
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet. , 26 July 2024
Sec. Behavioral and Psychiatric Genetics
Volume 15 - 2024 | https://doi.org/10.3389/fgene.2024.1421870
 Luis Alberto Vázquez-Pérez1Mónica Hattori-Hara2Gloria Arankowsky-Sandoval1Gerardo Pérez-Mendoza1Rodrigo Rubi-Castellanos1
Luis Alberto Vázquez-Pérez1Mónica Hattori-Hara2Gloria Arankowsky-Sandoval1Gerardo Pérez-Mendoza1Rodrigo Rubi-Castellanos1 Jorge Aarón Rangel-Méndez1
Jorge Aarón Rangel-Méndez1 Doris Pinto-Escalante1Thelma Canto-Cetina1
Doris Pinto-Escalante1Thelma Canto-Cetina1 Lizbeth González-Herrera1*
Lizbeth González-Herrera1*Introduction: Genetic variants that control dopamine have been associated with obesity in children through loss of control of satiety and impulses, the manifestation of addictive eating behaviors, and specific personality traits. The variants include FTO-rs9939609 and the MAO-A 30 pb u-VNTR low-transcription alleles (LTA).
Objective: To evaluate the genetic association of FTO-rs9939609 and the MAO-A LTA, along with personality traits and eating behavior with obesity in Mayan children from Mexico.
Methods: We cross-sectionally evaluated 186 children (70 with obesity and 116 with normal weight) 6–12 years old from Yucatan, Mexico. Nutritional status was defined with body mass index (BMI) percentiles. Personality traits were evaluated with the Conners and TMCQ tests; eating behavior was evaluated with the CEBQ test. Genotyping with real-time PCR and TaqMan probes was used for FTO-rs9939609, whereas PCR amplification was used for MAO-A u-VNTR.
Results: High-intensity pleasure (p = 0.013) and moderate appetite (p = 0.032) differed according to nutritional status. Heterozygous FTO-rs9939609 T/A children showed higher mean scores of low-intensity pleasure (p = 0.002) and moderate appetite (p = 0.027) than homozygous T/T. Hemizygous boys having MAO-A LTA showed significantly higher mean scores of anxiety (p = 0.001) and impulsivity (p = 0.008). In multivariate models, only LTA alleles of MAO-A explained obesity in boys (OR = 4.44; 95% CI = 1.18–16.63).
Conclusion: In the present study, MAO-A u-VNTR alleles were associated with obesity in multivariate models only in boys. These alleles might also have a role in personality traits such as anxiety and impulsivity, which secondly contribute to developing obesity in Mayan boys.
Overweight and obesity are defined as an abnormal and excessive accumulation of fat that might lead to health impairment. Fat accumulation in the early stages of development is related to later non-communicable diseases onset, such as diabetes, arterial hypertension, cardiovascular disease, and cancer (Umer et al., 2017; Hidayat et al., 2018). Thus, investigating the origin of childhood obesity would prevent morbidity in adolescents, adults, and older adults. Accordingly, some research has focused on evaluating the genetic and psychological aspects of childhood obesity, although from a non-integrative perspective.
It has been demonstrated that the psychological characteristics of children lead to behavior-related body fat excess (Flores-Dorantes et al., 2020). Such factors are configured by genetic-environment interactions that have shown paradoxical results: obese children have common personality traits that are not distinctive of a unified profile or a specific mental disease when compared with their normal weight counterparts (Shaker et al., 2014). However, obese children exhibit more significant anxiety and depressive symptoms arising from low self-esteem, distorted body image, and being victims of bullying (Lindberg et al., 2020). Taking together, anxiety, impulsivity, and the search for stimulation, reward, gratification, or novelty become the personality traits related to obesity.
On the other hand, eating behavior refers to the set of actions that underlies the relationship of the human being with food. This factor is mainly learned from parent behavior, although these patterns are associated with impulsivity, anxiety, and the search for stimulation and reward (Kininmonth et al., 2021). Moreover, the extensive study of genetic impairment has led to identifying genes related to impulsivity and searching for reward or gratification, particularly those associated with the serotoninergic and dopaminergic systems. Given the primary role of the dopaminergic rewarding process in appetite modulation, it has been shown that genetic variants of dopamine-related enzymes, namely, the fat mass and obesity-related (FTO) gene and the monoamine oxidase A (MAO-A) gene, might be associated with palatable food and, therefore, with obesity physiopathology (Fuemmeler et al., 2008; Qi et al., 2014; Dias et al., 2016; Krishnan et al., 2017). It has also been suggested that genetic variants associated with obesity partly exert their influence on BMI and waist circumference through undesirable eating behaviors such as disinhibition and susceptibility to hunger (Jacob et al., 2018).
The FTO gene is located on the locus 16q12.2 and consists of nine exons and eight introns. It is expressed in the hypothalamus, hypophysis and suprarenal glands, muscle, and adipose tissue. The latter is relevant since the FTO has been implied in fat mass regulation through the lipolysis mechanism (Jacobsson et al., 2008) and the adipocyte early differentiation process (Claussnitzer et al., 2015). The most studied variant from FTO is the SNP rs9939609 (g.87653T>A), in which carriers of the risk allele A (homozygous or heterozygous) have increased odds for obesity and higher calorie, fat, and carbohydrate intake (Daya et al., 2019; Mehrdad et al., 2020). Moreover, overweight children carrying the A allele have shown higher scores on the Food Responsiveness, Emotional Overeating, Enjoyment of Food, and Food Choice subscales and lower scores on the Satiety-Responsiveness and Slowness in Eating subscales. Obese children have shown higher scores on the Cognitive Restrained subscale and lower Food choice (Obregón Rivas et al., 2018). Minor allele frequency of FTO-rs9939609 has been reported in 16.8% of individuals from Yucatan, Mexico (Hernandez-Escalante et al., 2014).
Otherwise, monoaminoxidase (MAO) is a mitochondrial isoenzyme that catalyzes amine and neurotransmitter oxidation such as serotonin, norepinephrine, dopamine, phenylethylamine, and others (Bortolato et al., 2008). Humans produce two forms, MAO-A (mainly in catecholaminergic brain neurons) and MAO-B (mainly in brain serotoninergic and histaminergic neurons), that possess selective affinity by neurotransmitters. For example, MAO-A exhibits a higher affinity for serotonin, norepinephrine, and dopamine, whereas MAO-B is selective for phenylethylamine (Bortolato et al., 2008). The MAO-A consists of 16 exons in chromosome X (Xp11.23). It contains a functional 30 bp upstream variable number of tandem repeats (30 bp u-VNTR) located in the promoter region and consists of 2, 3, 3.5, 4, or 5 repeated copies, with the 3- (3R) and 4-repeat (4R) alleles as the most common. Given the central role of MAO-A in modulating neurotransmitters, it might be hypothesized that an impact on eating behavior, such as impulsivity, anxiety, and neuroticism, might result in obesity. In this respect, the lower activity 3R allele has been reported to be associated with obesity in Caucasian populations (Fuemmeler et al., 2008; Wallmeier et al., 2013; Dias et al., 2016); conversely, the 4R allele has been associated with higher consumption of lipid and sugar dense foods in Brazilian boys (Galvão et al., 2012).
In Mexico, 75.2% of adults (≥20 years) are overweight (39.1%) or obese (36.1%). Strikingly, 37.3% of children (5–11 years) exhibit the former (19.2%) or latter (18.1%) condition (Shamah-Levy et al., 2023). Of note is the case of Yucatan, Mexico, where 30% of children (5–11 years) are overweight or obese. This situation ranks this Mexican State among the 10 States with higher rates of these conditions (Shamah-Levy et al., 2023). Data from a national survey showed an elevated prevalence of consumption of “not recommended” food in children, mainly sugar-sweetened beverages (92.9%) (Shamah-Levy et al., 2023). Moreover, it has been reported that 87% of Mexican children and adolescents follow a dietary pattern high in fat and sugar (Galvan-Portillo et al., 2018). Even the FTO-rs9939609 has been found to be associated with obesity in Mayan girls (González-Herrera et al., 2019b). Thus, it might be hypothesized that the excessive consumption of such foods might be triggered by genetic impairment of the serotoninergic and dopaminergic pathways related to personality traits and eating behavior. Hence, the present study aimed to evaluate the genetic association between FTO-rs9939609 and MAO-A 30 bp u-VNTR variants, along with personality traits and eating behavior with obesity in Mayan children from Mexico.
School-aged children with Mayan ethnicity (6–12 years old) attending public elementary schools at Southeast, Mexico (Yucatan), were enrolled from January and June 2017. A total of 186 children participated, 52.7% were girls and 47.3% were boys. We included 116 children with normal weight and 70 children with obesity according to the World Health Organization (WHO) growth charts for children aged 5–19 years (De Onis et al., 2007).
All children were measured for weight with a digital scale (Seca, Hamburg, Germany) and height with a portable stadiometer (Seca 225, Hamburg, Germany) in a standing position without shoes. BMI-for-age was obtained from the WHO growth reference data tables for 5–19 years, and then it was z-scored for standard deviation (SD) calculation. Normal-weight infants were those with a BMI-for-age between 15th and 84th percentiles (BMIpc) or between −1 and < +1 SD, whereas infants with obesity were those with a BMI-for-age >97th percentile or ≥ +2 SD (De Onis et al., 2007).
Children with chronic diseases, monogenic obesity, as well as children with a BMI-for-age between +1 SD and +2 SD (overweight) or less than −1 SD (underweight) were excluded. Selected children were born in Yucatan, having familial ancestors back for at least the third generation born in Yucatan. Children with Mayan ethnicity were selected using anthropological and demographic parameters, such as language, birthplace, surnames, and genealogy, to match ethnically all children. We previously reported the genetic ancestry of the population from the Yucatan Peninsula, where Mayan is the main Amerindian genetic background (González-Herrera et al., 2019a; Lara-Riegos et al., 2020).
This study was conducted according to the Declaration of Helsinki and its later amendments. It was approved by the Institutional Review Board of Bioethics of the Centro de Investigaciones Regionales “Dr. Hideyo Noguchi,” Universidad Autónoma de Yucatán, Mexico (CEI/03/2015). Patients’ personal information was kept strictly confidential, and only the principal investigator could access it; parents of selected children signed informed consent. However, they had to provide assent for the venipuncture procedure.
Personality traits and eating behavior in children were evaluated from parents’ perspective. Questionnaires were answered by 86.6% of mothers and 13.4% of fathers. Impulsivity and anxiety were evaluated with a rigorous selection of items from the Conners test (Keith Conners et al., 1998), whereas low (LIP) and high (HIP) intensity pleasure were measured with some items from the Temperament in Middle Childhood Questionnaire (TMCQ) (Rothbart et al., 2001).
Eating behavior was evaluated through the Child Eating Behavior Questionnaire (CEBQ), translated and validated by our research group for its application in the Mexican population (Vázquez-Pérez et al., 2020). The adapted version included 34 items for six dimensions: 1) food responsiveness (FR) and emotional overeating (EOE) were condensed into the new “voracity and emotional intake (VEI)” category; 2) satiety responsiveness (SR) and slowness in eating (SE) were condensed into the new “moderate appetite (MA)”; 3) enjoyment of food (EF); 4) desire to drink (DD); 5) emotional undereating (EUE); and 6) food fussiness (FF).
Genomic DNA was extracted from peripheral whole blood using a conventional in-house method. The FTO-rs9939609 was genotyped using the TaqMan® Allelic Discrimination Assay (Applied Biosystems, Foster City, CA, United States) C_30090620_10 in a real-time PCR StepOne device (Applied Biosystems) following supplier specifications. All PCR reactions were done in 48-well plates with a final volume of 10 μL. The allelic discrimination was done with the StepOne v.2.1 software.
The MAO-A 30 bp u-VNTR was evaluated using the Lung et al. method (Lung et al., 2011). Briefly, the PCR reaction was performed, according to manufacturer instructions, in a final volume of 25 μL with the Dream Taq Green PCR Master Mix (Thermo Scientific) and the forward (5′-ACAGCCTCGCCGTGGAGAAG-3′) and reverse (5′-GAACGGACGCTCCATTCGGA-3′) primers. The PCR products were visualized in 10% acrylamide gels stained with silver nitrate. With this method, the sizes for the 2R, 3R, 4R, and 5R allelic variants were 320 bp, 350 bp, 380 bp, and 410 bp, respectively.
FTO-rs9939609 was genotyped in 186 DNA samples, whereas MAO-A 30 bp u-VNTR was genotyped in 134 samples due to DNA quality.
Genotypic and allelic frequencies of FTO-rs9939609 and MAO-A u-VNTR polymorphisms were stratified by sex and BMI status and described with frequencies and percentages. Otherwise, the MAO-A 30 bp u-VNTR was stratified according to its associated transcriptional activity: the hemizygous boys with 3R and 4R alleles were included in the low (LTA) or high (HTA) transcriptional activity group, respectively. Girls with the 3R/3R genotype were considered the LTA, whereas the 3R/4R and 4R/4R genotypes were HTA. The Hardy-Weinberg equilibrium (HWE) was analyzed with a χ2 test at p > 0.05.
Personality traits and eating behavior scores were obtained from parent recalls and are presented as mean ± standard deviation (SD). These mean ± SD scores were compared between BMI status (obese vs. normal weight) and MAO-A 30 bp u-VNTR with Student’s t-test at p < 0.05; one-way ANOVA with Tukey’s post-hoc test at p < 0.05 was employed for comparisons between FTO-rs9939609 genotypes. Frequency comparisons were done with a χ2 test at p < 0.05.
Finally, the combined effect of sex, age, genetic variants, personality traits, and eating behavior on BMI status was evaluated in a multivariate logistic regression model in which odds ratio (OR) and 95% confidence intervals (95% CI) at p < 0.05 were calculated.
Anthropometric description of the studied group according to BMI status is shown on Table 1. The mean ± SD age and BMIpc were 8.69 ± 1.4 years and 72.59 ± 27.83, respectively. BMIpc and weight significantly differed between obese and normal-weight infants (p < 0.001). Accordingly, 58.57% of boys and 41.43% of girls were obese. There was a significant difference for the frequency of obesity between boys and girls (p = 0.017), being the girls with a higher frequency of normal weight [N = 69 (59.48%)].
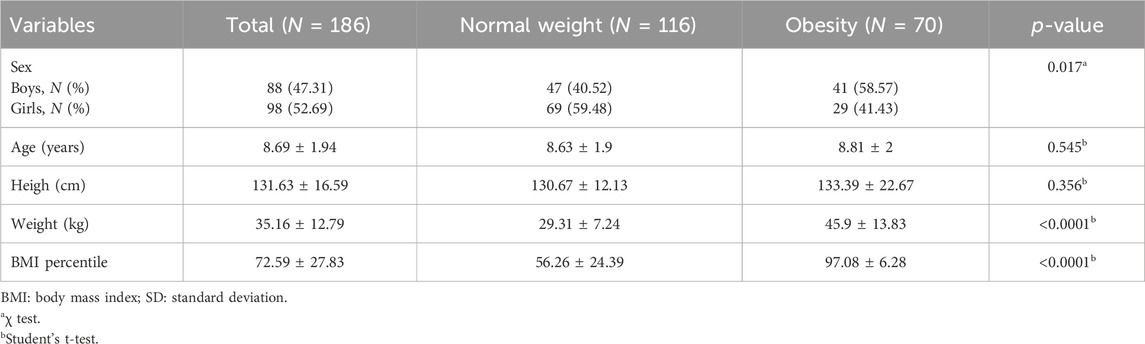
Table 1. Anthropometric variables according to BMI status. Mean ± SD.
The genotypic and allelic frequencies of FTO-rs9939609 according to sex are shown in Supplementary Table S1. The T/T genotype and the T allele were prevalent among the general sample; the polymorphism was not in HWE (p = 0.04), particularly in girls (p = 0.001). It has been reported that when the HWE deviations cannot be attributed to genotyping errors, selection, or non-random mating and may be caused by an unknown factor, the χ2 test for trend (Armitage trend test) should be investigated to reduce the chance of false-positive associations. Hence, after conducting the Armitage test, no deviations from the HWE were observed (p ≥ 0.93) (Alnafjan et al., 2022). Table 2 depicts the distribution of genotypic and allelic frequencies of FTO-rs9939609 according to sex and BMI status. There were no significant differences in genotypes/alleles according to BMI in girls, boys, or the total sample (p ≥ 0.11).
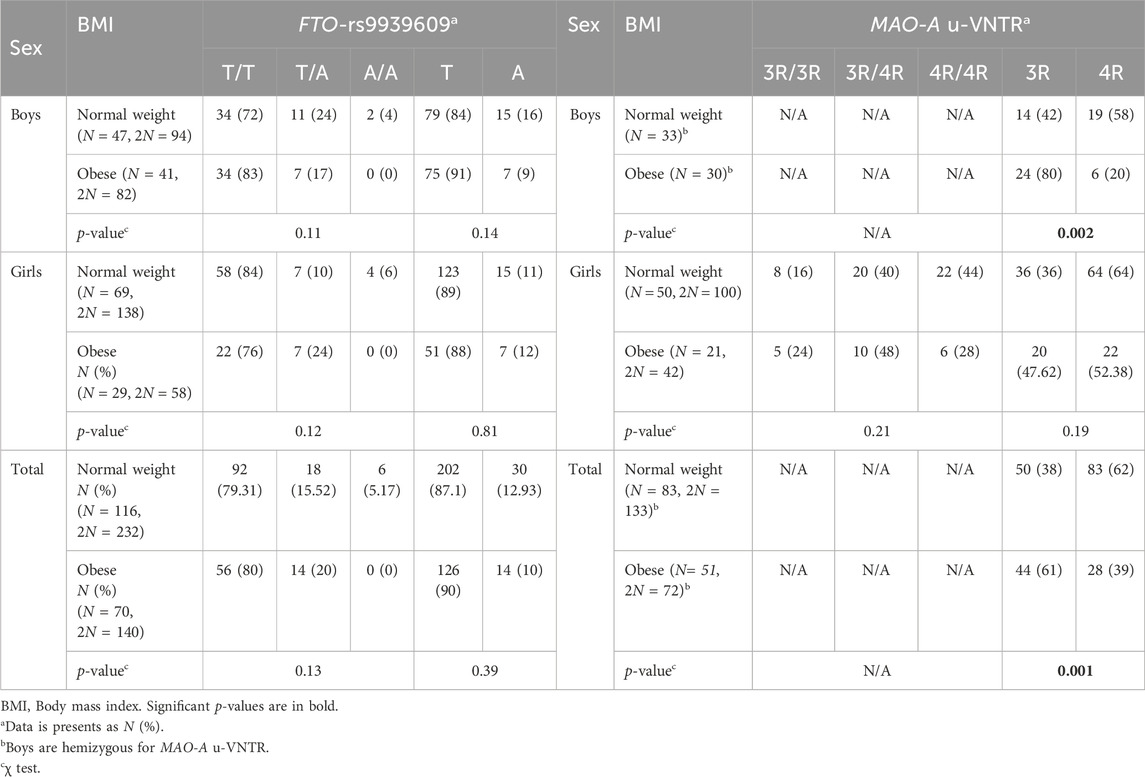
Table 2. FTO-rs9939609 and MAO-A u-VNTR according to sex and BMI status.
Regarding the MAO-A u-VNTR, the most frequent genotype was the heterozygous 3R/4R (42.25%), and was in HWE (p = 0.33) (Supplementary Table S1). After stratifying by BMI status (Table 2), hemizygous boys with the 3R allele, which conferred the LTA, showed the highest prevalence of obesity (p = 0.002). These results were consistent in the total sample (p = 0.001) but not in girls (p ≥ 0.19). In terms of odds, boys carrying the 3R allele were 5.43 times more likely to be obese than those with the 4R allele (OR: 5.43, 95%CI: 1.75–16.81).
We did not find statistically significant differences in mean scores of personality traits and eating behavior according to BMI status, except for items of HIP and MA (Table 3). Children with normal weight obtained significantly higher mean scores than children with obesity for HIP (p = 0.013) and MA (p = 0.032), suggesting an eating behavior compatible with adequate satiety in non-obese children.
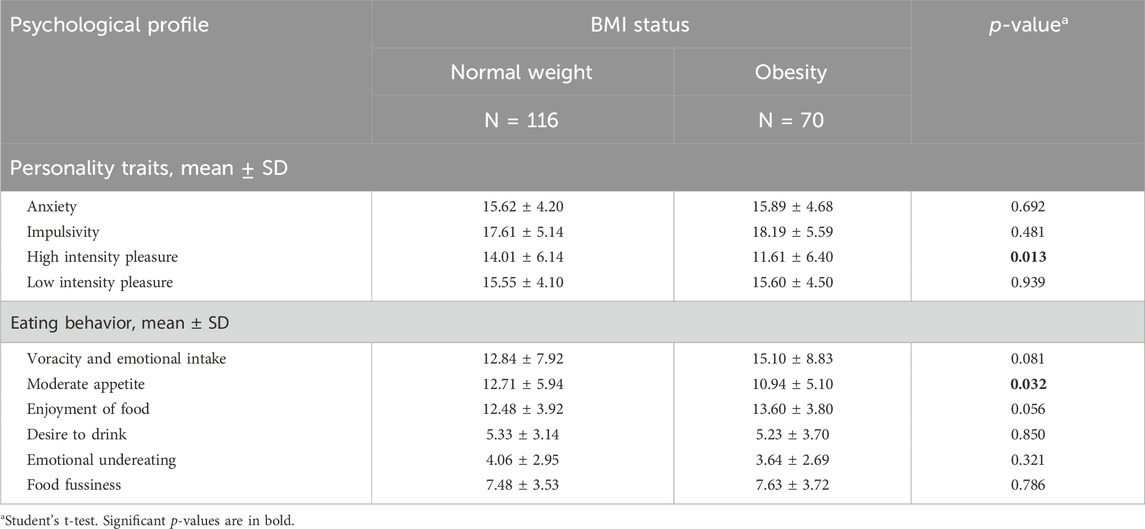
Table 3. Personality traits and eating behavior scores according to BMI status.
Personality traits and eating behavior according to FTO-rs9939609 genotyping are shown in Supplementary Table S2. We found that children with heterozygous T/A genotype showed significantly higher mean scores in LIP (p = 0.002) and moderate appetite (p = 0.027) than homozygous T/T children. Homozygous A/A FTO-rs9939609 children showed the highest or the lowest score for all studied personality traits and eating behaviors, except EF; however, no significant difference was found compared with T/T wildtype FTO children (p > 0.05), which may be due to the low frequency of homozygous A/A in the studied population.
Data regarding MAO-A 30 bp u-VNTR and psychological features are shown in Supplementary Table S3. For the total population, only impulsivity was significantly higher in children with LTA alleles compared to the HTA (p = 0.027), suggesting that children carrying LTA alleles might develop a more impulsive personality. After stratifying by sex, girls did not exhibit any significant difference in mean scores of any personality trait nor any eating behavior after comparing HTA vs. LTA alleles (p > 0.05); the same was seen for girls with obesity (Supplementary Table S4) or normal weight (Supplementary Table S5). Otherwise, boys with LTA alleles exhibited significantly higher mean scores of anxiety (p = 0.001) and impulsivity (p = 0.008) than boys with HTA alleles, suggesting that LTA alleles might affect these personality traits only in boys (Supplementary Table S3). After stratifying by BMI status, only boys with obesity having LTA alleles showed significantly higher mean scores of the personality traits, impulsivity (p = 0.026), EUE (p = 0.002), and FF (p = 0.022) than boys with obesity having HTA alleles (Supplementary Table S4). Noticeably, boys with normal weight did not show significant differences in mean scores of any personal trait nor any eating behavior (p > 0.05), neither girls with normal weight nor girls with obesity for MAO-A 30 bp u-VNTR (Supplementary Tables S4, S5).
The multivariate binary logistic regression model included those variables that showed significant differences during the analysis, namely, sex, age, HIP and LIP, impulsivity, MA, and MAO-A 30 bp u-VNTR transcriptional activity. The FTO-rs9939609 was also included based on its reported clinical relevance (Table 4). The stratified multivariate model suggested that boys having LTA alleles exhibited four times higher odds of being obese compared with HTA of MAO-A 30 bp u-VNTR (OR = 4.44; 95% CI = 1.18–16.63; p = 0.027).
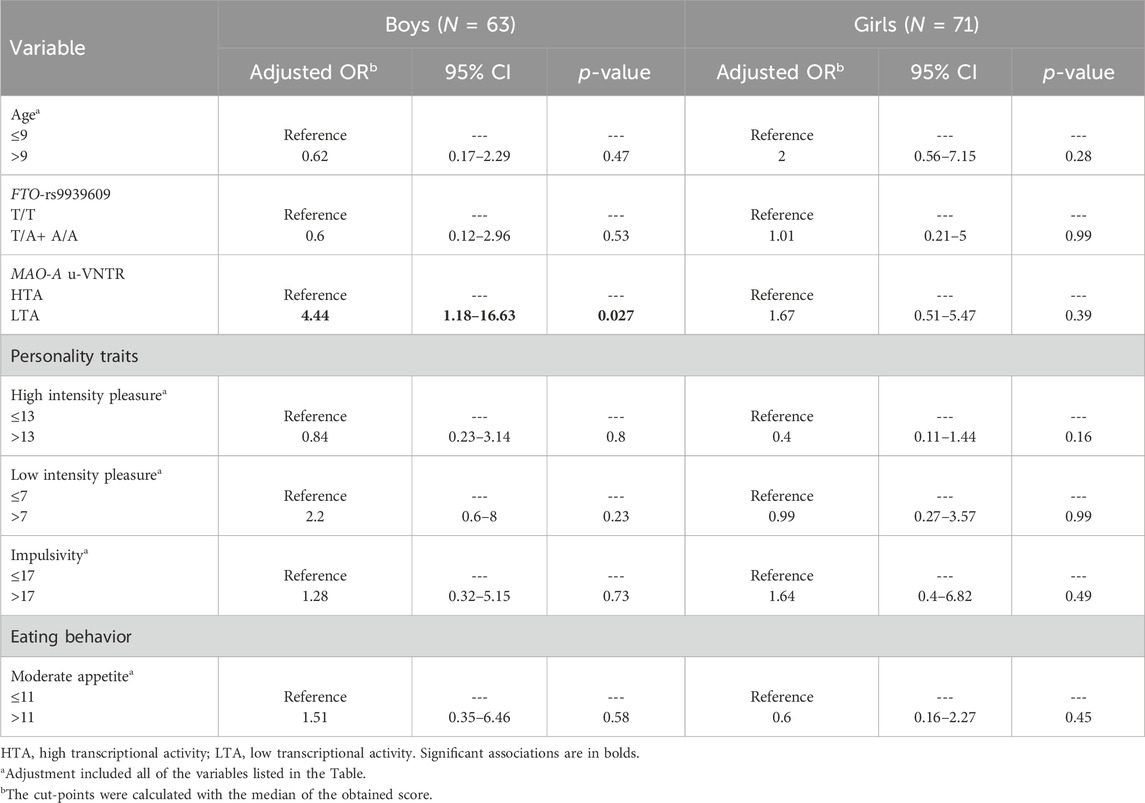
Table 4. Multivariate binary logistic regression for obesity status stratified by sex.
The present study evaluated the association of the genetic variants FTO-rs9939609 and MAO-A 30 bp u-VNTR, along with eating behavior and personality traits with obesity in Mayan children from Mexico. We found significantly higher mean scores in the personality trait HIP and in the eating behavior MA in normal-weight children vs. obese. Heterozygous children T/A of FTO-rs9939609 exhibited significantly lower LIP (personality trait) and MA (eating behavior) scores than children with wild-type FTO. Additionally, only boys with LTA alleles of MAO-A 30 bp u-VNTR showed higher mean scores for the personality traits of impulsivity and anxiety. Moreover, only obese boys with LTA alleles showed significantly higher mean scores of impulsivity, as well as for the eating behaviors of EUE and FF. However, in multivariate analysis, only the LTA alleles of the MAO-A u-VNTR were associated with obesity, particularly in boys of this Mayan children population.
By 2015, the accumulated evidence gave birth to the behavioral susceptibility theory (BST), which proposes that “obesity-related genes” influence weight gain partly through their effects on appetite and that variations in appetite emerge in the latter stages of life (Llewellyn and Wardle, 2015). The behavioral component of the BST is evaluated through the CEBQ. This instrument assesses food approach traits [VEI (FR and EOE), EF, and DD] and food avoidance traits [MA (SR and SE), FF and EUE) in our local validation of the instrument. In this respect, some of the CEBQ traits have shown to be positively (EF and FR) or negatively (SR and FF) associated with BMI or BMI z-scores (WHO or CDC) (Sánchez et al., 2016; Boswell et al., 2018; Pesch et al., 2018). Such information has also been synthesized in a recent comprehensive meta-analysis that found higher CEBQ food approach trait scores and lower food-avoidant trait scores in obese children (Kininmonth et al., 2021).
Conversely, a few reports show no association between obesity (measured as BMI) and eating behavior (Hayes et al., 2016; Lora et al., 2016). Our results partially agree with these findings since one of the six CEBQ dimensions differed between obese and normal-weight children, with MA showing higher scores in children with normal weight. As expected, parents of children with normal weights describe them as having more moderate eating habits, which might contribute to their healthy weight maintenance.
Otherwise, independently of BMI status, the MA mean score was higher in wildtype T/T children than T/A heterozygous for FTO-rs9939609, suggesting the MA as an eating behavior consistent with adequate satiety in children with wildtype FTO. The MA score is lower in those carrying one FTO risk allele, as in the T/A genotype, implying a lower threshold for satiety in heterozygous children (according to the parent’s testimony). Taking into account that MA in our validated CEBQ questionnaire estimates SR, other studies, such as that of Obregon-Rivas et al., 2018 (Obregón Rivas et al., 2018), demonstrated association of the FTO-rs9939609 gene with eating behavior traits, dependent of sex and nutritional status in Chilean children: normal weight girls carriers of the A allele showed higher SR and SE scores; normal weight boy carriers of the A allele showed higher scores on the negative affect subscale; overweight boys with the A allele showed higher FR, EOE, and lower SR and SE. In contrast, in preschool children from the Generation R study, it was found that the minor allele A leads to increased food responsiveness and better emotional control (Velders et al., 2012).
In adults, the FTO-rs9939609 has been associated with poorer eating behavior in an age-dependent fashion. For example, it has been reported a higher decline of emotional eating with age in the A/A+ T/A genotype group compared to the T/T group (Abdella et al., 2019). The FTO gene variation (i.e., rs9939609 A/A+ T/A vs. T/T) has been associated with increased dietary intake from unhealthy eating behaviors such as sweet, fat, and carbohydrate cravings from fast food (Abdella et al., 2019). Also, FTO A allele has also been associated with increased severity of eating disorders like binge eating behavior or psychopathological features, including emotional eating and disorder of corporeality (Castellini et al., 2017). In Mexican young adults (20.14 ± 3.95 years) from Guadalajara, Mexico, the FTO-rs9939609 genotype T/T was associated only with higher EUE in bivariate models (OR: 1.8; 95% CI: 1.1–9.1; p = 0.014) (Rivera-Iñiguez et al., 2024). In the same Mexican State, volunteers aged 18–25 years carrying the FTO-rs9939609T/A + A/A genotypes showed a higher consumption of products with added sugars than those participants with the T/T genotype (Madrigal-Juarez et al., 2023). In the context of metabolic syndrome components, a study in women from Mayan communities of Chiapas, Mexico, found an association of hyperglycemia with FTO-rs9939609 in the dominant model (T/T vs. T/A + A/A; ORadjusted: 2.6; 95% CI: 1.3–5.3; p = 0.007) (Núñez-Ortega et al., 2021).
The effect of FTO genotypes on psychological aspects might be modified by vitamin D intake; insufficient vitamin D intake might be an additional contributor to the effect of FTO genotypes on eating behaviors and mental health (Mehrdad et al., 2022). One pathway of the FTO effect on adiposity is by influencing dopamine signaling, given that FTO variants associated with obesity are also associated with several behaviors or disorders dependent on dopamine. For example, FTO knockout in mice has been shown to impair control of dopamine receptors D2 of neuronal activation and DA-dependent-regulation of locomotor activity and reward sensitivity (Sun et al., 2017). A neuro-image study showed that carriers of FTO variants have reduced longitudinal functioning in the medial prefrontal cortex, consistent with higher rates of impulsivity, behavioral disinhibition, and risky decision-making (Chuang et al., 2015). In this study, LIP (personality trait) differed according to FTO-rs9939609 genotypes, observing higher scores in T/T than in heterozygous T/A.
The abovementioned is striking since the higher LIP score suggests high sensitivity to the enjoyment of situations related to low stimuli, complexity, and novelty, suggesting that children with the A allele with lower LIP scores need more stimuli to get the optimal satiety threshold. Prediction analysis in silico has suggested that FTO risk variants could directly affect modulating binding sites of transcription factors as a regulatory function during brain development (Saucedo-Uribe et al., 2019). Even so, it is suggested that FTO in the basal ganglia regulates a circuit that steers movements and regulates reactions to novelty, exploration, motor function, and timing behavior via D2R medium spiny neurons. However, these changes do not predispose to weight gain or altered food reward in mice (Ruud et al., 2019). The latter might explain our finding on the FTO contribution independently of BMI status.
Notably, the FTO-rs9939609 was not associated (in uni- and multivariate models) with obesity in the studied children. A meta-analysis found 37% higher odds of being obese children in the group of the allele of risk A compared with the wild type T (OR = 1.37; 95% CI = 1.25–1.51; p ≤ 0.001). Such a result was still significative after stratifying by Caucasian (OR = 1.37; 95% CI = 1.18–1.6; p ≤ 0.001) or Asian (OR = 1.34; 95% CI = 1.25–1.43; p ≤ 0.001) ethnicity (Dastgheib et al., 2021). However, the association did not remain in the mixed population group (OR = 1.23; 95% CI = 0.77–1.95; p = 0.38). The latter was observed for Brazilian and Chilean children, two Latin-American populations that share a genetic admixture background with Mexico. Some studies in the Mexican population also found no association of BMI or other indicators of obesity [i.e., waist circumference, body fat (%), and energy intake] with the risk allele A from FTO-rs9939609 (Villalobos-Comparán et al., 2017; Chama-Avilés et al., 2023). In Yucatan, Mexico, our group previously reported the univariate association of obesity with the presence of the heterozygous AT and the risk allele A only in girls (González-Herrera et al., 2019b). The lack of association in the present sample of children might be due to limitations on sample size, given that none of the obese children carried the A/A genotype, whereas 5% of normal-weight cases did. However, it has been demonstrated that the association between FTO variation and obesity is sex-dependent. For example, the rs1421085 and rs9939609 affect genetic susceptibility for obesity only in girls, whereas rs8057044 and CNV are associated with overweight status only in boys. Furthermore, the SNP rs1421085 of the FTO gene was strongly associated with obesity in Mayan children instead of the rs9939609 (González-Herrera et al., 2019b).
A careful inspection of the evidence related to the BST allows the identification of the genetic predisposition as an unmodifiable factor for obesity. In this way, some polymorphisms in genes involved in the dopaminergic pathway, such as the 30 bp u-VNTR of the MAO-A, have shown consistent associations. The MAO-A is found on chromosome Xp11.3, thus generating heterozygous or homozygous females and hemizygous males. Further research showed that 3R and 4R alleles conferred low (LTA) and high (HTA) transcriptional activity, respectively. A report on alcoholic males showed significantly higher BMI in those cases with LTA than in those with HTA (Ducci et al., 2006). Also, a study on Chinese adolescents found that males with obesity had 1.85 times the odds of being in the LTA group compared with those in the HTA (OR = 1.85; 95% CI = 1.18–2.94; p = 0008) (Fuemmeler et al., 2008). Our findings agree with these previous reports since boys with LTA alleles exhibited four times the odds of being obese than boys with HTA alleles, suggesting the role of the dopaminergic pathway through the MAO-A gene in obesity pathogenesis, particularly in hemizygous boys. Human MAO-A is strongly expressed in adipocytes, where the non-neuronal transporter SLC22A3 allows the transportation and degradation of catecholamines. It has also been suggested that adipocytes are involved in establishing adrenergic tone and controlling thermogenic activity in humans since inhibition of MAOA can induce human adipocyte browning in vivo (Solivan-Rivera et al., 2022). MAO-A is the predominant isoform in human abdominal adipose and vascular tissues; it is overexpressed during inflammation and contributes to endothelial dysfunction (Sturza et al., 2019).
In the present study, we found significant differences in some personality traits, namely, the HIP and LIP, and impulsivity according to BMI, FTO-rs9939609, and MAO-A 30 bp u-VNTR. However, in multivariate logistic regression models, neither the FTO-rs9939609 nor the personality traits were associated with obesity. In this regard, two reports in large Dutch cohorts of children found no linear association between BMI z-scores and impulsivity (from TMCQ) among non- and overweight participants (Scholten et al., 2014; Sleddens et al., 2016). As a qualitative contribution of the present study, we observed that girls’ parents described their daughters as “audacious” and active seekers of intense, long-lasting, incongruent, and novel sensations related to pleasure or enjoyment. A possible explanation is that all girls look for stimulation, although normal-weight ones look outside their bodies. In contrast, obese ones look inside their bodies, using food as a vehicle for such stimuli.
On the other hand, we found significantly higher impulsivity scores in boys with MAO-A 30 bp u-VNTR LTA than those in the HTA group. Although it might be hypothesized that obese boys are impulsive and eat by anxiety rather than appetite, none of these “pre-conceived” ideas were corroborated in multivariate models. This finding agrees with previous reports in which boys with higher neuroticism (where anxiety and impulsivity are contained) showed higher consumption of sweetened beverages and low consumption of fruits and vegetables (Vollrath et al., 2012). It also might be hypothesized that parent beliefs regarding the “natural impulsivity” of boys promote weight gain unconsciously. Finally, we cannot discard additional unexplored factors mediating between the MAO-A 30 bp u-VNTR LTA and obesity. For example, a recent study found a hindbrain dopaminergic circuit in the caudal ventral tegmental area innervating DRD1-expressing neurons within the lateral parabrachial nucleus that regulates satiety and meal structure (Han et al., 2021). Therefore, mutations in such a critical circuit might explain the phenotype related to weight excess. The MAO-A 30 pb VNTR associated with LTA might operate differently in boys compared to girls, or there is also the possibility of different upbringings for boys and girls. This upbringing would consist of what food refers to and what is taught and allowed. In men, there may be a tendency toward greater indulgence and care. Conversely, women might receive better nutritional education, as excess weight and obesity are less tolerated in them compared to men. However, further studies and evidence are necessary to fully understand these differences.
To our knowledge, the present study is the first to evaluate the association of the genetic variants of FTO and MAO-A, along with eating behavior and personality traits with childhood obesity in Mexico. Also, some residual confusion generated by overweight children was ruled out during the design stage. Nevertheless, some limitations must be acknowledged. Firstly, the psychological tests employed to evaluate personality traits and eating behavior in this sample of children might be influenced by parental perceptions, including gender stereotypes or the inadvertent normalization of certain behaviors. Secondly, the sample size might have influenced the low prevalence of the A/A genotype of FTO-rs9939609 and the sex-stratified analyses. Thirdly, although the nutritional perspective was indirectly evaluated through the evaluated tests, a more integrative approach should evaluate at least 24-h dietary records. Finally, the study’s observational nature makes it impossible to draw causality-related conclusions, although some associations persisted even in multivariate models. Despite these limitations, and given the instrument’s inherent limitations related to parent perception, future analyses in a larger sample are required to incorporate eating behavior as an explicative obesity variable.
In the present study, some dimensions of the personality traits and eating behavior tests differed by sex, nutritional status, FTO-rs9939609, and MAO-A 30 bp u-VNTR. However, in multivariate analysis, only the latter retained its association with obesity in boys. Childhood obesity still represents a complex phenomenon that should be evaluated, at least from the psychological, genetic, nutritional, sex, and sociocultural perspectives. Moreover, the findings support that the factors associated with childhood obesity in a determined population might not necessarily be explicative in others.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The studies involving humans were approved by Institutional Review Board of Bioethics of the Centro de Investigaciones Regionales “Dr. Hideyo Noguchi,” Universidad Autónoma de Yucatán, Mexico (CEI/03/2015). The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent for participation in this study was provided by the participants’ legal guardians/next of kin.
LV-P: Conceptualization, Formal Analysis, Investigation, Writing–review and editing. MH-H: Supervision, Writing–review and editing. A-GA-S: Methodology, Supervision, Writing–review and editing. GP-M: Methodology, Supervision, Writing–review and editing. R-R-C: Data curation, Supervision, Writing–review and editing. JR-M: Data curation, Software, Writing–original draft, Writing–review and editing. DP-E: Methodology, Supervision, Writing–review and editing. TC-C: Visualization, Writing–review and editing. L-G-H: Conceptualization, Funding acquisition, Project administration, Writing–review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was funded by the Fondo Sectorial en Salud of the CONAHCYT in Mexico (No. FONSEC-SALUD-2012-180512), and by institutional funds from the Universidad Autónoma de Yucatán (UADY).
The authors thank the children, their parents, and elementary school teachers of Merida for their willingness to participate in the study. Special thanks are due to the students from Psychology and Nutrition who contributed to the questionnaire application and provided logistic support. V-PL received a graduate student fellowship from the Consejo Nacional de Humanidades, Ciencias y Tecnologías (CONAHCYT), Mexico.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2024.1421870/full#supplementary-material
Abdella, H. M., Farssi, H. O. E., Broom, D. R., Hadden, D. A., and Dalton, C. F. (2019). Eating behaviours and food cravings; influence of age, sex, BMI and FTO genotype. Nutrients 11, 377. doi:10.3390/NU11020377
Alnafjan, A. A., Alkhuriji, A. F., Alobaid, H. M., Babay, Z. A., and Khalil, M. I. (2022). Association of FTO gene variants rs9939609 and rs1421085 with polycystic ovary syndrome. Egypt. J. Med. Hum. Genet. 23, 45–48. doi:10.1186/s43042-022-00263-3
Bortolato, M., Chen, K., and Shih, J. C. (2008). Monoamine oxidase inactivation: from pathophysiology to therapeutics. Adv. Drug Deliv. Rev. 60, 1527–1533. doi:10.1016/J.ADDR.2008.06.002
Boswell, N., Byrne, R., and Davies, P. S. W. (2018). Eating behavior traits associated with demographic variables and implications for obesity outcomes in early childhood. Appetite 120, 482–490. doi:10.1016/J.APPET.2017.10.012
Castellini, G., Franzago, M., Bagnoli, S., Lelli, L., Balsamo, M., Mancini, M., et al. (2017). Fat mass and obesity-associated gene (FTO) is associated to eating disorders susceptibility and moderates the expression of psychopathological traits. PLoS One 12, e0173560. doi:10.1371/JOURNAL.PONE.0173560
Chama-Avilés, A., Flores-Viveros, K. L., Cabrera-Ayala, J. A., Aguilar-Galarza, A., García-Muñoz, W., Haddad-Talancón, L., et al. (2023). Identification and association of single nucleotide polymorphisms of the FTO gene with indicators of overweight and obesity in a young Mexican population. Genes. (Basel) 14, 159. doi:10.3390/genes14010159
Chuang, Y. F., Tanaka, T., Beason-Held, L. L., An, Y., Terracciano, A., Sutin, A. R., et al. (2015). FTO genotype and aging: pleiotropic longitudinal effects on adiposity, brain function, impulsivity and diet. Mol. Psychiatry 20, 133–139. doi:10.1038/MP.2014.49
Claussnitzer, M., Dankel, S. N., Kim, K.-H., Quon, G., Meuleman, W., Haugen, C., et al. (2015). FTO obesity variant circuitry and adipocyte browning in humans. N. Engl. J. Med. 373, 895–907. doi:10.1056/NEJMOA1502214
Dastgheib, S. A., Bahrami, R., Setayesh, S., Salari, S., Mirjalili, S. R., Noorishadkam, M., et al. (2021). Evidence from a meta-analysis for association of MC4R rs17782313 and FTO rs9939609 polymorphisms with susceptibility to obesity in children. Diabetes Metab. Syndr. 15, 102234. doi:10.1016/J.DSX.2021.102234
Daya, M., Pujianto, D. A., Witjaksono, F., Priliani, L., Susanto, J., Lukito, W., et al. (2019). Obesity risk and preference for high dietary fat intake are determined by FTO rs9939609 gene polymorphism in selected Indonesian adults. Asia Pac. J. Clin. Nutr. 28, 183–191. doi:10.6133/APJCN.201903_28(1).0024
De Onis, M., Onyango, A. W., Borghi, E., Siyam, A., Nishida, C., and Siekmann, J. (2007). Development of a WHO growth reference for school-aged children and adolescents. Bull. World Health Organ. 85, 660–667. doi:10.2471/BLT.07.043497
Dias, H., Muc, M., Padez, C., and Manco, L. (2016). Association of polymorphisms in 5-HTT (SLC6A4) and MAOA genes with measures of obesity in young adults of Portuguese origin. Arch. Physiol. Biochem. 122, 8–13. doi:10.3109/13813455.2015.1111390
Ducci, F., Newman, T. K., Funt, S., Brown, G. L., Virkkunen, M., and Goldman, D. (2006). A functional polymorphism in the MAOA gene promoter (MAOA-LPR) predicts central dopamine function and body mass index. Mol. Psychiatry 119 (11), 858–866. doi:10.1038/sj.mp.4001856
Flores-Dorantes, M. T., Díaz-López, Y. E., and Gutiérrez-Aguilar, R. (2020). Environment and gene association with obesity and their impact on neurodegenerative and neurodevelopmental diseases. Front. Neurosci. 14, 863. doi:10.3389/FNINS.2020.00863
Fuemmeler, B. F., Agurs-Collins, T. D., McClernon, F. J., Kollins, S. H., Kail, M. E., Bergen, A. W., et al. (2008). Genes implicated in serotonergic and dopaminergic functioning predict BMI categories. Obes. (Silver Spring) 16, 348–355. doi:10.1038/OBY.2007.65
Galvan-Portillo, M., Sánchez, E., Cárdenas-Cárdenas, L. M., Karam, R., Claudio, L., Cruz, M., et al. (2018). Dietary patterns in Mexican children and adolescents: characterization and relation with socioeconomic and home environment factors. Appetite 121, 275–284. doi:10.1016/J.APPET.2017.11.088
Galvão, A. C. S., Krüger, R. C., Campagnolo, P. D. B., Mattevi, V. S., Vitolo, M. R., and Almeida, S. (2012). Association of MAOA and COMT gene polymorphisms with palatable food intake in children. J. Nutr. Biochem. 23, 272–277. doi:10.1016/J.JNUTBIO.2010.12.004
González-Herrera, L., Sosa-Escalante, J. E., López-González, P., López-González, M. J., Gamboa-Magaña, R. Y., Herrera-Diaz, R. G., et al. (2019a). Ancestral proportions based on 22 autosomal STRs of an admixed population (Mestizos) from the Península of Yucatán, México. Forensic Sci. Int. Genet. Suppl. Ser. 7, 429–431. doi:10.1016/J.FSIGSS.2019.10.039
González-Herrera, L., Zavala-Castro, J., Ayala-Cáceres, C., Pérez-Mendoza, G., López-González, M. J., Pinto-Escalante, D., et al. (2019b). Genetic variation of FTO: rs1421085 T>C, rs8057044 G>A, rs9939609 T>A, and copy number (CNV) in Mexican Mayan school-aged children with obesity/overweight and with normal weight. Am. J. Hum. Biol. 31, e23192. doi:10.1002/AJHB.23192
Han, Y., Xia, G., He, Y., He, Y., Farias, M., Xu, Y., et al. (2021). A hindbrain dopaminergic neural circuit prevents weight gain by reinforcing food satiation. Sci. Adv. 7, eabf8719. doi:10.1126/SCIADV.ABF8719
Hayes, J. F., Altman, M., Kolko, R. P., Balantekin, K. N., Holland, J. C., Stein, R. I., et al. (2016). Decreasing food fussiness in children with obesity leads to greater weight loss in family-based treatment. Obes. (Silver Spring) 24, 2158–2163. doi:10.1002/OBY.21622
Hernandez-Escalante, V. M., Nava-Gonzalez, E. J., Voruganti, V. S., Kent, J. W., Haack, K., Laviada-Molina, H. A., et al. (2014). Replication of obesity and diabetes-related SNP associations in individuals from Yucatán, México. Front. Genet. 5, 380. doi:10.3389/FGENE.2014.00380
Hidayat, K., Du, X., and Shi, B. M. (2018). Body fatness at a young age and risks of eight types of cancer: systematic review and meta-analysis of observational studies. Obes. Rev. 19, 1385–1394. doi:10.1111/OBR.12705
Jacob, R., Drapeau, V., Tremblay, A., Provencher, V., Bouchard, C., and Pérusse, L. (2018). The role of eating behavior traits in mediating genetic susceptibility to obesity. Am. J. Clin. Nutr. 108, 445–452. doi:10.1093/AJCN/NQY130
Jacobsson, J. A., Klovins, J., Kapa, I., Danielsson, P., Svensson, V., Ridderstråle, M., et al. (2008). Novel genetic variant in FTO influences insulin levels and insulin resistance in severely obese children and adolescents. Int. J. Obes. (Lond). 32, 1730–1735. doi:10.1038/IJO.2008.168
Keith Conners, C., Sitarenios, G., Parker, J. D. A., and Epstein, J. N. (1998). The revised Conners’ Parent Rating Scale (CPRS-R): factor structure, reliability, and criterion validity. J. Abnorm. Child. Psychol. 26, 257–268. doi:10.1023/A:1022602400621
Kininmonth, A., Smith, A., Carnell, S., Steinsbekk, S., Fildes, A., and Llewellyn, C. (2021). The association between childhood adiposity and appetite assessed using the Child Eating Behavior Questionnaire and Baby Eating Behavior Questionnaire: a systematic review and meta-analysis. Obes. Rev. 22, e13169. doi:10.1111/OBR.13169
Krishnan, M., Thompson, J. M. D., Mitchell, E. A., Murphy, R., McCowan, L. M. E., Shelling, A. N., et al. (2017). Analysis of association of gene variants with obesity traits in New Zealand European children at 6 years of age. Mol. Biosyst. 13, 1524–1533. doi:10.1039/C7MB00104E
Lara-Riegos, J., Barquera, R., Castillo-Chávez, O. del, Medina-Escobedo, C. E., Hernández-Zaragoza, D. I., Arrieta-Bolaños, E., et al. (2020). Genetic diversity of HLA system in two populations from Yucatán, Mexico: Mérida and rural Yucatán. Hum. Immunol. 81, 569–572. doi:10.1016/J.HUMIMM.2019.07.280
Lindberg, L., Hagman, E., Danielsson, P., Marcus, C., and Persson, M. (2020). Anxiety and depression in children and adolescents with obesity: a nationwide study in Sweden. BMC Med. 18, 30. doi:10.1186/S12916-020-1498-Z
Llewellyn, C., and Wardle, J. (2015). Behavioral susceptibility to obesity: gene-environment interplay in the development of weight. Physiol. Behav. 152, 494–501. doi:10.1016/J.PHYSBEH.2015.07.006
Lora, K. R., Hubbs-Tait, L., Ferris, A. M., and Wakefield, D. (2016). African-American and hispanic children’s beverage intake: differences in associations with desire to drink, fathers’ feeding practices, and weight concerns. Appetite 107, 558–567. doi:10.1016/J.APPET.2016.09.012
Lung, F. W., Tzeng, D. S., Huang, M. F., and Lee, M. B. (2011). Association of the maoa promoter uvntr polymorphism with suicide attempts in patients with major depressive disorder. BMC Med. Genet. 12, 74–11. doi:10.1186/1471-2350-12-74
Madrigal-Juarez, A., Martínez-López, E., Sanchez-Murguia, T., Magaña-De La Vega, L., Rodriguez-Echevarria, R., Sepulveda-Villegas, M., et al. (2023). FTO genotypes (rs9939609 T>A) are associated with increased added sugar intake in healthy young adults. Lifestyle genomics 16, 214–223. doi:10.1159/000534741
Mehrdad, M., Doaei, S., Gholamalizadeh, M., and Eftekhari, M. H. (2020). The association between FTO genotype with macronutrients and calorie intake in overweight adults. Lipids Health Dis. 19, 197–206. doi:10.1186/s12944-020-01372-x
Mehrdad, M., Eftekhari, M. H., Jafari, F., Nikbakht, H. A., and Gholamalizadeh, M. (2022). Associations between FTO rs9939609 polymorphism, serum vitamin D, mental health, and eating behaviors in overweight adults. Nutr. Neurosci. 25, 1889–1897. doi:10.1080/1028415X.2021.1913316
Núñez-Ortega, P. E., Meneses, M. E., Delgado-Enciso, I., Irecta-Nájera, C. A., Castro-Quezada, I., Solís-Hernández, R., et al. (2021). Association of rs9939609-FTO with metabolic syndrome components among women from Mayan communities of Chiapas, Mexico. J. Physiol. Anthropol. 40, 11. doi:10.1186/S40101-021-00259-9
Obregón Rivas, A. M., Santos, J. L., Valladares, M. A., Cameron, J., and Goldfield, G. (2018). Association of the FTO fat mass and obesity-associated gene rs9939609 polymorphism with rewarding value of food and eating behavior in Chilean children. Nutrition 54, 105–110. doi:10.1016/J.NUT.2018.03.001
Pesch, M. H., Appugliese, D. P., Miller, A. L., Rosenblum, K. L., Lumeng, J. C., and Bauer, K. W. (2018). Approaches to restrictive feeding: associations with child weight and eating behavior. Eat. Behav. 31, 74–79. doi:10.1016/J.EATBEH.2018.08.006
Qi, Q., Kilpeläinen, T. O., Downer, M. K., Tanaka, T., Smith, C. E., Sluijs, I., et al. (2014). FTO genetic variants, dietary intake and body mass index: insights from 177,330 individuals. Hum. Mol. Genet. 23, 6961–6972. doi:10.1093/HMG/DDU411
Rivera-Iñiguez, I., Hunot-Alexander, C., Sepúlveda-Villegas, M., Campos-Medina, L., and Roman, S. (2024). Relationship between energy balance and reward system gene polymorphisms and appetitive traits in young Mexican subjects. Front. Nutr. 11, 1373578. doi:10.3389/fnut.2024.1373578
Rothbart, M. K., Ahadi, S. A., Hershey, K. L., and Fisher, P. (2001). Investigations of temperament at three to seven years: the Children’s Behavior Questionnaire. Child. Dev. 72, 1394–1408. doi:10.1111/1467-8624.00355
Ruud, J., Alber, J., Tokarska, A., Engström Ruud, L., Nolte, H., Biglari, N., et al. (2019). The fat mass and obesity-associated protein (FTO) regulates locomotor responses to novelty via D2R medium spiny neurons. Cell. Rep. 27, 3182–3198. doi:10.1016/J.CELREP.2019.05.037
Sánchez, U., Weisstaub, G., Santos, J. L., Corvalán, C., and Uauy, R. (2016). GOCS cohort: children’s eating behavior scores and BMI. Eur. J. Clin. Nutr. 70, 925–928. doi:10.1038/EJCN.2016.18
Saucedo-Uribe, E., Genis-Mendoza, A. D., Díaz-Anzaldúa, A., Martínez-Magaña, J. J., Tovilla-Zarate, C. A., Juárez-Rojop, I., et al. (2019). Differential effects on neurodevelopment of FTO variants in obesity and bipolar disorder suggested by in silico prediction of functional impact: an analysis in Mexican population. Brain Behav. 9, e01249. doi:10.1002/BRB3.1249
Scholten, E. W. M., Schrijvers, C. T. M., Nederkoorn, C., Kremers, S. P. J., and Rodenburg, G. (2014). Relationship between impulsivity, snack consumption and children’s weight. PLoS One 9, e88851. doi:10.1371/JOURNAL.PONE.0088851
Shaker, N. M., Elshahawi, H. H., Rabie, M., and Hamza, R. T. (2014). Personality profile of obese children and adolescents: relation to body mass index and metabolic complications. Middle East Curr. Psychiatry 21, 193–201. doi:10.1097/01.XME.0000449842.71358.E6
Shamah-Levy, T., Gaona-Pineda, E. B., Cuevas-Nasu, L., Morales-Ruan, C., Valenzuela-Bravo, D. G., Humarán, I. M. G., et al. (2023). Prevalencias de sobrepeso y obesidad en población escolar y adolescente de México. Ensanut Continua 2020-2022. Salud Publica Mex. 65, s218–s224. doi:10.21149/14762
Sleddens, E. F. C., ten Hoor, G. A., Kok, G., and Kremers, S. P. J. (2016). Associations between parental impulsivity and child body mass index. Springerplus 5, 1422. doi:10.1186/S40064-016-3048-X
Solivan-Rivera, J., Loureiro, Z. Y., Desouza, T., Desai, A., Pallat, S., Yang, Q., et al. (2022). A neurogenic signature involving monoamine Oxidase-A controls human thermogenic adipose tissue development. Elife 11, e78945. doi:10.7554/ELIFE.78945
Sturza, A., Olariu, S., Ionică, M., Duicu, O. M., Văduva, A. O., Boia, E., et al. (2019). Monoamine oxidase is a source of oxidative stress in obese patients with chronic inflammation 1. Can. J. Physiol. Pharmacol. 97, 844–849. doi:10.1139/CJPP-2019-0028
Sun, X., Luquet, S., and Small, D. M. (2017). DRD2: bridging the genome and ingestive behavior. Trends Cogn. Sci. 21, 372–384. doi:10.1016/J.TICS.2017.03.004
Umer, A., Kelley, G. A., Cottrell, L. E., Giacobbi, P., Innes, K. E., and Lilly, C. L. (2017). Childhood obesity and adult cardiovascular disease risk factors: a systematic review with meta-analysis. BMC Public Health 17, 683. doi:10.1186/S12889-017-4691-Z
Vázquez-Pérez, L. A., González-Herrera, L., Hattori-Hara, M., and Zavala-Cervantes, A. (2020). Traducción y Validación del Children´s Eating Behavior Questionnaire (CEBQ). Cienc. Humanismo Salud 7, 1–9.
Velders, F. P., de Wit, J. E., Jansen, P. W., Jaddoe, V. W. V., Hofman, A., Verhulst, F. C., et al. (2012). FTO at rs9939609, food responsiveness, emotional control and symptoms of ADHD in preschool children. PLoS One 7, e49131. doi:10.1371/JOURNAL.PONE.0049131
Villalobos-Comparán, M., Antuna-Puente, B., Villarreal-Molina, M. T., Canizales-Quinteros, S., Velázquez-Cruz, R., León-Mimila, P., et al. (2017). Interaction between FTO rs9939609 and the Native American-origin ABCA1 rs9282541 affects BMI in the admixed Mexican population. BMC Med. Genet. 18, 46. doi:10.1186/S12881-017-0410-Y
Vollrath, M. E., Hampson, S. E., and Júlíusson, P. B. (2012). Children and eating. Personality and gender are associated with obesogenic food consumption and overweight in 6- to 12-year-olds. Appetite 58, 1113–1117. doi:10.1016/J.APPET.2012.02.056
Keywords: childhood obesity, eating behavior, personality traits, FTO-rs9939609, MAO-A 30 pb u-VNTR, Mayan children
Citation: Vázquez-Pérez LA, Hattori-Hara M, Arankowsky-Sandoval G, Pérez-Mendoza G, Rubi-Castellanos R, Rangel-Méndez JA, Pinto-Escalante D, Canto-Cetina T and González-Herrera L (2024) Association between personality traits, eating behaviors, and the genetic polymorphisms FTO-rs9939609 and MAO-A 30 bp u-VNTR with obesity in Mexican Mayan children. Front. Genet. 15:1421870. doi: 10.3389/fgene.2024.1421870
Received: 23 April 2024; Accepted: 09 July 2024;
Published: 26 July 2024.
Edited by:
Beatriz Camarena, National Institute of Psychiatry Ramon de la Fuente Muñiz (INPRFM), MexicoReviewed by:
Sonia Roman, University of Guadalajara, MexicoCopyright © 2024 Vázquez-Pérez, Hattori-Hara, Arankowsky-Sandoval, Pérez-Mendoza, Rubi-Castellanos, Rangel-Méndez, Pinto-Escalante, Canto-Cetina and González-Herrera. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lizbeth González-Herrera, bGl6YmV0aEBjb3JyZW8udWFkeS5teA==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.