 Nina Gorlé
Nina Gorlé Eva Bauwens
Eva Bauwens Freddy Haesebrouck
Freddy Haesebrouck Annemieke Smet
Annemieke Smet Roosmarijn E. Vandenbroucke
Roosmarijn E. Vandenbroucke- 1VIB Center for Inflammation Research, Ghent, Belgium
- 2Department of Biomedical Molecular Biology, Faculty of Sciences, Ghent University, Ghent, Belgium
- 3Department of Pathology, Bacteriology and Avian Diseases, Faculty of Veterinary Medicine, Ghent University, Merelbeke, Belgium
- 4Laboratory of Experimental Medicine and Pediatrics, Faculty of Medicine and Health Sciences, University of Antwerp, Antwerp, Belgium
Trillions of symbiotic microbial cells colonize our body, of which the larger part is present in the human gut. These microbes play an essential role in our health and a shift in the microbiome is linked to several diseases. Recent studies also suggest a link between changes in gut microbiota and neurological disorders. Gut microbiota can communicate with the brain via several routes, together called the microbiome–gut–brain axis: the neuronal route, the endocrine route, the metabolic route and the immunological route. Helicobacter is a genus of Gram-negative bacteria colonizing the stomach, intestine and liver. Several papers show the role of H. pylori in the development and progression of neurological disorders, while hardly anything is known about other Helicobacter species and the brain. We recently reported a high prevalence of H. suis in patients with Parkinson’s disease and showed an effect of a gastric H. suis infection on the mouse brain homeostasis. Here, we discuss the potential role of H. suis in neurological disorders and how it may affect the brain via the microbiome–gut–brain axis.
Introduction
The human microbiota contains trillions of symbiotic microbial cells that live in and on our body of which the vast majority are present in the human gut (1–4). These commensal microbes perform several functions essential to our health and survival, including food digestion (5, 6), activation of certain drugs (4), prevention of infections (7–9), and they might play a role in the maturation of our immune system (10, 11).
Already for a few decades, changes in the gastrointestinal microbiota have been associated with a wide range of health problems including rheumatoid arthritis, inflammatory bowel diseases, asthma, and cancer, et cetera (12–17). Moreover, it has been shown that gastrointestinal changes are able to influence neurological disorders such as depression, anxiety, Alzheimer’s disease, Parkinson’s disease, and multiple sclerosis (MS) (18–29). Recently, it became clear that the gut microbiome can signal to the brain via several pathways, together called the microbiome–gut–brain axis (30–34). In general, communication between microbiota and the brain is divided into four categories: the neuronal route (enteric nervous system and vagus nerve), the endocrine route (e.g. cortisol), the metabolic route (e.g. short chain fatty acids (SCFAs) and tryptophan), and the immunological route (e.g. cytokines and immune cells) (35, 36). Bacteria can also affect the composition of the gut microbiota, thereby indirectly affecting gut-brain signaling [Cryan and Dinan (35)].
Helicobacter pylori and Neurological Disorders
A gastric spiral-shaped, Gram-negative microorganism, called H. pylori, colonizes the stomach of more than half of the world’s human population albeit with large geographical variations. Next to gastritis, peptic ulcer disease, mucosa-associated lymphoid tissue (MALT) -lymphoma, and adenocarcinoma, H. pylori infection has also been associated with neurological diseases.
Even though both innate and acquired immune responses are activated in individuals infected with H. pylori, the host is unable to eradicate the bacteria, leading to a chronic lifelong infection (37, 38). To escape the host’s immune response and to survive in the hostile conditions found in the stomach, H. pylori has developed several strategies, including manipulating innate immune receptors and inhibiting effector T-cell responses (39, 40). The mechanism to evade the immune system depends on the presence or absence of certain bacterial virulence factors (39). The evoked immune response by the host can lead to the local secretion of various inflammatory mediators, such as interleukin (IL) 8, -6, -1β, -10, and -12, tumor necrosis factor (TNF) and interferon (IFN) γ, which might reach the circulation causing a systemic effect (41, 42). The persistence of noticeable local and systemic concentrations of these pro-inflammatory factors can induce neuroinflammation and -toxicity (41). Next to this, H. pylori infection leads to the release of several neurotransmitters, such as acetylcholine, adrenaline, noradrenaline, serotonin, and dopamine (43, 44). Moreover, H. pylori infection might lead to axonal/neuronal damage, production of free radicals, and changes in neuropeptide expression, such as vasoactive intestinal peptide (VIP) and c-fos (43). Lastly, H. pylori infection is associated with changes in the composition of the gastrointestinal microbiome (43, 45). These changes, illustrated in Figure 1A, can potentially alter the outcome of neurological disorders.
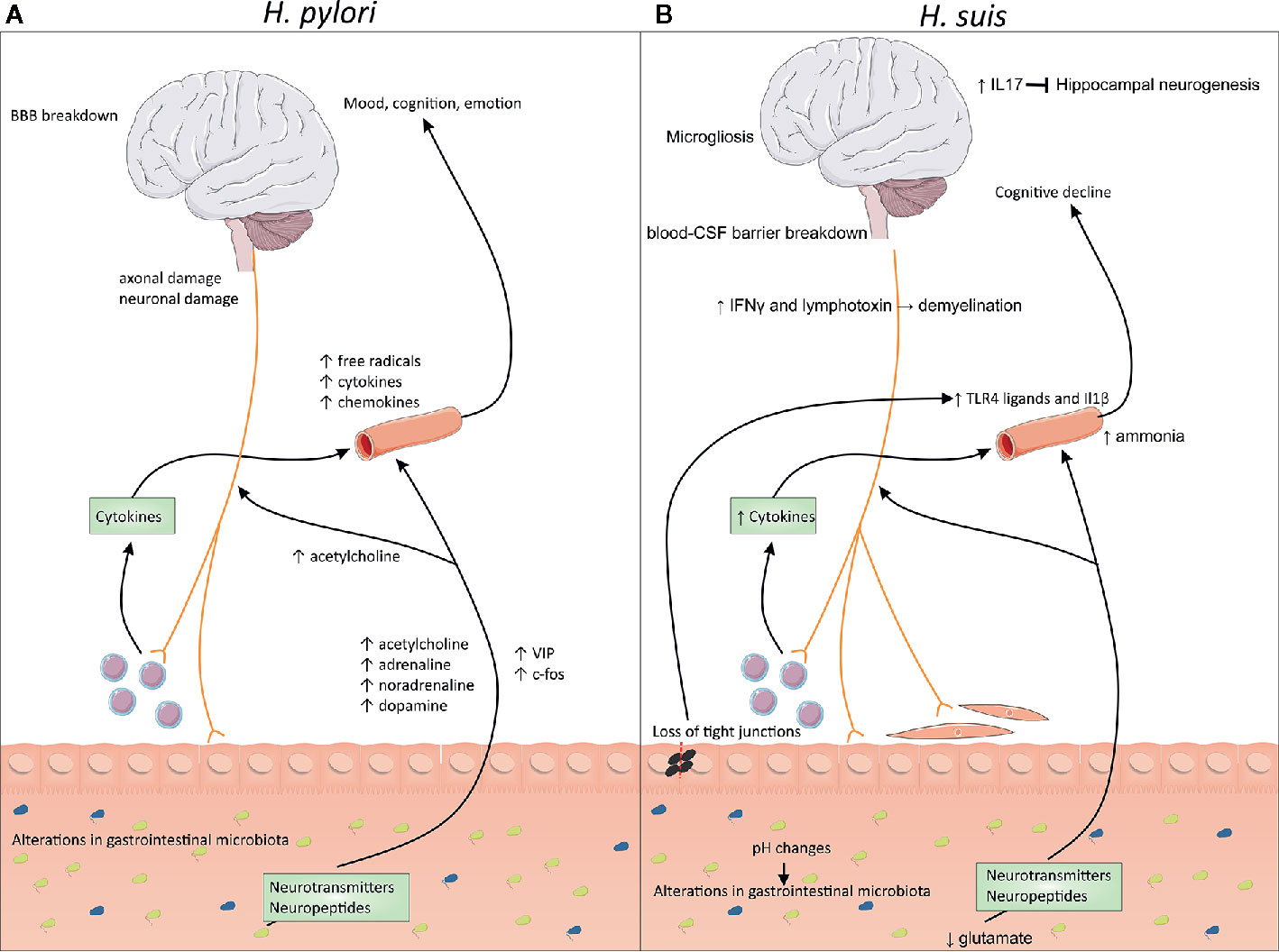
Figure 1 Changes at the microbiome–gut–brain axis during Helicobacter pylori and Non-H. pylori Helicobacter (NHPH) infection. (A) H. pylori is associated with persistent local inflammation, which might lead to systemic inflammation, characterized by increased levels of free radicals, cytokines and chemokines in the blood. Infection also leads to the release of several neurotransmitters, such as acetylcholine, adrenaline, noradrenaline and dopamine, but also increased levels of neuropeptides, such as vasoactive intestinal peptide (VIP) and c-fos. Moreover, H. pylori can lead to blood–brain barrier breakdown and axonal/neuronal damage. (B) H. suis is associated with inflammation of the stomach, associated with loss of the gastrointestinal barrier function, leading to leakage of TLR4 ligands into the blood. This leads to the breakdown of the blood-CSF barrier, combined with microgliosis and cognitive decline. H. suis-induced changes in the pH possibly leads to changes in the gastrointestinal microbiome. Moreover, lower levels of glutamate are present, which could influence the production of several neurotransmitters. Higher levels of IL-17 can block hippocampal neurogenesis while IFN-γ and lymphotoxins could lead to demyelination. BBB, blood–brain barrier; CSF, cerebrospinal fluid; IL, interleukin; IFN, interferon.
Indeed, seropositivity for H. pylori has been associated with poor cognition (46), neurologic impairment (47), and cerebrovascular disease (48) and is recognized as a significant risk factor for the development of dementia (21, 49). Next to an association of H. pylori with Parkinson’s disease (50), it has also been shown that infection with H. pylori increases the risk of developing Parkinson’s disease (41, 51, 52). Moreover, it has been shown that eradication of H. pylori improves the motor symptoms associated with Parkinson’s disease (53, 54). Interestingly, H. pylori might influence the bioavailability of L-3,4-dihydroxyphenylalaline (L-DOPA), the most common treatment for Parkinson’s disease (52, 55).
H. pylori might also play a role in Alzheimer’s disease as discussed in a review by Doulberis et al. (56). H. pylori infection is associated with mild cognitive impairment, a prodromal phase of Alzheimer’s disease (57, 58) and with Alzheimer’s disease itself (59). Higher levels of neuro-inflammation have been found in Alzheimer’s disease patients infected with H. pylori, which correlated with cognitive decline (60, 61), whereas eradication of H. pylori improved the cognitive and functional abilities (62, 63).
In multiple sclerosis (MS), however, H. pylori is found less in patients compared to control ones (64) and infection is even thought to be beneficial (65). Lower clinical signs were found in mice infected with H. pylori compared to control animals (66).
There is More in the Stomach Than Helicobacter pylori
Since the description of H. pylori, many other gastric species in the genus Helicobacter have been described. These gastric non-H. pylori Helicobacter (NHPH) species have been reported in the stomach of various hosts, including pigs, dogs, cats, and non-human primates and some of them have a zoonotic potential (67, 68). The most prevalent gastric NHPH species in humans is Helicobacter suis which naturally colonizes the stomach of pigs and non-human primates (67, 68). The bacterium is of zoonotic importance, infecting 0.2–6% of the human population, causing gastritis, peptic ulcers, and MALT lymphoma (67). However, since some infections with this microorganism remain subclinical, their true prevalence in humans is probably underestimated (67). Furthermore, these spiral-shaped bacteria are not always found in the human stomach after investigation of a small biopsy sample due to their focal and patchy colonization pattern (67, 69–71). Like H. pylori, H. suis may lead to a life-long infection, associated with a tolerogenic immune response (24, 72).
In literature, hardly any data is available on the association between an infection with NHPH species and neurological disorders. Indeed, there are no papers describing the association of NHPH with neurodegenerative or -immunological disorders like amyotrophic lateral sclerosis, spinocerebellar degeneration, acute disseminated encephalomyelitis, and Guillain-Barré syndrome. One study showed that mice infected with Helicobacter felis display both gastric and neuroinflammation (73). In another study, a remarkable high presence of H. suis DNA (27%) was found in gastric biopsies from idiopathic Parkinson’s disease patients compared to a control group without clinical symptoms of Parkinson’s disease (2%) (74). This was not the case for other zoonotically important gastric NHPH species. Additionally, H. suis DNA was found in a blood sample of a patient simultaneously affected by Parkinson’s and Alzheimer’s disease. After eradication of the H. suis infection, the patient’s gastric and neurological symptoms improved remarkably (74). Moreover, H. suis infection in Parkinson’s patients has recently been linked with higher mortality (75). To our knowledge, there are no other papers describing a role for H. suis in neurological disorders. Here, we will discuss several possible ways H. suis might influence the brain. These changes are summarized in Figure 1B.
Helicobacter suis and the Microbiome–Gut–Brain Axis
In the first part, inflammatory changes in the stomach and how they might affect the brain via the systemic circulation are discussed. In the second part, changes due to virulence factors of H. suis and the effect on the microbiome are discussed.
Inflammatory Changes and Gastrointestinal Barrier Functioning
Infection with H. suis in pigs and mice is associated with increased inflammation in the stomach, characterized by the higher expression of IL-8, -10, -1β, and -4, keratinocyte chemoattractant (KC), lipopolysaccharide-induced CXC chemokine (LIX), and macrophage inflammatory protein (MIP2) depending on the host (72, 76–78). This leads to the infiltration of B- and T-cells and macrophages in mice, inducing a Th2 response.
Gastritis is accompanied by mucosal edema (67) and gastric epithelial cell death (79), all of which could compromise the integrity of the gastrointestinal barrier. The gastrointestinal barrier consists of two layers: the epithelial cell layer, connected by tight junctions, and a mucus layer. In pigs, significant downregulation of claudin 18 (CLDN18) was found in the stomach of H. suis infected animals (72). In a recent mouse study, we found increased permeability of the gastrointestinal barrier after H. suis infection, accompanied by increased expression of mucine 13 (Muc13) and aberrant localization of zonula occludens 1 (ZO1) (77). This further progressed to systemic inflammation, characterized by the leakage of TLR4 ligands into the blood, affecting the brain homeostasis via the blood–cerebrospinal fluid barrier (77). Next to TLR4 ligands, also IL1β was found in the serum of H. suis-infected mice, which is shown to induce inflammatory gene expression in the hippocampus and hypothalamus associated with sickness behavior (80). As discussed below, also other molecules that are observed in the stomach upon H. suis infection might affect the brain when reaching the systemic circulation due to a leaky gut.
Next to the Th2 response, also a Th17 response has been associated with H. suis infection in the different hosts (mice, gerbils, pigs, and humans), characterized by the presence of Th17 cells and/or increased levels of IL-17 in the stomach (76, 78, 81, 82). IL-17 is known to block adult hippocampus neurogenesis (83) and is linked to depression in MS (84). In gerbils, but not mice, also increased levels of IFN-γ were found in the stomach of H. suis infected animals (81). IFN-γ is shown to be a regulator of the neural precursor pool in the non-inflamed brain (85) but is also linked with demyelination due to the reduced proliferation and viability of oligodendroglial cells (86, 87).
H. suis is also associated with increased levels of lymphotoxin (LT)-α and -β in the stomach of mice (88). These cytokines are not only involved in the generation of follicular dendritic cells (89), but also regulate neuronal and glial lineage differentiation (90). Lymphotoxins have been shown to play a role in MS, causing demyelination due to oligodendrocyte toxicity (91). Blocking lymphotoxin in experimental autoimmune encephalomyelitis (EAE), a mouse model of MS, reduces disease symptoms, which is accompanied with lower levels of the chemokine CXCL13 (92). This chemokine plays a role in the recruitment of B-cells and its expression is increased in the stomach after H. suis infection in both pigs, mice, and gerbils (72, 81), as are other chemokines such as C-X-C motif chemokine receptor (CXCR) 7, 15 and 4, C-C motif chemokine ligand (CCL) 19 and 21, and C-X-C motif chemokine ligand 12 (CXCL12) (88). In MS, higher levels of CXCL13 have been observed in B-cell aggregates in the inflamed meninges (92) and correlate with demyelination, neural cell loss, and rapid disease progression (93). Thus, higher levels of CXCL13 caused by a H. suis infection can potentially lead to accelerated disease progression.
Changes Due to Virulence Factors, Metabolism and Microbiome
H. suis affects the presence of glutamine and glutathione by its virulence factor γ-glutamyl transpeptidase (GGT), in this way damaging epithelial cells (81, 82, 94). Glutamine and glutathione are not only important for the health of gastrointestinal tissue (95), they are also precursors for the neurotransmitters glutamate, aspartate, and γ-amino butyric acid (GABA), which are important neurotransmitters. Depletion of glutamine, caused by H. suis infection, could thus lead to changes in these neurotransmitters, affecting gut–brain signaling.
Urea is converted by H. suis to ammonia by the presence of urease (96, 97). High levels of ammonia are linked to encephalopathy, associated with neuropsychiatric and neurological symptoms (98, 99). Although it is unlikely that an H. suis infection leads to high levels of ammonia, the continuous exposure of slightly higher levels could also interfere with normal brain functioning.
Parietal cells are also affected by H. suis-associated inflammation. This leads to changes in the expression and functioning of H+/K+-ATPase and subsequent changes in pH, which is associated with more fluid gastric content (72). These changes can subsequently influence the gastric microbiota. Indeed, more Fusobacterium gastrosuis was found in H. suis infected pigs (100). Infection with H. felis, another NHPH known to infect humans, is associated with a decrease in Lactobacillus and an increase in Clostridium, Bacteroidetes, Prevotella, Eubacterium, Ruminococcus, Streptococcus, and E. coli in the stomach (94, 101). Lactobacillus has been shown to secrete acetylcholine, which is important in regulating memory, attention, and learning, and has therapeutic effects in mental illnesses, reducing anxiety and depression (102). Lower numbers of Lactobacillus due to H. suis could thus possibly affect mood. Increased levels of Clostridium has been linked to autism (103), indicating that increased presence of Clostridium in H. suis-infected animals might affect brain homeostasis.
Conclusion
Numerous studies have been published about the possible effect of a H. pylori infection on neurological diseases, while other Helicobacter species have hardly been studied. However, recent studies report on a possible link between H. suis infection and Parkinson’s disease. Here, we describe several possible pathways in the microbiome–gut–brain axis which could be influenced by H. suis infection. Altogether, this highlights the importance of gaining more insights in the role of non-Helicobacter pylori Helicobacter species in neurological diseases.
Author Contributions
NG wrote the manuscript. EV, RV, AS, and FH advised and reviewed the manuscript. All authors contributed to the article and approved the submitted version.
Funding
NG is supported by FWO Vlaanderen. Research in the author’s lab is sponsored by FWO Vlaanderen, Ghent University, VIB, and the Baillet Latour Fund.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the Servier Medical Art for the use of Powerpoint images from their images bank.
References
1. Kverka M, Tlaskalova-Hogenova H. Intestinal Microbiota: Facts and Fiction. Dig Dis (2017) 35(1-2):139–47. doi: 10.1159/000449095
2. Sender R, Fuchs S, Milo R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol (2016) 14(8):e1002533. doi: 10.1371/journal.pbio.1002533
3. Ursell LK, Metcalf JL, Parfrey LW, Knight R. Defining the human microbiome. Nutr Rev (2012) 70 Suppl 1:S38–44. doi: 10.1111/j.1753-4887.2012.00493.x
4. Moos WH, Faller DV, Harpp DN, Kanara I, Pernokas J, Powers WR, et al. Microbiota and Neurological Disorders: A Gut Feeling. Biores Open Access (2016) 5(1):137–45. doi: 10.1089/biores.2016.0010
5. Bergman EN. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol Rev (1990) 70(2):567–90. doi: 10.1152/physrev.1990.70.2.567
6. Hill MJ. Intestinal flora and endogenous vitamin synthesis. Eur J Cancer Prev (1997) 6 Suppl 1:S43–5. doi: 10.1097/00008469-199703001-00009
7. Brandl K, Plitas G, Mihu CN, Ubeda C, Jia T, Fleisher M, et al. Vancomycin-resistant enterococci exploit antibiotic-induced innate immune deficits. Nature (2008) 455(7214):804–7. doi: 10.1038/nature07250
8. Royet J, Gupta D, Dziarski R. Peptidoglycan recognition proteins: modulators of the microbiome and inflammation. Nat Rev Immunol (2011) 11(12):837–51. doi: 10.1038/nri3089
9. Spitz JC, Ghandi S, Taveras M, Aoys E, Alverdy JC. Characteristics of the intestinal epithelial barrier during dietary manipulation and glucocorticoid stress. Crit Care Med (1996) 24(4):635–41. doi: 10.1097/00003246-199604000-00015
10. Cebra JJ. Influences of microbiota on intestinal immune system development. Am J Clin Nutr (1999) 69(5):1046S–51S. doi: 10.1093/ajcn/69.5.1046s
11. Cebra JJ, Logan AC, Weinstein PD. The preference for switching to expression of the IgA isotype of antibody exhibited by B lymphocytes in Peyer’s patches is likely due to intrinsic properties of their microenvironment. Immunol Res (1991) 10(3-4):393–5. doi: 10.1007/BF02919728
12. Wu X, He B, Liu J, Feng H, Ma Y, Li D, et al. Molecular Insight into Gut Microbiota and Rheumatoid Arthritis. Int J Mol Sci (2016) 17(3):431. doi: 10.3390/ijms17030431
13. Abraham C, Cho JH. Inflammatory bowel disease. N Engl J Med (2009) 361(21):2066–78. doi: 10.1056/NEJMra0804647
14. Halfvarson J, Brislawn CJ, Lamendella R, Vazquez-Baeza Y, Walters WA, Bramer LM, et al. Dynamics of the human gut microbiome in inflammatory bowel disease. Nat Microbiol (2017) 2:17004. doi: 10.1038/nmicrobiol.2017.4
15. Huang YJ, Boushey HA. The microbiome in asthma. J Allergy Clin Immunol (2015) 135(1):25–30. doi: 10.1016/j.jaci.2014.11.011
16. Hahn DL, Dodge RW, Golubjatnikov R. Association of Chlamydia pneumoniae (strain TWAR) infection with wheezing, asthmatic bronchitis, and adult-onset asthma. JAMA (1991) 266(2):225–30. doi: 10.1001/jama.266.2.225
17. Trinchieri G. Cancer and inflammation: an old intuition with rapidly evolving new concepts. Annu Rev Immunol (2012) 30:677–706. doi: 10.1146/annurev-immunol-020711-075008
18. Bercik P, Denou E, Collins J, Jackson W, Lu J, Jury J, et al. The intestinal microbiota affect central levels of brain-derived neurotropic factor and behavior in mice. Gastroenterology (2011) 141(2):599–609, e1-3. doi: 10.1053/j.gastro.2011.04.052
19. Bravo JA, Forsythe P, Chew MV, Escaravage E, Savignac HM, Dinan TG, et al. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc Natl Acad Sci USA (2011) 108(38):16050–5. doi: 10.1073/pnas.1102999108
20. Dinan TG, Cryan JF. Gut-brain axis in 2016: Brain-gut-microbiota axis - mood, metabolism and behaviour. Nat Rev Gastroenterol Hepatol (2017) 14(2):69–70. doi: 10.1038/nrgastro.2016.200
21. Hu X, Wang T, Jin F. Alzheimer’s disease and gut microbiota. Sci China Life Sci (2016) 59(10):1006–23. doi: 10.1007/s11427-016-5083-9
22. Mulak A, Bonaz B. Brain-gut-microbiota axis in Parkinson’s disease. World J Gastroenterol (2015) 21(37):10609–20. doi: 10.3748/wjg.v21.i37.10609
23. Parracho HM, Bingham MO, Gibson GR, McCartney AL. Differences between the gut microflora of children with autistic spectrum disorders and that of healthy children. J Med Microbiol (2005) 54(Pt 10):987–91. doi: 10.1099/jmm.0.46101-0
24. Zhang YJ, Li S, Gan RY, Zhou T, Xu DP, Li HB. Impacts of gut bacteria on human health and diseases. Int J Mol Sci (2015) 16(4):7493–519. doi: 10.3390/ijms16047493
25. Boulange CL, Neves AL, Chilloux J, Nicholson JK, Dumas ME. Impact of the gut microbiota on inflammation, obesity, and metabolic disease. Genome Med (2016) 8(1):42. doi: 10.1186/s13073-016-0303-2
26. Clemente JC, Ursell LK, Parfrey LW, Knight R. The impact of the gut microbiota on human health: an integrative view. Cell (2012) 148(6):1258–70. doi: 10.1016/j.cell.2012.01.035
27. Dinan TG, Cryan JF. The Microbiome-Gut-Brain Axis in Health and Disease. Gastroenterol Clin North Am (2017) 46(1):77–89. doi: 10.1016/j.gtc.2016.09.007
28. Walters WA, Xu Z, Knight R. Meta-analyses of human gut microbes associated with obesity and IBD. FEBS Lett (2014) 588(22):4223–33. doi: 10.1016/j.febslet.2014.09.039
29. Selber-Hnatiw S, Rukundo B, Ahmadi M, Akoubi H, Al-Bizri H, Aliu AF, et al. Human Gut Microbiota: Toward an Ecology of Disease. Front Microbiol (2017) 8:1265. doi: 10.3389/fmicb.2017.01265
30. Mayer EA, Knight R, Mazmanian SK, Cryan JF, Tillisch K. Gut microbes and the brain: paradigm shift in neuroscience. J Neurosci (2014) 34(46):15490–6. doi: 10.1523/JNEUROSCI.3299-14.2014
31. Martin CR, Osadchiy V, Kalani A, Mayer EA. The Brain-Gut-Microbiome Axis. Cell Mol Gastroenterol Hepatol (2018) 6(2):133–48. doi: 10.1016/j.jcmgh.2018.04.003
32. Galland L. The gut microbiome and the brain. J Med Food (2014) 17(12):1261–72. doi: 10.1089/jmf.2014.7000
33. Kelly JR, Minuto C, Cryan JF, Clarke G, Dinan TG. Cross Talk: The Microbiota and Neurodevelopmental Disorders. Front Neurosci (2017) 11:490:490. doi: 10.3389/fnins.2017.00490
34. Cryan JF, O’Riordan KJ, Cowan CSM, Sandhu KV, Bastiaanssen TFS, Boehme M, et al. The Microbiota-Gut-Brain Axis. Physiol Rev (2019) 99(4):1877–2013. doi: 10.1152/physrev.00018.2018
35. Cryan JF, Dinan TG. Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci (2012) 13(10):701–12. doi: 10.1038/nrn3346
36. Grenham S, Clarke G, Cryan JF, Dinan TG. Brain-gut-microbe communication in health and disease. Front Physiol (2011) 2:94. doi: 10.3389/fphys.2011.00094
37. Abadi ATB. Strategies used by helicobacter pylori to establish persistent infection. World J Gastroenterol (2017) 23(16):2870–82. doi: 10.3748/wjg.v23.i16.2870
38. Robinson K, Kaneko K, Andersen LP. Helicobacter: Inflammation, immunology and vaccines. Helicobacter (2017) 22 Suppl 1. doi: 10.1111/hel.12406
39. Mejias-Luque R, Gerhard M. Immune Evasion Strategies and Persistence of Helicobacter pylori. Curr Top Microbiol Immunol (2017) 400:53–71. doi: 10.1007/978-3-319-50520-6_3
40. Lundgren A, Suri-Payer E, Enarsson K, Svennerholm AM, Lundin BS. Helicobacter pylori-specific CD4+ CD25high regulatory T cells suppress memory T-cell responses to H. pylori in infected individuals. Infect Immun (2003) 71(4):1755–62. doi: 10.1128/IAI.71.4.1755-1762.2003
41. Alvarez-Arellano L, Maldonado-Bernal C. Helicobacter pylori and neurological diseases: Married by the laws of inflammation. World J Gastrointest Pathophysiol (2014) 5(4):400–4. doi: 10.4291/wjgp.v5.i4.400
42. Peek RM Jr., Fiske C, Wilson KT. Role of innate immunity in Helicobacter pylori-induced gastric malignancy. Physiol Rev (2010) 90(3):831–58. doi: 10.1152/physrev.00039.2009
43. Budzynski J, Klopocka M. Brain-gut axis in the pathogenesis of Helicobacter pylori infection. World J Gastroenterol (2014) 20(18):5212–25. doi: 10.3748/wjg.v20.i18.5212
44. Meng WP, Wang ZQ, Deng JQ, Liu Y, Deng MM, Lu MH. The Role of H. pylori CagA in Regulating Hormones of Functional Dyspepsia Patients. Gastroenterol Res Pract (2016) 2016:7150959. doi: 10.1155/2016/7150959
45. Engstrand L, Lindberg M. Helicobacter pylori and the gastric microbiota. Best Pract Res Clin Gastroenterol (2013) 27(1):39–45. doi: 10.1016/j.bpg.2013.03.016
46. Beydoun MA, Beydoun HA, Shroff MR, Kitner-Triolo MH, Zonderman AB. Helicobacter pylori seropositivity and cognitive performance among US adults: evidence from a large national survey. Psychosom Med (2013) 75(5):486–96. doi: 10.1097/PSY.0b013e31829108c3
47. Kimura A, Matsubasa T, Kinoshita H, Kuriya N, Yamashita Y, Fujisawa T, et al. Helicobacter pylori seropositivity in patients with severe neurologic impairment. Brain Dev (1999) 21(2):113–7. doi: 10.1016/S0387-7604(98)00086-2
48. Tsang KW, Lam SK. Helicobacter pylori and extra-digestive diseases. J Gastroenterol Hepatol (1999) 14(9):844–50. doi: 10.1046/j.1440-1746.1999.01974.x
49. Roubaud Baudron C, Letenneur L, Langlais A, Buissonniere A, Megraud F, Dartigues JF, et al. Does Helicobacter pylori infection increase incidence of dementia? The Personnes Agees QUID Study. J Am Geriatr Soc (2013) 61(1):74–8. doi: 10.1111/jgs.12065
50. Shen X, Yang H, Wu Y, Zhang D, Jiang H. Meta-analysis: Association of Helicobacter pylori infection with Parkinson’s diseases. Helicobacter (2017) 22(5). doi: 10.1111/hel.12398
51. Nielsen HH, Qiu J, Friis S, Wermuth L, Ritz B. Treatment for Helicobacter pylori infection and risk of Parkinson’s disease in Denmark. Eur J Neurol (2012) 19(6):864–9. doi: 10.1111/j.1468-1331.2011.03643.x
52. Mridula KR, Borgohain R, Chandrasekhar Reddy V, Bandaru V, Suryaprabha T. Association of Helicobacter pylori with Parkinson’s Disease. J Clin Neurol (2017) 13(2):181–6. doi: 10.3988/jcn.2017.13.2.181
53. Dobbs SM, Dobbs RJ, Weller C, Charlett A, Bjarnason IT, Lawson AJ, et al. Differential effect of Helicobacter pylori eradication on time-trends in brady/hypokinesia and rigidity in idiopathic parkinsonism. Helicobacter (2010) 15(4):279–94. doi: 10.1111/j.1523-5378.2010.00768.x
54. Bjarnason IT, Charlett A, Dobbs RJ, Dobbs SM, Ibrahim MA, Kerwin RW, et al. Role of chronic infection and inflammation in the gastrointestinal tract in the etiology and pathogenesis of idiopathic parkinsonism. Part 2: response of facets of clinical idiopathic parkinsonism to Helicobacter pylori eradication. A randomized, double-blind, placebo-controlled efficacy study. Helicobacter (2005) 10(4):276–87. doi: 10.1111/j.1523-5378.2005.00330.x
55. Hashim H, Azmin S, Razlan H, Yahya NW, Tan HJ, Manaf MR, et al. Eradication of Helicobacter pylori infection improves levodopa action, clinical symptoms and quality of life in patients with Parkinson’s disease. PLoS One (2014) 9(11):e112330. doi: 10.1371/journal.pone.0112330
56. Doulberis M, Kotronis G, Thomann R, Polyzos SA, Boziki M, Gialamprinou D, et al. Review: Impact of Helicobacter pylori on Alzheimer’s disease: What do we know so far? Helicobacter (2018) 23(1). doi: 10.1111/hel.12454
57. Kountouras J, Tsolaki M, Boziki M, Gavalas E, Zavos C, Stergiopoulos C, et al. Association between Helicobacter pylori infection and mild cognitive impairment. Eur J Neurol (2007) 14(9):976–82. doi: 10.1111/j.1468-1331.2007.01827.x
58. Han ML, Chen JH, Tsai MK, Liou JM, Chiou JM, Chiu MJ, et al. Association between Helicobacter pylori infection and cognitive impairment in the elderly. J Formos Med Assoc (2018) 117(11):994–1002. doi: 10.1016/j.jfma.2017.11.005
59. Kountouras J, Doulberis M, Polyzos SA, Katsinelos T, Vardaka E, Kountouras C, et al. Impact of Helicobacter pylori and/or Helicobacter pylori-related metabolic syndrome on incidence of all-cause and Alzheimer’s dementia. Alzheimers Dement (2019) 15(5):723–5. doi: 10.1016/j.jalz.2019.01.008
60. Kountouras J, Boziki M, Gavalas E, Zavos C, Deretzi G, Grigoriadis N, et al. Increased cerebrospinal fluid Helicobacter pylori antibody in Alzheimer’s disease. Int J Neurosci (2009) 119(6):765–77. doi: 10.1080/00207450902782083
61. Roubaud-Baudron C, Krolak-Salmon P, Quadrio I, Megraud F, Salles N. Impact of chronic Helicobacter pylori infection on Alzheimer’s disease: preliminary results. Neurobiol Aging (2012) 33(5):1009.e11–9. doi: 10.1016/j.neurobiolaging.2011.10.021
62. Kountouras J, Boziki M, Gavalas E, Zavos C, Deretzi G, Chatzigeorgiou S, et al. Five-year survival after Helicobacter pylori eradication in Alzheimer disease patients. Cognit Behav Neurol (2010) 23(3):199–204. doi: 10.1097/WNN.0b013e3181df3034
63. Kountouras J, Boziki M, Gavalas E, Zavos C, Grigoriadis N, Deretzi G, et al. Eradication of Helicobacter pylori may be beneficial in the management of Alzheimer’s disease. J Neurol (2009) 256(5):758–67. doi: 10.1007/s00415-009-5011-z
64. Li W, Minohara M, Su JJ, Matsuoka T, Osoegawa M, Ishizu T, et al. Helicobacter pylori infection is a potential protective factor against conventional multiple sclerosis in the Japanese population. J Neuroimmunol (2007) 184(1-2):227–31. doi: 10.1016/j.jneuroim.2006.12.010
65. Kira J. Helicobacter pylori infection might prove the hygiene hypothesis in multiple sclerosis. J Neurol Neurosurg Psychiatry (2015) 86(6):591–2. doi: 10.1136/jnnp-2014-309759
66. Cook KW, Crooks J, Hussain K, O’Brien K, Braitch M, Kareem H, et al. Helicobacter pylori infection reduces disease severity in an experimental model of multiple sclerosis. Front Microbiol (2015) 6:52. doi: 10.3389/fmicb.2015.00052
67. Haesebrouck F, Pasmans F, Flahou B, Chiers K, Baele M, Meyns T, et al. Gastric helicobacters in domestic animals and nonhuman primates and their significance for human health. Clin Microbiol Rev (2009) 22(2):202–23. doi: 10.1128/CMR.00041-08. Table of Contents.
68. Flahou B, Haesebrouck F, Smet A. Non-Helicobacter pylori Helicobacter Infections in Humans and Animals. In: Backert S, Yamaoka Y, editors. Helicobacter pylori Research: From Bench to Bedside. Tokyo: Springer Japan (2016). p. 233–69.
69. Yakoob J, Abbas Z, Khan R, Naz S, Ahmad Z, Islam M, et al. Prevalence of non Helicobacter pylori species in patients presenting with dyspepsia. BMC Gastroenterol (2012) 12:3. doi: 10.1186/1471-230X-12-3
70. Trebesius K, Adler K, Vieth M, Stolte M, Haas R. Specific detection and prevalence of Helicobacter heilmannii-like organisms in the human gastric mucosa by fluorescent in situ hybridization and partial 16S ribosomal DNA sequencing. J Clin Microbiol (2001) 39(4):1510–6. doi: 10.1128/JCM.39.4.1510-1516.2001
71. De Groote D, Van Doorn LJ, Van den Bulck K, Vandamme P, Vieth M, Stolte M, et al. Detection of non-pylori Helicobacter species in “Helicobacter heilmannii”-infected humans. Helicobacter (2005) 10(5):398–406. doi: 10.1111/j.1523-5378.2005.00347.x
72. De Witte C, Devriendt B, Flahou B, Bosschem I, Ducatelle R, Smet A, et al. Helicobacter suis induces changes in gastric inflammation and acid secretion markers in pigs of different ages. Vet Res (2017) 48(1):34. doi: 10.1186/s13567-017-0441-6
73. Albaret G, Sifre E, Floch P, Laye S, Aubert A, Dubus P, et al. Alzheimer’s Disease and Helicobacter pylori Infection: Inflammation from Stomach to Brain? J Alzheimers Dis (2020) 73(2):801–9. doi: 10.3233/JAD-190496
74. Blaecher C, Smet A, Flahou B, Pasmans F, Ducatelle R, Taylor D, et al. Significantly higher frequency of Helicobacter suis in patients with idiopathic parkinsonism than in control patients. Aliment Pharmacol Ther (2013) 38(11-12):1347–53. doi: 10.1111/apt.12520
75. Augustin AD, Savio A, Nevel A, Ellis RJ, Weller C, Taylor D, et al. Helicobacter suis Is Associated With Mortality in Parkinson’s Disease. Front Med (Lausanne) (2019) 6:188. doi: 10.3389/fmed.2019.00188
76. Bosschem I, Bayry J, De Bruyne E, Van Deun K, Smet A, Vercauteren G, et al. Effect of Different Adjuvants on Protection and Side-Effects Induced by Helicobacter suis Whole-Cell Lysate Vaccination. PLoS One (2015) 10(6):e0131364. doi: 10.1371/journal.pone.0131364
77. Gorle N, Blaecher C, Bauwens E, Vandendriessche C, Balusu S, Vandewalle J, et al. The choroid plexus epithelium as a novel player in the stomach-brain axis during Helicobacter infection. Brain Behav Immun (2017) 69:35–47. doi: 10.1016/j.bbi.2017.12.010
78. Flahou B, Deun KV, Pasmans F, Smet A, Volf J, Rychlik I, et al. The local immune response of mice after Helicobacter suis infection: strain differences and distinction with Helicobacter pylori. Vet Res (2012) 43:75. doi: 10.1186/1297-9716-43-75
79. Flahou B, Haesebrouck F, Chiers K, Van Deun K, De Smet L, Devreese B, et al. Gastric epithelial cell death caused by Helicobacter suis and Helicobacter pylori gamma-glutamyl transpeptidase is mainly glutathione degradation-dependent. Cell Microbiol (2011) 13(12):1933–55. doi: 10.1111/j.1462-5822.2011.01682.x
80. Skelly DT, Hennessy E, Dansereau MA, Cunningham C. A systematic analysis of the peripheral and CNS effects of systemic LPS, IL-1beta, [corrected] TNF-alpha and IL-6 challenges in C57BL/6 mice. PLoS One (2013) 8(7):e69123. doi: 10.1371/journal.pone.0069123
81. Zhang G, Ducatelle R, De Bruyne E, Joosten M, Bosschem I, Smet A, et al. Role of gamma-glutamyltranspeptidase in the pathogenesis of Helicobacter suis and Helicobacter pylori infections. Vet Res (2015) 46:31. doi: 10.1186/s13567-015-0163-6
82. Vermoote M, Van Steendam K, Flahou B, Smet A, Pasmans F, Glibert P, et al. Immunization with the immunodominant Helicobacter suis urease subunit B induces partial protection against H. suis infection in a mouse model. Vet Res (2012) 43:72. doi: 10.1186/1297-9716-43-72
83. Liu Q, Xin W, He P, Turner D, Yin J, Gan Y, et al. Interleukin-17 inhibits adult hippocampal neurogenesis. Sci Rep (2014) 4:7554. doi: 10.1038/srep07554
84. Waisman A, Hauptmann J, Regen T. The role of IL-17 in CNS diseases. Acta Neuropathol (2015) 129(5):625–37. doi: 10.1007/s00401-015-1402-7
85. Li L, Walker TL, Zhang Y, Mackay EW, Bartlett PF. Endogenous interferon gamma directly regulates neural precursors in the non-inflammatory brain. J Neurosci (2010) 30(27):9038–50. doi: 10.1523/JNEUROSCI.5691-09.2010
86. Hansen-Pupp I, Harling S, Berg AC, Cilio C, Hellstrom-Westas L, Ley D. Circulating interferon-gamma and white matter brain damage in preterm infants. Pediatr Res (2005) 58(5):946–52. doi: 10.1203/01.PDR.0000182592.76702.E8
87. Baerwald KD, Popko B. Developing and mature oligodendrocytes respond differently to the immune cytokine interferon-gamma. J Neurosci Res (1998) 52(2):230–9. doi: 10.1002/(SICI)1097-4547(19980415)52:2<230::AID-JNR11>3.0.CO;2-B
88. Zhao WJ, Tian ZB, Yao SS, Yu YN, Zhang CP, Li XY, et al. High-fat-diet-induced obesity upregulates the expression of lymphoid chemokines and promotes the formation of gastric lymphoid follicles after Helicobacter suis infection. Pathog Dis (2017) 75(8). doi: 10.1093/femspd/ftx101
89. Oldstone MB, Race R, Thomas D, Lewicki H, Homann D, Smelt S, et al. Lymphotoxin-alpha- and lymphotoxin-beta-deficient mice differ in susceptibility to scrapie: evidence against dendritic cell involvement in neuroinvasion. J Virol (2002) 76(9):4357–63. doi: 10.1128/JVI.76.9.4357-4363.2002
90. Xiao X, Putatunda R, Zhang Y, Soni PV, Li F, Zhang T, et al. Lymphotoxin beta receptor-mediated NFkappaB signaling promotes glial lineage differentiation and inhibits neuronal lineage differentiation in mouse brain neural stem/progenitor cells. J Neuroinflamm (2018) 15(1):49. doi: 10.1186/s12974-018-1074-z
91. Lock C, Oksenberg J, Steinman L. The role of TNFalpha and lymphotoxin in demyelinating disease. Ann Rheum Dis (1999) 58 Suppl 1:I121–8. doi: 10.1136/ard.58.2008.i121
92. Huber AK, Irani DN. Targeting CXCL13 During Neuroinflammation. Adv Neuroimmune Biol (2015) 6(1):1–8. doi: 10.3233/NIB-150101
93. Haugen M, Frederiksen JL, Degn M. B cell follicle-like structures in multiple sclerosis-with focus on the role of B cell activating factor. J Neuroimmunol (2014) 273(1-2):1–7. doi: 10.1016/j.jneuroim.2014.05.010
94. De Witte C, Taminiau B, Flahou B, Hautekiet V, Daube G, Ducatelle R, et al. In-feed bambermycin medication induces anti-inflammatory effects and prevents parietal cell loss without influencing Helicobacter suis colonization in the stomach of mice. Vet Res (2018) 49(1):35. doi: 10.1186/s13567-018-0530-1
95. Shibayama K, Kamachi K, Nagata N, Yagi T, Nada T, Doi Y, et al. A novel apoptosis-inducing protein from Helicobacter pylori. Mol Microbiol (2003) 47(2):443–51. doi: 10.1046/j.1365-2958.2003.03305.x
96. Burne RA, Chen YY. Bacterial ureases in infectious diseases. Microbes Infect (2000) 2(5):533–42. doi: 10.1016/S1286-4579(00)00312-9
97. Eaton KA, Brooks CL, Morgan DR, Krakowka S. Essential role of urease in pathogenesis of gastritis induced by Helicobacter pylori in gnotobiotic piglets. Infect Immun (1991) 59(7):2470–5. doi: 10.1128/IAI.59.7.2470-2475.1991
98. Braissant O, McLin VA, Cudalbu C. Ammonia toxicity to the brain. J Inherit Metab Dis (2013) 36(4):595–612. doi: 10.1007/s10545-012-9546-2
99. Bosoi CR, Rose CF. Identifying the direct effects of ammonia on the brain. Metab Brain Dis (2009) 24(1):95–102. doi: 10.1007/s11011-008-9112-7
100. De Witte C, Flahou B, Ducatelle R, Smet A, De Bruyne E, Cnockaert M, et al. Detection, isolation and characterization of Fusobacterium gastrosuis sp. nov. colonizing the stomach of pigs. Syst Appl Microbiol (2017) 40(1):42–50. doi: 10.1016/j.syapm.2016.10.001
101. Schmitz JM, Durham CG, Schoeb TR, Soltau TD, Wolf KJ, Tanner SM, et al. Helicobacter felis–associated gastric disease in microbiota-restricted mice. J Histochem Cytochem (2011) 59(9):826–41. doi: 10.1369/0022155411416242
102. Liu L, Zhu G. Gut-Brain Axis and Mood Disorder. Front Psychiatry (2018) 9:223:223. doi: 10.3389/fpsyt.2018.00223
Keywords: Helicobacter pylori, Helicobacter suis, microbiome–gut–brain axis, gut microbiota, neurological disorders
Citation: Gorlé N, Bauwens E, Haesebrouck F, Smet A and Vandenbroucke RE (2021) Helicobacter and the Potential Role in Neurological Disorders: There Is More Than Helicobacter pylori. Front. Immunol. 11:584165. doi: 10.3389/fimmu.2020.584165
Received: 16 July 2020; Accepted: 04 December 2020;
Published: 28 January 2021.
Edited by:
Francesca Ronchi, University of Bern, SwitzerlandReviewed by:
Jorge Tolivia, University of Oviedo, SpainYoshiro Ohara, Kanazawa Medical University, Japan
Copyright © 2021 Gorlé, Bauwens, Haesebrouck, Smet and Vandenbroucke. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Roosmarijn E. Vandenbroucke, Um9vc21hcmlqbi5WYW5kZW5icm91Y2tlQGlyYy5WSUItVUdlbnQuYmU=
†These authors have contributed equally to this work and share senior authorship