 Sophie P. Ewert
Sophie P. Ewert Mirjam Knörnschild
Mirjam Knörnschild Kirsten Jung
Kirsten Jung Karl-Heinz Frommolt
Karl-Heinz Frommolt
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 27 January 2023
Sec. Conservation and Restoration Ecology
Volume 11 - 2023 | https://doi.org/10.3389/fevo.2023.995133
This article is part of the Research Topic Origin, Conservation, and Restoration of the Threatened European Grassland Ecosystem in the Anthropocene View all 16 articles
Agricultural intensification has caused decrease and fragmentation of European semi-natural dry grasslands. While a high biodiversity value of dry grasslands is acknowledged for plants and insects, locally and on landscape level, their relevance for mobile species, such as bats, is unknown. Here we investigate the use of dry grassland fragments by bats in an agriculturally intensified region in Germany and evaluate local and landscape factors influencing bat activity and assemblages. Specifically, we predicted that a combination of local dry grassland structural richness and landscape features as well as their interactions affect bat activity and foraging above dry grasslands. We also expected that these features influence compositions of local bat assemblages. We repeatedly sampled at 12 dry grassland plots with acoustic monitoring and assessed activity and foraging of bat species/sonotypes, which we grouped into guilds known for foraging in open land, at vegetation edges and in narrow spaces. We determined structural richness of the dry grassland plots in field and derived landscape features from digital landscape data. A relatively high proportion of bat species/sonotypes used dry grasslands regularly. The edge space foragers responded positively to higher local structural richness. Their dry grassland use increased when surrounding forests and woody features were less available, but they foraged more on dry grasslands closer to water bodies. Narrow space bat activity on dry grasslands decreased with less landscape connectivity. Open and narrow space foragers responded to local structural richness only in landscape context. For all bat guilds we found increased use of structurally richer dry grasslands when there was more open farmland in the surroundings. This was also the case for edge space foragers, when landscapes were more homogeneous. Lastly, with increasing structural richness, bat assemblages were more dominated by edge space foragers. We show the importance of European dry grassland fragments for the highly mobile group of bats under certain local structural and landscape compositional conditions. Our results underline the value of heterogeneous dry grassland fragments as potential stepping stones in intensively used farmland areas and contribute to evidence based decision making in dry grassland management and bat conservation.
The agricultural intensification and industrialization, starting in the second half of the 20th century, have led to significant changes in land use and ecosystems worldwide. Both belong to the major suspected causes for global biodiversity loss (Díaz et al., 2019; Raven and Wagner, 2021) and population declines of various plant, insect and animal taxa (Burns et al., 2016; Young et al., 2016). Among the drivers of these detrimental effects are homogenization and simplification of landscapes, increased patch sizes of agricultural fields, reduced landscape structures (such as hedgerows) and loss of natural and semi-natural habitats (Matson et al., 1997; Stoate et al., 2001; Tilman et al., 2001; Jongman, 2002).
Semi-natural dry grasslands are open habitats, dominated by herbaceous (vascular) plants, mosses and lichens with occasional heterogeneous shrub and tree growth. In central Europe they have been significantly reduced during the 20th century, mainly due to land conversion, abandonment and forest encroachment, leaving small remnants fragmented in agricultural areas (Poschlod and Wallis de Vries, 2002; Habel et al., 2013; Vrahnakis et al., 2013; Leuschner and Ellenberg, 2017). Dry grasslands are known for their extremely high biodiversity in special plant communities, resulting from extremely nutrient poor, dry and sun exposed local conditions (Dengler et al., 2012; Wilson et al., 2012), and a rich insect fauna (Wallis de Vries and Van Swaay, 2009; Rasran et al., 2018; Schuldt et al., 2019). Therefore, they are now protected under the EU Habitats Directive (Council Directive 92/43/EEC). Conservation management targets these characteristic dry grassland plant species communities, by aiming to reduce nutrient inflow from agriculture (e.g., buffer zones) and by the application of grazing regimes, to reduce succession of shrubs and trees (Vrahnakis et al., 2013; Leuschner and Ellenberg, 2017). Especially smaller dry grassland fragments surrounded by intensively managed land are at greatest risk of further habitat degradation and loss of associated organisms due to external environmental influences (Butaye et al., 2005; Hodgson et al., 2005). However, exactly those small fragments have shown a great importance for species richness and communities of plants (Deák et al., 2020) and insects (Tscharntke et al., 2002; Rösch et al., 2015). Such positive effects on biodiversity are known to reach beyond their borders, affecting the species communities of the surrounding landscape. Lindborg et al. (2014) for example found increased plant richness on green islands in agricultural landscapes when dry grasslands were present in the vicinity. Their presence itself, as additional habitats in crop-dominated land, facilitates occurrence of various organisms and their exchange with adjacent landscape. This has been shown for insect communities (Tscharntke et al., 2002; Duelli and Obrist, 2003) and multiple taxa of insects and plants (Zulka et al., 2014). Dry grassland fragments can thus improve overall biodiversity of the landscape by providing local suitable conditions for specific species communities, which at the same time are influenced by the surrounding landscape. While research on plants and insects on dry grasslands has been plentiful so far, other more mobile organisms from higher trophic taxa have barely been taken into account. Therefore, researchers have recommended investigating the value of dry grasslands and their management regimes for other species and highlighted the importance of multi-scale approaches including local as well as surrounding landscape factors (Söderström et al., 2001; Zulka et al., 2014; Loos et al., 2021).
European bats are highly mobile insectivorous animals, protected by the EU Habitats Directive (Council Directive 92/43/EEC). Consequences of agricultural intensification, such as habitat and connectivity losses, are among the major drivers of bat population declines (Browning et al., 2021). While mainly associated with forests and partly urban areas for roosting, bats utilize a variety of habitat types during their nightly foraging trips of several kilometers (Dietz and Kiefer, 2016). Their habitat use is thereby not only directed by insect food availability, but also by suitability of flight pathways (Kusch et al., 2004; Müller et al., 2012). The latter is defined for each bat species by their functional traits (e.g., echolocation system and wing morphology) which determine the abilities to navigate, maneuver and forage in different degrees of vegetation clutter (Aldridge and Rautenbach, 1987; Norberg and Rayner, 1987). Based on that, bat species can be categorized into three functional guilds related to their preferred commuting and foraging habitat of open space, edge structures (such as forest edges, hedgerows) or narrow space (for example inside forests; Fenton, 1990; Schnitzler and Kalko, 2001). Local clutter conditions, determined by the structural compositions of the surroundings, can thus facilitate or hinder movement of bats from functional guilds depending on their requirements. The composition of vegetation structures, such as shrubs and trees, has shown to be of influence on bat species occurrence and activity in studies in forest systems. These studies have found species-specific activity responses of bats to various horizontal and vertical structural forest stand features (Jung et al., 2012; Froidevaux et al., 2016; Blakey et al., 2017). Highest bat activity and species diversity have been recorded in heterogeneous forest plots with intermediate vegetation clutter on different height levels, offering various structural niches (Adams and Law, 2011; Hanspach et al., 2012; Jung et al., 2012; Starik et al., 2018). Hence, on dry grasslands, one would expect a positive influence of local heterogeneous shrub and tree encroachment on bat occurrence and activity. However, dry grassland fragments, especially in open and homogeneous agricultural areas, might often be isolated from other main habitats for roosting and foraging or from structural landscape features, such as hedgerows, that offer connectivity networks as sonar guidelines, with higher prey availability and as possible protection from predators (Verboom and Huitema, 1997). When commuting through these kind of landscapes, open habitat adapted bat species do not face major constraints, while others, which are adapted to commuting and hunting closer to edges or cluttered vegetation, are likely to reach a limit in how far they move through open areas (Frey-Ehrenbold et al., 2013). These sensory constrains and habitat associations underline the necessity of a multi-scale approach when investigating the use of dry grasslands by bats embedded in open agricultural landscapes. After all, while local dry grassland conditions can offer abundant resources for commuting and foraging, the features of the surrounding landscape predominantly determine the accessibility of dry grassland fragments (Boughey et al., 2011b; Kalda et al., 2015a,b; Froidevaux et al., 2022).
A landscape should enable dispersal and support of a diverse species pool by providing a variety of resources in non-crop habitats, which is however usually limited in agriculturally intensified regions (Tscharntke et al., 2005). It is therefore crucial to understand how remaining (semi-) natural habitat fragments in farmland, such as dry grassland systems, affect endangered species. The value of dry grassland fragments for highly mobile protected bats is however still unknown. Identifying which conditions at local and landscape scales are important for bats to occur and forage on dry grasslands, adds evidence to the biodiversity value of dry grassland fragments and contributes to informed decision making for dry grassland management and bat conservation efforts. Our study therefore focuses on investigating the use of dry grassland fragments by bats in context of features of local and landscape scales as well as their interactions in an agriculturally rich region in Germany. We described the local conditions of dry grasslands with structural richness provided by shrubs and trees. In addition, we evaluated features describing openness, homogeneity and connectivity of the surrounding landscape as well as the availability of other main habitat types associated with foraging and roosting of bat species. We hypothesized that a combination of local and landscape effects as well as their interactions determines the use of dry grasslands by bats. Specifically, we expected that higher local structural richness on dry grassland fragments would increase bat activity and foraging [as in forests in Jung et al. (2012)]. Furthermore, we predicted effects of landscape features [as in Heim et al. (2015)] on use of dry grasslands by bats, and anticipated particularly the edge and narrow space adapted bats to show greater accessibility limitations (Frey-Ehrenbold et al., 2013). Most importantly however, we expected that landscape features would influence the correlation of bat activity with local structural richness [such as Froidevaux et al. (2022)]. Lastly, we predicted a dominance shift of functional bat guilds in bat assemblages above dry grasslands with changes in dry grassland and landscape features [given their different habitat preferences as in Schnitzler and Kalko (2001)].
This study has been conducted on designated protected dry grassland fragments (Figure 1) in the study region North-East Brandenburg in Northeastern Germany. Brandenburg is characterized by a comparatively low human population density. It is a young glacial landscape with many wetlands. The district Uckermark (Statistical Office Berlin-Brandenburg, 2020), where the majority of dry grassland fragments from this study are located, consists of roughly 60 percent of agricultural land (mainly used for cereal grains and a low proportion for pastures). About a quarter of the landscape is covered by forest, predominantly coniferous forests, followed by deciduous and a few floodplain forests. Although there are some fragmented forest patches, the majority of forests in the study region are rather continuous and extensive. This also means that in-between, large areas are covered by open land. The dry grassland fragments are typically rather small (in this study: min. 2 ha, max. 41 ha, mean 16 ha) and sometimes long and narrow, as they have been pushed to margins of agriculturally unusable land (e.g., slopes). They are found on dry, nutrient poor and sun exposed locations, reaching sub-continental extreme microclimatic conditions. The prominent type of management to reduce shrub and tree encroachment is grazing by sheep for a short period once a year. Brandenburg carries great conservation responsibility, containing Germany’s largest proportion of xeric sand calcareous and sub-pannonic steppic grassland (Landesamt für Umwelt Brandenburg, 2022). The latter type is dominating in the study region and is characterized by an open landscape with a high number of vascular plants, mosses and lichens. The dry grassland fragments chosen for this study showed a range of cover of woody structures (e.g., shrubs, small trees) from none up to 30%. These conditions are described as excellent to mediocre, respectively, in conservation value (Landesamt für Umwelt Brandenburg, 2022) and underline the heterogeneity of dry grasslands. This is especially relevant when seen in comparison to other grassland types (pastures and meadows) in the surroundings, which are mainly used for cattle grazing and are usually very homogeneous and lacking structural richness. Our dry grassland fragments differed not only in structural compositions but also in embedment in the landscape (regarding for example distances to other habitats).
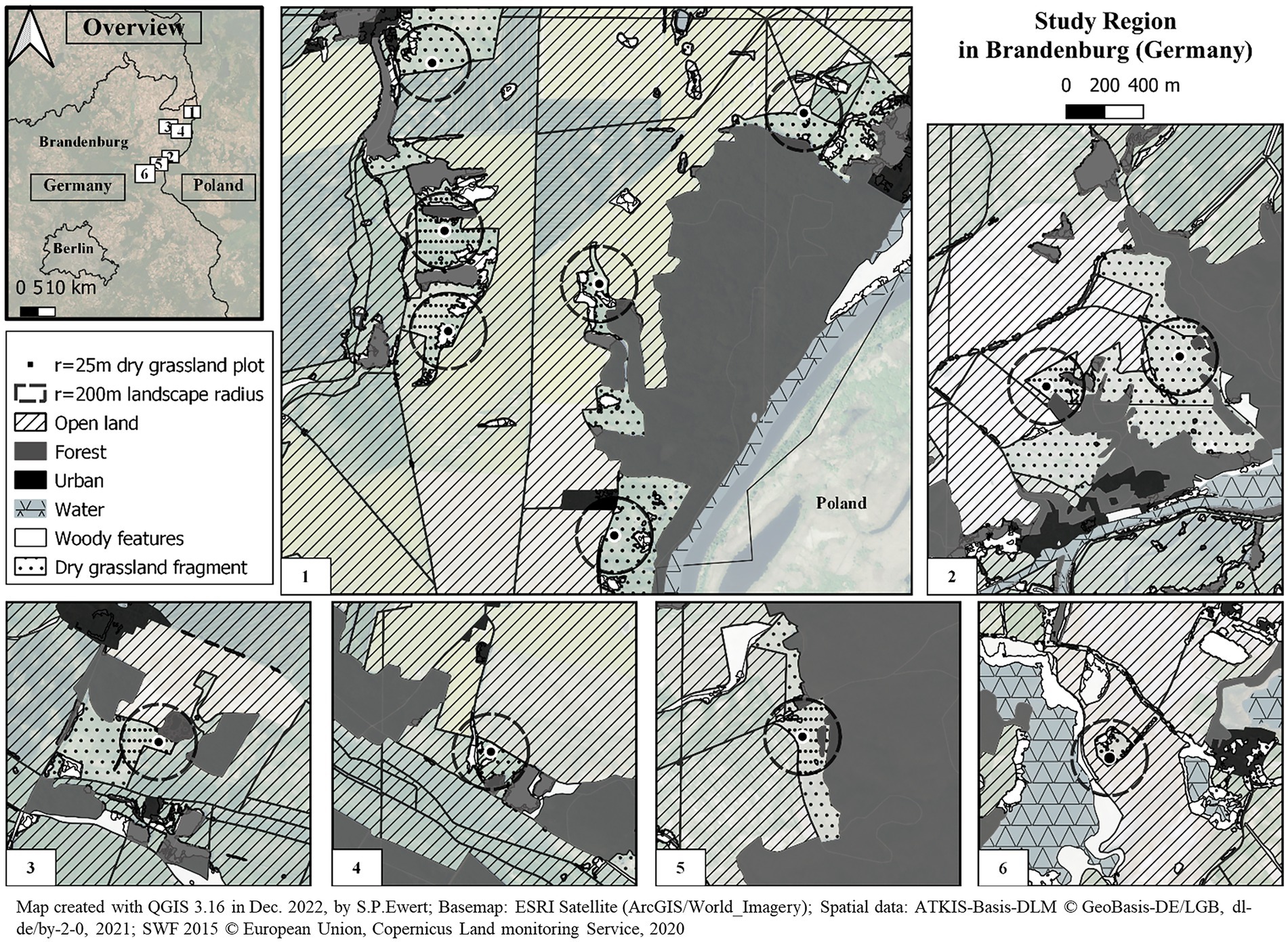
Figure 1. Map of study region in Brandenburg in Germany with the landscape types, locations of the dry grassland plots (n = 12) and the landscape radii for analysis.
We used Batlogger (Batlogger A, Elekon AG, Luzern, Switzerland) for repeated passive acoustic monitoring to assess bat activity above dry grassland fragments. Batlogger microphones were mounted on poles in the center of 12 designated study plots, which were intended for vegetation assessment (see 2.4), two meters above the ground and at least two meters away from larger trees to avoid strong echoes. We mounted one Batlogger per dry grassland fragment due to their small sizes, except on two larger fragments, where we used two plots each (see Figure 1). The locations of the plots were chosen in field depending on local structural conditions to cover the wide range of structural compositions. Their centers, hence Batlogger locations, were at least 500 m away from each other to prevent spatial autocorrelation and at least 30 m away from adjacent habitat types (e.g., crop and forest) to reduce edge effects. Note, however, that effects of other habitats were accounted for in the following landscape analysis. Each Batlogger sampled four nights (except at one plot, which was sampled three times due to access restrictions) throughout the main bat activity season (five plots in 2019, seven plots in 2020). Two sampling nights took place between June – July and (one) two between August – October. This resulted in 47 sampling nights. Recording started around 30 min before sunset and continued until approximately 30 min after sunrise. For triggering of sound recordings, we used the Crest Advanced setting that ensured the recording of quiet bat calls and reduced sensitivity to disturbing noise (Supplementary Datasheet S1). Sampling was avoided in nights with prolonged rainfall, high wind speed (>BFT 4; Martin et al., 2017) and low temperatures <8°C. We also avoided recording during sheep and cattle grazing, as movement of cattle can potentially increase bat activity (Downs and Sanderson, 2010; Ancillotto et al., 2017).
We performed semi-automatic call analysis in the program Batexplorer 2.1 (Elekon AG, Luzern, Switzerland) to assign recordings with bat echolocation calls to species level or to an identifiable sonotype (groups of bat species with very similar call structures). This was based on a self-created list of identification criteria (Supplementary Table S1) derived from various sources (Parsons and Jones, 2000; Russo and Jones, 2002; Pfalzer and Kusch, 2003; Obrist et al., 2004; Skiba, 2009; Barataud, 2020). All recordings were checked by hand to reduce misidentification (Russo and Voigt, 2016). We analyzed recordings with sequences containing two or more recognizable bat echolocation signals. Non-identifiable bat calls were omitted from further analysis. We also identified recordings with one or more feeding buzzes (distinct phases with rapid, steep calls with a high frequency bandwidth) as indications for insect prey encounter and therefore foraging activity. We grouped all bat species/sonotypes into three functional bat guilds by similarities in eco-morphological adaptations and in their preferred vegetation clutter conditions of habitats (Schnitzler and Kalko, 2001; Zeale et al., 2012; Dietz and Kiefer, 2016; Barataud, 2020): (i) open space guild (open habitats, high and fast flying bats with low maneuverability, narrowband and low-frequency echolocation calls), (ii) edge space guild (uses background cluttered habitats, such as forest edges, but is flexible in open and clutter; bats with intermediate speed and maneuverability; medium frequency echolocation calls with steep broadband and narrowband components), and (iii) narrow space guild (uses cluttered habitats close to vegetation; bats with low flight speed and high maneuverability; steep broadband echolocation calls). For a measure of habitat use of each guild, we took bat activity, which was defined by the recorded number of minutes of one night per plot, in which we found at least one echolocation recording (with a minimum of two calls) of the respective bat guild (Miller, 2001). We did the same for recordings including feeding buzzes as a measure of foraging activity. Feeding buzzes were not sampled for bats from the narrow space guild, as they mainly use gleaning and passive listening to find insects, instead of feeding buzzes (Entwistle et al., 1996; Schnitzler and Kalko, 2001).
From forest systems it is known, that higher three-dimensional structural heterogeneity supports a higher number of bat species and their activity (Adams and Law, 2011; Hanspach et al., 2012; Jung et al., 2012; Starik et al., 2018). We thus used an index that describes vertical as well as horizontal structural richness of the dry grassland plots. Therefore, we used the foliage height diversity index (FHD), which was originally proposed by MacArthur and MacArthur (1961) as a description of heterogeneity in vegetation layer coverage and to predict bird species richness. It was later adapted in other studies for birds (Wood et al., 2013) and for bat occurrence in forest systems (Froidevaux et al., 2016). For calculating the FHD as a measure of structural richness, we applied the Shannon-Wiener Index [in R 4.0.3. by R Core Team (2020); package: vegan, Oksanen et al., 2020; function: diversity], as proposed in Froidevaux et al., (2016). For that, we used cover values of foliage height classes: ground 0–1 m, shrub 1–5 m, tree low 5–10 m, tree high >10 m. To assess vegetation cover per height class, we did in-field vegetation sampling of the study plots within a 25 m radius around the Batlogger locations. We identified the positions and average heights of woody vegetation structures (shrubs and trees) that were taller than 0.5 m and drew the vegetation layout true to scale on a grid map. For height estimation of larger trees, we used a tree-height-measuring app (App: “Baumhöhenmesser,” Foresttools, Göttingen, Germany). In the following, looking at the drawn map and knowing the heights of the woody vegetation, we estimated the amount of square meters covered by woody vegetation in each defined height class and then calculated the percentage of the total plot covered by each height class. The FHD correlated positively with other descriptives of the dry grassland plots (cover values of height classes and total foliage cover) with Pearson coefficients from 0.8 and higher [in R 4.0.3. by R Core Team (2020), package: Ggally, by Schloerke et al. (2021), function “ggcorr”]. We therefore considered the FHD as an appropriate proxy for the description of local structural richness.
For the landscape analysis, we considered landscape features that are of known relevance for bats and would aid in describing the landscape in terms of openness, homogeneity, connectivity and availability of resources for bats. We thus included forests and urban areas, as important roosting and foraging habitats (e.g., Dietz and Kiefer 2016), as well as water bodies, offering valuable foraging sites especially in open land (e.g., Heim et al., 2018). We also considered woody features (e.g., hedges, tree lines and small woodland patches), as important connectivity features for many bat species using them for sonar guidance, protection and foraging (e.g., Verboom and Huitema, 1997). In addition, we included open land, consisting of pastures and predominantly cropland, known to negatively affect bat activity (e.g., Put et al., 2019). Lastly, we considered patchiness of the landscape as a measure for homogeneity. A homogeneous landscape, with less patches, can be detrimental for some bat species (e.g., Mendes et al., 2017) as it provides less heterogeneous niches and edge habitats for commuting and hunting (Ethier and Fahrig, 2011). The landscape features and the used measurements can be found in Table 1. To derive measurements of landscape features we performed landscape analysis (Supplementary image S1) within a 200 m radius around the Batlogger position using the geographic information system QGIS 3.16.3 (QGIS Development Team, 2021). We adapted a 200 m radius, as this distance has already shown significant effects of landscape features on bat activity and species composition, for example in Heim et al. (2015). We used layers with detailed landscape data (spatial data: ATKIS-Basis-DLM, source: ©GeoBasis-DE/LGB, dl-de/by-2-0, 2021). The categories of the landscape layers were then reclassified into broader categories: urban area, water body, forest and open land (pastures + cropland; Supplementary Table S2) and used for subsequent spatial analysis in QGIS. Furthermore, we used a high-resolution layer of woody features, which consisted of patchy, linear and additional woody structures, such as hedges and tree lines (spatial data: Small Woody Features 2015, source: ©European Union, Copernicus Land monitoring Service, 2020). Woody features directly connected to forest were reclassified as forest when they were protruding not more than 5 m from the forest. Further, we corrected major misclassification errors in landscape categorizations by checking in the field. We then calculated within the 200 m radius the cover of forest, open land (pastures + cropland) and woody features in percentage. Additionally, we counted the number of landscape patches regardless of their types within the 200 m radius. Separate patches could be for example landscape features distinct from their surroundings (e.g., forest or woody patches) or when divided by paths or roads (e.g., separate fields of crop or pastures). Lastly, we measured the linear distances from the Batlogger locations to the nearest urban areas, woody features and water bodies. We checked all variables for collinearity and found Pearson coefficients only between −0.6 and 0.6, indicating no notable correlations [in R 4.0.3. by R Core Team (2020), package: Ggally, by Schloerke et al. (2021), function “ggcorr”].
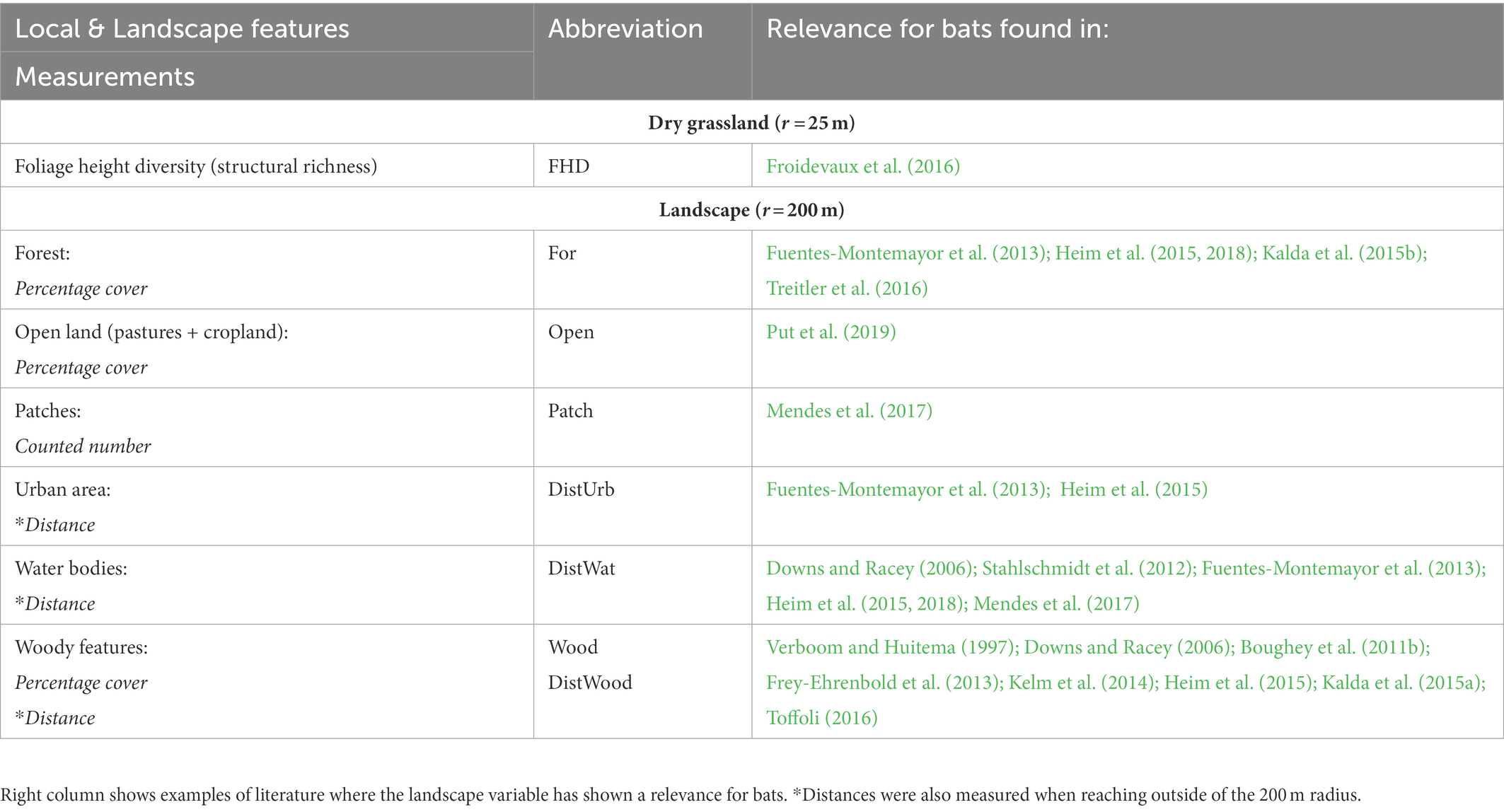
Table 1. Final choice of local and landscape features and their abbreviations used for further analysis.
The following statistical analyses were performed in R (version 4.0.3, R Core Team, 2020).
To assess whether bat activity and foraging of each functional guild responded to increased structural richness (FHD) of dry grassland plots, as well as to landscape features and/or the interaction of both, we applied information-theoretic model selection with generalized linear mixed models (R package: glmmTMB, Brooks et al., 2017). This approach was chosen over an a priori set of candidate models, as we expected all predictors to have effects on bat activity and foraging to some extent (see “Landscape features”) and it allowed us to find the most explanatory set of predictors (Bolker et al., 2009; Frey-Ehrenbold et al., 2013; Kalda et al., 2015a; Mendes et al., 2017). We used a negative binomial link due to overdispersion. FHD and landscape features were taken as fixed effects and mean nightly temperatures and the IDs of the dry grassland plots were taken as random effects. All predictors were scaled using the scale function (Becker et al., 1988). We conducted stepwise forward selection while keeping FHD in every model and in a stepwise manner adding landscape features as main effects and then as interaction terms with FHD, looking for the model with the best AIC value indicating the most parsimonious model (Supplementary Table S3). We chose forward selection for the following reasons: first, to reduce the risk of convergence issues due to overly complex models; secondly, to mirror our research questions approach, by initially testing for effects of dry grassland structural richness and then adding features from the broader landscape scale. Model selection was performed for all guilds and their activity as well as their foraging activity (except for the narrow space guild).
From the cumulative total bat activity of all three functional guilds per night and plot, we calculated the percentages of nightly activity per guild. From there we derived the median activity per dry grassland plot for each guild. This made the dry grassland plots comparable in their proportions of functional guild activity counts. To investigate the proportional differences on dry grassland fragments we performed non-metric multidimensional scaling (NMDS in R package: vegan; function: metaNMDS, Oksanen et al., 2020). We then applied environmental fitting of the dry grassland and landscape features onto the ordination (R package: vegan, Oksanen et al., 2020; function: envfit) to investigate their potential to explain proportional changes of bat assemblages between dry grassland plots. Results were supported by using permutational multivariate analysis of variance [PERMANOVA in R package: vegan, Oksanen et al., 2020; function: adonis, based on Anderson (2001)].
At 12 dry grassland plots, during a total of 47 sampling nights, we found 13 bat species/sonotypes that we had grouped into the three functional guilds (Table 2) and 5,488 minutes with echolocation activity of one or more bat species/sonotypes. Bats were active during 19% of the total recording time (28,560 min) in which one or more bats could have been recorded. Per night at a plot, we counted on average almost 2 hours of bat activity (mean: 117 min). Eight percent of activity minutes contained feeding buzzes of one or more bat species/sonotypes from the open and edge space guild, indicating foraging activity. Nightly activity (Figure 2) mainly consisted of the open and the edge space guild. Activity of bats from the narrow space guild was generally much lower. The activity of the open space guild was dominated by Nyctalus noctula (75%). The edge space guild showed shared dominances of Pipistrellus nathusii (37%), Pipistrellus pygmaeus (29%) and Pipistrellus pipistrellus (23%). Lastly, the Myotis group (81%) formed the largest part of the narrow space guild. Each dry grassland plot was visited by an average of 11 bat species/sonotypes throughout the year, with a nightly mean of eight, indicating a regular use by a high proportion of the recorded bat species/sonotypes. Additionally, we recorded at least three species/sonotypes of higher conservation concerns according to the German Red List (Meinig et al., 2020): Eptesicus serotinus (threatened) on 11 plots, Plecotus spp. (threatened) on 9 plots and Barbastella barbastellus (highly threatened) on one plot.
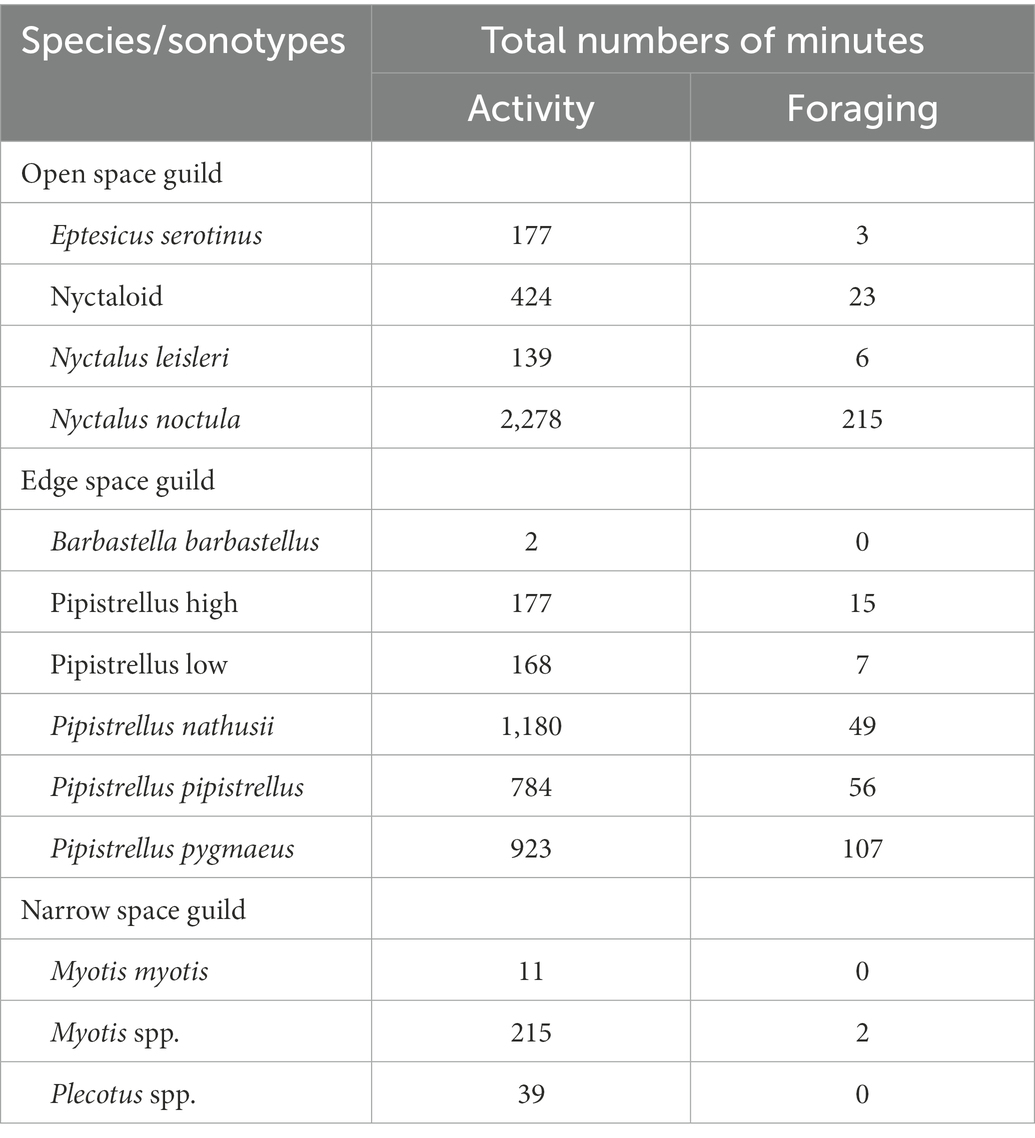
Table 2. List of identified species/sonotypes in their associated functional guilds and the accumulated number of activity/foraging minutes based on 47 sampling nights.
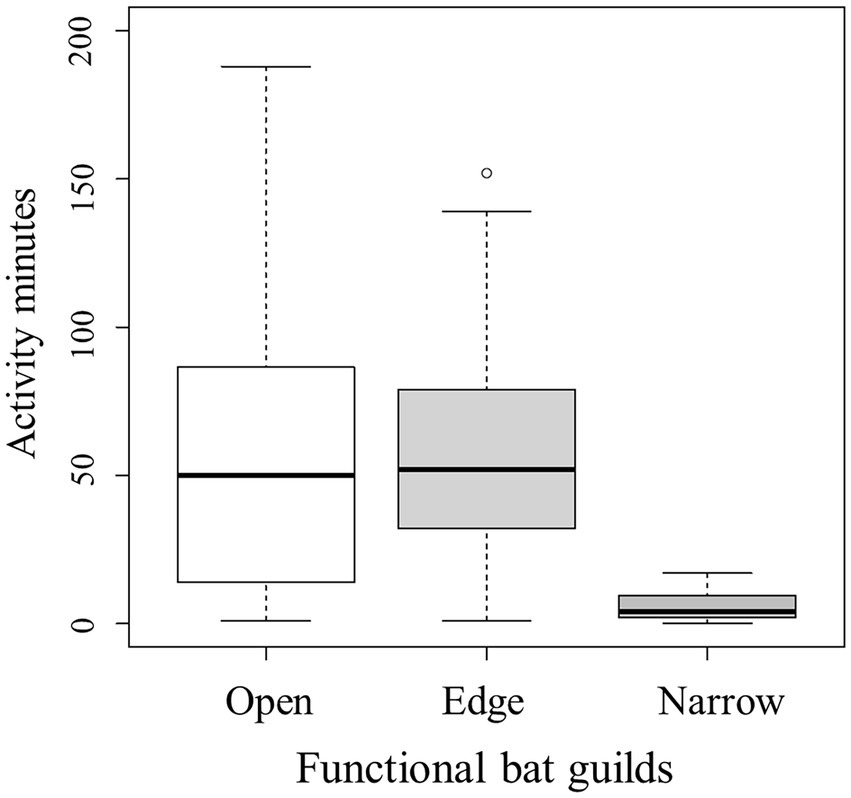
Figure 2. Nightly activity minutes of bat guilds (open, edge, and narrow space) counted during 47 nights on 12 dry grassland plots.
Model selections revealed that a combination of both local (plot level) and landscape scales, and especially their interactions, is important for explaining activity (Table 3) and foraging (Table 4) of bat functional guilds at the dry grasslands.
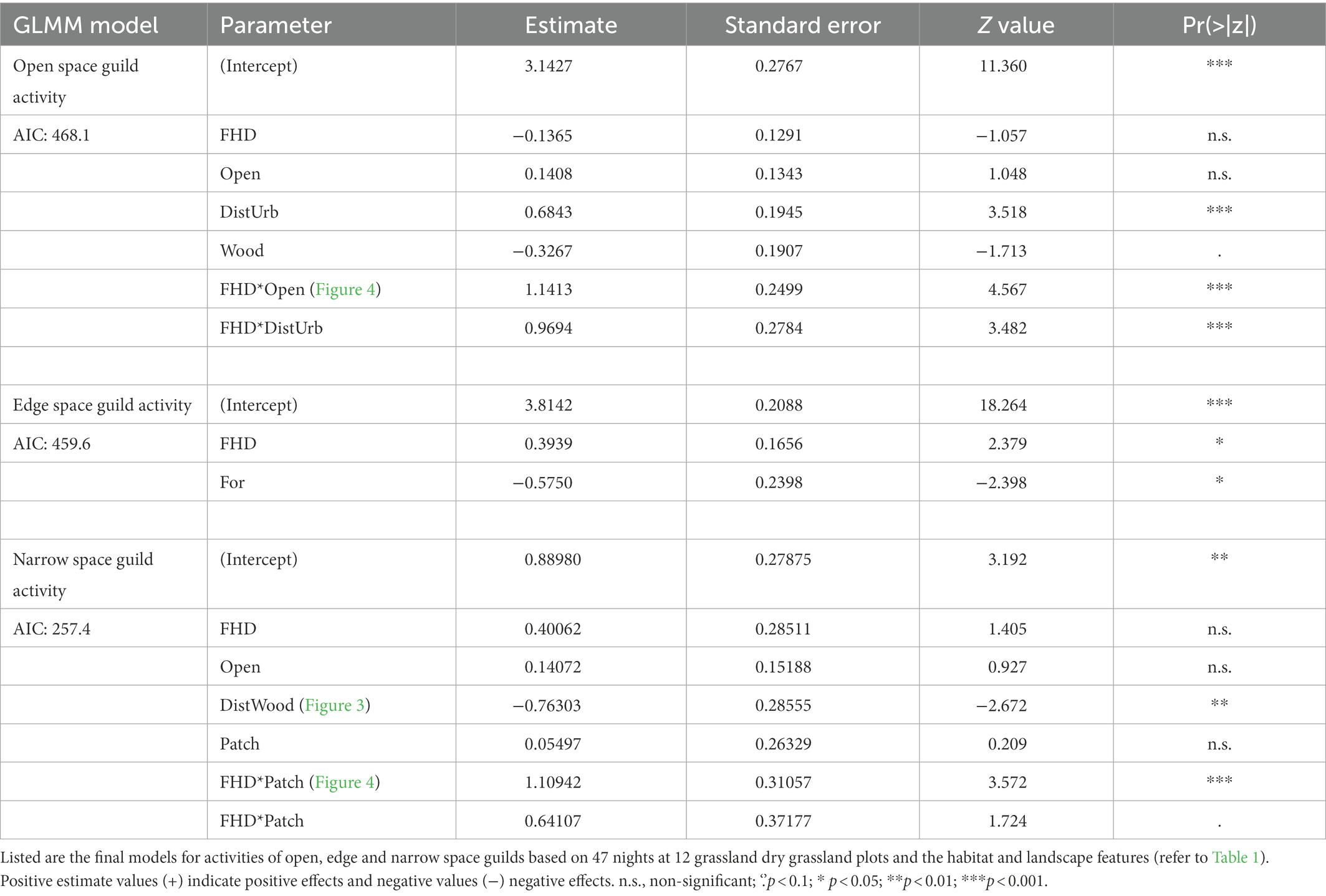
Table 3. Results of bat guild activity predictions from the forward selections of generalized linear mixed models (GLMM) with negative binomial links based on selection of the best Akaike’s information criterion (AIC).
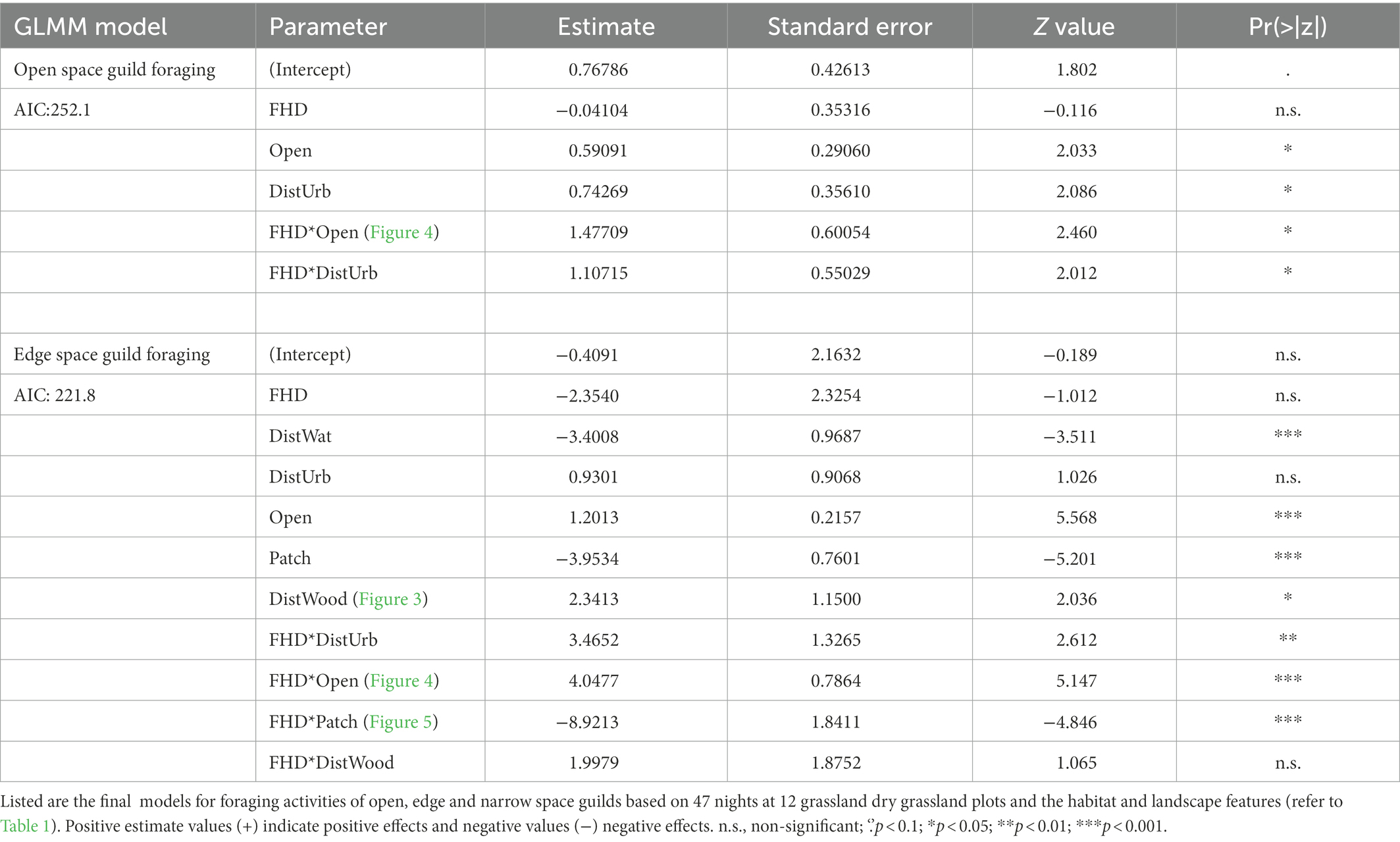
Table 4. Results of bat guild foraging activity predictions from the forward selections of generalized linear mixed models (GLMM) with negative binomial links based on selection of the best Akaike’s information criterion (AIC).
Bats from the edge space guild showed strong associations with local structural conditions. Their activity generally increased significantly with higher structural richness (FHD), regardless of landscape interactions. We also found significant additive effects of features on the landscape scale on edge space bats. They foraged more above dry grassland plots when water bodies were closer by, indicating associations with other main habitat types. However, they were more active on dry grasslands plots when there was less forest in the surrounding and their foraging activity increased with greater distances to woody features (Figure 3), suggesting an increased use of dry grasslands when other habitat sources are less available. Activity of bats from the narrow space guild decreased at dry grassland plots further away from woody features (Figure 3), showing a stronger accessibility limitation.
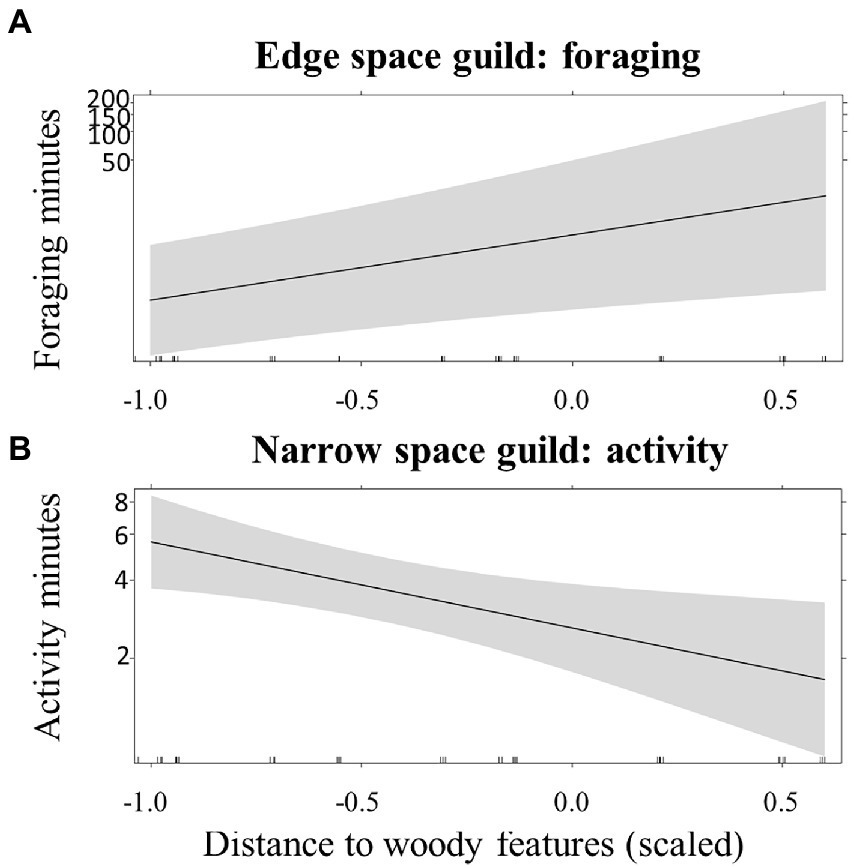
Figure 3. Effect plot for the predicted effects of distance to woody features (scaled) on (A) edge space guild foraging and (B) narrow space guild activity.
For all functional guilds, we found significant interaction effects of local structural richness in combination with landscape features describing openness, homogeneity and availability of main habitats. The activity of open and narrow space bats (Table 3) as well as the foraging activity of open and edge space bats (Table 4) increased above dry grassland plots with more local structural richness, when they were surrounded by a higher percentage of open land (pastures and cropland). In less open landscapes, structural richness was less important for the open space guild (Figures 4A,B) and the narrow space guild (Figure 4D), while in the foraging activity of the edge space guild, the effect was still positive but less prominent (Figure 4C). Additionally, edge space bat foraging increased on structurally richer dry grassland plots, when the number of patches in the surrounding decreased (Figure 5). Lastly, structurally richer dry grassland plots were used more by open space bats (increase in activity and foraging) and foraged more by edge space bats, when urban areas were further away.
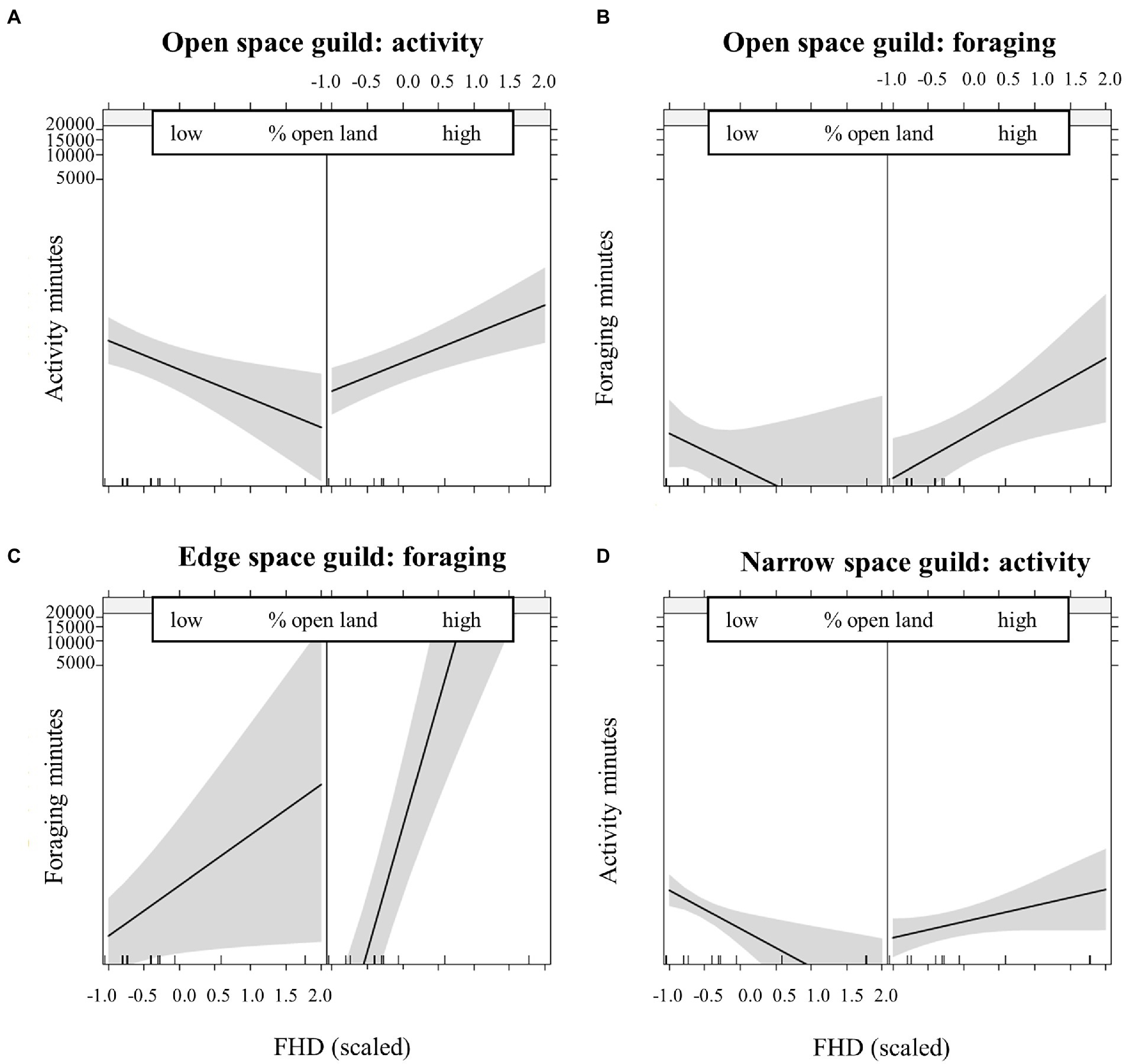
Figure 4. Effect plots of the interaction of the foliage height diversity (FHD, scaled) and % open land (scaled). Boxes show the predicted effects of FHD in low and high % of open land on (A) open space guild activity and (B) foraging, (C) edge space guild foraging, and (D) narrow space guild activity. Low and high values are represented by the first and third quartile of the open land data (scaled) respectively.
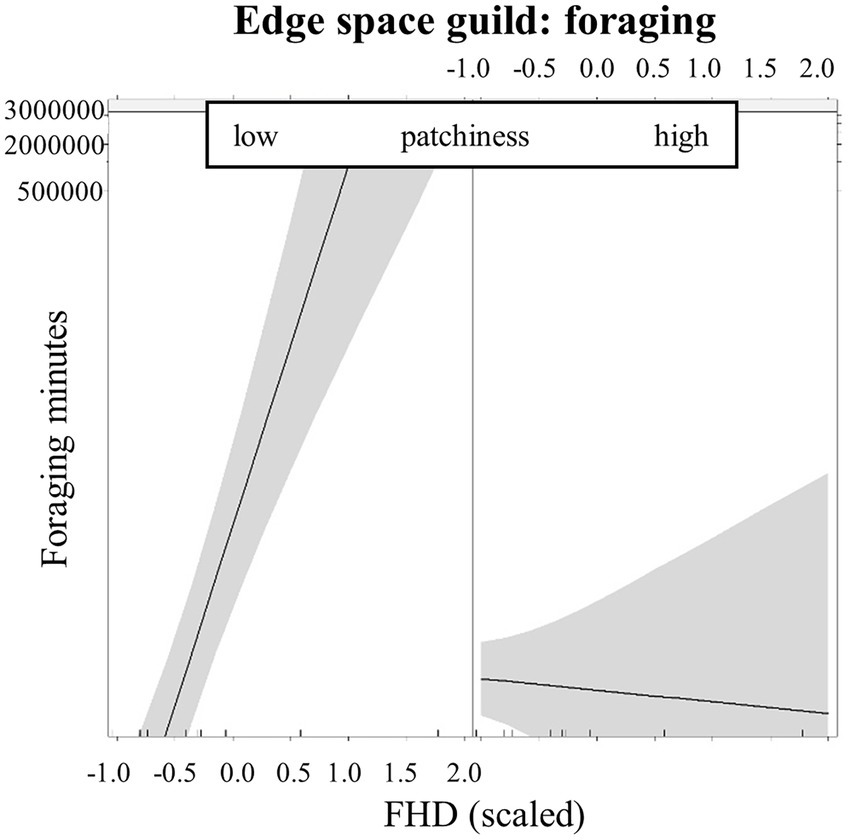
Figure 5. Effect plots of the interaction of the foliage height diversity (FHD, scaled) and number of patches (scaled). Boxes show the predicted effects of FHD in low and high amount of number of patches for foraging activity of the edge space guild. Low and high values are represented by the first and third quartile of the number of patches (scaled) respectively.
Non-metric multidimensional scaling separated dry grassland plots based on the median percentage of nightly activities of the function guilds (Figure 6; NMDS: k = 2, stress = 0.006, perm = 999, Supplementary Table S4). Environmental fitting including local and landscape features showed that the structural richness (FHD) significantly explained separation of dry grassland plots along the y-axis (r2 = 0.68, p = 0.010). Especially the activity proportion of edge space guild increased with higher structural richness. This was supported by results of the PERMANOVA (FHD: r2 = 0.54, p = 0.022, Supplementary Table S5) and suggests that structural composition of dry grassland fragments influences assemblage composition of bats. Landscape features did not show significant effects.
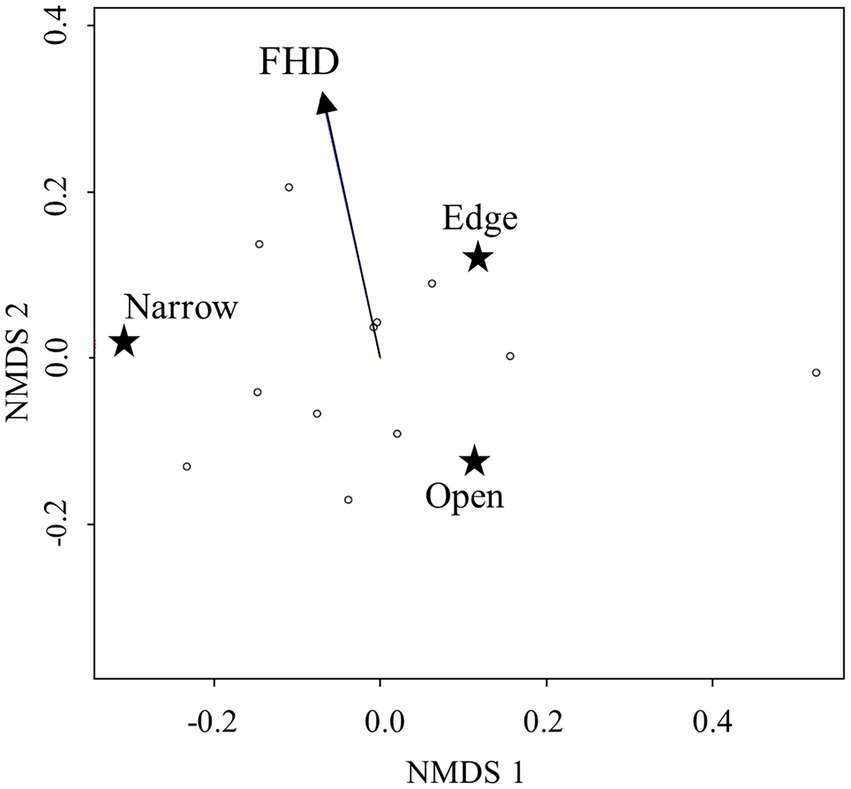
Figure 6. NMDS ordination plot showing the 12 dry grassland plots in k = 2 dimensions. Placement of plots is based on Bray–Curtis dissimilarity of the activity compositions of functional bat guilds (indicated by stars). Environmental fitting was used to find explanatory variables for differences in guild composition. The significant variable (FHD) is represented as a vector.
Loss of (semi-) natural habitats and ecosystem changes are consequences of agricultural intensification, which is globally identified as the major cause of the current biodiversity crisis (Burns et al., 2016; Young et al., 2016; Díaz et al., 2019; Raven and Wagner, 2021). The presence of non-crop habitats in intensified agro-ecosystems is crucial to enable occurrence and dispersal of a variety of organisms that are necessary to sustain a resilient species pool and associated ecosystem services (Tscharntke et al., 2005; Fischer et al., 2006). In this study we focused on fragmented, protected European semi-natural dry grasslands, which are known for a very high plant biodiversity (Dengler et al., 2012; Wilson et al., 2012) and a rich insect fauna (Wallis de Vries and Van Swaay, 2009; Rasran et al., 2018; Schuldt et al., 2019). An increased value has been particularly proven for small dry grassland fragments in agro-ecosystems, locally (Rösch et al., 2015; Deák et al., 2020) and in exchange with the surrounding landscape (Tscharntke et al., 2002; Duelli and Obrist, 2003; Lindborg et al., 2014; Zulka et al., 2014). However, their importance for highly mobile species, such as bats, is not yet understood, even though many bat species face challenges of agricultural intensification, such as habitat and connectivity losses (Browning et al., 2021) and when moving through open land (Frey-Ehrenbold et al., 2013). Revealing which local structural conditions and landscape features as well as their interactions increase commuting and foraging of bats on dry grassland fragments, is of utmost importance for bat conservation and for evidence-based decision making in management and conservation efforts of dry grasslands. Our results shed light onto the importance of dry grassland fragments for bats in a farmland-dominated region in Germany in relation to local vegetation structural richness and the availability of other important habitats. This is especially relevant in agriculturally rich regions, where other non-crop habitats are reduced (Burns et al., 2016) or more distant. We show regular visitations by a relatively high number of bat species/sonotypes throughout the year, also by species/sonotypes of higher conservation concern. Our results clearly show, that dry grassland use by bats is determined by local structural richness, the features of the surrounding landscape and the interaction of both.
As predicted, our results show additive effects of local and landscape features on dry grassland use by bats. This is in accordance with findings of other studies that highlighted the influence of landscape features at different scales on bat occurrence and their activities (Boughey et al., 2011a; Kalda et al., 2015b; Heim et al., 2017; Mendes et al., 2017).
Bats from the edge space guild appeared to be the most responsive to local conditions of dry grassland fragments. Their activity above dry grassland plots generally increased with a higher structural richness, hence more heterogeneous shrubs and trees. This is corroborated by findings of previous studies, showing that edge space bats prefer moving along structural features (Verboom and Huitema, 1997; Downs and Racey, 2006; Toffoli, 2016), specifically close to taller trees (Froidevaux et al., 2022) and it suggests that structurally richer dry grasslands have a higher value than very open ones, for edge space bats commuting in relatively open agricultural landscapes settings. When looking at landscape features, edge space bats were more active above dry grassland plots when less forest was available in the surrounding, and they foraged more at dry grassland plots when woody features (such as hedgerows) were further away. This is opposite to findings of other studies that show higher activity of the edge space bats (such as Pipistrellus species) close to forest (Davidson-Watts et al., 2006), forest edges (Heim et al., 2018) and woody features (Verboom and Huitema, 1997; Downs and Racey, 2006; Frey-Ehrenbold et al., 2013). However, the latter studies also highlight that Pipistrellus species can cover greater distances in open land away from favorite habitats to reach other favorable land patches. According to Kelm et al. (2014), the biggest activity drop of Pipistrellus species away from structural edges into open land is usually within the first 50 m. Since we show increased activity at dry grassland plots away from woody features (here average distance: 44 m) and with less surrounding forest cover, we propose that dry grassland fragments are possibly favorable habitats for bats from the edge space guild (see also Davidson-Watts et al., 2006 for Pipistrellus species in unimproved grasslands) and potentially fulfill an important function for commuting and foraging. These results are not supporting our expectation of accessibility limitations for edge space bats, although this could possibly become the case in regions where dry grasslands fragments are too isolated from other connecting landscape features (Frey-Ehrenbold et al., 2013). In contrast, but as expected, the narrow space guild exhibited stronger accessibility restrictions by showing reduced activity on dry grassland plots further away from woody features. This is in accordance with findings from Frey-Ehrenbold et al. (2013) showing the greatest limitations in open land for the narrow space bats of the Myotis and Plecotus groups. Therefore, isolated dry grasslands fragments might eventually fall out of their reach, if they are beyond their accessibility threshold.
Our results also highlight the importance of other habitat types in the landscape. Edge space bats foraged more on dry grassland plots closer to waterbodies. The high value of the latter for various bat species as foraging ground has been supported by other studies before (Stahlschmidt et al., 2012; Kalda et al., 2015a; Heim et al., 2018). Wetlands are also among the most preferred habitats for the edge space bat species Pipistrellus pipistrellus and Pipistrellus pygmaeus (Davidson-Watts et al., 2006; Dietz and Kiefer, 2016).
We did not find independent effects of local structural richness on bats from the open and the narrow space guild, but only in interaction with landscape features, indicating that for these bats surrounding conditions determine the importance of dry grasslands and their structures (discussed below). We also did not find significant effects of surrounding forest cover on bat activity and foraging in the open and narrow space guilds, even though other studies have shown positive associations of several bat species with forest proximity (Frey-Ehrenbold et al., 2013; Treitler et al., 2016) and cover (Boughey et al., 2011a; Heim et al., 2015; Mendes et al., 2017). We suppose that changes in the overall low forest cover in the 200 m radius (mean: 7%) were irrelevant for the fast flying open space bats and that the long distances (mean: 153 m) excluded many woodland dwelling narrow space species, keeping their activity low.
Our results here indicate the use of dry grasslands by bats in relation to local conditions and to availability of other main bat habitats and landscape features, which mainly affected bats from the edge and narrow space guild. While dry grassland fragments appear beneficial for commuting and foraging of bats from the edge space guild, especially when containing heterogeneous vegetation structures, narrow space bats show a dependency on higher connectivity to other structural landscape features (such as hedgerows). Our findings therefore also underline the necessity of connectivity networks and corridors in open farmland (Rosenberg et al., 1997) and the availability of other important habitats (Heim et al., 2015) to facilitate bat dispersal and provide additional resources.
We furthermore confirmed our prediction that landscape features would influence the correlations with local structural conditions and therefore determine dry grassland use. Key findings in this study are the impacts of the amount of open land (pastures and cropland) and of the number of landscape patches on the bat activity correlation with local structural richness. Bats used structurally richer dry grassland plots more for commuting and/or foraging, when the surroundings were more open (all guilds) and less patchy, hence more homogeneous (edge space guild). Both parameters, openness and homogeneity of landscapes, are of special importance, since they usually are negative consequences of agricultural intensification (Matson et al., 1997; Stoate et al., 2001; Tilman et al., 2001; Jongman, 2002; Burns et al., 2016). They very likely determine resource availability for bats in the landscape due to the following reasons: A higher amount of open farmland means the reduction of other main non-crop habitats, necessary for commuting and foraging. Similarly, homogeneous landscapes with less patches usually contain fewer crop types with reduced insect diversity (Olimpi and Philpott, 2018), and fewer habitat edges (Ethier and Fahrig, 2011). Consequently, previous studies have found decreasing bat activities of several bat species in farmland with greater amounts of open agricultural land (Put et al., 2019), above improved meadows and pastures when further away from other important habitats (Heim et al., 2015; Treitler et al., 2016), and with less landscape heterogeneity (Monck-Whipp et al., 2018; Froidevaux et al., 2022). The latter especially facilitates edge bats movement (Mendes et al., 2017), which could explain why patchiness only appeared in results for the edge guild. Despite the just named known negative effects of a high amount of open farmland and heterogeneity loss, we found locally increased bat activity and foraging above dry grassland plots, when these provided higher structural richness. This is also contrary to findings of Froidevaux et al. (2022), who showed that local features become more relevant for many bat species only when the landscape is more heterogeneous. We therefore argue that the value of dry grassland fragments with diverse shrub and tree vegetation in intensive farmland increases, when the surrounding landscape becomes more unfavorable. For that, we provide the following explanations: first, the higher richness in vegetation structures can offer more niches, aiding a high variety of bat species with different sensory requirements and habitat adaptations, as it is known from forest systems (Adams and Law, 2011; Hanspach et al., 2012; Jung et al., 2012; Müller et al., 2013; Froidevaux et al., 2016; Blakey et al., 2017; Starik et al., 2018). Secondly, dry grasslands contain a rich insect fauna (Wallis de Vries and Van Swaay, 2009; Rasran et al., 2018) as potential food sources. The presence of more and denser vertical woody vegetation structures, such as trees, can further increase insect abundance (Müller et al., 2012; Fuentes-Montemayor et al., 2013), especially in agricultural regions [such as at trees and hedgerows in Grüebler et al. (2008)]. In the surrounding landscape, certain types of farmland, such as orchards or vineyards, could also provide diverse vegetation structures. Additionally, even open cropland can offer occasional plentiful insect food sources for bats. However, the latter is usually ephemeral and depending on crop growth and season (Heim et al., 2016, 2017; Stahlschmidt et al., 2017). Moreover, in conventional farmland, feeding on insects will lead to direct contamination of bats with pesticides (Stahlschmidt and Brühl, 2012; Stahlschmidt et al., 2017). Dry grasslands on the other hand, as semi-natural habitats, can provide stable food supplies for bats throughout the growing season (Davidai et al., 2015) and thus buffer negative effects of intensive farming practices.
We additionally found prominent interaction effects of distance to urban areas and local dry grassland conditions in the open space (activity and foraging) and edge space (foraging) guild, as their use of dry grassland plots increased with higher structural richness and with greater distance to urban areas. It is known that some species especially from open and edge space guilds regularly utilize urban areas for roosting and foraging (Jung and Threlfall, 2018) and that the proximity to anthropogenic areas can positively affect bat species occurrence and activity in grasslands nearby (Heim et al., 2015). Our study region contained mainly rural, low-density settlements, which have great potential as roosting or feeding sites (Gehrt and Chelsvig, 2004; Dixon, 2012). We therefore argue, that structurally richer dry grasslands increase in value when further away from such urban sites.
Our results here could only be obtained by investigating interaction terms of landscape and local features. As other studies suggested, effects of landscape conditions on bats at a broader scale are usually dominant over local conditions (Kalda et al., 2015a,b) but interaction terms can reveal scenarios where local features become relevant. Hence, the availability of resources for commuting and foraging provided by the surroundings determine the importance of local features. We thus conclude that in simpler and more open agricultural regions, structurally richer dry grasslands are favored by bats over less structured ones.
In accordance to our prediction, we found that local structural conditions on dry grasslands change bat guild activity compositions. Our results show a proportional shift from open space to edge space guild activity with higher structural richness of plots. This again highlights the previously discussed positive affinity of edge space bats with structurally richer dry grasslands, which makes them the dominating guild, when diverse shrubs and trees are available. This corroborates the results of Davidson-Watts et al. (2006), who found that (scrubby) grasslanda are among favored habitat types for Pipistrellus pipistrellus and P. pygmaeus. We did not clearly see dominance changes along the structural richness gradient in the narrow space guild, which seems counterintuitive given their affinity to vegetation clutter. The activity of the narrow space guild however was generally low, which might have been caused by the overall open character of the landscape, and proportional changes were likely to be minor, compared to the other dominant guilds.
Heim et al. (2015) found shifts in species composition above pastures and meadows in correlation with cover of and distances to landscape features in a 200 m radius. In contrast, we did not find any explanation for shifts of bat assemblages by any landscape features. Here the limited sample size of dry grassland plots embedded in not extremely varying landscapes (given that most dry grassland fragments are located in farmland), might have obscured possible evidences.
To our knowledge, we show for the first time the importance of European dry grassland fragments for bats under certain local and landscape compositional conditions in an agriculturally rich region in Germany. While we confirm a constant use of dry grasslands by a high number of bat species/sonotypes throughout the season, we also show that effects of local and surrounding landscape features on bat activity and foraging vary between functional bat guilds. Nonetheless, our study emphasizes for all bat guilds the significant value of dry grasslands with increased local structural richness in unfavorable, open and homogeneous farmland. We thus argue that dry grassland fragments especially with heterogeneous shrub and tree vegetation have a high potential as stepping stones for bats in intensive agricultural regions. By that means, we underline the necessity of heterogeneity on different spatial scales for biodiversity in such landscapes (Benton et al., 2003). Our findings are of high ecological importance and conservation concern, as many bat species are facing increased challenges in agriculturally intensified regions through habitat and connectivity loss (Browning et al., 2021), but are providing important services as insect pest control (Cleveland et al., 2006; Kalka and Kalko, 2006; Kalka et al., 2008; Williams-Guillén et al., 2008). With our results, we add evidence to the high biodiversity value of European dry grassland fragments in agricultural regions, which was previously mainly shown for plants (Lindborg et al., 2014; Deák et al., 2020) and insects (Tscharntke et al., 2002; Duelli and Obrist, 2003; Rösch et al., 2015). Lastly, our findings contribute to informed decision making for dry grassland management and bat conservation.
For further applications, we highlight the importance of careful considerations before removal of heterogeneous shrub and tree vegetation on dry grassland fragments. This is contradicting the current management goals of dry grasslands, aiming to reduce shrub and tree encroachment. However, in this study we showed positive correlations of bat activity with increased structural richness, while the maximum structural richness corresponded with a rather low foliage cover of 25% (summed up cover of shrub and tree foliage), which is still equivalent to a mediocre management state for the dry grassland types found in Brandenburg, Germany (Landesamt für Umwelt Brandenburg, 2022). We thus propose considering leaving heterogeneous shrubs and trees on dry grassland fragments particularly in open, homogenous and intensively used agricultural regions, while this appears less important in heterogeneous landscapes. In line with previous studies, we highlight the need of connectivity networks (such as hedgerows) in landscapes to allow accessibility of edge and narrow space bat species (Frey-Ehrenbold et al., 2013; Heim et al., 2015). More importantly, we strongly advise to co-evaluate interactions of local and landscape features, as this leads to a better understanding of the landscape settings that determine the importance of dry grassland conditions for bats. Finally, we encourage more research on the relevance of these habitat types for single bat species, in different regions and subsequently also for other non-target organisms.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethical review and approval was not required for the animal study because acoustic monitoring of bats is non-invasive and does not involve any animal contact. However, fieldwork and methodological approaches were approved by the State Office for the Environment and access was permitted by the nature conservation authorities of Barnim and Uckermark (districts in Brandenburg, Germany).
SE conceptualized and designed the study. Data collection in the field was performed by SE and K-HF. SE did the data processing and statistical analysis. The manuscript was written by SE. MK, KJ, and K-HF contributed to all previous steps through guidance and supervision. All authors further contributed to manuscript revision and read and approved the submitted version.
This research was funded as a part of the project INPEDIV (SAW-2019-ZFMK-3-INPEDIV). Open Access Funding was provided by the Freie Universität Berlin.
We thank Livia Schäffler (Zoological Research Museum Alexander Koenig, Bonn, Germany) for her contribution as project lead and coordination in INPEDIV and in acquisition of fieldwork permissions for dry grassland sites. We greatly appreciate the assistance and efforts of Artur and Anna Ewert in fieldwork. We thank the various landowners and managers as well as cattle herders and farmers for their cooperation as well as their great interests and openness toward our work. We would like to thank participants of INPEDIV for their exchange and specifically Carsten Brühl, Bodo Möseler, and Benito Schoepke for their support and input. We greatly appreciate Neil Middleton for his exchange about bat acoustics. Lastly, we acknowledge support by the Open Access Publication Initiative of Freie Universität Berlin.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.995133/full#supplementary-material
Adams, M. D., and Law, B. S. (2011). “A preliminary assessment of the impact of forest thinning on bat activity: towards improved clutter-based hypotheses,” in The Biology and Conservation of Australasian Bats, ed. Law, B., Eby, P., Lunney, D. and Lumsden, D. (Sydney, NSW: Royal Zoological Society of New South Wales), 363–379.
Aldridge, H., and Rautenbach, I. (1987). Morphology, echolocation and resource partitioning in insectivorous bats. J. Anim. Ecol. 56:763. doi: 10.2307/4947
Ancillotto, L., Ariano, A., Nardone, V., Budinski, I., Rydell, J., and Russo, D. (2017). Effects of free-ranging cattle and landscape complexity on bat foraging: implications for bat conservation and livestock management. Agric. Ecosyst. Environ. 241, 54–61. doi: 10.1016/j.agee.2017.03.001
Anderson, M. J. (2001). A new method for non-parametric multivariate analysis of variance. Austral Ecol. 26, 32–46. doi: 10.1111/j.1442-9993.2001.01070.pp.x
Barataud, M. (2020). Acoustic Ecology of European Bats. Paris: Museum national d'Historie naturelle.
Becker, R. A., Chambers, J. M., and Wilks, A. R. (1988). The New S Language, a Programming Environment for Data Analysis and Graphics. London: Wadsworth & Brooks/Cole Computer Science Series by Chapman & Hall.
Benton, T. G., Vickery, J. A., and Wilson, J. D. (2003). Farmland biodiversity: is habitat heterogeneity the key? Trends Ecol. Evol. 18, 182–188. doi: 10.1016/S0169-5347(03)00011-9
Blakey, R. V., Law, B. S., Kingsford, R. T., and Stoklosa, J. (2017). Terrestrial laser scanning reveals below-canopy bat trait relationships with forest structure. Remote Sens. Environ. 198, 40–51. doi: 10.1016/j.rse.2017.05.038
Bolker, B. M., Brooks, M. E., Clark, C. J., Geange, S. W., Poulsen, J. R., Stevens, M. H. H., et al. (2009). Generalized linear mixed models: a practical guide for ecology and evolution. Trends Ecol. Evol. 24, 127–135. doi: 10.1016/j.tree.2008.10.008
Boughey, K. L., Lake, I. R., Haysom, K. A., and Dolman, P. M. (2011a). Effects of landscape-scale broadleaved woodland configuration and extent on roost location for six bat species across the UK. Biol. Conserv. 144, 2300–2310. doi: 10.1016/j.biocon.2011.06.008
Boughey, K. L., Lake, I. R., Haysom, K. A., and Dolman, P. M. (2011b). Improving the biodiversity benefits of hedgerows: how physical characteristics and the proximity of foraging habitat affect the use of linear features by bats. Biol. Conserv. 144, 1790–1798. doi: 10.1016/j.biocon.2011.02.017
Brooks, M. E., Kristensen, K., van Benthem, K. J., Magnusson, A., Berg, C., Nielsen, A., et al. (2017). glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. The R Journal, 9:2, 378–400. doi: 10.32614/RJ-2017-066
Browning, E., Barlow, K. E., Burns, F., Hawkins, C., and Boughey, K. (2021). Drivers of European bat population change: a review reveals evidence gaps. Mammal Rev. 51, 353–368. doi: 10.1111/mam.12239
Burns, F., Eaton, M. A., Barlow, K. E., Beckmann, B. C., Brereton, T., Brooks, D. R., et al. (2016). Agricultural management and climatic change are the major drivers of biodiversity change in the UK. PLoS One 11:e0151595. doi: 10.1371/journal.pone.0151595
Butaye, J., Adriaens, D., and Honnay, O. (2005). Conservation and restoration of calcareous grasslands: a concise review of the effects of fragmentation and management on plant species. BASE 9
Cleveland, C. J., Betke, M., Federico, P., Frank, J. D., Hallam, T. G., Horn, J., et al. (2006). Economic value of the pest control service provided by Brazilian free-tailed bats in south-Central Texas. Front. Ecol. Environ. 4, 238–243. doi: 10.1890/1540-9295(2006)004[0238:evotpc]2.0.co;2
Davidai, N., Westbrook, J. K., Lessard, J.-P., Hallam, T. G., and McCracken, G. F. (2015). The importance of natural habitats to Brazilian free-tailed bats in intensive agricultural landscapes in the winter garden region of Texas, United States. Biol. Conserv. 190, 107–114. doi: 10.1016/j.biocon.2015.05.015
Davidson-Watts, I., Walls, S., and Jones, G. (2006). Differential habitat selection by Pipistrellus pipistrellus and Pipistrellus pygmaeus identifies distinct conservation needs for cryptic species of echolocating bats. Biol. Conserv. 133, 118–127. doi: 10.1016/j.biocon.2006.05.027
Deák, B., Rádai, Z., Lukács, K., Kelemen, A., Kiss, R., Bátori, Z., et al. (2020). Fragmented dry grasslands preserve unique components of plant species and phylogenetic diversity in agricultural landscapes. Biodivers. Conserv. 29, 4091–4110. doi: 10.1007/s10531-020-02066-7
Dengler, J., Becker, T., Ruprecht, E., Szabó, A., Becker, U., Beldean, M., et al. (2012). Festuco-Brometea communities of the Transylvanian plateau (Romania)–a preliminary overview on syntaxonomy, ecology, and biodiversity. Tuexenia 32, 319–359.
Díaz, S., Settele, J., Brondízio, E. S., Ngo, H. T., Guèze, M., Agard, J., et al. (2019). Summary for policymakers of the global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. Bonn: IPBES secretariat. doi: 10.5281/zenodo.3553579
Dixon, M. D. (2012). Relationship between land cover and insectivorous bat activity in an urban landscape. Urban Ecosyst. 15, 683–695. doi: 10.1007/s11252-011-0219-y
Downs, N. C., and Racey, P. A. (2006). The use by bats of habitat features in mixed farmland in Scotland. Acta Chiropterologica 8, 169–185. doi: 10.3161/150811006777070893
Downs, N. C., and Sanderson, L. J. (2010). Do bats forage over cattle dung or over cattle? Acta Chiropterologica 12, 349–358. doi: 10.3161/150811010x537936
Duelli, P., and Obrist, M. K. (2003). Regional biodiversity in an agricultural landscape: the contribution of seminatural habitat islands. Basic Appl. Ecol. 4, 129–138. doi: 10.1078/1439-1791-00140
Entwistle, A., Racey, P., and Speakman, J. (1996). Habitat exploitation by a gleaning bat, Plecotus auritus. Philos. Trans. R. Soc. Lond. B 351, 921–931. doi: 10.1098/rstb.1996.0085
Ethier, K., and Fahrig, L. (2011). Positive effects of forest fragmentation, independent of forest amount, on bat abundance in eastern Ontario, Canada. Landsc. Ecol. 26, 865–876. doi: 10.1007/s10980-011-9614-2
Fenton, M. B. (1990). The foraging behaviour and ecology of animal-eating bats. Can. J. Zool. 68, 411–422. doi: 10.1139/z90-061
Fischer, J., Lindenmayer, D. B., and Manning, A. D. (2006). Biodiversity, ecosystem function, and resilience: ten guiding principles for commodity production landscapes. Front. Ecol. Environ. 4, 80–86. doi: 10.1890/1540-9295(2006)004[0080:BEFART]2.0.CO;
Frey-Ehrenbold, A., Bontadina, F., Arlettaz, R., and Obrist, M. K. (2013). Landscape connectivity, habitat structure and activity of bat guilds in farmland-dominated matrices. J. Appl. Ecol. 50, 252–261. doi: 10.1111/1365-2664.12034
Froidevaux, J. S., Laforge, A., Larrieu, L., Barbaro, L., Park, K., Fialas, P. C., et al. (2022). Tree size, microhabitat diversity and landscape structure determine the value of isolated trees for bats in farmland. Biol. Conserv. 267:109476. doi: 10.1016/j.biocon.2022.109476
Froidevaux, J. S. P., Zellweger, F., Bollmann, K., Jones, G., and Obrist, M. K. (2016). From field surveys to LiDAR: shining a light on how bats respond to forest structure. Remote Sens. Environ. 175, 242–250. doi: 10.1016/j.rse.2015.12.038
Fuentes-Montemayor, E., Goulson, D., Cavin, L., Wallace, J. M., and Park, K. J. (2013). Fragmented woodlands in agricultural landscapes: the influence of woodland character and landscape context on bats and their insect prey. Agric. Ecosyst. Environ. 172, 6–15. doi: 10.1016/j.agee.2013.03.019
Gehrt, S. D., and Chelsvig, J. E. (2004). Species-specific patterns of bat activity in an urban landscape. Ecol. Appl. 14, 625–635. doi: 10.1890/03-5013
Grüebler, M. U., Morand, M., and Naef-Daenzer, B. (2008). A predictive model of the density of airborne insects in agricultural environments. Agric. Ecosyst. Environ. 123, 75–80. doi: 10.1016/j.agee.2007.05.001
Habel, J. C., Dengler, J., Janišová, M., Török, P., Wellstein, C., and Wiezik, M. (2013). European grassland ecosystems: threatened hotspots of biodiversity. Biodivers. Conserv. 22, 2131–2138. doi: 10.1007/s10531-013-0537-x
Hanspach, J., Fischer, J., Ikin, K., Stott, J., and Law, B. S. (2012). Using trait-based filtering as a predictive framework for conservation: a case study of bats on farms in southeastern Australia. J. Appl. Ecol. 49, 842–850. doi: 10.1111/j.1365-2664.2012.02159.x
Heim, O., Lenski, J., Schulze, J., Jung, K., Kramer-Schadt, S., Eccard, J. A., et al. (2018). The relevance of vegetation structures and small water bodies for bats foraging above farmland. Basic Appl. Ecol. 27, 9–19. doi: 10.1016/j.baae.2017.12.001
Heim, O., Lorenz, L., Kramer-Schadt, S., Jung, K., Voigt, C. C., and Eccard, J. A. (2017). Landscape and scale-dependent spatial niches of bats foraging above intensively used arable fields. Ecol. Process. 6:24. doi: 10.1186/s13717-017-0091-7
Heim, O., Schroder, A., Eccard, J., Jung, K., and Voigt, C. C. (2016). Seasonal activity patterns of European bats above intensively used farmland. Agric. Ecosyst. Environ. 233, 130–139. doi: 10.1016/j.agee.2016.09.002
Heim, O., Treitler, J. T., Tschapka, M., Knornschild, M., and Jung, K. (2015). The importance of landscape elements for bat activity and species richness in agricultural areas. PLoS One 10:e0134443. doi: 10.1371/journal.pone.0134443
Hodgson, J., Grime, J., Wilson, P., Thompson, K., and Band, S. (2005). The impacts of agricultural change (1963–2003) on the grassland flora of Central England: processes and prospects. Basic Appl. Ecol. 6, 107–118. doi: 10.1016/j.baae.2005.01.009
Jongman, R. H. (2002). Homogenisation and fragmentation of the European landscape: ecological consequences and solutions. Landsc. Urban Plan. 58, 211–221. doi: 10.1016/S0169-2046(01)00222-5
Jung, K., Kaiser, S., Bohm, S., Nieschulze, J., and Kalko, E. K. V. (2012). Moving in three dimensions: effects of structural complexity on occurrence and activity of insectivorous bats in managed forest stands. J. Appl. Ecol. 49, 523–531. doi: 10.1111/j.1365-2664.2012.02116.x
Jung, K., and Threlfall, C. G. (2018). Trait-dependent tolerance of bats to urbanization: a global meta-analysis. Proc. R. Soc. B 285:20181222. doi: 10.1098/rspb.2018.1222
Kalda, O., Kalda, R., and Liira, P. (2015a). Multi-scale ecology of insectivorous bats in agricultural landscapes. Agric. Ecosyst. Environ. 199, 105–113. doi: 10.1016/j.agee.2014.08.028
Kalda, R., Kalda, O., Lõhmus, K., and Liira, J. (2015b). Multi-scale ecology of woodland bat the role of species pool, landscape complexity and stand structure. Biodivers. Conserv. 24, 337–353. doi: 10.1007/s10531-014-0811-6
Kalka, M., and Kalko, E. K. (2006). Gleaning bats as underestimated predators of herbivorous insects: diet of Micronycteris microtis (Phyllostomidae) in Panama. J. Trop. Ecol. 22, 1–10. doi: 10.1017/S0266467405002920
Kalka, M. B., Smith, A. R., and Kalko, E. K. V. (2008). Bats limit arthropods and herbivory in a tropical Forest. Science 320:71. doi: 10.1126/science.1153352
Kelm, D. H., Lenski, J., Kelm, V., Toelch, U., and Dziock, F. (2014). Seasonal bat activity in relation to distance to hedgerows in an agricultural landscape in Central Europe and implications for wind energy development. Acta Chiropterologica 16, 65–73. doi: 10.3161/150811014x683273
Kusch, J., Weber, C., Idelberger, S., and Koob, T. (2004). Foraging habitat preferences of bats in relation to food supply and spatial vegetation structures in a western European low mountain range forest. Folia Zool. 53, 113–128.
Landesamt für Umwelt Brandenburg (2022). Lebensraumtypen [Online]. https://lfu.brandenburg.de/lfu/de/aufgaben/natur/biotopschutz/lebensraumtypen/# (Accessed May 4, 2022).
Leuschner, C., and Ellenberg, H. (2017). Ecology of Central European Non-Forest Vegetation: Coastal to Alpine, Natural to Man-Made Habitats: Vegetation Ecology of Central Europe. Berlin: Springer.
Lindborg, R., Plue, J., Andersson, K., and Cousins, S. A. (2014). Function of small habitat elements for enhancing plant diversity in different agricultural landscapes. Biol. Conserv. 169, 206–213. doi: 10.1016/j.biocon.2013.11.015
Loos, J., Krauss, J., Lyons, A., Föst, S., Ohlendorf, C., Racky, S., et al. (2021). Local and landscape responses of biodiversity in calcareous grasslands. Biodivers. Conserv. 30, 2415–2432. doi: 10.1007/s10531-021-02201-y
MacArthur, R. H., and MacArthur, J. W. (1961). On bird species diversity. Ecology 42, 594–598. doi: 10.2307/1932254
Martin, C. M., Arnett, E. B., Stevens, R. D., and Wallace, M. C. (2017). Reducing bat fatalities at wind facilities while improving the economic efficiency of operational mitigation. J. Mammal. 98, 378–385. doi: 10.1093/jmammal/gyx0005
Matson, P. A., Parton, W. J., Power, A. G., and Swift, M. J. (1997). Agricultural intensification and ecosystem properties. Science 277, 504–509. doi: 10.1126/science.277.5325.504
Meinig, H., Boye, P., Dähne, M., Hutterer, R., and Lang, J. (2020). Rote Liste und Gesamtartenliste der Säugetiere (Mammalia) Deutschlands. Naturschutz und Biologische Vielfalt 170:2. doi: 10.19213/972172
Mendes, E. S., Fonseca, C., Marques, S. F., Maia, D., and Pereira, M. J. R. (2017). Bat richness and activity in heterogeneous landscapes: guild-specific and scale-dependent? Landsc. Ecol. 32, 295–311. doi: 10.1007/s10980-016-0444-0
Miller, B. W. (2001). A method for determining relative activity of free flying bats using a new activity index for acoustic monitoring. Acta Chiropterologica 3, 93–105.
Monck-Whipp, L., Martin, A. E., Francis, C. M., and Fahrig, L. (2018). Farmland heterogeneity benefits bats in agricultural landscapes. Agric. Ecosyst. Environ. 253, 131–139. doi: 10.1016/j.agee.2017.11.001
Müller, J., Brandl, R., Buchner, J., Pretzsch, H., Seifert, S., Strätz, C., et al. (2013). From ground to above canopy—bat activity in mature forests is driven by vegetation density and height. For. Ecol. Manag. 306, 179–184. doi: 10.1016/j.foreco.2013.06.043
Müller, J., Mehr, M., Bässler, C., Fenton, M. B., Hothorn, T., Pretzsch, H., et al. (2012). Aggregative response in bats: prey abundance versus habitat. Oecologia 169, 673–684. doi: 10.1007/s00442-011-2247-y
Norberg, U. M., and Rayner, J. M. (1987). Ecological morphology and flight in bats (Mammalia; Chiroptera): wing adaptations, flight performance, foraging strategy and echolocation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 316, 335–427. doi: 10.1098/rstb.1987.0030
Obrist, M. K., Boesch, R., and Flückiger, P. F. (2004). Variability in echolocation call design of 26 Swiss bat species: consequences, limits and options for automated field identification with a synergetic pattern recognition approach. Mammalia 68, 307–322. doi: 10.1515/mamm.2004.030
Oksanen, J., Blanchet, F. G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D., et al. (2020). “vegan: community ecology package". R package version 2.5-7 ed. Available at: https://CRAN.R-project.org/package=vegan (Accessed May 05, 2022).
Olimpi, E. M., and Philpott, S. M. (2018). Agroecological farming practices promote bats. Agric. Ecosyst. Environ. 265, 282–291. doi: 10.1016/j.agee.2018.06.008
Parsons, S., and Jones, G. (2000). Acoustic identification of twelve species of echolocating bat by discriminant function analysis and artificial neural networks. J. Exp. Biol. 203, 2641–2656. doi: 10.1242/jeb.203.17.2641
Pfalzer, G., and Kusch, J. (2003). Structure and variability of bat social calls: implications for specificity and individual recognition. J. Zool. 261, 21–33. doi: 10.1017/S0952836903003935
Poschlod, P., and Wallis de Vries, M. F. (2002). The historical and socioeconomic perspective of calcareous grasslands—lessons from the distant and recent past. Biol. Conserv. 104, 361–376. doi: 10.1016/S0006-3207(01)00201-4
Put, J. E., Fahrig, L., and Mitchell, G. W. (2019). Bats respond negatively to increases in the amount and homogenization of agricultural land cover. Landsc. Ecol. 34, 1889–1903. doi: 10.1007/s10980-019-00855-2
QGIS Development Team (2021). “QGIS geographic information system”. Open source geospatial foundation project. Available at: http://qgis.osgeo.org (Accessed February 03, 2021).
R Core Team (2020). "R: A language and environment for statistical computing. R foundation for statistical computing". (Vienna, Austria). Available at: www.r-project.org (Accessed February 03, 2021).
Rasran, L., Diener, A., Pachinger, B., and Bernhardt, K.-G. (2018). Diversity of flower visiting insects in dry grasslands and vineyards close to the City of Vienna with special focus on wild bees. Sociobiology 65, 603–611. doi: 10.13102/sociobiology.v65i4.3370
Raven, P. H., and Wagner, D. L. (2021). Agricultural intensification and climate change are rapidly decreasing insect biodiversity. Proc. Natl. Acad. Sci. 118:e2002548117. doi: 10.1073/pnas.2002548117
Rösch, V., Tscharntke, T., Scherber, C., and Batáry, P. (2015). Biodiversity conservation across taxa and landscapes requires many small as well as single large habitat fragments. Oecologia 179, 209–222. doi: 10.1007/s00442-015-3315-5
Rosenberg, D. K., Noon, B. R., and Meslow, E. C. (1997). Biological corridors: form, function, and efficacy. Bioscience 47, 677–687. doi: 10.2307/1313208
Russo, D., and Jones, G. (2002). Identification of twenty-two bat species (Mammalia: Chiroptera) from Italy by analysis of time-expanded recordings of echolocation calls. J. Zool. 258, 91–103. doi: 10.1017/S0952836902001231
Russo, D., and Voigt, C. C. (2016). The use of automated identification of bat echolocation calls in acoustic monitoring: a cautionary note for a sound analysis. Ecol. Indic. 66, 598–602. doi: 10.1016/j.ecolind.2016.02.036
Schloerke, B., Cook, D., Larmarange, J., Briatte, F., Marbach, M., Thoen, E., et al. (2021). ‘GGally’: Extension to 'ggplot2'. Available at: https://CRAN.R-project.org/package=GGally (Accessed April 24, 2022).
Schnitzler, H.-U., and Kalko, E. K. (2001). Echolocation by insect-eating bats: we define four distinct functional groups of bats and find differences in signal structure that correlate with the typical echolocation tasks faced by each group. Bioscience 51, 557–569. doi: 10.1641/0006-3568(2001)051[0557:EBIEB]2.0.CO;2
Schuldt, A., Ebeling, A., Kunz, M., Staab, M., Guimarães-Steinicke, C., Bachmann, D., et al. (2019). Multiple plant diversity components drive consumer communities across ecosystems. Nat. Commun. 10, 1–11. doi: 10.1038/s41467-019-09448-8
Söderström, B., Svensson, B., Vessby, K., and Glimskär, A. (2001). Plants, insects and birds in semi-natural pastures in relation to local habitat and landscape factors. Biodivers. Conserv. 10, 1839–1863. doi: 10.1023/A:1013153427422
Stahlschmidt, P., and Brühl, C. A. (2012). Bats at risk? Bat activity and insecticide residue analysis of food items in an apple orchard. Environ. Toxicol. Chem. 31, 1556–1563. doi: 10.1002/etc.1834
Stahlschmidt, P., Hahn, M., and Brühl, C. A. (2017). Nocturnal risks-high bat activity in the agricultural landscape indicates potential pesticide exposure. Frontiers in environmental. Science 5:62. doi: 10.3389/fenvs.2017.00062
Stahlschmidt, P., Pätzold, A., Ressl, L., Schulz, R., and Brühl, C. A. (2012). Constructed wetlands support bats in agricultural landscapes. Basic Appl. Ecol. 13, 196–203. doi: 10.1016/j.baae.2012.02.001
Starik, N., Göttert, T., Heitlinger, E., and Zeller, U. (2018). Bat Community responses to structural habitat complexity resulting from management practices within different land use types—a case study from north-eastern Germany. Acta Chiropterologica 20, 387–405. doi: 10.3161/15081109ACC2018.20.2.010
Stoate, C., Boatman, N. D., Borralho, R., Carvalho, C. R., De Snoo, G., and Eden, P. (2001). Ecological impacts of arable intensification in Europe. J. Environ. Manag. 63, 337–365. doi: 10.1006/jema.2001.0473
Tilman, D., Fargione, J., Wolff, B., D'antonio, C., Dobson, A., Howarth, R., et al. (2001). Forecasting agriculturally driven global environmental change. Science 292, 281–284. doi: 10.1126/science.1057544
Toffoli, R. (2016). The importance of linear landscape elements for bats in a farmland area: the influence of height on activity. J. Landsc. Ecol. 9, 49–62. doi: 10.1515/jlecol-2016-0004
Treitler, J. T., Heim, O., Tschapka, M., and Jung, K. (2016). The effect of local land use and loss of forests on bats and nocturnal insects. Ecol. Evol. 6, 4289–4297. doi: 10.1002/ece3.2160
Tscharntke, T., Klein, A. M., Kruess, A., Steffan-Dewenter, I., and Thies, C. (2005). Landscape perspectives on agricultural intensification and biodiversity–ecosystem service management. Ecol. Lett. 8, 857–874. doi: 10.1111/j.1461-0248.2005.00782.x
Tscharntke, T., Steffan-Dewenter, I., Kruess, A., and Thies, C. (2002). Contribution of small habitat fragments to conservation of insect communities of grassland–cropland landscapes. Ecol. Appl. 12, 354–363. doi: 10.1890/1051-0761(2002)012[0354:COSHFT]2.0.CO;2
Verboom, B., and Huitema, H. (1997). The importance of linear landscape elements for the pipistrelle Pipistrellus pipistrellus and the serotine bat Eptesicus serotinus. Landsc. Ecol. 12, 117–125. doi: 10.1007/Bf02698211
Vrahnakis, M., Janišová, M., Rūsiņa, S., Török, P., Venn, S., and Dengler, J. (2013). “The European dry grassland group (EDGG): stewarding Europe’s most diverse habitat type” in Steppenlebensräume Europas, ed. Baumbach, H, and Pfützenreuter, S. (Thüringen: Ministry for agriculture, Foerstry, Environment and Nature Protection), 417–434.
Wallis de Vries, M. F., and Van Swaay, C. A. (2009). “Grasslands as habitats for butterflies in Europe” in Grasslands in Europe, ed. Veen, P., Jedderson R., de Scmidt, J., and van der Straaten, J. (Leiden: KNNV publishing), 26–34. doi: 10.1163/9789004278103_004
Williams-Guillén, K., Perfecto, I., and Vandermeer, J. (2008). Bats limit insects in a neotropical agroforestry system. Science 320:70. doi: 10.1126/science.1152944
Wilson, J. B., Peet, R. K., Dengler, J., and Pärtel, M. (2012). Plant species richness: the world records. J. Veg. Sci. 23, 796–802. doi: 10.1111/j.1654-1103.2012.01400.x
Wood, E. M., Pidgeon, A. M., Radeloff, V. C., and Keuler, N. S. (2013). Image texture predicts avian density and species richness. PLoS One 8:e63211. doi: 10.1371/journal.pone.0063211
Young, H. S., McCauley, D. J., Galetti, M., and Dirzo, R. (2016). Patterns, causes, and consequences of anthropocene defaunation. Annu. Rev. Ecol. Evol. Syst. 47, 333–358. doi: 10.1146/annurev-ecolsys-112414-054142
Zeale, M. R., Davidson-Watts, I., and Jones, G. (2012). Home range use and habitat selection by barbastelle bats (Barbastella barbastellus): implications for conservation. J. Mammal. 93, 1110–1118. doi: 10.1644/11-MAMM-A-366.1
Zulka, K. P., Abensperg-Traun, M., Milasowszky, N., Bieringer, G., Gereben-Krenn, B.-A., Holzinger, W., et al. (2014). Species richness in dry grassland patches of eastern Austria: a multi-taxon study on the role of local, landscape and habitat quality variables. Agric. Ecosyst. Environ. 182, 25–36. doi: 10.1016/j.agee.2013.11.016
Keywords: dry grasslands, bats (Chiroptera), agricultural intensification, landscape, structural richness, acoustic monitoring, habitat fragments
Citation: Ewert SP, Knörnschild M, Jung K and Frommolt K-H (2023) Structurally rich dry grasslands – Potential stepping stones for bats in open farmland. Front. Ecol. Evol. 11:995133. doi: 10.3389/fevo.2023.995133
Edited by:
Péter Török, University of Debrecen, HungaryReviewed by:
Katherina Ng, Australian National University, AustraliaCopyright © 2023 Ewert, Knörnschild, Jung and Frommolt. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sophie P. Ewert,  U29waGllLmV3ZXJ0QHdlYi5kZQ==
U29waGllLmV3ZXJ0QHdlYi5kZQ==
†ORCID: Mirjam Knörnschild, https://orcid.org/0000-0003-0448-9600
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.