 Jérôme Belliard
Jérôme Belliard- 1Université Paris-Saclay, INRAE, UR HYCAR, Paris, France
- 2Independent Researcher, Cherbourg, France
- 3Union des Fédérations de pêche et de protection des milieux aquatiques du Bassin Seine Normandie, Montreuil, France
The spread of non-native species is nowadays recognized as a major threat to the biodiversity of freshwater ecosystems. However, for a very long time the introduction and acclimatization of new species has been perceived mainly as a source of wealth for human societies. Here, we examined the establishment of non-native fish species in the Seine River basin from a historical perspective by adopting a twofold approach. In a first step, at the whole basin scale, considering various written and archeological sources, we traced the chronology, over the last millennium, of the establishments of non-native species. In a second step, by analyzing fish monitoring from several hundred sites covering the diversity of rivers and streams, we examined the changes in numbers and abundance of non-native species in local fish communities over the last three decades. The first documented species introduction dates back to the 13th century but it is from the middle of the 19th century that the introduction attempts accelerated. Today, these introductions have reached an unprecedented level and 46% of the species recorded in the basin are non-native. During the last three decades, non-native species have continued to increase within fish communities both in terms of number of species and abundance of individuals. The most pronounced increases are noted on large rivers and sites where anthropic pressures are strong. Waterways connecting European basins, globalization of trade, and ongoing climate change provide a general background suggesting that the increase in the proportion of non-native species in the fish communities of the Seine River basin is likely to continue for several decades.
Introduction
The establishment of non-native species is now pervasive around the world and has progressively emerged as one of the major threats to biodiversity (IPBES, 2019; Pyšek et al., 2020). The incidences of non-native species on ecosystems can be numerous and extensive, especially when they exhibit a high degree of invasiveness (Mooney and Cleland, 2001; Simberloff, 2011). Non-native species can cause massive declines in some native species through competitive or predation interactions, even leading to their extinction (Corbin and D’Antonio, 2004; Bellard et al., 2016; Doherty et al., 2016). They can alter environment and consequently deeply reshape pre-existing biological communities and ecosystems (Dassonville et al., 2008; Emery-Butcher et al., 2020). They can induce genetic modifications in native populations through hybridization processes (Rhymer and Simberloff, 1996; Muhlfeld et al., 2009). They can be vectors for pathogens and parasites that may adversely affect native species (Lymbery et al., 2014). They can lead to deep restructuring in food webs notably through cascading effects (David et al., 2017). Beyond these, non-native species are also likely to generate socio-economic damages, for example by reducing crop production or fisheries yields, disrupting transport or water supply infrastructures or directly affecting human health (Pejchar and Mooney, 2009; Mazza et al., 2014; Pyšek et al., 2020).
These issues are especially acute in freshwater ecosystems, where non-native species seem to generate more significant ecological and socioeconomic impacts compared to their terrestrial counterparts (Sala et al., 2000; Moorhouse and Macdonald, 2015). Yet, while the ascertained or anticipated negative consequences of species introductions are increasingly highlighted, this has not always been the case, and for a long time the acclimatization of new non-native species was primarily perceived as a source of wealth for human societies, by providing greater production of livestock and crops, developing natural environments and as recreational resources (Anderson, 1992; Gozlan, 2008). This applies particularly to freshwater ecosystems where, very early on, some fish species were translocated outside their native range, in association with the emergence and development of fish farming (Hoffmann, 1995, 2005; Balon, 2004).
In this article we investigated the long-term dynamics of non-native fish establishment in the Seine River basin, a Western European territory where freshwater ecosystems were early used and transformed by human societies. For this purpose, we adopted an approach considering two nested and complementary temporal and spatial scales to address this issue. In a first step, considering the whole basin, we traced the chronology, over the last millennium, of the establishments or attempted introductions of non-native fish species by compiling various written and archeological sources. This approach provides a comprehensive but coarse overview of the occurrence of non-native species over the past centuries in the basin due to the difficulty in obtaining quantitative and spatially extensive information for the earliest periods. In a second step, by analyzing fish monitoring data from several hundred sites covering the diversity of rivers and streams within the Seine River basin, we examined the changes in numbers and abundance of non-native species in local fish communities over the last three decades. By providing accurate quantitative and spatialized data, this second approach allows us to detail and add more nuance to the temporal trends reported by the longer-term approach by focusing on the most recent period.
Over the past millennium, we hypothesized that the establishment of non-native fish species has steadily increased and even accelerated. For the most recent period, and in line with the previous hypothesis, we postulated that the increase in non-native species within local communities also amplified during the last three decades despite increasing regulations to restrict their introduction. Further, we believe that these trends could vary greatly among rivers, with stronger increases on larger rivers and those subject to heavy anthropogenic pressures, potentially more prone to non-native species establishment.
Materials and Methods
The Seine River Basin
The Seine River basin extends over more than 76,000 km2 in the northwest of France. Its average flow is nowadays 450 m3 s–1 in the upstream part of its estuary, with a pluvial/oceanic hydrological regime. More than 95% of the basin is within a large sedimentary basin at low elevation (<500 m) (Flipo et al., 2021). The natural conditions in the basin facilitated an early development of a large human population. As early as the Middle Ages, man had a strong imprint on watercourses, in particular with the widespread establishment of water mills, the expansion of fish ponds, and later the construction of canals connecting the Seine basin with its neighboring basins (Billen et al., 2007; Leuven et al., 2009). The human impact increased considerably from the 19th century onward with the industrialization, the systematic channelization of large rivers and the regulation of their flows, the intensification of agricultural practices and the strong growth of human populations, particularly in the large urban areas located around Paris and its suburbs and along the major waterways (Meybeck et al., 2018; Flipo et al., 2021). The total human population of the Seine basin increased from ca. 3 million people in the 13th century, to 8 million around 1900 and to 16.7 in 2015 (Billen et al., 2009; Flipo et al., 2021).
Comprehensive Approach: The Whole Basin During the Last Millennium
In order to list the non-native species recorded in the basin and document their timing of introduction, we used historical written and archeological sources. In particular we explore the CHIPS database which compiles information retrieved from the analysis of historical archives and includes data related to past fish species distribution in the Seine River basin for the period between the 16th and the mid-20th century (Beslagic et al., 2013). More recent complementary sources of information have been mobilized (species distribution atlases, mentions from anglers, reports, etc.) for the most recently introduced species. Occasionally, we also integrated archeological data to consolidate or to complement the information provided by written sources (Beslagic and Belliard, 2014). Thereafter, following Copp et al. (2005) we used the word “introduction” in a broad sense, meaning the appearance of a species in a new place because of mechanical transfer (intentional or unintentional) by man, or after overcoming or removing colonization barriers (e.g., following the creation of a canal connecting initially isolated basins).
For each species we determined the date of introduction. For some species the available sources did not allow to establish a precise year of introduction or observation. In such cases, the date was specified in a more approximate way (generally the corresponding decade). In some cases, the different sources available provided conflicting information concerning the possible dates (for example, between archeological data, mentions to voluntary attempts of introductions and first observations of individuals in the wild). In such cases we privileged the first observations of individuals in rivers or lakes to establish the date of introduction. We also recorded whether the species was still currently present in the basin, if it was naturalized (self-sustaining population(s) with successful natural reproduction) or not and its biogeographical origin (European vs. non-European origin). For species formerly introduced but no longer occurring in the basin, in the absence of precise information, we considered that their extirpation took place in the decade following their last record.
Detailed Approach: Local Communities in the Last Three Decades
To investigate in greater detail the most recent changes in distribution and abundance of non-native species in different watercourses, we examined the results of electrofishing carried out between 1990 and 2018 in the Seine River basin (Observatoire des Poissons du bassin Seine Normandie).1 We selected sites with samples available for at least two different years during this period, resulting in a dataset including 3,412 fish samples from 453 different sites. The number of fish samples varied widely among sites, ranging from 2 for 144 sites to 44 for the most-monitored site.
Electrofishing was carried out at low flow periods and followed the guidelines of European standard (CEN, 2003). The sampling protocols implemented could vary between sites and occasionally could change over time for a given site. For each sample, the sampling conditions were described by three qualitative variables: (1) the sampling strategy (3 modalities: entire sampling of the study reach; partial sampling of habitat areas with heterogeneous sizes; point abundance sampling), (2) the sampling mode (3 modalities: by wading; from a boat; mix of wading and boat sampling), and (3) the number of successive passes (3 modalities: 1 passage; 2 successive passages; 3 successive passages). In addition, for each sample, the sampled area (in m2) was also determined.
The sites were characterized by the following two variables: the catchment area (CA) (km2) and the density of inhabitants in the municipality where the site was located (HDens) (inhab./km2). The densities of inhabitants were calculated for the year 2015 and assigned regardless of the sampling year. By doing so, we assumed that the density of inhabitants had changed relatively little for a given site over the last 30 years and that the most significant variations in this variable were related to differences among sites. Thereafter CA and HDens were used as proxies for river size and local anthropic pressure intensity, respectively. Across all sites in our dataset, the CA varied between 1.5 and 72,462.4 km2 and the human population density between 1 and 21,146 inhab./km2.
We retrieved a set of metrics from each sample to be used as indicator variables for analyzing the temporal trends of non-native fish in local communities: number of non-native species (Snnat), proportion of non-native species (S%nnat) (i.e., number of non-native species/total number of species), absolute abundance of individuals from non-native species (Nnnat) and proportion of individuals from non-native species (N%nnat) (i.e., number of individuals from non-native species/total number of individuals) in the sample.
We used generalised linear mixed models (GLMMs) to explore changes in non-native species metrics over years (Bolker et al., 2009; Zuur et al., 2009). In these models we considered both the site and the sampling conditions as random effects to account for the non-independence of observations within sampling sites and protocols. A reasonably high number of levels is recommended for random variables to properly estimate their associated variance (Crawley, 2002), which was clearly not the case with each of our initial sampling condition variables (three levels for each variable). We therefore constructed a new variable (Samp) by crossing the three initial variables, reflecting the various sampling modalities represented in our dataset. Note that since some modalities of the former variables were never associated, this new variable only included nine different levels.
In a first step we analyzed the variations in the different non-native species metrics with GLMMs including the sampling year (Year) and its quadratic term (Year2) as fixed effects, and two crossed random effects, the site identity (Site) and the sampling mode (Samp). This step was designed to detect overall temporal trends in non-native species variables over the period 1990–2018. Term Year2 was used to test a possible non-monotonic temporal trend.
In a second step we extended the previous models by adding CA and HDens, their quadratic terms (CA2 and HDens2) and their respective interactions with Year as fixed effects. This step was conducted to test whether temporal trends in the non-native species variables could vary according to site attributes (stream size or level of anthropogenic pressures).
For all GLMMs, the selection of the most relevant fixed effects was based on AIC values. A given fixed effect was included in the final model only if it resulted in a decrease in AIC value >2 in comparison with the alternative model excluding this fixed effect.
We implemented binomial GLMMs to predict S%nnat and N%nnat and Poisson GLMMs to predict Snnat and Nnnat. For models predicting Nnnat, we added the sample area as offset variable (log-transformed because Poisson GLMM uses logarithmic link) to account for its heterogeneity among samples (Zuur et al., 2009). Prior to their use in the models, the fixed effects variables were standardized and transformed to avoid problems in parameters computation during model setting. The year was centered to the 2004 value and then divided by the total number of sampling years (as a result, the transformed variable varied between −1, corresponding to 1990, and +1, corresponding to 2018). CA and density of inhabitants were first log10-transformed and then centered and standardized (leading to a final variable with a mean value of 0 and a standard deviation of 1). GLMMs were implemented using the R packages lme4 version 1.1–26 (Bates et al., 2015) and MuMIn version 1.43.17 (for marginal and conditional pseudo-R2 calculation).
Results
Long Term Trends
We identified 37 different non-native fish species established in the Seine River basin or which experienced introduction attempts (Supplementary Table 1). Among them, 28 species are still recorded from 2000 to 2020, representing 46% of all species currently present in the entire catchment. However, 6 of these 28 species (Acipenser baerii, Acipenser ruthenus, Ctenopharyngodon idella, Hypophthalmichthys molitrix, Oncorhynchus mykiss, and Salvelinus fontinalis) seem to be maintained only by recurrent release of new individuals (no known self-sustaining populations).
The common carp (Cyprinus carpio), now widespread, is the first documented introduced non-native species which was spread throughout the basin from the 13th century onward with development of fish farming. Intentional or accidental introductions of non-native species remained fairly low for a long time and experienced a sharp acceleration during the second half of the 19th century (Figures 1, 2). After a phase of decline in the first half of the 20th century especially between the two World Wars, these introductions resumed with even greater intensity in the second half of the 20th century and further in the last two decades (Figure 2), with a mean number of four new species arriving per decade for the period 2000–2020 (3.5 species if we consider the species still present in 2020) (Figure 1). It should be noted that many species introduced during the second half of the 19th century did not establish and were rapidly extirpated, which was not the case later on and particularly during the second wave of introductions starting in the middle of the 20th century (Figure 2). Interestingly, nearly 60% of the species introduced during the 19th century came from outside Europe (mainly North America), whereas since the 20th century they only represented 38%.
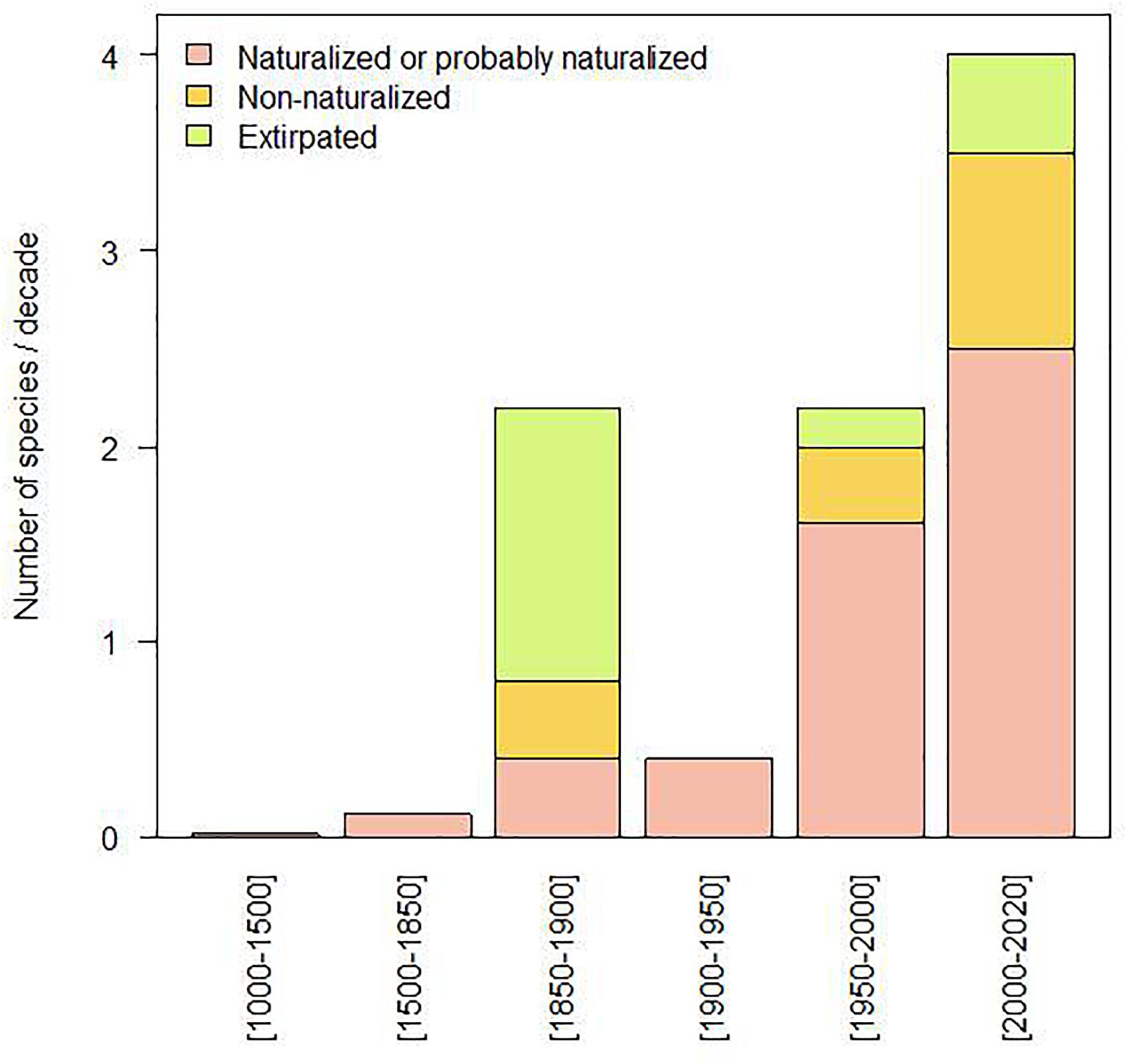
Figure 1. Number of new non-native species recorded per decade in the Seine River basin for successive periods. Species are distinguished according to whether they have established self-sustaining populations (naturalized), remain present through repeated introductions of new specimens (non-naturalized) or are no longer observed (extirpated).
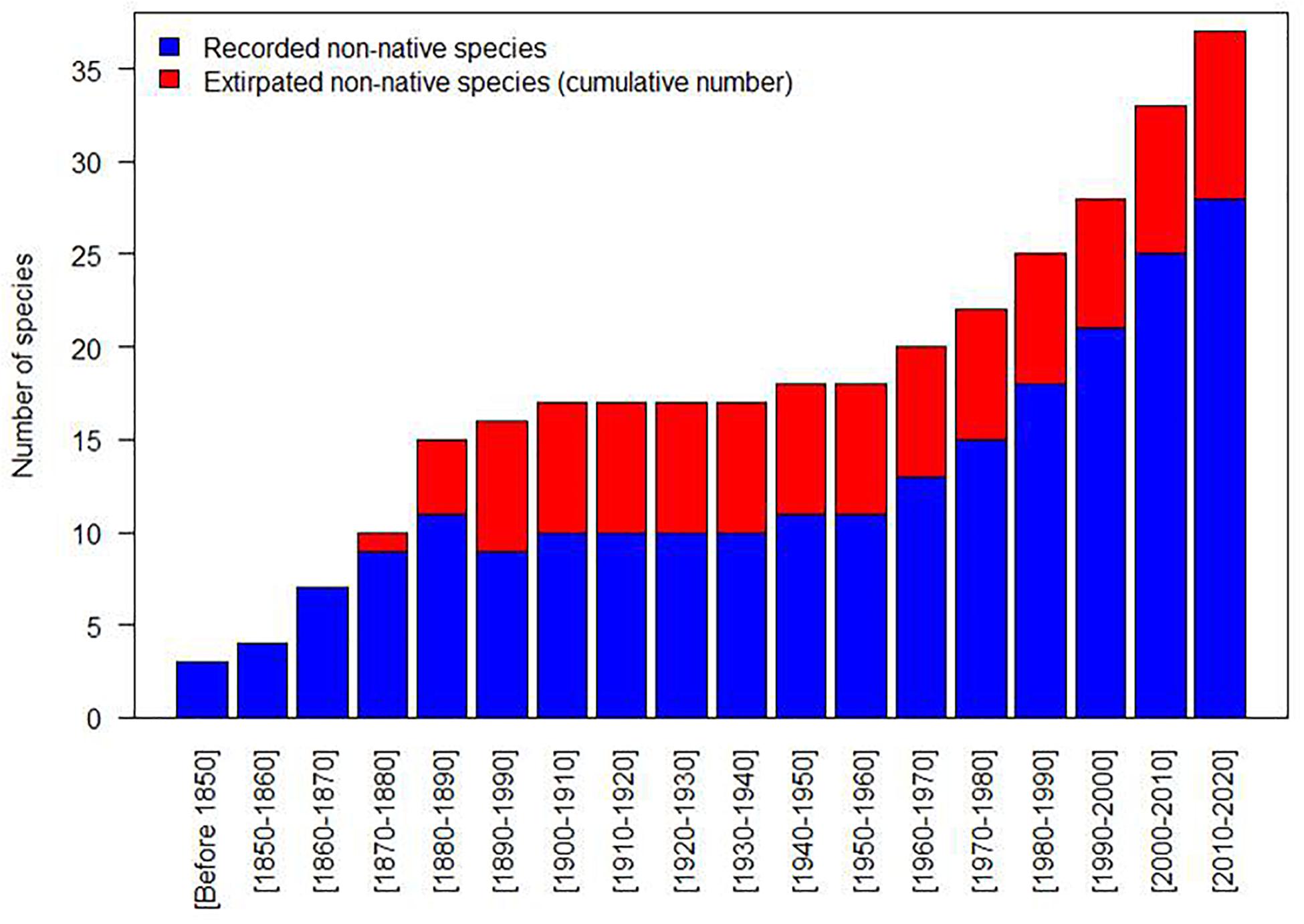
Figure 2. Number of recorded non-native species and cumulative number of extirpated non-native species per decade between 1850 and 2020.
Trends of the Last Decades
Twenty-two different non-native species were identified in electrofishing samples, representing a substantial part of the overall non-native species recorded in the entire basin. The importance of non-native species varied greatly between stations and years. 70% of the study sites hosted non-native species at least one year. The maximum number of non-native species in a sample was 6 and the maximum number of individuals caught was 2.8/m2. The maximum proportions of non-native fish in samples were 50 and 90% when considering species number or abundance of individuals, respectively.
Simple models showed a significant trend toward increase in the number and abundance of non-native species in fish communities over time (Table 1 and Figure 3). However, the inclusion of Year2 in the models (with a negative coefficient) suggests that this trend slowed down, and even slightly reversed in the case of number of non-native species, in recent years (Figure 3). In a typical situation, models predicted an increase from 0.40 to 0.67 species (i.e., an increase of 70% compared to the initial value) and from 0.52 to 2.28 individuals per 1,000 m2 sampled (i.e., an increase of 340% compared to the initial value) between 1990 and 2018. Although there was a trend toward an increase in the number and abundance of native species during the same period (results not shown), our models also proved that the proportion of non-native species and the proportion of individuals of non-native species increased significantly over time with, again, a slight reversal of the trend in the last decade (Table 1 and Figure 3). These changes were nevertheless very limited in magnitude for proportion of species (predicted values of 0.056 and 0.072 for 1990 and 2018, respectively) and even more for proportion of individuals (predicted values of 0.0030 and 0.0034 for 1990 and 2018, respectively). Marginal pseudo-R2 of models showed that temporal fixed effects explained a very small, although significant, part of the variance of the metrics (2.6% at most), especially for metrics representing proportions (Table 1).
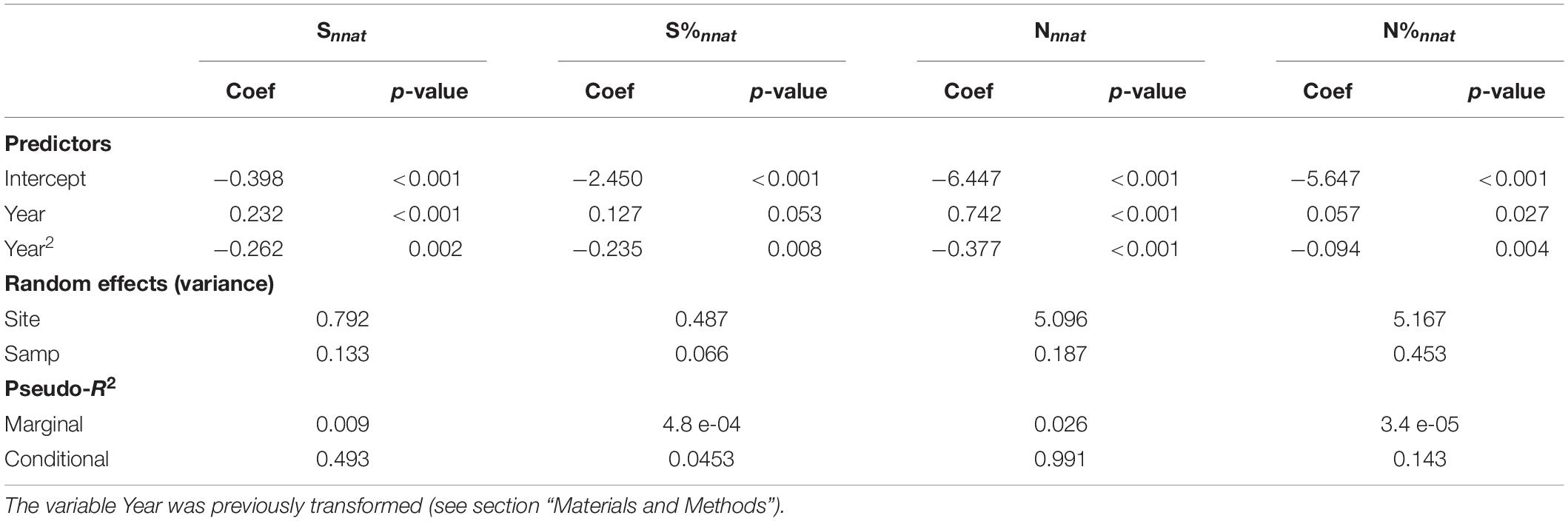
Table 1. Summary of the GLMMs examining variations in the four metrics describing non-native species richness and abundance according to the sampling year (Year and its quadratic term Year2) Site identity (Site) and sampling mode (Samp) are used as Random effects.
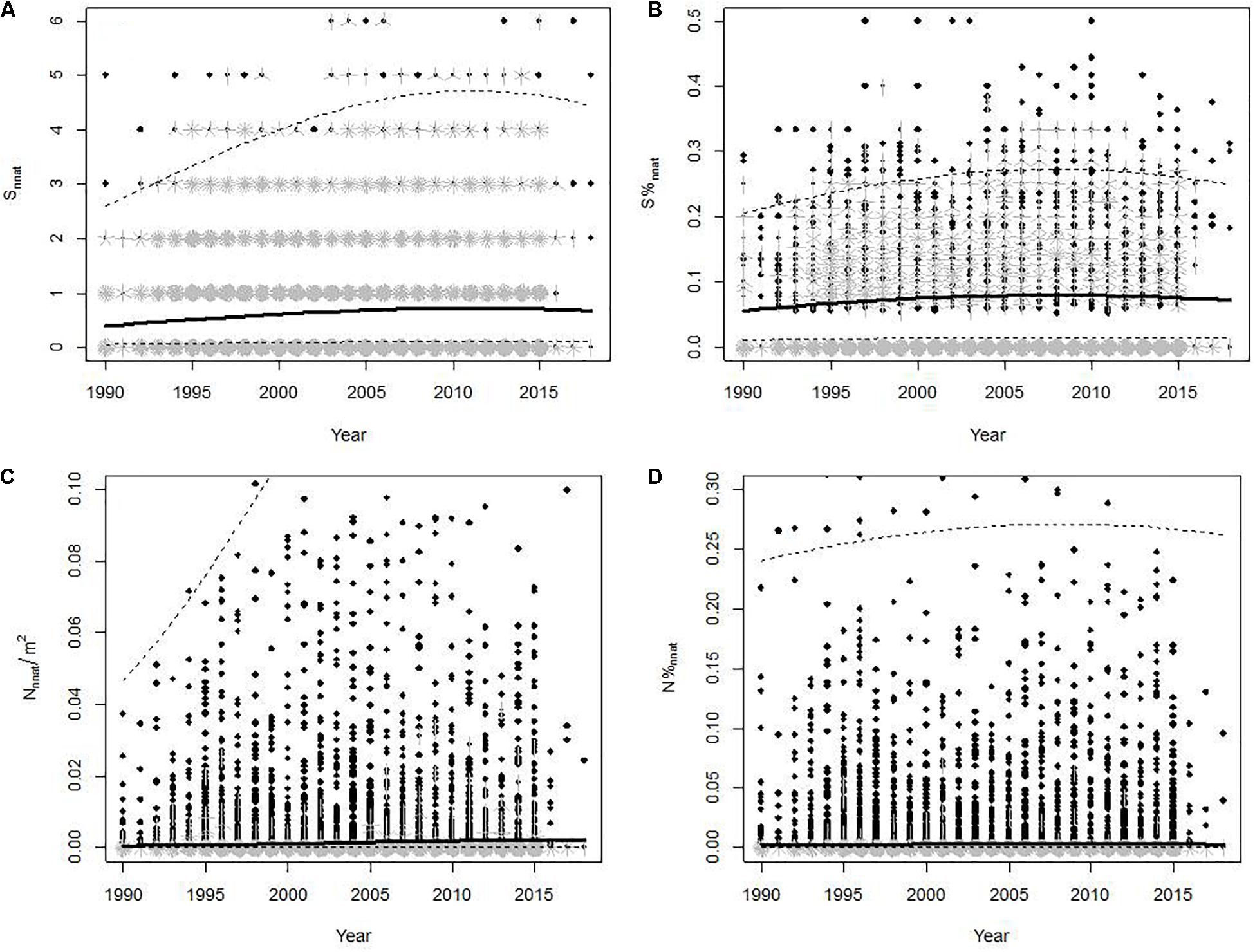
Figure 3. Variations in non-native species number (A), proportion of non-native species (B), abundance of individuals from non-native species (per m2) (C) and proportion of individuals from non-native species (D) according to the sampling year. When several observations are superimposed, each star branch represents one point. The bold lines correspond to the values predicted by the GLMMs presented in Table 2. The dashed lines reflect the variability of predicted values due to site and sampling random effects (95% interval). In panels (C,D) the scale of the vertical axis has been narrowed to better display the model’s predictions.
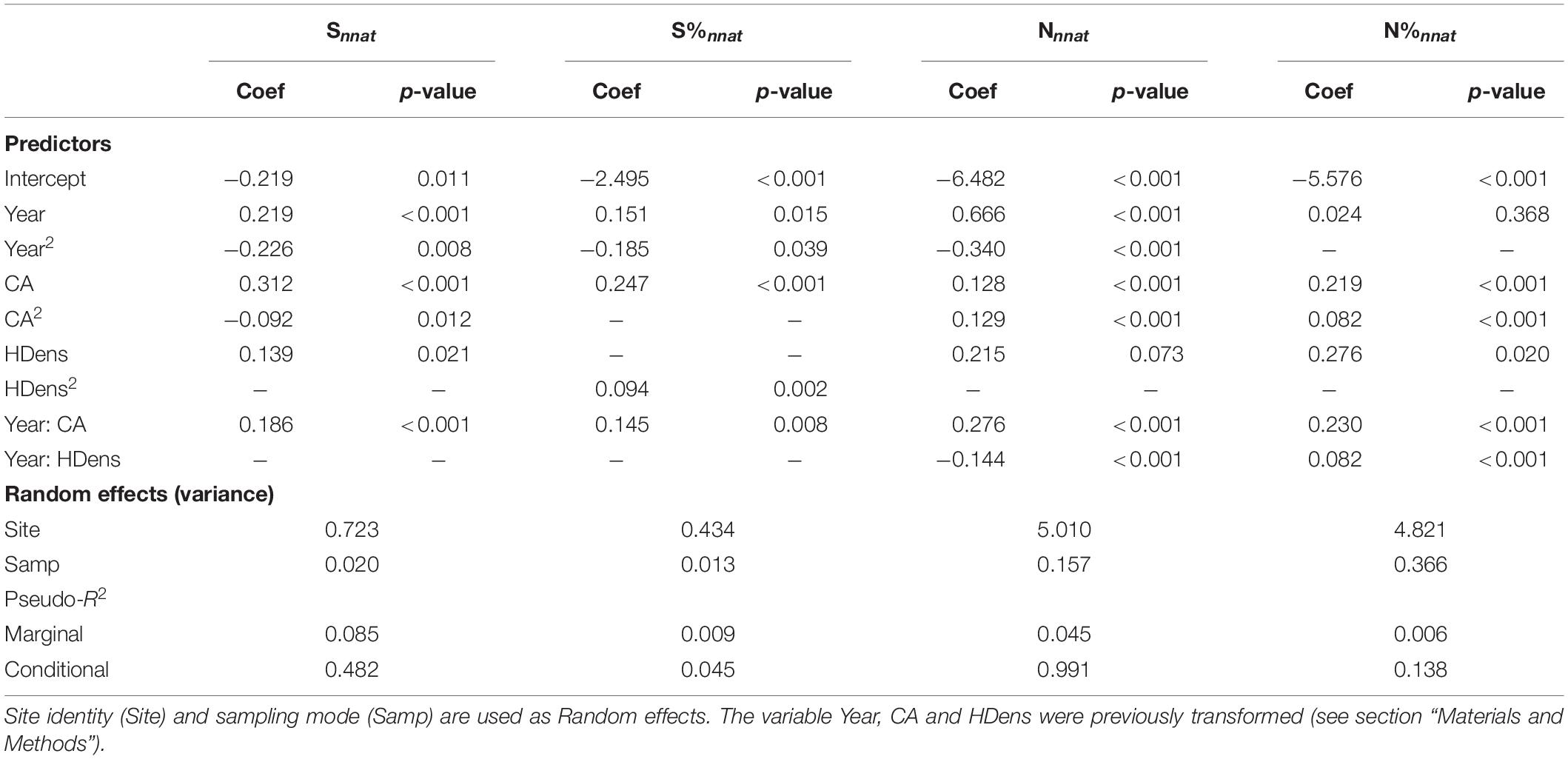
Table 2. Summary of the GLMMs examining variations in the four metrics describing non-native species richness and abundance according to the sampling year (Year and Year2), the catchment area (CA and CA2), the density of inhabitants (HDens and HDens2), and their interactions.
Environmental parameters and their interactions with year explained a significant part of variation in the importance of non-native species within fish communities. Overall, non-native species metrics exhibited higher values as CA and site human population density increased (Figures 4–6). However, this general pattern was subject to occasional exceptions such as the case of the abundance and proportion of individuals from non-native species which did not show clear variations with the CA at the beginning of the monitoring (Figure 5). The importance of interaction effects between year and CA (all models) and year and density of inhabitants (Nnnat and N%nnat models only) indicated that the temporal dynamics of non-native species varied strongly according to the environmental characteristics of the sites, with a stronger progressions when river size and level of anthropization increased (Figures 4–6). Model responses even suggested a slight decline in all non-native species metric over time for smaller watersheds (a few tens of km2 or less) (Figures 4, 5).
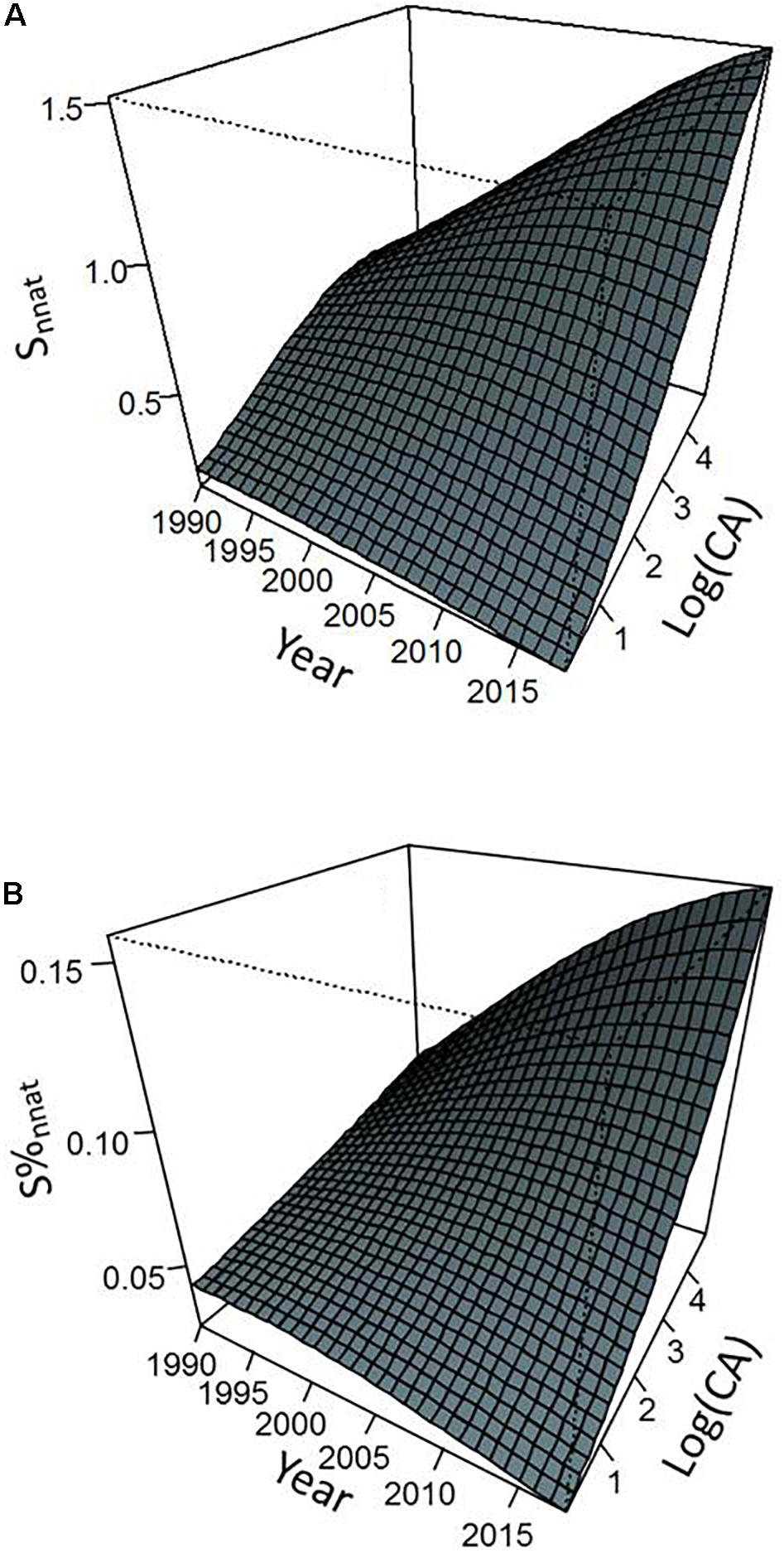
Figure 4. Predicted values of non-native species number (A) and proportion of non-native species (B) as a function of sampling year and catchment area from the models presented in Table 2. Predictions were calculated by considering a density of inhabitants equal to the average value observed in our data set.
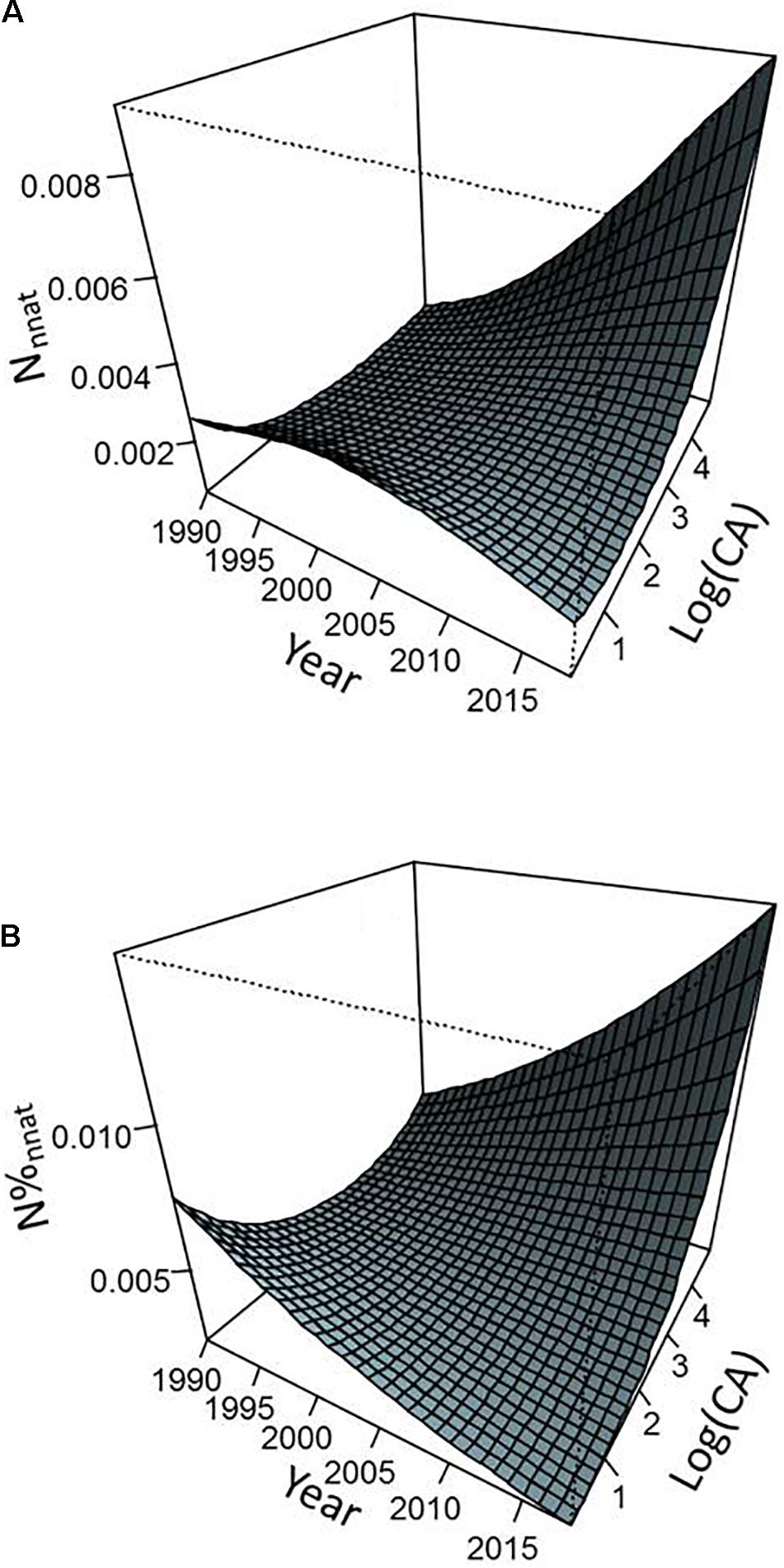
Figure 5. Predicted values of abundance of individuals from non-native species (per m2) (A) and proportion of individuals from non-native species (B) as a function of sampling year and catchment area from the models presented in Table 2. Predictions were calculated by considering a density of inhabitants equal to the average value observed in our data set.
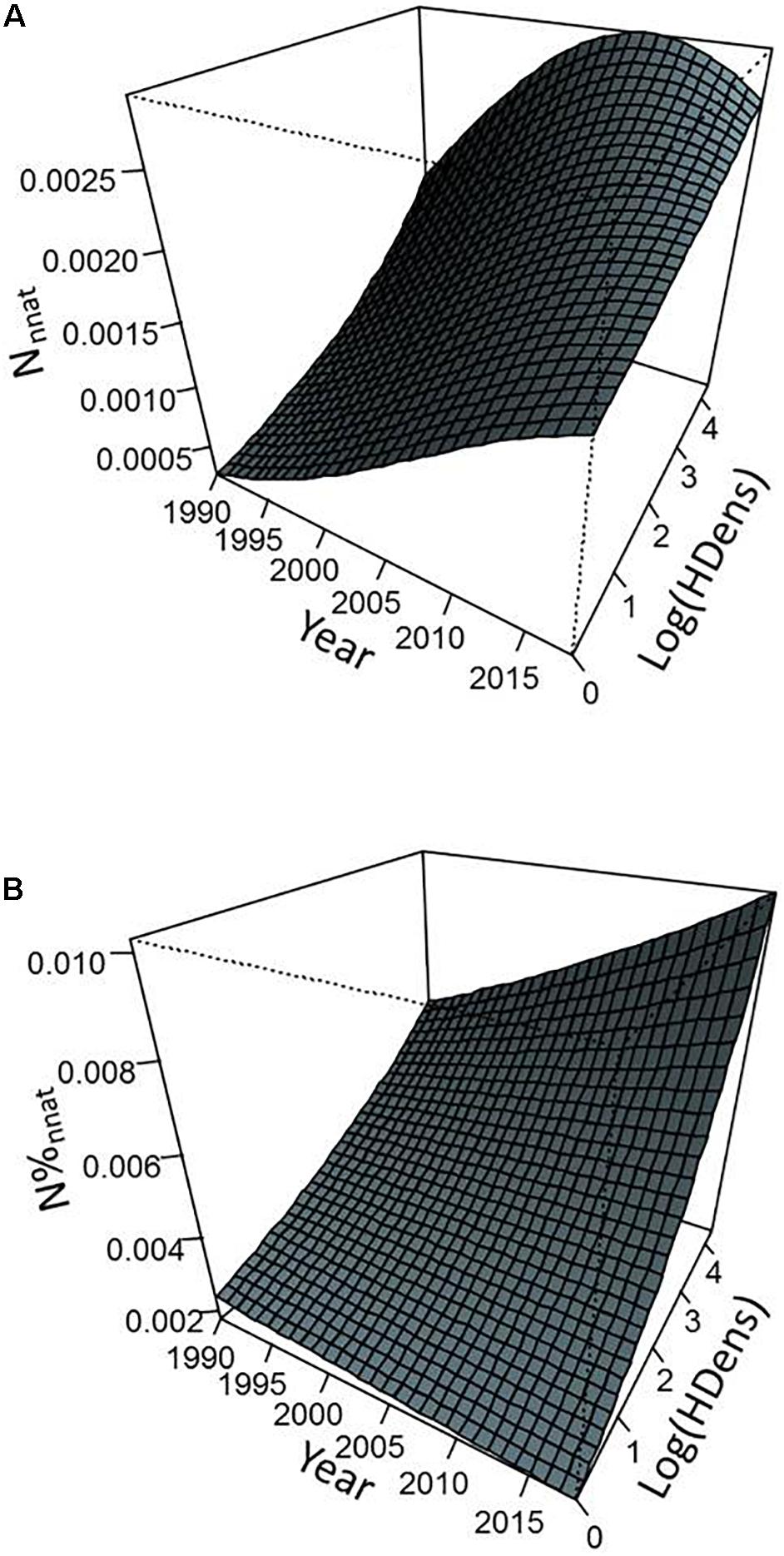
Figure 6. Predicted values of abundance of individuals from non-native species (per m2) (A) and proportion of individuals from non-native species (B) as a function of sampling year and density of inhabitants from the models presented in Table 2. Predictions were calculated by considering a catchment area equal to the average value observed in our data set.
Discussion
Long Term Evolution at Basin Scale
For a long time, the fish biology literature mentioned the precursory role of Roman civilization in the diffusion of non-native fish species throughout Europe, and particularly common carp (Balon, 2004; Keith et al., 2011). By crossing historical and archeological sources, Hoffmann questioned this standpoint and demonstrated that the diffusion of the carp in Europe took place later and progressively extended during medieval times in connection with the development of pond fish farming (Hoffmann, 1995, 1996). More specifically, for the Seine River basin the first written and archeological mentions date back to the 13th century and suggest that the carp is most likely the first fish species that was intentionally introduced into the watershed (Beslagic, 2013). Furthermore, it is worth noting that the native status of some species remains unclear, in particular for some widely distributed European species that could have been translocated from one basin to another in a remote past. As an example, following Kottelat and Freyhof (2007) and Olivier and Carrel (2020), we considered the European bitterling as native, but recent work suggests that this species could have been introduced in Western Europe following the diffusion of the common carp in fish ponds (Van Damme et al., 2007).
We hypothesized that, over the long term, the establishment of non-native fish species had steadily increased and even accelerated. Our results only partially support this hypothesis insofar as we observed two phases of accelerating fish introductions (the second being indeed more pronounced than the first) separated by a period of relative slowdown. The second half of the 19th century, constituted a first phase in accelerating the establishment of new species over the basin, driven both by scientific and technical advancements and a collective desire led by the public authorities to improve the production of natural systems through the establishment of species from other countries. This period was indeed marked by important advancements in fish biology and physiology, allowing a better control of their rearing conditions. The control of artificial reproduction has considerably facilitated the diffusion of fish species in the form of embryonated eggs, in particular for salmonids which are difficult to transport as juveniles or adults (Haxo, 1853; Vivier, 1956). In 1854, French Zoological Society of Acclimatisation was founded, with one of its main sections dedicated to freshwater species (Luglia, 2015). This resulted in numerous attempts to introduce new fish species, often from North American origin, motivated both by scientific curiosity and the desire to improve production in rivers, lakes and ponds (Keith et al., 2011). Remarkably, many of these attempts have failed more or less quickly and few of the species introduced at that time are still present today. It shows that although the rearing and spawning of species were well known and managed, their ecological requirements were generally neglected, leading to numerous introduction attempts into poorly suitable ecosystems. The number of attempts to introduce new species decreased considerably at the beginning of the 20th century (even if, contrary to the previous period, all these attempts resulted in a lasting establishment) probably in connection with the disruptions generated by the two World Wars. Furthermore, since the end of the 19th century, the scientific community has been stirred up by debates questioning the relevance of non-native species introductions, firstly in terms of their real economic benefits and then more broadly regarding the threats they may represent for ecosystems and native species (Luglia, 2015). Thus, at the turn of the 19th and 20th centuries, the French Zoological Society of Acclimatization gradually approached fish introductions with increasing caution, favoring instead measures such as reducing pollution or restoring ecological continuity to ensure a proper management of freshwaters (Luglia, 2014).
The establishment of non-native species started again, with even more strength, in the second half of the 20th century and since the beginning of the 21st century has reached an unprecedented level with the arrival, on average, of four new species per decade (even if not all of them will probably become established). Contrary to the 19th century, when there was a coordinated joint determination of public authorities and the scientific community, recent introductions appear to occur according to much more disparate modalities and motivations (Keith et al., 2011). Some of them were driven by zootechnical concerns such as bighead carp (H. molitrix) and grass carp (C. idella) initially introduced to regulate the development of algae and macrophytes in water bodies or exotic sturgeons to diversify fish farming production. Others were promoted by anglers as in the case of pikeperch (Sander lucioperca), grayling (Thymallus thymallus), or European catfish (Silurus glanis). However, several introductions seem to have occurred unintentionally, in particular through the transfer of restocking fish containing unwanted species (e.g., Pseudorasbora parva or Umbra pygmaea).
In addition to intentional and unintentional direct introductions, the transport of biological organisms by commercial shipping and opportunities for colonization through navigation waterways connecting previously isolated river basins appear as a major driver for the spread of non-native freshwater species. This phenomenon is becoming all the more relevant since most of the major rivers in western and central Europe are nowadays interconnected, favoring the exchange of numerous species (Copp et al., 2005; Keller et al., 2011). For the Seine River basin, the role of canals connecting basins in the colonization of the ruffe (Gymnocephalus cernua) and the nase (Chondrostoma nasus) during the 18th and 19th centuries had already been described (Nelva, 1997). The very recent arrivals of the asp (Aspius aspius), the round goby (Neogobius melanostomus), and tubenose goby (Proterorhinus semilunaris) seem associated with this phenomenon and more broadly with commercial river traffic. Indeed, the first areas where these species were observed in the basin closely match the river reaches mostly affected by commercial navigation and are in line with a general East-West extension across Europe along the main waterways (Beisel et al., 2017).
In some ways, the considerable increase in non-native species since the middle of the 20th century may seem paradoxical insofar as it takes place during a period when regulatory actions and measures have been progressively strengthened to limit or even prohibit the introduction of non-native species (Copp et al., 2005; Sarat et al., 2015). In fact, this trend observed for fish from the Seine River basin illustrates of a much more global phenomenon combining globalization of commodities and people circulation and artificialization of ecosystems leading to a rapid expansion of a wide variety of non-native species in most continents (Meyerson and Mooney, 2007). Moreover, the temporal pattern of non-native fish introduction marked by a first wave at the end of the 19th century and a second acceleration phase even more pronounced from the middle of the 20th century is not specific to the Seine basin and is found in a rather similar way in other European or North American areas (Copp et al., 2005; Wolter and Röhr, 2010).
A commonly cited rule of thumb, the so-called tens rule, postulates that only 10% of species introduction attempts result in a successful establishment and that only 10% of these established species become invasive (Williamson and Fitter, 1996). Our long-term results for fish from the Seine basin clearly conflict with the first postulate of the tens rule regarding the rate of successful introductions, since the number of species that seem to be naturalized in the basin exceeds 50% of the known attempts at introduction or colonizations. It is possible that some introductions escaped from our census, especially when they are unintentional or illegally conducted, and that some species currently considered as well established could eventually become extinct. These two possibilities would lead to a lower estimate of the rate of successful introductions, but very unlikely enough to approach the 10% level.
An invasive species is a species that, by colonizing a new environment (with or without the help of humans), modifies living communities, the functioning of ecosystems and potentially affects the human activities occurring therein (Mooney and Hobbs, 2000). A list of invasive exotic species has been established at the European level (European Commission, 2016, 2019) including four freshwater fish species, two of which occurring in the Seine River basin. There is also a list for metropolitan France established by the Muséum National d’Histoire Naturel – INPN,2 which includes eleven species, eight of which being present in the Seine basin (see Supplementary Table 1). Considering the French list, which seems better suited to the context of the Seine basin, it may be considered that 36% of the fish species naturalized in the basin are potentially invasive. Once again, this assessment is substantially higher than statement of the tens rule. These observations are consistent with several previous studies suggesting that invasiveness potential would be higher in freshwater fish compared to commonly accepted assessments (García-Berthou et al., 2005; Ruesink, 2005; Leprieur et al., 2009; Lapointe et al., 2012).
Recent Changes in Local Communities
While, at the scale of the whole Seine River basin, non-native species represent almost half of overall recorded species, their proportion in local communities remains much lower. Typically, non-native fish currently account for a few percent of the total species number and less than 1% of individuals in local communities, but there are considerable disparities between sites. This discrepancy between basin and local scales in the fraction of non-native species illustrates the fact that many of these species still have very restricted distribution ranges and small population sizes. Interestingly, we found that 30% of the study sites never hosted any non-native species during monitoring, suggesting that a substantial portion of the rivers in the basin have not yet been colonized by them.
We postulated an amplification of the increase in non-native species within local communities during the last three decades. Our results only partially support this hypothesis. We found indeed a general increase in non-native species in communities over the past three decades both in terms of number of species and abundance of fish individuals, but this trend tended to slow down, and even reverse slightly for some metrics, in recent years. While the values of the species numbers and individual abundance of non-native fish predicted by our models remain modest, their increases from the initial 1990 levels appear to be quite dramatic (increases of 70 and 340% for the number of species and abundance of individuals, respectively). Although over the same period native species have also increased, the proportion of non-native fish species and individuals in communities have grown overall.
The progressive increase of non-native species within river fish communities is a widespread phenomenon reported worldwide (see Taylor et al., 2001; Mercado-Silva et al., 2006; Matsuzaki et al., 2013; Carosi et al., 2015; Liu et al., 2017, e.g., for few examples). In French river fish communities, several recent studies show an increase in species number and density of individuals over the last decades related to multiple factors including global warming, improved water quality and the expansion of non-native species (Daufresne and Boët, 2007; Poulet et al., 2011; Maire et al., 2019; Santos et al., 2021). Our results are in line with these findings and suggest, for the Seine River basin, a stronger progression of non-native species compared to native ones in local fish communities over the last three decades. This trend could be explained by the regular arrival of new species and their spread by the progressive colonization of favorable areas remaining unoccupied until now. But it could also result from long-term environmental changes (e.g., hydroclimatic changes, changes in land use, etc.) which would make rivers more favorable to some species and which would have benefited non-native species more than native ones. On the basis of our results, it is not possible to determine the contribution of these two phenomena, but it is likely that they acted jointly.
In accordance with our starting assumption, our results suggest that sites located on large rivers and those heavily exposed to anthropogenic pressures support more non-native fish species and have also experienced a greater increase in these species over the past few decades. The positive relationship between non-native fish species and river size is a widespread pattern encountered on a global or continental scale between river catchments and on a regional scale between different river sections (Marchetti et al., 2004; Blanchet et al., 2009; Milardi et al., 2019). Two hypotheses are generally put forward to explain this positive relationship. The first one considers that when the size of a system is greater, its chances of being reached and thus colonized by a new species increase. The second hypotheses considers that areas supporting more native species offer better ecological conditions and more abundant resources – which is the case for large rivers compared to smaller ones – and are consequently also more favorable to the establishment of non-native species (Fridley et al., 2007).
The link between the establishment of non-native species and the intensity of human pressures and activities is also a widely acknowledged pattern (Meyerson and Mooney, 2007). Regarding more specifically freshwater fish, this situation has led to the use of non-native species as indicator of health and integrity of rivers and lakes, as their presence can both provide a symptom of anthropogenic alteration and become a source of disturbance for ecosystems (Kennard et al., 2005). The greater incidence of non-native species in highly human impacted ecosystems can first be explained by the higher human densities and activities which logically increase the probability for transferring new species either by direct introductions or by facilitating the transport and diffusion of biological propagules. It can also be explained by the weakening of the anthropized ecosystems and their biological communities which then become less resistant to the colonization of new species. Moreover, regarding freshwater ecosystems specifically, human impact frequently results in eutrophication, which seems to promote the establishment of tolerant non-native species (Strayer, 2010). The population density we used in our analyses is a global and very coarse indicator of human pressures, closely linked to urbanization issues. Consequently it probably fails to account for certain pressures such as diffuse agricultural pollution, which can be particularly intense in some sparsely inhabited parts of the basin. In addition, human population density alone does not allow identifying which type of activities or facilities might be particularly involved in the establishment of non-native fish. Numerous studies have pointed out the determining role of navigation and its associated infrastructures in the arrival and diffusion of non-native freshwater species (Copp et al., 2005). Ponds and reservoirs also appear as key habitats for the naturalization of certain species that are subsequently likely to colonize the hydrographic network (Havel et al., 2005; Clavero and Hermoso, 2011). These mechanisms are certainly operating in the Seine basin and their relative contribution deserves to be addressed in further details.
Perspectives
Our results clearly indicate that in the Seine River basin, the establishment of non-native species has tended to strengthen and even accelerate both on the scale of the entire basin over the last millennium. Considering that the spread of non-native species is favored by the globalization of the circulation of people and commodities and by the artificialization of ecosystems, it is very likely that the arrival of new fish species in the Seine Basin will continue at a steady rate, at least for the coming decades. In Europe, we are currently witnessing a spreading process of aquatic species from east to west of the continent favored by the progressive implementation of connections between the large river basins previously isolated (Copp et al., 2005). In this context, several fish species have already reached the Seine basin, some of them very recently. Others, such as Romanogobio belingi, Ballerus sapa, Ponticola kessleri, or Neogobius fluviatilis, are now present on the neighboring Rhine or the Maas catchments (Keith et al., 2020) and could colonize the Seine River in the near future. The Amur sleeper Perccottus glenii in full expanding phase in Eastern and Central Europe could also arrive quite soon (Reshetnikov, 2010).
Since the first half of the 20th century, French regulations have been progressively strengthened to restrain or even prevent direct introductions of non-native species into rivers. Our results show that these regulation systems have not been sufficient to slow down the arrival of new species in the Seine River basin. Nevertheless, it can be assumed that without their implementation, the number of non-native species would currently be much higher. Furthermore, it should be noted that non-native species introduction are still possible in closed water bodies from which they can escape and reach the hydrographic network. This concern becomes even more critical as the trade in ornamental species (for aquariums and garden ponds) is growing (Evers et al., 2019).
The results obtained at local fish community level for the last three decades add nuance to the whole-basin view of the extent of non-native species establishment, with implications on management. The slight decline in the establishment of non-native species in fish communities in recent years raises questions. Is this an encouraging result of the regulatory and management measures already in place, or is it just the result of natural fluctuations in a general trend of increase? The recent or probable arrival of several species with a high invasive potential, mentioned above, would rather lead to support the second hypothesis. Large rivers and sites under high anthropogenic pressures support more non-native species and have experienced the greatest increases. These environments should therefore deserve special attention in order to limit their potential role as a source for non-native species, contributing to their wider expansion in the hydrographic network. Our results also suggest that reducing specific anthropogenic pressures could help to mitigate the establishment of non-native species. However, the proxy we used to describe the pressure level is too coarse to identify precisely the human pressures and activities that would constitute the best drivers. The decrease over the last three decades in the importance of non-native species in small streams is a noteworthy observation suggesting that their progression is not an absolutely inexorable trend, at least on a local scale. Our analysis did not allow for the determination of the factors responsible for this decrease. Their identification would be important to support more informed management measures that can be applied to other rivers.
Ongoing climate change is likely to deeply affect the distribution and colonization dynamics of non-native species in the basin (Rahel and Olden, 2008). Warming of water temperatures should negatively affect the few cold-water non-native species, but for many warm-water non-native species, these new conditions should result in population growth and spatial spread, particularly in the upstream direction (Carosi et al., 2021). In this context, large and highly anthropized rivers which represent reservoirs of non-native species could play a determining role in this colonization dynamic. In the long term, climate change could also result in new ecological conditions that would become favorable to species that are not yet able to establish in the region. Finally, warmer and drier conditions could encourage the construction of infrastructures such as dams or reservoirs to ensure water supply and irrigation, which potentially constitute hotspots for the development of invasive species (Havel et al., 2005).
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics Statement
Ethical review and approval was not required for this study, since fish historical sources and monitoring data were pre-existing and available on public sites.
Author Contributions
JBe conceptualized the study and performed the statistical analysis. SB performed the statistical analysis. JBo and AZ compiled and formatted monitoring data on fish communities, associated environmental variables, and contributed to their analysis. All authors contributed to the study and critically contributed to writing of manuscript.
Funding
This study did not receive specific funding. However, it benefited from data and ideas developed in previous projects which received financial support from the PIREN-Seine research program (https://www.piren-seine.fr/en), the Seine-Normandie Water Agency and ONEMA (The French National Agency for Water and Aquatic Environments – now AFB French Biodiversity Agency).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We are especially grateful to all persons, known or anonymous, who contributed, over time, to the collection of fish data. We thank Evelyne Tales for her comments and suggestions on a first draft of the manuscript. We are grateful to the two reviewers for their helpful comments. We thank Mike Wheatley for improving the English on the early manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.687451/full#supplementary-material
Supplementary Table 1 | List of non-native species recorded in the Seine River basin.
Footnotes
- ^ https://www.observatoire-poissons-seine-normandie.fr
- ^ https://inpn.mnhn.fr/espece/listeEspeces/statut/metropole/J accessed on January 20, 2021.
References
Anderson, W. (1992). Climates of opinion: acclimatization in nineteenth-century France and England. Vic. Stud. 35, 135–157.
Balon, E. K. (2004). About the oldest domesticates among fishes. J. Fish Biol. 65, 1–27. doi: 10.1111/j.1095-8649.2004.00563.x
Bates, D., Mächler, M., Bolker, B., and Walker, S. (2015). Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 67, 1–48. doi: 10.18637/jss.v067.i01
Beisel, J.-N., Peltre, M.-C., Kaldonski, N., Hermann, A., and Muller, S. (2017). Spatiotemporal trends for exotic species in French freshwater ecosystems: where are we now? Hydrobiologia 785, 293–305. doi: 10.1007/s10750-016-2933-1
Bellard, C., Cassey, P., and Blackburn, T. M. (2016). Alien species as a driver of recent extinctions. Biol. Lett. 12:20150623. doi: 10.1098/rsbl.2015.0623
Beslagic, S. (2013). Histoire des interactions entre les sociétés humaines et le milieu aquatique durant l’Anthropocène. Évolutions des peuplements piscicoles dans le bassin de la Seine. Ph.D. Thesis. Paris: Panthéon-Sorbonne Université.
Beslagic, S., and Belliard, J. (2014). “L’apport des sciences de l’environnement à la compréhension de l’histoire des milieux: l’exemple des peuplements de poissons du bassin de la Seine au regard des données archéologiques et historiques,” in Journées archéologiques de Haute-Normandie, eds S. Beslagic and J. Belliard (Rouen: Presses universitaires de Rouen et du Havre).
Beslagic, S., Marinval, M. C., and Belliard, J. (2013). CHIPS: a database of historic fish distribution in the Seine River basin (France). Cybium 37, 75–93.
Billen, G., Barles, S., Garnier, J., Rouillard, J., and Benoit, P. (2009). The food-print of Paris: long-term reconstruction of the nitrogen flows imported into the city from its rural hinterland. Reg. Environ. Change 9, 13–24. doi: 10.1007/s10113-008-0051-y
Billen, G., Garnier, J., Némery, J., Sebilo, M., Sferratore, A., Barles, S., et al. (2007). A long-term view of nutrient transfers through the Seine river continuum. Sci. Total Environ. 375, 80–97. doi: 10.1016/j.scitotenv.2006.12.005
Blanchet, S., Leprieur, F., Beauchard, O., Staes, J., Oberdorff, T., and Brosse, S. (2009). Broad-scale determinants of non-native fish species richness are context-dependent. Proc. R. Soc. B 276, 2385–2394. doi: 10.1098/rspb.2009.0156
Bolker, B. M., Brooks, M. E., Clark, C. J., Geange, S. W., Poulsen, J. R., Stevens, M. H. H., et al. (2009). Generalized linear mixed models: a practical guide for ecology and evolution. Trends Ecol. Evol. 24, 127–135. doi: 10.1016/j.tree.2008.10.008
Carosi, A., Ghetti, L., and Lorenzoni, M. (2021). The role of climate changes in the spread of freshwater fishes: implications for alien cool and warm-water species in a mediterranean basin. Water 13:347. doi: 10.3390/w13030347
Carosi, A., Ghetti, L., Forconi, A., and Lorenzoni, M. (2015). Fish community of the river Tiber basin (Umbria-Italy): temporal changes and possible threats to native biodiversity. Knowl. Manag. Aquat. Ecosyst. 416:16. doi: 10.1051/kmae/2015018
CEN (2003). Water quality – sampling of fish with electricity European Standard – EN 14011. Brussels: European Committee for Standardization.
Clavero, M., and Hermoso, V. (2011). Reservoirs promote the taxonomic homogenization of fish communities within river basins. Biodivers. Conserv. 20, 41–57. doi: 10.1007/s10531-010-9945-3
Copp, G. H., Bianco, P. G., Bogutskaya, N. G., Eros, T., Falka, I., Ferreira, M. T., et al. (2005). To be, or not to be, a non-native freshwater fish? J. Appl. Ichthyol. 21, 242–262. doi: 10.1111/j.1439-0426.2005.00690.x
Corbin, J. D., and D’Antonio, C. M. (2004). Competition between native perennial and exotic annual grasses: implications for an historical invasion. Ecology 85, 1273–1283. doi: 10.1890/02-0744
Crawley, M. J. (2002). Statistical Computing: An Introduction to Data Analysis Using S-PLUS. Hoboken, NJ: John Wiley & Sons.
Dassonville, N., Vanderhoeven, S., Vanparys, V., Hayez, M., Gruber, W., and Meerts, P. (2008). Impacts of alien invasive plants on soil nutrients are correlated with initial site conditions in NW Europe. Oecologia 157, 131–140. doi: 10.1007/s00442-008-1054-6
Daufresne, M., and Boët, P. (2007). Climate change impacts on structure and diversity of fish communities in rivers. Global Change Biol. 13, 2467–2478. doi: 10.1111/j.1365-2486.2007.01449.x
David, P., Thébault, E., Anneville, O., Duyck, P. F., Chapuis, E., and Loeuille, N. (2017). Impacts of invasive species on food webs: a review of empirical data. Adv. Ecol.Res. 56, 1–60. doi: 10.1016/bs.aecr.2016.10.001
Doherty, T. S., Glen, A. S., Nimmo, D. G., Ritchie, E. G., and Dickman, C. R. (2016). Invasive predators and global biodiversity loss. Proc. Natl. Acad. Sci. U. S. A. 113, 11261–11265. doi: 10.1073/pnas.1602480113
Emery-Butcher, H. E., Beatty, S. J., and Robson, B. J. (2020). The impacts of invasive ecosystem engineers in freshwaters: a review. Freshwater Biol. 65, 999–1015. doi: 10.1111/fwb.13479
European Commission (2016). Implementing Regulation (EU) 2016/1141 of 13 July 2016 adopting a list of invasive alien species of Union concern pursuant to Regulation (EU) No 1143/2014 of the European Parliament and of the Council. Brussels: European Commission.
European Commission (2019). Implementing Regulation (EU) 2019/1262 of 25 July 2019 amending Implementing Regulation (EU) 2016/1141 to update the list of invasive alien species of Union concern. Brussels: European Commission.
Evers, H. G., Pinnegar, J. K., and Taylor, M. I. (2019). Where are they all from? – sources and sustainability in the ornamental freshwater fish trade. J. Fish Biol. 94, 909–916. doi: 10.1111/jfb.13930
Flipo, N., Lestel, L., Labadie, P., Meybeck, M., and Garnier, J. (2021). “Trajectories of the Seine River Basin,” in Handbook of Environmental Chemistry 90. The Seine River Basin, eds N. Flipo, P. Labadie, and L. Lestel (Cham: Springer), 1–28. doi: 10.1007/698_2019_437
Fridley, J. D., Stachowicz, J. J., Naeem, S., Sax, D. F., Seabloom, E. W., Smith, M. D., et al. (2007). The invasion paradox: reconciling pattern and process in species invasions. Ecology 88, 3–17. doi: 10.1890/0012-9658(2007)88[3:tiprpa]2.0.co;2
García-Berthou, E., Alcaraz, C., Pou-Rovira, Q., Zamora, L., Coenders, G., and Feo, C. (2005). Introduction pathways and establishment rates of invasive aquatic species in Europe. Can. J. Fish. Aquat.Sci. 62, 453–463. doi: 10.1139/f05-017
Gozlan, R. E. (2008). Introduction of non-native freshwater fish: is it all bad? Fish Fish. 9, 106–115. doi: 10.1111/j.1467-2979.2007.00267.x
Havel, J. E., Lee, C. E., and Vander Zanden, M. J. (2005). Do reservoirs facilitate invasions into landscapes? BioScience 55, 518–525. doi: 10.1641/0006-3568(2005)055[0518:drfiil]2.0.co;2
Haxo, J. (1853). De la fécondation artificielle des oeufs de poissons et de leur éclosion au moyen des procédés découverts par MM. Remy et Géhin, de la Bresse (Vosges), pour assurer le repeuplement des cours d’eau. Annales de la Société d’émulation du département des Vosges 8, 13–93.
Hoffmann, R. C. (1995). Environmental change and the culture of common carp in medieval Europe. Guelph Ichthyol. Rev. 3, 57–85.
Hoffmann, R. C. (1996). Economic development and aquatic ecosystems in medieval Europe. Am. Hist. Revi. 101, 631–668. doi: 10.2307/2169418
Hoffmann, R. C. (2005). A brief history of aquatic resource use in medieval Europe. Helgol. Mar. Res. 59, 22–30. doi: 10.1007/s10152-004-0203-5
IPBES (2019). “Summary for policymakers of the global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Service,” in IPBES, eds S. Díaz, J. Settele, E. S. Brondízio, H. T. Ngo, M. Guèze, J. Agard, et al. (Bonn: Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services).
Keith, P., Persat, H., Feunteun, E., and Allardi, J. (2011). Les poissons d’eua douce de France. Mèze, Paris: Biotope, Museum national d’Histoire Naturelle.
Keith, P., Poulet, N., Denys, G., Changeux, T., Feunteun, E., and Persat, H. (2020). Les poissons d’eua douce de France. Mèze, Paris: Biotope, Museum national d’Histoire Naturelle.
Keller, R. P., Drake, J. M., Drew, M. B., and Lodge, D. M. (2011). Linking environmental conditions and ship movements to estimate invasive species transport across the global shipping network. Divers. Distrib. 17, 93–102. doi: 10.1111/j.1472-4642.2010.00696.x
Kennard, M. J., Arthington, A. H., Pusey, B. J., and Harch, B. D. (2005). Are alien fish a reliable indicator of river health? Freshwater Biol. 50, 174–193. doi: 10.1111/j.1365-2427.2004.01293.x
Kottelat, M., and Freyhof, J. (2007). Handbook of European freshwater fishes. Cornol, Switzerland - Berlin, Germany: Kottelat and Freyhof.
Lapointe, N. W. R., Pendleton, R. M., and Angermeier, P. L. (2012). A comparison of approaches for estimating relative impacts of non-native fishes. Environ. Manag. 49, 82–95. doi: 10.1007/s00267-011-9767-4
Leprieur, F., Brosse, S., García-Berthou, E., Oberdorff, T., Olden, J. D., and Townsend, C. R. (2009). Scientific uncertainty and the assessment of risks posed by non-native freshwater fishes. Fish Fish. 10, 88–97. doi: 10.1111/j.1467-2979.2008.00314.x
Leuven, R. S. E. W., van der Velde, G., Baijens, I., Snijders, J., van der Zwart, C., Lenders, H. J. R., et al. (2009). The river Rhine: a global highway for dispersal of aquatic invasive species. Biol. Invasions 11, 1989–2008. doi: 10.1007/s10530-009-9491-7
Liu, C., He, D., Chen, Y., and Olden, J. D. (2017). Species invasions threaten the antiquity of China’s freshwater fish fauna. Divers. Distrib. 23, 556–566. doi: 10.1111/ddi.12541
Luglia, R. (2014). Le savant, le saumon et l’ingénieur. La Société d’acclimatation, l’État et le dépeuplement des cours d’eau à la fin du XIXe siècle. Pour Mémoire, revue du Comité d’histoire du ministère de l’Écologie 14, 88–97.
Luglia, R. (2015). Des savant pour protéger la nature. La Société d’acclimatation (1854-1960). Rennes: Presses universitaires de Rennes.
Lymbery, A. J., Morine, M., Kanani, H. G., Beatty, S. J., and Morgan, D. L. (2014). Co-invaders: the effects of alien parasites on native hosts. Int. J. Parasitol. Parasites Wildl. 3, 171–177. doi: 10.1016/j.ijppaw.2014.04.002
Maire, A., Thierry, E., Viechtbauer, W., and Daufresne, M. (2019). Poleward shift in large-river fish communities detected with a novel meta-analysis framework. Freshwater Biol. 64, 1143–1156. doi: 10.1111/fwb.13291
Marchetti, M. P., Light, T., Moyle, P. B., and Viers, J. H. (2004). Fish invasions in California watersheds: testing hypotheses using landscape patterns. Ecol. Appl. 14, 1507–1525. doi: 10.1890/03-5173
Matsuzaki, S. S., Sasaki, T., and Akasaka, M. (2013). Consequences of the introduction of exotic and translocated species and future extirpations on the functional diversity of freshwater fish assemblages. Global Ecol. Biogeogr. 22, 1071–1082. doi: 10.1111/geb.12067
Mazza, G., Tricarico, E., Genovesi, P., and Gherardi, F. (2014). Biological invaders are threats to human health: an overview. Ethol.Ecol. Evol. 26, 112–129. doi: 10.1080/03949370.2013.863225
Mercado-Silva, N., Lyons, J., Díaz-Pardo, E., Gutiérrez-Hernández, A., Ornelas-García, C. P., Pedraza-Lara, C., et al. (2006). Long-term changes in the fish assemblage of the Laja River. Guanajuato, central Mexico. Aquat. Conserv. Mar. Freshwater Ecosyst. 16, 533–546. doi: 10.1002/aqc.737
Meybeck, M., Lestel, L., Carré, C., Bouleau, G., Garnier, J., and Mouchel, J. M. (2018). Trajectories of river chemical quality issues over the Longue Durée: the Seine River (1900S–2010). Environ. Sci. Pollut. Res. 25, 23468–23484. doi: 10.1007/s11356-016-7124-0
Meyerson, L. A., and Mooney, H. A. (2007). Invasive alien species in an era of globalization. Front. Ecol. Environ. 5, 199–208. doi: 10.1890/1540-929520075[199:IASIAE]2.0.CO;2
Milardi, M., Gavioli, A., Soininen, J., and Castaldelli, G. (2019). Exotic species invasions undermine regional functional diversity of freshwater fish. Sci. Rep. 9:17921. doi: 10.1038/s41598-019-54210-1
Mooney, H. A., and Cleland, E. E. (2001). The evolutionary impact of invasive species. Proc. Natl. Acad. Sci. U. S. A. 98, 5446–5451. doi: 10.1073/pnas.091093398
Mooney, H. A., and Hobbs, R. J. (2000). Invasive Species In A Changing World. Washington: Island Press.
Moorhouse, T. P., and Macdonald, D. W. (2015). Are invasives worse in freshwater than terrestrial ecosystems? WIREs Water 2, 1–8. doi: 10.1002/wat2.1059
Muhlfeld, C. C., Kalinowski, S. T., McMahon, T. E., Taper, M. L., Painter, S., Leary, R. F., et al. (2009). Hybridization rapidly reduces fitness of a native trout in the wild. Biol. Lett. 5, 328–331. doi: 10.1098/rsbl.2009.0033
Nelva, A. (1997). The penetration of the nase, chondrostoma nasus nasus (pisces, cyprinidae), in the french hydrographic network and its consequences. B. Fr. Pêche Piscic. 34, 253–269. doi: 10.1051/kmae:1997027
Olivier, J.-M., and Carrel, G. (2020). “La bouvière Rhodeus amarus (Block, 1782),” in Les poissons d’eau douce de France, eds P. Keith, N. Poulet, G. Denys, T. Changeux, E. Feunteun, and H. Persat (Mèze, Paris: Biotope Editions, Muséum national d’Histoire Naturelle), 307–309.
Pejchar, L., and Mooney, H. A. (2009). Invasive species, ecosystem services and human well-being. Trends Ecol. Evol. 24, 497–504. doi: 10.1016/j.tree.2009.03.016
Poulet, N., Beaulaton, L., and Dembski, S. (2011). Time trends in fish populations in metropolitan France: insights from national monitoring data. J. Fish Biol. 79, 1436–1452. doi: 10.1111/j.1095-8649.2011.03084.x
Pyšek, P., Hulme, P. E., Simberloff, D., Bacher, S., Blackburn, T. M., Carlton, J. T., et al. (2020). Scientists’ warning on invasive alien species. Biol. Rev. 95, 1511–1534. doi: 10.1111/brv.12627
Rahel, F. J., and Olden, J. D. (2008). Assessing the effects of climate change on aquatic invasive species. Conserv. Biol. 22, 521–533. doi: 10.1111/j.1523-1739.2008.00950.x
Reshetnikov, A. N. (2010). The current range of Amur sleeper Perccottus glenii Dybowski, 1877 (Odontobutidae, Pisces) in Eurasia. Russ. J. Biol. Invasions 1, 119–126. doi: 10.1134/s2075111710020116
Rhymer, J. M., and Simberloff, D. (1996). Extinction by hybridization and introgression. Annu. Rev. Ecol. and Syst. 27, 83–109. doi: 10.1146/annurev.ecolsys.27.1.83
Ruesink, J. L. (2005). Global analysis of factors affecting the outcome of freshwater fish introductions. Conserv. Biol. 19, 1883–1893. doi: 10.1111/j.1523-1739.2005.00267.x-i1
Sala, O. E., Chapin Iii, F. S., Armesto, J. J., Berlow, E., Bloomfield, J., Dirzo, R., et al. (2000). Global biodiversity scenarios for the year 2100. Science 287, 1770–1774. doi: 10.1126/science.287.5459.1770
Santos, R., Poulet, N., and Besnard, A. (2021). Life-history traits correlate with temporal trends in freshwater fish populations for common European species. Freshwater Biol. 66, 317–331. doi: 10.1111/fwb.13640
Sarat, E., Mazaubert, E., Dutartre, A., Poulet, N., and Soubeyran, Y. (2015). Les espèces exotiques envahissantes dans les milieux aquatiques: connaissances pratiques et expériences de gestion, Vol. 1. Vincennes: Onema.
Simberloff, D. (2011). How common are invasion-induced ecosystem impacts? Biol. Invasions 13, 1255–1268. doi: 10.1007/s10530-011-9956-3
Strayer, D. L. (2010). Alien species in fresh waters: ecological effects, interactions with other stressors, and prospects for the future. Freshwater Biol. 55, 152–174. doi: 10.1111/j.1365-2427.2009.02380.x
Taylor, C. A., Knouft, J. H., and Hiland, T. M. (2001). Consequences of stream impoundment on fish communities in a small North American drainage. River Res. Appl. 17, 687–698. doi: 10.1002/rrr.629
Van Damme, D., Bogutskaya, N., Hoffmann, R. C., and Smith, C. (2007). The introduction of the European bitterling (Rhodeus amarus) to west and central Europe. Fish Fish. 8, 79–106. doi: 10.1111/j.1467-2679.2007.00239.x
Vivier, P. (1956). Un important centenaire: Remy, Géhin, Haxo, Coste et l’établissement domanial de pisciculture d’Huningue (1843-1853-1953). B. Fr. Pisc. 181, 121–139. doi: 10.1051/kmae:1956004
Williamson, M., and Fitter, A. (1996). The varying success of invaders. Ecology 77, 1661–1666. doi: 10.2307/2265769
Wolter, C., and Röhr, F. (2010). Distribution history of non-native freshwater fish species in Germany: how invasive are they? J. Appl. Ichthyol 26, 19–27. doi: 10.1111/j.1439-0426.2010.01505.x
Keywords: alien species, exotic species, historical biodiversity, ecological trajectory, Seine River, river fish community
Citation: Belliard J, Beslagic S, Boucault J and Zahm A (2021) Increasing Establishment of Non-native Fish Species in the Seine River Basin: Insights From Medium- and Long-Term Approaches. Front. Ecol. Evol. 9:687451. doi: 10.3389/fevo.2021.687451
Received: 29 March 2021; Accepted: 20 July 2021;
Published: 03 September 2021.
Edited by:
Gertrud Haidvogl, University of Natural Resources and Life Sciences Vienna, AustriaReviewed by:
Pedro Segurado, University of Lisbon, PortugalRob Lenders, Radboud University Nijmegen, Netherlands
Copyright © 2021 Belliard, Beslagic, Boucault and Zahm. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jérôme Belliard, jerome.belliard@inrae.fr