
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Hortic., 11 January 2024
Sec. Floriculture and Landscapes
Volume 2 - 2023 | https://doi.org/10.3389/fhort.2023.1248785
This article is part of the Research TopicEditors' Showcase: Floriculture and LandscapesView all 3 articles
 Francesca Bretzel1,2*
Francesca Bretzel1,2* Francesca Vannucchi1,2
Francesca Vannucchi1,2 Beatrice Pezzarossa1
Beatrice Pezzarossa1 Angeliki Paraskevopoulou3
Angeliki Paraskevopoulou3 Daniela Romano4
Daniela Romano4In anthropized sites, the establishment and management of sustainable and nature-based wildflower meadow solutions exclude fertilization, weeding, and watering, and reduce the frequency of mowing. Once such inputs have been reduced, the ecosystem services generated by the vegetation result in an increase in biodiversity in terms of plant and animal species, an improvement in soil quality and biota, the mitigation of air pollutants and carbon sequestration, as well as a reduction in soil temperature. Environmental education and citizen science are also some of the benefits of this type of vegetation, especially in urban areas. Herbaceous meadow-like communities, which are composed of forbs only or a combination of graminoid and forb species, are usually established by sowing. The seeds are available for purchase from specialized seed producers as single species or in a mix, some companies offer mixes composed of garden species with botanical species. Donor meadows are the sources of the genetic material and the template to reproduce the meadow herbaceous community at different sites through seed harvest, hay harvest, and the displacement of soil with its seed bank. Management techniques, such as soil preparation, determining the appropriate sowing and mowing time, can help in establishing and maintaining the most desirable species. The suspension of urban turf mowing allows the growth of the natural seed bank, which in unproductive soils facilitates the appearance of many pollinator-friendly herbaceous species. With the appropriate pedological conditions and management, the natural urban herbaceous vegetation thus provides beautiful and rich plant communities, which are also easy to maintain. This review identifies the best practices through a qualitative selection of case studies related to the various methods to establish wildflower meadows in the urban infrastructure and the main constraints such as the lack of a suitable seed market.
Fifty-five percent of the world’s population now lives in cities (UN, 2019), which is a major cause of habitat loss and fragmentation, and the challenge is to identify new strategies and appropriate management systems to conserve biodiversity in this increasingly urban environment (Fischer et al., 2018; Elmqvist et al., 2019; Colding et al., 2020; Kueffer, 2020). However, cities can be considered as biodiversity hotspots, due to the presence of endemic and threatened species (Derby Lewis et al., 2019), and urban nature can help citizens to get involved with environmental issues and connect to nature (Kueffer, 2020). The biodiversity value of the different types of urban vegetation mainly depends on plant size and connectivity (Liu et al., 2019), soil management (Lerman et al., 2018), and the socioeconomic characteristics of the neighbourhood (Leong et al., 2018). There is a growing interest in understanding how these factors interact and what mechanisms influence urban vegetation diversity (Fratarcangeli et al., 2019).
Given the concerns regarding the losses in global biodiversity and recognizing the importance of ecosystem services, there is increasing political will for cities to enhance and expand their green infrastructure (Andersson et al., 2014; Aronson et al., 2017; Lepczyk et al., 2017; Fischer et al., 2018). Cities are mosaics of different habitats, which also contribute to the relatively high urban biodiversity (Zhou et al., 2018). Urban green spaces can therefore take many forms: woodlands, brownfields and ruderal sites, lawns, ornamental plantings, and meadows.
Wildflower meadows include a mix of flowering dicotyledons and monocotyledons, and in some cases, grasses, generally established by sowing. They are managed with low cultivation inputs, i.e., mowing once annually, and without irrigation or fertilization, so flowering has a seasonal duration, determined by the climatic conditions of the site and the coexistence of species (Figure 1).

Figure 1 The flow chart reports the main steps to be followed for the establishment of a biodiverse wildflower meadows.
The green spaces devoted to wildflower meadows can contribute significantly to species diversity, despite the high fragmentation of urban environments (Atkins, 2018; Vega and Küffer, 2021; Griffiths-Lee et al., 2022). Converting a lawn into a wildflower-rich vegetation, in some cases, can be obtained simply by reducing the cultivation intensity, such as agrochemicals and mowing (Bertoncini et al., 2012). Alternatively, scalping the turf and sowing wildflower mixes, provides great environmental advantages, in terms of biodiversity, and at the same time satisfying people’s sense of the beauty of nature (Norton et al., 2019).
Wildflower meadows involve different ecosystem services, such as carbon storage, biotic nitrogen retention, nutrient recycling, biodiversity increase in flora and fauna, and the creation of pollinator habitats (Wang et al., 2018; Vega and Küffer, 2021). Planting wildflower meadows creates visual continuity with the landscape of the surrounding countryside. and ecological connectivity by constituting corridors and stepping stones for arthropods, small fauna and birds (Marshall et al., 2023). The low-input management respects the nature and functionality of the soil, and the reduction in mowing allows vegetation to grow tall and capture pollutants (Przybysz et al., 2021). At the social level, urban wildflower meadows enable citizens to get closer to biodiversity and to be involved in institutional or grassroots educational and citizen science programs, thus boosting their awareness of environmental issues (Steiner, 2023).
Intensive agriculture represents a threat to biodiversity. On the other hand, extensive cultivation allows the coexistence of crops with many spontaneous entomogamous species, that provide insects with nectar and pollen in exchange for pollination, and a habitat for many other organisms (Haaland et al., 2011; Benvenuti and Bretzel, 2017). Semi-natural herbaceous communities, rich in plant diversity, represent a model habitat, which can be reproduced in anthropogenic sites, mimicking the ecological dynamics of nature.
Native seed markets where the germplasm for establishing wildflower meadows can be purchased, have not yet been fully developed, however the new regulations in many sectors such as nature conservation, and agriculture politics mean that such markets are urgently needed (Gibson-Roy et al., 2021).
The aim of the present study is to identify the crucial elements in the establishment of wildflower meadows i.e. the choice of the species and the associated phytocoenosis, the origin of the propagation materials, and the availability in the market. An analysis of the literature was thus carried out, selecting the papers dealing with applicative methods related to seed production, meadow establishment techniques, and species employed. The research articles were selected through Scopus and the Web of Science, using the following keywords: “wildflower meadows”, “wildflower management”, “wildflower seed production and quality”, “wildflower planting”, “wildflower and agroecology”, and “wildflower and people”. The selection was also based on the authors’ scientific background and expertise. Papers published in the last ten years were considered as well as less recent papers that are still valid in terms of practice. The literature search did not focus on a specific geographical area, but primarily selected studies dealing with the establishment of wildflower meadows in dry and hot lands. This practice has been developed mainly in Northern Europe and North America, and thus these geographical areas predominate the list of cited papers. This review therefore presents a qualitatively selected collection of the methods, with a discussion of these, rather than a quantitative systematic review.
Urban planning structure and architecture vary across the world and have been influenced by globalisation, which in several cases has led to the homogenization of urban environments including the suppression of local indigenous plant communities (Ignatieva et al., 2020). Biodiversity, and particularly indigenous species, play a pivotal role in protecting the local ecological and cultural identity, as well as in creating a ‘sense of place’ (Ignatieva et al., 2020). The Green Infrastructure Strategy developed by the European Commission also considers green infrastructures as a “tool for providing ecological, economic and social benefits through natural solutions” (EC, 2013).
Biodiversity and ecosystem protection are essential elements of the green infrastructure aimed at sustainable urban development (UN Habitat, 2020). Carefully designed planting compositions with herbaceous species increase biodiversity and help reduce water use and promote sustainable agriculture and human wellbeing in line with the UN Sustainable Development Goals (Table 1).
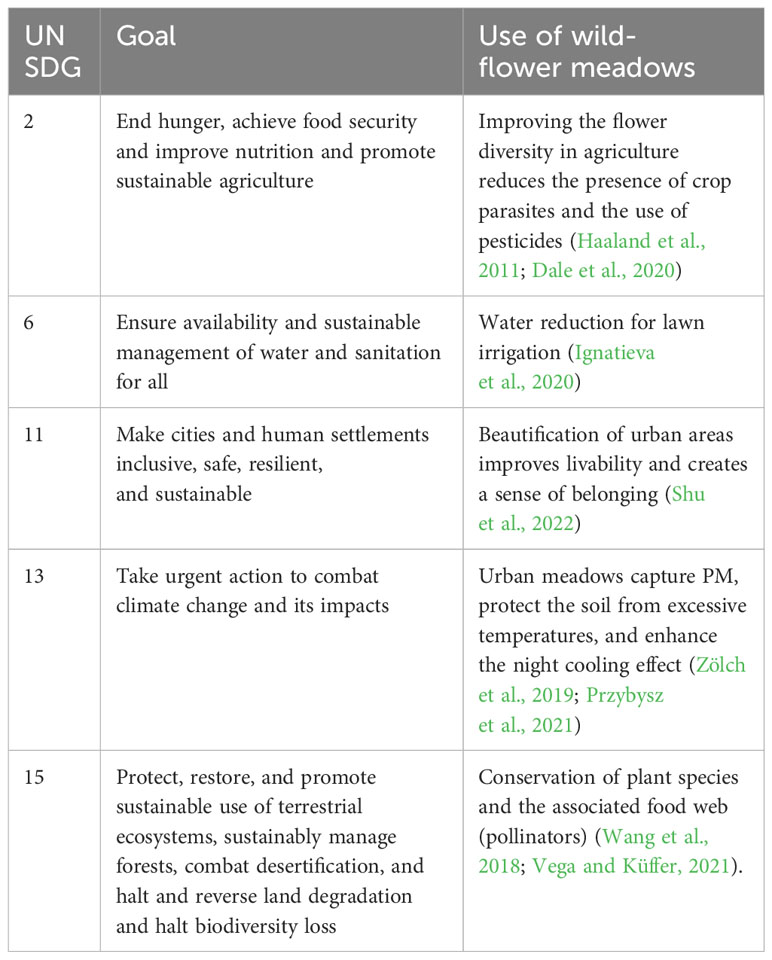
Table 1 The Sustainable Development Goals associated with wildflower meadows.
Herbaceous plant species, such as perennials, annuals, geophytes, and grasses, are often used in planting schemes by renowned urban landscape designers such as Gilles Clément, Nigel Dunnet, James Hitchmough, Noel Kingsbury, and Piet Oudolf, thus influencing the design of planting schemes across the world. These plants tend to be relatively small and occupy less space than woody plant species, therefore increasing the number of species per square meter. Also planting compositions with herbaceous species reflects seasonal changes and often provides colourful plant displays. The compositions of wildflower meadows can increase and conserve biodiversity in cities, attracting other plant species and animals: pollinators, birds, and small animals (Dale et al., 2020) (Figure 2).

Figure 2 Wildflower meadows support the food web, attracting pollinators, birds, and small animals.
The variety of flowers in a meadow attracts different insect species (Grass et al., 2016), and depending on the surrounding rural areas and season, green belts can create green corridors and stepping stones, to enhance the trophic network. Dale et al. (2020) highlighted that wildflower meadows in the vicinity of green sports areas, such as golf courses, can attract beneficial arthropods that control phytophagous pests.
An important aim in urban planning and design is to provide high-quality outdoor green spaces that meet the needs of various groups of people (Figure 3). Meadows also provide children with opportunities for play, experiential learning, exploration, socialisation, interaction with nature, and the development of aesthetic and restorative experiences, thus raising awareness of the various ecosystem services provided by green spaces (Shu et al., 2022).

Figure 3 Milan municipality planted a wildflower meadow of about 1 ha near to the “Bosco Verticale”, in the very populated neighbourhood of Isola. The residents spend their Sundays relaxing in the park, enjoying the colourful landscape (Photo F. Bretzel).
Given that urban areas with a high population and density of buildings are subjected to higher temperatures and air pollution, open meadow areas can provide unobstructed wind flow and reduce heat storage, thus enhancing the night cooling (Zölch et al., 2019).
Wildflower meadows can also be used as a sustainable urban drainage system (SuDS) to help tackle stormwater and flooding within cities caused by impervious surfaces, thus reducing overflows to the urban drainage systems (Meenar et al., 2022).
In cities, green roofs are also suitable for developing wildflower meadows and grasslands, provided that appropriate plant communities are used to meet the adverse environmental conditions (Benvenuti, 2014; Catalano et al., 2016; Vannucchi et al., 2018). However, in some cases the public are unwilling to have wildflower meadows in their urban neighbourhoods, thus highlighting the need to raise awareness through environmental education (Meenar et al., 2022). The combination of a wildflower meadow framed by mown-grass all around strengthens the impression that the wildflower meadow has been purposely designed and managed (Meenar et al., 2022). Patches of diverse wildflower meadows within the urban green infrastructure, such as in road verges and wastelands (Figure 4), can significantly improve wildlife and increase human wellbeing (Barry and Hodge, 2023) by encouraging people to directly immerse themselves in the beauty of the colours and shapes of flowers (Steiner, 2023).

Figure 4 The pedological conditions of many urban degraded sites such as roadsides are suitable for wildflower meadows to thrive. The figure shows a roundabout in Livorno (Italy) with a sown-on-purpose mix of wildflowers (Photo F. Bretzel).
Wildflower meadows can also help reconcile conservation needs with the demands of the local population for aesthetically pleasing spaces. These spaces can be used for wider community participation, socializing, recreation, and education. Sometimes the presence of exotic plant species, with a long-lasting, high visual impact, increases the benefits to human wellbeing (Vega et al., 2021), and non-native plant species often offer further food resources for pollinators (Kovács-Hostyánszki et al., 2022). Consequently, the presence of non-native plant species in a commercial wildflower mix in the right context (e.g., urban pollinator habitat) is acceptable in cases where a local product cannot be found on the market (Barry and Hodge, 2023). Only a few studies have evaluated the use of non-native wildflower species for urban parks and gardens. One example for ornamental and demonstrative purposes is the London Olympic Park (Hitchmough, 2016) where meadows from different geographical areas of the world were reproduced. In a study conducted in four UK cities native and exotic species were compared to verify the pollinator attractiveness through pollen production (Hicks et al., 2016; Mody et al., 2020).
Urban green areas can help to restore the biodiversity that intensive agriculture has reduced in the agroecosystems, leading to the loss of wildflowers and many associated pollinator species, such as bees and butterflies (Albrecht, 2003; Baessler and Klotz, 2006). Weed killers and the industrial cleaning of crop seeds have reduced the presence of flowering forbs next to cultivated fields (Spahillari et al., 1999) and have led to the loss of one of the most important ecosystem services for agriculture: pollination (Grab et al., 2019). Research has been aimed at increasing agroecology biodiversity, introducing wildflower meadows, and buffer flowered strips to combat the loss of entomofauna (Haaland et al., 2011). Perennial wildflowers help in controlling crop pests, and at the same time increase crop yield (Tschumi et al., 2016). Self-seeding is established in two-four meters wide strips of wildflowers that function as a habitat and food for many pollinators (Fenster et al., 2004). Studies conducted in vineyards with naturally grown wildflowers in the intercrop space compared the cut with the none-cut intercrop space (Figure 5), and the results highlighted the additional advantage of tall herbaceous species in protecting the grapes during extreme summer temperatures (Paraskevopoulou et al., 2022). Managing wildflower strips can be challenging, as mowing may take place exactly when there is a high presence of pollinators. Moreover, interventions are needed twice a year to improve the presence of flowers and control the weeds (Piqueray et al., 2019). However, a recent Directive of the EU Common Agricultural Policy (CAP) (EU Commission, 2021) promotes the establishment of meadows aimed at the conservation of pollinators, in accordance with the list of species that each country needs to provide, and imposes non-mowing during the active period of pollinators (spring to autumn). Finding sufficiently large amounts of seeds on the seed-market to cover farmlands, especially for organic farms that need organically-produced seeds, is still critical.

Figure 5 The intercrop spaces of ‘Sultanina’ vineyards in Perigiali, Corinth, (Greece) where the spontaneous wildflowers were not cut during spring providing shade from the extremely strong effect of the sun on the grapes (Photo E. Pappous).
As stated by the Plant Conservation Alliance (PCA) (2021) in the last report, a fundamental strategy is to get ‘the right seed in the right place at the right time’. Plant species selection is important for the establishment of reliable wildflower meadows. The effectiveness of wildflower meadows in increasing biodiversity is related to the composition of the seed mix and its adaptation to the local conditions, which depends on the number of species, flower abundance, plant species diversity, and vegetation structure (Haaland and Gyllin, 2011). Using a wide range of species in urban wildflower meadows with different life forms and survival strategies favors biodiversity and improves the establishment capacities of the plant community (Bretzel et al., 2016; Schröder and Kiehl, 2020).
The Supplementary Material reports some examples of mixes developed in different research studies along with technical details (Supplementary Table 1). However, the best approach is to choose species that grow in the wild in spontaneous associations, as they have adapted to coexist. Native species are recommended because they have adapted to specific climatic and edaphic conditions, minimising the risk of invasion, and helping to conserve genetic diversity (Gann et al., 2019).
Plant species selection should consider the species’ ability to survive in semi-arid and/or disturbed environments, the long period of flowering ensured by the species mix, the different life type (Bretzel et al., 2016), the germination characteristics (Bhatt et al., 2019), and the establishment performance of the plant species (Scheper et al., 2021). Adopting seed mixes with annual, biennial, and perennial species provides the benefits of different life strategies. Annual species act as nurse crops during the first year of sowing, improve the ornamental value, and can persist by self-seeding. On the other hand, biennial and perennial species persist and flower over different years, thus reducing the need for re-sowing (Hoyle, 2016). Using species with different flowering periods prolongs the enjoyment people can derive from them as well as the resources available for pollinators (Hoyle et al., 2018; Fernandes et al., 2023).
Research on wildflowers has also been aimed at increasing pollinators and bees in agroecosystems (Karamaouna et al., 2019). In Spain experiments have focused on revegetating olive groves with herbaceous entomogamous species (Jiménez-Alfaro et al., 2020). Species that flower in summer such as Cephalaria transsylvanica (L.) Schrad., are helpful in wildflower strips to feed bees when other resources are scarce, since this species easily germinates, grows, and adapts to drought (Benelli et al., 2014).
Wildflower strips are generally made up of annual and perennial forbs that are tolerant to disturbance and insect pollination, while the composition of some ecological models is associated with annual archeophytes, such as Agrostemma githago L., Nigella damascena L., Centaurea cyanus L., and Papaver rhoeas L. These species have coexisted with cultivation over time (Benvenuti and Bretzel, 2017), germinate and grow quickly, prefer fertile soil, and need seeding every year (Cameron, 2016) (Figure 6). Other species with long summer flowering are Delphinium consolida L., Malva sylvestris L., Malva punctata (All.) Alef., Scabiosa columbaria L., Dianthus carthusianorum L., Verbascum sinuatum L. and Cichorium intybus L. Some novel wildflower mix compositions based on scientific evidence have been shown to be more effective in attracting wild bees (Nichols et al., 2023).

Figure 6 Annual wildflower strip sown in a cultivation of winter wheat at Lavoria, Pisa (Italy). The species are Centaurea cyanus L., Papaver rhoeas L., Anthemis cotula L., Agrostemma githago L., Glebionis coronaria (L.) Cass. ex Spach (Photo S. Benvenuti).
Some of the native species listed in studies on seed production for grassland restoration (Kiehl et al., 2014) can be used in wildflower meadows in urban areas, although the pedoclimatic conditions are also different due to the extreme temperatures of cities under the effects of climate change.
Grassland and wildflower meadows are generally sown, but they can also be created using other methods. Green or dry hay transfer from species-rich donor sites is a good solution for the environmental restoration of species-rich grassland meadows (Valkó et al., 2022), which can be applied to anthropized habitats, e.g., green roofs (Baumann et al., 2021). The main limitation of the transfer of mown green or dry grassland cuttings is that certain species tend to fail, due to the varying stages of maturity from one seed species to another. The use of multiple donor sites, different cutting periods, and the combination of hay transfer with seed sowing may help to overcome some of these failures (Baumann et al., 2021; Valkó et al., 2022).
When using seeds, the selection of the donor site is important because the performance of seeds from different sites can vary in relation to the recipient sites and to the characteristics of the recipient habitat (Wallin et al., 2009). In the case of restoration, a donor meadow needs to be identified in order to preserve both inter- and intra- species genetic diversity, and to guarantee conservation and ecosystem resilience to climate change (Scotton et al., 2012). Most information on the production and germination of seeds of native herbaceous species is related to ecological restoration (Pedrini and Dixon, 2020; Shaw et al., 2020; Gibson-Roy et al., 2021), for which guidelines have been published (De Vitis et al., 2018).
The production of native seeds in cultivated settings reduces the impact on the natural populations, leads to higher productivity and improves the quality and reliability of the seeds (Figure 7).

Figure 7 Native wildflower seed production for commercial purposes, the company is based in Udine (Italy). The cultivation of wildflower species is carried out in monospecific plots, and later assembled in adequate percentages to obtain the mixes (Photo S. Assolari – Semenostrum).
It is important to minimize unintended selection and maintain genetic diversity in order to enhance the resilience of the plant community. High inter- and intra- specific genetic diversity guarantees the prevention of genetic erosion, inbreeding depression, and the creation of weak plant populations. Repeated propagation of mother plants, which is often necessary to obtain sufficient seeds, can further reduce genetic diversity and can involuntarily select specific genotypes (Vander Mijnsbrugge et al., 2010). While in nature great intraspecific variability is a valid strategy for coping with climate change and safeguarding the survival of the species, for the creation of wildflower meadows this variability needs to be restricted through certifications in order to prevent the diffusion of seeds of unknown origin (Abbandonato et al., 2018).
In Australia the main focus has been on returning native plants to the urban environment and on restoring native vegetation. In fact, Australian flora has high endemicity, and the issue of naturalisation of non-native plants is very acute.
In the United States and Australia, the market for native seeds is worth over 250 million US dollars. These seeds are supplied by a few companies that operate on a small scale, which have no link with the academic sector and also do not possess the basic knowledge, from an ecological point of view, of the main plant species of interest. On the other hand, the European market is worth less than 2% of that of the United States and Australia (De Vitis et al., 2018).
Often the seeds of spontaneous plants are dispersed as soon as they are ripe, thus the harvest needs to take place earlier, irrespective of the stage of seed ripening, which varies from species to species and even at an intraspecific level (Laverack et al., 2006). During collection and storage, the seeds need careful cleaning, to eliminate weed seeds, and drying to guarantee their germinability. Coating is used for rare or hard to germinate seeds, which increases the costs but protects the seeds and improves germinability (Pedrini et al., 2017). Mechanical harvesting, fertilization, and irrigation are all agronomic practices and part of the domestication process, which affect the variability by selecting some individuals rather than others. On the other hand, cultivation is necessary to provide a higher quantity of seeds to sell in the market and to stop people collecting seeds for themselves in the fields, thereby impoverishing the natural ecosystems. Germination plays a key role in the successful establishment of wildflower communities under field conditions, and it is important for creating a dense vegetation cover, which controls weeds and provides the ecological benefits of rich flowering vegetation (Kollmann and Bassin, 2001). Consequently, some specific features, such as rapid and uniform germination and plant growth, and flower colours, are essential in the selection of wildflower species (Bratcher et al., 1993).
It is estimated that 50-90% of wild plants produce seeds that are dormant at maturity, with dormancy depending on environmental conditions, geographical distribution, growth form, and genetics (Baskin and Baskin, 2014). Seed dormancy is an evolutionary adaptation that favors long-term plant survival in nature (Willis et al., 2014). However, in the context of wildflower meadows, where rapid plant establishment is essential to prevent the effects of competition among individuals related to high density, dormancy can pose significant problems (Turner et al., 2013). Little information is available in the literature on the requirements for breaking seed dormancy and on the germination of the various plant species used to create wildflower meadows (Baskin and Baskin, 2004). Furthermore, in wild species, germination is strictly controlled by water, temperature, and light. Adverse environmental factors promote the formation of seed banks and the growth of the embryo only when the external conditions ensure maximum survival of the young seedlings; this is essential to improve the survival of a species in nature (Baskin and Baskin, 2014).
Temperature is one of the most critical environmental factors for the germination and establishment of herbaceous species (Larsen and Bibby, 2005). The amount of natural light required is also important and varies considerably between wild species, especially in relation to seed size. Many native wildflower species have small seeds and require light for germination (Colbach et al., 2002), others are light-indifferent or germinate better in the dark (Baskin and Baskin, 1998). The need for light can depend on the temperature because some species require constant light and a constant temperature. Others are indifferent to light, but need fluctuating temperatures to germinate, and in some species, stratification or high temperatures can substitute light requirements (Toscano et al., 2018). Toscano et al. (2018) analyzed the seed germination behavior of 35 Mediterranean plant species to determine the influence of light and temperature on seed germination. In relation to the high number of plant species potentially used for the creation of flower meadows, further studies are needed to better identify the germinal requirements of the native species used. The Native Seed Science, Technology and Conservation Initial Training Network (NASSTEC) investigated the effects of a wide range of treatments to improve the germination of 28 wildflower species (De Vitis et al., 2018). The objective was to produce information on germination characteristics, test germination protocols for wild species, and improve the vigor and stress tolerance of seeds and seedlings, after the application of seed pre-treatments. The results showed how the conditions in which the seeds are produced impacts on the characteristics of the seeds, which can drastically change the germination requirements and the response to treatments.
Seed cleaning improves storage capacity and duration as non-separated dispersal organs may rot (De Vitis et al., 2020). The diversity of seed shapes requires different cleaning approaches for each species (Frischie et al., 2020). Seed size also affects the response to soil texture, which can markedly reduce seed germination and seedling emergence, partially in the case of sandy-loam soils, and substantially in clayey-loam soils (Benvenuti, 2023). In any case, seed size is a species-specific response that can reduce the number of species in the meadow (Figure 8), therefore removing the most important benefit of wildflower meadows which is the development of a habitat for pollinators (Nichols et al., 2022).

Figure 8 Different size and weight of wildflower seeds can be the cause of unpredictable and/or erratic germination (Photo S. Benvenuti).
Seed testing ensures the value of a seed lot, and Ryan et al. (2008) in the ISTA Bulletin, reported that ‘the establishment of appropriate and repeatable germination protocols for wildflower species is the first step toward quality control in these species’. To date, information on wildflower seed germination is lacking. If dormancy and germination characteristics are not taken into account, this could lead to the failure of immediate plant establishment. Because seeds of many native plant species exhibit dormancy (Baskin and Baskin, 2014), it is important to adopt other measures besides germination to determine the quality of seeds (Frischie et al., 2020; Pedrini and Dixon, 2020), and to be aware that germination will start when the right environmental conditions occur.
The agricultural seed industry has developed guidelines, rules, and protocols for testing seed quality (Association of Official Seed Analysts, 2018; International Seed Testing Association, 2023). On the other hand, the native seed industry needs to take into account the high interspecific variability (Hamasha and Hensen, 2009), since the complex morphological and physiological seed traits (Baskin and Baskin, 2014) have limited the application of seed quality testing within the international frameworks of the International Seed Testing Association (ISTA) (Ryan et al., 2008; Pedrini and Dixon, 2020).
Non fertile soils are generally preferred over fertile soils, as the spread of possible unwanted or invasive species is limited. Consequently, in fertile soils, ruderals become dominant and produce a large biomass which excludes slow-growing flowering species by competition (Bretzel et al., 2016). Many urban sites, such as the marginal areas of parks, gardens, cemeteries, didactic gardens, school gardens, road verges, railway embankment borders, and roundabouts, have low fertility soil which makes them suitable for wildflower meadows.
Anthropized soil can be revegetated through spontaneous colonization if meadow species pools are available in the soil seed bank or in surrounding habitats, and the soil conditions are suitable. Whether to handle the existing seed bank using mechanical or chemical methods is still under debate. When the soil is too fertile, tillage and management help to i) control the dominant weeds that compete with the seedlings and cause species failure; and ii) increase biodiversity when weed killers, fertilisers and pesticides are not employed (Bertoncini et al., 2012).
To improve the establishment of new wildflower meadows, mulching with straw or hay promotes the number of individuals per m2 and removing the sward in about 10 cm wide strips is efficient in creating gaps for the establishment of species in the long-term after new sowing (Mårtensson, 2017). The use of compost was evaluated when wildflower meadows were established in the soil of a mine, and the results demonstrated that the addition of organic matter may not be necessary for successful wildflower establishment and low levels of compost are sufficient to promote the restoration of ecosystem functions, such as water and nutrient cycling (Rothman et al., 2021).
In order to adopt sustainable cultivation methods, native soil microbes have been used because they can potentially improve the seed germination and growth of wildflowers (Barrera et al., 2021). Three different planting depths (2-cm, 6-cm, and 12-cm) were also adopted, and the response appeared to be species-specific, underlining the importance of the suitable preparation of the planting site (Barrera et al., 2021). Techniques such as topsoil removal have an excessive impact on the environment, and it is more important to exploit low fertility soils.
Reference frameworks given in the literature for setting up the planting and management of flower meadows on a rational basis are not always adequate for non-temperate climatic areas (e.g., hot and dry such as the Mediterranean basin). To promote the diffusion of wildflower meadows in urban environments, the agronomic aspects need to be adequately defined - from the choice of species to the composition of the mixture, the characteristics of the soils, and the agricultural interventions required.
A key aspect is to identify the most appropriate time for sowing and cutting, and this depends on the geographical area where the wildflower meadow is located. In dry and hot Mediterranean climates, sowing should generally be carried out before winter to take advantage of the autumn-winter rainy season. In fact, autumn sowing is suitable for perennials to break dormancy and to also bring forward the flowering season (Aldrich, 2002). In Nordic regions, spring sowing may be more suitable, and to prevent damage from frost, annuals can be sown until late spring (May) (Scott, 2003).
In relation to mowing time, in cool and wet summers the meadow flowering starts in late May-June and throughout the summer provides a very colourful display. On the other hand, in dry and hot climates flowering can start in March, and depending on the yearly trend, by the end of May the vegetation may have already gone dry, so it needs to be mowed as it is flammable and collects rubbish.
In the UK, Prentis and Norton (1992) recommend sowing in August to early October or March to mid-May. Operating in Gembloux, Belgium, Piqueray et al. (2021) found that autumn sowing and early cut improve flowering abundance and diversity and ensure a better control of annuals in the first year after sowing. Because environmental conditions differ, using the phenology of the plants, and in particular flowering, can be useful to understand if and when mowing should be suspended (Brom et al., 2023).
Sowing is carried out by mixing the seeds with inert material (sawdust, sand), raking, and in some cases covering with a jute or geotextile matting after sowing (Hitchmough, 2016). Sowing density affects the establishment of the meadow. If the soil is fertile, a high seed density is critical for the risk of inter- and intra- species competition. On the other hand, in unproductive and weed-free soils, and when the first year flowering is not required, low seed density is more successful for long-term establishment (Jiang and Hitchmough, 2022). However, lower sowing densities lead to spontaneous colonization by the local native flora (Burton et al., 2006).
The different size of the seeds is another critical aspect. Fine seeds are often light-dependent and need to be on the surface, while large seeds are less dependent on light intensity and need good contact with the soil particles in order to absorb water and start germinating, which can only be achieved if the seeds are buried in the soil. Good results were achieved by distributing a layer of sand on the soil surface, as a sowing environment, which decreases the seed bank plant competition (Hitchmough, 2016), however this is probably more suited to drought-free latitudes.
Mowing is fundamental in order to increase biodiversity, however, maintaining the diversity of planting for several years is difficult due to the vegetation dynamics in productive soils (Piqueray et al., 2019).
Turf can be converted to species-rich meadows by reducing the frequency of mowing, especially in critical periods such as spring and early summer, thus allowing the flowering of many species which are tolerant to disturbance (e.g., Bellis perennis, Taraxacum spp., Crepis spp., Aiuga reptans, Trifolium spp., Hypochoeris radicata, Ranunculus bulbosus L.), (Figure 9) and can be a habitat for pollinators and small fauna (Chollet et al., 2018; Sehrt et al., 2020).

Figure 9 Reducing the mowing of turf in spring, surprisingly, turns what has been defined as a “green desert” into a rich habitat which, allowing abundant flowering of the dicotyledonous species, provides food and shelter to insects, amphibians, and birds, and provides a pleasant sensory experience for people (Photo R. Pini).
In relation to mowing time, species life-form composition is important. For instance, July mowing was shown to be better than October in terms of increasing the biodiversity of plant species in an only-once-mown meadow composed of perennial forbs, (Vannucchi et al., 2015). September mowing was more effective in reducing the biomass of perennials at the right time to improve the seedling recruitment of annuals, and increase the overall diversity (Bretzel et al., 2012). Annuals, perennials, and bulbs added to warm-season lawn grass Cynodon dactylon (L.) Pers. can withstand frequent mowing and competition in the summer (grass growing season), and flower during the winter (grass dormancy). This helps extend the ornamental value and diversity of the lawn over a longer period (Mirabile et al., 2016).
In some cases, geophytes and forb species, which are particularly ornamental or difficult to reproduce by seed in situ, can be transplanted (Hitchmough, 2000). Wallin et al. (2009) compared seed sowing with the planting of plug-plants, i.e., grown from seed in the greenhouse, for the restoration of plant populations in semi-natural grasslands. They found that for two perennial herbs commonly growing in meadows, plug-planted transplants were twice the size of the seed-sown plants in both species. A similar result was found with plug planting in grass swards in Sweden (Mårtensson, 2017).
Earlier research included chemical weeding with glyphosate and burning, while the most recent techniques only use mowing to control undesired weeds. As the aim is generally related to the improvement in biodiversity as an ecosystem service, the most important issue is the total number of species naturalized in the meadow.
Sowing has also been associated with mycorrhizal inoculation, however no increase in the establishment has been observed, while the foliage cover and sowing time can affect the presence of target species (Fischer et al., 2013). Pre-sown biodegradable mats can be used, especially in areas where tilling is not possible or difficult (Aldrich, 2002). Portions of turf can be transplanted either for restoration purposes (Klimeš et al., 2010) or for new plantings. If natural sources of plant seeds are lacking, transplanting turf has been found to be effective for establishing a meadow, although the target species was not found to expand much from the turf to the surrounding areas. However, the success of transplanting should be based on long-term observations (Mudrák et al., 2017).
Planting meadows in anthropogenic landscapes, urban and rural areas, compensates for the loss of habitats for pollinators and the food web, offers a conservation opportunity for many plant species, and provides the possibility for people to enjoy nature and citizen science programs. Meadow vegetation provides many ecosystem services, which are now more necessary than ever for a healthy life on the planet. The methods used for establishing wildflower meadows have been widely investigated in northern Europe and America, and now studies are being carried out in different climatic regions such as the Mediterranean. The connection to the geographical area is key if conservation is a primary aim. Constraints are represented by the difficulties in finding the germplasm, as in many countries the seed market is not yet ready to offer the most appropriate materials, i.e. seeds of species from local populations. Nevertheless, reducing the mowing regimes of turf in cities, when the conditions are favorable for a large percentage of flowering species, can represent a win-win solution to conserve biodiversity, to provide ecosystem services and satisfy people’s need for nature. Further studies on plant life forms, cycles and phenological phases in wildflower meadows under climate change need to address the shift from perennial to annual cycles in drought climates as well as the ecosystem services related to soil protection and pollution reduction.
FB: Writing – original draft, conceptualization, review & editing. FV: Writing – review & editing. BP: writing – review & editing. AP: writing – review & editing. DR: Supervision, writing – review & editing. All authors contributed to the article and approved the submitted version.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was funded under the National Recovery and Resilience Plan (NRRP), Mission 4 Component 2 Investment 1.4 - Call for tender No. 3138 of 16 December 2021, rectified by Decree n.3175 of 18 December 2021 of Italian Ministry of University and Research funded by the European Union – NextGenerationEU; Award Number: Project code CN_00000033, Concession Decree No. 1034 of 17 June 2022 adopted by the Italian Ministry of University and Research, CUP, H43C22000530001 Project title "National Biodiversity Future Center" – NBFC.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors DR and FB declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fhort.2023.1248785/full#supplementary-material
Abbandonato H., Pedrini S., Pritchard H. W., De Vitis M., Bonomi C. (2018). Native seed trade of herbaceous species for restoration: a European policy perspective with global implications. Restor. Ecol. 26 (5), 820–826. doi: 10.1111/rec.12641
Albrecht H. (2003). Suitability of arable weeds as indicator organism to evaluate species conservation effects of management in the agricultural ecosystems. Agric. Ecosyst. Environ. 98, 201–211. doi: 10.1016/S0167-8809(03)00081-1
Aldrich J. H. (2002). Factors and benefits in the establishment of modest-sized wildflower plantings: a review. Native Plants J. 3 (1), 67–86. doi: 10.3368/npj.3.1.67
Andersson E., Barthel S., Borgström S., Colding J., Elmqvist T., Folke C., et al. (2014). Reconnecting cities to the biosphere: stewardship of green infrastructure and urban ecosystem services. Ambio 43, 445–453. doi: 10.1007/s13280-014-0506-y
Aronson M. F., Lepczyk C. A., Evans K. L., Goddard M. A., Lerman S. B., MacIvor J. S., et al. (2017). Biodiversity in the city: key challenges for urban green space management. Front. Ecol. Environ. 15 (4), 189–196. doi: 10.1002/fee.1480
Association of Official Seed Analysts. (2018). AOSA rules for testing seeds. Available at: https://www.nist.gov/system/files/documents/2017/04/28/11-appd-11-hb133-final.pdf (Accessed 16 May 2023).
Atkins E. (2018). “Green streets as habitat for biodiversity,” in Nature based strategies for urban and building sustainability. Eds. Pérez G., Perin K. (Butterworth-Heinemann: Elsevier), 251–260.
Baessler C., Klotz S. (2006). Effects of changes in agricultural land-use on landscape structure and arable weed vegetation over the last 50 years. Agric. Ecosyst. Environ. 115, 43–50. doi: 10.1016/j.agee.2005.12.007
Barrera D., Luera J., Lavallee K., Soti P. (2021). Influence of microbial priming and seeding depth on germination and growth of native wildflowers. Ecol. Process 10, 1–8. doi: 10.1186/s13717-021-00287-4
Barry C., Hodge S. (2023). You reap what you sow: A botanical and economic assessment of wildflower seed mixes available in Ireland. Conservation 3 (1), 73–86. doi: 10.3390/conservation3010007
Baskin C. C., Baskin J. M. (1998). Seeds: ecology, biogeography, and evolution of dormancy and germination (San Diego, California: Academic Press).
Baskin C. C., Baskin J. M. (2004). Germinating seeds of wildflowers, an ecological perspective. HortTechnology 14 (4), 467–473. doi: 10.21273/HORTTECH.14.4.0467
Baskin C. C., Baskin J. M. (2014). Seeds: ecology, biogeography, and evolution of dormancy and germination. 2nd edition (San Diego, CA: Elsevier, Academic Press).
Baumann N., Catalano C., Pasta S. (2021). “Improving extensive green roofs for endangered ground-nesting birds,” in Urban services to ecosystems. Eds. Catalano C., Andreucci M. B., Guarino R., Bretzel F., Leone M., Pasta S. (Cham: Springer), 13–29.
Benelli G., Benvenuti S., Desneux N., Canale A. (2014). Cephalaria transsylvanica-based flower strips as potential food source for bees during dry periods in European Mediterranean basin countries. PloS One 9 (3), e93153. doi: 10.1371/journal.pone.0093153
Benvenuti S. (2014). Wildflower green roofs for urban landscaping, ecological sustainability and biodiversity. Landsc. Urban Plan. 124, 151–161. doi: 10.1016/j.landurbplan.2014.01.004
Benvenuti S. (2023). Soil texture involvement in wildflower strip ecosystem services delivery in Mediterranean agro-environment. Eur. J. Agron. 145, 126793. doi: 10.1016/j.eja.2023.126793
Benvenuti S., Bretzel F. (2017). Agro-biodiversity restoration using wildflowers: What is the appropriate weed management for their long-term sustainability? Ecol. Eng. 102, 519–526. doi: 10.1016/j.ecoleng.2017.02.062
Bertoncini A. P., Machon N., Pavoine S., Muratet A. (2012). Local gardening practices shape urban lawn floristic communities. Landsc. Urban Plan. 105 (1-2), 53–61. doi: 10.1016/j.landurbplan.2011.11.017
Bhatt A., Bhat N. R., Santo A., Phartyal S. S. (2019). Influence of temperature, light and salt on the germination of Deverra triradiata seeds. Seed Sci. Technol. 47 (1), 25–31. doi: 10.15258/sst.2019.47.1.03
Bratcher C. B., Dole J. M., Cole J. C. (1993). Stratification improves seed germination of five native wildflower species. HortScience 28 (9), 899–901. doi: 10.21273/HORTSCI.28.9.899
Bretzel F., Malorgio F., Paoletti L., Pezzarossa B. (2012). Response of sowed, flowering herbaceous communities suitable for anthropic Mediterranean areas under different mowing regimes. Landsc. Urban Plan. 107 (2), 80–88. doi: 10.1016/j.landurbplan.2012.05.002
Bretzel F., Vannucchi F., Romano D., Malorgio F., Benvenuti S., Pezzarossa B. (2016). Wildflowers: from conserving biodiversity to urban greening - a review. Urban For. Urban Green. 20, 428–426. doi: 10.1016/j.ufug.2016.10.008
Brom P. D., Underhill L. G., Winter K., Colville J. F. (2023). A mowing strategy for urban parks to support spring flowers in a Mediterranean climate city in South Africa. Urban Ecosyst. 26, 1143–1153. doi: 10.1007/s11252-023-01369-1
Burton C. M., Burton P. J., Hebda R., Turner N. J. (2006). Determining the optimal sowing density for a mixture of native plants used to revegetate degraded ecosystems. Restor. Ecol. 14 (3), 379–390. doi: 10.1111/j.1526-100X.2006.00146.x
Cameron R. W. F. (2016). “Bedding and annual flowering plants,” in Environmental horticulture: science and management of green landscapes. Eds. Cameron R. W. F., Hitchmough J. D. (Wallingford UK: Cabi), 175–191.
Catalano C., Marcenò C., Laudicina V. A., Guarino R. (2016). Thirty years unmanaged green roofs: Ecological research and design implications. Landsc. Urban Plan. 149, 11–19. doi: 10.1016/j.landurbplan.2016.01.003
Chollet S., Brabant C., Tessier S., Jung V. (2018). From urban lawns to urban meadows: Reduction of mowing frequency increases plant taxonomic, functional and phylogenetic diversity. Landsc. Urban Plan. 180, 121–124. doi: 10.1016/j.landurbplan.2018.08.009
Colbach N., Chauvel B., Dürr C., Richard G. (2002). Effect of environmental conditions on Alopecurus myosuroides germination. I. Effect of temperature and light. Weed Res. 42 (3), 210–221. doi: 10.1046/j.1365-3180.2002.00279.x
Colding J., Colding M., Barthel S. (2020). The smart city model: A new panacea for urban sustainability or unmanageable complexity? Environ. Plan. B Urban Anal. City Sci. 47 (1), 179–187. doi: 10.1177/2399808318763164
Dale A. G., Perry R. L., Cope G. C., Benda N. (2020). Floral abundance and richness drive beneficial arthropod conservation and biological control on golf courses. Urban Ecosyst. 23 (1), 55–66. doi: 10.1007/s11252-019-00907-0
Derby Lewis A., Bouman M. J., Winter A. M., Hasle E. A., Stotz D. F., Johnston M. K., et al. (2019). Does nature need cities? Pollinators reveal a role for cities in wildlife conservation. Front. Ecol. Evol. 7. doi: 10.3389/fevo.2019.00220
De Vitis M., Hay F. R., Dickie J. B., Trivedi C., Choi J., Fiegener R. (2020). Seed storage: maintaining seed viability and vigor for restoration use. Restor. Ecol. 28, S249–S255. doi: 10.1111/rec.13174
De Vitis M., Mondoni A., Pritchard H. W., Laverack G., Bonomi C. (Eds.) (2018). Native Seed Ecology, Production & Policy – Advancing knowledge and technology in Europe (Trento: MUSE).
EC. (2013). Communication from the Commission to the European Parliament, The Council, The European Economic and Social Committee and the Committee of the Regions. Green Infrastructure (GI) — Enhancing Europe’s Natural Capital. COM/2013/0249 final (Brussels). Available at: https://eur-lex.europa.eu/resource.html?uri=cellar:d41348f2-01d5-4abe-b817-4c73e6f1b2df.0014.03/DOC_1&format=PDF (Accessed 5 May 2023).
Elmqvist T., Andersson E., Frantzeskaki N., McPhearson T., Olsson P., Gaffney O., et al. (2019). Sustainability and resilience for transformation in the urban century. Nat. Sustain. 2 (4), 267–273. doi: 10.1038/s41893-019-0250-1
European Commission Directorate-General for Agriculture and Rural Development (DG AGRI). (2021). List of potential AGRICULTURAL PRACTICES that ECO-SCHEMES could support. Fact Sheets, January 2021. Available at: https://agriculture.ec.europa.eu/system/files/2021-01/factsheet-agri-practices-under-ecoscheme_en_0.pdf (Accessed 21 April 2023).
Fenster C. B., Armbruster W. S., Wilson P., Dudash M. R., Thomson J. D. (2004). Pollination syndromes and floral specialization. Annu. Rev. Ecol. Evol. Syst. 35, 375–403. doi: 10.1146/annurev.ecolsys.34.011802.132347
Fernandes M. P., Matono P., Almeida E., Pinto-Cruz C., Belo A. D. (2023). Sowing wildflower meadows in Mediterranean peri-urban green areas to promote grassland diversity. Front. Ecol. Evol. 11. doi: 10.3389/fevo.2023.1112596
Fischer L. K., Honold J., Cvejić R., Delshammar T., Hilbert S., Lafortezza R., et al. (2018). Beyond green: Broad support for biodiversity in multicultural European cities. Glob. Environ. Change 49, 35–45. doi: 10.1016/j.gloenvcha.2018.02.001
Fischer L. K., von der Lippe M., Rillig M. C., Kowarik I. (2013). Creating novel urban grasslands by reintroducing native species in wasteland vegetation. Biol. Conserv. 159, 119–126. doi: 10.1016/j.biocon.2012.11.028
Fratarcangeli C., Fanelli G., Franceschini S., De Sanctis M., Travaglini A. (2019). Beyond the urban-rural gradient: Self-organizing map detects the nine landscape types of the city of Rome. Urban For. Urban Green. 38, 354–370. doi: 10.1016/j.ufug.2019.01.012
Frischie S., Miller A. L., Pedrini S., Kildisheva O. A. (2020). Ensuring seed quality in ecological restoration: native seed cleaning and testing. Restor. Ecol. 28, S239–S248. doi: 10.1111/rec.13217
Gann G. D., McDonald T., Walder B., Aronson J., Nelson C. R., Jonson J., et al. (2019). International principles and standards for the practice of ecological restoration. Restor. Ecol. 27 (S1), S1–S46. doi: 10.1111/rec.13035
Gibson-Roy P., Hancock N., Broadhurst L., Driver M. (2021). Australian native seed sector practice and behavior could limit ecological restoration success: further insights from the Australian Native Seed Report. Restor. Ecol. 29 (7), e13429. doi: 10.1111/rec.13429
Grab H., Branstetter M. G., Amon N., Urban-Mead K. R., Park M. G., Gibbs J., et al. (2019). Agriculturally dominated landscapes reduce bee phylogenetic diversity and pollination services. Science 363 (6424), 282–284. doi: 10.1126/science.aat6016
Grass I., Albrecht J., Jauker F., Diekötter T., Warzecha D., Wolters V., et al. (2016). Much more than bees—Wildflower plantings support highly diverse flower-visitor communities from complex to structurally simple agricultural landscapes. Agric. Ecosyst. Environ. 225, 45–53. doi: 10.1016/j.agee.2016.04.001
Griffiths-Lee J., Nicholls E., Goulson D. (2022). Sown mini-meadows increase pollinator diversity in gardens. J. Insect Conserv. 26 (2), 299–314. doi: 10.1007/s10841-022-00387-2
Haaland C., Gyllin M. (2011). “Sown wildflower strips–a strategy to enhance biodiversity and amenity in intensively used agricultural areas,” in The importance of biological interactions in the study of biodiversity. Ed. López-Pujol J. (Rijeka: InTech), 155–172.
Haaland C., Naisbit R. E., Bersier L. F. (2011). Sown wildflower strips for insect conservation: a review. Insect Conserv. Divers. 4 (1), 60–80. doi: 10.1111/j.1752-4598.2010.00098.x
Hamasha H. R., Hensen I. (2009). Seed germination of four Jordanian Stipa spp: differences in temperature regimes and seed provenances. Plant Species Biol. 24 (2), 127–132. doi: 10.1111/j.1442-1984.2009.00247.x
Hicks D. M., Ouvrard P., Baldock K. C., Baude M., Goddard M. A., Kunin W. E., et al. (2016). Food for pollinators: quantifying the nectar and pollen resources of urban flower meadows. PloS One 11 (6), e0158117. doi: 10.1371/journal.pone.0158117
Hitchmough J. D. (2000). Establishment of cultivated herbaceous perennials in purpose-sown native wildflower meadows in south-west Scotland. Landsc. Urban Plan. 51 (1), 37–51. doi: 10.1016/S0169-2046(00)00092-X
Hitchmough J. D. (2016). “Semi-natural grasslands and meadows,” in Environmental horticulture: Science and management of green landscapes. Eds. Cameron R. W. F., Hitchmough J. D. (Wallingford UK: Cabi), 175–191.
Hoyle H. (2016). Improving urban grassland for people and wildlife. Available at: http://www.relu.ac.uk/landbridge/LWEC%20PP%20Note%2032.pdf (Accessed April 5, 2023).
Hoyle H., Norton B., Dunnett N., Richards J. P., Russell J. M., Warren P. (2018). Plant species or flower colour diversity? Identifying the drivers of public and invertebrate response to designed annual meadows. Landsc. Urban Plan. 180, 103–113. doi: 10.1016/j.landurbplan.2018.08.017
Ignatieva M., Haase D., Dushkova D., Haase A. (2020). Lawns in cities: from a globalised urban green space phenomenon to sustainable nature-based solutions. Land 9 (3), 73. doi: 10.3390/land9030073
International Seed Testing Association. (2023). ISTA 2020 rules. Available at: https://www.seedtest.org/api/rm/39WYX6A534HB99Y/ista-rules-2023-00-introduction-final.pdf (Accessed 16 May 2023).
Jiang M., Hitchmough J. D. (2022). Can sowing density facilitate a higher level of forb abundance, biomass, and richness in urban, perennial “wildflower” meadows? Urban For. Urban Green. 74, 127657. doi: 10.1016/j.ufug.2022.127657
Jiménez-Alfaro B., Frischie S., Stolz J., Gálvez-Ramírez C. (2020). Native plants for greening Mediterranean agroecosystems. Nat. Plants 6 (3), 209–214. doi: 10.1038/s41477-020-0617-3
Karamaouna F., Kati V., Volakakis N., Varikou K., Garantonakis N., Economou L., et al. (2019). Ground cover management with mixtures of flowering plants to enhance insect pollinators and natural enemies of pests in olive groves. Agric. Ecosyst. Environ. 274, 76–89. doi: 10.1016/j.agee.2019.01.004
Kiehl K., Kirmer A., Shaw N. (Eds.) (2014). Guidelines for native seed production and grassland restoration (Cambridge: Scholars Publishing).
Klimeš L., Jongepierová I., Doležal J., Klimešová J. (2010). Restoration of a species-rich meadow on arable land by transferring meadow blocks. Appl. Veg. Sci. 13 (4), 403–411. doi: 10.1111/j.1654-109X.2010.01084.x
Kollmann J., Bassin S. (2001). Effects of management on seed predation in wildflower strips in northern Switzerland. Agric. Ecosyst. Environ. 83 (3), 285–296. doi: 10.1016/S0167-8809(00)00202-4
Kovács-Hostyánszki A., Piross I. S., Shebl M. A. (2022). Non-native plant species integrate well into plant-pollinator networks in a diverse man-made flowering plant community. Urban Ecosyst. 25 (5), 1491–1502. doi: 10.1007/s11252-022-01242-7
Kueffer C. (2020). Plant sciences for the Anthropocene: What can we learn from research in urban areas? Plants People Planet 2 (4), 286–289. doi: 10.1002/ppp3.10124
Larsen S. U., Bibby B. M. (2005). Differences in thermal time requirement for germination of three turfgrass species. Crop Sci. 45 (5), 2030–2037. doi: 10.2135/cropsci2004.0731
Laverack G., Matthews S., Powell A. A., Hosseini M. K. (2006). Scottish wildflower seeds: Production and use. Bot. J. Scotl. 58 (1), 49–58. doi: 10.1080/03746600608685106
Leong M., Dunn R. R., Trautwein M. D. (2018). Biodiversity and socioeconomics in the city: a review of the luxury effect. Biol. Lett. 14 (5), 20180082. doi: 10.1098/rsbl.2018.0082
Lepczyk C. A., Aronson M. F., Evans K. L., Goddard M. A., Lerman S. B., MacIvor J. S. (2017). Biodiversity in the city: fundamental questions for understanding the ecology of urban green spaces for biodiversity conservation. BioScience 67 (9), 799–807. doi: 10.1093/biosci/bix079
Lerman S. B., Contosta A. R., Milam J., Bang C. (2018). To mow or to mow less: Lawn mowing frequency affects bee abundance and diversity in suburban yards. Biol. Conserv. 221, 160–174. doi: 10.1016/j.biocon.2018.01.025
Liu J., Coomes D. A., Gibson L., Hu G., Liu J., Luo Y., et al. (2019). Forest fragmentation in China and its effect on biodiversity. Biol. Rev. 94 (5), 1636–1657. doi: 10.1111/brv.12519
Marshall C. A., Wilkinson M. T., Hadfield P. M., Rogers S. M., Shanklin J. D., Eversham B. C., et al. (2023). Urban wildflower meadow planting for biodiversity, climate and society: An evaluation at King’s College, Cambridge. Ecol. Solutions Evidence 4 (2), e12243. doi: 10.1002/2688-8319.12243
Mårtensson L. M. (2017). Methods of establishing species-rich meadow biotopes in urban areas. Ecol. Eng. 103, 134–140. doi: 10.1016/j.ecoleng.2017.03.016
Meenar M., Heckert M., Adlakha D. (2022). “Green enough ain’t good enough:” Public perceptions and emotions related to green infrastructure in environmental justice communities. Int. J. Environ. Res. Public Health 19, 1448. doi: 10.3390/ijerph19031448
Mirabile M., Bretzel F., Gaetani M., Lulli F., Volterrani M. (2016). Improving aesthetic and diversity of Bermudagrass lawn in its dormancy period. Urban For. Urban Green. 18, 190–197. doi: 10.1016/j.ufug.2016.06.007
Mody K., Lerch D., Müller A. K., Simons N. K., Blüthgen N., Harnisch M. (2020). Flower power in the city: Replacing roadside shrubs by wildflower meadows increases insect numbers and reduces maintenance costs. PloS One 15 (6), e0234327. doi: 10.1371/journal.pone.0234327
Mudrák O., Fajmon K., Jongepierová I., Doležal J. (2017). Restoring species-rich meadow by means of turf transplantation: long-term colonization of ex-arable land. Appl. Veg. Sci. 20, 62–73. doi: 10.1111/avsc.12281
Nichols R. N., Holland J. M., Goulson D. (2022). Can novel seed mixes provide a more diverse, abundant, earlier, and longer-lasting floral resource for bees than current mixes?Basic Appl. Ecol. 60, 34–47. doi: 10.1016/j.baae.2022.02.002
Nichols R. N., Holland J. M., Goulson D. (2023). A novel farmland wildflower seed mix attracts a greater abundance and richness of pollinating insects than standard mixes. Insect Conserv. Divers. 16 (2), 190–204. doi: 10.1111/icad.12624
Norton B. A., Bending G. D., Clark R., Corstanje R., Dunnett N., Evans K. L., et al. (2019). Urban meadows as an alternative to short mown grassland: effects of composition and height on biodiversity. Ecol. Appl. 29 (6), e01946. doi: 10.1002/eap.1946
Paraskevopoulou A. T., Pappous E., Biniari K., Bertsouklis K. F., Daskalakis I., Perdikis D. (2022). Enhancing the rural landscape character: the low frequency of inter-row wildflower meadow harvest positively affects biodiversity while maintaining grape quantitative and qualitative traits in a ‘Sultanina’Vineyard in Greece. Agronomy 12 (3), 550. doi: 10.3390/agronomy12030550
Pedrini S., Dixon K. W. (2020). International principles and standards for native seeds in ecological restoration. Restor. Ecol. 28, S286–S303. doi: 10.1111/rec.13155
Pedrini S., Merritt D. J., Stevens J., Dixon K. (2017). Seed coating: science or marketing spin? Trends Plant Sci. 22 (2), 106–116. doi: 10.1016/j.tplants.2016.11.002
Piqueray J., Gilliaux V., Bodson B., Mahy G. (2021). Autumn sowing and first-year mowing enhance flowering species abundance and diversity in wildflower strips. BASE 25 (1), 1–7. doi: 10.25518/1780-4507.18812
Piqueray J., Gilliaux V., Decruyenaere V., Cornelis J. T., Uyttenbroeck R., Mahy G. (2019). Management of grassland-like wildflower strips sown on nutrient-rich arable soils: the role of grass density and mowing regime. Environ. Manage. 63 (5), 647–657. doi: 10.1007/s00267-019-01153-y
Plant Conservation Alliance. (2021). National seed strategy progress report 2015–2020 (Washington (DC: US Department of the Interior, Bureau of Land Management). Available at: https://www.blm.gov/sites/default/files/docs/2021-08/Progress%20Report%2026Jul21.pdf (Accessed 5 May 2023).
Prentis E., Norton G. A. (1992). “MEADOWS”—an expert system for the establishment of diverse wildflower grasslands on derelict land in urban areas in the UK. Ecol. Eng. 1 (3), 213–228. doi: 10.1016/0925-8574(92)90003-K
Przybysz A., Popek R., Stankiewicz-Kosyl M., Zhu C. Y., Małecka-Przybysz M., Maulidyawati T., et al. (2021). Where trees cannot grow–Particulate matter accumulation by urban meadows. Sci. Total Environ. 785, 147310. doi: 10.1016/j.scitotenv.2021.147310
Rothman S. E., Cole C. A., Bruns M. A., Hall M. (2021). The influence of soil amendments on a native wildflower seed mix in surface mine restoration. Restor. Ecol. 29 (7), e13440. doi: 10.1111/rec.13440
Ryan N., Laverack G., Powell A. (2008). Establishing quality control in UK wildflower seed production. Seed Testing Int. 135, 49–53.
Scheper J., Bukovinszky T., Huigens M. E., Kleijn D. (2021). Attractiveness of sown wildflower strips to flower-visiting insects depends on seed mixture and establishment success. Basic Appl. Ecol. 56, 401–415. doi: 10.1016/j.baae.2021.08.014
Schröder R., Kiehl K. (2020). Ecological restoration of an urban demolition site through introduction of native forb species. Urban For. Urban Green. 47, 126509. doi: 10.1016/j.ufug.2019.126509
Scott R. (2004). “Wildflower landscapes in the urban environment”, in Plant User Handbook: A Guide to Effective Specifying, eds. Hitchmough J., Fieldhouse K. (Oxford, UK: Blackwell), 258–266.
Scotton M., Kirmer A., Krautzer B. (2012). Practical handbook for seed harvest and ecological restoration of species-rich grasslands (Padova: CLUEB, Cooperativa Libraria Editrice Università di Padova), ISBN: ISBN 978-88-6129-800-2.
Sehrt M., Bossdorf O., Freitag M., Bucharova A. (2020). Less is more! Rapid increase in plant species richness after reduced mowing in urban grasslands. Basic Appl. Ecol. 42, 47–53. doi: 10.1016/j.baae.2019.10.008
Shaw N., Barak R. S., Campbell R. E., Kirmer A., Pedrini S., Dixon K., et al. (2020). Seed use in the field: delivering seeds for restoration success. Restor. Ecol. 28, S276–S285. doi: 10.1111/rec.13210
Shu X., Mesimäki M., Kotze D. J., Wales M., Xie L., Benicke R., et al. (2022). Needs and expectations of German and Chinese children for livable urban green spaces revealed by the method of empathy-based stories. Urban For. Urban Green. 68, 127476. doi: 10.1016/j.ufug.2022.127476
Spahillari M., Hammer K., Gladis T., Diederichsen A. (1999). Weeds as part of agrobiodiversity. Outlook Agric. 28 (4), 227–232. doi: 10.1177/003072709902800405
Steiner H. (2023). I never promised you a rose garden. When landscape architecture becomes a laboratory for the Anthropocene. Hist. Hum. Sci. 36 (2), 178–201. doi: 10.1177/09526951221103077
Toscano S., Romano D., Tribulato A., Cavallaro V. (2018). Assessing and modeling seed germination of Mediterranean wildflowers for low input landscape restoration. Restor. Ecol. 26 (3), 525–536. doi: 10.1111/rec.12608
Tschumi M., Albrecht M., Bärtschi C., Collatz J., Entling M. H., Jacot K. (2016). Perennial, species-rich wildflower strips enhance pest control and crop yield. Agric. Ecosyst. Environ. 220, 97–103. doi: 10.1016/j.agee.2016.01.001
Turner S. R., Steadman K. J., Vlahos S., Koch J. M., Dixon K. W. (2013). Seed treatment optimizes benefits of seed bank storage for restoration-ready seeds: The feasibility of prestorage dormancy alleviation for mine-site revegetation. Restor. Ecol. 21 (2), 186–192. doi: 10.1111/j.1526-100X.2012.00879.x
UN Habitat. (2020). The new urban agenda (Nairobi, Kenya: United Nations Human Settlements Programme (UN-Habitat).
United Nations. (2019). World population prospects: the 2019 revision (New York: UN Department of Economic and Social Affairs Population Division). Available at: https://www.un.org/development/desa/pd/news/world-population-prospects-2019-0 (Accessed 5 May 2023).
Valkó O., Rádai Z., Deák B. (2022). Hay transfer is a nature-based and sustainable solution for restoring grassland biodiversity. J. Environ. Manage. 311, 114816. doi: 10.1016/j.jenvman.2022.114816
Vander Mijnsbrugge K., Bischoff A., Smith B. (2010). A question of origin: where and how to collect seed for ecological restoration. Basic Appl. Ecol. 11 (4), 300–311. doi: 10.1016/j.baae.2009.09.002
Vannucchi F., Malorgio F., Pezzarossa B., Pini R., Bretzel F. (2015). Effects of compost and mowing on the productivity and density of a purpose-sown mixture of native herbaceous species to revegetate degraded soil in anthropized areas. Ecol. Eng. 74, 60–67. doi: 10.1016/j.ecoleng.2014.09.121
Vannucchi F., Pini R., Scatena M., Benelli G., Canale A., Bretzel F. (2018). Deinking sludge in the substrate reduces the fertility and enhances the plant species richness of extensive green roofs. Ecol. Eng. 116, 87–96. doi: 10.1016/j.ecoleng.2018.02.027
Vega K. A., Küffer C. (2021). Promoting wildflower biodiversity in dense and green cities: The important role of small vegetation patches. Urban For. Urban Green. 62, 127165. doi: 10.1016/j.ufug.2021.127165
Vega K. A., Schläpfer-Miller J., Kueffer C. (2021). Discovering the wild side of urban plants through public engagement. Plants People Planet 3 (4), 389–401. doi: 10.1002/ppp3.10191
Wallin L., Svensson B. M., Lönn M. (2009). Artificial dispersal as a restoration tool in meadows: sowing or planting? Restor. Ecol. 17 (2), 270–279. doi: 10.1111/j.1526-100X.2007.00350.x
Wang F., Shi G., Nicholas O., Yao B., Ji M., Wang W., et al. (2018). Ecosystem nitrogen retention is regulated by plant community trait interactions with nutrient status in an alpine meadow. J. Ecol. 106 (4), 1570–1581. doi: 10.1111/1365-2745.12924
Willis C. G., Baskin C. C., Baskin J. M., Auld J. R., Venable D. L., Cavender-Bares J., et al. (2014). The evolution of seed dormancy: environmental cues, evolutionary hubs, and diversification of the seed plants. New Phytol. 203 (1), 300–309. doi: 10.1111/nph.12782
Zhou W., Wang J., Qian Y., Pickett S. T., Li W., Han L. (2018). The rapid but “invisible” changes in urban greenspace: A comparative study of nine Chinese cities. Sci. Total Environ. 627, 1572–1584. doi: 10.1016/j.scitotenv.2018.01.335
Keywords: seed characteristics, seed production, germination, green infrastructure planning, biodiversity, ecosystem services, plant species selection, cultivation practices
Citation: Bretzel F, Vannucchi F, Pezzarossa B, Paraskevopoulou A and Romano D (2024) Establishing wildflower meadows in anthropogenic landscapes. Front. Hortic. 2:1248785. doi: 10.3389/fhort.2023.1248785
Received: 27 June 2023; Accepted: 19 December 2023;
Published: 11 January 2024.
Edited by:
Francesco Ferrini, University of Florence, ItalyReviewed by:
Mirjana Ljubojević, University of Novi Sad, SerbiaCopyright © 2024 Bretzel, Vannucchi, Pezzarossa, Paraskevopoulou and Romano. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Francesca Bretzel, ZnJhbmNlc2NhLmJyZXR6ZWxAY25yLml0
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.