levantamento Herpetofaunístico Em Uma Localidade No Município De Cacoal, Rondônia, Brasil. In: Títulos Não-Correntes (2012). Available at: http://periodicos.puc-campinas.edu.br/seer/index.php/bioikos/article/view/737 (Accessed June 10, 2021).
56. Moraes L, Almeida A, de Fraga R, Zamora R, Pirani R, Silva A, et al. Integrative Overview of the Herpetofauna From Serra Da Mocidade, a Granitic Mountain Range in Northern Brazil. ZK (2017) 715:103–59. doi: 10.3897/zookeys.715.20288
57. França RC, Morais M, França FGR, Rödder D, Solé M. Snakes of the Pernambuco Endemism Center, Brazil: Diversity, Natural History and Conservation. ZK (2020) 1002:115–58. doi: 10.3897/zookeys.1002.50997
58. Silva-Soares T, Ferreira RB, Salles R deOL, Rocha CFD. Continental, Insular and Coastal Marine Reptiles From the Municipality of Vitória, State of Espírito Santo, Southeastern Brazil | Silva-Soares | Check List. Check List (2016) 7:290–8. doi: 10.15560/7.3.290
59. Martins M, Marques OAV, Sazima I, Höggren M, Douglas ME, Greene HW. Ecological and Phylogenetic Correlates of Feeding Habits in Neotropical Pitvipers (Genus Bothrops). In: Schuett GW, editor. Biology of the Vipers. Eagle Mountain, Utah, USA: Eagle Mountain Publishing (2002). p. 307–28.
60. Beebe W. Field Notes on the Snakes of Kartabo, British Guiana, and Caripito, Venezuela. Zoologica New York (1964) 31:11–52. doi: 10.5962/p.203521
61. Cunha OR, Nascimento FP. Ofídios Da Amazônia X - As Cobras Da Região Leste do Pará. Publ Avul Mus Par Emílio Goeldi (1978) 31:1–218.
62. Carrillo de Espinoza N. Contribución Al Conocimiento De Las Serpientes Venenosas Del Perú De Las Familias Viperidae, Elapidae E Hydrophiidae (Ophidia: Reptilia). Publico Museo Hist Natural “Javier Prado” (1983) 30:1–55.
63. Valencia JH, Garzón-Tello K, Barragán-Paladines ME. Serpientes Venenosas Del Ecuador: Sistemática, Taxonomía, Historia Natural, Conservación, Envenenamiento Y Aspectos Antropológicos. Quito: Fundación Herpetológica Gustavo Orcés, Universidad de Texas (2016).
64. da Fonseca WL, Oliveira A de S, Correa RR, Bernarde PS. Caudal Luring in the Neotropical Two-Striped Forest Pitviper Bothrops Bilineatus Smaragdinus Hoge, 1966 in the Western Amazon. Herpetology Notes (2019) 12:365–74.
65. Venegas PJ, Chávez-Arribasplata JC, Almora E, Grilli P, Duran V. New Observations on Diet of the South American Two-Striped Forest-Pitviper Bothrops Bilineatus Smaragdinus (Hoge, 1966). Cuad Herpetol (2019) 33:29–31. doi: 10.31017/CdH.2019.(2017-031
66. Greene HW, Campbell JA. Notes on the Use of Caudal Lures in Arboreal Green Pit Vipers. Herpetologica (1972) 28:32–4.
67. Sazima I. Natural History of the Jararaca Pitviper, Bothrops Jararaca, in South-Eastern Brazil. In: Campbell JA, Brodie ED, editors. Biology of the Pitvipers. Selva: Tyler (1992). p. 199–216.
68. Oliveira ME, Martins M. When and Where to Find a Pitviper: Activity Patterns and Hábitat Use of the Lancehead, Bothrops Atrox, in Central Amazonia, Brazil. Herpetological Natural History (2001) 8:101–10.
69. Dubeux MJM, Vilela HALS, Andrade AB, Bernarde PS. Bothrops Bilineatus Bilineatus (Two-Striped Forest Pitviper): Habitat Use. Herpetological Rev (2019) 50:385–5.
70. Eterovic A, Marques O, Endo W. Seasonal Activity of Snakes in the Atlantic Forest in Southeastern Brazil. Amphib Reptilia (2001) 22:103–11. doi: 10.1163/156853801750096213
71. Roriz KRPS, Zaqueo KD, Setubal SS, Katsuragawa TH, da Silva RR, Fernandes CFC, et al. Epidemiological Study of Snakebite Cases in Brazilian Western Amazonia. Rev Soc Bras Med Trop (2018) 51:338–46. doi: 10.1590/0037-8682-0489-2017
72. Mota-da-Silva A, Colombini M, Moura-da-Silva AM, de Souza RM, Monteiro WM, Bernarde PS. Ethno-Knowledge and Attitudes Regarding Snakebites in the Alto Juruá Region, Western Brazilian Amazonia. Toxicon (2019) 171:66–77. doi: 10.1016/j.toxicon.2019.10.238
73. de Oliveira LP, Moreira JGdoV, Sachett JdeAG, Monteiro WM, Meneguetti DU deO, Bernarde PS. Snakebites in Rio Branco and Surrounding Region, Acre, Western Brazilian Amazon. Rev Soc Bras Med Trop (2020) 53:e20200214. doi: 10.1590/0037-8682-0214-2020
74. Almeida-Santos SM, Salomão MG. Reproduction in Neotropical Pitvipers, With Emphasis on Species of the Genus Bothrops. In: Schuett GW, Höggren M, Douglas ME, Greene HW, editors. Biology of the Vipers. Carmel: Eagle Mountain Publishing (2002). p. 445–62.
75. Instituto Chico Mendes de Conservação da Biodiversidade. Livro Vermelho Da Fauna Brasileira Ameaçada De Extinção. Brasilia: ICMBio/MMA (2018).
76. International Union for Conservation of Nature and Natural Resources. The IUCN Red List of Threatened Species. Version 2019−2 (2019). Available at: http://www.iucnredlist.org (Accessed June 21, 2021).
77. Tozetti AM, Sawaya RJ, Molina FB, Bérnils RS, Barbo FE, Leite JCM, et al. Répteis. In: Conte CE, editor. Revisões Em Zoologia: Mata Atlântica. Curitiba: UFPR: Emygdio Leite de Araujo Monteiro Filho (2017). p. 309–58.
78. Bérnils RS, Castro TMC, Almeida AP, Argôlo AJS, Costa HC, Oliveira JCF, et al. Répteis Ameaçados De Extinção No Estado do Espírito Santo. In: Fraga CN, Formigoni MH, Chaves FG, editors. Fauna E Flora Ameaçadas De Extinção No Estado do Espírito Santo. Santa Teresa: Instituto Nacional da Mata Atlântica (2019). p. 271–93.
79. Feio RN, Caramaschi U. Contribuição Ao Conhecimento Da Herpetofauna do Nordeste do Estado De Minas Gerais, Brasil. Phyllomedusa (2002) 1:105–11. doi: 10.11606/issn.2316-9079.v1i2p105-111
80. Dias LG, Feio RN, Santos PS. New Record of Bothriopsis Bilineata (Wied, 1825) (Serpentes, Viperidae) in the Atlantic Forest of Minas Gerais, With a Discussion on Its Conservation. Lundiana: Int J Biodiversity (2008) 9:75–6. doi: 10.35699/2675-5327.2008.23218
81. Fry BG, Roelants K, Champagne DE, Scheib H, Tyndall JDA, King GF, et al. The Toxicogenomic Multiverse: Convergent Recruitment of Proteins Into Animal Venoms. Annu Rev Genom Hum Genet (2009) 10:483–511. doi: 10.1146/annurev.genom.9.081307.164356
82. Yu C, Yu H, Li P. Highlights of Animal Venom Research on the Geographical Variations of Toxin Components, Toxicities and Envenomation Therapy. Int J Biol Macromolecules (2020) 165:2994–3006. doi: 10.1016/j.ijbiomac.2020.10.190
83. Sanz L, Quesada-Bernat S, Pérez A, De Morais-Zani K, SantˈAnna SS, Hatakeyama DM, et al. Danger in the Canopy. Comparative Proteomics and Bioactivities of the Venoms of the South American Palm Pit Viper Bothrops Bilineatus Subspecies Bilineatus and Smaragdinus and Antivenomics of B. B. Bilineatus (Rondônia) Venom Against the Brazilian Pentabothropic Antivenom. J Proteome Res (2020) 19:3518–32. doi: 10.1021/acs.jproteome.0c00337
84. Rodrigues CR, Teixeira-Ferreira A, Vargas FFR, Guerra-Duarte C, Costal-Oliveira F, Stransky S, et al. Proteomic Profile, Biological Activities and Antigenic Analysis of the Venom From Bothriopsis Bilineata Smaragdina (“Loro Machaco”), a Pitviper Snake From Peru. J Proteomics (2018) 187:171–81. doi: 10.1016/j.jprot.2018.07.016
85. Fox JW, Serrano SMT. Insights Into and Speculations About Snake Venom Metalloproteinase (SVMP) Synthesis, Folding and Disulfide Bond Formation and Their Contribution to Venom Complexity. FEBS J (2008) 275:3016–30. doi: 10.1111/j.1742-4658.2008.06466.x
86. Sousa LF, Nicolau CA, Peixoto PS, Bernardoni JL, Oliveira SS, Portes-Junior JA, et al. Comparison of Phylogeny, Venom Composition and Neutralization by Antivenom in Diverse Species of Bothrops Complex. PloS Neglected Trop Dis (2013) 7:e2442. doi: 10.1371/journal.pntd.0002442
87. Moura-da-Silva AM, Butera D, Tanjoni I. Importance of Snake Venom Metalloproteinases in Cell Biology: Effects on Platelets, Inflammatory and Endothelial Cells. Curr Pharm Des (2007) 13:2893–905. doi: 10.2174/138161207782023711
88. Freitas-de-Sousa LA, Colombini M, Lopes-Ferreira M, Serrano SMT, Moura-da-Silva AM. Insights Into the Mechanisms Involved in Strong Hemorrhage and Dermonecrosis Induced by Atroxlysin-Ia, A PI-Class Snake Venom Metalloproteinase. Toxins (Basel) (2017) 9:239. doi: 10.3390/toxins9080239
89. Oliveira SS, Freitas-de-Sousa LA, Alves EC, de Lima Ferreira LC, da Silva IM, de Lacerda MVG, et al. Fatal Stroke After Bothrops Snakebite in the Amazonas State, Brazil: A Case Report. Toxicon (2017) 138:102–6. doi: 10.1016/j.toxicon.2017.08.021
90. Higuchi DA, Almeida MC, Barros CC, Sanchez EF, Pesquero PR, Lang EAS, et al. Leucurogin, a New Recombinant Disintegrin Cloned From Bothrops Leucurus (White-Tailed-Jararaca) With Potent Activity Upon Platelet Aggregation and Tumor Growth. Toxicon (2011) 58:123–9. doi: 10.1016/j.toxicon.2011.05.013
91. Kamiguti AS, Hay CR, Zuzel M. Inhibition of Collagen-Induced Platelet Aggregation as the Result of Cleavage of Alpha 2 Beta 1-Integrin by the Snake Venom Metalloproteinase Jararhagin. Biochem J (1996) 320:635–41. doi: 10.1042/bj3200635
92. Moura-da-Silva AM, Marcinkiewicz C, Marcinkiewicz M, Niewarowski S. Selective Recognition of Alpha 2 Beta 1 Integrin by Jararhagin, a Metalloproteinase-/Disintegrin From Bothrops Jararaca Venom. Thromb Res (2001) 102:153–9. doi: 10.1016/S0049-3848(01)00216-X
93. Bernardoni JL, Sousa LF, Wermelinger LS, Lopes AS, Prezoto BC, Serrano SMT, et al. Functional Variability of Snake Venom Metalloproteinases: Adaptive Advantages in Targeting Different Prey and Implications for Human Envenomation. PloS One (2014) 9:e109651. doi: 10.1371/journal.pone.0109651
94. Kini RM, Rao VS, Joseph JS. Procoagulant Proteins From Snake Venoms. Haemostasis (2001) 31:218–24. doi: 10.1159/000048066
95. Modesto JC de A, Junqueira-de-Azevedo ILM, Neves-Ferreira AGC, Fritzen M, Oliva MLV, Ho PL, et al. Insularinase A, A Prothrombin Activator From Bothrops Insularis Venom, Is a Metalloprotease Derived From a Gene Encoding Protease and Disintegrin Domains. Biol Chem (2005) 386:589–600. doi: 10.1515/BC.2005.069
96. de Almeida MT, Freitas-de-Sousa LA, Colombini M, Gimenes SNC, Kitano ES, Faquim-Mauro EL, et al. Inflammatory Reaction Induced by Two Metalloproteinases Isolated From Bothrops Atrox Venom and by Fragments Generated From the Hydrolysis of Basement Membrane Components. Toxins (2020) 12:96. doi: 10.3390/toxins12020096
97. Rucavado A, Nicolau CA, Escalante T, Kim J, Herrera C, Gutiérrez JM, et al. Viperid Envenomation Wound Exudate Contributes to Increased Vascular Permeability via a DAMPs/TLR-4 Mediated Pathway. Toxins (2016) 8:349. doi: 10.3390/toxins8120349
98. Serrano SMT. The Long Road of Research on Snake Venom Serine Proteinases. Toxicon (2013) 62:19–26. doi: 10.1016/j.toxicon.2012.09.003
99. Clemetson KJ, Lu Q, Clemetson JM. Snake Venom Proteins Affecting Platelets and Their Applications to Anti-Thrombotic Research. Curr Pharm Des (2007) 13:2887–92. doi: 10.2174/138161207782023702
100. de Oliveira SS, Alves EC, dos S Santos A, Nascimento EF, Tavares Pereira JP, Mendonça da Silva I, et al. Bothrops Snakebites in the Amazon: Recovery From Hemostatic Disorders After Brazilian Antivenom Therapy. Clin Toxicol (2020) 58:266–74. doi: 10.1080/15563650.2019.1634273
101. Assafim M, Frattani FS, Ferreira MS, Silva DM, Monteiro RQ, Zingali RB. Exploiting the Antithrombotic Effect of the (Pro)Thrombin Inhibitor Bothrojaracin. Toxicon (2016) 119:46–51. doi: 10.1016/j.toxicon.2016.05.007
102. Zingali RB, Ferreira MS, Assafim M, Frattani FS, Monteiro RQ. Bothrojaracin, A Bothrops Jararaca Snake Venom-Derived (Pro)Thrombin Inhibitor, as an Anti-Thrombotic Molecule. Pathophysiol Haemos Thromb (2005) 34:160–3. doi: 10.1159/000092416
103. Lomonte B, Angulo Y, Sasa M, Gutiérrez JM. The Phospholipase A2 Homologues of Snake Venoms: Biological Activities and Their Possible Adaptive Roles. Protein Pept Lett (2009) 16:860–76. doi: 10.2174/092986609788923356
104. Lomonte B, Rangel J. Snake Venom Lys49 Myotoxins: From Phospholipases A(2) to Non-Enzymatic Membrane Disruptors. Toxicon (2012) 60:520–30. doi: 10.1016/j.toxicon.2012.02.007
106. Torrez PPQ, Said R, Quiroga MMM, Duarte MR, França FOS. Forest Pit Viper (Bothriopsis Bilineata Bilineata) Bite in the Brazilian Amazon With Acute Kidney Injury and Persistent Thrombocytopenia. Toxicon (2014) 85:27–30. doi: 10.1016/j.toxicon.2014.04.001
107. Mamede CCN, de Sousa Simamoto BB, da Cunha Pereira DF, de Oliveira Costa J, Ribeiro MSM, de Oliveira F. Edema, Hyperalgesia and Myonecrosis Induced by Brazilian Bothropic Venoms: Overview of the Last Decade. Toxicon (2020) 187:10–8. doi: 10.1016/j.toxicon.2020.08.016
108. Laustsen AH, María Gutiérrez J, Knudsen C, Johansen KH, Bermúdez-Méndez E, Cerni FA, et al. Pros and Cons of Different Therapeutic Antibody Formats for Recombinant Antivenom Development. Toxicon (2018) 146:151–75. doi: 10.1016/j.toxicon.2018.03.004
109. Silva FS, Ibiapina HNS, Neves JCF, Coelho KF, Barbosa FBA, Lacerda MVG, et al. Severe Tissue Complications in Patients of Bothrops Snakebite at a Tertiary Health Unit in the Brazilian Amazon: Clinical Characteristics and Associated Factors. Rev Soc Bras Med Trop (2021) 54:e0374–2020. doi: 10.1590/0037-8682-0374-2020
110. Gutiérrez JM, Escalante T, Hernández R, Gastaldello S, Saravia-Otten P, Rucavado A. Why Is Skeletal Muscle Regeneration Impaired After Myonecrosis Induced by Viperid Snake Venoms? Toxins (Basel) (2018) 10:182. doi: 10.3390/toxins10050182
111. Cadillo EM, Ferreyra CB, Zvealeta A. Hemolytic Activity of Venoms From Snakes of the Genera Bothrop, Lachesis, Crotalus, and Micrurus (Serpentes: Viperidae and Elapidae). Rev Biol Trop (1991) 39:311–4.
112. Carregari VC, Stuani Floriano R, Rodrigues-Simioni L, Winck FV, Baldasso PA, Ponce-Soto LA, et al. Biochemical, Pharmacological, and Structural Characterization of New Basic PLA2 Bbil-TX From Bothriopsis Bilineata Snake Venom. BioMed Res Int (2013) 2013:1–12. doi: 10.1155/2013/612649
113. Floriano RS, Carregari VC, de Abreu VA, Kenzo-Kagawa B, Ponce-Soto LA, da Cruz-Höfling MA, et al. Pharmacological Study of a New Asp49 Phospholipase A2 (Bbil-TX) Isolated From Bothriopsis Bilineata Smargadina (Forest Viper) Venom in Vertebrate Neuromuscular Preparations. Toxicon (2013) 69:191–9. doi: 10.1016/j.toxicon.2013.03.006
114. Melaré R, Floriano RS, Gracia M, Rodrigues-Simioni L, da Cruz-Höfling MA, Rocha T. Ultrastructural Aspects of Mouse Nerve-Muscle Preparation Exposed to Bothrops Jararacussu and Bothrops Bilineatus Venoms and Their Toxins BthTX-I and Bbil-TX: Unknown Myotoxic Effects: Melaré Et al. Microsc Res Tech (2016) 79:1082–9. doi: 10.1002/jemt.22748
115. Rodrigues-Simioni L, Floriano RS, Rostelato-Ferreira S, Sousa NC, Marangoni S, Ponce-Soto LA, et al. Presynaptic Action of Bothriopsis Bilineata Smargadina (Forest Viper) Venom In Vitro. Toxicon (2011) 58:140–5. doi: 10.1016/j.toxicon.2011.05.002
116. Floriano RS, Rocha T, Carregari VC, Marangoni S, da Cruz-Höfling MA, Hyslop S, et al. The Neuromuscular Activity of Bothriopsis Bilineata Smaragdina (Forest Viper) Venom and Its Toxin Bbil-TX (Asp49 Phospholipase A2) on Isolated Mouse Nerve-Muscle Preparations. Toxicon (2015) 96:24–37. doi: 10.1016/j.toxicon.2015.01.001
117. Zoccal KF, Bitencourt C da S, Paula-Silva FWG, Sorgi CA, de Castro Figueiredo Bordon K, Arantes EC, et al. TLR2, TLR4 and CD14 Recognize Venom-Associated Molecular Patterns From Tityus Serrulatus to Induce Macrophage-Derived Inflammatory Mediators. PloS One (2014) 9:e88174. doi: 10.1371/journal.pone.0088174
118. Porto BN, Telli CA, Dutra TP, Alves LS, Bozza MT, Fin CA, et al. Biochemical and Biological Characterization of the Venoms of Bothriopsis Bilineata and Bothriopsis Taeniata (Serpentes: Viperidae). Toxicon (2007) 50:270–7. doi: 10.1016/j.toxicon.2007.03.020
119. Cardoso DF, Lopes-Ferreira M, Faquim-Mauro EL, Macedo MS, Farsky SHP. Role of Crotoxin, a Phospholipase A2 Isolated From Crotalus Durissus Terrificus Snake Venom, on Inflammatory and Immune Reactions. Mediators Inflamm (2001) 10:125–33. doi: 10.1080/09629350124986
120. Oliveira IS, Cardoso IA, Bordon KCF, Carone SEI, Boldrini-França J, Pucca MB, et al. Global Proteomic and Functional Analysis of Crotalus Durissus Collilineatus Individual Venom Variation and Its Impact on Envenoming. J Proteomics (2019) 191:153–65. doi: 10.1016/j.jprot.2018.02.020
121. Ribeiro CB, dos Santos JC, Silva JM, de Godoi PHS, Magalhães MR, Spadafora-Ferreira M, et al. Crotalus Durissus Collilineatus Venom Induces TNF- α and IL-10 Production in Human Peripheral Blood Mononuclear Cells. ISRN Inflammation (2014) 2014:1–7. doi: 10.1155/2014/563628
122. Setubal S da S, Pontes AS, Nery NM, Bastos JSF, Castro OB, Pires WL, et al. Effect of Bothrops Bilineata Snake Venom on Neutrophil Function. Toxicon (2013) 76:143–9. doi: 10.1016/j.toxicon.2013.09.019
123. Zuliani JP, Soares AM, Gutiérrez JM. Polymorphonuclear Neutrophil Leukocytes in Snakebite Envenoming. Toxicon (2020) 187:188–97. doi: 10.1016/j.toxicon.2020.09.006
124. Kini RM. Toxins in Thrombosis and Haemostasis: Potential Beyond Imagination: Toxins in Thrombosis and Haemostasis. J Thromb Haemostasis (2011) 9:195–208. doi: 10.1111/j.1538-7836.2011.04279.x
125. Berling I, Isbister GK. Hematologic Effects and Complications of Snake Envenoming. Transfus Med Rev (2015) 29:82–9. doi: 10.1016/j.tmrv.2014.09.005
126. Rodríguez E, Cahuana G, Sandoval GA, Yarlequé M, Yarlequé A. Evaluación Preliminar De La Actividad Coagulante Del Veneno De La Serpiente Peruana Bothrops Bilineatus “Loro Machaco.” Rev Soc Quím Perú (2010) 76:7.
127. Rodríguez E, Sandoval GA, Yarlequé A. Aislamiento Y Caracterización Bioquímica De La Bilinearina, Un Factor Activador De Protrombina Del Veneno De La Serpiente Peruana Bothrops Bilineatus (Loro Machaco). Rev Soc Quím Perú (2012) 78:3–13.
128. Silva JJ. Las Serpientes Del Género Bothrops En La Amazonia Colombiana. Acta Médica Colombiana (1989) 14:148–65.
129. Haad JS. Accidentes Humanos Por Las Serpientes De Los Géneros Bothrops Y Lachesis. Mem Inst Butantan (1981) 44/45:403–23.
130. Avila-Pires TCS, Vitt LJ, Sartorius SS, Zan PA. Squamata (Reptilia) From Four Sites in Southern Amazonia, With a Biogeographic Analysis of Amazonian Lizards. Boletim do Museu Paraense Emílio Goeldi - Ciências Naturais (2009) 4:99–118.
132. Wen FH, Monteiro WM, Moura da Silva AM, Tambourgi DV, Mendonça da Silva I, Sampaio VS, et al. Snakebites and Scorpion Stings in the Brazilian Amazon: Identifying Research Priorities for a Largely Neglected Problem. PloS Negl Trop Dis (2015) 9:e0003701. doi: 10.1371/journal.pntd.0003701
133. Muniz EG, Maria WS, Estevão-Costa MI, Buhrnheim P, Chávez-Olórtegui C. Neutralizing Potency of Horse Antibothropic Brazilian Antivenom Against Bothrops Snake Venoms From the Amazonian Rain Forest. Toxicon (2000) 38:1859–63. doi: 10.1016/S0041-0101(00)00082-9
134. Albuquerque PLMM, Jacinto CN, Silva Junior GB, Lima JB, Veras M doSB, Daher EF. Acute Kidney Injury Caused by Crotalus and Bothrops Snake Venom: A Review of Epidemiology, Clinical Manifestations and Treatment. . Rev Inst Med Trop S Paulo (2013) 55:295–301. doi: 10.1590/S0036-46652013000500001
135. Albuquerque PLMM, da Silva Junior GB, Meneses GC, Martins AMC, Lima DB, Raubenheimer J, et al. Acute Kidney Injury Induced by Bothrops Venom: Insights Into the Pathogenic Mechanisms. Toxins (Basel) (2019) 11:148. doi: 10.3390/toxins11030148
136. Otero-Patiño R. Epidemiological, Clinical and Therapeutic Aspects of Bothrops Asper Bites. Toxicon (2009) 54:998–1011. doi: 10.1016/j.toxicon.2009.07.001
137. Gras S, Plantefève G, Baud F, Chippaux J. Snakebite on the Hand: Lessons From Two Clinical Cases Illustrating Difficulties of Surgical Indication. J Venom Anim Toxins incl Trop Dis (2012) 18:467–77. doi: 10.1590/S1678-91992012000400019
Keywords: two-striped forest-pitviper, Amazon palm pitviper, snakebite, envenoming, venom, antivenom
Citation: Bernarde PS, Pucca MB, Mota-da-Silva A, da Fonseca WL, de Almeida MRN, de Oliveira IS, Cerni FA, Gobbi Grazziotin F, Sartim MA, Sachett J, Wen FH, Moura-da-Silva AM and Monteiro WM (2021) Bothrops bilineatus: An Arboreal Pitviper in the Amazon and Atlantic Forest. Front. Immunol. 12:778302. doi: 10.3389/fimmu.2021.778302
Received: 16 September 2021; Accepted: 25 November 2021;
Published: 15 December 2021.
Edited by:
Juliana Pavan Zuliani, Oswaldo Cruz Foundation (Fiocruz), BrazilReviewed by:
Bhargab Kalita, Amrita Vishwa Vidyapeetham (kochi campus), IndiaCopyright © 2021 Bernarde, Pucca, Mota-da-Silva, da Fonseca, de Almeida, de Oliveira, Cerni, Gobbi Grazziotin, Sartim, Sachett, Wen, Moura-da-Silva and Monteiro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paulo Sérgio Bernarde, U25ha2VCZXJuYXJkZUBob3RtYWlsLmNvbQ==; Wuelton M. Monteiro, d3VlbHRvbm1tQGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.